用于改进生物分析和生物加工特性的治疗性免疫球蛋白G4的设计的制作方法
相关申请的引用
本申请要求2016年12月23日提交的美国临时申请no.62/438684的权益,出于所有目的将其完整并入本文。
发明背景
单克隆抗体(mab)及其衍生产物(例如fc-融合蛋白)在治疗一些最具挑战性的人类疾病中起重要作用,原因在于这些类型的生物制剂的安全性、功效和高质量。单克隆抗体市场正在显著快速增长,据估计,到2020年单克隆抗体产品的全球总销售额将达到大约1250亿美元。免疫球蛋白g(igg)是治疗性抗体最常用的骨架。其有四个亚类,igg1、igg2、igg3和igg4。由于igg4抗体的抗炎活性,对于在不希望免疫效应子功能募集情况下的应用,igg4是优选的亚类。
免疫球蛋白g同种型4(igg4)是一种动态的分子,能够通过将重链和附接的轻链(半分子)与来自另一分子的重/轻链对交换而更换fab臂,从而产生双特异性抗体。目前认为igg4fab-臂交换的原因是igg4核心-铰链序列以及ch3结构域中的决定簇。据报道,治疗性野生型igg4可与内源性人igg4交换fab臂,这可能影响它们的治疗活性。为了在设计治疗性igg4时防止体内fab-臂交换,经常将铰链区中的丝氨酸228突变为脯氨酸(按照eu编号为s228p)来产生治疗用igg4。然而,这样的突变可能引起不希望的生物分析和生物加工行为。
因此,仍然需要设计具有改进的生物分析和生物加工特性的治疗性igg4分子。
发明概述
在某些实施方案中,本公开提供了包含重链和轻链的igg4抗体,其中所述重链包含:(a)修饰的igg4ch1区,其具有第196位的赖氨酸残基的取代;或(b)修饰的igg4铰链区,其具有第217位的丝氨酸残基、第220位的甘氨酸残基、第224位的脯氨酸残基或第225位的脯氨酸残基的取代。任选地,igg4抗体进一步包含重链铰链区中第228位的丝氨酸残基的取代。优选地,相对于不包含取代的相应igg4分子(也称为“未修饰的igg4抗体”或“野生型抗体”),所述igg4抗体具有改进的生物分析或生物加工特性。例如,第228位的丝氨酸残基变为脯氨酸(s228p)。举例说明,在第196位的赖氨酸残基变为脯氨酸(k196p);第217位的丝氨酸残基变为脯氨酸(s217p);第220位的甘氨酸残基变为苏氨酸(g220t);第224位的脯氨酸残基变为组氨酸(p224h);第225位的脯氨酸残基变为苏氨酸(p225t)。任选地,igg4抗体是人源化的或人的。任选地,igg4抗体是全长抗体或抗体片段,比如f(ab)2。在某些特定的方面,igg4抗体与选自cd137、cxcr4、etau、csf1r、lag3、pd1、pdl1或kir的靶分子结合。例如,igg4抗体包括在包含氨基酸序列seqidno:1的重链中进行的取代。举例说明,igg4抗体包含选自seqidno:22、25、27、28或29的氨基酸序列。例如,改进的生物分析特性包括分析型阳离子交换色谱(cex)树脂上的单峰洗脱行为,所述树脂包括但不限于propacscx-10、propacwcx-10、
在某些实施方案中,本发明提供一种融合蛋白,其包含:(a)修饰的igg4重链ch1区,其具有第196位的赖氨酸残基的取代;或(b)修饰的igg4重链铰链区,其具有第217位的丝氨酸残基、第220位的甘氨酸残基、第224位的脯氨酸残基或第225位的脯氨酸残基的取代。任选地,igg4抗体包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,融合蛋白进一步包含igg4重链铰链区中第228位的丝氨酸残基的取代(例如,s228突变)。优选地,融合蛋白相对于未修饰(即,不包含igg4重链的第196、217、220、224或225位的氨基酸修饰)的融合蛋白具有改进的生物分析和生物加工特性。
在某些实施方案中,本发明提供了用于改进igg4抗体的生物分析或生物加工特性的方法,所述方法包括通过取代人igg4重链的第196、217、220、224或225位的至少一个残基来修饰igg4抗体,其中,修饰的igg4抗体相对于未修饰的igg4抗体具有改进的生物分析或生物加工特性。例如,修饰的igg4抗体包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,igg4抗体包含s228p突变。任选地,igg4抗体是人源化的或人的。任选地,igg4抗体是全长抗体或抗体片段,比如f(ab)2。在某些特定的方面,igg4抗体与选自cd137、cxcr4、etau、csf1r、lag3、pd1、pdl1或kir的靶分子结合。举例说明,改进的生物分析特性包括分析型阳离子交换色谱(cex)树脂上的单峰洗脱行为,所述树脂包括但不限于propacscx-10、propacwcx-10、
在某些实施方案中,本发明提供用于改进包含igg4重链ch1或铰链区的融合蛋白的生物分析或生物加工特性的方法,所述方法包括通过取代igg4重链的第196、217、220、224或225位的至少一个残基来修饰igg4重链ch1或铰链区,其中修饰的融合蛋白相对于未修饰的融合蛋白具有改进的生物分析或生物加工特性。例如,修饰的融合蛋白包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,融合蛋白包含s228p突变。优选地,改进的生物分析或生物加工特性包括阳离子交换色谱(cex)树脂上的单峰洗脱行为。
在某些实施方案中,本发明提供药物组合物,其包含如上所述的igg4抗体或融合蛋白和(2)药学上可接受的载体。例如,药物组合物是液体制剂。例如,药物组合物被配制为用于肠胃外、皮下、静脉内、肌内、鼻内或肺部递送。任选地,药物组合物进一步包含另一种用于联合治疗的单克隆抗体。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的核酸。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的载体。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列(与启动子可操作连接)的宿主细胞。例如,宿主细胞是哺乳动物细胞。
在某些实施方案中,本发明提供产生如上所述的igg4抗体或融合蛋白的方法,其包括在适于表达igg4抗体或融合蛋白的条件下培养包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的宿主细胞。任选地,所述方法进一步包括分离igg4抗体或融合蛋白。例如,宿主细胞是哺乳动物细胞。
在某些实施方案中,本发明提供了治疗受试者(例如人)的癌症的方法,其包括向受试者施用治疗有效量的如上所述的igg4抗体或融合蛋白。
在某些实施方案中,本发明提供了治疗受试者(例如人)的自身免疫性疾病或炎性疾病的方法,其包括向受试者施用治疗有效量的如上所述的igg4抗体或融合蛋白。
附图简述
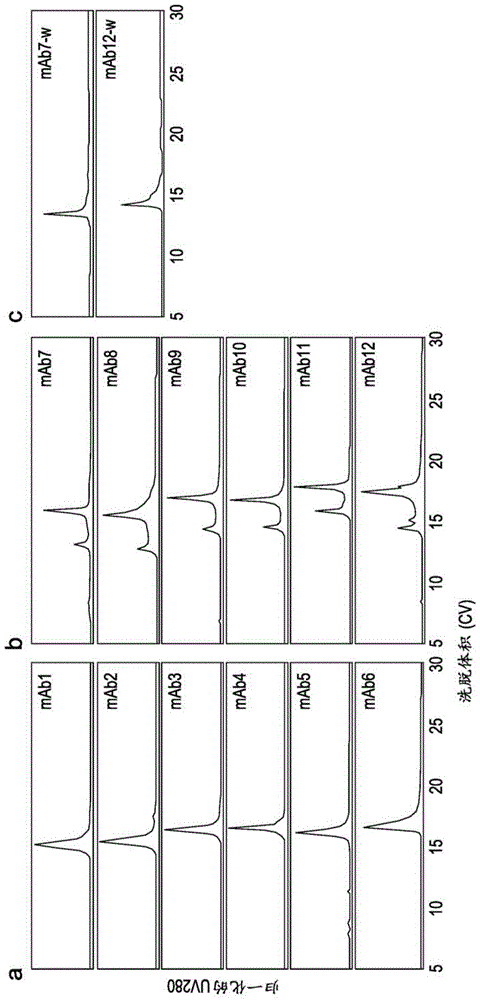
图1显示了治疗性igg4分子的cex-hplc色谱图。所有在铰链中具有丝氨酸到脯氨酸(cppc)突变的治疗性igg4分子均在cex-hplc柱上观察到双峰洗脱行为。所有治疗性igg1分子和野生型igg4均在cex-hplc柱上观察到单峰洗脱行为。cex-hplc方法使用20mmmes(ph5.0)至20mmmes、1mnacl(ph5.0)的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。
图2显示了mab11的sec-hplc色谱图,cex-hplc的峰1和峰2。两个色谱图非常好地重叠,表明两个峰没有大小差异或聚集形成。使用安装在waterscorporation(milford,ma,usa)的watershplc系统上的tosohbioscience(kingofprussia,pa,usa)的tskgelg3000swxl柱进行分析性sec。所述方法使用100mm磷酸钠、100mm硫酸钠(ph6.8),流速为1ml/min,恒定的总注入蛋白质量为100μg。通过uv280nm监测洗脱的蛋白质。
图3显示了治疗性mab11在cex-hplc上形成的峰1和峰2的ice280谱。两个色谱图非常好地重叠,表明这两个峰没有电荷差异。
图4显示了mab11以及峰1和峰2的再注入的cex-hplc色谱图。再注入峰1和峰2后仍然产生两个相似的洗脱模式。cex-hplc方法使用20mmmes至20mmmes、1mnacl在不同ph下的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。
图5a显示了在不同ph下mab11的cex-hplc色谱图。在ph5.0观察到双峰洗脱行为。改变ph值导致两个峰的比例改变。增加ph导致第一峰的百分比更高,降低ph导致第二峰的百分比更高。
图5b显示了在不同ph下mab3的cex-hplc色谱图。伊匹单抗在不同ph下观察到单峰洗脱行为。cex-hplc方法使用20mmmes至20mmmes、1mnacl在不同ph下的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。
图6a-6b显示了mab4和mab12的示例性cex-hplc电泳图(e-gram),其中分别注入抗体(a)或以1:1的比例组合抗体(b)。mab12具有宽的双峰洗脱行为,因此与mab4重叠,使得当以1:1的比例将两种组合抗体注入到cex柱上时,难于进行电荷变体表征。
图6c-6d显示了mab4和mab12的示例性cex-hplc电泳图(e-gram),其中分别注入抗体(c)或以1:1的比例组合抗体(d)。mab12-w具有窄的单峰洗脱行为,因此与mab4不重叠,使得当以1:1的比例将两种组合抗体注入射到cex柱上时,易于进行电荷变体表征。因此保留并且可以清楚地表征两种抗体的电荷变体谱。cex-hplc方法使用20mmmes(ph5.0)至20mmmes、600mmnacl(ph5.0)的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。
图7显示在制备型cex树脂上也观察到igg4的双峰洗脱行为,但igg1则不然。30cv的40mm乙酸钠和氯化钠(0-1.0m),ph=5.0;树脂:porosxs,树脂装填量10g/l。
图8a-8b显示了不同电导率下mab10的盐步骤洗脱。(a)较低电导率下的盐步骤洗脱色谱图;(b)较高电导率下的盐步骤洗脱色谱图。两种电导率可见于图7中的ph5.0的盐梯度洗脱期间。
图9显示了mab11及其片段的sec-hplc色谱图。sec谱指示,使用胃蛋白酶和木瓜蛋白酶分别成功生成高纯度f(ab’)2和f(ab)片段。
图10显示了mab11及其片段的cex-hplc色谱图。使用固定化的胃蛋白酶(thermofisherscientific)生成了mab11f(ab’)2片段。使用固定化的木瓜蛋白酶(thermofisherscientific)生成了mab11f(ab)片段。如预期的,观察到mab11的双峰洗脱行为。对于mab11f(ab’)2片段也观察到双峰洗脱行为,表明可能是f(ab’)2,而不是fc,对双峰洗脱行为做贡献。还在cex-hplc柱上测试了mab13,一种具有igg4(丝氨酸到脯氨酸取代(cppc))fc尾的fc融合蛋白。mab13在cex柱上的单峰洗脱行为进一步证实fc尾不参与双峰洗脱行为。为了进一步缩小可能造成双峰洗脱行为的原因的igg4片段的范围,还将f(ab)注射到cex-hplc柱上。有趣的是,对于f(ab),观察到单峰洗脱行为,而不是双峰洗脱行为。该结果表明igg4(丝氨酸到脯氨酸取代(cppc))铰链在双峰洗脱行为中起重要作用。因此得出结论,igg4(丝氨酸到脯氨酸取代(cppc))f(ab')2的存在是造成双峰洗脱行为的原因。
图11显示了igg4(丝氨酸到脯氨酸取代(cppc))和igg1的恒定重链1(ch1)和铰链区的氨基酸序列比对。在igg4(丝氨酸到脯氨酸取代(cppc))和igg1之间鉴定出总共12个氨基酸差异,用绿色和黄色突出显示。igg4中第一个鉴定出的半胱氨酸氨基酸形成重链和轻链之间的链间二硫键,因此未将半胱氨酸突变为相应的丝氨酸,以便保持igg4的完整结构。将igg4(丝氨酸到脯氨酸取代(cppc))中其余11个氨基酸各自突变为igg1中的相应氨基酸,以评估它们对cex-hplc柱行为的影响。
图12显示了突变体的cex-hplc色谱图。观察到mab12-4c、mab12-7、mab12-9、mab12-10、mab12-11和mab12-w的单峰洗脱行为。mab12-w是野生型mab12,在铰链区中没有丝氨酸到脯氨酸的突变。cex-hplc方法使用20mmmes(ph5.0)至20mmmes、600mmnacl(ph5.0)的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。
图13a-13h显示了突变体的未经还原的完整质量。未经还原的完整质谱证实了mab12、mab12-w、mab12-4c、mab12-7、mab12-9、mab12-10、mab12-11和hca195(bio-rad产的野生型重组人igg4κ)的成功表达。
图14显示了hca195和mab12-w的fab臂交换现象的未经还原的完整质量。mab12-w和hca195两者都是野生型igg4,在铰链区中没有cppc突变。fab臂交换作为一个新的双特异性抗体峰被检测到。将mab12-w与hca195混合,并在脱气的pbs(ph7.2)中与0.5mm还原型谷胱甘肽(gsh;sigma)一起温育。mab12-w和hca195两者的终浓度均为20μg/mm。将混合物(70μl)在37℃温育24小时。然后在完整质量测试之前将样品储存在4℃。
图15显示了hca195和mab12的fab臂交换结果的未还原的完整质量。由于mab12在铰链区中含有cppc基序,未检测到fab臂交换。
图16显示了hca195和mab12-4c的fab臂交换结果的未还原的完整质量。由于mab12-4c在铰链区中含有cppc基序,未检测到fab臂交换。
图17显示了hca195和mab12-7的fab臂交换结果的未还原的完整质量。由于mab12-7在铰链区中含有cppc基序,未检测到fab臂交换。
图18显示了hca195和mab12-9的fab臂交换结果的未还原的完整质量。由于mab12-9在铰链区中含有cppc基序,未检测到fab臂交换。
图19显示了hca195和mab12-10的fab臂交换结果的未还原的完整质量。由于mab12-10在铰链区中含有cppc基序,未检测到fab臂交换。
图20显示了hca195和mab12-11的fab臂交换结果的未还原的完整质量。由于mab12-11在铰链区中含有cppc基序,未检测到fab臂交换。
图21显示了mab12突变的elisa结合测定。elisa结合测定表明,所有突变体都具有如同mab12的合理结合。所述突变没有改变mab12的结合活性。
发明详述
在某些方面,本发明至少部分基于以下发现:具有s228p突变(按照eu编号)的治疗性igg4分子具有不期望的生物分析和生物加工特性。为了解决这个问题,申请人设计了具有改进的生物分析和生物加工特性的变体igg4分子。
1.定义
术语“免疫球蛋白”是指一类结构相关的糖蛋白,其由两对多肽链组成:一对轻(l)的低分子量链和一对重(h)链,所有这四条链都通过二硫键相互连接。免疫球蛋白的结构已充分表征。参见例如fundamentalimmunology第7章(paul,w.编辑,第版,ravenpress,n.y.(1989))。简言之,每条重链通常包含重链可变区(在本文中缩写为vh或vh)和重链恒定区。重链恒定区通常包含三个结构域ch1、ch2和ch3。每条轻链包含轻链可变区(在本文中缩写为vl或vl)和轻链恒定区。轻链恒定区包含一个结构域cl或cl。vh和vl区可以进一步细分为高变性区域(或高变区,其可以在序列上是高变的和/或为在结构上定义的环形式),也称为互补决定区(cdr),所述互补决定区散布有更保守的区域,称为框架区(fr)。每个vh和vl通常由三个cdr和四个fr组成,按照以下顺序从氨基末端到羧基末端排列:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4(还参见chothia和leskj.mol.biol.196,901-917(1987))。
通常,通过kabat等人在sequencesofproteinsofimmunologicalinterest,5thed.publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991)中描述的方法进行氨基酸残基编号。使用该编号系统,肽的实际线性氨基酸序列可以含有对应于可变结构域的fr或hvr的缩短或插入其中的较少或另外的氨基酸。例如,重链可变结构域可以包括在vhcdr2的残基52之后的单个氨基酸插入物(依照kabat的残基52a)和在重链fr残基82之后的插入残基(例如,依照kabat的残基82a、82b和82c等)。通过抗体序列与“标准”kabat编号序列的同源区比对,可以确定给定抗体的残基的kabat编号。
或者,通过kabat等人也在sequencesofproteinsofimmunologicalinterest,5thed.publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991)中描述的eu索引进行氨基酸残基编号。在本申请各处使用该编号。
在本发明上下文中的术语“抗体”(ab)是指免疫球蛋白分子、免疫球蛋白分子的片段或其任一者的衍生物,其具有在典型生理条件下与抗原特异性地结合的能力。免疫球蛋白分子的重链和轻链的可变区含有与抗原相互作用的结合结构域。抗体(ab)的恒定区可介导免疫球蛋白与宿主组织或因子((包括免疫系统的各种细胞(比如,效应细胞)和补体系统的组分比如c1q(补体激活经典途径的第一组分))结合。除非另有说明,否则术语“抗体”也包括通过任何已知技术比如酶切割、肽合成和重组技术提供的多克隆抗体、单克隆抗体(mab)、抗体样多肽比如嵌合抗体,以及人源化抗体、双特异性抗体和保留与抗原特异性地结合的能力的抗体片段(抗原结合片段)。特别是fc结构域中的恒定区结构域(当存在时)优选为igg4同种型,在这种情况下不需要抗体效应子功能。因此,每条重链优选地包含igg4ch2结构域和ch3结构域。
如本文中使用的术语“人抗体”预期包括具有源于人种系免疫球蛋白序列的可变区和恒定区的抗体。本发明的人抗体可以包括并非由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机诱变或定点诱变或者通过体内体细胞突变引入的突变)。然而,如本文中使用的,术语“人抗体”预期不包括这样的抗体:其中源自另一个哺乳动物物种种系,如小鼠的cdr序列被移植到人框架序列上。
术语“嵌合抗体”是指含有来自一种抗体的一个或多个区域和来自一种或多种其他抗体的一个或多个区域的抗体。术语“嵌合抗体”包括二价和多价抗体。通过本领域熟知的重组过程产生嵌合抗体(参见例如cabilly等人,pnasusa81,3273-3277(1984),morrison等人,pnasusa81,6851-6855(1984),boulianne等人,nature312,643-646(1984),ep125023,neuberger等人,nature314,268-270(1985),ep171496,ep173494,wo86/01533,ep184187,sahagan等人,j.immunol.137,1066-1074(1986),wo87/02671,liu等人,pnasusa84,3439-3443(1987),sun等人,pnasusa84,214-218(1987),better等人,science240,1041-1043(1988)和harlow等人,antibodies:alaboratorymanual,coldspringharborlaboratorypress,coldspringharbor,n.y.,(1988))。
“人源化抗体”是衍生自非人物种的抗体,其中重链和轻链的框架和恒定结构域中的某些氨基酸已经被突变,以避免或消除人类的免疫应答。非人(例如,鼠)抗体的人源化形式是含有衍生自非人免疫球蛋白的最小序列的嵌合抗体。通常,人源化抗体是人免疫球蛋白(受体抗体),其中来自受体的高变区的残基被来自具有期望的特异性、亲和力和能力的诸如小鼠、大鼠、兔或非人灵长类动物等非人物种的高变区(供体抗体)的残基替换。在一些情况下,人免疫球蛋白的fv框架区(fr)残基被相应的非人残基替换。此外,人源化抗体可以包含在受体抗体中或在供体抗体中未发现的残基。进行这些修饰以进一步改善抗体性能。一般而言,人源化抗体将包含至少一个(通常两个)可变结构域中的基本上所有的可变结构域,其中所有或基本上所有的高变环对应于非人免疫球蛋白的高变环,并且所有或基本上所有的fr区是人免疫球蛋白序列的fr区。人源化抗体通常还将包含免疫球蛋白恒定区(fc)(一般是人免疫球蛋白的恒定区)的至少一部分。关于更多详情,参见jones等人,nature321,522-525(1986),riechmann等人,nature332,323-329(1988)和presta,curr.op.struct.biol.2,593-596(1992)。
如本文中使用的“分离的抗体”意指基本上不含具有不同抗原特异性的其他抗体的抗体。然而,与特定人靶抗原的表位、同工型或变体的特异性地结合的分离的抗体可以与其他相关抗原(例如来自其他物种(比如物种同源物))具有交叉反应性。而且,分离的抗体可基本不含其他细胞物质和/或化学品。
如本文中使用的术语“单克隆抗体”或“单克隆抗体组合物”是指具有单分子组成的抗体分子制剂。单克隆抗体组合物对特定表位展示单一结合特异性和亲和力。因此,术语“人单克隆抗体”是指展示单一结合特异性的具有衍生自人种系免疫球蛋白序列的可变区和恒定区的抗体。人单克隆抗体可以由杂交瘤产生,所述杂交瘤包括与永生化细胞融合的获自转基因或转染色体非人动物(比如转基因小鼠)的b细胞,所述b细胞具有包含人重链转基因和轻链转基因的基因组。
如本文中使用的,在抗体与预定抗原结合背景下的术语“结合”通常是具有一定亲和力的结合,在通过例如在biacore3000仪器中使用抗原作为配体并且抗体作为分析物的表面等离子体共振(spr)技术确定时,所述亲和力对应于约10-7m或更低,比如约10-8m或更低,比如约10-9m或更低、约10-10m或更低、或约10-11m或甚至更低的kd。
如本文中使用的,“同种型”是指由重链恒定区基因编码的免疫球蛋白(亚)类,例如igg1、igg2、igg3、igg4、igd、iga、ige或igm。
术语“双特异性抗体”预期包括具有两种不同结合特异性的任何抗体,例如,结合两种不同的表位的抗体,所述表位可位于相同的靶抗原上,或更常见地位于不同的靶抗原上。
如本文中使用的,术语“效应细胞”是指涉及免疫应答的效应阶段而不是免疫应答的识别和激活阶段的免疫细胞。示例性免疫细胞包括髓样或淋巴样起源的细胞,例如淋巴细胞(比如b细胞和t细胞,包括溶细胞性t细胞(ctl))、杀伤细胞、自然杀伤细胞、巨噬细胞、单核细胞、嗜酸性粒细胞、多形核细胞比如中性粒细胞、粒细胞、肥大细胞和嗜碱性粒细胞。一些效应细胞表达特异性fc受体并执行特异性免疫功能。在一些实施方案中,效应细胞能够诱导抗体依赖性细胞介导的细胞毒性(adcc),比如自然杀伤细胞能够诱导adcc。例如,表达fcr的单核细胞、巨噬细胞涉及靶细胞的特异性杀伤并将抗原呈递给免疫系统的其他组分或与呈递抗原的细胞结合。在一些实施方案中,效应细胞可以吞噬靶抗原或靶细胞。特定fcr在效应细胞上的表达可受到体液因子比如细胞因子的调节。
术语“治疗”(“treatment”)或“治疗”(“treating”)是指为了减轻、改善、阻止或根除(治愈)症状或疾病状态而施用有效量的本发明治疗活性分子。
“有效量”是指在剂量和时间段方面有效实现期望的治疗结果所需的量。抗体的治疗有效量可以根据个体的疾病状态、年龄、性别和体重等因素以及抗体以在个体中引出期望应答的能力而变化。治疗有效量是抗体或抗体部分的治疗有益效果超过任何毒性或有害作用的量。
术语“fab臂交换”和“半分子交换”在本文中可互换使用并且是指针对人igg4的蛋白质修饰类型,其中igg4重链和附接的轻链(半分子)被交换为来自另一个igg4分子的重-轻链对。因此,igg4分子可以获得识别两种相异的抗原的两个相异的fab臂(产生双特异性分子),而它们的fc结构域结构保持不变。fab臂交换在体内天然地发生,并且可以通过纯化的血细胞或还原剂比如还原型谷胱甘肽在体外诱导。
除非上下文另外指出,否则术语“蛋白质”和“多肽”在本文中可互换使用。
除非上下文另外指出,否则术语“多核苷酸”包括基因、dna、cdna、rna、mrna。
在本说明书上下文中的术语“包含”应该被解释为“包括”。
在本发明上下文中的术语“野生型”意指可以在自然界中存在或可以从环境中分离的igg4分子,其不包含任何基因工程突变。
2.本发明进一步的方面和实施方案
在某些实施方案中,本发明提供了用于治疗疾病的修饰的igg4分子(也称为修饰的igg4抗体或抗体变体)或修饰的融合蛋白。本发明还提供了编码修饰的igg4抗体或融合蛋白的多核苷酸以及表达修饰的igg4抗体或融合蛋白的细胞系。进一步提供了治疗疾病的方法,其包括施用治疗有效量的修饰的igg4抗体或融合蛋白。
在某些实施方案中,本文中取代突变体的名称由字母后跟数字后跟字母组成。第一个字母表示野生型蛋白质中的氨基酸。数字是指进行氨基酸取代的氨基酸位置,第二个字母表示用于取代野生型氨基酸的氨基酸。常规地根据kabat等人设计的系统将抗体(例如,igg4分子)可变区和恒定区中的残基编号。这个系统于1987年在kabat等人的sequencesofproteinsofimmunologicalinterest,usdepartmentofhealthandhumanservices,nih,usa中提出(此后“kabat等人”)。通过抗体序列与“标准”kabat编号序列的同源残基比对,可以确定给定抗体(例如igg4分子)的残基的正确kabat编号。或者,可以通过eu索引或eu编号系统(同样描述于kabat等人,sequencesofproteinsofimmunologicalinterest,5thed.publichealthservice,nationalinstitutesofhealth,bethesda,md.(1991))进行氨基酸残基编号。抗体中氨基酸残基的另一个编号系统是imgt编号系统(lefranc,m.-p.等人,dev.comp.immunol.,29,185-203(2005))。在本说明书中,使用eu编号系统,另外指出使用kabat编号系统的情况除外。
表1显示了一些示例性的修饰的igg4抗体,其在铰链或ch1区中包含一个或多个突变。
表1.在igg4分子中的突变的代表性清单.
在某些实施方案中,本公开提供了包含重链和轻链的igg4抗体,其中所述重链包含:(a)修饰的igg4ch1区,其具有第196位的赖氨酸残基的取代;或(b)修饰的igg4铰链区,其具有第217位的丝氨酸残基、第220位的甘氨酸残基、第224位的脯氨酸残基或第225位的脯氨酸残基的取代。任选地,igg4抗体进一步包含重链铰链区中第228位的丝氨酸残基的取代。优选地,相对于不包含取代的相应igg4分子(也称为“未修饰的igg4抗体”或“野生型抗体”),所述igg4抗体具有改进的生物分析或生物加工特性。例如,在第228位的丝氨酸残基变为脯氨酸(s228p)。举例说明,第196位的赖氨酸残基变为脯氨酸(k196p);第217位的丝氨酸残基变为脯氨酸(s217p);第220位的甘氨酸残基变为苏氨酸(g220t);第224位的脯氨酸残基变为组氨酸(p224h);第225位的脯氨酸残基变为苏氨酸(p225t)。任选地,igg4抗体是人源化的或人的。任选地,igg4抗体是全长抗体或抗体片段,比如f(ab)2。在某些特定的方面,igg4抗体与选自cd137、cxcr4、etau、csf1r、lag3、pd1、pdl1或kir的靶分子结合。例如,igg4抗体包括在包含氨基酸序列seqidno:1的重链中进行的取代。举例说明,igg4抗体包含选自seqidno:22、25、27、28或29的氨基酸序列。例如,改进的生物分析特性包括在分析型阳离子交换色谱(cex)树脂上的单峰洗脱行为,所述树脂包括但不限于propacscx-10、propacwcx-10、
在某些实施方案中,本发明提供了融合蛋白,其包含:(a)修饰的igg4ch1区,其具有在第196位的赖氨酸残基的取代;或(b)修饰的igg4重链铰链区,其具有第217位的丝氨酸残基、第220位的甘氨酸残基、第224位的脯氨酸残基或第225位的脯氨酸残基的取代。任选地,igg4抗体包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,融合蛋白进一步包含在igg4重链铰链区中第228位的丝氨酸残基的取代(例如,s228突变)。优选地,融合蛋白相对于未修饰的融合蛋白(即,不包含在igg4重链的第196、217、220、224或225位的氨基酸修饰)具有改进的生物分析和生物加工特性。
在某些实施方案中,本发明提供了用于改进igg4抗体的生物分析或生物加工特性的方法,所述方法包括通过取代人igg4重链的第196、217、220、224或225位的至少一个残基来修饰igg4抗体,其中,修饰的igg4抗体相对于未修饰的igg4抗体具有改进的生物分析或生物加工特性。例如,修饰的igg4抗体包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,igg4抗体包含s228p突变。任选地,igg4抗体是人源化的或人的。任选地,igg4抗体是全长抗体或抗体片段,比如f(ab)2。在某些特定的方面,igg4抗体与选自cd137、cxcr4、etau、csf1r、lag3、pd1、pdl1或kir的靶分子结合。举例说明,改进的生物分析特性包括在分析型阳离子交换色谱(cex)树脂上的单峰洗脱行为,所述树脂包括但不限于propacscx-10、propacwcx-10、
在某些实施方案中,本发明提供了用于改进融合蛋白的生物分析或生物加工特性的方法,所述融合蛋白包含igg4重链ch1或铰链区,所述方法包括通过取代igg4重链的第196、217、220、224或225位的至少一个残基来修饰igg4重链ch1或铰链区,其中修饰的融合蛋白相对于未修饰的融合蛋白具有改进的生物分析或生物加工特性。例如,修饰的融合蛋白包含至少一个选自k196p、s217p、g220t、p224h或p225t的突变。任选地,融合蛋白包含s228p突变。优选地,改进的生物分析或生物加工特性包括在阳离子交换色谱(cex)树脂上的单峰洗脱行为。
在某些实施方案中,本发明提供了药物组合物,其包含如上所述的igg4抗体或融合蛋白和(2)药学上可接受的载体。例如,药物组合物是液体制剂。例如,药物组合物被配制为用于肠胃外、皮下、静脉内、肌内、鼻内或肺部递送。任选地,药物组合物进一步包含另一种用于联合治疗的单克隆抗体。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的核酸。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的载体。
在某些实施方案中,本发明提供了包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列(与启动子可操作连接)的宿主细胞。例如,宿主细胞是哺乳动物细胞。
在某些实施方案中,本发明提供产生如上所述的igg4抗体或融合蛋白的方法,其包括在适于表达igg4抗体或融合蛋白的条件下培养包含编码如上所述的igg4抗体或融合蛋白的核苷酸序列的宿主细胞。任选地,所述方法进一步包括分离igg4抗体或融合蛋白。例如,宿主细胞是哺乳动物细胞。
3.制造抗体的方法
产生抗体的方法是本领域熟知的。在优选的实施方案中,本发明抗体是单克隆抗体。单克隆抗体可以例如通过kohler等人首次描述于nature256,495(1975)的杂交瘤方法产生,或者可以通过重组dna方法产生。还可以使用例如clackson等人在nature352,624-628(1991)中和marks等人在j.mol.biol.222,581-597(1991)中描述的技术从噬菌体抗体文库中分离单克隆抗体。可以从任何适合的来源获得单克隆抗体。因此,例如,可以从鼠脾脏b细胞制备的杂交瘤获得单克隆抗体,所述鼠脾脏b细胞获自用感兴趣的抗原(例如呈在表面上表达抗原的细胞的形式)或编码感兴趣的抗原的核酸免疫的小鼠。还可以从衍生自免疫的人或非人哺乳动物(比如大鼠、狗或灵长类动物)的抗体表达细胞的杂交瘤获得单克隆抗体。
可以使用本领域熟知的标准重组dna技术进行进一步的修饰,例如如上所述的氨基酸取代、缺失或插入。
在一个实施方案中,本发明抗体是人抗体。可使用携带人免疫系统而非小鼠系统的部分的转基因或转染色体小鼠来生成定向的人单克隆抗体。这样的转基因和转染色体小鼠包括在本文中分别称为humab小鼠和km小鼠的小鼠,且在本文中统称为“转基因小鼠”。
humab小鼠含有编码未重排的人重链(μ和γ)及κ轻链免疫球蛋白序列的人免疫球蛋白基因迷你基因座以及使内源性μ和κ链基因座失活的靶向突变(lonberg,n.等人,nature368,856-859(1994))。因此,小鼠响应于免疫展现的小鼠igm或κ表达降低,并且,引入的人重链和轻链转基因经历类别转换和体细胞突变而生成高亲和力人iggκ单克隆抗体(lonberg,n.等人,(1994),上文,综述于lonberg,n.handbookofexperimentalpharmacology113,49-101(1994),lonberg,n.和huszar,d.,intern.rev.immunol.vol.1365-93(1995)以及harding,f.和lonberg,n.ann.n.y.acad.sci764536-546(1995))。humab小鼠的制备详细描述于taylor,l.等人,nucleicacidsresearch20,6287-6295(1992),chen,j.等人,internationalimmunology5,647-656(1993),tuaillon等人,j.immunol.152,2912-2920(1994),taylor,l.等人,internationalimmunology6,579-591(1994),fishwild,d.等人,naturebiotechnology14,845-851(1996)。还参见美国专利no.5,545,806、美国专利no.5,569,825、美国专利no.5,625,126、美国专利no.5,633,425、美国专利no.5,789,650、美国专利no.5,877,397、美国专利no.5,661,016、美国专利no.5,814,318、美国专利no.5,874,299、美国专利no.5,770,429、美国专利no.5,545,807、wo98/24884、wo94/25585、wo93/1227、wo92/22645、wo92/03918和wo01/09187。
hco7小鼠在其内源轻链(κ)基因中具有jkd破坏(如描述于chen等人,emboj.12,821-830(1993))、在其内源性重链基因中具有cmd破坏(如在wo01/14424的实施例1中所述)、且具有kco5人κ轻链转基因(如描述于fishwild等人,naturebiotechnology14,845-851(1996))和hco7人重链转基因(如美国专利no.5,770,429中所述)。
hco12小鼠在其内源轻链(κ)基因中具有jkd破坏(如描述于chen等人,emboj.12,821-830(1993))、在其内源性重链基因中具有cmd破坏(如在wo01/14424的实施例1中所述)、还具有kco5人κ轻链转基因(如描述于fishwild等人,naturebiotechnology14,845-851(1996))和hco12人重链转基因(如wo01/14424的实施例2中所述)。
在km小鼠品系中,内源性小鼠κ轻链基因已经如chen等人在emboj.12,811-820(1993)中所述的那样被纯合破坏,并且内源性小鼠重链基因已经如wo01/09187的实施例1中所述的那样被纯合破坏。这个小鼠品系携带人κ轻链转基因kco5,如fishwild等人在naturebiotechnology14,845-851(1996)中所述。这个小鼠品系还携带由染色体14片段hcf(sc20)组成的人重链转染色体,如wo02/43478中所述。
根据熟知的技术,可以利用来自这些转基因小鼠的脾细胞生成分泌人单克隆抗体的杂交瘤。这样的转基因非人动物、包含编码在本发明中使用的抗体的表达的可操作核酸序列的非人动物、用一个或多个靶编码核酸序列稳定转染的非人动物等等,是本发明另外的特征。
在本发明中使用的人单克隆或多克隆抗体、或在本发明中使用的源自其他物种的抗体也可以通过转基因手段产生:生成另一种用感兴趣的免疫球蛋白重链和轻链序列转基因的非人哺乳动物或植物,并且以可回收的形式从其产生抗体。关于在哺乳动物中的转基因产生,可以在山羊、奶牛或其他哺乳动物的乳汁中产生抗体并从中回收抗体。参见例如美国专利no.5,827,690、美国专利no.5,756,687、美国专利no.5,750,172和美国专利no.5,741,957。
此外,要在本发明中使用的人抗体或其他抗体可以使用本领域熟知的技术通过展示型技术来生成,所述展示型技术包括但不限于噬菌体展示、逆转录病毒展示、核糖体展示和其他技术,得到的分子可以经历另外的成熟过程,比如亲和力成熟,因为这样的技术是本领域熟知的(参见例如hoogenboom等人,j.mol.biol.227,381(1991)(噬菌体展示),vaughan等人,naturebiotech14,309(1996)(噬菌体展示),hanes和pluckthun,pnasusa94,4937-4942(1997)(核糖体展示),parmley和smith,gene73,305-318(1988)(噬菌体展示),scotttibs17,241-245(1992),cwirla等人,pnasusa87,6378-6382(1990),russel等人,nucl.acidsresearch21,1081-1085(1993),hoogenboom等人,immunol.reviews130,43-68(1992),chiswell和mccaffertytibtech10,80-84(1992),以及美国专利no.5,733,743)。如果利用展示技术产生非人抗体,则可以将这样的抗体人源化。
在一个另外的方面,本发明涉及产生本发明的修饰的抗体或融合蛋白的方法,所述方法包括在宿主细胞中表达编码所述修饰的抗体或融合蛋白的核酸构建体,并且任选地纯化所述修饰的抗体或融合蛋白。
在进一步的实施方案中,本发明的修饰的抗体或融合蛋白与选自下组的化合物连接:细胞毒剂;放射性同位素;前药或药物,如紫杉烷;细胞因子;和趋化因子。将这样的化合物与抗体连接(缀合)的方法是本领域熟知的。
4.药物组合物和治疗方法
在一个另外的方面,本发明涉及包含如上文定义的修饰的抗体或融合蛋白的药物组合物。可以用药学上可接受的载体或稀释剂以及任何其他已知的佐剂和赋形剂根据常规技术配制所述药物组合物,所述常规技术比如在remington:thescienceandpracticeofpharmacy,19thedition,gennaro,ed.,mackpublishingco.,easton,pa.,1995中公开的技术。
药学上可接受的载体或稀释剂以及任何其他已知的佐剂和赋形剂应该适合于所选择的本发明化合物和所选择的给药方式。载体和药物组合物的其他组分的适合性是基于对本发明所选择的化合物或药物组合物的期望生物学性质没有显著的负面影响(例如,不足以对抗原结合有实质性影响(10%或更低的相对抑制、5%或更低的相对抑制等等))而确定的。
本发明药物组合物还可包括稀释剂、填充剂、盐、缓冲剂、洗涤剂(例如,非离子洗涤剂,比如吐温80)、稳定剂、稳定剂(例如,糖或无蛋白质的氨基酸)、防腐剂、组织固定剂、增溶剂和/或适合包含在药物组合物中的其他物质。
本发明药物组合物中活性成分的实际剂量水平可以变化,以获得针对特定的患者、组合物和给药方式可有效地实现期望的治疗反应、而对患者无毒的活性成分的量。选定的剂量水平取决于多种药代动力学因素,包括所用本发明特定组合物或其酯、盐或酰胺的活性,给药途径,给药时间,所用特定化合物的排泄速率,治疗的持续时间,与所用特定组合物联合使用的其他药物、化合物和/或材料,被治疗患者的年龄、性别、体重、状况、一般健康和之前的医疗史,以及医学领域众所周知的类似因素。
具有本领域普通技能的医师或兽医可以容易地确定和处方药物组合物所需的有效量。例如,医师或兽医可以以低于为了实现期望的疗效所需的水平开始在药物组合物中使用的本发明化合物的剂量,并逐渐增加剂量直到实现期望的效果时为止。一般而言,本发明组合物的适合日剂量是化合物的这样的量,其为有效产生疗效的最低剂量。这样的有效剂量通常取决于上文描述的因素。任选地,给药是静脉内、肌内、腹膜内、通过吸入或皮下给药。治疗组合物的有效日剂量可以任选地以单位剂型在一天中以适当的间隔按照两个、三个、四个、五个、六个或更多个亚剂量分开施用。
在一个实施方案中,本发明的药物组合物是通过肠胃外施用的。如本文中使用的,短语“肠胃外施用”或“通过肠胃外施用”表示肠内和局部施用之外的施用方式,通常通过注射;包括表皮、静脉内、肌内、动脉内、鞘内、囊内、眶内、心内、皮内、腹膜内、腱内、经气管、皮下、表皮下、关节内、囊下、蛛网膜下、脊柱内、颅内、胸内、硬膜外和胸骨内注射和输注。
本发明的修饰的抗体或融合蛋白可用于治疗和/或预防许多疾病,并且可针对选自多种适合的靶分子的任何抗原。在本发明的一个实施方案中,修饰的抗体或融合蛋白结合选自下组的抗原:促红细胞生成素、β-淀粉样蛋白、tau、血小板生成素、干扰素-α(2a和2b)、干扰素-β(1b)、干扰素-γ、tnfri(cd120a)、tnfrii(cd120b)、cd137、il-1r1型(cd121a)、il-1r2型(cd121b)、il-2、il2r(cd25)、il-2r-β(cd123)、il-3、il-4、il-3r(cd123)、il-4r(cd124)、il-5r(cd125)、il-6r-α(cd126)、il-6r-β(cd130)、il-8、il-10、il-11、il-15、il-15bp、il-15r、il-20、il-21、tcr可变链、rank、rank-l、ctla4、cxcr4、cxcr4r、ccr5r、tgf-β1、tgf-β2、tgf-β3、g-csf、gm-csf、mif-r(cd74)、m-csf-r(cd115)、gm-csfr(cd116)、可溶性fcri、sfcrii、sfcriii、fcrn、因子vii、因子viii、因子ix、vegf、α-4整合素、cd11a、cd18、cd20、cd38、cd25、cd74、fcαri、fcεri、乙酰胆碱受体、fas、fasl、trail、肝炎病毒、丙型肝炎病毒、丙型肝炎病毒包膜e2、组织因子、组织因子和因子vii的复合物、egfr、cd4、cd28、vla-1,2,3或4、lfa-1、mac-1、1-选择素、psgl-1、icam-1、p-选择素、骨膜蛋白、cd33(唾液酸结合性免疫球蛋白样凝集素3)、唾液酸结合性免疫球蛋白样凝集素8、tnf、ccl1、ccl2、ccl3、ccl4、ccl5、ccl11、ccl13、ccl17、ccl18、ccl20、ccl22、ccl26、ccl27、cx3cl1、egf、vegf、tgfα、hgf、pdgf、ngf、cd32b、cd200、cd200r、杀伤细胞免疫球蛋白样受体(kir)、nkg2d、白细胞相关免疫球蛋白样受体(lair)、iy49、pd-l1、pd-l2、cd26、bst-2、ml-iap(黑色素瘤凋亡蛋白抑制剂)、组织蛋白酶d、cd40、cd40r、cd86、b细胞受体、cd79、pd-1、lag3和t细胞受体。
在某些特定的方面,修饰的igg4抗体或融合蛋白与选自cd137、cxcr4、etau、csf1r、lag3、pd1、pdl1或kir的靶分子结合。
在另一个特定的实施方案中,本发明的修饰的抗体或融合蛋白用于治疗恶性疾病和/或转移性疾病,比如黑色素瘤、卵巢癌、子宫内膜癌、nsclc、胶质母细胞瘤、脑相关肿瘤、乳腺癌、oscc、结肠癌、肾癌(renalcancer)、胰腺癌、hnscc、肾脏癌(kidneycancer)、胸腺瘤、肺癌、皮肤癌、喉癌、肝癌、腮腺肿瘤、胃癌、食道癌、前列腺癌、膀胱癌、睾丸的癌症、白血病和淋巴瘤。
在一个特定的实施方案中,本发明的修饰的抗体或融合蛋白用于治疗炎性和自身免疫性疾病,比如类风湿性关节炎、多发性硬化、炎性肠病、哮喘、1型糖尿病、sle、银屑病、特应性皮炎、移植物抗宿主病(gvhd)和copd。
通过下列实施例进一步展示了本发明,这些实施例不应该解释为进一步的限制。所有附图和所有参考文献的内容、在本申请各处引证的专利和公开的专利申请都通过提述明确并入本文。
实施例
实施例1
为改进生物分析和生物加工特性的治疗性igg4的设计
i.简介
对治疗性igg4抗体的铰链区加以工程改造使之具有cys-pro-pro-cys(cppc)基序以防止fab臂交换。[10,15,16]然而,治疗性igg4的核心-铰链区中的丝氨酸到脯氨酸取代对开发稳健的化学、制造和控制(cmc)造成了难题。对于具有cppc修饰的所有治疗性igg4抗体,在阳离子交换色谱(cex)上都观察到双峰洗脱行为,这对为了稳健制造的生物分析方法开发和生物加工开发造成了障碍。在这里,为了通过结合蛋白质工程和诱变来改进生物分析和生物加工特性,我们提出了在ch1结构域或铰链区中具有单点突变的下一代治疗性igg4抗体的设计。分子建模和模拟表明,即使对于igg4中的单点突变,也可能发生显著的二级结构和/或铰链区柔性变化,这可能导致意外的cmc行为。因此,在设计治疗性igg4抗体以及其他igg同种型时,应当仔细考虑cmc行为。
免疫球蛋白g(igg)是治疗性抗体开发最常用的骨架,并且细分为四个亚类:igg1、igg2、igg3和igg4。在igg同种型中,由于igg4不能激活经典补体途径,对于在不希望募集免疫效应子功能的情况下的应用,igg4是优选的亚类[1]。igg4与其他igg同种型表现不同之处在于其不能交联血浆中的两个相同的抗原,这也被称为功能性单价[2]。体内研究揭示,igg4通过将重链和附接的轻链(半分子)与来自另一个分子的重轻链对交换(称为fab臂交换),能够[3]交联两个不同的抗原而成为双特异性抗体[4]。由于fab臂组合的不可预测性,fab臂交换可能是igg4人类免疫疗法的一个主要缺陷。治疗性igg4和内源性人igg4之间的体内交换可产生具有未知的、不期望的特异性的双特异性抗体。功能性单价双特异性抗体的形成可改变治疗性igg4交联最初靶向抗原的能力,这可能影响人类免疫疗法的药代动力学和功效以及药效学[5]。
目前认为fab臂交换归因于igg4的两个独特结构特征:核心-铰链区中的cys-pro-ser-cys(cpsc)基序,以及ch3结构域中的r409氨基酸[4,6]。r409和k409分别是igg4和igg1的两个ch3结构域之间的主要界面。与具有k409残基的igg1中ch3结构域之间强的非共价相互作用不同,igg4的ch3-ch3相互作用弱得多,因为r409阻止链间氢键网络的正确形成[7],由此容许解离并使igg4具有fab臂交换的倾向[8]。igg4的ch3结构域中r409k的突变有助于使抗体稳定。核心-铰链区中的cppc和cpsc基序是igg1和igg4之间的另一个主要差异。igg1中的cppc基序的脯氨酸所诱导的相对刚性可妨碍链内二硫键的形成[9]。与之不同,igg4中的cpsc基序允许更好的铰链柔性,并使得形成链内二硫键成为可能。结果,如非还原sds-page证明的,形成了非共价缔合的半分子[9,10],这样的半分子然后将与其他半分子相互作用以形成双特异性抗体。为了解决这个问题,引入丝氨酸到脯氨酸的取代使铰链区与igg1中的更相似[10]。具有cppc基序的修饰的igg4铰链更稳定并且避免在体内与内源性人igg4进行fab臂交换。纳武单抗和派姆单抗,两种最近由fda批准的抗pd-1(程序性死亡蛋白-1)igg4癌症治疗剂,都含有具有cppc基序的修饰的铰链[11-13]。此外,许多在铰链区中具有丝氨酸到脯氨酸取代的其他igg4抗体目前正在进行临床前或临床试验[14]。逐渐得到公认的是,单纯的丝氨酸到脯氨酸突变(cppc)可以阻止体内fab臂交换,而不会增加治疗性igg4的任何风险或挑战。然而,这样的突变可能引起不希望的生物分析和生物加工行为。因此,仍然需要设计具有改进的生物分析和生物加工特性的治疗性igg4分子。
ii.材料与方法
1.cex-hplc方法
分析性cex-hplc使用thermofisherscientific(wilmington,de,usa)生产的propacscx-10lccolumns柱,安装waterscorporation(milford,ma,usa)生产的watershplc系统上进行。所述方法使用20mmmes(ph5.0)至20mmmes、1m或0.6mnacl(ph5.0)的盐梯度洗脱(0-60min),其中恒定的总注入蛋白质量为10μg。通过uv280nm监测洗脱的蛋白质。
2.用于分析聚集物的sec方法
分析性sec使用tosohbioscience(kingofprussia,pa,usa)生产的tskgelg3000swxl柱,安装在waterscorporation(milford,ma,usa)生产的watershplc系统上进行。所述方法使用100mm磷酸钠、100mm硫酸钠(ph6.8),流速为1ml/min,恒定的总注入蛋白质量为100μg。通过uv280nm监测洗脱的蛋白质。
3.色谱仪器和方法
使用安装有unicorn软件版本6.3(piscataway,ny,usa)的gehealthcare
4.ice280方法
使用ice280分析仪(convergentbioscience,toronto,canada)进行成像毛细管等电聚焦(icief)来检查电荷变体。分离盒与毛细管一起购自convergentbioscience。将这种毛细管固定在玻璃基板上,并通过两片中空纤维膜与阴极电解液和阳极电解液分离。通过将2mg/mligg与不同pi的标志物、1%甲基纤维素溶液和pharmalyte3-10混合来制备igg样品,然后用去离子水稀释。将混合物离心并在工作站中将上清液聚焦持续不同的时间长度以达到最佳分辨率。通过280nm氘灯检测器捕获ief痕迹的最终图像。
5.使用胃蛋白酶消化制备抗cd137(s228p)的f(ab’)2结构域
琼脂糖固定化的胃蛋白酶购自thermoscientific(waltham,ma,usa)。将抗cd137(s228p)用胃蛋白酶-琼脂糖(每20mg抗cd137(s228p)0.25ml胃蛋白酶-琼脂糖)在ph4.5在37℃水浴中消化4小时。将消化的产物调节至ph7.0,然后通过流过在ph7.0预平衡的mabselectsure柱纯化。在10mmtris·hcl洗涤过程中洗掉小的fc片段,并在25mm磷酸盐缓冲液洗涤过程中收集f(ab’)2结构域,未消化的mab和fc结构域保留在mabselectsure柱上。
6.使用木瓜蛋白酶消化制备抗cd137(s228p)的f(ab)结构域
琼脂糖固定化的木瓜蛋白酶购自thermoscientific(waltham,ma,usa)。在ph7将抗cd137(s228p)用在20mm磷酸钠、20mm半胱氨酸·hcl、10mmedta中的木瓜蛋白酶-琼脂糖在ph7.0在37℃水浴中消化4小时(每10mg抗cd137(s228p)0.25ml木瓜蛋白酶琼脂糖)。使用固定化的蛋白a柱将fab片段与未消化的mab和fc片段分离。
7.细胞系和培养条件
将expicho-s细胞(thermofisherscientific)在确定的无血清expicho表达培养基(thermofisherscientific)中培养,并且每三天在摇瓶中传代一次。将细胞在回转式振荡器平台上130rpm下在具有8%co2潮湿气氛的37℃培养箱中温育。
8.igg4-抗cd137突变体的构建
对ch1和铰链区中的单个氨基酸取代进行定点诱变。在诱变反应之前将igg4-抗cd137质粒的模板dna甲基化,其中使用两个重叠引物,并且其中一个含有靶突变。将诱变产物转化到大肠杆菌感受态细胞中,在该细胞中未甲基化的线性突变dna被环化和复制。用purelinkhipureplasmidmaxiprep试剂盒(thermofisherscientific)分离和纯化质粒dna。
9.抗cd137及其变体在expicho-s悬浮细胞中的瞬时表达
在转染前一天,将细胞以3.0百万个细胞/ml接种,并且在37℃、8%co2下在以130rpm旋转的回转式振荡器平台上搅拌。在转染当天(第0天),将细胞稀释至6.0百万个细胞/ml,加入250ml摇瓶中至50ml培养体积。将五十微克质粒dna稀释到opti-prosfm(thermofisherscientific)中,至总体积为2.0ml,混合。在另一个管中,将160μl的expifectaminecho试剂(thermofisherscientific)也用opti-prosfm稀释至总体积为2.0ml。将稀释的dna溶液和稀释的转染试剂轻轻混合,并且在室温下温育5分钟。然后将dna转染试剂混合物缓慢加入250ml烧瓶中的新稀释细胞中。将转染的细胞在37℃、8%co2下在以130rpm旋转的回旋式振荡器平台上温育。在第1天,向培养物供应10ml的expicho补充剂(feed)(thermofisherscientific)和300μl的expicho增效剂(enhancer)(thermofisherscientific);培养温度和co2分别改变为32℃和5%。培养物在第6天接受第二次补充剂推注,在第13天收获。抗cd137及其突变体都在expicho-s瞬时表达系统中稳健表达,表明靶区域中的氨基酸置换不影响igg4合成。
10.抗cd137结合测定
用碳酸氢盐缓冲液中的100μl(2μg/ml)的人cd137鼠iggb2将测定板在室温下包被1小时。然后用300μl的seablock溶液(pierce#37527)将板在室温下封闭1小时。将参考物质和样品在teknova测定稀释缓冲液(teknova-#d5120)中稀释至1μg/ml,然后进行2倍连续稀释。
倾出封闭缓冲液,用300μl的洗涤缓冲液(pbs-t)洗板三次。将样品和参考标准品(100μl)加入板中并在室温下温育1小时。洗涤3次后,加入100μl的第二抗体(抗人igg4fc,hrp;teknova稀释剂中1:1000),在室温下温育1小时。
洗涤后,加入100μl的dakotmb温育10分钟。用100μl的1n硫酸终止反应。使用softmax软件在m5酶标仪(moleculardevices)上使吸光度重新适应450nm,以650nm作为背景校正。根据相对于样品的对数浓度的吸光度值以及在每个稀释度的参考值计算ec50值。通过将参考物质ec50除以测试样品ec50的百分比来计算相对结合效力百分比。
11.完整质量(intactmass)分析
使用与四极杆飞行时间(q-tof)质谱仪(waters,milford,ma)偶联的acquityuhplc进行lc-ms。q-tof的毛细管电压设定为3000v,样品锥孔电压设定为80v。分析方法与前述相同。[17]简言之,将纯化样品与5mmdtt在37℃温育20分钟进行还原,通过加入甲酸至终浓度0.2%(v:v)终止。将还原后的样品注射0.5μg到用含有0.1%甲酸的20%乙腈平衡的反相(rp)柱(10μm,2.1x100mm
fab臂交换条件:将mab12突变与hca195混合,并与脱气的pbs(ph7.2)中的0.5mm还原型谷胱甘肽(gsh;sigma)一起温育。mab12突变和hca195两者的终浓度均为20μg/mm。将混合物(70μl)在37℃温育24小时。然后在完整质量测试之前将样品储存在4℃[4]。
iii.结果和讨论
我们在此报告,铰链区中的丝氨酸到脯氨酸(cppc)突变对各种igg4cmc开发造成了巨大挑战(参见表2)。
表2.抗体及其同种型列表
对于所有具有cppc基序修饰突变的治疗性igg4,在分析型cex柱上均观察到双峰洗脱行为(图1b),而对于所有igg1和野生型igg4(mab7-w和mab12-2)均观察到单峰洗脱行为(图1a,c)。其他研究先前也报道了在cex柱上的双峰洗脱行为,并且将其归因于特定分子[18]或特定树脂[19]。luo等人[18]报道了一种在互补决定区(cdr)的一级结构中具有被4个疏水残基包围的组氨酸残基的igg4在cex上的双峰洗脱行为,并且将其与cdr中的基于组氨酸质子化的电荷变体的分离关联起来。然而,我们的抗体序列分析表明,mab8、mab9、mab11和mab12在cdr中并不含相似的组氨酸残基,但显示出双峰洗脱行为;而mab4和mab6确实在cdr中含有相似的组氨酸残基,但显示出单峰洗脱行为。guo等人[19]报道了单克隆抗体在porosxs树脂上的表面诱导的三峰洗脱行为,并得出结论,porosxs树脂的双模态孔径分布诱导了蛋白质的两个结合位点,因而是导致两个峰的行为的原因。然而,在我们研究中使用的所有突变的igg4分子在无孔的树脂propacscx-10树脂上也观察到双峰洗脱行为。突变的igg4与野生型igg4之间的唯一区别是铰链区中的cppc和cpsc基序。显然,铰链区中的丝氨酸到脯氨酸(cppc)突变导致了cex柱上的这些双峰洗脱行为。
分离上述两个峰,并将峰级分注入到尺寸排阻色谱(sec)柱上。两个sec色谱图(图2)重叠(overlay)非常好,纯度超过99%,表明两个峰没有大小差异或聚集物形成。对于峰1和峰2观察到相似的电荷分布图(图3),表明这两个峰没有重大的电荷差异。还在相同条件下将双峰级分各自重新注入到cex柱上。重新注入两个级分产生与原始突变的igg4完全相同的双峰洗脱行为(图4),表明突变的igg4可能在树脂上存在两种不同的结合构象,而且两个群体在洗脱之后可互换。当改变cex操作ph时,双峰比率改变。增加ph导致第一峰的百分比更高,而降低ph导致第二峰的百分比更高(图5)。作为一个例子,突变igg4的双峰洗脱行为对用于电荷变异分析的分析型cex-hplc方法的开发造成了难题。双峰洗脱使得难以量化电荷变体谱,因为两个峰没有基于电荷差异分离,而且双峰的比率可能随着改变操作条件(例如,ph)而改变。此外,联合治疗近来已成为最有前景的免疫刺激性癌症理论,开发用于联合电荷变体分析的稳健分析方法成为了迫切需求。igg4(丝氨酸到脯氨酸取代(cppc))的双峰洗脱行为导致cex-hplc上的洗脱谱宽,这通常会导致与组合分子的色谱图产生干扰,使得不可能表征组合药物电荷变体(图6)。在制备型cex树脂上也观察到双峰洗脱行为(图7),使得难以开发稳健的商用cex纯化方法。在这里使用mab10作为例子。由于在盐梯度洗脱过程中观察到双峰洗脱行为,因此在该步骤洗脱过程中可以使用不同的电导率条件。如图8所示,较低的洗脱电导率得到的洗脱cv宽(图8a),而较高的电导率得到的洗脱cv小(图8b)。然而,越高的电导率导致洗脱池中杂质水平和hmw水平越高,如表3所示。洗脱条件是基于ph5的盐梯度洗脱选择的。使用了两种不同的洗脱条件,对应于盐梯度洗脱过程中的两个峰电导率。
表3.双峰洗脱行为对商用cex纯化过程开发的影响.
为了从结构角度更好地了解igg4(丝氨酸到脯氨酸取代(cppc))的双峰洗脱行为,进行了mab片段的酶促生成。使用固定化的胃蛋白酶和固定化的木瓜蛋白酶来生成mab11的f(ab')2和f(ab)片段(在方法部分中详述)。f(ab')2与f(ab)的不同之处在于前者是两个f(ab)与igg4铰链的连接体。sec-hplc谱(图9)提示,分别使用胃蛋白酶和木瓜蛋白酶成功生成了高纯度的f(ab')2和f(ab)。首先在与全mab11相同的条件下将mab11f(ab’)2注射到cex-hplc柱上。如图10所示,mab11f(ab')2片段也观察到两个峰洗脱行为,这表明f(ab')2,而不是fc尾,可能是双峰洗脱行为的原因。还在cex-hplc柱上测试了具有igg4(丝氨酸到脯氨酸取代(cppc))fc尾的fc融合蛋白mab13。mab13在cex柱上的单峰洗脱行为(图10)进一步证实fc尾不涉及双峰洗脱行为。为了进一步缩小可能造成双峰洗脱行为的原因的igg4片段的范围,还将f(ab)注射到cex-hplc柱上。有趣的是,对于f(ab),观察到单峰洗脱行为,而不是双峰洗脱行为。该结果提示,igg4(丝氨酸到脯氨酸(cppc))铰链在双峰洗脱行为中扮演重要角色。因此得出结论,igg4(丝氨酸到脯氨酸取代(cppc))f(ab')2的存在是造成双峰洗脱行为的原因。
为了可能针对改进的生物分析和生物加工特性设计下一代治疗性igg4,对当前igg4(丝氨酸到脯氨酸取代(cppc))序列的单点诱变进行了探索。首先,设计野生型igg4的丝氨酸到脯氨酸(cppc)突变,以便模拟具有cppc基序的igg1铰链来防止fab臂交换,但这种突变也在某些溶液条件下在cex柱上产生了不希望的双峰洗脱行为。希望的是,对现有igg4(丝氨酸到脯氨酸取代(cppc))的另一个单点突变将阻止fab臂交换并改善生物分析和生物加工特性。尝试了一种与铰链丝氨酸到脯氨酸(cppc)突变相似的途径,通过将igg4(丝氨酸到脯氨酸取代(cppc))中的单个氨基酸突变为igg1中的相应氨基酸来实现这样的特性。由于igg4(丝氨酸到脯氨酸取代(cppc))f(ab')2已经被鉴定为造成双峰洗脱行为的原因,因此在序列比对过程中不考虑fc区。图1中的所有igg1和igg4具有相同的κ轻链,因此轻链序列未予比对。由于每种抗体具有其自身独特的可变重链(vh)区,igg4的vh区不可能是造成双峰洗脱行为的原因,也被排除在序列比对之外。如此,在igg4(丝氨酸到脯氨酸取代(cppc))与igg1之间就恒定重链1(ch1)和铰链区进行了氨基酸序列比对,如图11所示。在igg4(丝氨酸到脯氨酸取代(cppc))和igg1之间鉴定出总共12个氨基酸差异,用绿色和黄色突出显示。igg4中第一个被鉴定出的半胱氨酸氨基酸形成重链和轻链之间的链间二硫键,因此不将半胱氨酸突变为相应的丝氨酸,以便保持igg4的完整结构。将igg4(丝氨酸到脯氨酸取代(cppc))中其余11个氨基酸各自突变为igg1中的相应氨基酸,以评估它们对cex-hplc柱行为的影响。
治疗性mab12突变体的cex-hplc色谱图显示在图12中。mab12-4c、mab12-7、mab12-9、mab12-10、mab12-11和mab12-w观察到了单峰洗脱行为。完整质量分析证实了这些突变体的成功突变(图13)。mab12-w是在铰链区中没有丝氨酸到脯氨酸的突变的野生型mab12;mab12-w可以与另一种野生型igg(hca195)交换fab臂以形成双特异性,如完整质量分析所证实的(图14)。将丝氨酸228突变为脯氨酸(mab12)以阻止fab臂交换(图15)。然而,这种突变在cex-hplc上产生了意外的双峰洗脱行为。通过附加赖氨酸196到谷氨酰胺(mab12-4c)、或丝氨酸217到脯氨酸(mab12-7)、或甘氨酸24到脯氨酸(mab12-9)、或脯氨酸224到组氨酸(mab12-10)、或脯氨酸225到苏氨酸(mab12-11)的单点突变,可以消除mab12的意外的双峰洗脱行为。完整质量分析也证实这些突变体不能与野生型igg交换fab臂,如图16-20所示。elisa结合测定(图21)表明,所有突变体都具有如同mab12的合理结合活性。突变没有改变mab12的结合活性。
参考文献
[1]y.m.lucisanovalim,p.j.lachmann,theeffectofantibodyisotypeandantigenicepitopedensityonthecomplement-fixingactivityofimmunecomplexes:asystematicstudyusingchimaericanti-nipantibodieswithhumanfcregions,clinicalandexperimentalimmunology,84(1991)1-8.
[2]j.s.vanderzee,p.vanswieten,r.c.aalberse,serologicaspectsofigg4antibodies.ii.igg4antibodiesformsmall,nonprecipitatingimmunecomplexesduetofunctionalmonovalency,thejournalofimmunology,137(1986)3566-3571.
[3]j.schuurman,r.vanree,g.j.perdok,h.r.vandoorn,k.y.tan,r.c.aalberse,normalhumanimmunoglobuling4isbispecific:ithastwodifferentantigen-combiningsites,immunology,97(1999)693-698.
[4]m.vanderneutkolfschoten,j.schuurman,m.losen,w.k.bleeker,p.martinez-martinez,e.vermeulen,t.h.denbleker,l.wiegman,t.vink,l.a.aarden,m.h.debaets,j.g.vandewinkel,r.c.aalberse,p.w.parren,anti-inflammatoryactivityofhumanigg4antibodiesbydynamicfabarmexchange,science(newyork,n.y.),317(2007)1554-1557.
[5]j.p.silva,o.vetterlein,j.jose,s.peters,h.kirby,thes229pmutationpreventsinvivoandinvitroigg4fab-armexchangeasdemonstratedusingacombinationofnovelquantitativeimmunoassaysandphysiologicalmatrixpreparation,jbiolchem,290(2015)5462-5469.
[6]t.rispens,p.ooijevaar-deheer,o.bende,r.c.aalberse,mechanismofimmunoglobuling4fab-armexchange,jamchemsoc,133(2011)10302-10311.
[7]a.m.davies,t.rispens,t.h.denbleker,j.m.mcdonnell,h.j.gould,r.c.aalberse,b.j.sutton,crystalstructureofthehumanigg4c(h)3dimerrevealstheroleofarg409inthemechanismoffab-armexchange,molimmunol,54(2013)1-7.
[8]a.m.davies,t.rispens,t.h.denbleker,j.m.mcdonnell,h.j.gould,r.c.aalberse,b.j.sutton,crystalstructureofthehumanigg4ch3dimerrevealstheroleofarg409inthemechanismoffab-armexchange,molecularimmunology,54(2013)1-7.
[9]j.w.bloom,m.s.madanat,d.marriott,t.wong,s.y.chan,intrachaindisulfidebondinthecorehingeregionofhumanigg4,proteinscience:apublicationoftheproteinsociety,6(1997)407-415.
[10]s.angal,d.j.king,m.w.bodmer,a.turner,a.d.g.lawson,g.roberts,b.pedley,j.r.adair,asingleaminoacidsubstitutionabolishestheheterogeneityofchimericmouse/human(igg4)antibody,molecularimmunology,30(1993)105-108.
[11]c.wang,k.b.thudium,m.han,x.t.wang,h.huang,d.feingersh,c.garcia,y.wu,m.kuhne,m.srinivasan,s.singh,s.wong,n.garner,h.leblanc,r.t.bunch,d.blanset,m.j.selby,a.j.korman,invitrocharacterizationoftheanti-pd-1antibodynivolumab,bms-936558,andinvivotoxicologyinnon-humanprimates,cancerimmunologyresearch,2(2014)846-856.
[12]a.m.davies,b.j.sutton,humanigg4:astructuralperspective,immunologicalreviews,268(2015)139-159.
[13]g.scapin,x.yang,w.w.prosise,m.mccoy,p.reichert,j.m.johnston,r.s.kashi,c.strickland,structureoffull-lengthhumananti-pd1therapeuticigg4antibodypembrolizumab,naturestructural&molecularbiology,22(2015)953-958.
[14]j.m.reichert,antibodiestowatchin2017,mabs,9(2017)167-181.
[15]j.g.salfeld,isotypeselectioninantibodyengineering,natbiotech,25(2007)1369-1372.
[16]a.f.labrijn,a.o.buijsse,e.t.vandenbremer,a.y.verwilligen,w.k.bleeker,s.j.thorpe,j.killestein,c.h.polman,r.c.aalberse,j.schuurman,j.g.vandewinkel,p.w.parren,therapeuticigg4antibodiesengageinfab-armexchangewithendogenoushumanigg4invivo,natbiotechnol,27(2009)767-771.
[17]j.fu,j.bongers,l.tao,d.huang,r.ludwig,y.huang,y.qian,j.basch,j.goldstein,r.krishnan,l.you,z.j.li,r.j.russell,characterizationandidentificationofalaninetoserinesequencevariantsinanigg4monoclonalantibodyproducedinmammaliancelllines,jchromatogrbanalyttechnolbiomedlifesci,908(2012)1-8.
[18]h.luo,m.cao,k.newell,c.afdahl,j.wang,w.k.wang,y.li,double-peakelutionprofileofamonoclonalantibodyincationexchangechromatographyiscausedbyhistidine-protonation-basedchargevariants,jchromatogra,1424(2015)92-101.
[19]j.guo,a.d.creasy,g.barker,g.carta,surfaceinducedthree-peakelutionbehaviorofamonoclonalantibodyduringcationexchangechromatography,jchromatogra,1474(2016)85-94.
序列表
抗cd137抗体野生型重链序列(seqidno:1):
qvqlqqwgagllkpsetlsltcavyggsfsgyywswirqspekglewigeinhggyvtynpslesrvtisvdtsknqfslklssvtaadtavyycardygpgnydwyfdlwgrgtlvtvssastkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcpscpapeflggpsvflfppkpkdtlmisrtpevtcvvvdvsqedpevqfnwyvdgvevhnaktkpreeqfnstyrvvsvltvlhqdwlngkeykckvsnkglpssiektiskakgqprepqvytlppsqeemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflysrltvdksrwqegnvfscsvmhealhnhytqkslslslgk
抗cd137野生型轻链序列(seqidno:2):
eivltqspatlslspgeratlscrasqsvssylawyqqkpgqaprlliydasnratgiparfsgsgsgtdftltisslepedfavyycqqrsnwppaltfgggtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec
mab1部分序列(seqidno:3):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcp
mab2部分序列(seqidno:4):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcp
mab3部分序列(seqidno:5):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcp
mab4部分序列(seqidno:6):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcp
mab5部分序列(seqidno:7):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcp
mab6部分序列(seqidno:8):
astkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcp
mab7部分序列(seqidno:9):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab8部分序列(seqidno:10):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab9部分序列(seqidno:11):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab10部分序列(seqidno:12):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab11部分序列(seqidno:13):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab12部分序列(seqidno:14):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab7-w部分序列(seqidno:15):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcpscp
mab12-w部分序列(seqidno:16):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcpscp
mab12-1部分序列(seqidno:17):
astkgpsvfplapcskstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab12-2部分序列(seqidno:18):
astkgpsvfplapcsrstsgstaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab12-3部分序列(seqidno:19):
astkgpsvfplapcsrstsegtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygppcppcp
mab12-4a部分序列(seqidno:20):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtytcnvdhkpsntkvdkrveskygppcppcp
mab12-4b部分序列(seqidno:21):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtrtytcnvdhkpsntkvdkrveskygppcppcp
mab12-4c部分序列(seqidno:22):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtptytcnvdhkpsntkvdkrveskygppcppcp
mab12-5部分序列(seqidno:23):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktyicnvdhkpsntkvdkrveskygppcppcp
mab12-6部分序列(seqidno:24):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvnhkpsntkvdkrveskygppcppcp
mab12-7部分序列(seqidno:25):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrvepkygppcppcp
mab12-8部分序列(seqidno:26):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskkgppcppcp
mab12-9部分序列(seqidno:27):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskytppcppcp
mab12-10部分序列(seqidno:28):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskyghpcppcp
mab12-11部分序列(seqidno:29):
astkgpsvfplapcsrstsestaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtktytcnvdhkpsntkvdkrveskygptcppcp
序列表
<110>百时美施贵宝公司
<120>用于改进生物分析和生物加工特性的治疗性免疫球蛋白g4的设计
<130>12883-wo-pct
<150>us62/438684
<151>2016-12-23
<160>29
<170>patentinversion3.5
<210>1
<211>448
<212>prt
<213>人工序列
<220>
<223>免疫球蛋白片段序列
<400>1
glnvalglnleuglnglntrpglyalaglyleuleulysproserglu
151015
thrleuserleuthrcysalavaltyrglyglyserpheserglytyr
202530
tyrtrpsertrpileargglnserproglulysglyleuglutrpile
354045
glygluileasnhisglyglytyrvalthrtyrasnproserleuglu
505560
serargvalthrileservalaspthrserlysasnglnpheserleu
65707580
lysleuserservalthralaalaaspthralavaltyrtyrcysala
859095
argasptyrglyproglyasntyrasptrptyrpheaspleutrpgly
100105110
argglythrleuvalthrvalserseralaserthrlysglyproser
115120125
valpheproleualaprocysserargserthrsergluserthrala
130135140
alaleuglycysleuvallysasptyrpheprogluprovalthrval
145150155160
sertrpasnserglyalaleuthrserglyvalhisthrpheproala
165170175
valleuglnserserglyleutyrserleuserservalvalthrval
180185190
proserserserleuglythrlysthrtyrthrcysasnvalasphis
195200205
lysproserasnthrlysvalasplysargvalgluserlystyrgly
210215220
proprocysprosercysproalaproglupheleuglyglyproser
225230235240
valpheleupheproprolysprolysaspthrleumetileserarg
245250255
thrprogluvalthrcysvalvalvalaspvalserglngluasppro
260265270
gluvalglnpheasntrptyrvalaspglyvalgluvalhisasnala
275280285
lysthrlysproargglugluglnpheasnserthrtyrargvalval
290295300
servalleuthrvalleuhisglnasptrpleuasnglylysglutyr
305310315320
lyscyslysvalserasnlysglyleuproserserileglulysthr
325330335
ileserlysalalysglyglnproarggluproglnvaltyrthrleu
340345350
proproserglngluglumetthrlysasnglnvalserleuthrcys
355360365
leuvallysglyphetyrproseraspilealavalglutrpgluser
370375380
asnglyglnprogluasnasntyrlysthrthrproprovalleuasp
385390395400
seraspglyserphepheleutyrserargleuthrvalasplysser
405410415
argtrpglngluglyasnvalphesercysservalmethisgluala
420425430
leuhisasnhistyrthrglnlysserleuserleuserleuglylys
435440445
<210>2
<211>216
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>2
gluilevalleuthrglnserproalathrleuserleuserprogly
151015
gluargalathrleusercysargalaserglnservalsersertyr
202530
leualatrptyrglnglnlysproglyglnalaproargleuleuile
354045
tyraspalaserasnargalathrglyileproalaargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglupro
65707580
gluaspphealavaltyrtyrcysglnglnargserasntrppropro
859095
alaleuthrpheglyglyglythrlysvalgluilelysargthrval
100105110
alaalaproservalpheilepheproproseraspgluglnleulys
115120125
serglythralaservalvalcysleuleuasnasnphetyrproarg
130135140
glualalysvalglntrplysvalaspasnalaleuglnserglyasn
145150155160
serglngluservalthrgluglnaspserlysaspserthrtyrser
165170175
leuserserthrleuthrleuserlysalaasptyrglulyshislys
180185190
valtyralacysgluvalthrhisglnglyleuserserprovalthr
195200205
lysserpheasnargglyglucys
210215
<210>3
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>3
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>4
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>4
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
lysvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>5
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>5
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>6
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>6
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>7
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>7
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>8
<211>113
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>8
alaserthrlysglyproservalpheproleualaproserserlys
151015
serthrserglyglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrilecysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluprolyssercysasplysthrhisthrcysproprocys
100105110
pro
<210>9
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>9
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>10
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>10
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>11
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>11
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>12
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>12
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>13
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>13
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>14
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>14
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>15
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>15
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysprosercyspro
100105110
<210>16
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>16
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysprosercyspro
100105110
<210>17
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>17
alaserthrlysglyproservalpheproleualaprocysserlys
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>18
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>18
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrserglyserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>19
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>19
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluglythralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>20
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>20
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrglnthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>21
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>21
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrargthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>22
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>22
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrprothr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>23
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>23
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrilecysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>24
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>24
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasnhislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyproprocysproprocyspro
100105110
<210>25
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>25
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluprolystyrglyproprocysproprocyspro
100105110
<210>26
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>26
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlyslysglyproprocysproprocyspro
100105110
<210>27
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>27
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrthrproprocysproprocyspro
100105110
<210>28
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>28
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyhisprocysproprocyspro
100105110
<210>29
<211>110
<212>prt
<213>人工
<220>
<223>免疫球蛋白片段序列
<400>29
alaserthrlysglyproservalpheproleualaprocysserarg
151015
serthrsergluserthralaalaleuglycysleuvallysasptyr
202530
pheprogluprovalthrvalsertrpasnserglyalaleuthrser
354045
glyvalhisthrpheproalavalleuglnserserglyleutyrser
505560
leuserservalvalthrvalproserserserleuglythrlysthr
65707580
tyrthrcysasnvalasphislysproserasnthrlysvalasplys
859095
argvalgluserlystyrglyprothrcysproprocyspro
100105110
- 还没有人留言评论。精彩留言会获得点赞!