一种玉米赤霉烯酮降解酶突变体及其编码基因和应用的制作方法
本发明属于生物技术领域,具体而言,涉及一种玉米赤霉烯酮降解酶、突变体及其编码基因和应用。
背景技术:
玉米赤霉烯酮首先是从玉米中分离出来的,是可以由许多镰孢属物种产生的一种非甾体雌激素霉菌毒素,作物在收获前后都会产生。玉米赤霉烯酮总是在包括玉米、大麦、小麦等许多作物和谷类副产品中被发现,尤其是在适合真菌生长的环境中。
玉米赤霉烯酮的衍生物有很多,例如玉米赤霉烯醇,它们会通过污染的作物进入食物链并积累在人体和动物体内,对生物造成损害。玉米赤霉烯酮及其衍生物的化学结构类似于天然雌激素,因此它们能够竞争性地结合雌激素受体,引起外部和内部生殖器改变和繁殖障碍,导致高雌性激素症和不孕症,此类毒素还会刺激乳腺癌细胞系的生长并在小鼠中致癌。
鉴于此类毒素的危害,玉米赤霉烯酮等在谷物、食品和饲料中的含量必须低于一定标准。由于玉米赤霉烯酮等是极端稳定的,使用传统的物理和化学方法去除此类毒素是低效的。为了解决这些问题,降低此类毒素污染的一个有希望的策略是酶降解。酶降解不仅可以高效地将毒素转化为无毒性产物,安全环保,而且酶催化反应专一性强、降解效率高,不会破坏谷物的营养物质。
迄今为止,已经有一些对于玉米赤霉烯酮降解酶的研究,得到了一些可以降解玉米赤霉烯酮毒素的酶,它们能够特异性地结合玉米赤霉烯酮并且降解它。但是,目前对筛选得到的微生物中与玉米赤霉烯酮降解相关的酶研究不多。
技术实现要素:
本发明的目的在于提供一种玉米赤霉烯酮降解酶、突变体及其编码基因,以及其在水解玉米赤霉烯酮及其衍生物中的应用。
为了实现本发明的目的,发明人通过大量试验研究并不懈努力,最终获得了如下技术方案:
一种玉米赤霉烯酮降解酶,该降解酶具有序列表中seqidno.1所示的氨基酸序列;或该降解酶是在seqidno.1所示的氨基酸序列基础上缺失、替换、插入或/和添加一个至几个氨基酸的保守性突变而获得的保守性变异体。所述的保守性变异体优选具有seqidno.3所示的氨基酸序列。
需要说明的是,本发明所提供的玉米赤霉烯酮降解酶或其突变体是一种内酯水解酶。seqidno.1或seqidno.3所示的氨基酸序列均由264个氨基酸残基组成。
为了使上述降解酶突变体蛋白质便于纯化,可在上述的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接上如表1所示的标签。
表1标签的序列
上述降解酶突变体蛋白质可以人工合成,也可先合成其编码基因,再进行生物表达得到。上述中的蛋白质的编码基因还可通过将seqidno.1所示的氨基酸序列中,缺失、置换、插入或添加一个至几个并保持原有酶活性,或者连上表1所示的标签的编码序列得到。
一种玉米赤霉烯酮降解酶的编码基因,该基因编码:
(a)具有seqidno.1所示的氨基酸序列的蛋白质;或
(b)具有衍生自缺失、置换、插入或/和添加一个至几个氨基酸的seqidno.1所示的氨基酸序列并具有玉米赤霉烯酮及其衍生物降解活性的蛋白质。
还需要说明的是,玉米赤霉烯酮及其衍生物降解活性是指可以切割底物的内酯键,随后产生具有开放侧链的二羟基苯基衍生物以及释放二氧化碳,作用于玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇这几种底物。
进一步,所述玉米赤霉烯酮降解酶的编码基因为(i)、(ii)或(iii)的dna分子:
(i)具有seqidno.2或seqidno.4所示的核苷酸序列的dna分子;
(ii)在严格条件下与(i)所述的核苷酸序列杂交且编码具有玉米赤霉烯酮及其几种衍生物降解活性的蛋白质的dna分子;
(iii)与(i)或(ii)所述的核苷酸序列具有90%以上同源性的核苷酸序列的dna分子。
seqidno.2所示的核苷酸序列由795个核苷酸组成。
进一步,所述严格条件为钠浓度为50-300mm的溶液中,反应温度为50-68℃。
例如:在进行分子杂交的过程中,可以为在6×ssc、质量分数为0.5%的sds的溶液中,在65℃下杂交,然后用2×ssc,质量分数为0.1%的sds和1×ssc、质量分数为0.1%的sds各洗膜一次。其中sds的中文名称为十二烷基硫酸钠,1×ssc包括0.15mol/lnacl和0.015mol/l柠檬酸;sds以及不同浓度倍数的ssc均为本领域的常用试剂。
含有上述任一所述编码基因的重组载体、表达盒、转基因细胞系或重组菌也属于本发明的保护范围。
本发明提供一种重组载体,该重组载体包含上述的玉米赤霉烯酮降解酶的编码基因。具体的,所述重组载体为将上述任一所述编码基因插入出发载体(例如:pet28a)的多克隆位点得到的重组表达载体。可用现有的表达载体构建含有所述基因的重组表达载体。使用所述基因构建重组表达载体时,在其转录起始核苷酸前可加上任何一种增强型启动子或组成型启动子,它们可单独使用或与其它的启动子结合使用;此外,使用本发明的基因构建重组表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。
本发明还提供一种转化体,该转化体包含上述的重组载体。转化体可以为重组菌,例如,将上述任一所述编码基因插入出发载体(例如:pet28a载体)的多克隆位点得到的重组表达载体转化至大肠杆菌bl21(de3),得到重组菌。
本发明还提供一种引物对,用于扩增上述的玉米赤霉烯酮降解酶的编码基因全长及其任意片段。例如:引物对的序列如seqidno.5和seqidno.6所示,或者如seqidno.7和seqidno.8所示。
上述任一所述蛋白质、上述任一所述编码基因、上述任一所述重组表达载体、所述表达盒、转基因细胞系或重组菌中的任意一种在降解玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇这几种底物的应用也属于本发明的保护范围。
在具体应用的过程中,可以采用下面的方法:以玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇这几种为底物,在偏碱性ph条件,利用玉米赤霉烯酮降解酶对玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇进行酶解。
所述的酶解条件包括:反应体系的温度20-55℃,优选为40℃,反应体系的ph值为6.0-11.0,优选为9.5。
本发明还提供了一种生产玉米赤霉烯酮降解酶的方法,该方法包括培养上述的转化体并由培养产物中收集玉米赤霉烯酮降解酶。收集的玉米赤霉烯酮降解酶可以进一步进行纯化。
需要说明的是,本发明提供的蛋白具有玉米赤霉烯酮及其几种衍生物降解活性,属于玉米赤霉烯酮降解酶。并且该蛋白与其他已表征的玉米赤霉烯酮降解酶的氨基酸序列相比,相似性不大于64%,属于一种全新的玉米赤霉烯酮降解酶,为人们降解玉米赤霉烯酮增加了一种新的选择。另外,本发明所提供的玉米赤霉烯酮降解酶最适天然底物为玉米赤霉烯酮,且具有在偏碱性ph条件下活性较高的特性,同时其在不同ph下的稳定性均较好。
与现有技术相比,本发明提供的玉米赤霉烯酮降解酶zhday3及其突变体具有显著的进步性,还主要体现在以下方面:
(1)文献报道,已经进行表征的玉米赤霉烯酮降解酶一个是zhd101,另外两种是zen-jjm和zlhy-6,zen-jjm和zlhy-6与zhd101的氨基酸同源性为99%和98%,性质基本一致;另外一个是zhd518。本发明中的zhday3与zhd101的氨基酸同源性为63%,与zhd518的氨基酸同源性为64%,确定是一种新型的玉米赤霉烯酮降解酶。另外,已经表征的玉米赤霉烯酮降解酶zhd101的最适反应温度是37℃、最适ph为9.5;玉米赤霉烯酮降解酶zhd518的最适反应温度是40℃、最适ph为8.0。本发明的zhday3的最适温度为40℃、最适ph为9.5。zhday3在温度为30-50℃的范围内依然具有60%以上的酶活,在ph为9.0-10.5的范围内具有60%以上的酶活。
(2)本发明提供的玉米赤霉烯酮降解酶zhday3对zen及其四种衍生物都具有降解活性,但降解能力有所差别。结果如下:将稀释酶液分别在相同浓度(反应体系中底物终浓度为20.0μg/ml)的不同底物条件下进行酶活测定。以玉米赤霉烯酮为底物测得酶活为参比(100%),以α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇为底物所测相对酶活分别为49.0%、44.2%、48.9%、32.7%。因此,该酶对玉米赤霉烯酮活性比较高,其他次之。
(3)本发明中提供的玉米赤霉烯酮降解酶zhday3在经过一个定点(n153h)突变后,对于底物α-玉米赤霉醇的底物特异性相对zhday3提高了2.1倍;对于β-玉米赤霉醇的底物特异性相对zhday3提高了1.4倍。这是一个全新的特性,之前报道的玉米赤霉烯酮降解酶zhd101的相应位置也进行突变后产生的突变酶zhd101(v153h)对α-玉米赤霉烯醇具有2.7倍的提高。两种突变体显示的底物特异性结果明显不同,这说明本发明中设计的突变具有唯一性、独特性,对于工业上的应用非常具有潜力。
附图说明
图1为玉米赤霉烯酮降解酶zhday3蛋白纯化前后的sds-page电泳图。
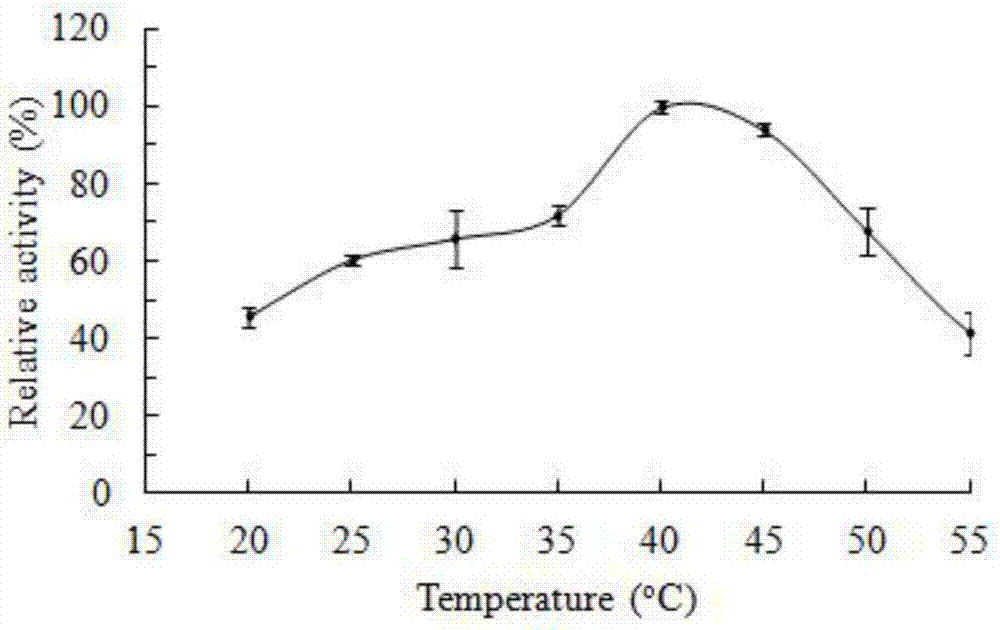
图2为玉米赤霉烯酮降解酶zhday3的活性随温度的变化的结果。
图3为玉米赤霉烯酮降解酶zhday3的活性随ph的变化的结果。
图4为玉米赤霉烯酮降解酶zhday3在不同温度下的活性变化结果。
图5为玉米赤霉烯酮降解酶zhday3在不同ph下的活性变化结果。
具体实施方式
以下结合附图对本发明的原理和特征进行描述,所举实例只用于解释本发明,并非用于限定本发明的范围。
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
实施例1、蛋白及基因的制备与纯化
1、基因序列的人工合成
seqidno.2所示的核苷酸序列委托武汉金开瑞生物工程有限公司按照本领域的常规技术进行基因人工合成,基因插入质粒载体puc57中,保存,备用。
2、基因序列的扩增
根据seqidno.2所示的核苷酸序列设计引物对如下:
正向引物:5′-cgcggatccatgcgcaccaggtccaatatcacc-3′,如seqidno.5所示;
反向引物:5′-ccgctcgagttacaagtactttcgagtcttttcc-3′,如seqidno.6所示;
正向引物的下划线部分为bamhi的酶切位点,反向引物的下划线部分为xhoi酶切位点。
pcr反应体系:
pcr反应条件:94℃预变性5min,然后94℃变性30s,55℃退火30s,72℃延伸1min,30个循环,最后72℃延伸10min。
pcr产物用质量分数为0.7%的琼脂糖凝胶电泳检测产量和特异性,并用dna纯化试剂盒(超薄离心柱型,天根公司生产)纯化。将纯化的pcr产物进行测序,结果表明pcr产物的序列包括seqidno.2所示1-795位,并将其命名为zhday3dna片段。
3、重组表达载体的构建
1)将上述测序正确的pcr产物用bamhi和xhoi双酶切,琼脂糖电泳回收酶切产物。
2)将质粒pet28a(cat.n069864-3,novogen)用bamhi和xhoi双酶切,琼脂糖电泳回收酶切产物。
3)将步骤1)的酶切产物和步骤2)的酶切产物进行连接,连接产物电击转化大肠杆菌dh5α后涂布于含有50μg/ml卡那霉素的lb平板,37℃过夜培养,将得到的转化子用上述的正向引物和反向引物进行菌落pcr,筛选到含有zhday3基因的重组菌,提取重组菌的质粒,进行测序验证,结果表明,在pet28a的bamhi和xhoi酶切位点之间插入了zhday3dna片段,该片段包括seqidno.2的自5′端起第1至795位的核苷酸,插入方向正确,将该重组质粒命名为pet28a-zhday3。
4、工程菌的制备
将质粒pet28a-zhday3电击转化大肠杆菌bl21(de3)(cat.n0cd601,全式金公司)后涂布于含有50μg/ml卡那霉素的lb平板,37℃过夜培养,得到含有质粒pet28a-zhday3的工程菌,记作bl21/pet28a-zhday3。
用pet28a代替pet28a-zhday3,转化大肠杆菌bl21(de3),步骤同上,得到含有pet28a的重组菌,作为对照菌。将转入bl21(de3)的阳性重组菌记作bl21/pet28a。
5、目标蛋白的表达和纯化
his60nisuperflowresin纯化柱购自takara公司,产品目录号为635660。
gehitrapdesalting纯化柱购自gehealthcare公司,产品目录号分别为17-1408-01。
将上述步骤4制备的阳性重组菌bl21/pet28a-zhday3培养于含有50μg/ml卡那霉素的lb培养基中,37℃培养3h;od600=0.7时,加入iptg至其在lb培养基中的终浓度0.8mm,转至18℃继续培养16h。
在3800rpm、15min条件下离心收集菌体,悬浮于缓冲溶液a(50mmglycine-naoh,ph9.5)中,于冰浴中超声破碎(60w,10min;超声1s,停止2s),之后12000rpm离心10min除去细胞碎片,取上清液;将上清液过his60nisuperflowresin纯化柱,用5ml超纯水冲洗,再用10ml溶液b(50mmglycine-naoh,ph9.5,25mm咪唑)漂洗,最后用5ml溶液c(50mmglycine-naoh,ph9.5,500mm咪唑)洗脱,收集洗脱液。然后将洗脱液用脱盐柱gehitrapdesalting进行脱盐处理,用溶液a(50mmglycine-naoh,ph9.5)进行洗脱,得到zhday3纯酶液。
将步骤4制备的对照菌采用相同的步骤进行培养和纯化,得到的溶液作为对照酶液。
sds-page电泳显示纯化的zhday3蛋白的分子量约为30kda,符合理论推断的29.3kda。结果如图1所示,图1中,泳道m表示蛋白分子量标准(250,150,100,75,50,37,25,15,10kda);泳道1表示大肠杆菌bl21/pet28a-zhday3破菌后的上清液;泳道2表示ni-nta柱纯化后的zhday3蛋白;泳道3表示gedesalting脱盐柱纯化后的zhday3蛋白。可以看出已经获得了zhday3蛋白。同时进行了对照组的实验,但对照菌并没有得到目的蛋白。
实施例2、以玉米赤霉烯酮为底物验证蛋白功能
酶活单位定义为1min内降解1μg底物玉米赤霉烯酮所需要的酶量作为一个酶活单位u。
(一)最适温度
用ph9.5的50mmglycine-naoh缓冲液稀释实施例1的步骤5中的zhday3纯酶液,用稀释后的酶液进行酶活测定。将稀释后的酶液记作稀释酶液。
溶液a组成:由50mm,ph9.5glycine-naoh缓冲液和玉米赤霉烯酮溶液组成;底物玉米赤霉烯酮在反应体系0.5ml中的终浓度为20.0μg/ml。
实验组:活性测定反应体系为0.5ml,由0.45ml溶液a和0.05ml稀释酶液;反应体系的ph值为9.5;反应体系在特定温度范围(20-55℃)内温育10min后,0.5ml色谱级甲醇终止反应,冷却后使用高效液相色谱仪(hplc)测定底物降解量。
结果如图2所示。图2表明,玉米赤霉烯酮降解酶具有降解玉米赤霉烯酮的活性。在40℃条件下,玉米赤霉烯酮降解酶具有最高的酶活性;将此温度下的酶活反应体系的底物玉米赤霉烯酮降解量作为相对活性100%,其他温度下酶活反应体系的底物玉米赤霉烯酮降解量与此最高酶活体系的底物玉米赤霉烯酮降解量的比值作为相对活性。在30-50℃条件下均具有60%以上的活性。
对照组:以对照菌bl21/pet28a获得的蛋白(记作对照酶液)进行上述实验,结果不管在哪个温度条件下,对照酶液都没有降解玉米赤霉烯酮的活性。
实验设3次重复,结果一致。
(二)最适ph
如下各组中的稀释酶液均是用各组中的缓冲液稀释实施例1的步骤5中的zhday3纯酶液得到的。
实验组:活性测定反应体系为0.5ml,分别由0.45ml溶液b(b1、b2、b3、b4、b5、b6、b7、b8、b9、b10、b11、b12、b13和b14)和0.05ml稀释酶液组成,底物玉米赤霉烯酮在反应体系0.5ml中的终浓度为20.0μg/ml。
溶液b1的组成:0.2mna2hpo4-柠檬酸缓冲液和底物玉米赤霉烯酮组成;溶液b1的ph值为5.5。
溶液b2的组成:与溶液b1的组成相同,不同的是溶液b2的ph值为6.0。
溶液b3的组成:与溶液b1的组成相同,不同的是溶液b3的ph值为6.5。
溶液b4的组成:与溶液b1的组成相同,不同的是溶液b4的ph值为7.0。
溶液b5的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmtris-hcl缓冲液。溶液b5的ph值为7.0。
溶液b6的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmtris-hcl缓冲液。溶液b6的ph值为7.5。
溶液b7的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmtris-hcl缓冲液。溶液b7的ph值为8.0。
溶液b8的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmtris-hcl缓冲液。溶液b8的ph值为8.5。
溶液b9的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmtris-hcl缓冲液。溶液b9的ph值为9.0。
溶液b10的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmglycine-naoh缓冲液。溶液b10的ph值为9.0。
溶液b11的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmglycine-naoh缓冲液。溶液b11的ph值为9.5。
溶液b12的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmglycine-naoh缓冲液。溶液b12的ph值为10.0。
溶液b13的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmglycine-naoh缓冲液。溶液b13的ph值为10.5。
溶液b14的组成:与溶液b1的组成相同,不同的是将0.2mna2hpo4-柠檬酸缓冲液替换为50mmglycine-naoh缓冲液。溶液b14的ph值为11.0
将反应体系在40℃温育10min后,加入0.5ml色谱级甲醇终止反应,冷却后使用高效液相色谱仪(hplc)测定底物降解量。
实验设三次重复。
结果如图3所示。
玉米赤霉烯酮降解酶突变体在ph为5.5至11.0之间的条件下均具有水解玉米赤霉烯酮的活性,即可以降解玉米赤霉烯酮。
图3表明玉米赤霉烯酮降解酶突变体在ph9.5条件下具有最高酶活性。以此最高酶活性体系的底物玉米赤霉烯酮降解量作为相对活性100%,其它反应体系的底物玉米赤霉烯酮降解量与此最高酶活性体系的底物玉米赤霉烯酮降解量的比值作为各自的相对活性。在ph9.0-ph10.5条件下均具有60%以上的活性。
对照组:以对照菌bl21/pet28a获得的蛋白(记作对照酶液)进行上述实验,结果不管在哪个ph条件下,对照酶液都没有降解玉米赤霉烯酮的活性。
实验设3次重复,结果一致。
(三)酶热稳定性
用ph9.5的50mmglycine-naoh缓冲液稀释实施例1的步骤5中的zhday3纯酶液,用稀释后的酶液以玉米赤霉烯酮为底物进行酶活测定。将稀释后的酶液记作稀释酶液。
将稀释酶液分别在20℃、30℃、35℃、40℃、45℃、50℃、55℃、60℃水浴放置10分钟,测定酶的残余活性。结果显示,酶在20℃至45℃的酶活相对稳定,在50℃处理10分钟丧失50%活性,在55℃处理10分钟丧失80%活性(参见图4)。
(四)ph耐受性
将稀释酶液分别在ph5.5、6.0、6.5、7.0、7.5、8.0、8.5、9.0、9.5、10.0、10.5、11.0条件下,温度4.0℃条件下放置16h后,以玉米赤霉烯酮为底物测定残余酶活。结果显示ph5.5-10.5条件下仍然残留60%以上的相对酶活(参见图5)。说明该酶具有良好的ph耐受性。
(五)底物特异性
将稀释酶液分别在相同浓度(反应体系中底物终浓度为20.0μg/ml)的不同底物条件下进行酶活测定,底物分别为玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇。
以玉米赤霉烯酮为底物测得酶活为参比(100%),以α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇为底物所测相对酶活分别为49.0%、44.2%、48.9%、32.7%。因此,该酶对玉米赤霉烯酮和α-玉米赤霉烯醇的活性比较高。
实施例3、以β-玉米赤霉烯醇为底物验证蛋白功能
酶活单位定义为1min内降解1μg底物玉米赤霉烯酮所需要的酶量作为一个酶活单位u。
如下所述的稀释酶液是用50mmglycine-naoh缓冲液稀释实施例1的步骤5中的zhday3纯酶液得到的。
实验组:以β-玉米赤霉烯醇为底物(底物在反应体系中的终浓度为20.0μg/ml),活性测定反应体系为0.5ml,由0.45ml底物溶液和0.05ml稀释酶液;反应体系的ph值为9.5;反应体系在最适温度40℃下反应10min后,0.5ml色谱级甲醇终止反应,冷却后使用高效液相色谱仪(hplc)测定底物降解量。
实验设三次重复,结果一致。
结果显示在40℃、ph9.5条件下,以β-玉米赤霉烯醇为底物具有一定的酶活。
实施例4、zhday3蛋白n153h突变体的构建
通过整个环状质粒pet28a-zhday3的反向聚合酶链式反应扩增进行突变。n153h的诱变引物是:n153h-f(5'gtcaagccatgtggttgtgggaagtg3'),如seqidno.7所示;n153h-r(5'caaccacatggcttgacattgcagcg3'),如seqidno.8所示。琼脂糖电泳回收pcr产物,之后用dpni酶处理以除去模板链,电击转化大肠杆菌dh5α后涂布于含有50μg/ml卡那霉素的lb平板,37℃过夜培养,将得到的转化子进行测序验证,结果表明,153位的n被突变成h而其他位置未发生突变,将该重组质粒命名为pet28a-zhday3(n153h)。
之后按照实施例1步骤4、5中方法制备目标蛋白,然后进行底物特异性的测定。将稀释酶液分别在相同浓度(反应体系中底物终浓度为20.0μg/ml)的不同底物条件下进行酶活测定,底物分别为玉米赤霉烯酮、α-玉米赤霉烯醇、β-玉米赤霉烯醇、α-玉米赤霉醇、β-玉米赤霉醇。
实验设三次重复。试验结果参见表2。
表2突变前后的酶活比较
通过表2的试验结果经计算可知,在40℃、ph9.5条件下,玉米赤霉烯酮降解酶zhday3在经过一个定点(n153h)突变后,对于底物α-玉米赤霉醇的底物特异性增加了2.1倍,对于β-玉米赤霉醇的底物特异性增加了1.4倍,对于玉米赤霉烯酮zen的活性没有提高,对于α-玉米赤霉烯醇的活性提高了33%,对于β-玉米赤霉烯醇的活性降低了22%(相比zhday3)。
以上所述仅为本发明的较佳实施例,并不用以限制本发明,凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
序列表
<110>湖北大学
<120>一种玉米赤霉烯酮降解酶突变体及其编码基因和应用
<160>8
<170>siposequencelisting1.0
<210>1
<211>264
<212>prt
<213>人工序列(artificialsequence)
<400>1
metargthrargserasnilethrthrlysasnglyilehistrptyr
151015
tyrgluglngluglyserglyprohisvalvalleuileproaspgly
202530
leuglyglucyslysmetpheasplysprometserleuilealaasn
354045
serglyphethrvalthrthrpheaspmetproglymetserargser
505560
serglualaproprogluthrtyrglngluilethralaglnlysleu
65707580
alasertyrvalileserilecysaspgluleualaileasplysala
859095
thrphetrpglycysserserglyglycysthrvalleualaleuval
100105110
alaasptyrprothrargvalargasnalaleualahisgluvalpro
115120125
thrtyrleumetgluaspleulysproleuleuglumetaspaspglu
130135140
alavalseralaalametserserasnvalvalvalglyservalgly
145150155160
aspilegluglysertrpglngluleuglygluglualahisalaarg
165170175
leutrplysasntyrproargtrpalaargglytyrproglytyrile
180185190
proglnserthrprovalserlysgluaspleuilelysalaproleu
195200205
asptrpthrvalglyalaserthrprothralaargpheleuaspasn
210215220
ilevalthralathrlyshisasnilepropheglnthrleuprogly
225230235240
methispheprotyrvalthrhisprogluvalphealaglutyrval
245250255
valglulysthrarglystyrleu
260
<210>2
<211>795
<212>dna
<213>人工序列(artificialsequence)
<400>2
atgcgcaccaggtccaatatcaccaccaaaaacggaatccattggtactacgagcaagag60
ggttctggtcctcacgtggtgctgataccagatggcttaggagagtgcaagatgttcgac120
aagcctatgtctcttatagcgaacagtggcttcaccgtcacaaccttcgatatgccaggg180
atgtcgaggtcgtctgaagcaccaccggagacataccaagagatcaccgcccagaagctg240
gccagctatgtcattagcatctgcgacgaattagcaatcgacaaggctacattctgggga300
tgcagctcaggcggctgtactgtgcttgctctggttgctgactatcctacaagggtgcgg360
aatgcgctggcccatgaagttccgacttatctcatggaagacttgaagcccctgcttgaa420
atggatgatgaggccgtctccgctgcaatgtcaagcaatgtggttgtgggaagtgtgggt480
gacatcgagggttcgtggcaagaactcggtgaggaggctcatgcaaggctttggaagaat540
tatccccggtgggctcgcggttaccctggatatataccacaatccaccccagtaagcaaa600
gaagacttgataaaggcaccgctggactggacagtcggcgcatcgacacccacggctcga660
ttcctggacaatatcgtgacggcgaccaaacacaacatcccctttcaaaccctcccggga720
atgcatttcccgtatgttacccacccggaagttttcgcggagtatgtggtggaaaagact780
cgaaagtacttgtaa795
<210>3
<211>264
<212>prt
<213>人工序列(artificialsequence)
<400>3
metargthrargserasnilethrthrlysasnglyilehistrptyr
151015
tyrgluglngluglyserglyprohisvalvalleuileproaspgly
202530
leuglyglucyslysmetpheasplysprometserleuilealaasn
354045
serglyphethrvalthrthrpheaspmetproglymetserargser
505560
serglualaproprogluthrtyrglngluilethralaglnlysleu
65707580
alasertyrvalileserilecysaspgluleualaileasplysala
859095
thrphetrpglycysserserglyglycysthrvalleualaleuval
100105110
alaasptyrprothrargvalargasnalaleualahisgluvalpro
115120125
thrtyrleumetgluaspleulysproleuleuglumetaspaspglu
130135140
alavalseralaalametserserhisvalvalvalglyservalgly
145150155160
aspilegluglysertrpglngluleuglygluglualahisalaarg
165170175
leutrplysasntyrproargtrpalaargglytyrproglytyrile
180185190
proglnserthrprovalserlysgluaspleuilelysalaproleu
195200205
asptrpthrvalglyalaserthrprothralaargpheleuaspasn
210215220
ilevalthralathrlyshisasnilepropheglnthrleuprogly
225230235240
methispheprotyrvalthrhisprogluvalphealaglutyrval
245250255
valglulysthrarglystyrleu
260
<210>4
<211>795
<212>dna
<213>人工序列(artificialsequence)
<400>4
atgcgcaccaggtccaatatcaccaccaaaaacggaatccattggtactacgagcaagag60
ggttctggtcctcacgtggtgctgataccagatggcttaggagagtgcaagatgttcgac120
aagcctatgtctcttatagcgaacagtggcttcaccgtcacaaccttcgatatgccaggg180
atgtcgaggtcgtctgaagcaccaccggagacataccaagagatcaccgcccagaagctg240
gccagctatgtcattagcatctgcgacgaattagcaatcgacaaggctacattctgggga300
tgcagctcaggcggctgtactgtgcttgctctggttgctgactatcctacaagggtgcgg360
aatgcgctggcccatgaagttccgacttatctcatggaagacttgaagcccctgcttgaa420
atggatgatgaggccgtctccgctgcaatgtcaagccatgtggttgtgggaagtgtgggt480
gacatcgagggttcgtggcaagaactcggtgaggaggctcatgcaaggctttggaagaat540
tatccccggtgggctcgcggttaccctggatatataccacaatccaccccagtaagcaaa600
gaagacttgataaaggcaccgctggactggacagtcggcgcatcgacacccacggctcga660
ttcctggacaatatcgtgacggcgaccaaacacaacatcccctttcaaaccctcccggga720
atgcatttcccgtatgttacccacccggaagttttcgcggagtatgtggtggaaaagact780
cgaaagtacttgtaa795
<210>5
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>5
cgcggatccatgcgcaccaggtccaatatcacc33
<210>6
<211>34
<212>dna
<213>人工序列(artificialsequence)
<400>6
ccgctcgagttacaagtactttcgagtcttttcc34
<210>7
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>7
gtcaagccatgtggttgtgggaagtg26
<210>8
<211>26
<212>dna
<213>人工序列(artificialsequence)
<400>8
caaccacatggcttgacattgcagcg26
- 还没有人留言评论。精彩留言会获得点赞!