一种调控黄瓜圆叶性状相关蛋白及其编码基因与应用的制作方法
本发明涉及基因工程和黄瓜的蛋白和基因,属于分子遗传育种领域,具体涉及一种调控黄瓜圆叶性状相关蛋白及其编码基因与应用。
背景技术:
黄瓜(Cucumis sativus L.2n=2x=14)属葫芦科甜瓜属一年生蔓生草本植物,一般叶片呈掌状3-5裂,裂片三角形并具锯齿。随着基因组学的发展,突变体材料成为研究基因功能直接、有效的途径之一。在用EMS构建的黄瓜突变体库中,发现一个可稳定遗传的单基因控制黄瓜圆叶突变体。突变体表现出叶片边缘平滑呈近圆形,无花萼,雌花不育,子叶不对称发育以及根异常发育等表型。叶片是光合作用和呼吸作用的主要器官,也是植物形态建成的重要指标之一,对植物的生长和发育具有极大地影响。叶形极大地影响着株型、光合面积、产量和商品性状等,叶缘形状很大程度上维持着叶形的多样性,而花器官的发育不仅可以影响植株繁育并且与黄瓜产量的大小也密切相关。目前黄瓜叶形分子调控机制仍然未知,花器官研究主要集中在性别决定基因上,形成调控基因的研究较少,因此,该突变体是研究黄瓜叶形和花器官发育调控基因及其调控机制的理想材料。随着新一代测序技术的发展,利用改进的MutMap法对圆叶突变体和野生型F2代群体进行混池测序,在一号染色体上初定位到一个19.1Mb到22.0Mb的区间,通过EMS诱变原则(G→A和C→T)以及ΔSNP指数≥0.8,经鉴定获得了一个SNP,SNP-1G19389144,位于基因Csa1M537400的第二个外显子上,引起编码蛋白保守结构域的氨基酸序列发生改变,最终确定调控黄瓜圆叶突变体的基因Csa1M537400,命名为CsPID。
叶片(包括子叶)是茎端分生组织产生的第一类侧生器官,在植物发育中具有重要地位。大量证据表明叶片发育受到体内遗传机制和体外环境因子的双重调节。植物激素,尤其是生长素在协调体内外调节机制中起着不可或缺的作用。生长素的稳态调控、极性运输和信号转导影响叶片发育的全过程。植物生长素作为一种重要的植物激素,参与了许多生理生化过程的调节与控制。如根的伸长生长和向地性、不定根的形成、维管组织的分化、顶端优势、形成层的细胞分裂、细胞的伸长生长、植物向性等。生长素的极性运输与植物的许多生长发育过程密切相关,但对其调控机制的认识仍不全面。研究表明,生长素对植物生长发育的调节主要依赖于极性运输形成的生长素浓度梯度。PIN蛋白可逆的磷酸化活性对调节生长素极性运输起着关键作用,而这一过程需要丝氨酸/苏氨酸蛋白激酶PINOID(PID)和PP2A磷酸化酶的共同参与。有证据表明PID是通过调节PIN蛋白的亚细胞定位来控制极性生长素的转运,CsPID表达异常可能改变了生长素稳态水平引起生长素极性运输严重缺陷从而导致黄瓜叶片形状的改变。
PID基因编码丝氨酸/苏氨酸的蛋白激酶(protein-serine/threonine kinase),属于AGC激酶家族,存在于子叶原基、叶片、花组织和正在发育的器官或分生组织近基端的微管组织中(Benjamins et al.,2001)。蛋白激酶PINOID((PID)起生长素极性运输的正调控剂作用(Friml et al.,2004)。AtPID,OsPID过量表达可产生与生长素运输增加的相关表型,可能是生长素极性运输的正调控因子(Morita and Kyozuka,2007)。PID参与生长素的极性运输和信号转导,其功能缺失突变体与其他多个生长素相关突变体组合导致胚胎和幼苗的子叶缺失。拟南芥插入突变体pid::En197和pid::En310表现出花通常包含很少或没有萼片,雌蕊小并且不产生或仅产生很少的种子,大约50%的突变体幼苗显示异常的子叶等表型,与本研究黄瓜圆叶突变体表型相似(Benjamins et al.,2001)。目前生长素在叶片发育过程中的功能,使我们得以窥见生长素是如何调控特定器官的发育,以及特定器官又是如何响应特定激素的调控。随着研究的深入,一个更为复杂而清晰的叶片发育和形态建成调控网络将被揭示。这些结果对黄瓜叶型改良、产量增加以及理想株型培育研究也具有重要的意义。
技术实现要素:
本发明所要解决的技术问题是提供一种调控黄瓜圆叶性状相关蛋白及其编码基因与应用。PID基因编码丝氨酸/苏氨酸的蛋白激酶,属于AGC激酶家族,在氨基酸取代的情况下,调控生长素极性运输,突变体造成生长素极性运输缺陷,表现出叶边缘平滑,锯齿不明显突出,叶片呈近圆形;子叶不对称发育,一般出现三片子叶;主根不发达,主侧根没有明显区别;雌雄花无花萼,雌花不育等表型。可以为黄瓜叶型改良、产量增加以及理想株型培育和生长素信号通路分子调控机制的研究创造条件。
本发明解决上述技术问题所采用的技术方案为:定位到一个黄瓜圆叶性状相关蛋白,命名为CsPID,来源于黄瓜,CsPID是如下a)或b)的蛋白:
a)由序列表中SEQ ID NO:2所示的氨基酸序列组成的蛋白质;
b)在序列表SEQ ID NO:2所示的氨基酸序列中经过取代一个氨基酸且与黄瓜圆叶突变相关的由a)衍生的蛋白质。
氨基酸序列由474个氨基酸残基组成,属于AGC激酶家族,调控生长素极性运输,突变体造成生长素极性运输缺陷,突变体表现出圆叶无花萼,雌花不育,子叶不对称发育以及根异常发育等表型。上述b)中的CsPID可人工合成,也可先合成其编码基因,再进行生物表达得到。上述b)中的CsPID的编码基因可以通过将序列表中SEQ ID NO:1所示的DNA序列中进行一个碱基对的非同义突变。
编码所述CsPID的基因也属于本发明的保护范围。
CsPID基因具体可分为1)或2)或3)的基因:
1)其核苷酸序列是序列表中SEQ ID NO:1所示的DNA分子;
2)在严格条件下可与序列表SEQ ID NO:1限定的DNA序列杂交且编码上述与黄瓜圆叶性状相关蛋白的DNA分子;
3)与1)的基因有90%以上的同源性,且编码上述与黄瓜圆叶性状相关蛋白的DNA分子。
序列由1452个碱基组成,自5端第1-3位为起始密码子,编码具有序列表中SEQ ID NO:2所示的氨基酸残基序列的蛋白质;该氨基酸序列为PID蛋白激酶,具有个2外显子和1个内含子。基因碱基序列的1092bp位原本的鸟嘌呤(G)被腺嘌呤(A)取代,导致编码的V(缬氨酸)突变为M(甲硫氨酸)。
含有CsPID基因的重组载体、转基因细胞系和重组菌也属于本发明的保护范围。
本发明的优点在于利用改进的MutMap的方法定位调控黄瓜圆叶无花萼突变体的基因,发现了一种调控黄瓜叶形相关蛋白及其编码基因,该蛋白的氨基酸序列如序列表2所示,属于AGC激酶家族,与生长素极性运输相关;该蛋白的编码基因如序列表SEQ ID NO:1所示的碱基序列,该碱基序列在1092bp位的鸟嘌呤(G)被腺嘌呤(A)取代,编码的氨基酸残基序列的346位的Val(缬氨酸)突变为Met(甲硫氨酸),该蛋白抑制生长素极性运输,导致生长素运输极性缺陷,具体植株表型为叶边缘平滑,中部没有明显突出,且锯齿整齐,叶片呈近圆形;主根不发达,主侧根没有明显区别;雌雄花无花萼,雌花不育等。该基因及其蛋白的发现,可以为黄瓜叶型改良、产量增加以及理想株型培育和生长素信号通路分子调控机制的研究提供基础。
附图说明
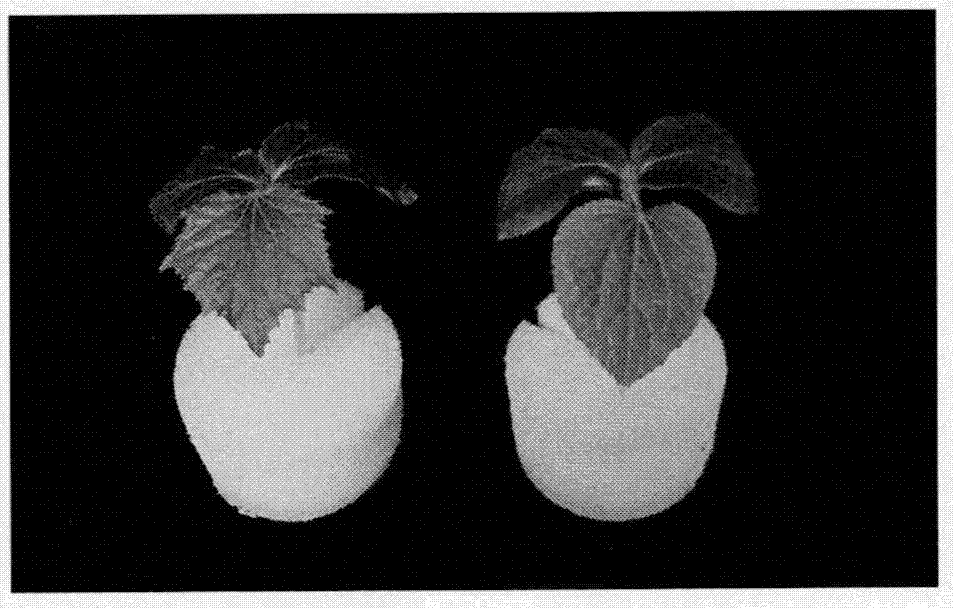
黄瓜圆叶突变体rl与野生型WT表型对比图。图1为成熟期株型表型,左边为野生型WT,右边为突变体rl表现出整株圆叶表型。图2为一叶期时叶型表型,左边为野生型表现出掌状浅裂,右边为突变体表现出近圆形;图3为突变体表现出子叶不对称现象;图4为雌花花萼的发育情况,图5为雄花花萼的发育情况,对比左边野生型WT,右边突变体rl都表现出无花萼现象;图6主根与侧根发育情况,左边为突变体rl,右为野生型WT;
具体实施方式
以下结合附图、实施例对本发明作进一步详细描述。
实施例1突变体的表型及遗传分析
从EMS(ethyl methane sulfonate)诱变的长春密刺突变体库中筛选到一个黄瓜圆叶突变体rl。该突变体叶发育初期就表现出圆形,雌花与雄花都无花萼,雌花不育,子叶不对称发育(三片子叶为最常见的表型)以及根异常发育等表型。
以突变体r1为父本,与野生型长春密刺杂交,获得子一代F1植株,所有F1后代均具有野生型表型。由于纯合的r1突变体植物是雌性不育,因此统计来自r1突变体和WT野生型之间杂交的F2代的野生型植物(RLRL或RLr1)和突变型植物(rlr1),在含有78株植物的F2群体中只有24株植物表现出圆叶表型,分离比为3∶1(χ2=0.19,P=0.66),表明该突变体由单隐性基因调控,命名该基因为CsPID。
实施例2 CsPID基因的定位
按常规方法-CTAB法提取F2分离群体的圆叶突变体和野生型叶片DNA。DNA混池用超声波破碎仪破碎到150bp~250bp,对基于Illumina HiSEQ 2500测序平台构建的小片段DNA文库进行全基因组重测序。通过质量控制去除低质量的reads后,利用BWA(版本:0.5.9rc1)和SAMtools软件(版本:0.1.19)将数据和黄瓜基因组序列(“9930”参考基因组)进行比对,将SNP数目除以该位点的测序深度计算allele index。在两个混池中筛选出相同的SNP,将对应的allele index相减计算Δallele index。采用同样的方法随机对1株野生型长春密刺单株进行高通量测序,得到野生型长春密刺与9930之间的SNP位点,作出野生型池的SNP指数图,这些差异主要是由参考序列测序错误造成的。在野生型混池筛选出allele index≥0.9的SNP,去掉突变体混池中与筛选出的SNP相同的SNP,这些SNP中SNP index≤0.3或者测序深度小于等于7的SNP被认为是不可信的。在可靠的SNP位点中,以1Mb为窗口,10kb为步移利用SNPindex计算SNPindices绘制折线图,SNP indices最高的区域即为候选基因的区域。理论上,造成突变性状的突变位点SNP指数应该等于1,即为纯合位点;由于遗传连锁,其附近的SNP位点指数应该等于或接近1。在初定位区域内寻找成簇分布的SNP位点,再根据EMS诱变通常产生的突变,即G→A或C→T突变,一共找到了27个SNP位点。27个SNPs中的25个SNPs位于基因间区或内含子区,仅有2个SNPs与编码区(CDS)区相对应。其中SNP(SNP-1G19389144)是一个发生在蛋白质编码区的非同义突变,原本的鸟嘌呤(G)被腺嘌呤(A)取代,导致编码的V(缬氨酸)到M(甲硫氨酸)的突变,从而确定导致突变的候选基因CsPID。
实施例3 CsPID基因克隆和鉴定
CsPID基因突变位于在Csa1M537400的第二个外显子1092bp处,碱基G突变为A。为了验证测序的结果,对Cucurbit Genomics Database中编号为Csa1M537400基因的cDNA进行测序,证实编码区cDNA原本的鸟嘌呤(G)突变为腺嘌呤(A),导致编码的V(缬氨酸)突变为M(甲硫氨酸),与测序结果相符,证实该基因的突变导致产生圆叶无花萼突变体,命名该基因为CsPID。
CsPID基因全长cDNA的获得:
黄瓜长春密刺和圆叶总RNA的提取采用TRIzol方法,并在25℃条件下进行DNA消化30分钟,从总RNA中吸取2mg为模板采用TaKaRa公司的反转录试剂盒PrimeScriptTM RT reagent Kit合成第一链cDNA。以此cDNA为模板,用引物Csa1M537400-F(5’-ATGTTACAACACCTTCCCCCAGA-3’)和引物Csa1M537400-R(5’-GAAATAATCGAACCCGGTCGACT-3’)采用TaKaRa公司的高保真酶GXL DNA polymerase进行PCR扩增反应,反应条件如下:
反应体积26ul,其中含有:
用双蒸水补足至26ul体积。
反应程序如下:95℃,变性5分钟,然后95℃变性30秒,57.8℃退火30秒,72℃延伸90秒,扩增35个循环;最后在72℃下延伸10分钟。
扩增产物用OMEGA公司的DNA凝胶回收试剂盒按产品说明书进行纯化,然后纯化的产物用rTaq酶(TaKaRa)进行加A反应,反应条件如下:
反应体积10ul,其中含有
反应程序:72℃连接10分钟。
将加A后的产物与pMD19-T载体(TaKaRa)在16℃下连接4小时,构建重组载体pMD19-CsPID。42℃热击70秒将重组载体pMD19-CsPID转化大肠杆菌DH5α(TaKaRa),转化物在含有氨苄青霉素的LB平板培养基上生长,挑取克隆,提取质粒,送交测序。测序结果表明,扩增到的片段的核苷酸序列如序列表SEQ ID NO:1所示,将该片段命名为CsPID基因,CsPID基因全长1425bp,其编码的氨基酸序列见序列表中序列SEQ ID NO:2所示。
参考文献:
[1]Benjamins,R.,Quint,A.,Weijers,D.,Hooykaas,P.,and Offringa,R.(2001)The PINOID protein kinase regulates organ development in Arabidopsis by enhancing polar auxin transport.Development 128:4057-4067.
[2]Friml,J.,Yang,X.,Michniewicz,M.,Weijers,D.,Quint,A.,Tietz,O.,et al.(2004)A PINOID-dependent binary switch in apical-basal PIN polar targeting directs auxin efflux.Science 306:862-865.
[3]Morita,Y.,and Kyozuka,J.(2007)Characterization of OsPID,the rice ortholog of PINOID,and its possible involvement in the control of polar auxin transport.Plant Cell Physiol.48:540-549.
- 还没有人留言评论。精彩留言会获得点赞!