一种里氏木霉及其在单宁酶生产中的应用的制作方法
本发明涉及基因工程技术领域,具体内容涉及一种里氏木霉及其在单宁酶生产中的应用。
背景技术
单宁酶全称是单宁酯酰水解酶(tannase,ec3.1.1.20),它可以水解没食子酸单宁中的酯键和缩酚羧键,生成没食子酸和葡萄糖。单宁酶在自然界中分布相当广泛,有关单宁酶较早的报道是20世纪20年代,freudenberg在进行黑曲霉培养时发现在其菌丝中含有单宁酶,以后又发现各类微生物,包括真菌、酵母、细菌都能可产生单宁酶;单宁酶还存在于一些植物材料中,例如茶叶提取物。单宁酶可广泛应用于食品工业、精细化工等领域。
在液态茶和速溶茶的生产中,由于多酚类、咖啡碱、蛋白质等物质生成络合物,形成所谓的“冷后浑”,因此需要进行“转溶”工艺。单宁酶就是酶法转溶的专一酶。它能断裂儿茶酚与没食子酸间的酯键,使苦涩味的酯型儿茶素水解,释放出的没食子酸阴离子又能与茶黄素、茶红素竞争咖啡碱,形成分子量较小的水溶性短链物质,从而降低茶汤的混浊度。单宁酶应用于红茶、绿茶、乌龙茶深加工中,还能使茶叶中可溶性金属元素含量增加。随着单宁酶在茶饮料业中的应用越来越广泛深入,美国食品与药物管理局(fda)已确认单宁酶为安全产品。
单宁酶还可用于啤酒和果汁的澄清、防止果酒和果汁中酚类引起的变质、咖啡风味的软饮料生产、麦芽多酚的稳定处理、葡萄酒风味的改进等方面。韩国科学家chaesoo-kyu和yutai-jong用单宁酶处理橡实,使之可以食用;还将米粉与橡实粉1:1混合,再经淀粉酶和单宁酶作用后可用于酿造橡实果酒。单宁酶还可用来除去天然食品中因单宁而引起的苦涩味。
此外,单宁酶还广泛应用于饲料工业领域。来源于植物的饲料往往含有单宁,它可以使植物蛋白以及家畜自身所分泌的消化酶蛋白凝固沉淀,从而减少家畜对摄入蛋白的消化率和增加粪便排泄中的氮源。因此,在饲料加工过程中如用单宁酶进行相应处理,将大大提高家畜对植物蛋白的利用和吸收率,降低畜牧业的生产成本。viverosa等研究发现,在对家鸡投放的豆类更壳饲料中添加单宁酶,将显著地增加鸡肉的获取量。
单宁酶主要是由微生物生产的,能够产生单宁酶的菌种十分丰富,除了传统的青霉、曲霉、酵母,还有巴斯德菌、寄生内座壳菌、绿色木霉和茄形镰刀菌等。目前,市售的单宁酶产品种类比较少,而且因生产菌株产量较低,导致该酶的价格居高不下,严重限制了该酶在食品、饲料等领域的广泛应用。因此,开发高产单宁酶的生产菌株迫在眉睫。
技术实现要素:
本发明为解决现有技术问题,提供了一种高产单宁酶的里氏木霉(trichodermareesei)菌株。申请人首先构建得到重组表达单宁酶的里氏木霉工程菌株,然后通过紫外诱变的方法筛选获得一株单宁酶产量得到显著提高的突变菌株,可广泛应用于单宁酶的生产。
本发明一方面提供了一种里氏木霉工程菌株,所述菌株携带有用于重组表达单宁酶基因的重组质粒。
所述单宁酶的氨基酸序列为seqidno:1,其编码核苷酸序列为seqidno:2。
本发明一方面提供了一种突变菌株里氏木霉4qt2(trichodermareesei4qt2),已于2018年6月21日保藏于中国武汉武汉大学的中国典型培养物保藏中心,保藏编号为cctccno:m2018389。
本发明还提供了所述里氏木霉突变株在生产单宁酶中的应用。
本发明将来源于米曲霉(aspergillusoryzae)的单宁酶基因在里氏木霉(trichodermareesei)宿主细胞中表达,构建得到重组菌株里氏木霉4qt,20l罐发酵160h后,其发酵上清液中单宁酶酶活达到101u/ml。申请人以里氏木霉4qt为出发菌株,通过多轮紫外诱变,最终筛选获得一株突变菌里氏木霉4qt2,能大幅度提高单宁酶的表达量,20l罐发酵160h后,突变菌株发酵上清液中单宁酶酶活高达158u/ml,比出发菌提高了56.4%,取得了意料不到的技术效果。所述里氏木霉菌株可广泛应用于单宁酶的生产,从而有利于降低单宁酶的生产成本,促进其在食品和饲料加工等领域中的推广与应用。
附图说明
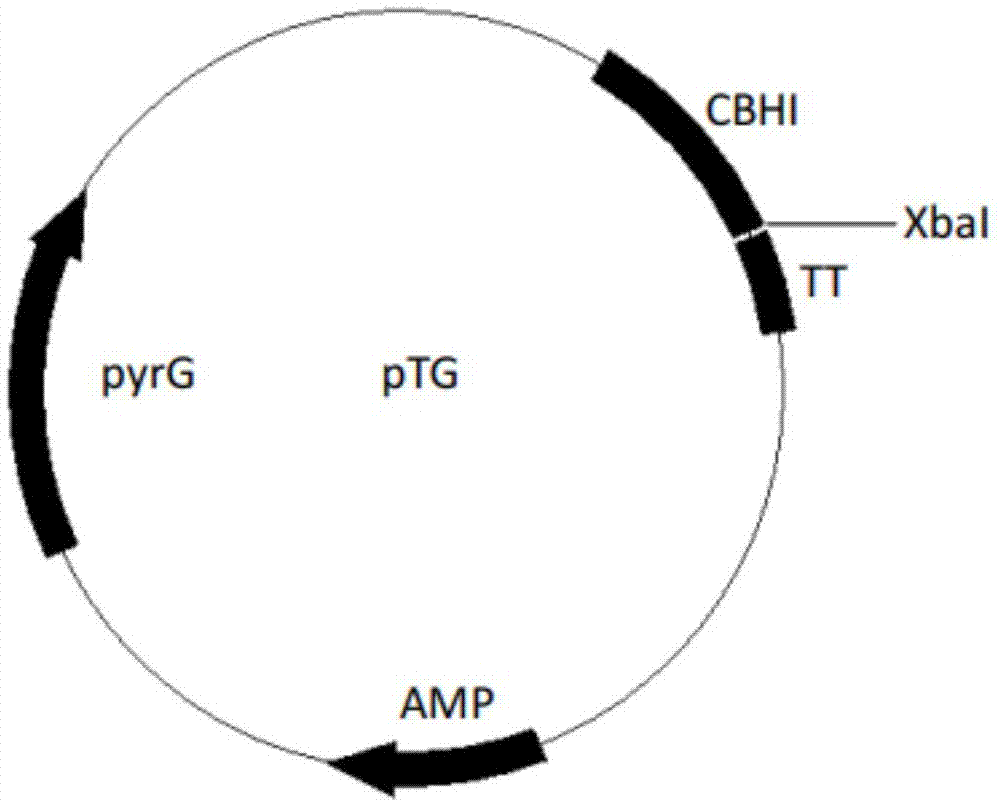
图1为质粒ptg图谱;
图2为20l罐发酵进程曲线;
图3为sds-page蛋白电泳图:其中:m为蛋白分子量marker,泳道1、2分别为突变菌里氏木霉4qt2、出发菌里氏木霉4qt发酵上清液;箭头所指65kda处的蛋白条带即为单宁酶。
具体实施方式
本发明用到了遗传工程和分子生物学领域使用的常规技术和方法,例如molecularcloning:alaboratorymanual,3nded.(sambrook,2001)和currentprotocolsinmolecularbiology(ausubel,2003)中所记载的方法。这些一般性参考文献提供了本领域技术人员已知的定义和方法。但是,本领域的技术人员可以在本发明所记载的技术方案的基础上,采用本领域其它常规的方法、实验方案和试剂,而不限于本发明具体实施例的限定。
下面结合具体实施方式对本发明进行详细描述。
实施例1单宁酶基因的克隆及重组质粒的构建
申请人根据里氏木霉密码子偏好性对来源于米曲霉(aspergillusoryzae)的单宁酶基因进行密码子优化,在第1个氨基酸密码子前增加6个碱基tctaga(xbai酶切位点),在其终止密码子taa后增加tctaga(xbai酶切位点),优化后的核苷酸序列为seqidno:2,由上海捷瑞公司合成,其编码的氨基酸序列为seqidno:1。
pcr扩增上述单宁酶基因。引物序列如下:
引物1(f):gctctagaatgcgccagcactcccgcatg
引物2(r):gctctagattagtagacggggaccttgaa
pcr反应条件为:94℃变性5min;然后94℃变性30s,56℃复性30s,72℃延伸110s,30个循环后,72℃保温10min。琼脂糖凝胶电泳结果显示,单宁酶基因大小为1767bp的片段。
将上述获得的单宁酶基因片段与表达质粒ptg分别进行限制性内切酶xbai单酶切,酶切条件如下:
37℃水浴酶切处理2h,电泳后分别回收两个目的片段,溶于20ulddh2o。用t4dna连接酶进行连接,连接体系如下:
22℃连接1h,转化大肠杆菌dh5a感受态,涂布lb+aap平板,37℃培养过夜后长出单菌落,菌落pcr验证连接正确的转化子提取质粒送测序,测序正确后,即得到含有单宁酶基因的重组质粒ptg-tan。
实施例2里氏木霉重组菌株的构建
1、原生质体制备:
接种里氏木霉宿主细胞于pda+u(马铃薯200g/l,煮沸20-30min后过滤除渣;葡萄糖2%;uridine1%;琼脂粉1.5%)平板,30℃培养5-7d;割取2cm×2cm大小的菌块,接种100ml液体pda+u(马铃薯200g/l,煮沸20-30min后过滤除渣;葡萄糖2%;uridine1%)培养基中,30℃培养16h生长菌丝体用于转化;将长好的菌丝体过滤后,用20ml1.2m硫酸镁溶液重悬;加入0.2g溶菌酶,30℃、100rpm培养2-3h;将裂解好的菌丝用2层擦镜纸过滤,3000rpm离心10min获得原生质体;将裂解好的菌丝用擦镜纸过滤,离心获得原生质体;再用适量的山梨醇溶液重悬。
2、转化:
将上述获得的里氏木霉原生质体用1.2m山梨醇溶液清洗2遍,再用适量的山梨醇溶液重悬,使原生质体浓度达到108个/ml;每200ul原生质体中分别加入10ul准备好的重组载体ptg-tan,加入50ul25%的peg6000,冰浴20min,再加入2ml25%的peg6000,室温放置5min;加入4ml山梨醇溶液颠倒混匀,倒入50ml转化上层培养基后,倾倒入4个转化下层平板中,待上层培养基凝固后,于30℃培养箱中倒置培养5d。
3、转化子筛选:
培养5d后,挑取长出的菌落,点种到转化下层平板进行复筛,30℃培养3d。将正常生长的转化子分别接种到新鲜的pda平板,30℃培养5-7d。每个转化子割取2cm×2cm大小的菌块,分别接种50ml液体摇瓶培养基(葡萄糖1%;乳糖2%;玉米浆1.5%;硫酸铵0.9%;硫酸镁0.15%;柠檬酸0.073%;氯化钙0.1125%;微量元素0.1%)中发酵,28℃培养5d。培养5d后,离心菌体获得上清液即为粗酶液,分别进行sds-page蛋白电泳检测和单宁酶酶活力检测。
检测阳性转化子发酵上清液中单宁酶活力,筛选出酶活最高的阳性转化子,命名为里氏木霉4qt(trichodermareesei4qt)。
实施例3诱变筛选
紫外诱变导致的突变随机性很强,突变产生的效果也是随机的,难以预测。因此,为了获得有效的正突变,技术人员通常需要进行多轮紫外诱变,筛选的工作量较大,而且存在无法获得有效正突变的可能性。但因为紫外诱变所需设备简单、费用少,并且可在短时间内获得大量突变体,因此,它现在仍是一种常用的诱变选育方法。
申请人以里氏木霉4qt为出发菌株,通过紫外诱变方法对其进行遗传学改造,进一步提高其单宁酶的产量。
1、确定致死率:
接种出发菌里氏木霉4qt于pda平板,30℃培养5-7d。待菌落表面产生大量孢子时,吸取5ml无菌水洗脱,获得孢子液,离心后用无菌水重悬,用血球计数板计数。取一个90mm培养皿,加入5ml稀释好的孢子悬液(浓度为1×107),加入转子并在磁力搅拌器上搅拌使孢子液处于均匀状态。在无菌超净工作台中,用功率为9w的紫外灯于垂直距离20cm的上方照射,分别照射30s、45s、60s、75s、90s、105s、120s,取照射后的孢子液稀释10、100、1000倍,取100ul涂布pda平板,30℃培养2-3d后计数,以未照射的孢子液为对照,计算致死率。其中照射90s时,致死率为95%,选取该照射时间进行后续诱变实验。
2、第一轮诱变筛选:
取一个90mm培养皿,加入5ml稀释好的孢子悬液(浓度为1×107),加入转子并在磁力搅拌器上搅拌使孢子液处于均匀状态。在无菌超净工作台中,用功率为9w的紫外灯于垂直距离20cm的上方照射,照射90s后稀释1000倍,取100ul涂布pda平板,30℃培养2-3d。
共涂布200块pda平板,30℃培养2-3d后,每个平板长出30-50个菌落,先通过菌落形态,筛选短分枝的突变体。申请人挑取出菌落形态较小、菌丝致密、菌落周围绒毛较短的突变菌共75个,分别接种到pda平板,30℃培养5-7d。每个转化子割取2cm×2cm大小的菌块,分别接种至50ml液体摇瓶培养基中发酵,28℃培养5d。培养5d后,离心菌体获得上清液即为粗酶液,分别进行蛋白电泳检测和单宁酶酶活力检测。
结果显示,第一轮紫外诱变筛选获得的75株突变菌中,没有一株突变菌发酵上清液酶中单宁酶的酶活高于出发菌;其中,46株突变菌的酶活与出发菌基本相当,其余29株突变菌的酶活甚至比出发菌普遍降低了6-18%,。
申请人按照上述方法继续进行了8轮诱变筛选,最终获得3株单宁酶产量显著高于出发菌的突变菌株,分别命名为里氏木霉4qt1,4qt2,4qt3,其单宁酶产量比出发菌分别提高了35%,58%,61%。
(1)单宁酶酶活单位的定义
在30℃、ph值为5.0的条件下,每分钟降解没食子酸丙酯(pg)溶液释放产生1μmol没食子酸所需的酶量,定义为一个酶活力单位u。
(2)酶活测定方法
没食子酸标准溶液(10mmol/l):秤取没食子酸0.18813g,用ph5.0的磷酸氢二钠-柠檬酸缓冲液溶解,定容至100ml。
没食子酸丙酯(10mmol/l):秤取没食子酸丙酯0.2122g,再加入80mlph5.0的磷酸氢二钠-柠檬酸缓冲液,加热直至完全溶解,冷却后调节ph至5.0,定容至100ml。
罗丹宁溶液(0.667%):秤取罗丹宁0.667g,加入80ml甲醇溶解,溶解后用甲醇定容至100ml,现配现用。
没食子酸标准曲线的绘制:用ph5.0的磷酸氢二钠-柠檬酸缓冲液配制不同浓度的没食子酸标准溶液,从40-240μmol/l配制9个不同浓度梯度。分别取0.5ml的没食子酸标准溶液与0.3ml甲醇绕罗丹宁混合,加入到所有试管中,30℃水浴5min,然后加入4.2ml的koh溶液(0.5mol/l),30℃保温10min。以缓冲液代替标准溶液作空白对照,于520nm处测定吸光值(a520)。以没食子酸浓度(mmol/l)为横坐标,a520为纵坐标绘制标准曲线。
测定:反应开始前,将10ml没食子酸丙酯溶液和待测酶液于30℃水浴中保温5-10min。
1)取3支试管,分别为空白管和测试管,向各管中加入0.25ml没食子酸丙酯溶液,然后向测定管中分别加入0.25ml待测酶液,30℃水浴反应5min。
2)向所有试管中加入0.3ml甲醇绕罗丹宁溶液,保温5min。
3)向所有试管中加入4.2ml的koh溶液(0.5mol/l),向空白管中加入0.25ml酶液,30℃保温10min,以空白管归零,于520nm处测定各管溶液吸光值a520。
酶活计算公式:
式中:
x——样品的酶活力,单位为u/ml;
a520——样品空白吸光值之差;
c0——标准曲线的截距;
0.5——没食子酸丙酯加上待测酶液总体积,0.5ml;
n——稀释倍数;
k——标准曲线的斜率;
1/0.25——换算成1ml酶液的酶活;
5——反应时间,5min;
实施例4发酵放大
申请人进一步将出发菌里氏木霉4qt和突变菌里氏木霉4qt1、4qt2、4qt3,分别在20l罐中进行发酵。发酵160h结束时,分别测定发酵上清液中单宁酶酶活。结果显示,出发菌里氏木霉4qt发酵上清液中单宁酶酶活达到101u/ml,而突变菌里氏木霉4qt1,4qt2,4qt3的发酵酶活分别为135u/ml,158u/ml,143u/ml,比出发菌分别提高了33.7%,56.4%,41.6%。
出发菌里氏木霉4qt和突变菌里氏木霉4qt2的发酵进程曲线如图2所示,发酵50h后,本发明所述突变菌里氏木霉4qt2的发酵液酶活开始显著高于出发菌;发酵160h结束时,出发菌里氏木霉4qt发酵上清液中单宁酶活达到101u/ml,而突变菌里氏木霉4qt2发酵上清液中单宁酶酶活高达158u/ml,比出发菌提高了56.4%。
同时,发酵结束后,将出发菌里氏木霉4qt和突变菌里氏木霉4qt2的发酵上清液进行sds-page电泳检测。结果如图3所示,箭头所指65kda处的蛋白条带即为单宁酶,泳道1突变菌里氏木霉4qt2发酵上清液中单宁酶的含量显著高于泳道2出发菌里氏木霉4qt发酵上清液中单宁酶的含量,取得了意料不到的技术效果。
申请人已于2018年6月21日将上述突变菌株里氏木霉4qt2(trichodermareesei4qt2)保藏于中国武汉武汉大学的中国典型培养物保藏中心,保藏编号为cctccno:m2018389。
序列表
<110>青岛蔚蓝生物集团有限公司
<120>一种里氏木霉及其在单宁酶生产中的应用
<160>2
<170>siposequencelisting1.0
<210>1
<211>588
<212>prt
<213>米曲霉(aspergillusoryzae)
<400>1
metargglnhisserargmetalavalalaalaleualaalaglyala
151015
asnalaalaserphethraspvalcysthrvalserasnvallysala
202530
alaleuproalaasnglythrleuleuglyilesermetleuproser
354045
alavalthralaasnproleutyrasnglnseralaglymetglyser
505560
thrthrthrtyrasptyrcysasnvalthrvalalatyrthrhisthr
65707580
glylysglyasplysvalvalilelystyralapheprolysproser
859095
asptyrgluasnargphetyrvalalaglyglyglyglypheserleu
100105110
serseraspalathrglyglyleualatyrglyalavalglyglyala
115120125
thraspalaglytyraspalapheaspasnsertyraspgluvalval
130135140
leutyrglyasnglythrileasntrpaspalathrtyrmetpheala
145150155160
tyrglnalaleuglyglumetthrargileglylystyrilethrlys
165170175
glyphetyrglyglnserseraspserlysvaltyrthrtyrtyrglu
180185190
glycysseraspglyglyarggluglymetserglnvalglnargtrp
195200205
glygluglutyraspglyalailethrglyalaproalapheargphe
210215220
alaglnglnglnvalhishisvalphesersergluvalgluglnthr
225230235240
leuasptyrtyrproproprocysgluleulyslysilevalasnala
245250255
thrilealaalacysaspproleuaspglyargthraspglyvalval
260265270
serargthraspleucyslysleuasnpheasnleuthrserileile
275280285
glygluprotyrtyrcysalaalaglythrserthrserleuglyphe
290295300
glypheserasnglylysargserasnvallysargglnalaglugly
305310315320
serthrthrsertyrglnproalaglnasnglythrvalthralaarg
325330335
glyvalalavalalaglnalailetyraspglyleuhisasnserlys
340345350
glygluargalatyrleusertrpglnilealasergluleuserasp
355360365
alagluthrglutyrasnseraspthrglylystrpgluleuasnile
370375380
proserthrglyglyglutyrvalthrlyspheileglnleuleuasn
385390395400
leuaspasnleuseraspleuasnasnvalthrtyraspthrleuval
405410415
asptrpmetasnthrglymetvalargtyrmetaspserleuglnthr
420425430
thrleuproaspleuthrpropheglnserserglyglylysleuleu
435440445
histyrhisglygluseraspproserileproalaalaserserval
450455460
histyrtrpglnalavalargservalmettyrglyasplysthrglu
465470475480
gluglualaleuglualaleugluasptrptyrglnphetyrleuile
485490495
proglyalaalahiscysglythrasnserleuglnproglyprotyr
500505510
progluasnasnmetgluilemetileasptrpvalgluasnglyasn
515520525
lysproserargleuasnalathrvalserserglythrtyralagly
530535540
gluthrglnmetleucysglntrpprolysargproleutrparggly
545550555560
asnserserpheaspcysvalasnaspglulysserileaspsertrp
565570575
thrtyrglupheproalaphelysvalprovaltyr
580585
<210>2
<211>1767
<212>dna
<213>米曲霉(aspergillusoryzae)
<400>2
atgcgccagcactcccgcatggccgtcgccgctctggctgctggcgctaacgccgcttcc60
ttcaccgacgtctgcaccgtctccaacgtcaaggccgccctgcccgccaacggcactctc120
ctcggcatctccatgctcccctccgccgtcaccgccaaccccctctacaaccagtccgcc180
ggcatgggctccaccaccacctacgactactgcaacgtcaccgtcgcctacacccacacc240
ggcaagggcgacaaggtcgtcatcaagtacgccttccccaagcccagcgactacgagaac300
cgcttctacgtcgccggcggcggcggctttagcctgtcttccgacgccaccggcggcctc360
gcttacggcgctgtcggcggcgctaccgacgctggctacgacgctttcgacaactcctac420
gacgaggtcgtcctctacggcaacggcaccatcaactgggacgccacctacatgttcgcc480
taccaggccctcggcgagatgacccgcatcggcaagtacatcaccaagggcttctacggc540
cagagcagcgacagcaaggtctacacctactacgagggctgctccgacggcggccgcgag600
ggcatgtctcaggtccagcgctggggcgaggagtacgacggcgctatcaccggcgccccc660
gctttccgattcgcccagcagcaggtccaccacgtcttctcctccgaggtcgagcagacc720
ctcgactactacccccccccctgcgagctgaagaagatcgtcaacgccaccatcgccgcc780
tgcgaccccctggatggccgaactgacggcgtcgtctcccgcaccgacctgtgcaagctg840
aacttcaacctcaccagcatcatcggcgagccctactactgcgccgccggcactagcacc900
agcctcggcttcggcttctccaacggcaagcgcagcaacgtcaagcgccaggccgagggc960
agcaccaccagctaccagcccgcccagaacggcaccgtcaccgctcgcggtgtcgccgtc1020
gctcaggctatctacgacggcctccacaacagcaagggcgagcgcgcctacctctcctgg1080
cagatcgcctccgagctgagcgacgccgagaccgagtacaacagcgacaccggcaagtgg1140
gagctgaacatcccctccaccggcggcgagtacgtcaccaagttcatccagctcctgaac1200
ctcgacaacctgagcgacctcaacaacgtcacctacgacaccctggtcgactggatgaac1260
accggcatggtccgctacatggactccctccagaccaccctccccgacctgacccccttc1320
cagtcctccggcggcaagctcctgcactaccacggcgagagcgaccccagcatccccgct1380
gcttcctccgtccactactggcaggccgtccgcagcgtcatgtacggcgacaagaccgag1440
gaggaggccctcgaagccctcgaagactggtatcaattctacctcatccccggcgccgcc1500
cactgcggcaccaacagcctccagcccggcccttaccccgagaacaacatggagatcatg1560
atcgactgggtcgagaacggcaacaagccctcccgcctcaacgccaccgtcagcagcggc1620
acctacgccggcgagacccagatgctctgccagtggcccaagcgccccctgtggcgaggc1680
aactccagcttcgactgcgtcaacgacgagaagtccatcgactcctggacctacgagttc1740
cccgccttcaaggtccccgtctactaa1767
- 还没有人留言评论。精彩留言会获得点赞!