一株无致病力灰葡萄孢菌QT5-19及应用的制作方法
本发明涉及农业微生物技术领域,具体地涉及一株无致病力灰葡萄孢菌qt5-19及应用。
背景技术:
随着我国设施农业的发展,大棚果蔬的栽培面积逐年扩大。由于果蔬大棚中温湿等环境条件适宜,因而由葡萄孢(botrytisspp.)引起的果蔬灰霉病发生呈现逐年上升的趋势,并从传统的次要病害逐渐上升为影响大棚作物产量和质量的主要病害,引起果蔬产量和品质下降,给农民带来严重的经济损失。葡萄孢除侵染果实外,还可侵染花、茎、叶等器官,造成严重减产减收,部分危害严重地区甚至完全绝收。此外,该病害除了在生产过程中侵染植物,还可在运输储藏的果蔬上进行危害,进一步造成经济损失,并成为多种果蔬产后储藏期的主要病害。
目前,灰霉病的防治方法仍以化学防治为主,但葡萄孢对杀菌剂极易产生抗药性,这一方面会加大灰霉病的防治难度;另一方面会提高农民的施药量,从而造成环境污染和危及食品安全。其他的防治方法,例如抗病育种由于尚未发现灰霉病的抗源故难以实施;国外以栽培防治为主的防治措施有些时候并不符合我国实际情况。因此,亟需有效的替代防治措施。近年来,生物防治作为灰霉病一种可能的替代防治措施被广泛研究,并有多种生防菌被用于灰霉病的生防并成功商业化,其中包括木霉菌、粉红粘帚霉和酵母菌等。
灰葡萄孢(botrytiscinerea)一般在马铃薯葡萄糖琼脂(pda)培养基上产生灰白色的菌丝,不能产生色素,培养后期,会在菌落表面形成大量的分生孢子及菌核,在人工接种条件下,一般可以侵染多种植物。本发明提供的灰葡萄孢菌qt5-19在pda上不能产生分生孢子及菌核,对多种植物不能致病或致病力很弱,并且能产生红色色素,此外其产生的挥发性物质具有抑制灰葡萄孢及核盘菌的作用,并有用于生物防治灰霉病的功能。
技术实现要素:
本发明的目的在于提供了一株无致病力的灰葡萄孢菌,所述的菌株为灰葡萄孢(botrytiscinerea)qt5-19,保藏编号为:cctccno:m2018610。
本发明的另一个目的在于提供了灰葡萄孢(botrytiscinerea)qt5-19的应用,包括用于制备成灰霉病菌杀菌剂或熏蒸剂,或者用于核盘菌的杀菌剂或熏蒸剂。
为了达到上述目的,本发明采取以下技术措施:
申请人于2015年从陕西省感染灰霉病的番茄果实上分离获得一株灰葡萄孢菌株qt5-19,该菌株已于2018年9月11日送至中国典型培养物保藏中心保藏,分类命名:灰葡萄孢(botrytiscinerea)qt5-19,保藏编号:cctccno:m2018610,地址:中国武汉武汉大学。
该菌株在pda上的培养物及在麦粒培养基上的发酵物可产生挥发性物质,且该挥发性物质具有抑制植物灰霉病发生的能力。与典型灰葡萄孢菌株相比,在pda培养基上生长速度显著较慢,产生大量气生灰白色菌丝,并在培养基上产生大量的红色色素,使整个菌落呈现红色,无大型分生孢子及黑色菌核产生,灰葡萄孢qt5-19菌株几乎不能在本生烟草叶片上引起病斑,在黄瓜、油菜、草莓、苹果、葡萄及番茄上不能致病或致病力很弱。
灰葡萄孢(botrytiscinerea)qt5-19的应用,包括将本发明提供的灰葡萄孢以唯一有效成分或有效成分之一制备成防治灰霉病的杀菌剂或熏蒸剂;或制备成核盘菌的杀菌剂或熏蒸剂。
一种含有灰葡萄孢(botrytiscinerea)qt5-19的抑菌剂,其通过将灰葡萄孢(botrytiscinerea)qt5-19接种至麦粒中获得。
与现有技术相比,本发明具有以下优点:
1.该菌株源于自然,其产生的挥发性气体对环境影响较人工合成化学农药小;
2.由于是活菌剂,其发酵物可以不断产生挥发物,因而其防效的持续性较一般化学农药久。
附图说明
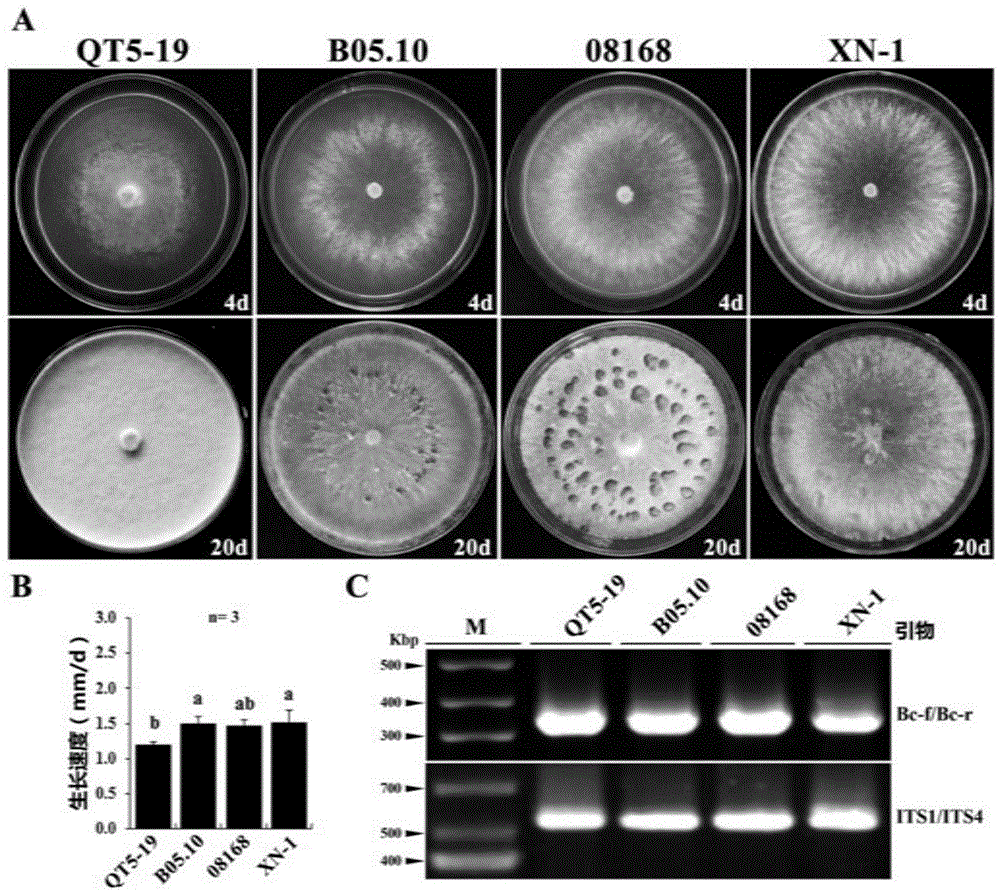
图1为灰葡萄孢菌株qt5-19、b05.10、08168、xn-1在pda培养基上4天和20天的培养结果;
其中,a图显示,与其他典型的灰葡萄孢菌株相比,菌株qt5-19无菌核及分生孢子产生,且菌落呈现红色;
b图显示,与典型的灰葡萄孢菌株相比,菌株qt5-19生长速度较慢,图中标有相同字母表示无显著性差异(p>0.05);
c图显示,四株灰葡萄孢菌均可扩增出灰葡萄孢的特异性条带。
图2为灰葡萄孢菌株qt5-19、b05.10、08168、xn-1在离体本生烟草上的致病情况;
其中,a图显示,与典型的灰葡萄孢菌株相比,灰葡萄孢菌株qt5-19致病力严重丧失,几乎不能在本生烟草叶片上引起病斑;
b图显示,四株灰葡萄孢菌株的病斑直径,图中标有相同字母表示无显著性差异(p>0.05)。
图3为灰葡萄孢菌株qt5-19和b05.10在不同植株上的致病情况示意图;
结果显示,在番茄、葡萄、苹果果实及离体油菜、黄瓜、草莓叶片上的致病情况及平均病斑直径(ld),与其他典型的灰葡萄孢菌株相比,菌株qt5-19致病力严重丧失,在上述植物上均不能致病或致病力很弱。
图4为灰葡萄孢菌株qt5-19菌丝中dsrna的检测示意图;
可以发现该菌株的菌丝中含有两条大小十分接近的dsrna条带,大小比2.0kb略小。
图5为灰葡萄孢菌株qt5-19菌丝中病毒botrytiscinereapartitivirus2的基因组模式图。
图6为病毒botrytiscinereapartitivirus2(bcpv2)的示意图;
其中,a为病毒的粒体形态;
b为病毒粒体中dsrna的电泳检测示意图;
c为病毒粒体衣壳蛋白的sds-page检测示意图。
图7为灰葡萄孢菌株qt5-19产生的挥发性气体对强致病力灰葡萄孢菌rosebc-3的抑制作用;
图7中(a)显示,在小皿中接种菌株qt5-19后会对大皿中的强致病力菌株rosebc-3的生长产生明显抑制;
图7中(b)显示,在小皿中接种菌株qt5-19后,大皿中的rosebc-3菌落直径明显小于两个对照处理中的菌落直径(b);
图7中(c)显示,在小皿中接种菌株qt5-19后,大皿中rosebc-3菌落表面无分生孢子产生。
图8为灰葡萄孢菌株qt5-19在pda上产生的挥发性气体对核盘菌菌株ep-1pna367的抑制作用示意图;
(a)显示,在小皿中接种菌株qt5-19后,均对核盘菌菌株ep-1pna367的菌落直径无明显抑制,但对大皿中的核盘菌菌核的产生有明显抑制;
(b)显示,3个处理均对核盘菌菌株ep-1pna367的菌落直径无明显抑制;
(c)显示,在小皿中接种菌株qt5-19后,大皿中的核盘菌几乎无菌核产生;图中的a367为ep-1pna367的缩写。
图9为灰葡萄孢菌株qt5-19在灭菌的麦粒培养基(awg)上产生的挥发性气体对核盘菌菌株ep-1pna367的抑制作用示意图;
其中,(a)图显示在小皿的awg中接种菌株qt5-19后对大皿中的核盘菌菌株ep-1pna367的生长产生明显抑制;(b)图显示在小皿中接种菌株qt5-19后,ep-1pna367菌落直径明显小于两个对照处理;(c)图显示在小皿中接种菌株qt5-19后,对大皿中的核盘菌菌核的产生有明显抑制,图中的a367为ep-1pna367的缩写。
图10为灰葡萄孢菌株qt5-19在灭菌麦粒培养基上产生的挥发性气体对草莓灰霉病的抑制作用示意图;
其中,(a)为实验设计图;(b)图显示经过密封20℃培养10天,含有菌株qt5-19发酵物处理的草莓果实上灰霉病发病明显减轻,无明显霉层出现;(c)图显示所有三个处理病情指数都明显下降。
具体实施方式
下述实施例中的实验方法,如无特别说明,均为报道的微生物学常规操作方法。所用试剂或材料,如未特别说明,均来源于商业渠道。
实施例1:
灰葡萄孢菌株qt5-19的获得、生物学特性及鉴定:
申请人于2015年从陕西省感染灰霉病的番茄果实上分离获得一株灰葡萄孢菌株qt5-19,该菌株已于2018年9月11日送至中国典型培养物保藏中心保藏,分类命名:灰葡萄孢(botrytiscinerea)qt5-19,保藏编号:cctccno:m2018610,地址:中国武汉武汉大学。
菌株qt5-19的培养形态与一般的灰葡萄孢有显著差异,申请人将其与灰葡萄孢菌株b05.10,08168和xn-1(haoetal.,2018a,b;zhouetal.,2017,2018)进行了对比实验,如下:
该菌株在马铃薯葡萄糖琼脂培养基(pda)平板上与另外三个典型灰葡萄孢菌株相比,生长速度显著较慢(图1中b),于20℃培养20天,产生大量气生灰白色菌丝,并在培养基上产生大量的红色色素,使整个菌落呈现红色,无大型分生孢子及黑色菌核产生(图1中a)。
此外,灰葡萄孢qt5-19菌株几乎不能在本生烟草叶片上引起病斑(图2),在黄瓜、油菜、草莓、苹果、葡萄及番茄上不能致病或致病力很弱(图3)。由于一般灰葡萄孢菌株会产生大量分生孢子和黑色菌核,不产色素,且对多种植物可致病;因此,红色色素、无分生孢子和菌核产生、无致病性为灰葡萄孢qt5-19菌株与其他灰葡萄孢菌株最为显著的区别。
由于该菌在形态上与典型的灰葡萄孢菌有明显区别,因此申请者在分子水平上对该菌株进行了鉴定。一方面,申请者通过its序列的测定及在ncbi网站(https://www.ncbi.nlm.nih.gov)上blast分析,发现与其its序列(登录号:kx822693)与模式灰葡萄孢菌株b05.10的its序列有100%的同源性。另一方面,申请者通过灰葡萄孢特异性引物bc-f/bc-r对其dna进行扩增发现,该菌株可以扩增出于其他灰葡萄孢一样大小的pcr条带(图1中c)。故将该菌株鉴定为灰葡萄孢(botrytiscinerea),并定名为灰葡萄孢(botrytiscinerea)qt5-19。
除上述特征以外,菌株qt5-19还感染了一种真菌病毒。将菌株qt5-19在覆盖有玻璃纸的pda上进行培养,收集菌丝,并进行dsrna检测,申请人可在该菌株的菌丝中检测到两条大小分别为1909bp和1883bp的dsrna条带(图4)。经过基因组测序分析,发现这两条dsrna条带为一种真菌病毒的基因组(图5),并名为botrytiscinereapartitivirus2(bcpv2)。bcpv2在分类上属于分体病毒科(partitiviridae),具有球型的病毒粒体,直径约为36nm(图6中a)。通过琼脂糖凝胶电泳及sds-page电泳分析,发现病毒粒体的衣壳蛋白大小约为68kda(图6中c),包含有病毒的两条dsrna(图6中b),且大小与在菌丝中检测到的大小一致。
该菌株的上述特征,包括其中病毒的感染,在继代培养过程中十分稳定,该菌株从分离至今已经稳定传代50代以上,在中国典型培养物保藏中心保藏的菌株qt5-19为稳定传代50代的菌株。此外,菌株qt5-19中的真菌病毒bcpv2在使用多种化学试剂和物理方法均无法去除,因此该病毒在菌株qt5-19中的存在十分稳定。
实施例2:
灰葡萄孢菌株qt5-19产生的挥发性气体对强致病力灰葡萄孢菌抑制活性的测定
将菌株qt5-19接种在直径为5cm的pda平板上,以不接种菌株的pda平板,和接种灰葡萄孢菌rosebc-3(范雷,2012)的平板为对照,并置于含有pda培养基的大皿(直径15cm)中,于20℃培养2天后,大皿内的培养基(小培养皿外)中接种强致病力的灰葡萄孢菌rosebc-3,于20℃培养20天后,观察结果。
结果表明(图7),两个对照处理(小皿未接菌或者接种同样的rosebc-3)中rosebc-3菌株将占据了整个小培皿的外围pda,其中未接菌的菌落直径为15cm,接种同样的rosebc-3菌落直径为14.4cm,而且产生了大量的分生孢子,其中未接菌的分生孢子产量为1.0×108个/皿,接种同样的rosebc-3分生孢子产量为8.0×107个/皿。与之相比,在小皿接种有菌株qt5-19的处理中,菌株rosebc-3在外围pda上生长受到明显抑制,菌落小,直径仅为5.3cm,且无分生孢子产生,不能占据全部的外围培养基质。这些试验结果表明,灰葡萄孢菌株qt5-19可以通过产生挥发性气体对强致病力灰葡萄孢菌株的生产及产孢产生明显的抑制作用。
实施例3:
灰葡萄孢菌株qt5-19产生的挥发性气体对核盘菌抑制活性的测定
将菌株qt5-19接种在直径为5cm的pda平板上(以不接种菌株的pda平板,和接种核盘菌菌株ep-1pna367(yuetal.,2010)的平板为对照),并置于含有pda培养基的大皿(直径15cm)中,于20℃培养2天后,大皿内的培养基(小培养皿外)中接种核盘菌菌株ep-1pna367,于20℃培养20天后,观察结果。
结果表明(图8中a所示),两个对照处理(小皿未接菌或者接种同样的ep-1pna367)中ep-1pna367菌株将占据了整个小培皿的外围pda,而且产生了大量的菌核。与之相比,在小皿接种有菌株qt5-19的处理中,菌株ep-1pna367虽然占据了外围全部的培养基质(3个处理的菌落平均直径均为15cm),但是菌落不能产生菌核。其中,两个对照处理,即小皿未接菌或者接种同样的ep-1pna367,菌核产量分别为82个/皿和83个/皿,而菌株qt5-19的处理无菌核产生。
为了测试更大量的菌株qt5-19培养物是否会对核盘菌的生长产生抑制,我们将上述装置的小皿中替换为接种有5天的菌株qt5-19的麦粒培养物,对照为未接种的麦粒培养基和接种有核盘菌菌株ep-1pna367的麦粒培养物,大皿内的培养基(小培养皿外)中接种核盘菌菌株ep-1pna367,20℃培养20天后,观察结果。
结果显示(图9),两个对照处理中ep-1pna367菌株将占据了整个小培皿的外围pda,菌落直径为15cm,而且产生了大量的菌核。与之相比,在小皿接种有菌株qt5-19的处理中,菌株ep-1pna367不仅生长受到抑制且未能占领外围全部的培养基质,菌落直径为10.5cm,而且在菌落上无菌核产生。其中,两个对照处理,即小皿的麦粒未接菌或者接种同样的ep-1pna367,菌核产量分别为81个/皿和82个/皿,而菌株qt5-19的处理无菌核产生。
本发明涉及到的麦粒发酵物(或称为麦粒培养物)的制备方法如下:
1.用清水将麦粒漂洗2遍,除去灰尘、麦糠等杂物;
2.将洗净的麦粒用蒸馏水浸泡过夜约12小时;
3.将浸泡过夜的小麦捞出后在煮麦锅内煮沸,水沸腾后再煮20分钟左右,当麦粒达到“无白心,不开花”的标准后及时捞出,沥干多余水分;
4.将沥干水的麦粒装入250ml三角瓶中,每瓶装麦粒100g,将装瓶的麦粒于121℃灭菌20分钟后取出晾凉;
5.向每个装有麦粒的三角瓶接种4块直径约6mm的待测菌株的菌丝块(菌株qt5-19或ep-1pna367),于20℃培养5天后用于生测试验。
以上试验结果表明,灰葡萄孢菌株qt5-19可以通过产生挥发性气体对核盘菌生长及菌核形成有明显的抑制作用,而且这种抑制作用呈现剂量效应,菌株qt5-19的培养物量越多,抑制效果越明显。
实施例4:
灰葡萄孢菌株qt5-19产生的挥发性气体对草莓灰霉病的抑制活性测定:
将干燥器底部分别放置100g,150g和200g的菌株qt5-19的麦粒发酵物,将接种强致病力菌株b05.10的草莓果实置于培养皿中并放置在干燥器的中间隔板上,以未接种菌株qt5-19的麦粒培养基作为对照,每个处理15个草莓果实,将玻璃盖盖好密封后置于20℃培养10天(图10中a)。
结果表明,有菌株qt5-19发酵物处理的草莓果实上灰霉病的病斑都较小,病情指数仅有20,与之相比,对照麦粒培养基质处理中灰霉病的病斑较大,几乎扩展到整个草莓果实,病情指数达到90(图10中b和c)。这些试验结果表明,灰葡萄孢菌株qt5-19可以通过产生挥发性气体抑制储藏期草莓果实上灰霉病的扩展,具有进一步应用于生物防治灰霉病的潜力。
- 还没有人留言评论。精彩留言会获得点赞!