T细胞和/或HLA受体的靶向破坏的制作方法
相关申请的交叉引用
本申请要求2017年6月16日提交的美国临时申请号62/521,132,2017年8月7日提交的美国临时申请62/542,052和2017年10月18日提交的美国临时申请号62/573,956的权益,其公开内容通过引用其全文的方式纳入本文。
本公开内容属于人细胞(包括淋巴细胞和干细胞)的基因组修饰的领域。
背景技术:
基因治疗对于人类治疗学的新时代拥有巨大的潜能。这些方法将实现对于标准医疗实践尚未解决的病症的治疗。基因治疗可包括基因组编辑技术的多种变化形式,例如基因座的破坏(失活)或校正,和/或可表达的转基因的插入,其可通过与转基因操作性连接的特定外源性启动子或通过存在于基因组插入位点处的内源性启动子来控制。
转基因的递送和插入是就该技术的任何实际实施而言必须解决的障碍的实例。例如,尽管许多基因递送方法对于治疗应用有潜在的可行性,但是它们所有都涉及安全性、持久性和表达水平之间大量的权衡。以附加体(例如,腺病毒(ad)、腺相关病毒(aav)和基于质粒的系统)形式提供转基因的方法能实现高初始表达水平,然而,这些方法缺少稳健的附加型复制,而这可能会限制有丝分裂活性组织中表达的持续时间。相比之下,导致所需转基因(例如,整合性慢病毒(lv))随机整合的递送方法提供了更持久的表达,但是由于随机插入的无靶向性质,这可能会在受体细胞中引起无调控的生长,可能会经随机整合的转基因盒附近致癌基因的活化而导致恶性肿瘤。此外,尽管转基因整合避免了复制造成的损失,但是其并不能阻止与转基因融合的外源性启动子的最终沉默。随着时间推移,这样的沉默导致对于大部分非特异性插入事件的转基因表达减少。另外,转基因的整合极少发生在每个靶细胞中,从而可能难以使感兴趣的转基因以足够高水平表达来实现所需疗效。
近年来,已经开发了用于遗传修饰(例如,灭活、校正和/或转基因整合)的新策略,其利用位点特异性核酸酶(例如,锌指核酸酶(zfn),转录激活剂样效应物结构域核酸酶(talen),具有经工程改造的crrna/tracrrna(″单向导rna″)以指导特异性切割的crispr/cas系统等)以在所选基因组基因座进行偏好编辑。参见例如,美国专利号9,937,207、9,255,250、9,045,763、9,005,973、8,956,828、8,945,868、8,703,489、8,586,526、6,534,261、6,599,692、6,503,717、6,689,558、7,067,317、7,262,054、7,888,121、7,972,854、7,914,796、7,951,925、8,110,379、8,409,861、美国专利公开号2017/0211075、2003/0232410、2005/0208489、2005/0026157、2005/0064474、2006/0063231、2008/0159996、2010/00218264、2012/0017290、2011/0265198、2013/0137104、2013/0122591、2013/0177983和2013/0177960以及2015/0056705。此外,开发中的靶向的核酸酶基于阿尔古(argonaute)系统(例如,来自嗜热栖热菌(t.thermophilus),称作`ttago′,参见swarts等(2014)nature507(7491):258-261),其也可具有用于基因组编辑和基因治疗的潜力。与经典的整合方法相比,该核酸酶介导的基因修饰方法提供了改善的转基因表达、增加的安全性和表达持久性的前景,因为其允许准确的转基因定位,从而使基因沉默或附近致癌基因活化的风险最小化。
t细胞受体(tcr)是t细胞选择性活化的必要部分。其具有与抗体的一些相似之处,tcr的抗原识别部分通常由α和β两条链构成,两者共组装形成异二聚体。其与抗体的相似之处在于,编码tcrα和β复合物的单个基因凑在一起。tcrα(tcrα)和tcrβ(tcrβ)链各自包含c-末端恒定区和n-末端可变区两个区域。编码tcrα和β链的基因组基因座与编码抗体的基因座类似,所述编码抗体的基因座中tcrα基因包含v和j区段,而β链基因座除了v和j区段以外还包括d区段。对于tcrβ基因座,在选择过程中还另选择了两个不同的恒定区。在t细胞发育过程中,各种区段重组,从而每个t细胞在α和β链中都包含独特的tcr可变部分,称为互补决定区(cdr),并且身体具有t细胞的大型储库(largerepertoire),它们因具有独特的cdr而能够与抗原呈递细胞展示的独特抗原相互作用。一旦tcrα或β基因重排发生,第二相应tcrα或tcrβ的表达就会被遏制,从而各t细胞在一种被称为″抗原受体等位基因排斥″的过程中仅表达一种独特的tcr结构(参见,brady等,(2010)jimmunol185:3801-3808)。
t细胞活化期间,tcr与抗原呈递细胞的主要组织相容性复合物(mhc)上以肽形式展示的抗原相互作用。通过tcr进行的抗原-mhc复合物识别导致t细胞刺激,这进而导致记忆和效应淋巴细胞中的t辅助细胞(cd4+)和细胞毒性t淋巴细胞(cd8+)的分化。然后,这些细胞可以克隆的方式扩增,以在整个t细胞群中产生能够与一种特定抗原反应的活化亚群。
mhc蛋白具有两种类型--i型和ii型。mhci型蛋白是两个蛋白质:α链和β2微球蛋白链(一些时候也称为b2m)的异二聚体,其中所述α链是由mhc1i类基因编码的跨膜蛋白,而β2微球蛋白链是由不位于mhc基因簇内的基因编码的小胞外蛋白。α链折叠成三个球状结构域,当β2微球蛋白链联合时,该球状结构复合物在细胞表面上呈现功能并表达。肽存在于两个最n末端的结构域上,这两个结构域也是可变性最高的。ii类mhc蛋白也是异二聚体,但这些异二聚体包含由mhc复合物内的基因编码的两个跨膜蛋白。i类mhc:抗原复合物与细胞毒性t细胞相互作用,而ii类mhc将抗原呈递给辅助t细胞。此外,i类mhc蛋白倾向于在几乎所有有核细胞和血小板(以及小鼠中的红细胞)中表达,而ii类mhc蛋白则更选择性地表达。通常,ii类mhc蛋白表达于b细胞,一些巨噬细胞和单核细胞,朗格汉斯细胞和树突细胞。
在人类中,主要组织相容性复合物(mhc)通常被称为人类白细胞抗原(hla)。人的i类hla基因簇包括三个主要基因座,b、c和a,以及几个次要基因座(包括e、g和f,全部存在于染色体6上的hla区域)。ii类hla簇还包括三个主要基因座:dp、dq和dr,并且i类和ii类基因簇都是多态性的,因此在该群体内存在i类和ii类基因的数种不同的等位基因。有几种辅助蛋白也在hla功能中起作用。β-2微球蛋白被用作伴侣(由b2m编码,位于染色体15上),使细胞表面上表达的hlaa、b或c蛋白稳定化,还使i类结构上的抗原展示槽(displaygroove)稳定化。其在血清和尿液中通常含量低。
hla在移植排斥中起主要作用。移植排斥的急性期可能在约1-3周内出现,并且由于宿主系统对供体i类和ii类hla分子的致敏作用,通常涉及宿主t淋巴细胞对供体组织的作用。在大多数情况下,触发抗原是i类hla。为了实现最大成功,供体针对hla进行分型,并且与患病受者尽可能完全匹配。但是,即使在可能共有高度hla相同性的家庭成员之间进行的捐赠也常常无法成功。因此,为了使移植组织留存于受者内,患者通常必须经历彻底的免疫抑制治疗以防止排斥反应。由于患者可能难以克服的机会性感染,此类治疗可导致并发症和高发病率。i类或ii类基因的调节可能会在一些肿瘤存在时被破坏,并且这样的破坏可能会影响患者的预后。例如,在转移性结直肠癌中发现了b2m表达的减少(shrout等(2008)brjcanc98:1999)。因为b2m在稳定mhci类复合物中具有关键作用,在某些实体癌症中,b2m的减少已经被假设为是免疫逃逸避开t细胞驱动的免疫监督的机制。已显示,遏制的b2m表达归因于正常ifnγb2m表达调节的抑制和/或b2m编码序列中导致基因敲除的特定突变(shrout等,同上)。令人困惑的是,b2m升高也与某些类型的癌症相关联。尿液中升高的b2m水平被用作多种癌症(包括前列腺,慢性淋巴细胞性白血病(cll)和非霍奇金淋巴瘤)的预测因素。
过继细胞治疗(act)是一种发展中形式的癌症治疗,其基于将肿瘤特异性免疫细胞递送至患者,以使递送的细胞攻击并清除患者的癌症。act可涉及肿瘤浸润性淋巴细胞(til)的应用,所述til是分离自患者自身的肿瘤肿块并经离体扩增以再灌注回该患者的t细胞。该方法在治疗转移性黑素瘤中具有前景,其中在一项研究中,观测到>50%的长期响应率(参见例如,rosenberg等(2011)clincancres17(13):4550)。til是有希望的细胞来源,因为它们是患者自身的、针对肿瘤上存在的肿瘤相关抗原(tta)具有特异性的t细胞受体(tcr)的细胞的混合群(wu等(2012)cancerj18(2):160)。其它方法涉及编辑分离自患者血液的t细胞,从而它们被工程改造为以一定方式对肿瘤具有响应性(kalos等(2011)scitranslmed3(95):95ra73)。
嵌合抗原受体(car)是被设计成将免疫细胞靶向细胞表面上表达的特定分子靶标的分子。它们最基本的形式是被引入细胞的受体,所述细胞将细胞外侧表达的特异性结构域与细胞内侧的信号转导途径偶联,从而在特异性结构域与其靶标相互作用时,细胞被活化。car常常由仿效t细胞受体(tcr)的功能结构域制得,其中抗原特异性结构域(例如scfv或一些类型的受体)与信号转导结构域(例如itam和其它共刺激结构域)融合。然后将这些构建体离体引入t细胞,以允许该t细胞在表达靶抗原的细胞存在时被活化,致使在t细胞被重新引入患者时,活化的t细胞以非mhc依赖性方式在被靶向的细胞上发起攻击(参见chicaybam等(2011)intrevimmunol30:294-311)。因此,使用由经工程改造的tcr或car离体改变的t细胞进行的过继细胞治疗对于多种类型的疾病而言是非常有前景的临床方法。例如,被靶向的癌症及其抗原包括滤泡性淋巴瘤(cd20或gd2)、成神经细胞瘤(cd171)、非霍奇金淋巴瘤(cd19和cd20)、淋巴瘤(cd19)、成胶质细胞瘤(il13rα2)、慢性淋巴白血病或cll和急性淋巴白血病或all(均为cd19)。还开发了病毒特异性car以攻击具有病毒(如hiv)的细胞。例如,启动了使用对gp100具有特异性的car来治疗hiv的临床试验(chicaybam,同上)。
actr(抗体偶联的t细胞受体)是经工程改造的t细胞组分,其能够结合至外来供给的抗体。抗体与actr组分的结合装配(arm)了t细胞,使之与由抗体识别的抗原相互作用,并且当遇到抗原时,触发包含actr的t细胞以与抗原相互作用(参见美国专利公开号2015/0139943)。
然而,过继细胞治疗的缺点之一是细胞产物的来源必须是患者特异性的(自体同源的),以避免移植细胞的潜在排斥。这已引导研究人员开发编辑患者自身t细胞的方法以避免这样的排斥。例如,患者的t细胞或造血干细胞可通过添加经工程改造的car、actr和/或t细胞受体(tcr)来离体操作,然后进一步使用经工程改造的核酸酶处理,以敲除t细胞检查点抑制剂,例如pd1和/或ctla4(参见,国际专利公开号wo2014/059173)。为了将该技术应用于更大群体的患者,开发细胞的通用群体(同种异体)将会是有利的。此外,tcr的敲除将导致在引入患者后无法引发移植物抗宿主疾病(gvhd)响应的细胞。
因此,仍然需要可用于修饰(例如敲除)效应t细胞,调节性t细胞,b细胞,nk细胞或干细胞(例如,造血干细胞、诱导型多能干细胞和胚胎干细胞)中的tcr和/或hla表达的方法和组合物。
技术实现要素:
本文公开了用于部分或完全灭活或破坏tcr和/或b2m基因的组合物和方法,以及用于在内源性tcr和/或b2m的破坏之后或同时在t淋巴细胞中引入并表达所需水平的外源性转基因的组合物和方法。本文还提供了用于删除(灭活)或遏制tcr和/或b2m基因以产生tcr无效t细胞或tcr和hlai类无效t细胞,b细胞,nk细胞,干细胞,组织或完整生物体(例如,在其表面上不表达一种或多种t细胞受体和/或一种或多种hlai类受体的细胞)的方法和组合物。本文所述的tcr和/或hlai类无效细胞中可能存在其它基因组修饰,包括但不限于,对另一不同基因的基因组修饰(所述基因例如程序性细胞死亡1(pd1)基因,细胞毒性t淋巴细胞抗原4(ctla-4)基因,cish基因,tet2基因,人白细胞抗原(hla)a基因,hlab基因,hlac基因,hla-dpa基因,hla-dq基因,hla-dra基因,lmp7基因,与抗原加工相关联的运输体(tap)1基因,tap2基因,胰蛋白酶基因(tapbp),ii类主要组织相容性复合物反式活化因子(ciita)基因,糖皮质激素受体基因(gr),il2rg基因,rfx5基因),这些或其它基因(例如,安全港基因)中一种或多种中的转基因(例如car)插入,和此类基因组修饰的任何组合。在某些实施方式中,tcr无效细胞(tcrnullcell)和/或hlai类无效细胞(hlaclassinullcell)或组织是有利于用于移植的人细胞或组织。在优选的实施方式中,tcr无效t细胞和/或hlai类无效细胞经制备用于过继t细胞治疗。
一方面,本文描述了锌指核酸酶,其包含:来自命名为68957、72678、72732或72748的zfn的zfp;和工程化的foki切割结构域;和foki切割结构域与zfp之间的接头。在某些实施方式中,zfn包含如下的第一zfn和第二zfn(实施例中公开的氨基酸和多核苷酸序列):包含来自命名为72678的zfn的zfp的zfn,和包含来自命名为72732的zfn的zfp的zfn。在某些实施方式中,zfn包含左和右(第一和第二)zfn,如下:命名为57531的zfn和命名为72732的zfn;命名为57531的zfn和命名为72748的zfn;命名为68957的zfn和命名为57071的zfn;命名为68957的zfn和命名为72732的zfn;命名为68957的zfn和命名为72748的zfn;命名为72678的zfn和命名为57071的zfn;命名为72678的zfn和命名为72732的zfn;和,包含zfp命名为72678的zfn和命名为727482的zfn。锌指核酸酶(zfn)包含左和右(第一和第二)zfn,如下:命名为68796的zfn和命名为68813的zfn;命名为68796的zfn和命名为68861的zfn;命名为68812的zfn和命名为68813的zfn;命名为68876的zfn和命名为68877的zfn;命名为68815的zfn和命名为55266的zfn;命名为68879的zfn和命名为55266的zfn;命名为68798的zfn和命名为68815的zfn;或命名为68846的zfn和命名为53853的zfn。还提供了本文所公开的编码zfn(包括一对)的多核苷酸(例如,mrna,质粒,病毒载体等),包括位于编码左链和zfn的序列之间的包含2a序列的多核苷酸。还公开了包含本文公开的zfn和/或多核苷酸的一种或多种的经遗传修饰的细胞(例如,干细胞,前体细胞,t细胞(效应细胞和调节细胞)等)和源自这些细胞的细胞(例如,不包含zfn但包含遗传修饰的经遗传修饰的细胞)。所述遗传修饰包括被zfn靶向的基因中的插入、缺失及其组合。其它基因组修饰可能存在于所述靶标处和/或一个或多个不同基因座处,所述其它基因组修饰例如t细胞受体(tcr)基因的修饰,hla-a基因的修饰,hla-b基因的修饰,hla-c基因的修饰,tap基因的修饰,ctla-4基因的修饰,pd1基因的修饰,cish基因的修饰,tet-2基因的修饰和/或转基因(例如car)的插入。还提供了包含本文所述的任何锌指核酸酶,多核苷酸和/或细胞的药物组合物。还提供了修饰细胞中内源性β-2-微球蛋白(b2m)和/或tcr基因的方法,该方法包括给予所述细胞本文所述的多核苷酸或药物组合物,从而该内源性基因被修饰(例如,缺失、外源性序列(例如转基因)的插入)。还提供了使用本文所述的zfn,多核苷酸,细胞和/或药物组合物治疗和/或预防癌症,自身免疫性疾病或移植物抗宿主疾病的方法。还提供了包含本文所述的任何zfn,多核苷酸,细胞和/或药物组合物的试剂盒。
在其它方面,本文描述了分离的细胞(例如,真核细胞,例如哺乳动物细胞,包括淋巴样细胞,干细胞(例如,ipsc,胚胎干细胞,msc或hsc)或祖细胞/前体细胞),其中tcr基因的表达通过修饰tcr基因的外显子序列来调节。在某些实施方式中,针对以下序列进行修饰:包含表1、2或6(seqidno:8-21和/或92-103)中一或多项所示的靶位点(如本文中的靶位点所示)的序列的9-25(包括9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25个核苷酸的靶位点)或更多个核苷酸(连续或不连续)的序列的序列;表1、2或6(seqidno:8-21和/或92-103)中所示靶位点任一侧的1-5个碱基对内、1-10个碱基对内或1-20个碱基对内(侧接基因组序列内);或在aacagt、agtgct、ctcct、ttgaaa、tggactt和aatcctc内或包含aacagt、agtgct、ctcct、ttgaaa、tggactt和aatcctc的靶位点内。或者或此外,还可对以下序列进行所述修饰:本文所述的成对靶位点(例如,表3所示的核酸酶对的靶位点)之间的序列(例如,基因组序列),包括,对于55204和53759的靶位点之间(seqidno:8和seqidn0:9之间);对于55229和53785的靶位点之间(seqidno:10和seqidno:11之间);对于53810和55255的靶位点之间(seqidno:12和seqidno:13之间);对于55248和55254/55260所示的靶位点之间(seqidno:14和seqidno:13之间);对于55266和53853的靶位点之间(seqidno:15和seqidno:16之间);对于53860和53863的靶位点之间(seqidno:17和seqidno:18之间);对于53856和55287的靶位点之间(seqidno:21和seqidno:18之间);或对于53885或52774和53909或52742的靶位点之间(seqidno:19和seqidno:20之间)。所述修饰可利用外源性融合分子进行,所述外源性融合分子包含功能结构域(例如,转录调节结构域,核酸酶结构域,包括与野生型相比具有一个或多个突变的任何foki切割结构域)和dna结合结构域,包括但不限于:(i)包含外源性转录因子的细胞,所述外源性转录因子包含与seqidno:8-21和/或92-103中任一所示的靶位点结合的dna结合结构域和其中转录因子修饰trac基因表达的转录调节结构域,和/或(ii)包含在以下位置有插入和/或缺失的细胞:本文所示靶位点(包括seqidno:8-21和/或92-103)的一或多个中;表1和2所示靶位点(seqidno:8-21和/或92-103)任一侧的1-5个碱基对内、1-10个碱基对内或1-20个碱基对内(侧接基因组序列内);aacagt、agtgct、ctcct、ttgaaa、tggactt和aatcctc内;和/或本文所述成对靶位点(例如,表3所示核酸酶对的靶位点)之间。还描述了包含对tcr基因的这些修饰和其它遗传修饰(例如b2m基因修饰,ctla,cish,pd1和/或tet2基因修饰,car,抗原特异性tcr(α和β链),在这些或包括编码抗体偶联的t细胞受体(actr)和/或编码抗体的转基因的其它基因座处的插入等)的细胞。
在另一方面,本文描述了分离的细胞(例如,真核细胞,例如哺乳动物细胞,包括淋巴样细胞,干细胞(例如,ipsc,胚胎干细胞,msc或hsc)或祖细胞/前体细胞),其中b2m基因的表达通过修饰b2m基因来调节。在某些实施方式中,针对以下序列进行修饰:包含表5和8(seqidno:117、123、126和/或127)中一或多者所示的靶位点的序列(如本文中的靶位点所示序列)的9-25(包括9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25个核苷酸的靶位点)或更多个核苷酸(连续或不连续)的序列;表5和8中所示靶位点(seqidno:117、123、126和/或127)任一侧1-5个碱基对内、1-10个碱基对内或1-20个碱基对内(侧接基因组序列内)的序列。或者或此外,还可对以下序列进行修饰:本文所述的成对靶位点(例如,对于表5和8中所示的核酸酶对的靶位点)之间,包括表8中所示的靶位点(seqidno:126和127)之间的序列(例如,基因组序列)。所述修饰可利用外源性融合分子进行,所述外源性融合分子包含功能性结构域(例如,转录调节结构域,核酸酶结构域,其包括与野生型相比具有一个或多个突变的任何foki切割结构域)和dna结合结构域(例如,如表8所示的zfp(命名为72732、72748、68957或72678的zfn的zfp组分(设计物)),包括但不限于:(i)包含外源性转录因子的细胞,所述外源性转录因子包含与表5或8中任一所示的靶位点(例如seqidno:126或127)结合的dna结合结构域和转录调节构域,其中所述转录因子修饰b2m基因表达,和/或(ii)在以下位置包含插入和/或缺失的细胞:本文(包括表5和8)所示靶位点之一或多者内;在任一侧上的1-5个碱基对内、1-10个碱基对内或1-20个碱基对内(侧接基因组序列内);和/或本文所述的成对靶位点(例如,对于表8中所示核酸酶对的靶位点)之间。还描述了包含对b2m基因的这些修饰和其它遗传修饰(例如tcr基因修饰,ctla,cish,pd1和/或tet2基因修饰,pd1修饰,car插入,抗原特异性tcr(α和β链),在这些或包括编码抗体偶联的t细胞受体(actr)和/或编码抗体的转基因的其它基因座处的插入等)的细胞。
本文所述的tcr和/或b2m修饰的细胞可包括其它修饰,例如b2m修饰的细胞中的一个或多个失活的t细胞受体基因,其它失活的tcr基因,pd1和/或ctla4基因和/或编码嵌合抗原受体(car)的转基因,编码抗体偶联的t细胞受体(actr)的转基因和/或编码抗体的转基因。还提供了包括本文所述任何细胞的药物组合物,以及在离体治疗中使用所述细胞或药物组合物用于治疗对象的病症(例如,癌症)的方法。在某些实施方式中,提供了本文所述的包含一个或多个修饰(tcr编辑,b2m编辑,pd1编辑,cish,tet2和/或ctla4编辑,hlai类基因编辑和/或这些或其它基因中的转基因(例如,car)插入)的细胞群,包括其中小于5%(例如0-5%或其间任何值),优选小于3%,甚至更优选小于2%的细胞包含任何其它修饰(例如,脱靶位点的修饰)的细胞群。在某些实施方式中,与用未如本文所述修饰的zfn(其中,未修饰的zfn也称为″亲本″或″亲代″zfn))修饰的细胞相比,所述细胞群在脱靶位点处具有背景水平的修饰(例如,2-10倍(或其间任何值)更少)。通过zfn做出的修饰是可遗传的,其中,在体内或培养中,源自包含所述zfn(和修饰)的细胞的细胞(包括分化的细胞)包括本文所述的修饰。
因此,在一个方面中,本文描述了细胞,其中tcr基因的表达被调节(例如,活化、遏制或失活)。在优选的实施方式中,tcr基因的外显子序列被调节。所述调节可以通过外源性分子(例如,包含dna结合结构域和转录活化或遏制结构域的经工程改造的转录因子)进行,所述外源性分子结合至tcr基因并调节tcr表达,和/或,所述调节可通过tcr基因的序列修饰进行(例如,使用切割tcr基因并通过插入和/或缺失来修饰该基因序列的核酸酶),包括例如表6中所示的zfn(例如左zfn和右zfn的zfn对)。在一些实施方式中,描述了细胞,其包含经工程改造的核酸酶以造成tcr基因的敲除。在其它实施方式中,描述了细胞,其包含经工程改造的转录因子(tf),从而tcr基因的表达被调节。在一些实施方式中,所述细胞是t细胞。还描述了细胞,其中tcr基因的表达被调节,并且其中所述细胞被进一步工程改造以包含至少一种外源性转基因和/或至少一种内源基因(例如,β2微球蛋白(b2m)和/或免疫检查点基因(例如pd1和/或ctla4))的附加敲除,或其组合。
在另一方面中,本文描述细胞,其中的b2m基因的表达被调节(例如,活化、遏制或失活)。所述调节可通过外源性分子(例如,包含dna结合结构域和转录活化或阻遏结构域的经工程改造的转录因子)来进行,所述外源性分子结合至b2m基因并调节b2m表达,和/或,可通过对b2m基因的序列修饰来进行(例如,使用切割b2m基因并通过插入和/或缺失来修饰该基因序列的核酸酶),包括例如,表8中所示的zfn(例如左zfn和右zfn的zfn对)或包含具有本文(例如表8)所述设计物(命名为72732、72748、68957或72678的zfn中的zfp的识别螺旋区和主链)的zfp的zfn,联合任何foki结构域(野生型或经工程改造的),以及任选地,foki结构域和zfp之间的任何接头(例如l0、n7a、n7c等)。在一些实施方式中,描述了细胞,其包含经工程改造的核酸酶以造成b2m基因的敲除。在其它实施方式中,描述了细胞,其包含经工程改造的转录因子(tf),从而b2m基因的表达被调节。在一些实施方式中,细胞是t细胞,包括效应t细胞和调节性t细胞。还描述了细胞,其中b2m基因的表达被调节,并且其中所述细胞被进一步工程改造以包含至少一种外源性转基因和/或至少一种内源基因(例如,一种或多种tcr基因和/或免疫检查点基因(例如pd1和/或ctla4))的附加敲除,或其组合。
在本文所述的包含外源性转基因的任何细胞中,外源性转基因可被整合到tcr和/或b2m基因中(例如,此时tcr和/或b2m基因被敲除)和/或可被整合到例如安全港基因的基因中。在一些情况下,外源性转基因编码actr,抗原特异性tcr和/或car。转基因构建体可通过hdr或nhej驱动的方法插入。在一些方面中,具有调节的tcr和/或b2m表达的细胞至少包含外源性actr,外源性tcr和外源性car。包含tcr调节剂的一些细胞还包含一个或多个检查点抑制剂基因的敲除。在一些实施方式中,检查点抑制剂是pd1。在其它实施方式中,检查点抑制剂是ctla4。在其它方面中,tcr和/或b2m调节的细胞包含pd1敲除和ctla4敲除。在一些实施方式中,调节的tcr基因是编码tcrβ的基因(tcrb)。在一些实施方式中,这通过对该基因的恒定区(tcrβ恒定区,或trbc)进行靶向切割来实现。在某些实施方式中,调节的tcr基因是编码tcrα的基因(tcra)。在其它实施方式中,插入通过对tcr基因的恒定区进行靶向切割来实现,包括对tcrα基因的恒定区(本文称之为″trac″序列)的靶向切割。在一些实施方式中,tcr基因修饰的细胞在b2m基因,hla-a、-b、-c基因或tap基因或其任何组合处被进一步修饰。在其它实施方式中,还修饰对于hlaii类的调节剂,ciita。
在某些实施方式中,本文所述的细胞包含对tcra基因的修饰(例如,缺失和/或插入,经工程改造的tf的结合,以阻遏tcr表达)(例如,外显子的修饰)。在某些实施方式中,修饰在表1、2或6(seqidno:8-21和/或92-103)所示的任何靶位点内和/或在成对的靶位点(例如表3所示的核酸酶对的靶位点)之间,包括通过结合至、切割、插入和/或删除这些序列中任一者内和/或侧接tcra基因中这些序列的基因(基因组)序列的1-50个碱基对(包括其间任何值,例如1-5、1-10或1-20个碱基对)内的一个或多个核苷酸进行的修饰。在某些实施方式中,使用表6中所示的zfn(例如,一个或多个zfn对)进行修饰。在某些实施方式中,细胞在以下序列之一或多者中包含修饰(结合至,切割,插入和/或缺失):tcra基因(例如,外显子,参见图1b)内的aacagt,agtgct,ctcct,ttgaaa,tggactt和aatcctc。在某些实施方式中,修饰包括本文所述的经工程改造的tf的结合,从而tcra基因表达被调节,例如,遏制或活化。
在某些实施方式中,本文所述的细胞包含对b2m基因的修饰(例如,缺失和/或插入,经工程改造的tf的结合,以阻遏b2m表达)。在某些实施方式中,修饰在表5或8所示的任何靶位点内和/或在成对的靶位点(例如表8所示的核酸酶对的靶位点)之间,包括通过结合至、切割、插入和/或删除这些序列中任一者内和/或侧接b2m基因中这些序列的基因(基因组)序列的1-50个碱基对(包括其间任何值,例如1-5、1-10或1-20个碱基对)内的一个或多个核苷酸进行的修饰。在某些实施方式中,使用zfn进行修饰,所述zfn包含zfp,foki结构域(任何野生型或经工程改造的foki结构域)和任选的接头(foki结构域的n末端或c末端,以及所示zfp设计物的n末端或c末端之间的任何接头,包括但不限于l0,n7a,n7c等),所述zfp包括表8所示zfn的zfp设计物的识别螺旋区和主链。在某些实施方式中,zfn包含如表8所示的zfn(例如,第一和第二zfn的对)。在某些实施方式中,细胞在以下序列之一或多者内包含修饰(结合至,切割,插入和/或缺失):seqidno:126和127。在某些实施方式中,修饰包括如本文所述的经工程改造的tf的结合,从而b2m基因表达被调节,例如被遏制或活化。
在其它实施方式中,修饰是位于(一个或多个)核酸酶结合(靶)位点和/或切割位点处或附近的遗传修饰(核苷酸序列改变),包括但不限于,对于所述切割和/或结合位点中一个或多个位点的上游、下游和/或包括所述位点的一个或多个碱基对的1-300个碱基对(或其间任何数量的碱基对)内序列的修饰;包括一个或多个结合和/或切割位点或位于其任一侧的1-100个碱基对(或其间任何数量的碱基对)内的修饰;包括一个或多个结合和/或切割位点和/或位于其任一侧的1-50个碱基对(例如,1-5、1-10、1-20或更多碱基对)内的修饰;和/或,对于核酸酶结合位点和/或切割位点内的一个或多个碱基对的修饰。在某些实施方式中,修饰位于本文公开的任何靶位点周围或之间的基因序列之处或附近(例如,1-300个碱基对、1-50、1-20、1-10或1-5个碱基对,或其间任何数量的碱基对)和/或所述基因序列的成对靶位点(例如表3或8)之间。在某些实施方式中,修饰包括表1、2和6(tcra)和/或表5和8(b2m)的靶位点中所示的一个或多个序列内的tcra和/或b2m基因的修饰,例如对这些序列之一或多者进行的一个或多个碱基对的修饰。在某些实施方式中,核酸酶介导的遗传修饰位于成对的靶位点之间(使用二聚体切割靶标的情况)。核酸酶介导的遗传修饰可以包括任何数量碱基对的插入和/或缺失,包括插入任何长度的非编码序列和/或任何长度的转基因和/或删除1个碱基对至超过1000kb(或其间的任何值,包括但不限于1-100个碱基对、1-50个碱基对、1-30个碱基对、1-20个碱基对、1-10个碱基对或1-5个碱基对)。
本发明的修饰的细胞可以是真核细胞,包括非人类哺乳动物和人类细胞,例如淋巴样细胞(例如,t细胞(包括效应t细胞(teff)和调节性t细胞(treg)),b细胞或nk细胞),干/祖细胞(例如,诱导型多能干细胞(ipsc),胚胎干细胞(例如人es),间充质干细胞(msc)或造血干细胞(hsc)。干细胞可以是全能的或多能的(例如,部分分化的,如hsc,其是多能髓样或淋巴样干细胞)。在其它实施方式中,本发明提供了用于产生tcr和/或hla表达的无效基因型(nullgenotype)的细胞的方法。然后,可以使本文所述的任何修饰的干细胞(在tcra和/或b2m基因座处修饰)分化以产生源自本文所述的干细胞的、具有本文所述的修饰(包括修饰的tcra和/或b2m基因表达)的、分化的(体内或体外(培养))细胞。
在另一方面,本文所述的组合物(经修饰的细胞)和方法可被用于例如治疗或预防或改善病症。所述方法通常包括(a)使用核酸酶(例如,zfn或talen)或核酸酶系统(例如采用工程改造的crrna/tracrrna的crispr/cas)或使用工程改造的转录因子(例如,zfn-tf、tale-tf、cfp1-tf或cas9-tf)在分离的细胞(例如,t细胞或其它淋巴细胞)中切割或下调内源性tcr基因和/或b2m基因,从而使tcr和/或b2m基因失活或下调;和(b)向对象引入所述细胞,由此治疗或预防病症。在一些实施方式中,编码tcrβ(tcrb)的基因失活或下调。在一些实施方式中,编码b2m的基因失活或下调。在一些实施方式中,失活通过对该基因的恒定区(tcrβ恒定区,或trbc)进行靶向切割来实现。在优选的实施方式中,编码tcrα(tcra)和/或b2m的基因失活或下调。在进一步优选的实施方式中,病症是癌症、感染性疾病或自身免疫疾病。在一些实施方式中,进行修饰以诱导免疫耐受。在进一步优选的实施方式中,失活通过对该基因的恒定区(tcrα恒定区,或缩写为trac)进行靶向切割来实现。在一些实施方式中,b2m基因被切割。在其它实施方式中,另外的基因(除tcr和/或b2m以外)被调节(敲除),例如,tcr/b2m双敲除,另外的tcr基因,pd1和/或ctla4和/或一种或多种治疗性转基因存在于细胞中(附加型,随机整合或通过靶向整合(例如核酸酶介导的整合)而整合)。修饰的细胞可包含本文所述的一个或多个zfn(例如,zfn对),包括但不限于,包含第一和第二zfn的锌指核酸酶(zfn),各zfn包含切割结构域(例如,任何野生型或经工程改造的foki切割结构域)和zfpdna结合结构域。在某些实施方式中,使用zfn进行修饰,所述zfn包含本文(例如,表6或表8,其包括命名为68846、53853、72732、72748、68957、55266、68798、68879、68815、68799或72678的zfn的zfp)所述的″设计物″的zfp(识别螺旋区和主链)、foki结构域(任何野生型或经工程改造的foki结构域)和任选的接头(foki结构域的n末端或c末端,和本文所述的zfp设计物的n末端或c末端之间的任何接头)。在一些实施方式中,zfn包含一对zfn,其中一个zfn包含操作性地连接至foki结构域的68846(seqidno:177)的zfp,对中的另一个zfn包含操作性地连接至foki结构域的53853(seqidno:178)的zfp。在一些实施方式中,zfn包含一对zfn,其中一个zfn包含操作性地连接至foki结构域的72732(seqidno:175)的zfp,对中的另一个zfn包含操作性地连接至foki结构域的72678(seqidno:176)的zfp。在某些实施方式中,zfn包含本文所述的zfn(例如,第一或第二(本文中也称作左和右)伴侣zfn的对),如下:命名为68796的zfn和命名为68813的zfn;命名为68796的zfn和命名为68861的zfn;命名为68812的zfn和命名为68813的zfn;命名为68876的zfn和命名为68877的zfn;命名为68815的zfn和命名为55266的zfn;命名为68879的zfn和命名为55266的zfn;命名为68798的zfn和命名为68815的zfn;或命名为68846的zfn和命名为53853的zfn;命名为57531的zfn和命名为72732的zfn;命名为57531的zfn和命名为72748的zfn;命名为68957的zfn和命名为57071的zfn;命名为68957的zfn和命名为72732的zfn;命名为68957的zfn和命名为72748的zfn;命名为72678的zfn和命名为57071的zfn;命名为72678的zfn和命名为72732的zfn;和包含命名为72678的zfn和命名为72748的zfn的zfp。因此,zfn(例如,成对zfn的各zfn伴侣)包含识别螺旋区,并且可包含下文所述的其它zfp修饰(例如,对主链区)(例如,表1、2、5、6和8所示的设计物),并且还包含任何野生型或经工程改造的foki切割结构域(包括foki取代、添加和/或缺失突变体的任何组合)。例如,zfn伴侣可包含与任何foki切割结构域融合的特异性锌指dna结合结构域,所述foki切割结构域包含来自突变序列(如实施例中所示,seqidno:140-174)或来自野生型蛋白的切割结构域(seqidno:139)。b2m特异性zfn伴侣可包含与选自seqidno:139-174的foki切割结构域融合的b2m特异性锌指dna结合结构域(例如72732)。进一步地,b2m特异性zfn伴侣可包含与选自seqidno:139-174的foki切割结构域融合的b2m特异性锌指dna结合结构域(例如72678)。同样,trac特异性zfn伴侣可包含与选自seqidno:139-174的foki切割结构域融合的trac特异性锌指dna结合结构域(例如68846),和,trac特异性锌指dna结合结构域53853可与选自任何所示的野生型或经工程改造的foki切割(例如所附实施例中所示的结构域(seqidno:139-174))的foki切割结构域融合。在一些实施方式中,foki结构域在zfpdna结合结构域的n末端融合,而在其它实施方式中,其与zfpdna结合结构域的c末端融合。此外,任何接头均可用于将dna结合结构域连接至foki切割结构域。
还提供了衍生自按本文所述修饰的细胞(例如,包含本文所述zfn的细胞)的细胞,包括但不限于,部分或完全分化自本文按所述修饰的干细胞的细胞。这些细胞通常不包含所述zfn,但确实包含由其产生的遗传修饰。
一种或多种转录因子和/或一种或多种核酸酶可以mrna形式、以蛋白质形式和/或以编码所述一种或多种核酸酶的dna序列形式引入细胞或周围培养基。在某些实施方式中,引入对象的分离的细胞还包含其它基因组修饰,例如,整合的外源性序列(整合进入切割的tcr和/或b2m基因或另一不同基因,例如安全港基因或基因座)和/或其它基因(例如一个或多个hla基因或ctla-4,cish,pd1或tet2基因)的失活(例如,由核酸酶介导)。外源性序列(例如,car或外源性tcr)或蛋白质可通过载体(例如,ad,aav,lv)或通过使用技术(例如电穿孔或瞬时转染)来引入。在一些实施方式中,蛋白质通过诱导机械应激(例如细胞挤压)而被引入细胞(参见kollmannsperger等.(2016)natcomm7,10372doi:10.1038/ncomms10372)。在一些方面,组合物可包括分离的细胞片段和/或(部分或完全)分化的细胞。
在一些方面,修饰的细胞可以被用于细胞治疗,例如,用于过继细胞转移。在其它实施方式中,用于t细胞移植的细胞包含感兴趣的其它基因修饰。一方面,t细胞包含对癌细胞上存在的标志物具有特异性的插入的嵌合抗原受体(car)。在另一方面,插入的car对b细胞(包括b细胞恶性肿瘤)的cd19标志物特征具有特异性。这样的细胞能够用于治疗组合物以治疗患者,而无需匹配hla,从而因此能够被用作针对任何有需要的患者的″现成的″治疗剂。在其它情况下,在引入之前,扩增包含本文所述修饰的干细胞或前体细胞,例如造血干细胞或前体细胞(hsc/pc)或诱导型多能干细胞(ipsc)。在其它方面,经遗传修饰的hsc/pc在骨髓移植物中给予对象,其中hsc/pc在体内植入、分化和成熟。在一些实施方式中,在g-csf诱导的调动(mobilization)、普乐沙弗(plerixafor)诱导的调动和g-csf与普乐沙弗诱导的调动组合之后从对象分离hsc/pc,并且在其它情况下,细胞从人骨髓或人脐带分离。在其它实施方式中,ipsc源自患者或健康供体细胞。在一些方面,在引入包含衍生自ipsc的经修饰的细胞或经修饰的hsc/pc的植入物之前,使对象经历轻度骨髓清除术操作,而在其它方面,对象用强力骨髓清除调理疗法进行治疗。在一些实施方式中,本发明的方法和组合物用于治疗或预防癌症。
另一方面,tcr和/或b2m调节的(修饰的)t细胞包含插入的抗体偶联的t细胞受体(actr)供体序列。在一些实施方式中,actr供体序列被插入tcr基因中,以在核酸酶诱导的切割之后破坏tcr基因表达。在其它实施方式中,供体序列被插入″安全港″基因座中,例如aavs1、hprt、白蛋白和ccr5基因。在一些实施方式中,actr序列通过靶向整合插入,其中actr供体序列包括侧接同源臂,与侧接经工程改造的核酸酶的切割位点的序列具有同源性。在一些实施方式中,actr供体序列还包括启动子和/或其它转录调节序列。在其它实施方式中,actr供体序列缺少启动子。在一些实施方式中,actr供体被插入tcrβ编码基因(tcrb)。在一些实施方式中,插入通过对该基因的恒定区(tcrβ恒定区,或trbc)进行靶向切割来实现。在优选的实施方式中,actr供体被插入tcrα编码基因(tcra)。在进一步优选的实施方式中,插入通过对该基因的恒定区(tcrα恒定区,缩写为trac)进行靶向切割来实现。在一些实施方式中,供体被插入tcra中的外显子序列,然而在其它实施方式中,供体被插入tcra中的内含子序列。在其它实施方式中,actr供体被插入b2m基因中。在一些实施方式中,b2m和/或tcr调节型细胞还包含car。在其它实施方式中,b2m和/或tcr调节型细胞还在hla基因或检查点抑制剂基因处被调节。
还提供了包含如本文所述的经修饰的细胞(例如,具有失活tcr基因的t细胞或干细胞)的药物组合物,或包含本文所述的一种或多种tcr和/或b2m基因结合分子(例如,经工程改造的转录因子和/或核酸酶)的药物组合物。在某些实施方式中,药物组合物还可包含一种或多种药学上可接受的赋形剂。经修饰的细胞,tcr和/或b2m基因结合分子(或编码这些分子的多核苷酸)和/或包含这些细胞或分子的药物组合物通过本领域已知的方法,例如通过静脉内输注、输注进入特定血管(例如肝动脉),或通过直接组织注射(例如,肌肉)来引入对象。在一些实施方式中,对象是患有可采用该组合物治疗或缓解的疾病或病症的成人。在其它实施方式中,对象是给予该组合物以预防、治疗或缓解疾病和病症(例如,癌症、移植物抗宿主疾病等)的儿科对象。
在一些方面,组合物(包含actr的tcr和/或b2m调节型细胞)还包含外源性抗体。还参见美国专利公开号2017/0196992。在一些方面,抗体可用于装配包含actr的t细胞,以预防或治疗病症。在一些实施方式中,抗体识别与肿瘤细胞或与癌症相关过程相关联的抗原,如epcam、cea、gpa33、粘蛋白、tag-72、caix、psma、叶酸-结合抗体、cd19、egfr、erbb2、erbb3、met、igf1r、epha3、trailr1、trailr2、rankl、fap、vegf、vegfr、αvβ3和α5β1整合素、cd20、cd30、cd33、cd52、ctla4和细胞粘合素(enascin)(scott等(2012)natreycancer12:278)。在其它实施方式中,抗体识别与感染性疾病(如hiv、hcv等)相关联的抗原。
在另一方面,本文提供了结合至tcr基因中的靶位点的tcr基因dna结合结构域(例如,zfp、tale和sgrna)。在某些实施方式中,dna结合结构域包含:zfp,其具有表1单行所示顺序的识别螺旋区;tal-效应物结构域dna-结合蛋白,其具有与表1第1列或表2第3列所示靶位点相结合的rvd;和/或表2单行所示的sgrna。这些dna结合蛋白可与转录调节结构域相关联,以形成经工程改造的转录因子,其调控tcr表达。或者,这些dna结合蛋白可与一个或多个核酸酶结构域结合,以形成经工程改造的锌指核酸酶(zfn)、talen和/或crispr/cas系统,其结合并切割tcr基因。在某些实施方式中,zfn、talen或crispr/cas系统的单向导rna(sgrna)结合人tcr基因中的靶位点。转录因子或核酸酶的dna结合结构域(例如zfp,tale,sgrna)可结合至tcra基因中的靶位点,其包含本文所示的任何靶位点(例如seqidno:8-21和/或92-103所示的表1或2的靶位点)的9、10、11、12或更多个(例如13、14、15、16、17、18、19、20或更多个)核苷酸。锌指蛋白可包括1、2、3、4、5、6或更多个锌指,各锌指具有识别螺旋,其与靶基因中的亚靶位点(targetsubsite)特异性地接触。在某些实施方式中,锌指蛋白包括4或5或6个指(将其命名为f1、f2、f3、f4、f5和f6,并且从n末端至c末端排列f1至f4或f5或f6),例如表1所示。本文所述的zfp还可包括锌指蛋白的磷酸接触残基(phosphatecontactresidue)的一个或多个突变,例如,美国专利公开号2018/0087072中描述的nr-5qabc突变体。在其它实施方式中,单向导rna或tal-效应物dna结合结构域可结合至本文所述的靶位点(例如,seqidno:8-21和/或92-103所示的表1或表2或表6的靶位点)或任何这些靶位点内或成对靶位点之间的12或更多个碱基对。示例性sgrna靶位点示于表2(seqidno:92-103)。还提供了与表1或表2所示靶位点的12或更多个核苷酸结合的sgrna。如美国专利号8,586,526和9,458,205所述,可以采用规范或非规范的rvd将talen设计成靶向本文所述的位点(表1或表2或表6的靶位点)。本文所述的核酸酶(包括zfp、tale或sgrnadna-结合结构域)能够在包括seqidno:8-21和/或92-103中任一者的tcra基因内产生遗传修饰,包括在这些序列(seqidno:8-21和/或92-103)中任一者内的修饰(插入和/或缺失)和/或对侧接seqidno:8-21和/或92-103所示靶位点序列的tcra基因序列进行的修饰,例如,以下序列之一或多种内的tcr基因的外显子序列内的修饰:aacagt、agtgct、ctcct、ttgaaa、tggactt和aatcctc。
在另一方面,本文提供了b2m基因dna-结合结构域(例如,zfp、tale和sgrna),其结合至b2m基因中的靶位点。在某些实施方式中,dna结合结构域包含:zfp,其具有识别螺旋区,顺序如表5或表8单行所示(标记为″设计物″的列,包括命名为72732、72748、68957或72678的的zfn的zfp);tal效应子结构域dna结合蛋白,其具有与表5或表8的第一列所示靶位点结合的rvd;和/或sgrna,其与本文(表5或表8)所述的b2m靶位点结合。这些dna-结合蛋白可与转录调节结构域相关联,以形成调节b2m表达的经工程改造的转录因子。或者,这些dna结合蛋白可与一个或多个核酸酶(切割)结构域结合,以形成经工程改造的锌指核酸酶(zfn)、talen和/或crispr/cas系统,其结合并切割b2m基因。在某些实施方式中,zfn、talen或crispr/cas系统的单向导rna(sgrna)结合人b2m基因中的靶位点。转录因子或核酸酶的dna结合结构域(例如zfp,tale,sgrna)可结合至b2m基因中的靶位点,其包含本文所示的任何靶位点(例如表5或表8,如seqidno:117、123、126或127所示)的9、10、11、12或更多个(例如13、14、15、16、17、18、19、20或更多个)核苷酸。锌指蛋白可包括1、2、3、4、5、6或更多个锌指,各锌指具有识别螺旋,其与靶基因中的亚靶位点(targetsubsite)特异性地接触。在某些实施方式中,锌指蛋白包括4或5或6个指(将其命名为f1、f2、f3、f4、f5和f6,并且从n末端至c末端排列f1至f4或f5或f6),例如表5或表8所示。本文所述的zfp还可包括锌指蛋白的磷酸接触残基的一个或多个突变,例如,美国专利公开号2018/0087072中描述的nr-5qabc突变体,包括表8的zfp设计物(识别螺旋区和主链突变体)。在其它实施方式中,单向导rna或tal-效应物dna结合结构域可结合至本文所述的靶位点(例如,表5或8的靶位点)或这些靶位点中任一者内或成对靶位点之间的12或更多个碱基对。如美国专利号8,586,526和9,458,205所述,可以采用规范或非规范的rvd将tale结构域设计成靶向本文所述的位点(表5或表8的靶位点)。本文所述的核酸酶(包含zfp、tale或sgrnadna结合结构域)能够在包含本文公开的任何b2m靶位点的b2m基因内产生遗传修饰,包括在这些序列中任一者内产生的修饰(插入和/或缺失)和/或对侧接表5和表8中所示靶位点序列(seqidno:117、123、126或127)的b2m基因序列进行的修饰。
本文所述的任何核酸酶均可包含本文所述的dna结合结构域(例如表6或8的zfp设计物,tale或sgrna)和切割结构域和/或切割半结构域(例如,野生型或经工程改造的foki切割半结构域)。因此,在本文所述的任何核酸酶(例如,zfn,talen,crispr/cas系统)中,核酸酶结构域可包含野生型核酸酶结构域或核酸酶半结构域(例如,foki切割半结构域)。在其它实施方式中,核酸酶(例如,zfn,talen,crispr/cas核酸酶)包含经工程改造的核酸酶结构域或半结构域,例如形成专性异二聚体的经工程改造的foki切割半结构域。参见例如美国专利号7,914,796和8,034,598。在某些实施方式中,本文描述的核酸酶的一个或多个foki核酸内切酶结构域还可包含如美国专利公开号2018/0087072中所述的磷酸接触突变体(例如r416s和/或k525s)。因此,本文所述的foki结构域(例如,包含以下部分的zfn:(i)表8所示的zfp设计物,包括命名为72732、72748、68957或72678的zfn的zfp;和(ii)foki结构域)可包含foki结构域(相对于全长foki编号的位置)的突变的任何结合,包括野生型foki催化结构域序列,还包括但不限于,表8所示的foki结构域,foki-sharkey(s418p+k441e);fokield(486位q->e,499位i->l,496位n->d);fokield,sharkey(486位q->e,499位i->l,496位n->d,s418p+k441e);fokield,r416e(486位q->e,499位i->l,496位n->d,r416e);fokield,sharkey,r416e(486位q->e,499位i->l,496位n->d,s418p+k441e,r416e);fokield,r416y(486位q->e,499位i->l,496位n->d,r416y);fokield,sharkey,r416e(486位q->e,499位i->l,496位n->d,s418p+k441e,r416e);fokield,s418e(486位q->e,499位i->l,496位n->d,s418e);fokield,sharkey部分,s418e(486位q->e,499位i->l,496位n->d,k441e,s418e);fokield,k525s(486位q->e,499位i->l,496位n->d,k525s);fokield,sharkeyk525s(486位q->e,499位i->l,496位n->d,s418p+k441e,k525s);fokield,i479t(486位q->e,499位i->l,496位n->d,i479t);fokield,sharkey,i479t(486位q->e,499位i->l,496位n->d,s418p+k441e,i479t);fokield,p478d(486位q->e,499位i->l,496位n->d,p478d);fokield,sharkey,p478d(486位q->e,499位i->l,496位n->d,s418p+k441e,p478d);fokield,q481d(486位q->e,499位i->l,496位n->d,q481d);fokield,sharkey,q481d(486位q->e,499位i->l,496位n->d,s418p+k441e,q481d);fokikkr(490位e->k,538位i->k,537位h->r);fokikkrsharkey,(490位e->k,538位i->k,537位h->r,s418p+k441e);fokikkr,q481e(490位e->k,538位i->k,537位h->r,q481e);fokikkr,sharkeyq481e(490位e->k,538位i->k,537位h->r,s418p+k441e,q481e);fokikkr,r416e(490位e->k,538位i->k,537位h->r,r416e);fokikkr,sharkey,r416e(490位e->k,538位i->k,537位h->r,s418p+k441e,r416e);fokikkr,k525s(490位e->k,538位i->k,537位h->r,k525s);fokikkr,sharkey,k525s(490位e->k,538位i->k,537位h->r,s418p+k441e,k525s);fokikkr,r416y(490位e->k,538位i->k,537位h->r,r416y);fokikkr,sharkey,r416y(490位e->k,538位i->k,537位h->r,s418p+k441e,r416y);foki,kkri479t(490位e->k,538位i->k,537位h->r,i479t);foki,kkrsharkeyi479t(490位e->k,538位i->k,537位h->r,s418p+k441e,i479t;foki,kkrp478d(490位e->k,538位i->k,537位h->r,p478d),fokikkrsharkeyp478d(490位e->k,538位i->k,537位h->r,p478d);fokidad(487位r->d,496位n->d,499位i->a);fokidadsharkey(487位r->d,496位n->d,499位i->a,s418p+k441e);fokirvr(483位d->r,537位h->r,538位i->v);fokirvrsharkey(483位d->r,537位h->r,538位i->v,s418p+k441e)。本文所述的zfn还可包括任何接头序列,包括但不限于美国专利号7,888,121、7,914,796、8,034,598、8,623,618、9,567,609和美国公开号2017/0218349中公开的序列,其可用于dna结合结构域(例如zfp)的n末端或c末端和foki切割结构域的n末端或c末端之间。
在另一个方面,本公开提供了编码本文所述的任何蛋白质、融合分子和/或其组分(例如,sgrna或其它dna结合结构域)的多核苷酸。多核苷酸可以是病毒载体、非病毒载体(例如,质粒)的部分,或是mrna形式。本文所述的任何多核苷酸也可包含用于靶向插入tcrα和/或tcrβ基因的序列(供体,同源臂或补丁序列)。而在另一个方面中,提供包含本文所述的任何多核苷酸的基因递送载体。在某些实施方式中,载体是腺病毒载体(例如,ad5/f35载体),或慢病毒载体(lv),包括有整合能力的或整合缺陷型慢病毒载体,或腺相关的载体(aav)。因此,本文还提供了病毒载体,其包括编码核酸酶(例如,zfn或talen)和/或核酸酶系统(crispr/cas或ttago)的序列和/或用于靶基因内的靶向整合的供体序列。在一些实施方式中,供体序列和编码核酸酶的序列位于不同的载体。在其它实施方式中,核酸酶以多肽形式提供。在优选的实施方式中,多核苷酸是mrna。在一些方面中,该mrna可经化学修饰(参见例如kormann等,(2011)naturebiotechhology29(2):154-157)。在其它方面中,该mrna可包含arca帽(参见美国专利号7,074,596和8,153,773)。在一些方面,mrna可包括通过酶促修饰引入的帽。酶促引入的帽可包括:cap0、cap1或cap2(参见例如smietanski等,(2014)naturecommunications5:3004)。在其它方面,mrna可通过化学修饰封端。在其它实施方式中,mrna可包含未经修饰和经修饰的核苷酸的混合物(参见美国专利公开号2012/0195936)。在另一个实施方式中,mrna可包含wpre元件(参见美国专利公开号2016/0326548)。在一些实施方式中,mrna是双链的(参见例如,kariko等.(2011)nuclacidres39:e142)。
而在另一方面,本公开提供了分离的细胞,其包含本文所述的任何蛋白质、多核苷酸和/或载体。在某些实施方式中,细胞选自下组:干/祖细胞或t细胞(例如有效或调节性t细胞)。在又一方面,本公开提供了一种细胞或细胞系,其衍生自包含本文所述的任何核酸酶、转录因子、多核苷酸和/或载体的细胞或细胞系,即,衍生自(培养中)其中tcr和/或b2m已经由一种或多种zfn失活和/或其中供体多核苷酸(例如actr和/或car)已被稳定整合到细胞的基因组中的细胞的细胞或细胞系。因此,本文所述细胞的衍生后代本身可能不包含本文所述分子,多核苷酸和/或载体,但在这些细胞中,tcr和/或b2m基因失活和/或供体多核苷酸整合到基因组中且/或表达。
在另一方面,本文描述了使细胞中的tcr和/或b2m基因失活的方法,所述方法通过将一种或多种蛋白质,多核苷酸和/或载体引入本文描述的细胞中来进行。在某些实施方式中,采用表6中所示的编码zfn(例如,zfn对)的一种或多种多核苷酸来修饰细胞中的tcr基因,而从这些细胞衍生的细胞(包括分化的细胞)包含所述修饰。在其它实施方式中,采用表8所示的编码zfn(例如,zfn对)的一种或多种多核苷酸来修饰细胞中的b2m基因,而从这些细胞衍生的细胞(包括分化的细胞)包含所述修饰。在本文所述任何方法中,核酸酶可诱导定向诱变,细胞dna序列的缺失,和/或促进预定染色体基因座处的靶向重组。因此,在某些实施方式中,核酸酶将一个或多个核苷酸从靶基因删除和/或将其插入靶基因。在一些实施方式中,tcr和/或b2m基因通过核酸酶切割随后非同源末端连接而失活。在其它实施方式中,靶基因(例如,tcr或b2m)中的基因组序列被替代,例如,采用本文所述的核酸酶(或编码所述核酸酶的载体)和在用所述核酸酶进行靶向切割之后插入该基因的″供体″序列。供体序列可存在于核酸酶载体中,也可存在于分开的载体中(例如,质粒,线性单链或双链dna,aav,ad或lv载体),或者可以使用不同的核酸递送机制引入细胞。在一些实施方式中,所述方法还包括使一种或多种其它基因(例如,b2m)失活和/或将一种或多种转基因整合进入细胞的基因组,包括但不限于,将一种或多种转基因整合进入失活的tcr和/或b2m基因和/或整合进入一个或多个安全港基因。在某些实施方式中,本文描述的方法产生细胞群,其中至少80-100%(或其间任何值),包括至少90-100%(或其间任何值)的细胞包括敲除和/或整合的转基因。
此外,本文所述的任何方法可体外、体内和/或离体实施。在某些实施方式中,所述方法离体实施,例如,以修饰t细胞(效应t细胞或调节性t细胞),以使其在同种异体环境中可用作治疗对象(例如患有癌症或自身免疫性疾病的对象)的治疗剂。可被治疗和/或预防的癌症的非限制示例包括:肺癌、胰腺癌、肝癌、骨癌、乳腺癌、结肠直肠癌、白血病、卵巢癌、淋巴瘤、脑癌等。自身免疫性疾病的非限制性示例包括移植排斥,1型糖尿病,肠易激疾病/病症,多发性硬化症,狼疮,硬皮病,类风湿性关节炎等。所述细胞也可用于诱导免疫耐受。
在另一方面,本文描述了将一种或多种转基因整合进入分离细胞的基因组的方法,该方法包括:向细胞中引入,(a)一种或多种供体载体(例如,质粒,线性单链或双链dna,aav,质粒,ad,mrna等),所述供体载体包含所述一种或多种转基因,和(b)以mrna形式存在的至少一种非天然产生的核酸酶,其中至少一种核酸酶切割所述细胞的基因组,从而所述一种或多种转基因被整合进入所述细胞的基因组(例如,整合进入tcr受体),其中所述供体载体在紧邻核酸酶电穿孔进入细胞之前或之后被引入包含所述分离细胞和所述mrna的电穿孔缓冲液。在某些实施方式中,在电穿孔之后且在将细胞转移至培养基之前将供体载体引入电穿孔缓冲液中。参见例如,美国专利公开号2015/0174169和2015/0110762。所述方法可用于将一种或多种转基因引入任何基因组位置,包括但不限于,tcr基因,b2m基因和/或安全港基因(例如,aavs1,rosa,白蛋白,ccr5,cxcr4等)。
附图说明
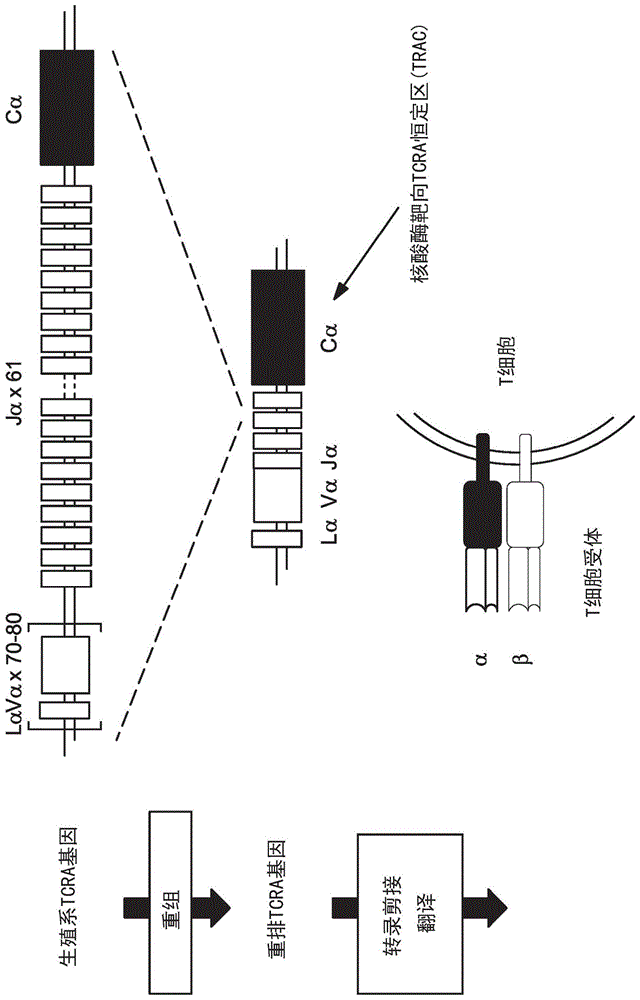
图1a和1b是tcra基因的示意图,其显示通过核酸酶靶向的位点的位置。图1a说明从种系形式到成熟t细胞形式的tcra基因加工,并且指示了核酸酶的一般靶标。图1b(seqidno:116(外显子c1)、117(外显子c2)和118(外显子c3))显示恒定区序列中靶位点之间的区域。以大写黑色字体显示的序列是指示的外显子序列的序列,而以小写灰色字体显示的序列是邻近的内含子序列。
图2a和2b的图描述t细胞中修饰的各位点百分比,所述t细胞用对tcra位点a、b和d(图2a)和位点e、f和g(图2b)具有特异性的zfn处理。许多对实现了80%或更高的修饰比例。
图3描述在用tcra特异性zfn对处理后,通过facs分析法所分析的cd3阴性t细胞的百分比。
图4的图表显示了在t细胞中经由高通量测序测量的tcra序列修饰水平和通过荧光活化细胞分选法测量的cd3表达的损失之间的高度相关性。
图5a至5d的图表描述了用根据tcra基因中靶位点分组的tcra特异性zfn处理后t细胞的生长。
图6显示来自trac(tcra)和b2m双敲除以及将供体靶向整合进入trac(tcra)或b2m基因座中的结果。
图7显示了来自trac(tcra)和b2m双敲除以及将供体靶向整合进入trac(tcra)或b2m基因座中的facs结果。显示了指示条件下的facs结果(上图从左到右:对照(假);无供体的trac和b2mzfn;有靶向至b2m的供体的trac和b2mzfn;有靶向至trac的供体的trac和b2mzfn)。fac图首行的左下象限显示具有双(trac/b2m)敲除的细胞,fac图底行的右半部分显示具有双敲除和靶向整合的细胞。细胞百分比也由指向fac图的合适区块的箭头指示。如箭头所示,85-90%或更多的细胞为双ko,并且对于靶向整合也是阳性的。
具体说明
本文公开了用于产生细胞的组合物和方法,其中tcr基因的表达被调节,从而所述细胞的细胞表面上不再包含tcr,和/或其中b2m基因的表达被调节,从而所述细胞不再表达b2m。以这种方式修饰的细胞可用作治疗剂,例如移植物,因为tcr复合物的缺少会阻止或减少基于hla的免疫反应。另外,可以将其它感兴趣的基因(例如,转基因)插入其中tcr和/或b2m基因已被操作的细胞中。可以通过敲除和/或靶向插入外源性序列来修饰一个或多个其它(非tcr和/或b2m)基因(例如其它tcr,b2m,pd1,ctla4,hla基因,安全港基因等)。外源性序列可包括用于整合进入经修饰细胞的嵌合抗原受体,其可用于治疗癌症和自身免疫疾病。
概述
除非另有说明,本方法的实施以及本文所述组合物的制备与应用采用本领域技术范围内的分子生物学、生物化学、染色质结构与分析、计算化学、细胞培养、重组dna与相关领域的常规技术。这些技术在文献中已有充分描述。参见例如,sambrook等,《分子克隆:实验室手册》(molecularcloning:alaboratorymanual)第2版,冷泉港实验室出版社(coldspringharborlaboratorypress),1989以及第3版,2001;ausubel等,《新编分子生物学实验指南》(currentprotocolsinmolecularbiology),约翰韦利父子公司(johnwiley&sons),纽约,1987及定期更新;《酶学方法》(methodsinenzymology)丛书,学术出版社(academicpress),圣迭戈;wolffe,《染色质结构与功能》(chromatinstructureandfunction),第3版,学术出版社,圣迭戈,1998;《酶学方法》(methodsinenzymology),第304卷,″染色质(chromatin)″(p.m.wassarman和a.p.wolffe编),学术出版社,圣迭戈,1999;和《分子生物学方法》(methodsinmolecularbiology),第119卷,″染色质方案(chromatinprotocols)″(p.b.becker编),休玛纳出版社(humanapress),托托瓦,1999。
定义
术语″核酸″,″多核苷酸″和″寡核苷酸″互换使用并指脱氧核糖核苷酸或核糖核甘酸聚合物,可以是直链或环状构型的,是单链或双链形式的。出于本公开目的,这些术语不意在限制聚合物的长度。所述术语可涵盖天然核苷酸的已知类似物,以及在碱基、糖和/或磷酸部分(例如,硫逐磷酸酯主链)经修饰的核苷酸。一般而言,具体核苷酸的类似物具有相同的碱基配对特异性;即,a的类似物将与t进行碱基配对。
术语〝多肽″、〝肽″和〝蛋白质″在本文中可互换使用,指氨基酸残基的聚合物。该术语还应用于其中一种或多种氨基酸是对应的天然产生的氨基酸的化学类似物或经修饰的衍生物的氨基酸聚合物。
″结合″指大分子之间(例如蛋白质与核酸之间)的序列特异性、非共价相互作用。只要相互作用作为整体为序列特异性,不要求结合相互作用的所有组分都是序列特异性(例如,与dna主链中的磷酸残基接触)。这样的相互作用通常表征为解离常数(kd)是10-6m-1或更低。″亲和性″指结合的强度:增加的结合亲和性与较低的kd关联。″非特异性结合″指不依赖于靶序列的发生在大分子(例如,dna)和任何感兴趣的分子(例如,经工程改造的核酸酶)之间的非共价相互作用。
″dna结合分子″是能够结合dna的分子。这样的dna结合分子可以是多肽,蛋白质的结构域,较大蛋白质内的结构域,或多核苷酸。在一些实施方式中,多核苷酸是dna,而在其它实施方式中,多核苷酸是rna。在一些实施方式中,dna结合分子是核酸酶的蛋白质结构域(例如,foki结构域),而在其它实施方式中,dna结合分子是rna引导的核酸酶的向导rna组分(例如,cas9或cfp1)。
″结合蛋白″是能与另一分子非共价结合的蛋白质。结合蛋白能够结合至,例如,dna分子(dna结合蛋白)、rna分子(rna结合蛋白)和/或蛋白质分子(蛋白质结合蛋白)。在蛋白质结合蛋白的情况中,其可与自身结合(形成同型二聚体、同型三聚体等)和/或其可与一种或多种不同蛋白的一个或多个分子结合。结合蛋白可具有多于一种类型的结合活性。例如,锌指蛋白具有dna结合、rna结合和蛋白结合活性。
″锌指dna结合蛋白″(或结合结构域)是能以序列特异性方式通过一个或多个锌指结合dna的蛋白质或较大蛋白质内的结构域,锌指是通过锌离子配位稳定其结构的结合结构域内氨基酸序列的区域。因此,多指zfp的各锌指都包含识别螺旋区,用于与主链内的dna结合。术语锌指dna结合蛋白常缩写为锌指蛋白或zfp。术语″锌指核酸酶″包括一个zfn以及一对zfn(该对成员称为″左和右″或〝第一和第二″或″对″),其二聚化以切割靶基因。
″taledna结合结构域″或〝tale″是包含一种或多种tale重复结构域/单元的多肽。各自包含重复可变双残基(rvd)的重复结构域参与tale与其关联靶dna序列的结合。单个″重复单元″(也称作″重复″)通常长33-35个氨基酸,并且与天然产生的tale蛋白中的其它tale重复序列显示至少一些序列同源性。tale蛋白可经设计以利用重复单元内典型或非典型rvd来结合靶位点。参见例如,美国专利号8,586,526和9,458,205。锌指和taledna结合结构域可以″经工程改造″以结合至预定的核苷酸序列,例如,通过对天然产生的锌指蛋白的识别螺旋区进行工程改造(改变一个或多个氨基酸)或通过对参与dna结合的氨基酸(重复可变双残基或rvd区)进行工程改造。因此,经工程改造的锌指蛋白或tale蛋白是非天然产生的蛋白质。用于工程改造锌指蛋白和tale的方法的非限制性示例是设计和选择。经设计的蛋白质是非天然产生的蛋白质,其设计/组成主要来自于合理标准。设计的合理标准包括应用替换规则和计算机算法来处理储存现有zfp或tale设计物(规范和非规范rvd)信息和结合数据的数据库中的信息。参见例如,美国专利号9,458,205、8,586,526、6,140,081、6,453,242和6,534,261;还参见国际专利公开号wo98/53058、wo98/53059、wo98/53060、wo02/016536和wo03/016496。术语″talen″包括一个talen以及一对talen(所述对的成员称为″左和右″或″第一和第二″或〝对″),其二聚化以切割靶基因。
〝选择的″锌指蛋白、tale蛋白或crispr/cas系统不存在于自然界中,其产生的结果主要来自经验过程,例如噬菌体展示、相互作用阱或杂交选择。参见例如,美国专利号5,789,538、5,925,523、6,007,988、6,013,453、6,200,759和国际专利公开号wo95/19431、wo96/06166、wo98/53057、wo98/54311、wo00/27878、wo01/60970、wo01/88197和wo02/099084。
″ttago″是原核阿尔古(argonaute)蛋白质,认为其参与基因沉默。ttago源自嗜热细菌(thermusthermophilus)。参见例如,swarts等,ibid,g.sheng等.(2013)proc.natl.acad.sci.u.s.a.111,652)。″ttago系统″是所需的全部组分,其包括例如用于被ttago酶切割的向导dna。
″重组″指两个多核苷酸之间交换遗传信息的过程。就本公开的目的而言,″同源重组(hr)″指发生这种交换的特定形式,例如在修复细胞内双链断裂期间通过同源导向修复机制发生。该过程要求核苷酸序列同源性,利用″供体″分子模板修复″靶″分子(即,经历双链断裂的分子),该过程也称作″非交叉基因转化″或″短道基因转化″,因为其导致遗传信息从供体向靶转移。不希望受限于任何特定理论,这种转移可以涉及断裂的靶与供体间形成的异双链体dna的错配校正,和/或采用供体再合成将成为部分靶的遗传信息的″合成依赖性链退火″,和/或相关过程。这种专门的hr通常导致靶分子序列的改变,从而供体多核苷酸的部分或全部序列被纳入靶多核苷酸中。
在本公开的方法中,本文所述的一种或多种靶向核酸酶在靶序列(例如,细胞染色质)中的预定位点(例如,感兴趣的基因或基因座)处产生双链断裂(dsb),并且可将与所述断裂区域中的核苷酸序列具有同源性的″供体″多核苷酸引入所述细胞。已显示dsb的存在有助于供体序列的整合。任选地,该构建体与该断裂区域内的核苷酸序列具有同源性。供体序列可被物理整合(physicallyintegrated),或者,供体多核苷酸被用作模板用于通过同源重组进行的断裂修复,导致将供体中的全部或部分核苷酸序列导入细胞染色质。因此,细胞染色质中的第一序列可改变,并且在某些实施方式中,可转化成供体多核苷酸中存在的序列。因此,所用术语″替换″或″置换″可理解为表示一种核苷酸序列被另一种核苷酸序列替换(即,信息意义上序列的替换),而不一定要求一种多核苷酸被另一种多核苷酸物理或化学替换。
在本文描述的任何方法中,其它锌指蛋白对可用于细胞内其它靶位点的另外的双链切割。
在用于靶向重组和/或替代和/或改变细胞染色质内感兴趣区域的序列的方法的某些实施方式中,通过采用外源性″供体″核苷酸序列的同源重组来改变染色体序列。如果存在与断裂的区域同源的序列,那么所述同源重组被细胞染色质中存在的双链断裂的刺激。
在本文所述的任何方法中,第一核苷酸序列(″供体序列″可包含与感兴趣区域中的基因组序列同源但不相同的序列,由此刺激同源重组以在感兴趣区域插入非相同序列。因此,在某些实施方式中,供体序列中与感兴趣区域序列同源的部分呈现与待替换的基因组序列有约80-99%(或其间任意整数)的序列相同性。在其它实施方式中,供体和基因组序列间的同源性超过99%,例如,超过100个连续碱基对的基因组序列与供体之间相差仅1个核苷酸。在某些情况中,供体序列的非同源部分可包含不存在于感兴趣区域的序列,从而将新序列导入感兴趣区域。在这些情况中,非同源序列一般侧接50-1,000个碱基对(或其间的任何整数值)或大于1,000的任何数量个碱基对的序列,其与感兴趣区域中的序列同源或相同。在其它实施方式中,供体序列与第一序列非同源,并且通过非同源重组机制插入基因组。
本文所述的任意方法可用于通过能破坏感兴趣基因表达的供体序列的靶向整合来使细胞内的一种或多种靶序列部分或完全失活。还提供具有部分或完全失活基因的细胞系。
此外,本文所述的靶向整合方法也可用于整合一种或多种外源性序列。外源性核酸序列可包含,例如,一种或多种基因或cdna分子,或任何类型的编码或非编码序列,以及一种或多种控制元件(例如,启动子)。此外,外源性核酸序列可产生一种或多种rna分子(例如,小发夹rna(shrna),抑制性rna(rnai),微小rna(mirna)等)。
″切割″指dna分子共价主链的断裂。切割可以由多种方法引发,包括但不限于,磷酸二酯键的酶促或化学水解。单链切割和双链切割均可采用,并且双链切割可由不同的单链切割事件所致。dna切割可导致产生钝端或交错末端。在某些实施方式中,将融合多肽用于靶向双链dna切割。
″切割半结构域″是能与第二多肽(两者相同或不同)形成具有切割活性(优选双链切割活性)的复合物的多肽序列。术语″第一和第二切割半结构域″、″+和-切割半结构域″和″右和左切割半结构域″可互换使用,指二聚化的切割半结构域的对。
″工程改造的切割半结构域″是经修饰以与另一切割半结构域(例如,另一工程改造的切割半结构域)形成专性异二聚体的切割半结构域。还参见美国专利号7,888,121;7,914,796;8,034,598;8,623,618和美国专利公开号2011/0201055,其全文通过引用方式纳入本文。
术语″序列″指任意长度的核苷酸序列,可以是dna或rna;可以是直链、环状或支链且可以是单链或双链。术语″供体序列″指插入基因组的核苷酸序列。供体序列可以是任意长度,例如长度为2-10,000个核苷酸(或其间或其上的任意整数值),优选长度为100-1,000个核苷酸(或其间的任意整数),更优选长度为200-500个核苷酸。
〝染色质″是包含细胞基因组的核蛋白结构。细胞染色质包含核酸和蛋白,核酸主要为dna,蛋白包括组蛋白和非组蛋白染色体蛋白。真核细胞染色质主要以核小体形式存在,其中核小体核心包含约150碱基对的dna与八聚体关联,所述八聚体包含组蛋白h2a、h2b、h3和h4各两份;以及在核小体核心之间延伸的接头dna(长度根据生物体而各有不同)。组蛋白h1分子通常与接头dna关联。就本公开的目的而言,术语″染色质″意在涵盖所有类型的细胞核蛋白,包括原核与真核的。细胞染色质包括染色体和附加体染色质。
″染色体″是包含细胞的全部或部分基因组的染色质复合物。细胞基因组通常由其核型表征,其为包含该细胞基因组的全部染色体的集合。细胞基因组可包含一条或多条染色体。
″附加体″是复制的核酸、核蛋白复合物或其它包含并非细胞染色体核型部分的核酸的结构。附加体的示例包括质粒和某些病毒基因组。
″靶位点″或″靶序列″是限定结合分子将结合的核酸部分的核酸序列,前提是存在结合的充分条件。例如,序列5′gaattc3′是ecori限制性内切核酸酶的靶位点。
〝外源性″分子是通常不存在于细胞内的分子,但可通过一种或多种遗传、生化或其它方法导入细胞。〝正常存在于细胞中″相对于细胞的具体发育阶段和环境条件确定。因此,例如,仅在肌肉的胚胎发育期间存在的分子对于成体肌肉细胞是外源性分子。类似地,相对于非热激的细胞,通过热激诱导的分子是外源性分子。例如,外源性分子可以包括功能失常的内源性分子的功能性形式或者正常功能的内源性分子的功能失常形式。
外源性分子可以是小分子或大分子等,小分子如由组合化学方法所产生,大分子如蛋白质、核酸、碳水化合物、脂质、糖蛋白、脂蛋白、多糖、上述分子的任何经修饰衍生物,或者是包含一种或多种上述分子的任何复合物。核酸包括dna和rna,其可以是单链或双链的;可以是直链、支链或环状的;并且可以具有任何长度。参见例如,美国专利号8,703,489和9,255,259。核酸包括能够形成双链体的那些,以及形成三链体的核酸。参见,例如美国专利号5,176,996和5,422,251。蛋白质可包括但不限于,dna结合蛋白、转录因子、染色质重构因子、甲基化dna结合蛋白、聚合酶、甲基化酶、脱甲基酶、乙酰基转移酶、脱乙酰酶、激酶、磷酸酶、整合酶、重组酶、连接酶、拓扑异构酶、旋转酶和解旋酶。
外源性分子可以是与内源性分子同一类型的分子,例如外源性蛋白或核酸。例如,外源性核酸可以包括感染性病毒基因组、导入细胞内的质粒或附加体,或包含通常不存在于细胞内的染色体。本领域技术人员已知将外源性分子导入细胞内的方法,包括但不限于脂质介导的转移(即脂质体,包括中性和阳离子脂质)、电转导、直接注射、细胞融合、粒子轰击、磷酸钙共沉淀、deae-葡聚糖介导的转移和病毒载体介导的转移。外源性分子也可以是与内源性分子相同类型但源自与该细胞来源不同物种的分子。例如,可将人核酸序列引入原始源自小鼠或仓鼠的细胞系。
相反,〝内源性″分子是通常存在于特定环境条件下特定发育阶段的特定细胞中的分子。例如,内源性核酸可以包括染色体、线粒体基因组、叶绿体或其它细胞器,或天然产生的附加型核酸。其它内源性分子可包括蛋白质,例如,转录因子和酶。
″融合″分子是其中两个或更多个亚基分子相连(优选共价相连)的分子。亚基分子可以是相同化学类型的分子,也可以是不同化学类型的分子。第一类型的融合分子的示例可包括但不限于,融合蛋白(例如,zfp或taledna结合结构域与一种或多种活化结构域之间的融合)和融合核酸(例如,编码上文所述的融合蛋白的核酸)。第二类融合分子的示例包括但不限于:形成三链体的核酸与多肽之间的融合体,以及小沟结合子与核酸之间的融合体。该术语还包括其中多核苷酸组分与多肽组分相关联以形成功能性分子(例如,crispr/cas系统,其中单一向导rna与功能结构域相关联以调节基因表达)。
细胞内融合蛋白的表达可由融合蛋白递送入细胞造成或通过向细胞递送编码融合蛋白的多核苷酸而引起,其中所述多核苷酸被转录,转录本经翻译产生所述融合蛋白。细胞中蛋白质的表达也可涉及反式剪接、多肽切割和多肽连接。用于将多核苷酸和多肽递送至细胞的方法在本公开内容中的他处呈现。
就本公开的目的而言,″基因″包括编码基因产物(见前文)的dna区域,以及调节基因产物生成的dna区域,不论这类调节序列是否毗邻编码和/或转录序列。因此,基因包括但不必限于,启动子序列、终止子、翻译调节序列,例如核糖体结合位点和内部核糖体进入位点、增强子、沉默子、绝缘子、边界元件、复制起点、基质连接位点和基因座控制区域。
″安全港″基因座是基因组内的基因座,其中,可插入基因且对宿主细胞没有任何有害影响。其中插入的基因序列的表达没有被来自相邻基因的任何通读表达所扰乱的安全港基因座是最有益的。由核酸酶靶向的安全港基因座的非限制性示例包括:ccr5、ccr5、hprt、aavs1、rosa和白蛋白。参见例如,美国专利号8,771,985、8,110,379、7,951,925、美国专利公开号2010/0218264、2011/0265198、2013/0137104、2013/0122591、2013/0177983、2013/0177960、2015/0056705和2015/0159172)。
″基因表达″指将基因所含信息转化成基因产物。基因产物可以是基因的直接转录产物(例如,mrna、trna、rrna、反义rna、核糖酶、结构rna或任何其它类型的rna)或通过mrna翻译产生的蛋白质。基因产物还包括经修饰的rna,通过如下加工修饰,例如加帽、聚腺苷酸化、甲基化,和编辑,以及通过如下加工修饰的蛋白质,例如,甲基化、乙酰化、磷酸化、泛素化、adp-核糖基化、豆蔻酰化和糖基化。
基因表达的〝调控″或〝修饰″指基因活性的改变。表达的调控可包括但不限于:基因活化和基因遏制,包括通过结合外源性分子(例如,经工程改造的转录因子)进行基因修饰来实现。调节可通过基因组编辑(例如,切割、改变、失活、随机突变)对基因序列进行修饰来实现。基因失活(或灭活)指,相较于未经本文所述修饰的细胞,基因表达的任何减少。因此,基因失活可以是部分或完全的。
″感兴趣的区域″是需要结合外源性分子的细胞染色质的任意区域,例如,基因或者基因内或与之毗邻的非编码序列。结合可以是出于靶向dna切割和/或靶向重组的目的。感兴趣的区域可以存在于例如染色体,附加体,细胞器基因组(例如,线粒体、叶绿体),或感染性病毒基因组中。感兴趣的区域可处于基因的编码区中,转录的非编码区域例如前导序列、尾随序列或内含子中,或非转录区域中编码区的上游或下游。感兴趣的区域可如单一核苷酸对一样小,或长达2,000个核苷酸对,或任意整数值的核苷酸对。
″真核″细胞可包括但不限于,真菌细胞(例如酵母)、植物细胞、动物细胞、哺乳动物细胞和人细胞(例如,t细胞)。
涉及两个或多个组件(例如序列元件)的并置,所述组件设置成组件都可正常发挥作用并允许组件中至少一种能介导施加于至少一种其它组件上的作用时,术语″操作性相连″和″操作性连接的″(或″操作地连接″)互换使用。例如,若转录调节序列控制编码序列响应一种或多种转录调节因子存在与否时的转录水平,则所述转录调节序列如启动子与所述编码序列操作性连接。转录调节序列一般与编码序列以顺式操作性地连接,但无需紧邻该序列。例如,增强子是操作性地连接编码序列的转录调节序列,尽管它们是不连续的。
对于融合多肽,术语″操作性连接″可指各组件在与其它组件的连接中所发挥的功能与其在未连接时的功能相同。例如,对于其中dna结合结构域(例如,zfp、tale)与活化结构域融合的融合多肽,如果在该融合多肽中,dna结合结构域部分能够结合其靶位点和/或其结合位点,同时所述活化结构域能够上调基因表达,那么所述dna结合结构域和活化结构域是操作性连接的。在其中dna结合结构域与切割结构域融合的融合多肽的情况中,如果在所述融合多肽中,dna结合结构域部分能结合其靶位点和/或其结合位点,同时切割结构域能切割靶位点附近的dna,那么所述dna结合结构域和切割结构域是操作性连接的。同理,对于其中dna结合结构域与活化或遏制结构域融合的融合多肽,如果在所述融合多肽中,dna结合结构域部分能够结合其靶位点和/或其结合位点,同时活化结构域能上调基因表达或遏制结构域能下调基因表达,那么所述dna结合结构域和活化或遏制结构域是操作性连接的。
蛋白质、多肽或核酸的″功能性片段(或功能片段)″是序列与全长蛋白质、多肽或核酸不相同但保留全长蛋白质、多肽或核酸的相同功能的蛋白质、多肽或核酸。功能片段可具有比相应的天然分子更多、更少或相同数量的残基,和/或可含有1个或多个氨基酸或核苷酸取代。用于确定核酸功能(例如,编码功能、与另一核酸杂交的能力)的方法是本领域熟知的。类似地,用于确定蛋白质功能的方法也是为本领域熟知的。例如,可测定多肽的dna结合功能,例如通过滤膜结合、电泳迁移率改变或免疫沉淀试验。dna切割可通过凝胶电泳分析。参见ausubel等,同上。可测定蛋白质与另一蛋白质相互作用的能力,例如,通过共免疫沉淀、双杂交试验或互补分析,既可以是遗传的也可以是生化的。参见例如,fields等.(1989)nature340:245-246;美国专利号5,585,245和国际专利公开号wo98/44350。
″载体″能将基因序列转移至靶细胞。″载体构建体″、″表达载体″和″基因转移载体″通常指能指导感兴趣基因表达并能将基因序列转移至靶细胞的核酸构建体。因此,该术语包括克隆和表达载体,以及整合载体。
″报告基因″或″报告序列″指的是产生这样蛋白质产物的任何序列,所述蛋白质产物容易被检测,其在常规试验中并非必需但是优选的。合适的报告基因包括但不限于,编码介导抗生素抗性(例如氨苄青霉素抗性,新霉素抗性,g418抗性,嘌呤霉素抗性)的蛋白质的序列,编码有色或荧光或发光蛋白(例如绿色荧光蛋白,增强型绿色荧光蛋白,红色荧光蛋白,荧光素酶)的序列,和介导增强的细胞生长和/或基因扩增的蛋白质(例如,二氢叶酸还原酶)。表位标签包括,例如,一个或多个拷贝的flag、his、myc、tap、ha或任何可检测的氨基酸序列。〝表达标签″包括编码可操作性地连接至所需基因序列以监测感兴趣的基因的表达的报告物的序列。
术语″对象″和″患者″可以互换使用,并且表示哺乳动物例如人类患者和非人类灵长类,以及实验室动物例如兔、狗、猫、大鼠、小鼠和其他动物。因此,本文所用术语〝对象″或″患者″表示可给予本发明表达盒的任何哺乳动物患者或对象。本发明的对象包括患有病症的那些或处于发展病症的风险中的那些。
本文所用的术语″治疗″和″处理″指减少症状的严重性和/或频率、消除症状和/或根本原因、预防症状的发生和/或它们的根本原因、以及改善或消除损伤。癌症和移植物抗宿主疾病是可以使用本文所述组合物和方法治疗的病症的非限制性示例。因此,〝处理″和〝治疗″包括:
(i)预防所述疾病或病症在哺乳动物中发生,具体是在所述哺乳动物倾向于患上该病症但尚未诊断患有该病症时;
(ii)抑制该疾病或病症,即阻止其发展;
(iii)减轻该疾病或病症,即,使该疾病或病症消退;或
(iv)减轻由该疾病或病症所致的症状,即,减轻痛苦但不解决潜在的疾病或病症。
本文中所用术语″疾病″和″病症″可互换使用,或可不同,其中具体的疾病或病症可不具有已知的致病物(从而病因尚不知晓),因此其尚未被视作疾病而仅仅是作用不希望的病症或综合征,其中由临床医师或多或少地鉴定出了具体的一组症状。
〝药物组合物″指一种制剂,其具有本发明的化合物和本领域通常接受的用于将生物活性化合物递送到哺乳动物(例如,人)的介质。这样的介质包括药学上可接受的运载体、稀释剂或赋形剂等等。
″有效量″或″治疗有效量″指:本发明化合物的一定量,当将其给予哺乳动物,优选人类时,足以提供对于该哺乳动物(优选人类)的有效治疗。构成″治疗有效量″的本发明化合物的量将根据化合物、病症及其严重性、给予方式,以及待治疗的哺乳动物的年龄而变化,但可由本领域技术人员根据其所掌握的知识和本发明内容来常规地确定。
dna结合结构域
本文描述了组合物,其包含dna结合结构域,所述dna结合结构域特异性地结合包括hla基因或hla调节物的任何基因中的靶位点。本文公开的方法和组合物中可以使用任何dna结合结构域,包括但不限于,锌指dna结合结构域,taledna结合结构域,crispr/cas核酸酶的dna结合部分(sgrna),或来自兆核酸酶(meganuclease)的dna结合结构域。dna结合结构域可结合基因内的任何靶序列,包括但不限于,本文公开的任何靶位点(seqidno:8-21和/或92-103)所示的12或更多个核苷酸的靶序列。
在某些实施方式中,dna结合结构域包含锌指蛋白。优选地,该锌指蛋白是非天然产生的,其中它经工程改造以结合至所选的靶位点。参见例如,beerli等(2002)naturebiotechnol.20:135-141;pabo等(2001)ann.rev.biochem.70:313-340;isalan等(2001)naturebiotechnol.19:656-660;segal等(2001)curr.opin.biotechnol.12:632-637;choo等(2000)curr.opin.struct.biol.10:411-416;美国专利号6,453,242、6,534,261、6,599,692、6,503,717、6,689,558、7,030,215、6,794,136、7,067,317、7,262,054、7,070,934、7,361,635和7,253,273;和美国专利公开号2005/0064474、2007/0218528、2005/0267061,其均通过引用其全文纳入本文。在某些实施方式中,dna结合结构域包括美国专利公开号2012/0060230公开的锌指蛋白(例如,表1),通过引用将其全部内容纳入本文。在其它实施方式中,dna结合结构域包含zfp组分(称为〝设计物″)并且包括如表1、2、4、5、6或8的zfn中所示的识别螺旋区和主链,包括但不限于zfn72732、72748、68957或72678的zfp结构域。
与天然产生的锌指蛋白相比,经工程改造的锌指结合结构域可具有新结合特异性。工程改造方法可包括但不限于,合理设计和不同选择类型。例如,合理设计包括利用包含三体(或四体)核苷酸序列和单独锌指氨基酸序列的数据库,其中各三体或四体核苷酸序列与一种或多种结合该特定三体或四体序列的锌指氨基酸序列相关联。参见例如,美国专利号6,453,242和6,534,261,通过引用其全文的方式纳入本文。
示例性选择方法包括噬菌体展示和双杂交系统,公开于美国专利号5,789,538、5,925,523、6,007,988、6,013,453、6,410,248、6,140,466、6,200,759和6,242,568,以及国际专利公开号wo98/37186、wo98/53057、wo00/27878和wo01/88197,以及gb2,338,237。此外,例如在美国专利号6,794,136中描述了对锌指结合结构域的结合特异性的增强。
此外,如这些及其它参考文献中所公开的,锌指结构域和/或多指的锌指蛋白可利用任何合适的接头序列连接在一起,包括例如长度为5个或更多个氨基酸的接头。长度为6个或更多个氨基酸的示例性接头序列还参见例如美国专利号6,479,626、6,903,185和7,153,949。本文所述的蛋白质可包括蛋白质的个体锌指之间的合适的接头的任何组合。此外,例如在美国专利号6,794,136中描述了对锌指结合结构域的结合特异性的增强。
靶位点选择;用于设计和构建融合蛋白(及其编码多核苷酸)的zfp和方法是本领域技术人员已知的,并且详细描述于美国专利号6,140,081、5,789,538、6,453,242、6,534,261、5,925,523、6,007,988、6,013,453、6,200,759;和国际专利公开号wo95/19431、wo96/06166、wo98/53057、wo98/54311、wo00/27878、wo01/60970、wo01/88197、wo02/099084、wo98/53058、wo98/53059、wo98/53060、wo02/016536和wo03/016496。
此外,如这些及其它参考文献中所公开的,锌指结构域和/或多指的锌指蛋白可利用任何合适的接头序列连接在一起,包括例如长度为5个或更多个氨基酸的接头。示例性接头序列还参见美国专利号6,479,626、6,903,185、7,153,949、7,888,121、7,914,796、8,034,598、8,623,618、9,567,609;和美国专利公开号2017/0218349。本文所述的蛋白质可包括蛋白质的个体锌指之间的合适接头的任何组合。
在某些实施方式中,dna结合结构域是经工程改造的锌指蛋白,其(以序列特异性方式)结合至tcr基因或tcr调节基因中的靶位点并调节tcr基因的表达。在一些实施方式中,锌指蛋白结合至tcra中的靶位点,而在其它实施方式中,锌指结合至trbc中的靶位点。在其它实施方式中,dna结合结构域是经工程改造的锌指蛋白,其以序列特异性方式结合至b2m基因中的靶位点并调节b2m基因的表达。这些dna结合结构域的非限制性示例性实施方式示于表1、2和6(tcr)以及表5和8(b2m)。在某些实施方式中,zfp包括命名为72732、72748、68957或72678的zfn的zfp部分。
zfp通常包括至少三个指。某些zfp包括四个、五个或六个指。包括三个指的zfp通常识别包括9或10个核苷酸的靶位点;包括四个指的zfp通常识别包括12-14个核苷酸的靶位点;而具有六个指的zfp能识别包括18-21个核苷酸的靶位点。zfp也可以是包括一个或多个调节结构域的融合蛋白,所述结构域可以是转录活化或阻遏结构域。
在一些实施方式中,dna结合结构域可衍生自核酸酶。例如,寻靶内切核酸酶和兆核酸酶的识别序列例如i-scei、i-ceui、pi-pspi、pi-sce、i-sceiv、i-csmi、i-pani、i-sceii、i-ppoi、i-sceiii、i-crei、i-tevi、i-tevii和i-teviii是已知的。还可参见美国专利号5,420,032;美国专利号6,833,252;belfort等.(1997)nucleicacidsres.25:3379-3388;dujon等.(1989)gene82:115-118;perler等.(1994)nucleicacidsres.22,1125-1127;jasin(1996)trendsgenet.12:224-228;gimble等.(1996)j.mol.biol.263:163-180;argast等.(1998)j.mol.biol.280:345-353和新英格兰生物实验室公司(newenglandbiolabs)产品目录。此外,寻靶内切核酸酶和兆核酸酶的dna结合特异性可被工程改造以结合非天然靶位点。参见例如chevalier等.(2002)molec.cell10:895-905;epinat等.(2003)nucleicacidsres.31:2952-2962;ashworth等.(2006)nature441:656-659;
在其它实施方式中,dna结合结构域包含来自tal效应物的经工程改造的结构域,其类似于源自植物病原体黄单胞菌(xanthomonas)(参见boch等,(2009)science326:1509-1512和moscou与bogdanove(2009)science326:1501)和青枯菌(ralstonia)(参见heuer等(2007)appliedandenvironmentalmicrobiology73(13):4379-4384)的那些;美国专利申请号2011/0301073和2011/0145940。已知黄单胞菌属(xanthomona)的植物病原菌在重要农作物中导致很多疾病。黄单胞菌属的病原性取决于保守的iii型分泌(t3s)系统,该系统向植物细胞内注入超过25种不同的效应物蛋白。这些注射的蛋白质包括模仿植物转录活化剂并操纵植物转录组的转录活化剂样效应物(tale)(参见kay等,(2007)science318:648-651)。这些蛋白质含有dna结合结构域和转录活化结构域。最为充分表征的tale之一是来自野油菜黄单胞菌辣椒斑点病致病变种(xanthomonascampestrispv.vesicatoria)的avrbs3(参见bonas等(1989)molgengenet218:127-136和国际专利公开号wo2010/079430)。tale含有串联重复的集中化结构域,各重复含有约34个氨基酸,它们是这些蛋白的dna结合特异性的关键。此外,它们含有核定位序列和酸性转录活化结构域(综述参见schornacks等(2006)jplantphysiol163(3):256-272)。此外,在致植物病细菌烟草青枯菌(ralstoniasolanacearum)中,已在烟草青枯菌生物变型1菌株gmi1000和生物变型4菌株rs1000中发现两个基因,称为brgll和hpx17,其与黄单胞菌属(xanthomonas)的avrbs3家族具有同源性(参见heuer等(2007)applandenvirmicro73(13):4379-4384)。这些基因在核苷酸序列上彼此具有98.9%的同一性,但区别为hpx17重复结构域中的1,575bp缺失。然而,两种基因产物均与黄单胞菌属的avrbs3家族蛋白质具有低于40%序列相同性。
这些tal效应物的特异性取决于串联重复中存在的序列。重复序列包含约102个碱基对,且所述重复彼此通常为91-100%同源(bonas等,同上)。重复的多态性通常位于12和13位,并且似乎在位置12和13的高可变双残基(重复可变双残基或rvd区域)的种类与tal效应物靶序列中连续核苷酸的种类之间存在一一对应(参见moscou和bogdanove,(2009)science326:1501以及boch等(2009)science326:1509-1512)。实验上,已确定用于这些tal效应物dna识别的天然编码,因此位于位置12与13的hd序列(重复可变双残基或rvd)导致对于胞嘧啶(c)的结合,ng结合至t、ni结合至a、c、g或t,nn结合至a或g,且ing结合至t。这些dna结合重复已被组装进入具有新的重复组合与数量的蛋白质,产生能在植物细胞中与新序列相互作用并活化非内源性报告基因的表达的人工转录因子(boch等,同上)。已将经工程改造的tal蛋白连接至foki切割半结构域,以产生tal效应物结构域核酸酶融合体(talen),包括具有非典型rvd的talen。参见例如,美国专利号8,586,526。
在一些实施方式中,talen包括内切核酸酶(例如,foki)切割结构域或切割半结构域。在其它实施方式中,tale-核酸酶是兆tal(megatal)。这些兆tal核酸酶是包含taledna结合结构域和兆核酸酶切割结构域的融合蛋白。兆核酸酶切割结构域作为单体具有活性,且不需要二聚化获得活性。(参见boissel等,(2013)nuclacidres:1-13,doi:10.1093/nar/gkt1224)。
在另一实施方式中,核酸酶包括紧凑型(compact)talen。这些是单链融合蛋白,其将taledna结合结构域与tevi核酸酶结构域相连。该融合蛋白可以作为通过tale区域定位的切口酶,或可以产生双链断裂,这取决于该taledna结合结构域相对于tevi核酸酶结构域所位于的位置(参见beurdeley等(2013)natcomm:41762doi:10.1038/ncomms2782)。此外,核酸酶结构域还可显示dna结合功能性。任何talen都可与其它talen联用,例如,一种或多种talen(ctalen或foki-talen)与一种或多种兆tale。
此外,如这些及其它参考文献中所公开的,锌指结构域和/或多指的锌指蛋白或tale可利用任何合适的接头序列(包括例如长度为5或更多个氨基酸的接头)连接在一起。长度为6或更多个氨基酸的示例性接头序列还参见例如美国专利号6,479,626、6,903,185和7,153,949。本文所述的蛋白质可包括蛋白质的个体锌指之间的合适接头的任何组合。此外,例如在美国专利号6,794,136中描述了对锌指结合结构域的结合特异性的增强。
在某些实施方式中,dna结合结构域是crispr/cas核酸酶系统的部分,包括与dna结合的单向导rna(sgrna)。参见例如,美国专利号8,697,359和美国专利公开号2015/0056705和2015/0159172。编码该系统的rna组分的crispr(规律成簇的间隔短回文重复序列)基因座,以及编码蛋白质的cas(crispr相关)基因座(jansen等.(2002)mol.microbiol.43:1565-1575;makarova等.(2002)nucleicacidsres.30:482-496;makarova等.(2006)biol.direct1:7;haft等.(2005)ploscomput.biol.1:e60)组成了crispr/cas核酸酶系统的基因序列。微生物宿主中的crispr基因座包含crispr-相关(cas)基因以及能够对crispr介导的核酸切割的特异性进行编程的非编码rna元件的组合。
ii型crispr是最充分表征的系统之一,并且以4个连续步骤进行靶向dna双链断裂。首先,从crispr基因座转录两个非编码rna:前-crrna阵列和tracrrna。第二,tracrrna杂交至前-crrna的重复序列区域,并且介导从前-crrna到含个体间隔子序列的成熟crrna的加工。第三,成熟的crrna:tracrrna复合物通过沃森-克里克碱基配对将功能结构域(例如,核酸酶,如cas)引导至靶dna,其位于crrna上的间隔子和靶dna上的原型间隔子之间,所述靶dna在原型间隔子邻近基序(protospaceradjacentmotif,pam)旁,其为靶向识别的附加要求。最后,cas9介导靶dna的切割,以在原型间隔子内产生双链断裂。crispr/cas系统活性包括3步:(i)通过称作″适应″的过程将外来dna序列插入crispr阵列,以防止未来的攻击,(ii)表达相关蛋白质,以及表达和处理所述阵列,然后(iii)采用外来核酸进行rna介导的干扰。因此,在细菌细胞中,多个所谓的″cas″蛋白参与crispr/cas系统的自然功能,并且在多种功能中发挥作用(例如外来dna的插入等)。
在某些实施方式中,cas蛋白可为天然产生的cas蛋白的″功能性衍生物″。天然序列多肽的″功能性衍生物″是与天然序列多肽具有共同定性生物性质的化合物。″功能性衍生物″包括但不限于天然序列的片段或天然序列多肽的衍生物及其片段,前提是它们与相应的天然序列多肽具有共同的生物活性。本文设想的生物活性是功能性衍生物将dna底物水解成片段的能力。术语″衍生物″包括多肽的氨基酸序列变体,共价修饰物,和其融合体,例如衍生的cas蛋白。cas多肽合适的衍生物或其片段包括但不限于cas蛋白的突变体、融合体、共价修饰形式或其片段。包括cas蛋白或其片段的cas蛋白以及cas蛋白或其片段的衍生物可获自细胞或通过化学合成或通过这两种方法的组合获得。所述细胞可以是天然产生cas蛋白的细胞,或天然产生cas蛋白并经遗传工程改造的细胞,所述遗传工程改造使所述细胞以较高表达水平产生内源性cas蛋白或从外源导入的核酸产生cas蛋白,其中所述核酸编码与内源性cas相同或不同的cas。在一些情况中,细胞天然不产生cas蛋白,但是经遗传工程改造以产生cas蛋白。在一些实施方式中,cas蛋白是小cas9同源物,用于经aav载体的递送(ran等(2015)nature510:186)。
在一些实施方式中,dna结合结构域是ttago系统的部分(参见swarts等,如上;sheng等,如上)。在真核细胞中,基因沉默通过阿尔古(argonaute,ago)家族的蛋白介导。在该范例中,ago与小(19-31nt)rna结合。该蛋白质-rna沉默复合物通过小rna与靶标之间的沃森-克里克碱基配对识别靶rna,并且通过内切核酸裂解活性切割靶rna(vogel(2014)science344:972-973)。相比之下,原核ago蛋白与小单链dna片段结合,并且可能作用于检测并移除外源性(通常是病毒)dna(yuan等,(2005)mol.cell19,405;olovnikov等(2013)mol.cell51,594;swarts等,同上)。示例性的原核ago蛋白包括:来自超嗜热菌(aquifexaeolicus)、类球红细菌(rhodobactersphaeroides)和嗜热栖热菌(thermusthermophilus)的那些。
最为充分表征的原核ago蛋白之一是来自嗜热栖热菌(t.thermophilus)的一种(ttago;swarts等,同上)。ttago与具有5′磷酸基团的15nt或13-25nt单链dna片段关联。被ttago结合的该″向导dna″可用于引导该蛋白质-dna复合物结合第三方(third-party)dna分子中的沃森-克里克互补dna序列。一旦这些向导dna中的序列信息允许鉴定靶dna,ttago-向导dna复合物即能切割该靶dna。这样的机制还被ttago-向导dna复合物的结构支持,尽管与其靶dna结合(g.sheng等,同上)。来自类球红细菌的ago(rsago)具有相似的性质(olovnikov等,同上)。
任何dna序列的外源性向导dna均可被加载到ttago蛋白上(swarts等,同上)。因为ttago切割的特异性由向导dna引导,所以与外源性研究物特异性的(investigator-specified)向导dna形成的ttago-dna复合物将引导ttago靶dna切割至互补性研究物特异性的靶dna。由此,人们可在dna中生成靶向的双链断裂。ttago-向导dna系统(或来自其它生物体的直系同源ago-向导dna系统)的应用允许细胞内基因组dna的靶向切割。这样的切割可以是单链的或双链的。对于哺乳动物基因组dna的切割,优选应用针对哺乳动物细胞中表达进行优化的ttago密码子形式。此外,可优选用体外形成的ttago-dna复合物处理细胞,其中ttago蛋白融合至细胞穿透肽。并且,可优选应用已通过诱变而被改变的tgago蛋白的形式以在37℃具有增强的活性。ago-rna介导的dna切割可用于影响结果的全面多重性(panopoly),包括基因敲除,靶向基因添加,基因校正,靶向基因缺失,使用本领域标准的利用dna断裂的技术。
因此,可使用任何dna结合结构域。
融合分子
还提供融合分子,所述融合分子包含与异源性调节(功能)结构域(或其功能性片段)相关联的本文所述的dna结合结构域(例如,zfp或tale,crispr/cas组分,例如单向导rna)。常见结构域包括,例如,转录因子结构域(活化剂,阻遏物,共活化剂,共阻遏物),沉默子,癌基因(例如myc,jun,fos,myb,max,mad,rel,ets,bcl,myb,mos家族成员等);dna修复酶及其相关因子和修饰物;dna重排酶及其相关因子和修饰物;染色质相关蛋白质及其修饰物(例如激酶,乙酰酶和脱乙酰酶);和dna修饰酶(例如甲基转移酶,拓扑异构酶,解旋酶,连接酶,激酶,磷酸酶,聚合酶,核酸内切酶)及其相关因子和修饰物。此类融合分子包括转录因子,其包含本文所述dna结合结构域和转录调节结构域,以及核酸酶,其包含dna结合结构域和一个或多个核酸酶结构域。
用于实现活化的合适结构域(转录活化结构域)包括hsvvp16活化结构域(参见例如,hagmann等.(1997)j.virol.71:5952-5962),核激素受体(参见例如,torchia等.(1998)curr.opin.cell.biol.10:373-383);核因子κb的p65亚基(bitko和barik(1998)j.virol.72:5610-5618,及doyle和hunt(1997)neuroreport8:2937-2942);liu等.(1998)cancergenether.5:3-28),或人工嵌合功能结构域如vp64(beerli等.(1998)proc.natl.acad.sci.usa95:14623-33),和降解决定子(molinari等.(1999)emboj.18,6439-6447)。其它示例性活化结构域包括,oct1,oct-2a,sp1,ap-2和ctf1(seipel等.(1992)emboj.11,4961-4968以及p300,cbp,pcaf,src1pvalf,athd2a和erf-2。参见例如robyr等.(2000)mol.endocrinol.14:329-347;collingwood等.(1999)j.mol.endocrinol.23:255-275;leo等.(2000)gene245:1-11;manteuffel-cymborowska(1999)actabiochim.pol.46:77-89;mckenna等.(1999)j.steroidbiochem.mol.biol.69:3-12;malik等.(2000)trendsbiochem.sci.25:277-283;和lemon等.(1999)curr.opin.genet.dev.9:499-504。其它示例性活化结构域包括但不限于,osgai,half-1,c1,ap1,arf-5,-6,-7和-8,cprf1,cprf4,myc-rp/gp,和trab1。参见例如,ogawa等.(2000)gene245:21-29;okanami等.(1996)genescells1:87-99;goff等.(1991)genesdev.5:298-309;cho等.(1999)plantmol.biol.40:419-429;ulmason等.(1999)proc.natl.acad.sci.usa96:5844-5849;sprenger-haussels等.(2000)plantj.22:1-8;gong等.(1999)plantmol.biol.41:33-44;和hobo等.(1999)proc.natl.acad.sci.usa96:15,348-15,353。
本领域技术人员应明了,在dna结合结构域与功能结构域之间的融合蛋白(或编码相同内容的核酸)的形成中,活化结构域或与活化结构域相互作用的分子适于作为功能结构域。基本上,能够招募活化复合物和/或活化活性(例如,组蛋白乙酰化)至靶基因的任何分子均可用作融合蛋白的活化结构域。适用作融合分子中的功能结构域的绝缘子结构域、定位结构域和染色质重构蛋白(如含iswi的结构域和/或甲基结合结构域的蛋白质)描述于,例如,美国专利号7,053,264。
示例性的阻遏结构域包括但不限于,kraba/b,kox,tgf-b-诱导型早期基因(tieg),v-erba,sid,mbd2,mbd3,dnmt家族成员(例如,dnmt1,dnmt3a,dnmt3b),rb和mecp2。参见例如,bird等.(1999)cell99:451-454;tyler等.(1999)cell99:443-446;knoepfler等.(1999)cell99:447-450;和robertson等.(2000)naturegenet.25:338-342。其它示例性阻遏结构域包括但不限于rom2和athd2a。参见例如,chem等.(1996)plantcell8:305-321;和wu等.(2000)plantj.22:19-27。
通过本领域技术人员熟知的克隆和生物化学偶联方法构建融合分子。融合分子包括dna结合结构域(例如,zfp、tale、sgrna),其与功能结构域(例如,转录活化或遏制结构域)相关联。融合分子还任选地包含核定位信号(例如,来自sv40介质t-抗原)和表位标签(例如,flag和血细胞凝集素)。设计融合蛋白(及其编码核酸),从而翻译阅读框保留于融合体的组分间。
一方面的功能结构域(或其功能片段)的多肽组分与另一方面的非蛋白质dna结合结构域(例如抗生素,嵌入剂,小沟结合物,核酸)之间的融合体通过本领域技术人员已知的生化偶联方法来构建。参见例如,皮尔斯化学公司(piercechemicalcompan)(伊利诺伊州罗克福德)产品目录。已描述用于生成小沟结合剂与多肽之间的融合的方法和组合物。mapp等.(2000)proc.natl.acad.sci.usa97:3930-3935。此外,crispr/cas系统的单一向导rna与功能结构域相关联,以形成活性转录调节剂和核酸酶。
在某些实施方式中,靶位点存在于细胞染色质的可及区域中。可及区域可以如例如美国专利号7,217,509和7,923,542所述确定。如果靶位点不存在于细胞染色质的可及区域中,则可按美国专利号7,785,792和8,071,370所述产生一个或多个可及区域。在其它实施方式中,融合分子的dna结合结构域能够结合至细胞染色质,无论其靶位点是否位于可及区域中。例如,所述dna结合结构域能够结合至接头dna和/或核小体dna。该类型的″先驱″dna结合结构域的示例可见于某些类固醇受体和肝细胞核因子3(hnf3)(cordingley等(1987)cell48:261-270;pina等(1990)cell60:719-731;和cirillo等(1998)emboj.17:244-254)。
融合分子可用药学上可接受的运载体配制,如本领域技术人员已知。参见例如《雷明顿药物科学》(remington′spharmaceuticalsciences),第17版,1985;和美国专利号6,453,242和6,534,261。
融合分子的功能性组分/结构域可选自任何多种不同的组分,所述组分一旦该融合分子通过其dna结合结构域结合至靶序列即能影响基因转录。因此,功能性组分可包括但不限于,各种转录因子结构域,例如活化物、阻遏物、共活化物、共阻遏物,和沉默子。
例如,美国专利号6,534,261和6,933,113中公开了其它示例性功能结构域。
还可选择通过外源性小分子或配体调节的功能结构域。例如,可使用
核酸酶
在某些实施方式中,融合分子包括与切割(核酸酶)结构域相关联的dna结合型结合结构域。如此,基因修饰可采用核酸酶,例如经工程改造的核酸酶来实现。经工程改造的核酸酶技术基于对天然产生的dna结合蛋白的工程改造。例如,已描述了具有调整的dna结合特异性的寻靶内切核酸酶的工程改造。chames等.(2005)nucleicacidsres33(20):e178;arnould等.(2006)j.mo1.biol.355:443-458。此外,zfp的工程改造已有描述。例如,参见美国专利号6,534,261;6,607,882;6,824,978;6,979,539;6,933,113;7,163,824;和7,013,219。
此外,zfp和/或tale可融合至核酸酶结构域,以产生zfn和talen,其为一种功能性实体,该功能性实体能够通过其经工程改造的(zfp或tale)dna结合结构域识别其意图针对的核酸靶标,并通过核酸酶活性在dna结合位点附近切割该dna。
因此,本文所述的方法和组合物应用面广,并且可涉及任何感兴趣的核酸酶。核酸酶的非限制性示例包括兆核酸酶、talen和锌指核酸酶。核酸酶可包含异源性dna结合结构域和切割结构域(例如,锌指核酸酶;具有异源性切割结构域的兆核酸酶dna结合结构域),或者替代地,天然产生的核酸酶的dna结合结构域可被改变以结合至选择的靶位点(例如,以经工程改造以结合不同于关联结合位点的位点的兆核酸酶)。
在本文所述的任何核酸酶中,核酸酶可包含经工程改造的taledna结合结构域和核酸酶结构域(例如,内切核酸酶和/或兆核酸酶结构域),也称为talen。已经公开了用于工程改造这些talen蛋白从而与用户选择的靶序列稳健且位点特异性地相互作用的方法和组合物(参见,美国专利号8,586,526)。在一些实施方式中,talen包括内切核酸酶(例如,foki)切割结构域或切割半结构域。在其它实施方式中,tale-核酸酶是兆tal。这些兆tal核酸酶包含taledna结合结构域和兆核酸酶切割结构域的融合蛋白。兆核酸酶切割结构域作为单体具有活性,且不需要二聚化获得活性。(参见boissel等,(2013)nuclacidres:1-13,doi:10.1093/nar/gkt1224)。此外,核酸酶结构域还可显示dna结合功能性。
在另一实施方式中,核酸酶包括紧凑型talen(ctalen)。存在单链融合蛋白,其将taledna结合结构域与tevi核酸酶结构域相连。该融合蛋白可以作为通过tale区域定位的切口酶,或可以产生双链断裂,这取决于该taledna结合结构域相对于tevi核酸酶结构域所位于的位置(参见beurdeley等(2013)natcomm:41762doi:10.1038/ncomms2782)。任何talen都可与其它talen(例如,一种或多种talen(ctalen或foki-talen)和一种或多种兆tal)或其它dna切割酶联用。
在其它实施方式中,核酸酶包括显示切割活性的兆核酸酶(寻靶内切核酸酶)或其部分。天然产生的兆核酸酶识别15-40碱基对的切割位点,并且通常分为四个家族:laglidadg家族(″laglidadg″,公开为seqidno:122)、giy-yig家族、his-cyst盒家族和hnh家族。示例性寻靶核酸内切酶包括i-scei、i-ceui、pi-pspi、pi-sce、i-sceiv、i-csmi、i-pani、i-sceii、i-ppoi、i-sceiii、i-crei、i-tevi、i-tevii和i-teviii。其识别序列是已知的。还参见美国专利号5,420,032;美国专利号6,833,252;belfort等.(1997)nucleicacidsres.25:3379-3388;dujon等.(1989)gene82:115-118;perler等.(1994)nucleicacidsres.22:1125-1127;jasin(1996)trendsgenet.12:224-228;gimble等.(1996)j.mol.biol.263:163-180;argast等.(1998)j.mol.biol.280:345-353和新英格兰生物实验室公司产品目录。
来自天然产生的兆核酸酶、主要来自laglidadg家族(〝laglidadg″,公开为seqidno:122)的dna结合结构域已被用于在植物、酵母、果蝇、哺乳动物细胞和小鼠中促进位点特异性基因组修饰,但该方法仅限于保留兆核酸酶识别序列的同源基因的修饰(monet等(1999),biochem.biophysics.res.common.255:88-93)或限于其中已引入识别序列的经预先工程改造的基因组(route等(1994),mol.cell.biol.14:8096-106;chilton等(2003),plantphysiology.133:956-65;puchta等(1996),proc.natl.acad.sci.usa93:5055-60;rong等(2002),genesdev.16:1568-81;gouble等(2006),j.genemed.8(5):616-622)。因此,人们已尝试工程改造兆核酸酶,使其在医学或生物技术相关位点显示新结合特异性(porteus等(2005),nat.biotechnol.23:967-73;sussman等(2004),j.mol.biol.342:31-41;epinat等(2003),nucleicacidsres.31:2952-62;chevalier等(2002)molec.cell10:895-905;epinat等(2003)nucleicacidsres.31:2952-2962;ashworth等(2006)nature441:656-659;paques等(2007)currentgenetherapy7:49-66;美国专利公开号2007/0117128、2006/0206949、2006/0153826、2006/0078552和2004/0002092)。此外,天然产生或经工程改造的来自兆核酸酶的dna结合结构域可以操作性地连接来自异源性核酸酶(例如,foki)的切割结构域,和/或,来自兆核酸酶的切割结构域可操作性地连接异源性dna结合结构域(例如,zfp或tale)。
在其它实施方式中,核酸酶是锌指核酸酶(zfn)或taledna结合结构域-核酸酶融合体(talen)。zfn和talen包括dna结合结构域(锌指蛋白或taledna结合结构域),其经工程改造以结合至所选基因和切割结构域或切割半结构域(例如,来自如本文所述的限制和/或兆核酸酶)中的靶位点。
如上详述,锌指结合结构域和taledna结合结构域可经工程改造以结合所选序列。参见例如,beerli等(2002)naturebiotechnol.20:135-141;pabo等(2001)ann.rev.biochem.70:313-340;isalan等(2001)naturebiotechnol.19:656-660;segal等(2001)curr.opin.biotechnol.12:632-637;choo等(2000)curr.opin.struct.biol.10:411-416。与天然产生的锌指蛋白相比,经工程改造的锌指结合结构域或tale蛋白可具有新结合特异性。工程改造方法包括但不限于合理设计和不同选择类型。合理设计包括,例如,利用包含三体(或四体)核苷酸序列和个体锌指或tale氨基酸序列的数据库,其中各三体或四体核苷酸序列与结合该特定三体或四体序列的锌指或tale重复单元的一个或多个氨基酸序列相关联。参见例如,美国专利号6,453,242和6,534,261,通过引用其全文的方式纳入本文。在某些实施方式中,dna结合结构域包含衍生自命名为68957、72678、72732、72748(b2m)或68846(tcr)的zfn的zfp(例如,zfp组分)。
靶位点的选择;和用于设计和构建融合蛋白(以及编码相同内容的多核苷酸)的方法是本领域技术人员已知的并详述于美国专利号7,888,121和8,409,861中,两者通过引用其全文的方式纳入本文。
此外,如这些和其它参考文献中所公开的,锌指结构域、tale和/或多指的锌指蛋白可利用任何合适的接头序列(包括例如长度为5或更多个氨基酸的接头)连接在一起。长度为6或更多个氨基酸的示例性接头序列参见例如美国专利号6,479,626、6,903,185和7,153,949。本文所述的蛋白质可包括蛋白质的个体锌指之间的合适接头的任何组合。还可参见美国专利号8,772,453。
因此,核酸酶(如zfn、talen和/或兆核酸酶)可包括任何dna结合结构域和任何核酸酶(切割)结构域(切割结构域,切割半结构域)。如上所述,切割结构域相对于dna结合结构域可以是异源的,例如,锌指或tal效应物dna结合结构域和来自某一核酸酶的切割结构域,或兆核酸酶dna结合域和来自另一不同核酸酶的切割结构域。异源切割结构域可获自任何内切核酸酶或外切核酸酶。可衍生出切割结构域的示范性核酸内切核酸酶,包括但不限于限制性内切核酸酶和寻靶内切核酸酶。参见例如,马萨诸塞州贝弗利(beverly,ma)的neb公司的2002-2003产品目录;和belfort等(1997)nucleicacidsres.25:3379-338。切割dna的其它酶是已知的(例如,s1核酸酶、绿豆核酸酶、胰dna酶i、微球菌核酸酶、酵母ho核酸内切酶;还参见linn等(编)《核酸酶》(nucleases),冷泉港实验室出版社(coldspringharborlaboratorypress),1993)。可将一种或多种这些酶(或其功能性片段)用作切割结构域和切割半结构域的来源。
类似地,切割半结构域可衍生自如上所述的任何核酸酶或其部分,其需要二聚化以用于切割活性。一般而言,若融合蛋白包含切割半结构域,则需要两种融合蛋白供于切割。或者,可使用包含两个切割半结构域的单个蛋白。这两个切割半结构域可衍生自同一核酸内切核酸酶(或其功能性片段),或各切割半结构域可衍生自不同的核酸内切核酸酶(或其功能性片段)。此外,优选两种融合蛋白的靶位点相对彼此布置,从而这两种融合蛋白与其各自靶位点的结合使切割半结构域彼此处于允许切割半结构域形成功能性切割结构域(例如,通过二聚化)的空间定位。因此,在某些实施方式中,这些靶位点的邻近边缘间隔有5-8个核苷酸或15-18个核苷酸。然而,可在两个靶位点之间介入任何整数个核苷酸或核苷酸对(例如,2-50个核苷酸对或更多)。一般而言,切割的位点位于靶位点之间,但可能位于距离该切割位点一或多个千碱基的位置,包括距离该切割位点1-50个碱基对(或其间的任何值,包括1-5个、1-10个和1-20个碱基对)之间、1-100个碱基对(或其间的任何值)之间、100-500个碱基对(或其间的任何值)之间、500-1000个碱基对(或其间的任何值)之间或甚至超过1kb之间。
限制性内切核酸酶(限制性酶)存在于许多物种中,其能够序列特异性结合dna(在识别位点处),并在结合位点处或其附近切割dna。某些限制性酶(例如,iis型)在从识别位点移除的位点处切割dna并具有可分开的结合与切割结构域。例如,iis型酶foki催化dna的双链切割,其中一条链在距离其识别位点9个核苷酸处被切割,另一条链在距离其识别位点13个核苷酸处被切割。参见例如,美国专利号5,356,802、5,436,150和5,487,994;以及li等,(1992)proc.natl.acad.sci.usa89:4275-4279;li等,(1993)proc.natl.acad.sci.usa90:2764-2768;kim等(1994a)proc.natl.acad.sci.usa91:883-887;kim等(1994b)j.biol.chem.269:31,978-31,982。因此,在一个实施方式中,融合蛋白包括来自至少一种iis型限制性酶的切割结构域(或切割半结构域)和一种或多种经或未经工程改造的锌指结合域。
切割结构域可与结合域分离的一个示例性iis型限制性酶是foki。该具体酶作为二聚体具有活性。bitinaite等,(1998)proc.natl.acad.sci.usa95:10,570-10,575。全长foki的序列如下所示。本文所述的核酸酶中所用的切割结构域以斜体和下划线(全长蛋白质的第384-579位)显示,其中完整蛋白质序列如下所示(seqidno:138):
因此,就本公开的目的而言,认为所述融合蛋白所用foki酶的部分是切割半结构域。因此,对于利用锌指-foki融合体的靶向双链切割和/或靶向细胞序列置换,可使用各自含有一个foki切割半结构域的两种融合蛋白重建催化活性的切割结构域。或者,也可使用包含锌指结合结构域和两个foki切割半结构域的单个多肽分子。使用锌指-foki融合体进行靶向切割和靶向序列改变的参数在本公开他处提供。
切割结构域或切割半结构域可以是蛋白质的任何部分,其保留了切割活性,或保留了多聚化(例如,二聚化)以形成功能性切割结构域的能力。
示例性iis型限制酶描述于国际专利公开号wo07/014275中,其通过引用其全文的方式纳入本文。其它限制性酶也包含可分开的结合和切割结构域,并且这些是本发明所设想的。参见例如roberts等.(2003)(2003)nucleicacidsres.31:418-420。
在某些实施方式中,切割结构域包含一个或多个经工程改造的切割半结构域(也称作二聚化结构域突变体),使同二聚作用最小化或被阻止,例如,如美国专利号7,914,796、8,034,598和8,623,618,以及美国专利公开号2011/0201055所述,其全部内容通过引用其全文方式纳入本文。如美国专利公开号2018/0087072中所述的〝sharkey″突变(例如418和441,相对于全长编号)和其它突变,例如对于残基416(例如r416s)和/或残基525(例如k525s)也可包括在此。因此,用于本发明的核酸酶的foki切割结构域可在以下一个或多个氨基酸残基位置(相对于全长编号)突变:416、418、441、446、447、479、483、484、486、487、490、491、496、498、499、500、525、531、534、537和/或538。
形成专性异二聚体的foki的示例性工程切割半结构域包括一对,其中第一切割半结构域包括foki的490和538位氨基酸残基处的突变,第二切割半结构域包括氨基酸残基486和499处的突变。
因此,在一实施方式中,490位的突变将glu(e)替换为lys(k);538位的突变将iso(i)替换为lys(k);486位的突变将gln(q)替换为glu(e);而499位的突变将iso(i)替换为lys(k)。具体地,制备本文所述经工程改造的切割半结构域是通过在一个切割半结构域中对位置490(e→k)和538(i→k)进行突变来产生名为″e490k:i538k″的经工程改造的切割半结构域,并通过另一切割半结构域中对位置486(q→e)和499(i→l)进行突变来产生名为″q486e:i499l″的经工程改造的切割半结构域。本文所述的经工程改造的切割半结构域是专性杂二聚体突变体,其中异常切割被最小化或废除。参见例如,美国专利号7,914,796和8,034,598,其公开内容通过引用其全文的方式纳入本文用于所有目的。在某些实施方式中,经工程改造的切割半结构域包括486、499和496位(根据野生型foki编号)的突变,例如突变将486位的野生型gln(q)残基替换为glu(e)残基,将499位的野生型iso(i)残基替换为leu(l)残基,以及将496位的野生型asn(n)残基替换为asp(d)或glu(e)残基(也分别称作〝eld″和〝ele″结构域)。在其它实施方式中,经工程改造的切割半结构域包括490、538和537位(相对于野生型foki编号)的突变,例如突变将490位的野生型glu(e)残基替换为lys(k)残基,将538位的野生型iso(i)残基替换为lys(k)残基,并且将537位的野生型his(h)残基替换为lys(k)或arg(r)残基(也分别称作〝kkk″和″kkr″结构域)。在其它实施方式中,经工程改造的切割半结构域包括在490和537位(相对于野生型foki编号)的突变,例如突变将490位的野生型glu(e)残基替换为lys(k)残基,并且将537位的野生型his(h)残基替换为lys(k)残基或arg(r)残基(也分别被称作″kik″和″kir″结构域)。
在其它实施方式中,经工程改造的切割半结构域包含487、499和496位(相对于野生型foki编号)处的突变,例如,突变将487位的野生型arg(r)残基替换为asp(d)残基,和499位的野生型ile(i)残基替换为ala(a),和496位的野生型asn(n)残基替换为asp(d)残基(也称为″dad″)和/或,483、538和537位(相对于野生型foki编号)处的突变,例如,突变将483位的野生型asp(d)残基替换为arg(r)残基,和538位的野生型ile(i)残基替换为val(v)残基,和537位的野生型his(h)残基替换为arg(r)残基(也称为″rvr″)。参见例如美国专利号8,962,281、7,914,796、8,034,598,其公开内容通过引用其全文的方式纳入本文用于所有目的。在其它实施方式中,经工程改造的切割半结构域包含″sharkey″和/或″sharkey″突变(参见guo等,(2010)j.mol.biol.400(1):96-107)。
因此,可用于本文所述的核酸酶的foki结构域的非限制性示例包括:表8所示的fok突变(例如eld、kkr等),foki-sharkey(s418p+k441e),fokield(486位q->e,499处i->l,496位n->d),fokield,sharkey(486位q->e,i->l499位,496位n->d,s418p+k441e),fokield,r416e(486位q->e,499位i->l,496位n->d,r416e),fokield,sharkey,r416e(486位q->e,499位i->l,496位n->d,s418p+k441e,r416e),fokield,r416y(486位q->e,499位i->l,496位n->d,r416y),fokield,sharkey,r416e(486位q->e,499位i->l,496位n->d,s418p+k441e,r416e),fokield,s418e(486位q->e,499位i->l,496位n->d,s418e),fokield,sharkey部分,s418e(486位q->e,499位i->l,496位n->d,k441e,s418e),fokield,k525s(486位q->e,499位i->l,496位n->d,k525s),fokield,sharkeyk525s(486位q->e,499位i->l,496位n->d,s418p+k441e,k525s),fokield,i479t(486位q->e,499位i->l,496位n->d,i479t),fokield,sharkey,i479t(486位q->e,499位i->l,496位n->d,s418p+k441e,i479t),fokield,p478d(486位q->e,499位i->l,496位n->d,p478d),fokield,sharkey,p478d(486位q->e,499位i->l,496位n->d,s418p+k441e,p478d),fokield,q481d(486位q->e,499位i->l,496位n->d,q481d),fokield,sharkey,q481d(486位q->e,499位i->l,496位n->d,s418p+k441e,q481d),fokikkr(490位e->k,538位i->k,537位h->r),fokikkrsharkey,(490位e->k,538位i->k,537位h->r,s418p+k441e),fokikkr,q481e(490位e->k,538位i->k,537位h->r,q481e),fokikkr,sharkeyq481e(490位e->k,538位i->k,537位h->r,s418p+k441e,q481e),fokikkr,r416e(490位e->k,538位i->k,537位h->r,r416e),fokikkr,sharkey,r416e(490位e->k,538位i->k,537位h->r,s418p+k441e,r416e),fokikkr,k525s(490位e->k,538位i->k,537位h->r,k525s),fokikkr,sharkey,k525s(490位e->k,538位i->k,537位h->r,s418p+k441e,k525s),fokikkr,r416y(490位e->k,538位i->k,537位h->r,r416y),fokikkr,sharkey,r416y(490位e->k,538位i->k,537位h->r,s418p+k441e,r416y),foki,kkri479t(490位e->k,538位i->k,537位h->r,i479t),foki,kkrsharkeyi479t(490位e->k,538位i->k,537位h->r,s418p+k441e,i479t,foki,kkrp478d(490位e->k,538位i->k,537位h->r,p478d),foki,kkrsharkeyp478d(490位e->k,538位i->k,537位h->r,p478d),fokidad(487位r->d,496位n->d,499位i->a),fokidadsharkey(487位r->d,496位n->d,499位i->a,s418p+k441e),fokirvr(483位d->r,537位h->r,538位i->v),fokirvrsharkey(483位d->r,537位h->r,538位i->v,s418p+k441e)。
本文所述的zfn还可包括任何接头序列,包括但不限于本文公开的序列(l0,n7a,n7c等)和/或美国专利号7,888,121、7,914,796、8,034,598、8,623,618、9,567,609;和美国公开号20170218349中公开的序列,其可用于dna结合结构域的n或c末端与foki切割结构域的n或c末端之间。
本文所述的zfn的zfp(包括经工程改造的和/或野生型切割结构域)还可包含修饰,以增加zfn(包括核酸酶对)对其预期靶标相对于其它非预期切割位点(称为脱靶位点)的特异性(参见美国专利公开号20180087072)。因此,本文所述的核酸酶可在dna结合结构域和切割结构域之间包含特异性接头;和/或可在其一个或多个dna结合结构域主链区中包含突变和/或在其核酸酶切割结构域中包含一个或多个突变,如上所述。这些核酸酶的zfp可包含zfpdna结合结构域(″zfp主链″)中的氨基酸突变,所述突变可与dna主链上的磷酸发生非特异性相互作用,但它们不包含dna识别螺旋的变化。因此,本发明包括zfp,其包含zfp主链中阳离子氨基酸残基的突变,这对于核苷酸靶标特异性不是必需的。在一些实施方式中,zfp主链中的这些突变包括将阳离子氨基酸残基突变为中性或阴离子氨基酸残基。在一些实施方式中,zfp主链中的这些突变包括将极性氨基酸残基突变为中性或非极性氨基酸残基。在优选的实施方式中,在相对于dna结合螺旋的(-5)位,(-9)位和/或(-14)位处发生突变。在一些实施方式中,锌指可在(-5),(-9)和/或(-14)处包含一个或多个突变。在其它实施方式中,多指型锌指蛋白中的一个或多个锌指可包含(-5),(-9)和/或(-14)中的突变。在一些实施方式中,将(-5),(-9)和/或(-14)处的氨基酸(例如精氨酸(r)或赖氨酸(k))突变为丙氨酸(a),亮氨酸(l),ser(s),asp(n),glu(e),tyr(y)和/或谷氨酰胺(q)。
在某些实施例中,zfn包括以下对中的至少一对:68796和68813;68796和68861;68812和68813;68876和68877;68815和55266;68879和55266;68798和68815;或68846和53853,如表6所示。在其它实施方式中,zfn包含以下对中的至少一对:57531和72732;57531和72748;68957和57071;68957和72732;68957和72748;72678和57071;72678和72732;或72678和72748,如表8中所示。
或者,核酸酶可利用称为″分裂-酶(split-enzyme)″的技术(参见例如,美国专利公开号2009/0068164)在核酸靶位点处体内组装。这类分裂酶的组件可在另外的表达构建物上表达,或者可以连接于单独的组件相互分开的某一开放读框中,例如,组件由自切割2a肽或ires序列分开。组件可以是单独的锌指结合结构域或兆核酸酶核酸结合结构域的结构域。
核酸酶(例如,zfn和/或talen)可在使用前针对活性进行筛选,例如,在如美国专利号8,563,314中所述的基于酵母的染色体系统中。
在某些实施方式中,核酸酶包括crispr/cas系统。编码该系统的rna组分的crispr(规律成簇的间隔短回文重复序列)基因座,以及编码蛋白质的cas(crispr相关)基因座(jansen等.(2002)mol.microbiol.43:1565-1575;makarova等.(2002)nucleicacidsres.30:482-496;makarova等.(2006)biol.direct1:7;haft等.(2005)ploscomput.biol.1:e60)组成了crispr/cas核酸酶系统的基因序列。微生物宿主中的crispr基因座包含crispr-相关(cas)基因以及能够对crispr介导的核酸切割的特异性进行编程的非编码rna元件的组合。
ii型crispr是最充分表征的系统之一,并且以4个连续步骤进行靶向dna双链断裂。首先,从crispr基因座转录两个非编码rna:前-crrna阵列和tracrrna。第二,tracrrna杂交至前-crrna的重复序列区域,并且介导从前-crrna到含个体间隔子序列的成熟crrna的过程。第三,成熟的crrna:tracrrna复合物经由沃森-克里克碱基配对指导cas至靶dna,其位于crrna上的间隔子和原型间隔子邻近基序(pam)旁边的靶dna上的原型间隔子之间,所述pam是靶标识别的额外要求。最后,cas9介导靶dna的切割,以在原型间隔子内产生双链断裂。crispr/cas系统活性包括3步:(i)通过称作〝适应″的过程将外来dna序列插入crispr阵列,以防止未来的攻击,(ii)表达相关蛋白质,以及表达和处理所述阵列,然后(iii)采用外来核酸进行rna介导的干扰。因此,在细菌细胞中,多个所谓的〝cas″蛋白参与crispr/cas系统的自然功能,并且在多种功能中发挥作用(例如外来dna的插入等)。
在某些实施方式中,cas蛋白可为天然产生的cas蛋白的″功能性衍生物″。天然序列多肽的〝功能性衍生物″是与天然序列多肽具有共同定性生物性质的化合物。″功能性衍生物″包括但不限于天然序列的片段或天然序列多肽的衍生物及其片段,前提是它们与相应的天然序列多肽具有共同的生物活性。本文设想的生物活性是功能性衍生物将dna底物水解成片段的能力。术语″衍生物″涵盖多肽的氨基酸序列变体,共价修饰形式,和其融合体。cas多肽的合适衍生物或其片段包括但不限于cas蛋白的突变体、融合体、共价修饰形式或其片段。包括cas蛋白或其片段的cas蛋白以及cas蛋白或其片段的衍生物可获自细胞或通过化学合成或通过这两种方法的组合获得。所述细胞可以是天然产生cas蛋白的细胞,或天然产生cas蛋白并经遗传工程改造的细胞,所述遗传工程改造使所述细胞以较高表达水平产生内源性cas蛋白或从外源导入的核酸产生cas蛋白,其中所述核酸编码与内源性cas相同或不同的cas。在一些情况中,细胞天然不产生cas蛋白,但是经遗传工程改造以产生cas蛋白。
靶向tcr基因及其它基因的示例性crispr/cas核酸酶系统公开于例如美国专利公开号2015/0056705中。
核酸酶可在靶位点中产生一个或多个双链和/或单链切割。在某些实施方式中,核酸酶包含催化灭活的切割结构域(例如,foki和/或cas蛋白)。参见例如美国专利号9,200,266和8,703,489,和guillinger等.(2014)naturebiotech.32(6):577-582。催化灭活的切割结构域可与具有催化活性的结构域组合,作为切口酶以产生单链切割。因此,可将两种切口酶联用以在特定区域产生双链切割。本领域已知其它切口酶,例如,amccaffery等(2016)nucleicacidsres.44(2):ell.doi:10.1093/nar/gkv878.电子出版于2015年10月19日。此外,可使无活(dead)cas(`dcas′)或cas切口酶融合至碱基修饰酶(例如,胞苷脱氨酶),以产生碱基编辑系统(komor等.(2016)nature533:420)。这些系统允许碱基编辑复合物改变dna碱基(修饰),而不会在dna中产生双链断裂。因此,在一些实施方式中,向导rna(表2)可用于在trac基因中引入突变以造成敲除。
递送
蛋白质(例如,转录因子、核酸酶、tcr和car分子)、多核苷酸和/或包含本文所述的蛋白质和/或多核苷酸的组合物,可通过任何合适的手段递送至靶细胞,包括例如,通过注射蛋白质和/或mrna组分。在一些实施方式中,蛋白质通过细胞挤压(cellsqueezing)引入细胞(参见kollmannsperger等(2016)natcomm7,10372doi:10.1038/ncomms10372)。
合适的细胞包括但不限于真核和原核细胞和/或细胞系。由此类细胞产生的此类细胞或细胞系的非限制性示例包括t细胞、cos、cho(例如,cho-s、cho-k1、cho-dg44、cho-duxb11、cho-dukx、chok1sv)、vero、mdck、wi38、v79、b14af28-g3、bhk、hak、ns0、sp2/0-ag14、hela、hek293(例如,hek293-f、hek293-h、hek293-t)和perc6细胞,以及昆虫细胞例如草地贪夜蛾(spodopterafugiperda)(sf),或真菌细胞例如酵母(saccharomyces)、毕赤酵母(pichia)和裂殖酵母(schizosaccharomyces)。在某些实施方式中,细胞系是cho-k1、mdck或hek293细胞系。合适的细胞还包括干细胞,例如,胚胎干细胞、诱导型多能干细胞(ips细胞)、造血干细胞、神经元干细胞和间充质干细胞。
递送含本文所述dna结合结构域的蛋白质的方法见述于,例如,美国专利号6,453,242;6,503,717;6,534,261;6,599,692;6,607,882;6,689,558;6,824,978;6,933,113;6,979,539;7,013,219和7,163,824,其全部公开内容通过引用其全文的方式纳入本文。
dna结合结构域和含本文所述这些dna结合结构域的融合蛋白还可采用包含编码一种或多种dna结合蛋白的序列的载体递送。此外,还可通过这些载体递送其它核酸(例如,供体)。可采用任何载体系统,包括但不限于,质粒载体、逆转录病毒载体、慢病毒载体、腺病毒载体、痘病毒载体、疱疹病毒载体和腺相关病毒载体等。还参见例如,美国专利号6,534,261、6,607,882、6,824,978、6,933,113、6,979,539、7,013,219;和7,163,824,其通过引用其全文的方式纳入本文。此外,应明了,这些载体中的任一种可包括一种或多种dna结合蛋白-编码序列和/或其它合适的核酸。因此,当本文所述的一种或多种dna结合蛋白被引入细胞时,可与其它合适的dna被相同载体或不同载体携载。当采用多重载体时,各载体可以包含编码一种或多种dna结合蛋白的序列,以及所需的其它核酸。
可采用传统的基于病毒和非病毒的转基因方法来将编码经工程改造的dna结合蛋白的核酸引入细胞(例如,哺乳动物细胞)和靶组织,并且共同引入所需的其它核苷酸序列。此类方法也可用于将核酸(例如,编码dna结合蛋白和/或供体的核酸)体外给予细胞。在某些实施方式中,给予的核酸用于体内或离体基因治疗应用。非病毒载体递送系统包括dna质粒、裸核酸和与递送载剂(例如脂质体、脂质纳米颗粒或泊咯沙姆(poloxamer))复合的核酸。病毒载体递送系统包括dna和rna病毒,其在递送至细胞后具有附加型或整合的基因组。基因治疗操作的综述见于anderson(1992)science256:808-813;nabel和felgner(1993)tibtech11:211-217;mitani和caskey(1993)tibtech11:162-166;dillon(1993)tibtech11:167-175;miller(1992)nature357:455-460;vanbrunt(1988)biotechnology6(10):1149-1154;vigne(1995)restorativeneurologyandneuroscience8:35-36;kremer和perricaudet(1995)britishmedicalbulletin51(1):31-44;haddada等.(1995)currenttopicsinmicrobiologyandimmunologydoerfler和
核酸的非病毒递送方法包括电穿孔、脂质转染、显微注射、基因枪、病毒颗粒、脂质体、免疫脂质体、聚阳离子或脂质:核酸偶联物、裸dna、mrna、人工病毒粒,和试剂增强的dna摄取。利用例如sonitron2000系统(rich-mar)的声孔效应也可用于递送核酸。在一个优选实施方式中,一种或多种核酸以mrna形式递送。还优选采用加帽的mrna来增加翻译效率和/或mrna稳定性。尤其优选arca(防倒转帽类似物)帽或其变化形式。参见美国专利号7,074,596和8,153,773,其通过引用纳入本文。
其它示例性的核酸递送系统包括amaxa生物系统公司(德国科隆)、迈克赛特公司(maxcyte,inc.)(马里兰州罗克韦尔)、btx分子递送系统公司(马萨诸塞州霍利斯顿)和哥白尼治疗公司(copernicustherapeuticsinc.)(参见例如美国专利号6,008,336)提供的那些。脂质转染描述于例如,美国专利号5,049,386、4,946,787和4,897,355,并且脂质转染试剂市售可得(例如,transfectamtm、lipofectintm和lipofectaminetmrnaimax)。适于多核苷酸的高效受体-识别脂质转染的阳离子和中性脂质包括felgner、国际专利公开号wo91/17424和wo91/16024的那些。可以递送至细胞(离体给予)或靶组织(体内给予)。
脂质:核酸复合物(包括靶向的脂质体,例如免疫脂质复合物)的制备是本领域技术人员熟知的(参见例如,crystal(1995)science270:404-410;blaese等.(1995)cancergenether.2:291-297;behr等.(1994)bioconjugatechem.5:382-389;remy等.(1994)bioconjugatechem.5:647-654;gao等.(1995)genetherapy2:710-722;ahmad等.(1992)cancerres.52:4817-4820;美国专利号4,186,183;4,217,344;4,235,871;4,261,975;4,485,054;4,501,728;4,774,085;4,837,028;和4,946,787)。
递送的其它方法包括将待被递送的核酸包装到engeneic递送载体(edv)中。采用双特异性抗体将这些edv被特异性递送至靶组织,其中,所述抗体的一个臂对靶组织具有特异性,而另一个臂对edv具有特异性。抗体将edv带至靶细胞表面,然后edv通过内吞作用进入细胞。一旦进入细胞,内容物即被释放(参见macdiarmid等(2009)naturebiotechhology27(7):643)。
使用基于rna或dna病毒的系统递送编码经工程改造的dna结合蛋白的核酸和/或所需要的供体(例如,car或actr)利用高度演化的过程来将病毒靶向身体中特定的细胞并且将该病毒负载物(payload)运输至核。病毒载体可直接给予患者(体内)或其可用于体外处理细胞,并将经修饰的细胞给予患者(离体)。用于递送核酸的常规的基于病毒的系统包括但不限于,用于基因传递的逆转录病毒、慢病毒、腺病毒、腺相关、牛痘和单纯性疱疹病毒载体。宿主基因组中的整合可采用逆转录病毒、慢病毒和腺相关病毒基因传递方法进行,通常导致插入的转基因的长期表达。此外,已在许多不同的细胞类型和靶组织中观察到了高转导功效。
逆转录病毒的趋性可通过引入外来包膜蛋白、扩大靶细胞的潜在靶群体来改变。慢病毒载体是能够转导或感染非分裂细胞的逆转录病毒载体,并且通常产生高病毒效价。逆转录病毒转基因系统的选择取决于靶组织。逆转录病毒载体包含顺式作用的长末端重复,其具有包装长达6-10kb的外来序列的能力。最小的顺式作用的ltr足以用于载体的复制和包装,其随后用于将治疗基因整合进入靶细胞,以提供永久的转基因表达。广泛应用的逆转录病毒载体包括基于鼠白血病病毒(mulv)、长臂猿白血病病毒(galv)、类人猿免疫缺陷型病毒(siv)、人类免疫缺陷型病毒(hiv),及其组合的那些(参见例如,buchscher等.(1992)j.virol.66:2731-2739;johann等.(1992)j.virol.66:1635-1640;sommerfelt等.(1990)virol.176:58-59;wilson等.(1989)j.virol.63:2374-2378;miller等.(1991)j.virol.65:2220-2224;国际专利公开号wo1994/026877)。
在其中优选瞬时表达的应用中,可采用基于腺病毒的系统。基于腺病毒的载体能够在许多细胞类型中获得极高的转导效率,并且不需要细胞分裂。采用这种载体,已获得了高效价和高水平的表达。该载体可在相对简单的系统中大量产生。腺相关病毒(″aav″)载体也用于用靶核酸转导细胞,例如,用于核酸和肽的体外生成,以及用于体内和离体基因治疗法(参见例如,west等.,virology160:38-47;美国专利号4,797,368;国际专利公开号wo93/24641;kotin(1994)humangenetherapy5:793-801;muzyczka(1994)j.clin.invest.94:1351。重组aav载体的构建描述于多个公开文本中,包括美国专利号5,173,414;tratschin等.(1985)mol.cell.biol.5:3251-3260;tratschin等.(1984)mol.cell.biol.4:2072-2081;hermonat和muzyczka(1984)pnasusa81:6466-6470;和semulski等.(1989)j.virol.63:03822-3828。
目前有至少六种病毒载体法可用于临床试验中的基因传递,其采用通过将基因插入辅助细胞系来产生转导剂进行涉及缺陷型载体的互补的方法。
plasn和mfg-s是已用于临床试验的逆转录病毒载体的示例(dunbar等.(1995)blood85:3048-305;kohn等.(1995)nat.med.1:1017-102;malech等.(1997)pnasusa94:2212133-12138)。pa317/plasn是用于基因治疗试验的第一治疗载体。(blaese等.science270:475-480)。已在mfg-s包装的载体中观察到50%或更高的转导功效。(ellem等.(1997)immunolimmunother.44(1):10-20;dranoff等.(1997)hum.genether.1:111-2。
重组腺相关病毒载体(raav)是一种具有前景的替代性基因递送系统,其基于缺陷型和非病原性细小病毒腺相关2型病毒。全部载体源自仅保留侧接转基因表达盒的aav145bp反向末端重复序列的质粒。高效的基因转移和稳定的转基因递送是该载体系统的关键特征,这归因于向转导的细胞的基因组的整合。(wagner等.(1998)lancet351(9117):1702-3,kearns等.(1996)genether.9:748-55)。根据本发明也可使用其它aav血清型,包括aav1、aav3、aav4、aav5、aav6、aav8、aav8.2、aav9和aavrh10和假型化的aav,如aav2/8、aav2/5和aav2/6。
复制缺陷型重组腺病毒载体(ad)可以高效价生成,并且容易地感染多种不同的细胞类型。大多数腺病毒载体是经工程改造的,从而转基因替代了ade1a、e1b和/或e3基因;随后该复制缺陷型载体在提供缺失反式基因功能的人293细胞中传播。ad载体可体内转导多种类型的组织,包括不分裂的、分化的细胞,例如发现于肝、肾和肌肉中的那些。传统的ad载体具有较大携载能力。临床试验中ad载体应用的一个示例涉及用于采用肌内注射进行抗肿瘤免疫的多核苷酸治疗(sterman等.(1998)hum.genether.7:1083-9)。临床试验中用于转基因的腺病毒载体的应用的其它示例包括rosenecker等.(1996)infection24(1):5-10;sterman等.(1998)hum.genether.9(7):1083-1089;welsh等.(1995)hum.genether.2:205-18;alvarez等.(1997)hum.genether.5:597-613;topf等.(1998)genether.5:507-513;sterman等.(1998)hum.genether.7:1083-1089。
采用包装细胞来形成能够感染宿主细胞的病毒颗粒。所述细胞包括293细胞,其包装腺病毒,和ψ2细胞或pa317细胞,其包装逆转录病毒。用于基因治疗的病毒载体通常由将核酸载体包装成病毒颗粒的生产细胞系产生。所述载体通常包含包装和后续整合进入宿主(若可行)所需的最小病毒序列,其它病毒序列被编码待表达的蛋白质的表达盒所替代。失去的病毒功能通过包装细胞系以反式提供。例如,用于基因治疗的aav载体通常仅加工来自aav基因组的反向末端重复(itr)序列,其为包装和整合进入宿主基因组所需。病毒dna被包装进入细胞系,其包含编码其它aav基因(即rep和cap)但缺乏itr序列的辅助质粒。所述细胞系也用腺病毒(作为辅助物)感染。辅助病毒促进aav载体的复制和aav基因从辅助质粒的表达。因为缺乏itr序列,辅助质粒不以显著的量包装。腺病毒的污染可通过,例如,热处理(相比aav,腺病毒对热处理更为敏感)来减少。此外,可以使用杆状病毒系统生产avv(参见例如,美国专利号6,723,551和7,271,002)。
来自293或杆状病毒系统的aav颗粒的纯化通常涉及使产生病毒的细胞生长,然后由细胞上清液收集病毒颗粒,或裂解细胞并从粗裂解物收集病毒。然后,aav通过本领域已知方法纯化,包括离子交换色谱(例如,参见美国专利号7,419,817和6,989,264),离子交换色谱和csc1密度离心(例如,国际专利公开号wo2011/094198a10),免疫亲和色谱(例如,国际专利公开号wo2016/128408)或采用avb琼脂糖凝胶(sepharose)(例如,通用电气医疗集团生命科学部(gehealthcarelifesciences))的纯化。
在许多基因治疗应用中,希望将基因治疗载体以高度特异性递送至特定组织类型。因此,病毒载体可经修饰以通过将配体表达成与病毒包衣蛋白融合的位于病毒外表面上的蛋白来具有针对给定细胞类型的特异性。选择对于已知存在于感兴趣的细胞类型上的受体具有亲和性的配体。例如,han等(1995),proc.natl.acad.sci.usa92:9747-9751(1995),报道了莫洛尼鼠白血病病毒可经修饰以表达融合至gp70的人调蛋白,并且该重组病毒感染表达人表皮生长因子受体的某些人乳腺癌细胞。该原理可延伸至其它病毒-靶细胞对,其中靶细胞表达受体,并且病毒表达包含针对该细胞表面受体的配体的融合蛋白。例如,丝状噬菌体可经工程改造以显示对于几乎任何所选细胞受体具有特异性结合亲和力的抗体片段(例如,fab或fv)。尽管上文描述主要应用至病毒载体,相同的原理可应用至非病毒载体。所述载体可经工程改造以包含特定的摄取序列,其有利于被特定靶细胞摄取。
基因治疗载体可通过给予个体患者来体内递送,通常通过全身性给予(例如,静脉内、腹膜内、肌内、皮下,或颅内输注)或局部施用,如下所述。或者,载体可离体递送至细胞,例如来自个体患者的外植的细胞(例如,淋巴细胞、骨髓抽出物、组织活检物)或通用供体造血干细胞,然后将所述细胞再植入患者,通常在选择已引入载体的细胞之后进行。
本文所述的细胞还可以用于细胞治疗,例如用于治疗和/或预防癌症的过继细胞治疗。细胞治疗是一种特殊类型的移植,其中将某种类型的细胞(例如,对肿瘤抗原具有反应性的t细胞或b细胞)提供给受体。细胞治疗可以使用自体同源(来自受体)或同种异体(来自供体)的细胞完成,并且细胞可以是未成熟的细胞(例如干细胞)或完全成熟且有功能的细胞(例如t细胞)。实际上,在某些疾病(如某些癌症)中,t细胞可经离体操纵以增加其对某些肿瘤抗原的亲和力,扩增,然后将其引入患有该癌症类型的患者中以尝试根除肿瘤。当内源性t细胞反应被肿瘤本身抑制时,这特别有用。
用于诊断、研究、移植或用于基因和/或细胞治疗(例如,通过重新输注转染的细胞进入宿主生物体)的离体细胞转染是本领域技术人员熟知的。在一个优选实施方式中,细胞从对象机体分离,用dna结合蛋白核酸(基因或cdna)转染,并重新输注回对象机体(例如,患者)。合适于离体转染的多种细胞类型是本领域技术人员熟知的(参见例如,freshney等,《动物细胞培养,基础技术手册》(cultureofanimalcells,amanualofbasictechnique)(第三版.1994)),本文所引的参考内容是关于讨论如何分离和培养来自患者的细胞)。
在一个实施方式中,细胞转染和基因治疗的离体方案中采用干细胞。采用干细胞的好处是,它们可以体外分化成其它细胞类型,或可被导入哺乳动物(例如细胞的供体),其中这些细胞将被植入骨髓。采用细胞因子例如gm-csf、ifn-γ和tnf-α将cd34+细胞体外分化成临床上重要的免疫细胞类型的方法是已知的(参见inaba等.(1992)j.exp.med.176:1693-1702)。
采用已知方法分离干细胞用于转导和分化。例如,干细胞也可通过骨髓细胞用结合不需要细胞的抗体筛选而从骨髓细胞中分离,所述细胞例如cd4+和cd8+(t细胞)、cd45+(panb细胞)、gr-1(粒细胞)和iad(分化抗原呈递细胞)(参见inaba等.(1992)j.exp.med.176:1693-1702)。
在一些实施方式中,可采用已经修饰的干细胞。例如,可将被制成抗凋亡的神经元干细胞用作治疗组合物,其中所述干细胞还包含本发明的zfptf。对于凋亡的抗性可来自于,例如,通过采用bax-或bak特异性zfn敲除干细胞中的bax和/或bak(参见,美国专利申请号8,597,912),或干扰胱冬酶的那些,同样采用例如胱冬酶6特异性zfn。可以用已知调节tcr的zfptf转染这些细胞。
也可将包含治疗性dna结合蛋白(或编码这些蛋白质的核酸)的载体(例如,逆转录病毒、腺病毒、脂质体等)直接给予机体用于体内细胞转导。或者,可给予裸dna。给予通过常用于导入分子并最终与血液或组织细胞接触的任何常规途径进行,包括但不限于,注射、输注、局部施用和电穿孔。合适的给予所述核酸的方法可得且为本领域技术人员熟知,并且,尽管可利用超过一种途径给予特定组合物,某特定途径通常能提供比另一途径更直接和更有效的反应。
用于将dna导入造血干细胞的方法公开于,例如,美国专利号5,928,638。可用于将转基因引入造血干细胞(例如,cd34+细胞)的载体包括腺病毒第35型。
合适于将转基因引入免疫细胞(例如,t细胞)的载体包括非整合型慢病毒载体。参见例如ory等.(1996)proc.natl.acad.sci.usa93:11382-11388;dull等.(1998)j.virol.72:8463-8471;zuffery等.(1998)j.virol.72:9873-9880;follenzi等.(2000)naturegenetics25:217-222。
药学上可接受的运载体部分地由所给予的特定组合物以及用于给予组合物的特定方法确定。因此,有多种合适的药物组合物制剂可用,如下所述(参见例如,《雷明顿药物科学》(remington′spharmaceuticalsciences);第17版,1989)。
如上所述,公开的方法和组合物可用于任何类型的细胞,包括但不限于,原核细胞、真菌细胞、古细菌细胞、植物细胞、昆虫细胞、动物细胞、脊椎动物细胞、哺乳动物细胞和人细胞,包括任何类型的t细胞和干细胞。用于蛋白质表达的合适的细胞系是本领域技术人员已知的,并且可包括但不限于cos、cho(例如,cho-s、cho-k1、cho-dg44、cho-duxb11)、vero、mdck、wi38、v79、b14af28-g3、bhk、hak、nso、sp2/0-agl4、hela、hek293(例如,hek293-f、hek293-h、hek293-t)、perc6、昆虫细胞例如草地贪夜蛾(spodopterafugiperda)(sf),和真菌细胞例如酵母(saccharomyces)、毕赤酵母(pichia)和裂殖酵母(schizosaccharomyces)。也可采用这些细胞系的子代、变体和衍生物。
应用
所公开的组合物和方法可用于需要调节tcr和/或b2m表达和/或功能的任何应用,包括但不限于其中期望tcr和/或b2m调节的治疗和研究应用。例如,公开的组合物可体内和/或离体(细胞治疗)使用,以破坏t细胞中内源性tcr和/或b2m的表达,所述t细胞经修饰以用于过继细胞治疗以表达一种或多种外源car,外源tcr,或其它癌症特异性受体分子,由此治疗和/或预防癌症。t细胞可以是效应t细胞或调节性t细胞。另外,在这种情况下,细胞内tcr表达的废除可消除或显著降低与健康的非靶向组织的不希望的交叉反应(即,移植物抗宿主反应)的风险。本文所述的经修饰细胞也可用于治疗癌症,包括但不限于前列腺癌,慢性淋巴细胞性白血病(cll)和非霍奇金淋巴瘤。
方法和组合物还包括干细胞组合物(例如,ipsc和hsc/hspc),其中干细胞内的b2m、tcra和/或tcrb基因已被调节(修饰),并且细胞进一步包含actr和/或car和/或分离或经工程改造的tcr。例如,可在骨髓消融后将tcr基因敲除或基因敲减调节的同种异体造血干细胞引入hla匹配的患者中。这些改变的hsc将允许患者的再定殖(re-colonization),但将不会引起潜在的gvhd。引入的细胞还可能具有其它改变以在后续治疗(例如,化疗耐药性)过程中协助治疗潜在的疾病。hlai类无效细胞还可以在有创伤患者的急诊室情况下用作〝现成″疗法。
本发明的方法和组合物也可用于设计和获得体外和体内模型,例如,tcr或b2m和相关病症的动物模型,其允许研究这些病症。
本文中提及的所有专利、专利申请和出版物均以参考的方式用全文纳入本文。
尽管出于理解和清楚的目的,本发明通过说明和示例的方式提供了一些细节,但本领域技术人员可理解在不偏离本发明的精神或范围的情况下可实施各种改变和改进。因此,上述说明和实施例不应理解为是限制性的。
实施例
实施例1:tcr特异性核酸酶的设计
构建tcr特异性zfn,以在tcrα(tcra)基因处位点特异性地引入双链断裂。zfn的设计基本见述于,urnov等.(2005)nature435(7042):646-651,lombardo等.(2007)natbiotechhol.25(11):1298-306,和美国专利公开号2008/0131962;2015/016495;2014/0120622;和2014/0301990和美国专利号8,956,828。zfn对靶向tcra基因恒定区的不同位点(参见图1)。示例性zfn对的识别螺旋以及靶序列示于下表1。tcra锌指设计物的靶位点示于第一列。被zfp识别螺旋靶向的靶位点中的核苷酸以大写字母表示;未被靶向的核苷酸以小写字母指示。用于连接foki核酸酶结构域和zfpdna结合结构域的接头也同样示出(参见,美国专利公开号2015/0132269)。例如,结构域接头l0的氨基酸序列是dna结合结构域-qlvks-foki核酸酶结构域(seqidno:5)。类似地,结构域接头n7a的氨基酸序列是foki核酸酶结构域-sgtphevgvytl-dna结合结构域(seqidno:6),并且n7c是foki核酸酶结构域-sgairchdefwf-dna结合结构域(seqidno:7)。
表1:tcr-α(tcra)锌指设计物
测试所有的zfn,且发现它们结合至它们的靶位点,并发现具有核酸酶活性。
本文所述的zfp还可包含锌指蛋白和/或foki结构域的磷酸接触残基的一个或多个突变,例如,nr-5qabc突变体(至zfp主链)和/或r416s和/或k525s突变体(至foki),描述于美国专利公开号20180087072。
还构建了用于酿脓链球菌(s.pyogenes)crispr/cas9系统的向导rna,用于靶向tcra基因。关于其它tcrα-靶向的向导rna,还参见美国专利公开号2015/00566705。下表2中指示了tcra基因中的靶序列以及向导rna序列。在crispr/cas9系统中测试所有的向导rna,并且发现其具有活性。
表2:针对人tcra恒定区(trac)的向导rna
因此,本文所述的核酸酶(例如,含zfp或sgrnadna结合结构域的核酸酶)结合至其靶位点,并切割tcra基因,由此产生包含seqidno:6-48或137-205任一者的tcra基因中的遗传修饰,包括这些序列之任一者(例如seqidno:8-21和/或92-103任一所示的靶序列;这些靶位点的12-25个核苷酸;和/或成对靶位点之间)内的修饰(插入和/或缺失),和/或以下序列内的修饰:aacagt、agtgct、ctcct、ttgaaa、tggactt和/或aatcctc(参见图1b)。还设计了靶向这些靶位点的tale核酸酶,并且发现其在结合和活性方面具有功能性。
此外,当与一个或多个转录调节结构域相关联时,所有与其靶位点结合的dna结合结构域(zfp和sgrna)以及识别这些靶位点的zfp、tale和srnadna结合结构域还形成有活性的经工程改造的转录因子。
实施例2:体外核酸酶活性
表1中描述的zfn被用于测试k562细胞中的核酸酶活性。为了测试切割活性,将编码上述成对人tcra特异性zfn对的质粒转染到具有质粒或mrna的k562细胞中。k562细胞获自美国典型培养物保藏中心,并按建议在补充有10%合格胎牛血清(fbs,cyclone)的rpmi培养基(英杰公司(invitrogen))中生长。对于转染,将表1所列的活性核酸酶的orf克隆到表达载体中,该表达载体针对mrna生产优化,包含5′和3′utr和合成的多聚a信号。按照生产商的说明,使用mmessagemmachinet7ultra试剂盒(安碧公司(ambion))生成mrna。核酸酶mrna的体外合成使用基于pvax的载体,或基于pgem的载体,或pcr扩增子,所述基于pvax的载体包含t7启动子、合适的核酸酶和多聚a基序,用于在体外转录反应后酶促添加多聚a尾,所述基于pgem的载体包含t7启动子、5′utr、合适的核酸酶、3′utr以及64bp的多聚a延伸段,所述pcr扩增子包含t7启动子、5′utr、合适的核酸酶、3′utr以及60bp的多聚a延伸段。将一百万个k562细胞与250ng或500ngzfn编码mrna混合。使用程序t-16将细胞转染至amaxanucleofectoriitm中,并且回收到1.4ml温热pmi培养基+10%fbs中。转染后三天,按照标准规程通过深度测序(miseq,illumina)评估核酸酶活性。结果见下表3。
表3:锌指核酸酶活性
高活性的tcra特异性talen也已于先前描述(参见国际专利公开号wo2014/153470)。
还测试了人tcra-特异性crispr/cas9系统。人k562细胞中crispr/cas9系统的活性通过miseq分析测量。通过cas9的内源性tcradna序列的切割通过高通量测序(miseq,亿明达(illumina))分析。
在这些实验中,cas9在pvax质粒上提供,sgrna在启动子(例如,u6启动子或cmv启动子)的控制下在质粒上提供。质粒以各100ng或各400ng混合,并且每轮与2e5个细胞混合。使用amaxa系统转染细胞。简言之,使用amaxa转染试剂盒,并使用标准的amaxa穿梭方案转染核酸。转染后,让细胞在室温下静置10分钟,然后在预热的rpmi中重悬。然后使细胞在标准条件下于37℃下生长。转染后7天分离基因组dna,并且进行miseq分析。
简言之,测试表2中所列的向导rna的活性。向导rna在三种不同的结构中测试:g0如上所述设置。g1使用pvax载体,其包括驱动cas9基因表达的cmv启动子以及u6-向导rna-示踪剂表达盒,其中两个阅读框的转录以相同的方向进行。g2与g1相似,不同之处在于cas9和u6-向导表达盒以相反的方向设置。使用100ng或400ng转染的dna对这三种设置进行了测试,结果列于下表4中。结果以″插入缺失(indel)百分比″或″nhej%″来表示,其中″插入缺失″表示由于核酸酶诱导的双链切割的位点处的易错nhej修复过程所致的小的插入和/或缺失。
表4:crispr/cas活性
如其所示,本文所述核酸酶包括位于靶向位点的切割和基因组修饰。
因此,本文所述核酸酶(例如,包括zfp、tale或sgrnadna结合结构域的核酸酶)结合至其靶位点并且切割tcra基因,由此在包括seqidno:8-21或92-103中任一者的tcra基因中产生遗传修饰,包括在这些序列(seqidno:8-21,92-103)之任一者内的修饰(插入和/或缺失);这些基因序列的1-50个(例如,1-10个)碱基对内的修饰;配对的靶位点(对于二聚体)的靶位点之间的修饰;和/或下述序列之一或多者内的修饰:aacagt、agtgct、ctcct、ttgaaa、tggactt和/或aatcctc(参见,图1b)。
此外,当与一个或多个转录调节结构域相关联时,所有dna结合结构域(zfp、tale和sgrna)与其靶位点结合,并且还形成有活性的经工程改造的转录因子。
实施例3:t细胞中的tcra特异性zfn活性
还在人t细胞中测试tcra特异性zfn对的核酸酶活性。将编码zfn的mrna转染到纯化的t细胞中。简言之,从白细胞除去法(leukopheresis)产物中获得t细胞,并且使用美天旎(miltenyi)公司的clinimacs系统(cd4和cd8双选)纯化。然后,根据生产商方案,使用dynabeads(赛默飞世尔公司(thermofisher))活化这些细胞。活化后3天,用三个剂量的mrna(60、120和250μg/ml)转染细胞,采用maxcyte电穿孔仪(maxcyte),oc-100,30e6个细胞/ml,体积为0.1ml。在转染后第10天,使用深度测序(miseq,illumina)分析细胞的中靶(ontarget)tcra修饰。在整个培养的13-14天中测量细胞活力和细胞生长(总细胞倍增)。此外,在针对cd3染色培养的第10天,使用标准facs分析,测量处理的细胞的细胞表面上的tcr。
所有的tcra特异性zfn对都在t细胞中具有活性,并且一些tcra特异性zfn对能在这些条件中造成超过80%的tcra等位基因修饰(参见,图2a和2b)。类似地,用zfn处理的t细胞失去了cd3的表达,其中facs分析显示在一些情况中,80-90%的t细胞呈cd3阴性(图3)。这些细胞中被zfn修饰的tcra百分比与cd3损失之间的比较显示出高度的相关性(图4)。细胞活力与模拟处理对照相当,并且tcra敲除细胞生长也与对照相当(参见图5a-5d)。
实施例4:采用靶向整合的b2m和tcra双敲除
采用如上所述的核酸酶和表5所述的b2m靶向核酸酶(也参见美国专利公开号2017/0173080)来使b2m和tcra失活,并通过靶向整合将供体(转基因)引入tcra或b2m基因座。b2m特异性zfn如下表5所示:
表5:b2m特异性zfn设计物
在该实验中,tcra特异性zfn对是sbs#55266/sbs#53853,其包含位于tcra特异性zfn靶位点(表1)之间的序列ttgaaa,而b2m对是sbs#57332/sbs#57327(表5),其包括b2m特异性zfn靶位点之间的序列tcaaat。
简言之,t细胞(ac-tc-006)被解冻并以cd3/28免疫磁珠(dynabead)(1:3细胞:珠比)在x-vivo15t细胞培养基中活化(第0天)。培养2天后(第2天),将aav供体(包括gfp转基因和tcra或b2m基因的同源臂)添加到细胞培养物,不同的是,还维持无供体的对照组。接下来的一天(第3天),通过mrna递送将tcra和b2mzfn添加到下述5组中:
(a)第1组(仅有tcra和b2mzfn,无供体):tcra120ug/ml:b2m仅60ug/ml;
(b)第2组(tcra和b2mzfn以及具有tcra同源臂的供体):tcra120ug/ml;b2m60uq/ml和aav(tcra-位点e-hpgk-egfp-克隆e2)1e5vg/细胞;
(c)第3组(tcra和b2mzfn以及具有tcra同源臂的供体):tcra120ug/ml;b2m60ug/ml:和aav(tcra-位点e-hpgk-egfp-克隆e2)3e4vg/细胞;
(d)第4组(tcra和b2mzfn以及具有b2m同源臂的供体):tcra120ug/ml;b2m60ug/ml和aav(paavb2m-hpgkgfp)1e5vg/细胞;
(e)第5组(tcra和b2mzfn以及具有b2m同源臂的供体):tcra120ug/ml;b2m60ug/ml和aav(paavb2m-hpgkgfp)3e4vg/细胞。
所有的实验都以3e7个细胞/ml的细胞密度使用如美国专利公开号2017/0137845所述方案(极端冷激)进行,并且在电穿孔后在30℃过夜培养以冷激。
接下来的一天(第4天),将细胞稀释至0.5e6个细胞/ml,并转移至37℃培养物。3天后(第7天),再次将细胞稀释至0.5e6个细胞/ml。另培养3和7天后(分别为第10天和14天),收获细胞用于facs和miseq分析(稀释至0.5e6个细胞/ml)。
如图6所示,gfp表达指示靶向整合成功并且获得了经遗传修饰的细胞,所述细胞包含核酸酶靶位点内(或核酸酶靶位点的1-50、1-20、1-10或1-5个碱基对内的),包括本文所公开的ttgaaa和tcaaat内(成对的靶位点之间)的b2m和tcra修饰(插入和/或缺失)。
进行了其它实验以产生具有trac和b2m双重敲除和供体载体的靶向整合的细胞。具体而言,将trac特异性zfn对sbs#55266/sbs#53853和b2m对sbs#57071/sbs#57531引入t细胞。简言之,在x-vivo15t细胞培养基中,将1∶1比例的cd4:cd8人t细胞解冻并用cd3/28
培养3天后(第3天),在存在zfnmrna的maxcyte电穿孔缓冲液中将细胞浓缩至3e7个细胞/ml,然后使用maxcyte装置电穿孔。然后将浓集的电穿孔的细胞置于组织培养孔中,然后将编码hpgk-gfp-bgh多聚a转基因供体的aav6加入浓集的细胞中,允许其恢复并在37℃孵育20分钟。或者,可将供体载体添加到装置中的电穿孔缓冲液。然后将细胞在培养基中稀释至3e6个细胞/ml,并在30℃下培养过夜。次日早晨,另在新培养基中将细胞稀释至0.5e6个细胞/ml。以下是对这些组的描述:
(a)假:未添加zfnmrna或aav供体的电穿孔细胞;
(b)仅trac和b2mzfn,无供体):trac120ug/ml:仅b2m30ug/ml;
(c)trac和b2mzfn和具有b2m同源臂的供体:trac120ug/ml;b2m30ug/ml和aav6(b2m-位点a-hpgk-egfp)3e4vg/细胞;
(d)tcac和b2mzfn和具有trac同源臂的供体:trac120uq/ml;b2m30ug/ml;和aav6(tcra-位点e-hpgk-egfp)3e4vg/细胞。
所有的实验都以3e7个细胞/ml的细胞密度使用如美国专利公开号2017/0137845所述方案(极端冷激)进行,并且在电穿孔后在30℃过夜培养以冷激。接下来的一天(第4天),将细胞稀释至0.5e6个细胞/ml,并且在37℃转移至培养物。3天后(第7天),再次将细胞稀释至0.5e6个细胞/ml。另培养3和7天后(分别为第10天和14天),收获细胞用于facs和miseq分析(稀释至0.5e6个细胞/ml)。
如图7所示,gfp表达(供体)指示,靶向整合成功,且以高频率(包括80-90%敲除和靶向整合率)获得了在本文公开的核酸酶靶位点内(或核酸酶靶位点的1-50、1-20、1-10或1-5个碱基对内,包括成对位点之间)包含b2m和trac修饰(插入和/或缺失)的经遗传修饰的细胞。
还进行了实验,其中将car转基因整合到b2m和tcra双敲除中,在b2m、tcra或另一基因座处,以产生表达car的双b2m/tcra敲除。
实施例5:tcra和b2mzfn的优化
为了减少脱靶切割,采用一种核酸酶优化的策略,其中非特异性磷酸接触被选择性地去除,以引起脱靶切割的整体遏制(guilinger等.(2014)natmethods.11(4):429-35.doi:10.1038/nmeth.2845;kleinstiver等.(2016)nature529(7587):490-5.doi:10.1038/nature16526;slaymaker等.(2016)science)351(6268):84-8.doi:10.1126/science.aad5227)(参见美国专利公开号2018/0087072)。在与dna的磷酸主链相互作用的锌指框架内的一个或多个关键位置(pavletich和pabo(1991)science252(5007):809-17;elrod-erickson等.(1996)structure4(10):1171-80)以及在同样预测会形成磷酸接触的右zfnfoki结构域中的位置进行了氨基酸取代。
在下表6中示出了各zfn的特征信息。从左侧开始,显示sbs编号(例如55254),其中zfn结合的dna靶标显示在sbs编号下方。接下来显示指1-6或1-5的氨基酸识别螺旋设计物(表6的细分第二列)。表6中还在合适螺旋设计物下显示对指示的指的zfp主链序列进行的突变,如美国专利申请号15/685,580所述。表6中使用的表示法中,″qm5″表示在指示的指的-5位(相对于编号为-1至+6的螺旋),该位置的精氨酸已被谷氨酰胺(q)替代,而〝qm14″表示正常存在于-14位的精氨酸(r)已被谷氨酰胺(q)替代。nqm5中的缩写〝n″表示突变位于用于构建5或6指蛋白质的二指模块的n末端指中。〝无″表示在识别螺旋区之外没有变化。因此,例如,sbs#68797在指1、3和5中包含nqm5突变,而指2、4和6对锌指主链没有突变(例如,在识别螺旋区之外的锌指序列)。
最后,表6的最右列显示用于将dna结合结构域连接至foki切割结构域的接头(例如,″l0″lrgsqlvks(seqidno:135),称为″标准″接头,例如描述于美国专利号9,567,609中),显示在该列的顶行,在接头名称下方的框中显示foki磷酸接触突变和二聚化突变的位点。其它接头包括n7c(sgairchdefwf,seqidno:136)和n7a(sgtphevgvytl,seqidno:137)。具体而言,在fok突变体框的顶行上指示的是在二聚化结构域中发现的突变类型(例如,如美国专利号8,962,281中所述的eld或kkr)。在二聚化突变体名称的下方显示foki结构域中存在的任何突变,所述突变用以去除底部显示的非特异性磷酸接触(例如,k525s或r416s,其中氨基酸525位或416位的丝氨酸残基已取代赖氨酸或精氨酸,分别描述于美国公开号20180087072中)。因此,例如,在sbs#68796中,接头是l0接头,并且foki切割结构域包括eld二聚化突变体且没有磷酸接触突变。另外,对于sbs#68812,接头是l0接头,并且foki切割结构域包括kkr二聚化突变,其中foki结构域还包括r416e取代突变。
可与本文所述的zfp(包括衍生自本文所述的zfn的zfp)一起使用的其它foki结构域变体包括添加sharkey突变(s418p+k441e,参见guo等.(2010)j.molbiol,doi:10.101b/j.jmb.2010.04.060)和dad和rvrfoki突变(参见美国专利号8,962,281)。可用的经工程改造的foki变体的非限制性示例包括:
野生型foki切割结构域(seqidno:139):
foki-sharkey(s418p+k441e,seqidno:140):
fokield(q->e@486,i->l@499,n->d@496,seqidno:141)
fokield,sharkey(q->e@486,i->l@499,n->d@496,s418p+k441eseqidno:142)
fokield,r416e(q->e@486,i->l@499,n->d@496,r416e,seqidno:143)
fokield,sharkey,r416e(q->e@486,i->l@499,n->d@496,s418p+k441e,r416e,seqidno:144)
fokield,r416y(q->e@486,i->l@499,n->d@496,r416y,seqidno:145)
fokield,sharkey,r416e(q->e@486,i->l@499,n->d@496,s418p+k441e,r416e,seqidno:146)
fokield,s418e(q->e@486,i->l@499,n->d@496,s418e,seqidno:147)
fokield,部分sharkey,s418e(q->e@486,i->l@499,n->d@496,k441e,s418e,seqidno:148)
fokield,k525s(q->e@486,i->l@499,n->d@496,k525s,seqidno:149)
fokield,sharkeyk525s(q->e@486,i->l@499,n->d@496,s418p+k441e,k525s,seqidno:150)
fokield,i479t(q->e@486,i->l@499,n->d@496,i479t,seqidno:151)
fokield,sharkey,i479t(q->e@486,i->l@499,n->d@496,s418p+k441e,i479t,seqidno:152)
fokield,p478d(q->e@486,i->l@499,n->d@496,p478d,seqidno:153)
fokield,sharkey,p478d(q->e@486,i->l@499,n->d@496,s418p+k441e,p478d,seqidno:154)
fokield,q481d(q->e@486,i->l@499,n->d@496,q481d,seqidn0:155)
fokield,sharkey,q481d(q->e@486,i->l@499,n->d@496,s418p+k441e,q481d,seqidno:156)
fokikkr(e->k@490,i->k@538,h->r@537,seqidno:157)
fokikkrsharkey,(e->k@490,i->k@538,h->r@537,s418p+k441e,seqidno:158)
fokikkr,q481e(e->k@490,i->k@538,h->r@537,q481e,seqidno:159)
fokikkr,sharkeyq481e(e->k@490,i->k@538,h->r@537,s418p+k441e,q481e,seqidno:160)
fokikkr,r416e(e->k@490,i->k@538,h->r@537,r416e,seqidno:161)
fokikkr,sharkey,r416e(e->k@490,i->k@538,h->r@537,s418p+k441e,r416e,seqidno:162)
fokikkr,k525s(e->k@490,i->k@538,h->r@537,k525s,seqidno:163)
fokikkr,sharkey,k525s(e->k@490,i->k@538,h->r@537,s418p+k441e,k525s,seqidno:164)
fokikkr,r416y(e->k@490,i->k@538,h->r@537,r416y,seqidno:165)
fokikkr,sharkey,r416y(e->k@490,i->k@538,h->r@537,s418p+k441e,r416y,seqidno:166)
foki,kkri479t(e->k@490,i->k@538,h->r@537,i479t,seqidno:167)
foki,kkrsharkeyi479t(e->k@490,i->k@538,h->r@537,s418p+k441e,i479t,seqidno:168)
foki,kkrp478d(e->k@490,i->k@538,h->r@537,p478d,seqidno:169)
foki,kkrsharkeyp478d(e->k@490,i->k@538,h->r@537,p478d,seqidno:170)
fokidad(r->d@487,n->d@496,i->a@499,seqidno:171)
fokidadsharkey(r->d@487,n->d@496,i->a@499,s418p+k441e,seqidno:172)
fokirvr(d->r@483,h->r@537,i->v@538,seqidno:173)
fokirvrsharkey(d->r@483,h->r@537,i->v@538,s418p+k441e,seqidno:174)
测试了zfn的所有成对组合的功能,发现它们均有活性。
表6:对tcra具有特异性的zfn对
将编码针对各位点的zfn的基因克隆到表达质粒中,作为被各靶位点组合中的2a自切割肽分开的左和右伴侣。编码zfn的mrna使用标准的体外转录方法衍生。然后,通过电穿孔,以3种不同剂量(100μl3e6个t细胞中的12、6或3μg)的各种mrna处理活化的t细胞(活化后3天)。电穿孔后4天,分析细胞在一个和多个靶位点处的切割。数据显示在下方的两张表中(各靶位点对应一张表)。
表7a:位点d处的中靶和脱靶切割
表7b:位点e处的中靶和脱靶切割
因此,修饰后,zfn试剂保持了优异的中靶切割活性,常常还同时消除对于背景的脱靶裂解活性(例如,将亲代55254/55248对经修饰的68861/68796对的中靶切割活性做比较),在12μg的饱和剂量下分别显示96.7%和99.3%的中靶裂解,而在亲代对中该剂量的总脱靶活性为42.22%,在经修饰的对中是1.19%,与1.28的对照水平类似。
与traczfn一样:修饰b2m蛋白的foki结构域中的潜在的磷酸接触氨基酸。zfp组分(″设计物″)的示例性修饰示于下表8。
表8:b2m特异性zfn优化
如上所述测试了经修饰的b2m试剂的活性,并使用对hla具有特异性的抗体通过fac分析分析表型敲除。发现所有成对的组合(57531/57071、57531/72732、57531/72748、68957/57071、68957/72732、68957/72748、72678/57071、72678/72732、72678/72748)均具有活性,指示对的示例性结果示于下表9中,并且显示经修饰的变体具有活性。
表9:b2m特异性zfn的表型分析
nt:未测。
还使用miseq对表9中如上所列的各对进行了中靶和脱靶分析。各对的结果如下示于表10a-10d中,显示这些试剂是高度特异性的。
表10a:57071/68957对的脱靶分析
表10b:72732/57531对的脱靶分析
表10c:72732/72678对的脱靶分析
表10d:72748/68957对的脱靶分析
通过miseq分析且通过表型分析经facs分析法分析cd3+或hla+细胞的量,经修饰的trac-和b2m-特异性zfn经联合测试并评估敲除效率。使用两种不同浓度的添加的zfn编码mrna(90μg/ml或120μg/ml)在t细胞中进行分析。结果显示在下表11中,并证明这些试剂是高效的。
表11:trac/b2m切割
还在存在或不存在由pgk启动子驱动的gfp供体构建体的情况下联合测试了试剂。结果示于表12,其中插入到裂解的b2m或trac基因座中。在各情况下,pgk-gfp供体均由aav6递送,并且包含具有侧接trac或b2m切割位点的同源性的同源臂。所用的trac特异性zfn对构建体为68846-2a-53853,而用于b2m特异性对的构建体为72732-2a-72678。
表12:两种t细胞供体中的双敲除活性。
因此,通过选择foki变体(参见上文)与zfpdna结合结构域的组合,构建了对b2m具有特异性的zfn的优化对。
b2mzfn72732和72678的dna结合结构域的优化氨基酸序列如下所示:
72732n末端:
72678c末端:
还使用不同foki和/或接头结构域产生包含本文所述zfn(例如seqidno:175和seqidno:176)的经修饰zfp的其它zfn。
类似地,通过选择foki变体(参见上文)与zfpdna结合结构域的组合来构建对trac具有特异性的zfn的优化对。b2mzfn68846和53853的dna结合结构域的优化氨基酸序列如下所示:
68846c末端:
53853n末端:
zfn可与foki结构域n末端的dna结合结构域一同组装,其中在dna结合结构域和foki结构域之间的接头序列是l0接头:lrgs。或者,如果组装zfn使foki结构域在dna结合结构域的n末端,则使用的接头是n7c接头:sgairchdefwf(seqidno:179)。
将其它特征添加到构建体中,包括在n末端区域的3xflagtag(dykdhdgdykdhdidykddddk,seqidno:180)和核定位序列(pkkkrkv,seqidno:181)。
另外,在一些构建体中,编码感兴趣的zfn对的序列在一个dna序列中连接在一起,其中各zfn伴侣的开放阅读框被2a序列隔开。对于68846-2a-53853,所述dna序列如下所示:
68846-2a-53853开放阅读框的氨基酸序列是:
下表13中,该多肽的特征突发。
表13:68846-2a-53853肽序列的特征
72732-2a-72678开放阅读框的序列如下所示:
72732-2a-72678开放阅读框的氨基酸序列如下所示。
72732-2a-72678氨基酸序列的特征示于下表14。
表14:72732-2a-72678氨基酸序列的特征
实施例6:zfn试剂的体内测试
将本文所述的t细胞给予移植物抗宿主疾病和/或癌症的动物模型(例如,注射有癌细胞系例如多发性骨髓瘤的裸鼠以建立肿瘤模型)。例如,活化的人t细胞用编码b2m和trac特异性zfn的mrna进行电穿孔,其中各对均由单一mrna编码,所述单一mrna被编码2a自裂解肽的序列隔开(macleod等(2017)molther.25(4):949-961)。还用包含car供体(例如,cd19car)的aav颗粒转导细胞。然后,细胞经培养并针对car表达和cd3+细胞的缺乏染色。通过磁力分离耗尽所有残留的cd3+细胞。nsg小鼠静脉注射萤火虫荧光素酶表达型raji细胞(raji-ffluc),并在四天后注射cd3-/抗cd19cart细胞。注射后第四天,raji-ffluc细胞的植入和生长明显,并且在未经处理的小鼠中明显增加。在第8天观察到经处理小鼠血液中cart细胞的峰值频率,达到高剂量组外周血细胞的约10%。到第17-19天,对照组中的所有小鼠都显示明显的肿瘤负荷,尤其是在脊柱和骨髓中,导致完全后肢麻痹,并被安乐死。相比之下,接受抗cd19cart细胞处理的所有小鼠组在第11天都没有肿瘤生长的迹象,并且到研究的第32天仍保持无肿瘤状态。
在接受本文所述的t细胞的动物组织(例如,骨髓,脾,肺,肝,心脏等)中未检测到或仅有最小残留疾病。相比之下,对照对象的大多数组织中具有可检测的肿瘤细胞。
本文中提及的所有专利、专利申请和出版物均以参考的方式用全文纳入本文。
尽管出于理解清楚的目的,本发明通过说明和示例的方式提供了一些细节,但本领域技术人员可理解在不偏离本发明的精神或范围的情况下可实施各种改变和改进。因此,上述说明和实施例不应理解为是限制性的。
- 还没有人留言评论。精彩留言会获得点赞!