载体构建体的制作方法
本发明涉及噬菌体展示,并且尤其是涉及荧光噬菌体颗粒和用于产生荧光噬菌体颗粒的核酸载体。更具体地,本发明涉及载体构建体,其中编码荧光团的序列与编码pviii噬菌体外壳蛋白的序列融合。
背景技术:
在过去的三十年中,噬菌体展示已经成为一种用于发现和进化新型结合蛋白的有力且有效的方法。尽管存在竞争性的组合技术,但没有一种技术显示出与易用性组合的高度的多功能性。然而,在噬菌体淘选后筛选所需的表型仍然面临着挑战,并且没有鉴定出最佳结合物。如由流式细胞术兼容的细胞组合技术(如酵母菌展示)提供的实时选择具有显著的优势,因为荧光标记的配体和荧光激活的细胞分选(facs)以结合其回收的定量的方式有效地将结合物与非结合物分离(boder和wittrup,1997,《自然生物技术(natbiotechnol)》,15,553-557和feldhaus等人,2003,《自然生物技术》21,163-170)。
由于以下多种原因,基于facs的有效方法尚未与噬菌体展示整合在一起:1)噬菌体颗粒太小以至于无法通过流动中的大小(正向和侧向散射)检测到,因此需要直接荧光标记或间接可视化,如通过标记的抗噬菌体抗体的直接荧光标记或间接可视化,这显著地限制了灵敏度和实用性(bowley等人,2009,《美国科学院院报(pnas)》106:1380-1385),2)facs需要可回收的物理颗粒,因此,由于单独的噬菌体颗粒太小,靶必须在细胞表面上表达或者被固定在固相如珠子上,3)当在结合位点处或在结合位点附近存在游离反应基团时,通过化学偶联直接荧光标记噬菌体可能会破坏靶结合,或者引入抗原结合的空间位阻,4)抗体检测需要额外的孵育和洗涤步骤,这些步骤导致克隆的潜在丢失并降低选择效率。
因此,将固有的荧光检测模块整合到噬菌体平台中将显著地改善使用基于facs的方法的能力,以及为基于使用荧光的检测的其他类型的测定提供高度有用的工具。
先前已经实现了在噬菌体颗粒的表面上荧光蛋白的功能展示。例如,velappan等人(《核酸研究(nucleicacidresearch)》,38(4):e22,2010)已经表明,一些形式的gfp可以被表达为与pix或piii噬菌体外壳蛋白的融合体,尽管最好的构建体在荧光团和噬菌体外壳蛋白之间没有直接的基因融合体,而是使用了卷曲螺旋排列以间接翻译后方式连接荧光团和噬菌体外壳蛋白,其中每种组分都被编码并表达为单独的独立组分。此外,speck等人(《蛋白质工程、设计与选择(proteinengineering,design&selection)》:peds,24,473-484,2011)已经表明,在噬菌粒系统中,gfp和一些gfp衍生物可以被表达为与截短形式(仅c端结构域)的piii噬菌体外壳蛋白的直接融合体。
pviii是丝状噬菌体上的主要外壳(衣壳)蛋白,并且因此因其每噬菌体颗粒的高拷贝数,荧光团与pviii外壳蛋白的融合是潜在有利的,并且不干扰在其余四种衣壳蛋白中的任一种上的末端展示。然而,有关基于经典噬菌体展示的异源分子与pviii外壳蛋白融合的报道仅限于短肽的展示。
此外,如上面所讨论的,使用经典的噬菌体展示技术的荧光团的功能性噬菌体展示可能是有问题的。其中的一个原因是荧光团蛋白非常敏感,并且通常在细胞质中表达和折叠以便发挥功能,这在获得表达的荧光团-噬菌体外壳蛋白融合蛋白向在噬菌体表面上展示的周质和功能性荧光团的转移方面可能存在问题(delisa等人,《生物化学(biol.chem.)》,277(33),29825-29831,2002)。
目前解决这一问题的技术包括使用荧光标记的抗pviii抗体,所述抗体进而可以用于荧光标记噬菌体颗粒,所述噬菌体颗粒然后可以使用标准技术检测,例如通过流式细胞术或荧光染色来检测。
最近,hess等人,2012(《生物共轭化学(bioconjugatechemistry)》,23:1478-1487),通过使用翻译后技术证明gfp在噬菌体pviii外壳蛋白上的融合,已经克服了这些问题中的一些,其中分选酶用于将gfp分子与噬菌体颗粒的表面上存在的pviii分子缀合。这已经通过使用标准噬菌体展示技术以获得表达pviii外壳蛋白的噬菌体颗粒来实现,所述pviii外壳蛋白具有可以被分选酶识别的短肽插入。然后这一肽插入可以经历标准分选酶技术,以便将噬菌体的表面上的修饰的pviii外壳蛋白翻译后连接至从标准来源获得的gfp分子。
因此,尽管hess等人(见上文)提供了其中pviii蛋白与gfp荧光团融合的噬菌体颗粒,但这不是通过经典的噬菌体展示技术来完成的,在所述经典的噬菌体展示技术中,在噬菌体颗粒内在编码pviii外壳蛋白的核酸序列与编码荧光团的序列之间存在基因融合体,使得将产生融合蛋白。这也意味着在hess系统中不存在基因型-表型联系,这对于筛选技术是不利的,在筛选技术中,经典的噬菌体展示方法的一个重要特征是展示感兴趣的蛋白(例如,与靶配体结合的蛋白质)的选定噬菌体颗粒含有编码这一展示的感兴趣的蛋白的遗传物质,其然后可以获得并进一步分析或操作。
技术实现要素:
在本发明中,令人惊讶地且与如上面所描述的现有技术教导的预期相反,本发明人已经发现荧光蛋白可以作为与pviii噬菌体外壳蛋白的基因融合体展示在噬菌体颗粒上。这已经通过提供一种载体构建体来实现,所述载体构建体将大肠杆菌(e.coli)tat分泌途径的使用与荧光团-pviii融合蛋白相结合。这样的载体有利地允许产生展示功能性和可检测的荧光蛋白的噬菌体颗粒。换句话说,噬菌体颗粒本质上(或固有地)是荧光的。
发明人还已经表明,这样的载体可以用于并入感兴趣的蛋白(poi)与可替代的(非pviii)噬菌体外壳蛋白的第二融合体,以便实现荧光-pviii融合蛋白和与另一种外壳蛋白融合的感兴趣的蛋白的同时表面展示。换句话说,本发明人在本文中已经证明了具有固有的荧光能力和由poi提供的另一种能力(例如,靶结合活性)的载体和噬菌体颗粒的产生。因此,载体的这一荧光噬菌体系统和使用所述载体产生的噬菌体(本文中也被称为荧光噬菌体(fluorophage))可以有利地用于选择和筛选,以便将荧光筛选的优点和容易性与使用来自经典噬菌体展示的表型-基因型联系立即且容易地研究选定poi的能力相结合。这一荧光噬菌体系统在非facs测定中具有用途,因为它本身与经典的基于微量滴定板的筛选方法直接兼容,所述经典的筛选方法传统上作为酶联免疫吸附测定(elisa)或类似的方法来完成,其现在可以被简化并且改为依赖于定量荧光测量,例如通过flisa(荧光连接免疫吸附测定)。其他用途在本文其他地方描述。
进一步地,发明人已经表明,通过建立现在具有作为噬菌体展示系统的固有部分的检测模块的遗传融合蛋白的能力,人们还可以使用组合分子进化来优化和改善融合蛋白。这是通过以下完成的:将编码荧光团和pviii部分的序列随机化,随后在facs中进行表型筛选,以用于与母体构建体相比增加的信号。这允许鉴定新的突变融合蛋白,所述新的突变融合蛋白增加了获得的信号,超出了用母体构建体所能看到的信号。效果似乎是由于异源蛋白更好地适应产生的大肠杆菌宿主,使得最终结果是功能性融合蛋白池的增加,其转化为来自产生的细胞的荧光信号的增加,以及由这一细胞所产生的噬菌体颗粒的增加。此外,本发明人已经表明在大肠杆菌宿主细胞中tat转运蛋白的过表达可以显著地改善荧光信号。
因此,在一个方面,本发明提供了一种载体构建体,所述载体构建体包含以下组分:
(i)编码将蛋白质导入tat分泌途径的信号肽的序列;和
(ii)与编码pviii噬菌体外壳蛋白的序列融合的编码荧光团的序列。
本发明的载体是表达载体或表达构建体,即通常由能够在合适的宿主细胞中表达(蛋白质合成)所需编码的蛋白质组分的核酸序列构成。
如上面所概述的,本发明属于噬菌体展示领域。因此,本发明的载体可以是噬菌体载体或噬菌粒载体(质粒),其基本结构和组分对于本领域技术人员来说是众所周知的,并且被选择以便实现噬菌体蛋白的表达和噬菌体颗粒在合适的宿主细胞中的包装,使得与各种噬菌体外壳蛋白融合的异源蛋白或外源蛋白被展示在噬菌体颗粒的表面上。
因此,当本发明的载体用于转化合适的宿主细胞如合适的大肠杆菌菌株时,产生了噬菌体颗粒,所述噬菌体颗粒含有许多与pviii噬菌体外壳蛋白融合并展示在噬菌体颗粒的表面上的功能性荧光团分子,其中载体序列或其他核酸序列编码噬菌体颗粒中含有的噬菌体基因组的各种噬菌体组分。展示在噬菌体的表面上的这种功能性荧光团可以通过常规技术检测,如荧光染色(可通过例如荧光显微镜或流式细胞术,或荧光读取器或检测器,例如板读取器或扫描仪或光度计检测)。
本发明扩展到可以形成本发明的载体的一部分的核酸序列或核酸分子。因此,本发明的另一方面提供了一种包含以下组分的核酸分子或核酸序列:
(i)编码将蛋白质导入tat分泌途径的信号肽的序列;和
(ii)与编码pviii噬菌体外壳蛋白的序列融合的编码荧光团的序列。
本发明进一步提供了噬菌体或噬菌体颗粒,其包含本发明的载体或核酸分子并且在表面上表达荧光团-pviii融合蛋白。这样的噬菌体或噬菌体颗粒可以是任何丝状噬菌体。优选的示例是肠杆菌噬菌体,例如m13、fd和f1噬菌体。
因此,本发明的载体包含编码信号肽的序列。所述信号肽(或信号序列,或前导序列,或周质前导肽,或周质前导序列)可以是任何合适的序列(氨基酸序列),其例如通过将蛋白质靶向或导向tat转运蛋白而将蛋白质导入或靶向到双精氨酸易位(tat)出口或分泌途径(或易位途径)。这样的信号肽在本文中也被称为tat信号肽。在本发明的载体中,这一编码信号肽的序列与编码荧光团-pviii融合蛋白(一种与pviii蛋白融合的荧光团蛋白)的序列可操作地连接,并且因此将蛋白(荧光团-pviii融合蛋白)导入tat分泌途径中。
tat途径是在细菌宿主细胞如大肠杆菌中发现的若干种易位或分泌途径之一。在这一途径中,在完全折叠的蛋白质跨过内膜易位至周质之前,蛋白质在细胞质的还原条件下被折叠。在到达周质后,信号肽然后被合适的信号肽酶除去。其他已知的在革兰氏阴性菌如大肠杆菌中的易位或分泌途径(其涉及蛋白质从胞质向周质区室的易位)是信号识别颗粒(srp)依赖性sec途径,即经典分泌(sec)途径或yidc依赖性途径(baneyx,2004,《自然生物技术》22:1399-1408)。与tat途径相反,sec途径将未折叠的蛋白质跨过内膜转运至周质,这涉及穿线机制,而yidc介导的转运也可能涉及使用翻转酶机制的至少部分折叠的转运(kumazaki等人,2014,《自然(nature)》509:516-520)。
用于tat途径的合适的信号序列或信号肽对于本领域技术人员而言将是众所周知的,并且可以使用其中的任何一种,或者实际上显示出保留的或改善的将蛋白质导入tat途径的能力的其衍生物或变体。事实上,tat途径的名称(双精氨酸易位)指的是高度保守的双精氨酸前导基序(s/trrxflk,seqno:33),所述基序在意图通过tat途径转运的蛋白质的n端区域中发现。在这一序列中,x可以是任何氨基酸,优选地任何极性氨基酸。因此,这将是可以形成本发明的载体中信号肽的一部分的示例性序列。
一个优选的示例是大肠杆菌三甲胺n-氧化物还原酶(tora)信号序列或其衍生物(或变体),与起始序列相比,其保留或具有改善的通过tat途径输出蛋白质的能力(例如,功能突变体的获得)。示例性的且优选的衍生物例如如在delisa等人(《生物化学杂志(j.biol.chem.)》,277(33),29825-29831,2002)中所描述的。
野生型tora前导序列的完整序列如下(mnnndlfqasrrrflaqlggltvagmlgpslltprrat,seqidno:32)。靶向作为tat转运蛋白的一部分的蛋白的这一信号肽序列或共享tora的特征的任何其他信号肽序列可以用于本发明的载体(例如,如在interpro条目ipr006311或bendtsen等人,2005,《bmc生物信息学(bmcbioinformatics)》6:167中所描述的)。在上面的tora野生型前导序列中,双精氨酸共有基序被加下划线,并且包含这一序列的信号肽可以用于本发明的载体,只要对tat途径的靶向性被保留。
用于本发明的特别优选的信号肽是torab7信号肽(mnnndlfqtsrqrflaqlggltvagmlgpslltprrat,seqidno:2),与野生型tora信号肽相比,所述torab7信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。torab7信号肽具有或包含序列tsrqrfla(seqidno:34),其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
可替代地,也可以使用如在delisa等人(见上文)的表ii中公开的torab6、e2、f1、f11或h2突变体。
因此,用于本发明的优选的信号肽是torab6信号肽(mnnndlfqtsrrrllaqlggltvagmlgpslltprrat,seqidno:35),与野生型tora信号肽相比,所述torab6信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。torab6信号肽具有或包含序列tsrrrlla,seqidno:36,其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
因此,用于本发明的优选的信号肽是torae2信号肽(mnnndifqasrrrflaqpggltvagmlgpslltprrat,seqidno:37),与野生型tora信号肽相比,所述torae2信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。torae2信号肽具有或包含序列ifqasrrrflaqp,seqidno:38,其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
因此,用于本发明的优选的信号肽是toraf1信号肽(mnnnelfqasrrrflaqlggltvagmlgpslltprrat,seqidno:39),与野生型tora信号肽相比,所述toraf1信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。toraf1信号肽具有或包含序列elfqasrrrfla,seqidno:40,其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
因此,用于本发明中的优选的信号肽是toraf11信号肽(mnnndlfqttrrrflaqlggltvagmlgpslltprrat,seqidno:41),与野生型tora信号肽相比,所述toraf11信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。toraf11信号肽具有或包含序列ttrrrfla,seqidno:42,其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
因此,用于本发明中的优选的信号肽是torah2信号肽(mnnndsfqtsrrrflaqlggltvagmlgpslltprrat,seqidno:43),与野生型tora信号肽相比,所述torah2信号肽已经被工程改造成用于通过tat途径改善蛋白质的周质靶向性。torah2信号肽具有或包含序列sfqtsrrrfla,seqidno:44,其中带下划线的残基是不同于野生型tora信号序列的残基,并且包含这一序列的信号肽是特别优选的,只要对tat途径的靶向性被保留。
此外,来自上述信号肽(特别是torab7信号肽)的进一步的衍生物或变体可以用于本发明的载体,只要这样的衍生物或变体序列显示出保留的或改善的将蛋白质导入tat途径的能力。使用任何方便的技术,例如使用大肠杆菌tatc和tate突变株,或者例如使用如在delisa等人(见上文)中所描述的蛋白质报告测定,可以容易地验证将蛋白质导入tat途径的能力。
合适的变体可以包括信号肽,所述信号肽包含氨基酸序列或编码氨基酸序列的核苷酸序列或由其组成,与上述信号肽序列具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。这些变体序列应该保留或具有充当可以将蛋白质导入tat分泌途径的信号肽的功能特性。也可以使用这些序列(或这些同源序列)的功能性截短或片段,只要编码保留或具有充当信号肽的功能特性的蛋白质的能力被保留。突变的信号肽的其他优选的示例是在上述信号肽中含有多达6个,例如多达5个、4个、3个、2个或1个改变的氨基酸的序列。
在本文其他地方概述的实施例中,其中本发明的载体可以编码第二种非pviii噬菌体外壳蛋白融合体,例如与感兴趣的蛋白(poi)融合的第二种(不同的)噬菌体外壳蛋白,于是在这些实施例的一些中,另一种类型的(即非tat)的信号序列可以与非pviii噬菌体外壳蛋白融合体一起使用(可以可操作地与非pviii噬菌体外壳蛋白融合体连接),或者可以不使用信号序列,例如在poi-pix融合体的情况下,不需要使用信号序列,并且有时优选地不使用信号序列。在其中不使用信号序列的实施例中,例如在poi-pix融合构建体的情况下,这样的pix融合蛋白通常将被导入yidc途径。在其中使用非tat信号序列的实施例中,非pviii噬菌体外壳蛋白融合体将被导入如由所选的信号序列的性质决定的非tat分泌途径,例如,被导入sec分泌途径。
在本发明的载体中,编码信号肽(tat信号肽)的序列与编码荧光团-pviii融合蛋白的序列连接,例如与编码荧光团-pviii融合蛋白的序列可操作地连接,使得其将荧光团-pviii融合蛋白导入tat分泌途径。
术语“融合蛋白”、“融合的”等在本文中用于描述同一多肽序列或同一开放阅读框(orf)中的两种或更多种蛋白质组分的功能连接。这样的融合蛋白也可以被描述为基因融合体,因为它们由相同的核酸序列(有时被称为“融合基因”或“融合核苷酸序列”)编码。虽然在这种融合蛋白中两种(或更多种)蛋白质组分(或编码核酸序列)可以彼此直接相邻,但是同样地,这些组分可以通过合适的肽间隔子或接头连接。如本领域中众所周知的,间隔子或接头对于允许每个单独的蛋白质组分以功能性方式表达(例如,允许它们形成适当的三维结构以实现或保持它们的天然功能)可以是重要的。
因此,在由本发明的载体编码的融合蛋白中,肽间隔子(或接头)通常被包括在荧光团和融合蛋白的pviii部分(有时被称为pviii融合蛋白)之间,或者被包括在感兴趣的蛋白(poi)和第二噬菌体外壳蛋白之间。在其他实施例中,不需要包括这样的接头或间隔子,或者可以仅被包括在一些组分之间。因此,在本发明的载体中,编码荧光团或poi的序列可以分别与编码pviii或其他噬菌体蛋白的序列融合,在组分之间有或没有间隔子或接头序列。所有这些可能性(即有或没有间隔子或接头序列的融合蛋白或编码核酸)仍然被认为是直接融合体或直接基因融合体。
尽管这一讨论集中在噬菌体外壳蛋白和荧光团或poi之间的接头或间隔子上,但接头序列可以在其他地方适当地被包括在本发明的载体中,例如在如本文所讨论的载体的其他组分之间,例如在信号肽和荧光团之间。
因此,术语“pviii融合蛋白”是指与外源肽或蛋白(在这种情况下是荧光团)融合的pviii蛋白(噬菌体pviii蛋白)或其片段。术语“piii融合蛋白”、“pvi融合蛋白”、“pvii融合蛋白”或“pix融合蛋白”是指与外源肽融合的piii、pvi、pvii或pix蛋白(piii、pvi、pvii或pix噬菌体蛋白)(视情况而定),或其片段,例如感兴趣的蛋白(poi)。
因此,本发明的载体包含编码荧光团的序列(核酸序列),所述序列与编码pviii噬菌体外壳蛋白(有时在本文中被称为荧光团-pviii)的序列融合(基因地融合)。荧光团和pviii在载体中可以处于任何合适的顺序或间隔,只要一旦表达并包装成噬菌体颗粒,就在荧光团和pviii之间形成功能性融合蛋白,其中融合蛋白的pviii组分形成噬菌体外壳的一部分,并且荧光团在噬菌体颗粒的表面上功能性表达或展示。因此,融合蛋白的荧光团部分与融合蛋白的pviii部分位于框内。这意味着荧光团和pviii在相同的多肽序列中表达,或者换句话说,作为直接融合体。在本发明的载体中,通常优选的是,融合蛋白的荧光团组分位于融合蛋白的pviii组分的n端(或在n端或其附近)。
当荧光团与主要噬菌体外壳蛋白融合时,采用经典的(8+8野生型(噬菌粒)或88野生型(基因组))互补表达方法的pviii,通过使用本发明的载体产生的最终噬菌体颗粒将具有沿着噬菌体颗粒的长度随机分布的荧光团的多个拷贝,以及野生型pviii蛋白的拷贝。例如,由于m13噬菌体在其表面上具有主要外壳蛋白pviii的大约2700个拷贝,因此可以存在荧光团的许多拷贝。因此,荧光团的这种功能性表达可以通过监测荧光(例如,可检测的荧光),通过任何合适和方便的测定被方便地评估。如在实例中所示出的,这种荧光监测可以方便地通过监测转化的宿主细胞如大肠杆菌中的荧光(例如,使用荧光显微镜法),或者通过使用板读取器或其他方便的设备分析噬菌体颗粒的样品例如液体样品的荧光来进行。也可以使用facs或flisa分析。
由于在荧光团和pviii噬菌体外壳蛋白之间具有基因融合体对于本发明是重要的,因此方便地,合适的荧光团将是生物荧光团或可以由核酸序列编码并因此被包括在本发明的载体中的其他荧光团。优选地,这种荧光团也是固有地发荧光的或可直接检测的。也可以使用可以被活化或可漂白的荧光团。
用于本发明的优选的荧光团是具有或包含β-桶结构或架构的生物荧光团。
这样的荧光团对于本领域技术人员来说是众所周知的(参见,例如shaner等人,2007,《细胞科学杂志(j.cellsci)》,120(24):4247-4260的综述),并且包括绿色荧光蛋白(gfp)或其与gfp具有相同折叠拓扑的衍生物(也被称为gfp衍生物)。换句话说,用于本发明的这样的荧光团包括绿色荧光蛋白(gfp)或其具有β-桶架构或结构或gfp-架构的荧光团衍生物。这样的非gfp荧光可以方便地被称为gfp样荧光团或具有gfp架构的gfp衍生物或荧光团。虽然绿色荧光蛋白现在已经从多种生物中分离出来,但术语gfp在传统上和在本文中是指首先从水母维多利亚水母(aequoreavictoria)中分离出来的蛋白。然而,这一野生型蛋白质已经经历各种修饰,以产生目前在文献中描述的各种各样的gfp衍生物或gfp样荧光团。
这样的荧光团含有一个或多个β-桶作为单元,例如可以含有单个β-桶(β-桶单体)或多个β-桶,例如β-桶同源四聚体。
所有这样的荧光团都具有或包含类似的3-d圆柱形结构,其中大部分多肽骨架被缠绕成通常包含11股β片层的β-桶结构,这些β片层围绕含有发色团的中心α螺旋。β-桶结构形成近乎完美的圆柱体,形成了一种可以被称为“β-can”的结构,其表征了gfp样荧光团家族(gfp衍生物)。因此,从另一个角度来看,含有这一“β-can”结构的荧光团也优选用于本发明。这一β桶形圆筒通常为大约为24至
荧光团的gfp家族(gfp加gfp变体或衍生物,或gfp样荧光团)包括蓝色变体(bfp)、青色变体(cfp)和黄色变体(yfp),这取决于原始gfp的光谱发射谱以及绿色变体。增强的gfp(egfp)、增强的cfp(ecfp)和增强的yfp(eyfp)是进一步的示例。
尽管荧光团的gfp家族(gfp加gfp变体或衍生物,或gfp样荧光团或gfp相关荧光团)具有一些固有的不同性质,例如发射颜色、荧光强度和其他性质,如稳定性(并且实际上可以被工程改造以便具有这样的不同性质),但上面描述的β-桶结构或架构的存在和保持意味着它们通常以类似的方式表现。因此,荧光团的gfp家族的成员(或其变体,例如具有至少70%序列同一性或如本文其他地方所描述的其他值的功能变体)是用于本发明的优选的荧光团。
编码这样的荧光团的核酸序列(和它们的氨基酸序列)在本领域中是众所周知并被描述,并且可以容易地从文献或标准数据库(参见,例如pfam条目pf01353或interpro条目ipr011584)中获得。
用于本发明的合适的荧光团或gfp样荧光团(或gfp变体荧光团)或含有β-桶的荧光团的一些具体示例是mneongreen(另一种具有极亮黄绿色荧光的绿色荧光蛋白,shaner等人,2013,《自然方法(naturemethods)》,10(5):407-409)、mgfpmut2(gfp的facs优化版本,cormack等人,1996,《基因(gene)》173:33-38)和mcherry(一种红色荧光蛋白,shaner等人,2004,《自然生物技术》,22,1567-1572)。“m”代表单体。其他示例可以是egfp(或ecfp或eyfp)。用于本发明的优选的荧光团是mneongreen或其衍生物或变体,以及gfp或其衍生物或变体,例如mgfpmut2或一些其他衍生物或变体,例如egfp(heim等人,1995,《自然》373:663-664)。用于本发明的更优选的荧光团是mneongreen或其衍生物或变体。
mneongreen(shaner等人,2013,见上文)是一种来源于文昌鱼属(branchiostomalanceolatum)的明亮的单体绿色荧光蛋白。来自文昌鱼属的原始天然蛋白质是四聚体,并且已经对其进行修饰(如在shaner等人,2013中所描述的,见上文),以获得用于本发明的单体(m)neongreen(mng)。最终的突变体含有相对于四聚体起始蛋白的21个取代,并且具有尖锐的激发和发射峰(在506nm和517nm)。因此,使用标准的绿色荧光蛋白带通或长通滤光片组,可以在基本不损失发射光子的情况下对mneongreen成像。用于mneongreen的合成构建体的序列具有genbank登录号kc295282.1并且为了完整起见在下面重复。核酸序列的长度是711个碱基对(seqidno:45):
用于编码的蛋白的蛋白标识符为agg56535.1,其具有如下所概述的序列(236个氨基酸,seqidno:46):
在其中载体编码mneongreen荧光团的本发明的实施例中,如上所概述的核酸序列可以被并入到本发明的载体中。如同在本发明的载体中一样,编码mneongreen的序列被编码为信号肽下游的融合蛋白,然后并入到本发明的载体中的mneongreen核苷酸序列已经方便地去除起始密码子(atg)和终止密码子(taa)。
在本发明的优选实施例中,使用这一核酸序列的密码子优化版本,其已经针对细菌表达(例如,在大肠杆菌中)被优化。尽管可以使用任何合适的密码子优化版本,但可以用于本发明的mneongreen的密码子优化版本的特定序列概述如下。(同样,为了更好地整合到载体中,起始密码子和终止密码子已经被省略)。
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcacatcttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggacgaactgtacaaa(seqidno:3)
这些密码子优化的版本都编码与上述agg56535.1相同的氨基酸序列。然而,因为省略了编码初始甲硫氨酸残基的核苷酸序列,所以编码的氨基酸序列是:
如上所述,并且如本文其他地方更详细地讨论的,在本发明的其他实施例中,载体包含编码mneongreen(mng)荧光团的衍生物或变体(例如,seqidno:4或46的衍生物或变体,或seqidno:45或3的衍生物或变体)的序列。
因此,在其他实施例中,编码的mng荧光团包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:4具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。这些变体mng序列应保留或具有充当荧光团的功能性质。也可以使用seqidno:4(或这些同源序列)或其他mng序列的功能性截短或片段,只要编码保留或具有充当荧光团的功能特性的蛋白质的能力被保留。
同样地,编码mng荧光团的核酸分子可以包含核苷酸序列或由核苷酸序列组成,所述核苷酸序列与seqidno:45或3具有至少70%、75%或80%的序列同一性,例如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。这些变体mng核苷酸序列应编码保留或具有充当荧光团的功能特性的蛋白质。还可以使用seqidno:45或3(或这些同源序列)或其他mng序列的功能性截短或片段,只要编码保留或具有充当荧光团的功能特性的蛋白质的能力被保留。
如本文中所使用的术语“pviii噬菌体外壳蛋白”或“pviii蛋白”或“pviii噬菌体蛋白”或“pviii外壳蛋白”等是指源自或衍生自丝状噬菌体(野生型或天然序列)的pviii蛋白,或具有对应于这种pviii蛋白的序列的序列的pviii蛋白。衍生出pviii蛋白或pviii蛋白所对应的优选的丝状噬菌体是m13、fd和f1噬菌体。可以使用任何合适的pviii蛋白,只要它具有将荧光团展示为pviii融合蛋白的能力。优选地,由本发明的载体编码的pviii蛋白对应于成熟的(全长)pviii蛋白,即缺少pviii信号肽。优选地,由本发明的载体编码的pviii蛋白包含以下氨基酸序列或由以下氨基酸序列组成,所述氨基酸序列对应于来自m13丝状噬菌体的野生型(全长)成熟pviii蛋白,而没有其信号序列。
aegddpakaafnslqasateyigyawamvvvivgatigiklfkkftskas
(seqidno:8)
用于包含在载体中的编码这一序列的示例性核酸序列在本文其他地方作为seqidno:7提供。
在其他实施例中,编码的pviii蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:8具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。如上所述,这些变体pviii序列应保留或具有将荧光团展示为pviii融合蛋白的功能能力。还可以使用seqidno:8(或这些同源序列)或其他pviii序列的功能性截短或片段,只要将荧光团展示为pviii融合蛋白的能力被保留。
同样,编码pviii蛋白的核酸分子包含核苷酸序列或由核苷酸序列组成,所述核苷酸序列与seqidno:7具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。这些变体pviii核苷酸序列应保留或具有编码荧光团作为可以在噬菌体颗粒上展示的pviii融合蛋白的功能能力。还可以使用seqidno:7(或这些同源序列)或其他pviii序列的功能性截短或片段,只要编码荧光团作为可以在噬菌体颗粒上展示的pviii融合蛋白的能力被保留。
优选的pviii变体在seqidno:8的位置33或对应于seqidno:8的位置33具有缬氨酸至异亮氨酸突变(即在seqidno:8的位置33或对应于seqidno:8的位置33具有异亮氨酸残基),或在seqidno:7的残基97至99处或对应于seqidno:7的残基97至99具有ata异亮氨酸密码子,如本文其他地方所描述的。特别优选的pviii变体是如本文其他地方所述的如在f03克隆中发现的pviii序列。优选的pviii片段或截短是功能性片段或截短,其包括在氨基酸或核苷酸水平上对应于seqidno:8的位置33的残基。
如本文其他地方所描述的,在一些实施例中,本发明的载体编码其他非pviii噬菌体外壳蛋白,如pviii、pvi、pvii或pix外壳蛋白。
如本文中所使用的术语“piii噬菌体外壳蛋白”或“piii蛋白”或“piii噬菌体蛋白”或“piii外壳蛋白”等是指来源于或衍生自丝状噬菌体(野生型或天然序列)的piii蛋白,或具有对应于这种piii蛋白的序列的序列的piii蛋白。衍生出piii蛋白或piii蛋白所对应的优选的丝状噬菌体是m13、fd和f1噬菌体。可以使用任何合适的piii蛋白,只要它具有将poi展示为piii融合蛋白的能力。优选地,由本发明的载体编码的piii蛋白对应于成熟的piii蛋白,即缺少piii信号肽。优选地,由本发明的载体编码的piii蛋白包含以下氨基酸序列或由以下氨基酸序列组成,所述氨基酸序列对应于来自m13丝状噬菌体的野生型成熟piii蛋白,而没有其信号序列(也参见(genbankay598820.1)。
>piii_1
aetvesclakphtensftnvwkddktldryanyegclwnatgvvvctgdetqcygtwvpiglaipenegggsegggsegggsegggtkppeygdtpipgytyinpldgtyppgteqnpanpnpsleesqplntfmfqnnrfrnrqgaltvytgtvtqgtdpvktyyqytpvsskamydaywngkfrdcafhsgfnedpfvceyqgqssdlpqppvnagggsgggsgggsegggsegggsegggsegggsgggsgsgdfdyekmanankgamtenadenalqsdakgkldsvatdygaaidgfigdvsglangngatgdfagsnsqmaqvgdgdnsplmnnfrqylpslpqsvecrpfvfgagkpyefsidcdkinlfrgvfafllyvatfmyvfstfanilrnkes
(seqidno:12)
用于包含在载体中的编码这一氨基酸序列的示例性核酸序列在下文中作为seqidno:11提供。
gctgaaactgttgaaagttgtttagcaaaaccccatacagaaaattcatttactaacgtctggaaagacgacaaaactttagatcgttacgctaactatgagggctgtctgtggaatgctacaggcgttgtagtttgtactggtgacgaaactcagtgttacggtacatgggttcctattgggcttgctatccctgaaaatgagggtggtggctctgagggtggcggttctgagggtggcggttctgagggtggcggtactaaacctcctgagtacggtgatacacctattccgggctatacttatatcaaccctctcgacggcacttatccgcctggtactgagcaaaaccccgctaatcctaatccttctcttgaggagtctcagcctcttaatactttcatgtttcagaataataggttccgaaataggcagggggcattaactgtttatacgggcactgttactcaaggcactgaccccgttaaaacttattaccagtacactcctgtatcatcaaaagccatgtatgacgcttactggaacggtaaattcagagactgcgctttccattctggctttaatgaggatccattcgtttgtgaatatcaaggccaatcgtctgacctgcctcaacctcctgtcaatgctggcggcggctctggtggtggttctggtggcggctctgagggtggtggctctgagggtggcggttctgagggtggcggctctgagggaggcggttccggtggtggctctggttccggtgattttgattatgaaaagatggcaaacgctaataagggggctatgaccgaaaatgccgatgaaaacgcgctacagtctgacgctaaaggcaaacttgattctgtcgctactgattacggtgctgctatcgatggtttcattggtgacgtttccggccttgctaatggtaatggtgctactggtgattttgctggctctaattcccaaatggctcaagtcggtgacggtgataattcacctttaatgaataatttccgtcaatatttaccttccctccctcaatcggttgaatgtcgcccttttgtctttggcgctggtaaaccatatgaattttctattgattgtgacaaaataaacttattccgtggtgtctttgcgtttcttttatatgttgccacctttatgtatgtattttctacgtttgctaacatactgcgtaataaggagtcttaa
(seqidno:11)
在其他实施例中,编码的piii蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:12具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。这些变体piii序列应保留或具有将poi展示为piii融合蛋白的功能能力。还可以使用seqidno:12(或这些同源序列)或其他piii序列的功能性截短或片段,只要将poi展示为piii融合蛋白的能力被保留。用于展示的合适的截短的piii蛋白的示例,例如仅含有piii的c端部分的截短,在本领域中是众所周知的并且被描述。
如本文中所使用的术语“pvi噬菌体外壳蛋白”或“pvi蛋白”或“pvi噬菌体蛋白”或“pvi外壳蛋白”等是指源自或衍生自丝状噬菌体(野生型或天然序列)的pvi蛋白,或具有对应于这种pvi蛋白的序列的序列的pvi蛋白。衍生出pvi蛋白或pvi蛋白所对应的优选的丝状噬菌体是m13、fd和f1噬菌体。可以使用任何合适的pvi蛋白,只要它具有将poi展示为pvi融合蛋白的能力。优选地,由本发明的载体编码的pvi蛋白包含以下氨基酸序列或由以下氨基酸序列组成,所述氨基酸序列对应于来自vcsm13辅助噬菌体(genbankay598820.1)的pvi蛋白。
mpvllgiplllrflgfllvtlfgylltflkkgfgkiaiaislflaliiglnsilvgylsdisaqlpsdfvqgvqlilpsnalpcfyvilsvkaaififdvkqkivsyldwdk
(seqidno:13)
在其他实施例中,编码的pvi蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:13具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。这些变体pvi序列应保留或具有将poi展示为pvi融合蛋白的功能能力。还可以使用seqidno:13(或这些同源序列)或其他pvi序列的功能性截短或片段,只要将poi展示为pvi融合蛋白的能力被保留。
如本文中所使用的术语“pvii噬菌体外壳蛋白”或“pvii蛋白”或“pvii噬菌体蛋白”或“pvii外壳蛋白”等是指源自或衍生自丝状噬菌体(野生型或天然序列)的pvii蛋白,或具有对应于这种pvii蛋白的序列的序列的pvii蛋白。衍生出pvii蛋白或pvii蛋白所对应的优选的丝状噬菌体是m13、fd和f1噬菌体。可以使用任何合适的pvii蛋白,只要它具有将poi展示为pvii融合蛋白的能力。优选地,由本发明的载体编码的pvii蛋白包含以下氨基酸序列或由以下氨基酸序列组成,所述氨基酸序列对应于来自vcsm13辅助噬菌体(genbankay598820.1)的野生型pvii蛋白。
meqvadfdtiyqamiqisvvlcfalgiiaggqr
(seqidno:14)
在其他实施例中,编码的pvii蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:14具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。这些变体pvii序列应保留或具有将poi展示为pvii融合蛋白的功能能力。还可以使用seqidno:14(或这些同源序列)或其他pvii序列的功能性截短或片段,只要将poi展示为pvii融合蛋白的能力被保留。
如本文中所使用的术语“pix噬菌体外壳蛋白”或“pix蛋白”或“pix噬菌体蛋白”或“pix外壳蛋白”等是指源自或衍生自丝状噬菌体(野生型或天然序列)的pix蛋白,或具有对应于这种pix蛋白的序列的序列的pix蛋白。衍生出pvii蛋白或pix蛋白所对应的优选的丝状噬菌体是m13、fd和f1噬菌体。可以使用任何合适的pix蛋白,只要它具有将poi展示为pix融合蛋白的能力。优选地,由本发明的载体编码的pix蛋白对应于pix蛋白。优选地,由本发明的载体编码的pix蛋白包含以下氨基酸序列或由以下氨基酸序列组成,所述氨基酸序列对应于来自vcsm13辅助噬菌体(genbankay598820.1)的野生型pix蛋白。
msvlvysfasfvlgwclrsgityftrlmetss
(seqidno:15)
在其他实施例中,编码的pix蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:15具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。这些变体pix序列应保留或具有将poi展示为pix融合蛋白的功能能力。还可以使用seqidno:15(或这些同源序列)或其他pix序列的功能性截短或片段,只要将poi展示为pix融合蛋白的能力被保留。
在本发明中,“序列同一性”是在氨基酸水平上的蛋白质之间的同一性的度量以及在核苷酸水平上的核酸之间的同一性的度量。当进行序列比对时,可以通过比较每个序列中的给定位置中的氨基酸序列来确定蛋白质序列同一性。类似地,当进行序列比对时,可以通过比较每个序列中的给定位置中的核苷酸序列来确定核酸序列同一性。当在本文中提及变体或衍生物分子时,例如通过百分比同一性值,例如至少70%等,那么应当理解,这样的变体或衍生物通常是功能性变体或衍生物,即展示出保留的或改善的功能。
确定两个氨基酸序列或两个核酸序列的百分比同一性的方法在本领域中是众所周知的并且被描述,并且可以使用这些方法中的任何一种。例如,为了确定两个氨基酸序列或两个核酸序列的百分比同一性,将序列比对以用于最佳比较的目的(例如,可以在第一个氨基酸或核酸序列中引入缺口,用于与第二个氨基酸或核酸序列进行最佳比对)。然后比较在相应的氨基酸位置或核苷酸位置的氨基酸残基或核苷酸。当第一序列中的位置被与第二序列中的相应位置相同的氨基酸残基或核苷酸占据时,分子在
那个位置是相同的。两个序列之间的百分比同一性是所述序列共享的相同位置数的函数(即,%同一性=相同位置数/位置总数(例如,重叠位置)×100)。在一些实施例中,两个序列是相同的长度。
可以手动地比对序列并计算相同氨基酸的数量。可替代地,可以使用数学算法来完成用于确定百分比同一性的两个序列的比对。这种算法被并入(altschul等人,1990)的nblast和xblast程序中。blast核苷酸搜索可以用nblast程序进行,得分=100,字长=12,以获得与本发明的核酸分子同源的核苷酸序列。blast蛋白质搜索可以用xblast程序进行,得分=50,字长=3,以获得与本发明的蛋白质分子同源的氨基酸序列。为了获得用于比较目的的缺口比对,可以利用gappedblast。可替代地,psi-blast可以用于执行迭代搜索,其检测分子之间的远距离关系。当使用nblast、xblast和gappedblast程序时,可以使用相应程序的默认参数。参见http://www.ncbi.nlm.nih.gov。可替代地,可以在序列已经例如通过embl数据库(www.ncbi.nlm.gov/cgi-bin/blast)中的blast程序被比对后,计算序列同一性。通常,关于例如“评分矩阵”和“缺口罚分”的默认设置可以用于比对。在本发明的上下文中,blastn和psiblast默认设置可以是有利的。在计算百分比同一性时,仅计算精确匹配。
在本发明的优选实施例中,在载体构建体中包括一个或多个核糖体(核糖体)结合位点(rbs)。这样的组分也可以被称为翻译起始区(tir)。
rbs序列在载体中位于rbs序列起作用的适当位置。rbs的作用是在蛋白质翻译的启动期间募集核糖体,并且因此可以方便地置于期望翻译的蛋白质的起始密码子的上游或者期望翻译的蛋白质的orf上游的适当距离处。因此,在本发明的载体中,rbs序列方便地置于编码信号肽的序列的上游(在其中信号肽是orf的一部分的实施例中,例如对于荧光团-pviii融合蛋白而言)。在其中orf中不存在信号肽的实施例中,例如在其中poi与第二非pviii外壳蛋白融合的一些实施例中,将rbs序列方便地置于编码poi或外壳蛋白的序列上游的适当距离处。适当的距离将是已知的或由本领域技术人员根据所选择的rbs容易确定的。示例性的距离可以是距atg起始密码子7个或8个核苷酸,但这可以变化。
rbs/tir序列调节位于下游的序列的翻译强度(蛋白质表达水平),并且不同类型的rbs可以产生不同水平的蛋白质表达,例如,弱表达或强表达。弱或强rbs/tir序列在本领域中是众所周知的,并且本领域技术人员可以根据所需的蛋白质表达水平容易地选择。如所预期的,与弱rbs相比,强rbs促进或诱导更多的翻译(强翻译)。如下面将更详细地解释的,在本发明的优选实施例中,弱rbs特别地用于驱动荧光团-pviii融合蛋白的翻译。
特别地,在本发明的优选的载体中,rbs被包括在编码信号肽的序列的起始密码子的上游(或5'或n端),所述信号肽将蛋白质导入tat分泌途径。用于本发明的优选的rbs是shinedalgarno(sd)序列或基于sd的序列,其可以包括在载体构建体中。sd序列在本领域中是众所周知的并被描述,并且可以使用这些中的任何一种。例如,核心sd序列是gagg(seqidno:47),而其他共有序列是aggagg(seqidno:48)或aggaggu(seqidno:49)。因此,可以使用包含这些核心或共有序列的sd序列。
弱或强sd序列在本领域中是众所周知的,并且本领域技术人员可以容易地选择,以便控制任何给定orf的蛋白质表达的水平或量。例如,强sd将能够很好地结合至核糖体。这种强sd通常将精确地或几乎没有改变地含有上面概述的sd核心序列或共有序列之一,和/或例如将在附近含有多个a和/或g残基。可替代地,可以通过插入其他组分与sd核心或共有序列(或高度相关的序列)结合来获得强sd。这样的另外的组分对于本领域技术人员而言是众所周知的,例如经典的或共有的sd序列可以与epsilon序列(例如,ttaacttta,seqidno:50)结合使用。例如,在本发明中,优选的强sd是t7g10tir(olins等人,1988,《基因》73:227-235),其还包括epsilon序列(例如,ttaacttta,seqidno:50)。因此,在其中使用强sd的本发明的载体的具体示例中,tat信号肽的atg起始位点的上游区域可以具有序列
相比之下,弱sd序列通常将含有与本文概述的核心或共有sd序列更加不同的序列。弱sd序列将仍然能够与核糖体结合,但以较低的水平或较低的效率结合。例如,用于本发明的弱sd序列可以含有一个或两个来自本文概述的核心共有sd序列的修饰。用于本发明的载体的优选的弱sd序列包含或由aggaga,seqidno:30组成(即含有与共有序列aggagg,seqidno:48的一个核苷酸差异)。这一弱sd序列优选地位于距离atg起始位点8个核苷酸的位置,尽管这一距离有时可以如上面所讨论的变化。因此,在其中使用弱sd的本发明的载体的具体示例中,tat信号肽的atg起始位点的上游区域可以具有序列
如所预期的,与将促进或诱导低量(或弱或低效)翻译的弱sd相比,强sd促进或诱导更多翻译(强或高翻译)。
强或弱rbs或sd序列均可以用于本发明的载体构建体中。本发明的目的之一是通过增加并入到噬菌体的外壳中(表面上)的荧光团-pviii融合蛋白的数量来增加噬菌体颗粒的荧光。这通常将有利于使用强sd序列,以便获得更多的翻译。因此,有些令人惊讶的是,在一些实施例中,弱sd(或弱rbs)序列产生更好的结果,例如,就荧光团-pviii融合蛋白的更好的功能展示,或荧光团-pviii融合蛋白的增加的展示(例如,每颗粒更多拷贝)而言,优选地导致噬菌体颗粒的增加的荧光。
因此,在一些实施例中,在编码包含tat信号肽和荧光团-pviii融合蛋白的orf的序列上游的载体构建体中使用弱sd(或弱rbs)序列是优选。换句话说,控制或驱动tat信号肽的翻译并因此控制或驱动荧光团-pviii融合蛋白的rbs(或sd)是弱sd(或弱rbs)。优选地,当与其中使用强sd(或强rbs)的构建体相比时,或者当使用共有sd序列,例如aggagg(或如上定义的其他共有序列)时,使用这种弱sd序列导致在噬菌体的外壳中并入完整荧光团-pviii融合蛋白的更多拷贝。
事实上,其中荧光团-pviii融合蛋白的翻译效率低、弱、降低或非最佳的载体构建体通常是优选的。例如,如本文其他地方所描述的优选的修饰/变体分子是其中例如与原始亲本序列相比翻译效率或速度,例如并且特别是荧光团-pviii融合蛋白的翻译效率或速度降低或显著降低的那些。优选地,这种降低的或弱的(或减少的)翻译效率或速度也导致噬菌体的表面上荧光团-pviii融合蛋白的更多拷贝和/或噬菌体的表面上功能性荧光团的增加的数量,以及因此增加的荧光。
在本发明的优选实施例中,尽管是任选的,但在载体构建体中可以包含一个或多个标签,例如蛋白质或肽标签。这样的标签可以方便地用于检测,并且因此,在这些实施例中,可以使用任何可检测的标签。标签也可以用于其他目的,如富集含标签的融合蛋白,例如在进行噬菌体荧光的筛选(例如,通过facs)之前。当使用facs技术或其他荧光检测技术时,抗原肽标签(或表位标签)如flag、c-myc、ha(血凝)、his、hat或v5标签特别优选地用于本发明的实施例中,因为这些标签易于允许通过使用已知的和容易获得的针对特定标签的荧光标记的抗体来标记。同样可以使用本领域中已知的其他抗原肽标签。
flag标签是用于本发明的载体的优选标签。flag标签是八肽标签并且包含序列dykddddk(seqidno:10)。还可以使用flag标签衍生物,其在本领域中是众所周知的且标准的。此外,可以识别各种flag标签并因此可以被荧光标记并用于检测flag标签的抗体在本领域中也已被很好地描述,例如m1和m2抗体。
用于本发明的其他标签(作为抗原肽标签并且特别是flag标签的替代物或除了抗原肽标签并且特别是flag标签外)是生物素标签,其可以例如通过结合至固相上的链霉亲和素或抗生物素蛋白或抗生物素蛋白样分子而被捕获,或者可以通过使用链霉亲和素或抗生物素蛋白或抗生物素蛋白样分子的荧光标记形式被标记,例如被荧光标记。用于本发明的构建体的合适的生物素标签对于技术人员而言将是众所周知的。用于本发明的方法中的生物素标签可以包含生物素分子本身,例如诸如通过化学缀合附着于噬菌体表面上的表达的荧光团-pviii融合蛋白的生物素分子。用于附着这样的生物素标签的方法在本领域中是众所周知的。
可替代地,生物素标签可以包含这样的部分,例如肽,它们可以充当生物素化反应的底物,并且从而变得附着于生物素分子。这样的肽标签可以容易地被并入到本发明的载体中。例如,用于本发明的示例性的生物素标签是avitagtm(msglndifeaqkiewhe,seqidno:52),其是来自美国科罗拉多州奥罗拉市艾维蒂有限公司(avidityllc)的市售标签,所述市售标签通过生物素连接酶在体外或体内变得生物素化。
优选的生物素标签的另一个示例是strep-tag,所述strep-tag还可以从德国哥廷根的ibagmbh商购获得,并且能够结合至链霉亲和素的生物素结合口袋。优选地,strep-tag包含8个氨基酸的序列trp-ser-his-pro-gln-phe-glu-lys(wshpqfek,seqidno:53)。同样可以使用能够结合至链霉亲和素或抗生物素蛋白的任何其他蛋白质标签或肽标签。
一种或多种上述标签的组合可以用于标记噬菌体颗粒,使得存在多个标记。
优选的标签是蛋白质标签或肽标签,其可以例如使用标准的重组和克隆技术容易地并入到本发明的载体构建体(例如,噬菌体展示构建体)中。
标签序列可以在载体中位于用于使标签起作用(例如,被表达和可检测)的任何适当的位置。因此,这样的标签可以在构建体的正在被标记并且将使用标签检测的部分的内部、n端或c端。在本发明中,标签可以方便地用于检测构建体的荧光团-pviii部分的表达。因此,标签可以方便地放置在编码荧光团-pviii融合蛋白的构建体(orf)的部分内,例如放置在荧光团-pviii融合蛋白的内部或者荧光团-pviii融合蛋白的n端(或附近),或者荧光团-pviii融合蛋白的c端(或附近)。
在本发明的载体中,用于编码蛋白质标签的序列的优选位置在荧光团-pviii融合体中的荧光团的n端处或附近。例如,编码蛋白质标签的序列可以位于信号肽和荧光团之间。
在本发明的优选实施例中,蛋白质标签是flag标签(或其衍生物)或另一种带负电荷的标签。不希望受到理论的束缚,据信具有带负电荷的标签,优选地位于荧光团的n端处或附近,例如在信号肽和荧光团之间,可以导致改善的稳定性并因此导致荧光团-pviii融合蛋白的折叠,这是在其他β-结构中观察到的现象(schaefer,2012,《分子生物学杂志(j.mol.biol.)》,417:309-335和dudgeon等人,2012,《美国科学院院报》,109:10879-10884)。
用于具有荧光团-pviii融合蛋白的本发明的构建体的示例性结构在图1中示出。如本文其他地方所描述的,如图1中所示的flag标签(或其他标签)是优选的,但是任选的。
在本发明的优选载体中,在编码荧光团的序列和编码pviii噬菌体外壳蛋白的序列之间包括编码间隔子或接头的序列。所述序列可以帮助折叠连接的蛋白质,特别是n端蛋白质(此处通常是荧光团),并且可以适当地调节间隔子或接头的长度,以使得能够实现两种组分的最佳或令人满意的功能折叠。适当的长度可以由本领域技术人员容易地确定。然而,示例性的长度将在5个和15个氨基酸之间(weiss等人,2000,《蛋白质科学(proteinsci.)》,9:647-654),例如,6个至10个氨基酸。用于本发明的特定接头为8个氨基酸长。接头的序列通常与长度无关。然而,在示例性的载体中使用的接头具有序列gggsgggs(seqidno:6,例如其由seqidno:5编码)。因此,对于本发明的一些实施例,这一接头是优选的,但是应当理解,也可以使用具有其他序列和长度的接头(间隔子)。
本发明的优选的载体(或核酸分子)编码包含seqidno:a或16(其包含mneongreen、接头和pviii的n端30个氨基酸)的序列或与其具有至少70%同一性的序列(用于百分比同一性的其他示例性值在本文其他地方描述)。
本发明的其他优选的载体(或核酸分子)编码包含seqidno:c或18(其包含mneongreen、接头和全长pviii)的序列或与其具有至少70%同一性的序列(用于百分比同一性的其他示例性值在本文其他地方描述)。
发明的其他优选的载体(或核酸分子)编码包含seqidno:e或20(其包含torab7、mneongreen、接头和全长pviii)的序列或与其具有至少70%同一性的序列(用于百分比同一性的其他示例性值在本文其他地方描述)。
本发明的其他优选的载体(或核酸分子)编码包含seqidno:g或22(其包含torab7、flag标签、mneongreen、接头和全长pviii)的序列或与其具有至少70%同一性的序列(用于百分比同一性的其他示例性值在本文其他地方描述)。
还提供了载体(或核酸分子),其编码包含seqidno:k或26(其包含torab7、mng、接头和pviii的n端30个氨基酸)或seqidno:m或28(其包含torab7、flag标签、mng、接头和pviii的n端30个氨基酸)的序列,或与其具有至少70%同一性的序列(用于百分比同一性的其他示例性值在本文其他地方描述)。
这些载体序列(例如,编码seqidno:a或16、c或18、e或20、g或22、k或26或m或28的序列,或其变体)中的任何一种是可以用于亲本载体并且经历如下所概述的进一步变化或修饰的序列的示例。
在图9中示出了用于本发明的进一步的示例性的载体序列(seqidno:54和55)。
例如在seqidno:c或18、e或20、g或22、54或55中的优选的pviii变体,在seqidno:8的位置33或对应于seqidno:8的33位具有缬氨酸至异亮氨酸突变(即,在seqidno:8的33位或在对应于seqidno:8的位置33具有异亮氨酸残基),或在seqidno:7的残基97至99处或在对应于seqidno:7的残基97至99处具有ata异亮氨酸密码子,如本文其他地方所描述的。特别优选的pviii变体是如本文其他地方所述的如在f03克隆中发现的pviii序列。
如上面所描述的本发明的噬菌体展示载体也可以用于产生进一步修饰的或变异的载体,所述修饰的或变异的载体可以用于产生荧光噬菌体颗粒。这样的修饰的(或变异或衍生的或突变的)载体也构成本发明的一部分。这样的修饰(或突变)涉及例如在亲本载体的核酸序列中添加、缺失、取代或插入一个或多个核苷酸,以形成新的载体,其中所述亲本载体是如上定义的本发明的载体之一,并且测试所得的新载体以鉴定可以用于产生具有改善性质的荧光噬菌体颗粒的载体。这样的方法可以用于形成多个新的载体(方便地是新载体的文库),可以测试所有这些新的载体产生荧光噬菌体颗粒,优选地改善的荧光噬菌体颗粒的能力。
所述修饰,例如以一种或多种核苷酸(以及因此编码的氨基酸)的添加、缺失、取代或插入的形式,可以发生在载体的任何功能结构域中,即发生在信号肽(优选地tora,更优选地torab7)、荧光团(优选地mneongreen)、荧光团和pviii、pviii之间的接头、或编码载体的部分的标签(优选地flag标签)中的一个或多个(或全部)中。优选地,修饰位于荧光团(优选地mneongreen)、荧光团和pviii之间的接头或pviii编码区中的一个或多个(或全部)中。在修饰位于pviii编码区的情况下,在一些实施例中,突变位于pviii的n端一半,例如在前90个核苷酸(或者pviii的前30个氨基酸,例如前25至28个或25至30个氨基酸)中。
对亲本载体的这样的修饰或突变可以使用本领域众所周知的和文献记录的技术以任何合适的方式进行,例如通过进行随机或定向诱变的方法。优选地,所产生的突变是取代,并且这些突变可以方便地并且优选地使用随机诱变来制备,特别是当需要产生和筛选多个(例如,一个文库)突变体以选择显示出改善的特性的突变体时。可以以任何合适的方式例如通过易错pcr或使用突变大肠杆菌菌株进行随机诱变。一种优选且方便的技术在示例中描述并且涉及通过pcr并入dntp类似物以将随机突变引入到载体(例如,噬菌粒)dna中。商业试剂盒可用于进行这样的随机诱变。在本文其他地方描述的实施例中,在进行单个或仅几个(例如,多达5个)突变,例如,如本文所描述的pviii蛋白中的缬氨酸至异亮氨酸(v至i)突变(或编码的核苷酸序列中的相应突变)的情况下,定向诱变或其中特定残基可以被靶向和修饰的其他类型的诱变是合适的。
通过这些方法产生的新载体,当被转化为合适的宿主细胞时,将优选地产生具有改善的功能特性,优选地改善的荧光特性的荧光噬菌体颗粒。
因此,本发明进一步提供了能够产生噬菌体颗粒的变体或突变的载体,所述噬菌体颗粒与原始的起始野生型或亲代噬菌体颗粒(即,由非突变的或野生型亲本载体产生的噬菌体颗粒)相比表现出荧光强度的改善(改善的亮度)。这样的改善可以通过以下来提供:a)增加细菌噬菌体颗粒的表面上的荧光团的数量,例如经荧光团与病毒衣壳蛋白融合,通过增加荧光团-pviii融合蛋白在噬菌体表面/外壳中的并入或整合,作为来自产生的大肠杆菌宿主的噬菌体颗粒组装的固有特性(换句话说,增加每颗粒的荧光团-pviii分子的数量或平均数量),或b)增加单个荧光染料(fluorochrome)的亮度,例如增加融合蛋白中荧光部分的固有亮度(荧光强度),或c)a)和b)的组合,即增加亮度和荧光团的数量。可以单独使用,或者与a)和/或b)结合使用的另一种选项将是改善荧光团的功能展示,例如通过改善(例如,更有效)折叠或增加的溶解度,因为大肠杆菌中的异源蛋白质表达通常导致一部分翻译的但非功能性的蛋白质产物,例如不溶性蛋白质产物。因此,还可以通过增加噬菌体表面上的功能性荧光团的数量来提供改善。
在a)的情况下,没有必要增加单个荧光团的亮度。因此,这一结果可以通过显示与起始荧光团分子相同的荧光强度的荧光团来实现,或者,实际上,甚至单个荧光团分子的降低的荧光强度可以是容许的,因为更多数量的荧光分子将展示在每个单个噬菌体的表面上,从而可以补偿这种降低。
可以使用任何适当的测定来筛选含有突变的载体的噬菌体颗粒,其在细菌噬菌体的表面上具有增加数量的荧光团。例如,某种可检测标签(其可在噬菌体的表面上检测到)可以方便地用于进行这种筛选。优选地,这些标签将独立于由荧光部分所显示的荧光水平,使得可以评估噬菌体的表面上的荧光部分的数量的指示。可检测标签的示例将是本领域技术人员众所周知的。例如,可以使用flag标签。可以使用针对所讨论的标签的抗体方便地进行检测。facs,例如使用荧光标记的抗体,例如针对flag标签的抗体,是特别优选的,因为它更容易地允许通过监测和选择在相关荧光轴上在正方向上移动的颗粒来筛选单个颗粒。除了使用可检测标签外,特异性地识别荧光团的抗体(如抗mneongreen32f6(chromotekgmbh))或类似抗体可以被用于facs。在实例中示出了示例性的方法,例如基于facs的方法,或能够测量来自所讨论的荧光团的荧光信号的类似方法(例如,flisa)。
在b)的情况下,没有必要增加待获得的单个噬菌体颗粒的表面上的荧光团的数量。因此,这一结果可以通过在单个噬菌体颗粒的表面上相同数量的荧光团来获得,或者实际上,甚至减少数量的单个荧光团分子可以是容许的,因为增加单个荧光染料的亮度,例如增加荧光部分的固有亮度(荧光强度)可以补偿这种减少。
任何合适的测定可以用于筛选含有突变的载体的噬菌体颗粒,其在细菌噬菌体的表面上表达的融合蛋白中具有单个荧光染料的增加的亮度。例如,结合荧光团的另一种定量方法,例如,使用蛋白质印迹,或使用针对荧光团的抗体,例如,如在实例中所示的facs分选中使用的抗neongreen纳米体,facs或一些其他类型的基于荧光的测定可以用于测量荧光的水平,例如,测量溶液中噬菌体颗粒的荧光水平。facs是特别优选的,因为它更容易允许通过监测和选择在相关荧光轴上在正向方向上移动的颗粒来筛选单个颗粒。在实例中示出了示例性的方法,例如基于facs的方法或flisa。
优选地,这样的增加(以及实际上如在本文中其他地方提到的其他增加、改善或积极效果)是可测量的增加等(视情况而定),更优选地,它们是显著的增加,优选地是统计上显著的增加,例如当与适当的对照水平或值相比(例如,与用非突变的或亲代的或野生型载体获得的荧光的水平相比)时,概率值≤0.05。荧光强度的优选的增加可以是比亲代载体大或至少大两倍或三倍,例如多达5倍、10倍或20倍的增加。可以用于评估这样的增加或改善的亲代载体的方便的比较器或基线水平可以是包含编码seqidno:e或20或seqidno:g或22或seqidno:c或18的序列(或者实际上是seqidno:a或16、k或26或m或28)的载体。
已经开发了显示出改善的荧光性质的载体的突变版本,所述载体的突变版本可以产生在表面上表达mneongreen-pviii融合蛋白的荧光噬菌体。这些突变版本中的一些在个体基础上显示出mneongreen荧光部分的改善的亮度(如通过实例2中所示的facs图谱上y轴上移所证明的)。这些突变版本中的其他版本在噬菌体的表面上显示出改善的展示,即荧光团的增加的数量(如通过实例2中所示的facs图谱上沿x轴右移所证明的,其通过用标记的抗体测量flag标签来评估)。这些突变版本中的其他版本显示出基于个体的mneongreen荧光部分的改善的亮度和在细菌噬菌体的表面上荧光团的增加的数量(如通过实例2中所示的facs图谱右上象限中克隆的存在所证明的)。
如上面所描述的,导致噬菌体颗粒的改善的荧光的突变残基可以位于载体的任何部分中,但是优选地位于mneongreen或融合蛋白的pviii部分中(更优选地位于pviii部分中),或者位于mneongreen和pviii之间的接头中。在突变位于pviii区域的情况下,在一些实施例中,突变位于pviii的n端一半,例如在前90个核苷酸(或pviii的前30个氨基酸,例如前25至28个或25至30个氨基酸)中。
如上面所描述的,在本发明的优选实施例中,用于载体的mneongreen部分的编码的氨基酸序列由seqidno:4提供(并且示例性的核苷酸序列由seqidno:3提供)。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的mneongreen部分中的实施例中,这样的变体mneongreen组分可以包含seqidno:3或seqidno:4,或在核苷酸或氨基酸水平上与其具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:3或seqidno:4中分别在核苷酸或氨基酸水平上,优选地在氨基酸水平上包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体mneongreen组分可以包含多达15%,优选地多达13%或10%,更优选地多达5%或4%的突变水平。
突变的mneongreen序列的其他优选的示例是在mneongreen序列例如seqidno:4中含有多达30个,例如多达25个、20个、15个、12个或10个,例如1个或多达2个、3个、4个、5个、6个、7个、8个、9个或10个改变的氨基酸的序列。
如上面所描述的,在本发明的优选实施例中,用于载体的pviii部分的编码的氨基酸序列由seqidno:8提供(并且示例性的核苷酸序列由seqidno:7提供)。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的pviii部分的实施例中,这样的变体pviii组分可以包含seqidno:7或seqidno:8,或在核苷酸或氨基酸水平上与其具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:7或seqidno:8中分别在核苷酸或氨基酸水平上,优选地在氨基酸水平上包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体pviii组分可以包含多达15%,优选地多达13%或10%,更优选地多达5%或4%的突变水平。
突变的pviii序列的其他优选的示例是在pviii序列例如seqidno:8中含有多达10个,例如多达9个、8个、7个、6个、5个、4个、3个、2个或1个改变的氨基酸的序列。
如本文其他地方所描述的,尽管可能存在其他突变残基,但优选的突变载体序列在seqidno:8的残基(或位置)33处,或与其对应的残基处含有缬氨酸至异亮氨酸的取代(即,在seqidno:8的位置33处或对应于seqidno:8的位置33具有异亮氨酸残基)。更优选地,编码异亮氨酸的密码子是ata。例如,缬氨酸密码子gtc可以被异亮氨酸密码子ata取代。因此,在上面讨论的实施例中,其中变体载体可以包含seqidno:8,或这样的序列,即与seqidno:8具有至少70%的同一性,或在seqidno:8中具有多达15%的突变水平,或一定数量例如多达35个改变的氨基酸的序列,在优选的实施例中,存在这一异亮氨酸残基或密码子。
还如上面所描述的,在本发明的一些实施例中,突变的残基存在于pviii蛋白的n端一半中,例如存在于如由seqidno:j或25提供的载体的pviii部分的编码的氨基酸序列中,其含有pviii的前30个氨基酸(具有如由seqidno:i或24提供的示例性核苷酸序列,pviii的前90个核苷酸)。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变的残基中的至少一些位于载体的pviii部分的实施例中,这样的变体pviii组分可以包含seqidno:i或24或seqidno:j或25,或在核苷酸或氨基酸水平上与其具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:i或24或seqidno:j或25中分别在核苷酸或氨基酸水平上,优选地在氨基酸水平上包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体pviii组分可以包含多达15%,优选地多达13%或10%,更优选地多达5%或4%的突变水平。
突变的pviii序列的其他优选示例是在pviii序列例如seqidno:j或25中含有多达5个,例如多达4个、3个、2个或1个改变的氨基酸的序列。
如上面所描述的,在本发明的优选实施例中,用于载体的接头部分(此处我们指的是荧光团和pviii组分之间的接头)的编码的氨基酸序列由seqidno:6提供(并且示例性的核苷酸序列由seqidno:5提供)。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的这一接头部分中的实施例中,这样的变体接头组分可以包含seqidno:5或seqidno:6,或在核苷酸或氨基酸水平上与其具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:5或seqidno:6中分别在核苷酸或氨基酸水平上,优选地在氨基酸水平上包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体接头组分可以包含多达15%,优选地多达13%或10%,更优选地多达5%或4%的突变水平。
突变的接头序列的其他优选示例是在接头序列例如seqidno:6中含有1个、2个或3个改变的氨基酸的序列。在其他实施例中,在接头序列例如seqidno:6中不存在突变。
如上面所描述的,在本发明的一些实施例中,用于载体的torab7部分的编码的氨基酸序列由seqidno:2提供(并且示例性的核苷酸序列由seqidno:1提供)。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的torab7部分中的实施例中,这样的变体torab7组分可以包含seqidno:2或seqidno:1或与其具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:1或seqidno:2中分别在核苷酸或氨基酸水平上,优选地在氨基酸水平上包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体torab7组分可以包含多达15%,优选地多达13%或10%,更优选地多达5%或4%的突变水平。
突变的torab7序列的其他优选的示例是在torab7序列例如seqidno:2中含有多达6个,例如多达5个、4个、3个、2个或1个改变的氨基酸的序列。
在本发明的其他优选实施例中,可以在mneongreen、mneongreen和pviii之间的接头以及pviii编码区中的一种或多种(或全部)中发现突变。在一些优选的实施例中,可以在pviii编码区中发现突变。在突变位于pviii区域的情况下,在一些实施例中,突变位于pviii的n端一半,例如在前90个核苷酸(或pviii的前30个氨基酸,例如前25至28个或25至30个氨基酸)中。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的这些区域的实施例中,这样的变体载体可以包含seqidno:a或16或seqidno:c或18或与seqidno:a或16或c或18具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分在seqidno:a或c中可以包含多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体组分可以包含多达15%,优选地13%或10%,更优选地多达5%或4%的突变水平。
突变的载体序列的其他优选的示例是在seqidno:a或16或c或18中含有多达30个,例如多达25个、20个、15个、12个或10个,例如1个或多达2个、3个、4个、5个、6个、7个、8个、9个或10个改变的氨基酸的序列。
如本文其他地方所描述的,尽管可以存在其他突变的残基,但是优选的突变的载体序列在seqidno:c或18的残基276处,或与之对应的残基处含有缬氨酸至异亮氨酸取代(即,在seqidno:c或18的位置276处或对应于seqidno:c或18的位置276处具有异亮氨酸残基)。更优选地,编码异亮氨酸的密码子是ata。例如,缬氨酸密码子gtc可以被异亮氨酸密码子ata取代。因此,在以上讨论的实施例中,其中变体载体可以包含seqidno:c或18,或这样的序列,即与seqidno:c或18具有至少70%的同一性,或在seqidno:c或18中具有多达15%的突变水平,或一定数量例如多达35个改变的氨基酸的序列,在优选的实施例中,存在这一异亮氨酸残基或密码子。
在本发明的其他优选实施例中,可以在mneongreen、mneongreen和pviii之间的接头、pviii编码区和torab7信号肽中的一种或多种(或全部)中发现突变。在一些优选的实施例中,可以在pviii编码区中发现突变。
在突变位于pviii区域的情况下,在一些实施例中,突变位于pviii的n端一半,例如在前90个核苷酸(或pviii的前30个氨基酸,例如前25至28个或25至30个氨基酸)中。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的这些区域中的实施例中,这样的变体载体可以包含seqidno:e或20或seqidno:k或26或在氨基酸水平上与seqidno:e或20或seqidno:k或26具有至少70%同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%同一性)的序列。换句话说,这样的变体组分可以在seqidno:e或20或seqidno:k或26中包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体组分可以包含多达15%,优选地13%或10%,更优选地多达5%或4%的突变水平。
突变的载体序列的其他优选的示例是在seqidno:e或20或k或26中含有多达35个,例如多达30个、25个、20个、17个、15个、12个或10个,例如1个或多达2个、3个、4个、5个、6个、7个、8个、9个或10个改变的氨基酸的序列。
如本文其他地方所描述的,尽管可以存在其他突变残基,但是优选的突变载体序列在seqidno:e或20的残基317,或与之对应的残基处含有缬氨酸至异亮氨酸取代(即,在seqidno:e或20的位置317处或对应于seqidno:e或20的位置317处具有异亮氨酸残基)。更优选地,编码异亮氨酸的密码子是ata。例如,缬氨酸密码子gtc可以被异亮氨酸密码子ata取代。因此,在以上讨论的实施例中,其中变体载体可以包含seqidno:e或20,或这样的序列,即与seqidno:e或20具有至少70%的同一性,或在seqidno:e或20中具有多达15%的突变水平,或一定数量例如多达35个改变的氨基酸的序列,在优选的实施例中,存在这一异亮氨酸残基或密码子。
在本发明的其他优选实施例中,可以在mneongreen、mneongreen和pviii之间的接头、pviii编码区、torab7信号肽和flag序列中的一种或多种(或全部)中发现突变。
在突变位于pviii区域的情况下,在一些实施例中,突变位于pviii的n端一半,例如在前90个核苷酸(或pviii的前30个氨基酸,例如前25至28个或25至30个氨基酸)中。因此,在其中导致噬菌体颗粒的保留或改善的荧光的突变残基中的至少一些位于载体的这些区域中的实施例中,这样的变体载体可以包含seqidno:g或22或seqidno:m或28或与seqidno:g或22或seqidno:m或28具有至少70%的同一性(例如,至少75%、80%、85%、87%、88%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性)的序列。换句话说,这样的变体组分在seqidno:g或22或seqidno:m或28中可以包括多达30%、25%、20%、15%、13%、12%、10%、9%、8%、7%、6%、5%、4%、3%、2%或1%的突变水平。优选的变体组分可以包含多达15%,优选地13%或10%,更优选地多达5%或4%的突变水平。
突变的载体序列的其他优选示例是在seqidno:g或22或m或28中含有多达35个,例如多达30个、25个、20个、17个、15个、12个或10个,例如1个或多达2个、3个、4个、5个、6个、7个、8个、9个或10个改变的氨基酸的序列。
如本文其他地方所描述的,尽管可以存在其他突变残基,但优选的突变的载体序列在seqidno:g或22的残基325,或与其对应的残基处含有缬氨酸至异亮氨酸取代(即,在seqidno:g或22的位置325处或对应于seqidno:g或22的位置325具有异亮氨酸残基)。更优选地,编码异亮氨酸的密码子是ata。例如,缬氨酸密码子gtc可以被异亮氨酸密码子ata取代。因此,在以上讨论的实施例中,其中变体载体可以包含seqidno:g或22,或这样的序列,即与seqidno:g或22具有至少70%的同一性,或在seqidno:g或22中具有多达15%的突变水平,或一定数量例如多达35个改变的氨基酸的序列,在优选的实施例中,存在这一异亮氨酸残基或密码子。
在其中载体含有flag标签的这样的实施例中,优选的是flag标签保持未修饰或未突变,例如,保留seqidno:10的序列(dykddddk)。
在特别优选的实施例中,本发明的载体含有突变,使得突变的序列或变体序列显示出降低的或显著降低的或较弱的或减小的翻译效率或速度。因此,其中荧光团-pviii融合蛋白的翻译效率低、弱、降低、减小或非最佳的载体构建体通常是优选的。优选的修饰/变体分子是这样的分子,即其中例如与原始亲本(未突变的或未修饰的)序列相比,翻译效率例如且特别是荧光团-pviii融合蛋白的翻译效率降低或显著降低(或减小),其具体示例在本文其他地方描述。举例来说,这可以通过具有突变的pviii组分来实现,使得pviii蛋白(以及因此含有荧光团-pviii融合蛋白的整个orf)的翻译效率降低或显著降低。然而,同样可以设想的是,荧光团-pviii融合蛋白的其他部分中的突变例如在荧光团(例如,mneongreen荧光团)中或在接头序列中或在信号肽中的突变可以导致降低的翻译效率。
具有降低的翻译效率的变体可以通过本领域技术人员设计或已知的适当方法来筛选或测试。例如,可以对归一化的宿主细胞数量和通过独立于蛋白质功能的手段(例如,通过蛋白质印记)定量的蛋白质产生进行时间追踪测定,以评估与相关对照(例如,非变体或野生型融合蛋白)相比,在被测试降低的翻译效率的系统中蛋白质的绝对量是否较低。
优选地,这样的减小(以及实际上如本文其他地方提到的其他减小、降低或负面影响)是可测量的减小等(视情况而定),更优选地,它们是显著的减小,优选地是统计学上显著的减小,例如当与适当的对照水平或值相比(例如,与用非突变的或亲本或野生型载体或其他相关对照获得的翻译水平,例如翻译效率或速度)相比时,概率值≤0.05。
尽管不希望受到理论的束缚,但据信这一降低的翻译效率或速度,以及例如融合蛋白在宿主细胞中的更逐渐的或更缓慢的积累,最终导致当与其中观察到正常或高翻译效率的载体构建体(例如亲本构建体,其具体实例在本文其他地方描述)相比时,更多荧光团-pviii融合蛋白的拷贝被并入噬菌体的外壳中。可替代地,噬菌体的外壳中功能性荧光团融合体的数量增加,而不必改变所并入的单位的绝对数量。由于本发明的目的之一是通过增加并入噬菌体的外壳(表面上)的荧光团-pviii融合蛋白的数量或通过增加噬菌体的表面上的功能性融合蛋白的数量来增加噬菌体颗粒的荧光,因此有些令人惊讶的是,在一些实施例中,降低的翻译效率,特别是荧光团-pviii融合蛋白的降低的翻译效率,产生更好的结果,例如,就荧光团pviii融合蛋白的更好的功能展示和/或荧光团pviii融合蛋白的增加的展示(例如,每颗粒更多的拷贝)而言,这可以导致噬菌体颗粒的增加的荧光。
导致噬菌体颗粒的改善的荧光的突变的一个示例是在seqidno:8中所示的pviii蛋白的残基33处将v修饰成i。如在实例中所描述的在克隆f03中发现了这一突变,与亲本序列相比,换句话说,与在这一位置不具有v至i突变的序列相比,所述克隆f03显示出具有改善的荧光。有趣的是,当检查核酸序列时,在亲本/野生型/未修饰的pviii中的位置33处的编码缬氨酸的密码子gtc已被改变为编码异亮氨酸的密码子ata。这是大肠杆菌中极为罕见的密码子,并且因此被认为有助于显著降低f03克隆中荧光团-pviii融合蛋白的翻译效率。因此,其他合适的突变将是那些导致在核酸序列中并入稀有密码子(在大肠杆菌中稀有的密码子)如ata的那些突变(在相应的rna序列中t当然是u)。适当的稀有密码子将是本领域技术人员众所周知的,然而一些示例可能是稀有的arg密码子,如agg、cga、aga或cgg、或cta(leu)、ccc(pro)或tcg(ser)。另一个示例可能是将正常的atg起始/启动密码子交换为一个更稀有的密码子,例如更稀有的gtg密码子。
如从实验实例中可以看出,在这一位置处v至i突变的存在,例如通过存在gtc缬氨酸密码子至ata异亮氨酸密码子的改变,导致显著改善的荧光,例如比用野生型/亲本/未修饰的pviii序列所观察到的荧光高2倍(或高100%)。观察到的荧光已经很亮。实际上,据信通过表达亲本载体而赋予的荧光对于本领域中已知的噬菌体颗粒表现出最亮的荧光。因此,使这一荧光加倍是明显有利的改善。
因此,如本文其他地方所描述的,尽管可以存在其他突变残基,但是优选的突变载体序列具有pviii序列,其在seqidno:8的残基33,或与其对应的残基处含有缬氨酸至异亮氨酸取代(即,在seqidno:8的位置33处或对应于seqidno:8的位置33具有异亮氨酸残基)。更优选地,编码异亮氨酸的密码子是ata。例如,缬氨酸密码子gtc可以被异亮氨酸密码子ata取代。因此,在如本文其他地方所描述的实施例中,其中变体载体可以包含seqidno:8,或seqidno:8的变体,例如这样的序列,即与seqidno:8具有至少70%的同一性,或在seqidno:8中具有多达15%的突变水平,或一定数量例如多达35个改变的氨基酸的序列,在优选的实施例中,存在这一异亮氨酸残基或密码子。因此,在优选的载体构建体中,在载体中含有(或编码)的pviii噬菌体外壳蛋白或变体序列在seqidno:8的位置33或相应位置处具有缬氨酸至异亮氨酸突变(异亮氨酸残基)。优选地,编码所述异亮氨酸残基的核酸序列包含密码子ata。
同样地,尽管可以存在其他突变残基,但是在核苷酸水平上,编码pviii蛋白的优选的突变载体序列在seqidno:7的残基97置99或与其对应的残基处含有gtc至ata取代。因此,在如本文其他地方所描述的实施例中,其中变体载体可以包含seqidno:7或seqidno:7的变体,例如与seqidno:7具有至少70%同一性的序列,于是在优选的实施例中,存在编码异亮氨酸的这一ata密码子。
因此,优选的载体构建体包含pviii噬菌体外壳蛋白,所述pviii噬菌体外壳蛋白包含seqidno:61,或与seqidno:61具有至少70%同一性的序列。可替代地,pviii噬菌体外壳蛋白由包含seqidno:60的序列或与seqidno:60具有至少70%同一性的序列编码。
例如,在一些实施例中,编码的pviii蛋白包含氨基酸序列或由氨基酸序列组成,所述氨基酸序列与seqidno:61具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。如上所述,这些变体pviii序列应保留或具有将荧光团展示为pviii融合蛋白的功能能力。还可以使用seqidno:61(或这些同源序列)或其他pviii序列的功能性截短或片段,只要将荧光团展示为pviii融合蛋白的能力被保留。
同样地,编码pviii蛋白的核酸分子包含核苷酸序列或由核苷酸序列组成,所述核苷酸序列与seqidno:60具有至少70%、75%或80%的序列同一性,如至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、或99%的同一性。这些变体pviii核苷酸序列应保留或具有编码荧光团作为可以在噬菌体颗粒上展示的pviii融合蛋白的功能能力。还可以使用seqidno:60(或这些同源序列)或其他pviii序列的功能性截短或片段,只要编码荧光团作为可以在噬菌体颗粒上展示的pviii融合蛋白的能力被保留。
如实例中所示的f03克隆包含seqidno:2的torab7序列、seqidno:10的flag标签、seqidno:4的mneongreen荧光团、seqidno:6的接头序列和seqidno:61(由seqidno:60编码)的pviii噬菌体外壳蛋白。因此,尽管flag标签是任选的,但是包含或编码这些序列中的一个或多个或全部的载体是优选的。包含编码seqidno:4的mneongreen荧光团、seqidno:6的接头序列和seqidno:61(由seqidno:60编码)的pviii噬菌体外壳蛋白的序列的载体是优选的,包含图9中所示的序列的载体同样也是优选的,但是其中pviii噬菌体外壳蛋白具有或包含seqidno:61(由seqidno:60编码)所示的氨基酸序列。如本文其他地方所描述的,也预期与这些序列中的一个或多个或全部具有至少70%同一性的变体。然而,在这样的变体序列中,优选的是异亮氨酸残基存在于seqidno:61或其变体序列的残基33(或相应的残基)处。还优选的是,这一异亮氨酸残基由密码子ata(如在seqidno:60中发现的)编码。
在一些其他实施例中,如上所述的通过荧光团-pviii融合蛋白(或编码核酸)的pviii(或其他)组分中的突变而表现出荧光团-pviii融合蛋白的降低的翻译效率的变体(特别是v至i变体/突变体)的使用,可以与导致荧光团-pviii融合蛋白的降低的翻译效率的其他特征结合,如在编码包含tat信号肽和荧光团-pviii融合蛋白的orf的序列上游的载体构建体中使用弱sd(或弱rbs)序列,如本文其他地方所述。
本发明的优选的载体可以编码第二噬菌体外壳蛋白融合体,所述第二噬菌体外壳蛋白融合体包含感兴趣的蛋白(poi)或与除pviii外的噬菌体外壳蛋白融合的靶向单元。这样的第二噬菌体外壳蛋白融合体优选地被设置在与荧光团-pviii融合蛋白相同的载体上。然而,第二外壳蛋白融合体可以被设置在单独的载体上。示例性的非pviii噬菌体外壳蛋白对于本领域技术人员而言将是众所周知的,例如可以使用与piii、pvi、pvii或pix的融合体。这样的实施例允许在噬菌体颗粒的两种不同外壳蛋白上同时展示poi,例如靶向蛋白和荧光团。优选的第二融合体将是感兴趣的蛋白(poi)或pix的靶向单元。
因此,本发明的载体可以进一步包含与编码非pviii噬菌体外壳蛋白的序列融合(基因地融合)的编码poi的序列(核酸序列)。poi和非pviii噬菌体外壳蛋白在载体中可以处于以任何合适的顺序或间隔,只要一旦表达并包装成噬菌体颗粒,在poi和非pviii外壳蛋白之间形成功能性融合蛋白,其中融合蛋白的非pviii外壳蛋白组分形成噬菌体外壳的一部分,并且poi在噬菌体颗粒的表面上被功能性地表达或展示。因此,融合蛋白的poi部分与融合蛋白的非pviii外壳蛋白部分位于框架内。在本发明的载体中,第二融合蛋白的poi组分可以位于第二融合蛋白的非pviii噬菌体外壳蛋白组分的n端(或在n端或在n端附近)或c端(或在c端或在c端附近)。这通常将基于所使用的非pviii噬菌体外壳蛋白的类型来确定,并且对于本领域技术人员来说将是显而易见的。
在其中使用poi与第二噬菌体外壳蛋白的融合的实施例中,优选的是外壳蛋白融合体(即pviii和非pviii融合体)中的每一个都被表达为单独的开放阅读框。因此,在这样的实施例中,载体可以含有两个独立的核糖体结合位点(rbs)或sd或tir区域,一个指导荧光团-pviii融合蛋白的翻译,而另一个指导poi-第二外壳蛋白融合蛋白的翻译。任何适当的rbs等可以用于指导poi-第二外壳蛋白融合蛋白的翻译(例如,如本文其他地方所描述的强序列、弱序列或共有序列)。因此,用于两个外壳蛋白融合体的rbs序列可以相同或不同,并且示例在本文其他地方描述。
用于具有两个融合蛋白的这种构建体的示例性结构在图2中示出。
在这样的载体中,编码两个融合蛋白的序列可以处于任何顺序,尽管在一些实施例中,poi融合蛋白位于荧光团-pviii融合蛋白的n端。
如本文其他地方所描述的,信号序列或信号肽可以被包括在非pviii融合蛋白的orf中。这将在很大程度上取决于所使用的非pviii融合蛋白的类型。piii融合蛋白通常将需要信号肽,而信号肽对于pvii和pix融合蛋白是任选的(即存在或不存在),而信号序列通常将不用于pvi。例如,pvi通常被用作c端融合体。在其中使用第二信号肽的本发明的一些实施例中,将使用除tat之外的信号肽。
在这样的实施例中,还优选的是两种外壳蛋白融合体在相同启动子序列的控制下表达,尽管在其他实施例中可以使用单独的或不同的(例如,2个)启动子序列。示例性的启动子序列对于本领域技术人员来说将是众所周知的,并且可以使用这些序列中的任何一种。示例性的启动子序列可以是lac启动子,其可以例如用iptg诱导。其他启动子可以包括tac、arabb或psp。
重要的是,本文示出的结果表明pviii-荧光团融合蛋白在噬菌体表面上的表达不干扰在同一噬菌体颗粒上的不同外壳蛋白(例如,pix)上展示的poi的结合能力,并且特别是靶向蛋白的结合能力(例如,抗体的抗原结合能力)。因此,pviii-荧光团融合蛋白可以与感兴趣的蛋白在任何其他噬菌体外壳蛋白上的表达结合使用。
任何感兴趣的蛋白(poi)都可以编码在本发明的载体中,只要其适合于在噬菌体上展示。优选的示例是靶向分子/靶向单元或结合配偶体,所述结合配偶体可以结合至(靶向)其他实体。poi的一些优选的示例将是抗体或其片段(例如fab、scfv、纳米体)、mhc分子(i类或ii类)、t细胞受体或非ig衍生的结合蛋白,如darpins、ankyrin家族、纤连蛋白家族、结蛋白(knottins)、抗运载蛋白(anticalin)等(hosse等人,2006,《蛋白质科学》15:14-27)和肽。
本发明的载体可以用于经典的噬菌体展示,以便为特定的靶实体(例如,靶蛋白)选择结合配偶体(例如,抗体)。在这样的应用中,poi的文库可以在噬菌体颗粒上被表达作为非pviii融合蛋白(第二外壳蛋白融合体)的一部分,并通过标准和众所周知的技术选择其与靶实体结合。由荧光-pviii融合蛋白提供的可检测噬菌体表面荧光可以用于筛选和检测。
因此,poi通常是外源的或异源的,就像荧光团一样。当提及外源或异源蛋白质时,是指最初不是相关噬菌体外壳蛋白的一部分的蛋白质或肽,例如piii、pvi、pvii、pix或pviii蛋白等,其与噬菌体外壳蛋白(或其片段)融合(具有或没有任何接头氨基酸),例如与噬菌体外壳蛋白的n端融合。
可以存在于本发明的载体(或核酸分子)中的其他任选特征对于本领域技术人员而言将是众所周知的。例如,载体,例如噬菌体载体或噬菌粒载体(其可以被统称为噬菌体展示载体或构建体)可以任选地另外含有其他合适的组分,例如复制起点、用于启动转录的诱导型或非诱导型启动子/操作子、增强子、终止序列、抗生素抗性基因和标记物,编码伴侣蛋白的序列(例如,周质伴侣蛋白,如fkpa)、信号序列、接头、蛋白酶位点、通用标签或报告分子、能够进行克隆和其他操作的限制性位点、能够通过例如pcr扩增构建体的引物结合位点或其他期望的序列元件,例如允许通过例如pcr区分不同文库的dna序列。在噬菌体展示构建体中这些额外的组分的适当的来源和定位使得它们发挥其所需的功能在本领域技术人员的正常实践中将是很好的。
可以用于表达本发明的载体和核酸序列并包装和产生荧光噬菌体颗粒的用于噬菌体展示的合适的细菌宿主对于本领域技术人员而言将是众所周知的,并且因此可以被选择。优选的细菌宿主细胞是革兰氏阴性菌,如大肠杆菌菌株。示例性的大肠杆菌菌株包括xl-1blue、tg1、er2738、avb100fmkii’、mc1061、ss320和top10f’。在一些实施例中,xl-1blue菌株是优选的。
用于本发明的优选的细菌宿主例如大肠杆菌宿主是已经被工程改造以过表达tat转运蛋白或tat转运蛋白的组分(例如,过表达蛋白tata、tatb和tatc)的宿主。用于这种过表达的方法对于本领域技术人员而言将是众所周知的,例如通过在表达载体上表达tat转运蛋白的组分,例如至少蛋白tata、tatb和tatc。这些组分(例如,蛋白tata、tatb和tatc)的序列和来源在本领域中是可,例如从标准数据库中容易地获得。示例性的方法在所附的实例中示出。这些宿主细胞然后可以被转染或以其他方式提供有本发明的载体,并且用于产生荧光噬菌体颗粒。
如实验实例中所示出的,除了在宿主细胞中产生的和由宿主细胞分泌的噬菌体颗粒的改善的荧光(增加的荧光强度)之外,例如,与当使用过表达非tat转运蛋白的细菌宿主细胞时所观察到的荧光相比,使用这样的宿主使得能够实现表达噬菌体颗粒的细菌细胞的改善的荧光(增加的荧光强度)。这一改善的荧光可以是显著的,例如荧光强度的所观察到的和优选的增加可以是比当使用过表达非-tat转运蛋白的宿主细胞时所观察到的荧光大或至少大两倍或三倍,例如多达5倍、10倍、20倍或30倍的增加。
这样的细菌宿主细胞,例如大肠杆菌,例如xl-1blue或如本文所述的其他细菌宿主细胞,形成了本发明的又一方面,这些细菌宿主细胞被工程改造以异源表达或过表达tat转运蛋白,例如至少异源表达或过表达蛋白tata、tatb和tatc。
如本文其他地方所描述的,本发明的载体用于噬菌体展示,并且因此可以是噬菌粒载体或噬菌体载体。
噬菌体展示是本领域众所周知和描述的技术。在这方面,在1985年,g.p.smith建立了一种在丝状噬菌体的表面上展示多肽的方法,所述丝状噬菌体是一种感染大肠杆菌细胞的病毒(smith,g.p.,1985,《科学》228,1315-1317)。从那时起,所谓的噬菌体展示已发展成为一种用于蛋白质工程以及结合特定靶的多肽和蛋白质的选择的强有力的技术(
在噬菌体展示中,通常将编码感兴趣的蛋白(poi)的基因置于编码外壳蛋白(最常见的是piii)的基因和其n端信号序列之间,以产生poi-外壳蛋白融合体,尽管在本发明的一些实施例中,信号序列并不总是存在。术语“噬菌体文库”是指独特的噬菌体的集合,所述独特的噬菌体在poi的氨基酸序列上不同,并且是通过标准分子克隆技术制备的。文库很有可能含有>1010个成员,并且可以用于特定的结合物的选择。
因此,本发明的另一方面提供了一种荧光噬菌体例如荧光丝状噬菌体的文库,其由如本文所描述的本发明的载体(或核酸分子)产生并因此包含本发明的载体(或核酸分子)。所述荧光噬菌体包含荧光团-pviii融合蛋白。任选地,所述丝状噬菌体还将poi或poi的文库展示为与piii、pvi、pvii或pix的融合体。与其他噬菌体展示文库一样,每个单独的噬菌体颗粒表达/展示同一poi,但是多个颗粒的存在允许展示多个(或一个文库或多个)不同的poi。
对于涉及噬菌体展示的本发明的方面,关于相关的技术和定义,可以参考通用噬菌体展示教科书,如sachdevs.sidhu的《生物技术和药物发现中的噬菌体展示(phagedisplayinbiotechnologyanddrugdiscovery)》,1995,或barbas等人的《噬菌体展示:实验室手册(phagedisplay:alaboratorymanual)》,1994。
荧光团-外壳蛋白融合体和poi-外壳蛋白融合体可以通过将编码荧光团或poi的序列插入到编码噬菌体基因组中相关外壳蛋白的序列中而被编码在完整的噬菌体基因组中(噬菌体载体展示),或者被编码在噬菌粒上(噬菌粒展示)。噬菌粒是一种高拷贝数质粒,其可以编码荧光团-pviii和任选地poi-非pviii外壳融合蛋白,并且需要与辅助噬菌体重叠感染,所述辅助噬菌体提供噬菌体产生所需的遗传物质。因此,在噬菌粒展示中,通常存在用于荧光团和任选地poi展示的两种来源的外壳蛋白;辅助噬菌体编码的野生型蛋白,和噬菌粒编码的荧光团-pviii和任选地poi-外壳蛋白融合体。新的病毒体于是将具有荧光团-pviii/poi-非pviii外壳蛋白融合体和野生型外壳蛋白的混合物。类似地,如果使用噬菌体基因组系统,那么野生型噬菌体蛋白通常也需要存在,尽管在本发明的一些实施例中,不存在野生型的poi-非pviii外壳蛋白。
然而,在本发明中,对噬菌体颗粒进行工程改造以具有在非pviii外壳蛋白上展示的poi的一个或多个拷贝是可能的。这可以通过所使用的辅助噬菌体来控制,并且在本发明的优选实施例中,使用了具有非pviii外壳蛋白上展示的poi的多个拷贝的噬菌体颗粒。这可以以任何适当的方式来实现。然而,特别地,为了增加展示水平并在噬菌体的表面上产生poi的多个拷贝(这也可以增加结合的亲和力),可以使用被称为δ噬菌体(deltaphage)的修饰型辅助噬菌体,所述修饰型辅助噬菌体允许在pix上进行高价展示。这样的修饰的辅助噬菌体与仅允许低价展示的常规辅助噬菌体如m13k07、vcsm13、r408或类似物的使用形成对比。
被称为δ噬菌体的辅助噬菌体是由nicolayrustadnilssen(nilssen等人,2012,《核酸研究(nucleicacidsresearch)》,40,e120;wo2011/036555)开发的。在这一辅助噬菌体中,在pix起始密码子附近插入至少一个(例如,两个)琥珀色突变,从而有条件地灭活辅助噬菌体编码的pix。如果这一辅助噬菌体然后被重叠感染到用编码poi-pix融合体的噬菌粒转化的宿主细胞(例如,大肠杆菌)中,则在抑制琥珀色突变的宿主细胞(例如,supe+菌株)中可以看到poi-pix的中间价展示,而在不抑制琥珀色突变的宿主细胞(例如,supe-菌株)中可以看到poi-pix的高价展示。这是因为,在supe+宿主细胞株(例如,supe+大肠杆菌)中,来自辅助噬菌体的野生型pix被翻译以给出中间价展示,因为野生型pix与poi-pix竞争展示,并且显示两者的混合物。然而,在辅助噬菌体上的野生型pix的产生在supe-菌株中被阻断,并且仅噬菌粒编码的poi-pix将存在,从而导致高价展示。
因此,在本发明的优选实施例中,载体构建体是噬菌粒载体,所述噬菌粒载体除了荧光团-pviii融合蛋白外还包含poi-pix融合蛋白,并且这种载体构建体与具有条件突变的辅助噬菌体结合使用,使得可以控制野生型pix噬菌体蛋白的表达,这进而可以使得能够控制噬菌体的表面上poi-pix融合蛋白的数量。在其中条件突变未被抑制的实施例中,野生型pix将不会被表达,并且仅poi-pix将被表达,从而导致在表面上的仅poi-pix融合蛋白和高价展示。在其中条件突变被抑制的实施例中,野生型pix蛋白将被表达,从而在表面上产生野生型pix和poi-pix融合蛋白的混合物。
合适的条件突变对于本领域技术人员而言将是众所周知的,并且可以很容易地设计辅助噬菌体载体,使得pix处于条件突变的控制之下。例如,如上面所描述,在被称为δ噬菌体的辅助噬菌体中,一个或多个可抑制终止密码子形式的条件突变(例如,琥珀色突变)与合适的宿主细胞结合使用,以抑制或不抑制条件突变。
术语“噬菌粒”是本领域的术语,并且指的是一种克隆载体,所述克隆载体作为丝状噬菌体ff和质粒的杂交体被开发,以产生可以作为质粒繁殖并且还被包装为病毒颗粒中的单链dna的载体。与质粒相似,噬菌粒可以用于克隆dna片段,并且通过一系列技术(转化、电穿孔)被引入到细菌宿主中。然而,用“辅助”噬菌体(例如vcsm13或m13k07)感染含有噬菌粒的细菌宿主提供了必要的病毒成分,以使得能够实现单链dna复制和将噬菌粒dna包装成噬菌体颗粒。
如本文其他地方所描述的,本发明的另一方面提供了噬菌体颗粒,其包含本发明的载体(或核酸分子),并在表面上表达功能性荧光团-pviii融合蛋白并且还任选地表达包含poi的第二外壳融合蛋白。噬菌体颗粒因此可以包含噬菌体基因组或噬菌粒。
噬菌体,通常被称为细菌噬菌体,在本文中以其本领域公认的形式使用,意指感染、复制并由细菌分泌的病毒。丝状细菌噬菌体,或丝状噬菌体,是具有单链dna基因组(ssdna基因组)的噬菌体,其包装有噬菌体外壳蛋白。分泌的丝状噬菌体颗粒在表型上具有丝状结构。
如本文所使用的术语噬菌体或丝状噬菌体涵盖噬菌体基因组衍生的病毒体和噬菌粒衍生的病毒体。
术语“辅助噬菌体”是本领域的术语,并且指的是这样的病毒,即所述病毒通过感染已经被缺陷型病毒(例如,噬菌粒)占据的同一宿主细胞并提供缺陷型病毒(例如,噬菌粒)缺失且需要完成其生命周期并形成病毒体(例如,含有噬菌粒)的蛋白质来帮助分离的和不相关的缺陷型病毒(例如,噬菌粒)繁殖,所述病毒本身不是噬菌体基因组或功能性病毒,而仅仅是含有一种或若干种衍生自噬菌体基因组的元件的质粒。
用于本发明的优选的辅助噬菌体在本文其他地方描述,并且包括m13k07(stratagene)、高噬菌体(hyperphage)(progenbiotechnikgmbh)、r408(安捷伦科技公司(agilenttechnologies))和vcsm13(stratagene)。在优选的实施例中,辅助噬菌体可以是具有如所描述的条件性(或可抑制性)突变的辅助噬菌体,例如,如本文所述的δ噬菌体构建体,或phaberge、ex-噬菌体、vcsm13d3或r408d3。
由本发明提供的另一方面是一种包含本发明的载体(或核酸分子)的噬菌体展示系统。优选的噬菌体展示系统包含本发明的载体(或核酸分子),例如本发明的噬菌粒载体和辅助噬菌体,例如本文所定义的。本发明的其他优选的噬菌体展示系统包含本发明的载体(例如,本发明的噬菌粒载体)和细菌宿主细胞(例如,大肠杆菌宿主细胞),所述细菌宿主细胞过表达tat转运蛋白,例如至少过表达蛋白tata、tatb和tatc。这样的宿主细胞也在本文其他地方描述,并且可以作为组分包括在本文描述的所有噬菌体展示系统、试剂盒、方法和用途中。
用于本发明的方法的本文描述的另一方面是包含本发明的载体(或核酸分子)的试剂盒或包含如上面所描述的噬菌体展示系统的试剂盒,其例如由本发明的噬菌粒和辅助噬菌体(优选地如本文所描述的辅助噬菌体,例如,如本文所描述的具有条件性(或可抑制)突变的辅助噬菌体)构成,或包含本发明的载体(例如,本发明的噬菌粒载体)和细菌宿主细胞(例如,大肠杆菌宿主细胞)的试剂盒,所述细菌宿主细胞过表达tat转运蛋白,例如至少过表达蛋白tata、tatb和tatc。所述试剂盒还可以包括附带有必要的使用说明。
包含在这样的试剂盒中的本发明的优选的载体(或核酸分子)可以包含如本文所述的荧光团-pviii载体,所述荧光团-pviii载体包含编码tat信号肽的序列和编码荧光团-pviii融合蛋白的序列。任选地,载体(或核酸分子)也可以包含例如,如本文所描述的编码第二噬菌体外壳蛋白(非pviii噬菌体蛋白)的序列,和一个或多个适合于在poi中克隆的克隆位点(例如,多克隆位点),其然后将与第二噬菌体蛋白融合。
因此,优选的试剂盒可以包含用于产生荧光噬菌体颗粒的试剂的集合或由所述集合组成,所述荧光噬菌体颗粒包含荧光团-pviii融合蛋白,任选地具有poi与非pviii外壳蛋白的第二融合蛋白。除了本发明的载体之外,试剂盒可以包括一种或多种选自以下的组分:其他噬菌粒、辅助噬菌体、细菌菌株和说明书。用于这样的附加组分的优选选项如本文其他地方所述。
本发明的又进一步的方面提供了本发明的载体构建体、核酸分子、噬菌体展示系统或试剂盒产生荧光噬菌体颗粒的用途,或在噬菌体展示中的用途。换句话说,本发明提供了一种用于产生荧光噬菌体颗粒的方法(或噬菌体展示的方法),所述方法包括使用本发明的载体构建体、核酸分子、噬菌体展示系统或试剂盒。
如本文定义的本发明的荧光噬菌体颗粒也可以用作用于体外应用和测定的分子工具。颗粒可以用于任何其中需要可检测荧光试剂的测定。
因为本发明的优选的噬菌体颗粒还展示可以是特异性结合配偶体或靶向单元例如如本文其他地方所述的抗体等的poi,这些可以用作特异性结合对或靶向试剂的成员,并且这样的荧光噬菌体颗粒可以用于任何其中需要特定结合对成员或靶向单元的测定。于是,荧光团可以容易地用于检测颗粒,并且因此检测poi与其靶的结合。对于这些用途来说,重要的是,本文已经表明,荧光团-pviii在噬菌体颗粒的表面上的展示不会干扰在另一种噬菌体外壳蛋白上展示的靶向模块。
本发明的荧光噬菌体颗粒也可以用于任何涉及荧光检测的测定,特别是荧光染色(例如,在溶液中或在细胞上)。因此,用于颗粒的优选的测定是facs分析或免疫荧光或flisa测定(其具有与facs分析可比较的检测系统)。在免疫荧光或flisa测定中使用颗粒的能力避免了对抗体染色(或检测)的需要,而仅依赖于荧光团-pviii融合体的固有荧光进行检测。在facs分析中使用颗粒的能力将提供显著的优点,即能够进行实时选择(例如,实时噬菌体展示),以避免对抗体染色(或检测)的需要,而仅依靠荧光团-pviii融合体的固有荧光进行检测。也考虑用于体外诊断的用途。
因此,本发明的又进一步的方面提供了一种包含如本文所定义的本发明的荧光噬菌体颗粒的试剂,和这样的荧光噬菌体颗粒作为分子工具的用途,例如在体外测定中。
在下表中总结了本文所提及的一些序列以及相关的标识符
根据本技术领域的惯例,此表中的所有序列均在本文中以5’至3’叙述。
附图说明
将参考以下非限制性实例并参考以下附图进一步描述本发明,在附图中:
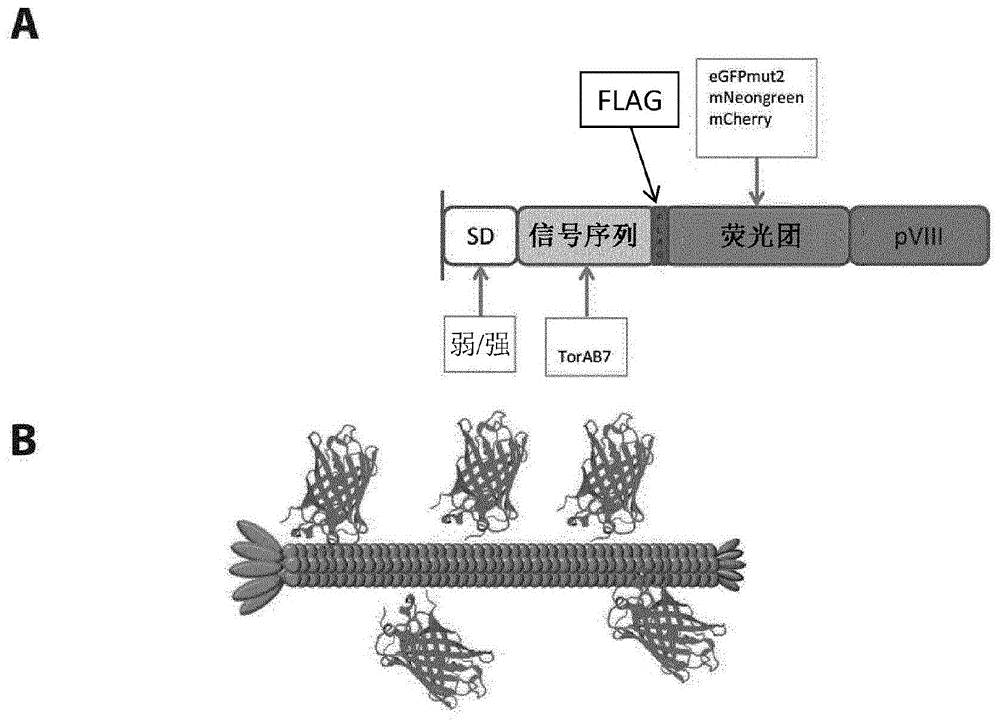
图1a示出了图示本文所描述的荧光团-pviii噬菌粒构建体的一些组分的示意图。sd=shinedalgarno序列(其可以是弱的或强的);sig序列=信号序列(此处显示为torab7,但可以是将蛋白质导向tat分泌途径的任何信号序列);flag=flag标签;pviii=噬菌体pviii外壳蛋白。方框示出了制造和测试的在载体的sd、信号序列和荧光团部分中的各种组分的变体。除了指定的变化外,所有变体在序列上是相同的。图1b示出了荧光团-pviii展示荧光噬菌体颗粒的草图。噬菌体颗粒将具有沿着噬菌体颗粒的长度随机分布的荧光团的多个拷贝的。
图2a示出了图示本文描述的双重展示噬菌粒构建体的一些组分的示意图。lac=lac启动子;sd=shinedalgarno序列;poi=感兴趣的蛋白;pix=噬菌体pix外壳蛋白;sig序列=信号序列;flag=flag标签;pviii=噬菌体pviii外壳蛋白。方框示出了制造和测试的在载体的sd、信号序列和荧光团部分中的各种组分的变体。除了指定的变化外,所有变体在序列上是相同的。图2b示出了双重展示荧光噬菌体颗粒的草图。取决于辅助噬菌体,噬菌体颗粒将具有在pix上展示的poi的一个或多个拷贝,和沿着噬菌体颗粒的长度随机分布的荧光团的多个拷贝。
图3示出了用噬菌粒构建体染色的大肠杆菌,所述噬菌粒构建体含有与pviii融合的荧光团mneongreen、mcherry和mgfpmut2。将转化的大肠杆菌细胞在30℃下生长过夜,以允许荧光团的表达。将细胞培养物加入到载玻片中,并且使用共聚焦显微镜观察荧光。mneongreen和egfpmut2在大多数细胞中显示出清晰和明亮的荧光。mcherry显示出弱的但可检测的荧光。
图4示出了在噬菌粒拯救中荧光团的评估。a)在大肠杆菌top10f’的100ml培养物中繁殖含有弱或强sd序列、torab7或gp8信号序列和mneongreen或mgfpmut2荧光团的荧光噬菌体噬菌粒的六种变体,随后通过m13k07或δ噬菌体辅助噬菌体进行拯救。噬菌体通过peg沉淀浓缩,随后进行感染性滴定,并且结果以cfuampr/ml给出。b)噬菌粒与辅助噬菌体的比率通过cfuampr/cfukanr来确定。c)将拯救的荧光噬菌体颗粒归一化至1×1012cfuampr,并使用fitc滤光片在victor3多标记板读取器中测试荧光。荧光强度以任意单位给出。
图5示出了通过使用不同大肠杆菌菌株对荧光强度的影响的评估。a)四种不同的大肠杆菌菌株用带有或不带有flag标签的荧光噬菌体进行转导,且生长过夜并在od600nm上归一化。细菌荧光用varioskan多模式板读取器测量,并且荧光强度以任意单位给出。b)使用δ噬菌体从四种不同菌株中拯救的归一化的荧光噬菌体样品(荧光噬菌体颗粒)在varioskan多模式板读取器中分析荧光强度。荧光强度以任意单位给出。
图6示出了在elisa和flisa中对荧光性能的评估。a)在victor3板读取器上分析归一化的噬菌体样品的荧光强度,并且强度以任意单位给出。b)将稀释系列的归一化噬菌体样品加入到用1μg/mlphox-bsa或牛奶块包被的孔中。捕获的噬菌体用抗m13hrpmab检测并用可溶性tmb进行显影。c)在victor3板读取器中平行分析相同的稀释系列的荧光信号。
图7示出了通过流式细胞术对bwtcr4b2a1细胞的荧光染色。始终使用5×1011cfu/ml到2x105个细胞的归一化噬菌体输入。a)转染的bw4b2a1细胞的染色。结合的噬菌体用pe缀合的抗fdigy检测。b)未转染的bw细胞的染色。结合的噬菌体用pe缀合的抗fdigy检测。c)bw4b2a1细胞的染色。结合的噬菌体通过固有噬菌体荧光来检测。d)在victor3板读取器上分析归一化的噬菌体样品的荧光强度,并且强度以任意单位给出。
图8示出了与突变体文库一起使用的分选/门控策略的概述。第一轮是基于细胞的分选和基于噬菌体的克隆富集的组合,具有高展示水平。第二轮集中于具有mneongreen信号和高水平的表面flag(双阳性细胞)的克隆。使用sanger测序、流式细胞术和荧光测量来分析单个克隆。
图9示出了flag_mng_pviii载体的序列(其是本发明的优选实施例);核苷酸序列以seqidno:54提供,并且氨基酸序列以seqidno:55提供。这一载体的torab7、flag、mng(mneongreen)、接头和pviii组分均被示出。
图10示出了原始荧光噬菌体(mngwt)与来自筛选的主要候选物(mngf03)的比较。(a)在没有葡萄糖的情况下,将含有不同噬菌粒的xl1-blue细胞在37℃培养16小时,以允许融合蛋白的表达。细胞在pbs中洗涤3次。测量细胞的光密度,并基于光密度将所有样品调节至1的a600值。制备以1:2步骤的系列稀释。(b)噬菌体按照标准方案使用δ噬菌体进行包装。样品浓度通过感染性滴定来确定,并且所有样品均被归一化为5,0e+12cfuamp/ml。在varioskanlux上以488ex/517em进行荧光测量。
图11:在phox-pix融合系统的上下文中对pviii-mngf03克隆的评估。(a)在包装过程期间取出细胞。测量来自起始培养物(含有葡萄糖)的荧光,以建立用于荧光测量(glu)的基线。在过夜包装(没有葡萄糖)后,测量相同细胞的荧光。基于光密度将所有样品均归一化至1的a600值。制备以1:2步骤的系列稀释。(b)噬菌体的浓度基于样品中存在的dna量来确定。将原始数据调整至最低值4,18e+12。
图12示出了通过流式细胞术对skw-3r12-c9细胞的荧光噬菌体染色。始终使用5×1012cfu/ml到5×104个细胞的归一化的噬菌体输入。a)转染的skw-3r12-c9细胞的染色。结合的噬菌体用apc-缀合的抗fdigy检测。b)未转染的skw-3细胞的染色。结合的噬菌体用apc-缀合的抗fdigy检测。c)skw-3r12-c9细胞的染色。结合的噬菌体通过固有噬菌体荧光来检测。d)在varioskanlux板读取器上分析归一化的噬菌体样品的荧光强度,并且强度以任意单位给出。
图13示出了在elisa和flisa中对荧光噬菌体性能的评估。a、b)将稀释系列的归一化的噬菌体样品(在pngf03中使用不同滴度,tat高在x轴上指示)添加到用1μg/mlphox-bsa或牛奶块包被的孔中。在varioskanlux板读取器中分析捕获的噬菌体的荧光信号。c)相同的样品用抗m13hrpmab检测,并用可溶性tmb显影。d)在varioskanlux板读取器上分析归一化的噬菌体(5.0e+12cfuamp/ml)样品的荧光强度,并且强度以任意单位给出。
图14示出了细菌荧光水平的比较。除mngf03噬菌粒外还含有用于tatac和b基因的表达载体的xl1-blue细胞在37℃下在不存在葡萄糖的情况下培养16小时,以允许融合蛋白的表达。没有噬菌粒的细胞用作阴性对照。将细胞在pbs中洗涤3次。测量细胞的光密度,并基于光密度将所有样品调节至1的a600值。制备以1:2步骤的系列稀释。在varioskanlux上以488ex/517em进行荧光测量。
具体实施方式
实例1-通过使用周质靶向在主要衣壳蛋白pviii上功能性生物荧光团的有效的m13展示
材料和方法
双重展示噬菌粒的构建
所有荧光团-pviii构建体均在计算机上组装,并从genscript(中国)订购。将所述片段插入到携带scfvgb113(nilssen,等人,2012,《核酸研究》,40,e120)的pgald9δlfn噬菌粒(
含有mneongreen的flag标签和n端部分的基因片段从genscript订购,并使用snabi和bsgi限制性酶切位点插入,以便产生flag-mneongreen。这一噬菌粒的一般结构在图2a中的示意图中示出。这一载体序列的一部分在图9中示出。
以类似的方式制备编码mcherry和mgfpmut2荧光团的噬菌粒。
噬菌体产生
将含有100μg/ml氨苄青霉素、20μg/ml四环素和0.1葡萄糖(yt-tag)的5ml的2×yt培养基用来自含有相应噬菌粒的大肠杆菌xl1-blue的甘油原液的细胞接种,并在37℃下在定轨摇床上生长过夜。在0.025的od600下用预培养物接种200ml的yt-tag,并在37℃下在摇动下生长。培养物在0.3至0.4的od600用辅助噬菌体δ噬菌体(nilssen等人,2012,见上文)的moi10感染,所述辅助噬菌体δ噬菌体是一种有条件地将其同源pix敲低的辅助噬菌体,允许pix-poi的高价展示。在孵育60分钟后,将培养物沉淀并重悬于补充有100μg/ml氨苄青霉素和50μg/ml卡那霉素的2×yt中,并在28℃下继续振荡16小时。通过离心去除细菌细胞,并通过0.2μm真空驱动过滤器(密理博(millipore))过滤,并与聚乙二醇(peg)/nacl溶液(20%peg8000,2.5mnacl)以1:5混合。在冰上孵育过夜后,将样品离心(5000×g,45分钟,4℃),并将沉淀溶解在25ml磷酸盐缓冲盐水(pbs)中。将样品与peg/nacl以1:5混合,并且在冰上孵育4小时,随后如前所述离心,并将沉淀重悬于1ml的pbs中。如koch,j.等人,2000,《生物技术》,29,1196-1198,2002中所述,通过感染性斑点滴定来评估病毒体滴度。
荧光测量
使用fp-8500分光荧光计(日本分光(jasco))确定激发/发射光谱。在平衡仪器的串扰与灵敏度时,我们将激发和发射的间隔设置为15nm,同时允许发射滤光片接受±2.5nm的变化。对于激发和发射,以5nm的间隔进行测量。
细菌的荧光测量
将带有荧光噬菌体mneongreen或荧光噬菌体mneongreen-flag噬菌粒的大肠杆菌的单个菌落接种到补充有100μg/ml氨苄青霉素的5ml2×yt中,并在37℃下孵育过夜。沉淀细胞,并重悬于1×pbs中,随后基于od600nm进行归一化。样品以1:2稀释系列制备,并用varioskan多模式板读取器(赛默飞世尔(thermofischer))测量荧光,其中在仪器的限制内(500/525)对mneongreen进行适当的ex/em设置。
载玻片上细菌的共聚焦显微镜
将带有mneongreen、mgfpmut2和mcherry荧光噬菌体噬菌粒的所有三种变体的大肠杆菌的单个菌落接种到5ml的补充有100μg/ml氨苄青霉素的2×yt中,并在37℃下孵育过夜。将3μl的细菌培养物吸移到载玻片上,并将载玻片放在上面。使用fv1000共聚焦激光扫描显微镜(奥林巴斯(olympus))将细菌可视化,每种荧光团均具有合适的可用激光波长(488nm:mneongreen和mgfpmut2,543nm:mcherry)。
噬菌体的荧光测量
噬菌体的荧光强度测量是通过将噬菌体样品(在1×pbs中稀释)进行归一化,并使用带有fitc滤光片(488/510)的victor3多标记读取器(珀金埃尔默(perkinelmer))在1s激发下或使用varioskan多模式板读取器(赛默飞世尔)测量荧光来进行的,其中在仪器的限制内(500/525)对mneongreen进行适当的ex/em设置。
噬菌体捕获elisa
将微量滴定板在4℃下用1μg/mlphox-bsa包被过夜,并用含0.1%吐温(x)和4%非脂脱脂奶粉(pbstm)的pbs封闭。一式三份加入在pbstm中稀释的以2.5×1012cfu/ml开始的噬菌体样品的1:4系列稀释。通过抗m13hrp(安玛西亚生物科学(amershambiosciences)),随后用tmb溶液显影和在610nm处的吸光度读数来检测结合的噬菌体,或者通过在victor3多标记读取器中进行荧光测量来检测结合的噬菌体。
流式细胞术
将2×105bw4b2a1tcr转染细胞和未转染的bw细胞(阴性对照)的等分试样分配到v形96孔培养皿(nunc)中。用5%w/vfcs/pbs(ph7.4)将总体积调节至250μl/孔。将板在室温下以300g/5分钟离心,并弃去上清液。加入在5%fcs/pbs中预先封闭的归一化滴度为5×1011cfuampr/ml的50μl/孔噬菌体的等分试样(对照孔仅接受5%fcs/pbs),并将板在4℃孵育1小时。通过加入200μl/孔用5%fcs/pbs洗涤细胞,通过以300g/5分钟/rt离心沉淀细胞,并且弃去上清液。将在5%fcs/pbs中的pe缀合的鸡抗fd(norwegian抗体)添加到适当的孔中,并且未染色的样品接受5%fcs/pbs,随后在4℃下孵育30分钟。如上洗涤孔,并用200μl/孔的2%pfa固定,并保持在黑暗中,直到在facscalibur(bd生物科学(bdbiosciences))上进行分析。使用flowjo(版本10.2)软件进行数据分析。
结果与讨论
构建
基于pgald9δlfn(
一些变化包括在构建体中(如图2a所示)。为了研究翻译强度的变化的影响,我们包括了弱sd(包含序列aggaga,其中tat信号肽的atg起始位点的上游区域具有序列aaggagacagtcata)或强sd(使用了t7g10tir,其也包括epsilon序列ttaacttta,其中tat信号肽的atg起始位点的上游区域具有序列ttaactttaagaaggagatatacat)。较强的翻译可能会迫使更大量的pviii融合体进入生长的颗粒中。为了研究易位途径的重要性,我们引入了gp8ss或torab7作为周质靶向前导肽。前导肽gp8ss提供了与分泌系统的sec途径的融合体的靶向性,蛋白质通过所述途径以未折叠的状态易位,并允许在周质的氧化环境中发生折叠(manting,e.h和driessen,a.j.,2000,《分子微生物(molmicrobiol)》,37,226-238)。torab7前导肽已经被工程改造以用于改善gfp的周质靶向性(delisa等人,2002,见上文),并将融合体靶向双精氨酸转运(tat)途径,在所述途径中蛋白质在易位前在细胞质的还原条件下折叠。然后将完全折叠的蛋白质易位到周质中(berks,b.c等人,2005,《微生物学的当前观点(curropinmicrobiol)》,8,174-181)。
为了研究各种荧光蛋白如何作为融合体与pviii融合,评估了三种不同的荧光蛋白:作为gfp的facs优化版本的gfpmut2(cormack,b.p等人,1996,《基因》,173,33-38),其被选择用于红移,以使用来自大肠杆菌文库的标准fitc滤光片在facs选择中进行最佳检测。当在大肠杆菌中表达时,突变gfp也被证明是极好的折叠体(folder)。选择了另一种绿色荧光蛋白mneongreen(shaner,n.c等人,2013,《自然方法(natmethods)》,10,407-409)。基于先前用生物荧光团的展示进行的研究的结果(speck等人,2011,见上文),我们还选择包括mcherry(shaner,n.c等人,2004,《自然生物技术》,22,1567-1572),其中无论易位路线如何,它都显示出在功能上被展示。除非另有说明,否则pix融合体是scfvgb113,一种抗体片段,它仅与鼠t细胞受体(tcr)4b2a1反应((bogen,b等人,1990,《欧洲免疫学杂志(eurjimmunol)》,20,2359-2362),并且之前已经示出当在噬菌体上展示时在流式细胞术中表现良好(nilssen,n.r等人,2012,《核酸研究》,40,e120)。
噬菌体产生和验证
为了确认pviii-荧光团融合确实产生了荧光蛋白,我们使用荧光显微镜法检查了转化的大肠杆菌细胞的培养物。为此,我们发现当去除葡萄糖阻遏物时,用带有强sd和sec靶向荧光蛋白的组合的噬菌粒转化的细菌在培养中并未生长,并且因此,这一噬菌粒版本在进一步的实验中被忽略(数据未示出)。
将具有荧光团mneongreen、mcherry和mgfpmut2(图2a)的噬菌粒变体转化到大肠杆菌中,然后使其在30℃下生长过夜,以允许荧光团的表达。这些噬菌粒变体均含有弱sd序列和torab7信号序列。将细胞培养物添加至载玻片,并使用共聚焦显微镜观察荧光。结果在图3中示出,其中可以看到mneongreen和mgfpmut2显示出清晰且明亮的荧光,而mcherry显示出弱荧光。在所有三种荧光团的情况下可检测水平的荧光的存在证明了荧光团被表达并正确折叠。通过蛋白质印迹证实了这一荧光团表达(数据未示出)。因此,上述噬菌粒系统正在展示功能性荧光团-pviii融合蛋白。
由于mneongreen和mgfpmut2比mcherry显示出更好的可检测荧光,因此用这些构建体进行进一步的实验。
为了评估向系统中添加第二外壳蛋白融合体是否会影响噬菌体的产生和/或噬菌粒的包装,我们生产了所有携带有绿色荧光蛋白的荧光噬菌体变体(mgfpmut2和mneongreen),并通过感染滴定法测量了噬菌体的产生和噬菌粒的包装(图4a)。所有噬菌体样品所达到的最终滴度都与之前在我们的pix系统中观察到的最终滴度可比(loset等人,2011,e17433,见上文,和nilssen等人,2012,见上文)。此外,如先前所观察到的,使用δ噬菌体(一种有条件地将其同源pix的辅助噬菌体敲低的辅助噬菌体,允许pix-poi的高价展示)的噬菌粒拯救似乎并未影响噬菌体的产生(nilssen等人,2012,见上文)。弱sd和sec靶向性的组合导致用于mneongreen的辅助噬菌体基因组的优先包装,并且针对mgfpmut2的强sd和tat靶向观察到相同的效果(图4b)。然而,这些结果表明,含有两个外壳蛋白融合体的双重展示噬菌粒被包装,并且允许噬菌体的产生的水平与当仅存在pix融合体时所看到的水平等同。
为了评估存在于荧光噬菌体颗粒上的功能性荧光蛋白的水平,使用fitc滤光片(488nm/510nm)在victor3多标记读取器中分析归一化的荧光噬菌体样品(图4c)。弱sd和tat靶向的mneongreen的组合脱颖而出,因为它超过背景荧光至少三倍。然而,具有强sd和tat靶向性的噬菌体还显示出比背景高大约两倍的强度,而例如具有弱sd和tat靶向性的mgfpmut2噬菌体的强度略高于背景信号。pixpoi的展示价的变化似乎并未影响荧光蛋白在pviii上的功能展示水平。达到高端滴度对于下游应用如文库选择非常重要,并且产生的荧光噬菌体的数量并不比之前在pix系统中观察到的少。荧光噬菌体构建体确实产生了荧光噬菌体颗粒,并且变化中的一种,即具有弱sd的tat靶向的mneongreen,达到了令人满意的信噪比。此处值得注意的是,mneongreen在506nm处具有尖锐的激发峰,并且在517nm处同样具有尖锐的发射峰,这与victor3(485nm/535nm)中使用的荧光素滤光片不匹配。使用这一滤光片测量的强度很可能被严重低估。无论如何,弱sd和tat靶向的mneongreen的组合呈现出是最佳表现的构建体,并且因此被选择用于下游表征。
为了研究和验证当在pviii上展示时mneongreen的激发和发射光谱,在分光荧光计上分析了2×1011个噬菌体颗粒/ml(数据未示出)。作为阳性对照,我们包括了2μg/ml的可溶性egfp,其在预期的波长(490nm/510nm)处显示出清晰和明确的峰。作为阴性对照,我们分析了展示scfvgb113但没有荧光团的噬菌体颗粒。此处,我们在440nm/455nm处观察到尖锐峰。当分析具有mneongreen展示的噬菌体样品时,除了在(505nm/520nm)处由正确折叠的mneongreen所预期的峰外,还出现了相同的尖锐峰(shaner等人,2013,见上文)。
功能展示的优化
我们想调查在标准实验室大肠杆菌菌株中的表达和折叠是否存在任何差异,以及任何差异是否会转化为功能展示水平。此外,我们希望包括用于荧光蛋白表达的检测标签,并向mneongreen的n端中添加flag标签。用荧光噬菌体±flag转化了四种大肠杆菌菌株xl1-blue、avb100f'、ss320和top10f'。
然后直接在活的大肠杆菌细胞中测量荧光强度(图5a)。菌株之间的荧光存在明显差异。以xl1-blue表达的蛋白质给出最高的强度,紧邻其后的是avb100f’。这两个菌株显示出比ss320和top10f’明显更高的荧光强度,尽管在这些菌株中仍清楚地观察到功能表达和折叠。令我们惊讶的是,我们观察到加入flag似乎转化为更高的总体荧光强度。然后这些含有flag的构建体用于测量来自在四种单独菌株中产生的噬菌体样品的荧光强度(图5b),并再次比较包装在四种不同菌株中的来自荧光噬菌体的mneongreen-flag的荧光(图5b),变得清楚的是,xl1-blue是优异的菌株,其次是avb100f'。包装在ss320和top10f'中的噬菌体给出较差的荧光强度。
即使它们密切相关,但不同菌株在荧光团的功能表达和折叠方面显示出相当大的差异。此外,尽管当单独产生pviii-荧光团融合体时,xl1-blue和avb100在细菌荧光方面非常相似,但是在xl1-blue中产生的荧光噬菌体颗粒给出了几乎两倍高的荧光强度。这些数据表明,在xl1-blue和avb100之间共享在胞质溶胶中功能上折叠荧光团的有利功能,这与ss320和top10f’形成对比。然而,与avb100相比,xl1-blue似乎还具有优异的将折叠的荧光团从胞质溶胶运输到周质并将其整合到病毒体中的能力。因此,尽管可以使用这些菌株(或实际上其他大肠杆菌菌株)中的任何一种,但在一些情况下,xl1-blue是优选的。
酶联免疫吸附测定(elisa)和荧光联免疫吸附测定(flisa)
为了评估我们是否可以将在同一噬菌体颗粒上在与pviii不同的衣壳上展示功能性荧光团-pviii融合体和不同功能性融合体的能力结合,我们选择将描述良好的抗phoxscfv(marks等人,1992,《生物技术(biotechnology)》,10:779-783)插入到表达噬菌粒的荧光团-pviii中作为pix融合体。然后,我们制备了噬菌体,并在如上所述的经典的elisa以及flisa中测试了它们的phox结合。为了比较,我们制备了展示没有荧光团-pviii融合体的噬菌体的抗phoxscfv-pix,并进行了平行测试。
在victor3板读取器中测量归一化量的噬菌体颗粒的荧光强度(图6a),且显示出非常高的荧光强度,并且仅来自显示荧光团-pviii融合体的噬菌体。重要的是,当在标准elisa中测试相同噬菌体样品的scfv靶结合时,观察到特异性和相同结合,而与荧光团-pviii融合体存在与否无关(图6b)。因此,数据表明荧光团在pviii上的展示不影响双显示噬菌体上的pix融合体中由靶向部分介导的抗原结合。当使用flisa采用victor3板读取器再次测试相同噬菌体样品的靶结合时,我们无法观察到来自任何样品的荧光信号(图6c)。因此,数据显示,测量应用于孔中的所有噬菌体的荧光是容易完成的,而在当前的设置中,我们不具有特异性检测在flisa中洗涤后保留在靶上的减少量的噬菌体的灵敏度。重要的是,数据不可逆转地表明,荧光团-pviii融合体和scfv-pix融合体在同一噬菌体颗粒上的双重展示不影响scfv-pix融合体的表型。
通过流式细胞术检测细胞表面表达的蛋白质
荧光噬菌体的强有力的应用将是用于facs的实时选择,不需要染色抗体,而仅依靠荧光团-pviii融合的固有荧光进行检测。为了评估这一点,我们使用流式细胞术测试了显示荧光噬菌体的gb113scfv-pix与在鼠4b2a1t细胞杂交瘤细胞上表达的t细胞受体的特异性结合。为了比较,我们制备了展示没有荧光团-pviii融合体的噬菌体的gb113scfv-pix,并进行了平行测试。
为了使用标准方法验证与表达tcr4b2a1的细胞的特异性结合,我们使用了pe缀合的抗fd抗体来检测结合的噬菌体。这也允许荧光噬菌体和常规噬菌体的并排比较,其结合至tcr(图7a),并且两者都不结合至tcr阴性的鼠bw58细胞(pmid:2558022)(图7b)。当试图在不添加检测scfv的情况下对细胞进行染色时,我们观察到与对照相比荧光强度没有增加(图7c),尽管当在溶液中时从荧光噬菌体scfvgb113观察到强荧光信号(图7d)。因此,即使荧光噬菌体在溶液中显示出强荧光信号,但是我们也无法使用流式细胞术检测到它与表面结合抗原的结合。这意味着尽管这一荧光噬菌体在许多测定中非常有用和有利,但这种形式的荧光噬菌体还没有达到允许细胞的直接染色和facs分选所需的每颗粒亮度。
结论评述
已经报道了尝试建立在噬菌体和细胞技术的结合优势上的先前努力,但尚未看到常规使用。先前已经实现了在噬菌体颗粒上荧光蛋白的功能展示,但是到目前为止收效甚微,并且在pviii外壳蛋白上还没有实现。在这一点上,荧光团与分选酶标记的pviii的体外间接偶联已经显示出最大的希望(hess等人,2012,见上文),但是这一程序效率低、费力且昂贵。
我们在此显示了具有固有荧光能力和靶结合活性的噬菌体颗粒的产生。mneongreen被证明是针对在pviii上展示所研究的那些的优异的荧光团。我们发现,用于在pviii上功能展示生物荧光团的最佳条件涉及融合体通过tat途径的易位以用于胞质折叠。我们还惊奇地发现,弱sd给出了更高的功能展示。这一情况的确切原因尚不清楚,但人们可能会推测,可能会出现某种形式的大肠杆菌折叠机制的过载。xl1-blue显示出为产生的噬菌体颗粒提供最高的荧光强度。
这一试剂的潜在最终用途不限于抗体的选择和筛选。可以在噬菌体上展示的任何蛋白质都可以被插入到荧光噬菌体噬菌粒中以产生单层检测试剂。
尽管荧光噬菌体表现出强荧光并且由于其固有的荧光而将在许多测定中具有有利的用途,从而避免标记的或染色试剂/抗体的使用,且具有通过与poi的第二外壳蛋白融合体而结合至目标分子的能力,但是当结合至抗原时,它仍然不够亮而不能被检测到。实现这一点的可能的方式可能是增加荧光噬菌体颗粒的亮度。从理论上讲,这可以通过以下来实现:1)增加每颗粒的pviii-mneongreen的数量,或2)通过改善折叠性来改善mneongreen的功能展示,或3)增加mneongreen的固有亮度,或全部的组合。
为此,我们已经能够有利地开发出显示改善的功效(改善的荧光性质)的荧光噬菌体构建体的变体。
实例2–荧光噬菌体变体的产生
进行了如本文所述的原始(亲本)mneongreen蛋白的序列(seqidno:3和4)中的氨基酸的随机突变,以评估是否可以产生比现有(亲本)蛋白更亮或显示出在噬菌体外壳中作为pviii融合体的改善的展示的版本。理想地,我们将鉴定在两个类别中均显示出改善的突变体。然而,我们不确定这是否可能。
我们构建了一个文库,所述文库由携带mneongreen基因中的突变的荧光噬菌体载体变体、接头和pviii基因的前90个核苷酸组成。
经历诱变的构建体的部分在下面示出(这三个部分在载体中被视为连续序列,但为清楚起见已在此处单独显示):
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacg
aactgcacatcttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggacgaactgtacaaa
(mneongreen)
ggcggtggcagcggcggtggcagc(接头)
gctgagggtgacgatcccgcaaaagcggcctttaactccc
tgcaagcctcagcgaccgaatatatcggttatgcgtgggcgatggttgttpviii(部分地,前90个核苷酸),(seqidno:56)
文库构建
我们通过pcr进行dntp类似物的并入,以将随机突变引入到噬菌粒dna中。基于zaccolo等人的论文(1996,《分子生物学杂志(jmolbiol)》255(4):589-603),jenabiosciences生产并销售在文库生成期间使用的基于pcr的诱变试剂盒。作为克隆工具,我们使用了
诱变:
我们基于在pcr期间用于并入dntp类似物的轮数/循环数,创建并表征了一个具有不同突变负载的片段的文库。我们选择了这样的片段,即所述片段含有完整的荧光噬菌体pviii融合基因(包括所有5个功能结构域,即torab7、flag、mng、接头和全长pviii)和其侧翼区域(参见下面的序列)。使用这一方法,我们将来有可能返回并扩增不同的区域以构建新的文库,而不必再次经历诱变过程:
下面示出了这一构建体的各个部分(这些部分在载体中被视为连续序列,但为清楚起见已在此处单独显示):
agtgttttagtgtattctttcgcctctttcgttttaggttggtgccttcgtagtggcattacgtattttacccgtttaat
ggaaacttcctcatgataagctagcaagcaaggagacagtcata(侧翼区域)
atgaacaataacgatctctttcagacatcacgtcag
cgttttttggcacaactcggcggcttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgact
(torab7)
gcggcg(氨基酸aa)
gattacaaggatgacgatgacaag(flag)
ggc(氨基酸g)
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacg
aactgcacatcttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggacgaactgtacaaa
(mneongreen)
ggcggtggcagcggcggtggcagc(接头)
gctgagggtgacgatcccgcaaaagcggcctttaactccc
tgcaagcctcagcgaccgaatatatcggttatgcgtgggcgatggttgttgtcattgtcggcgcaactatcggtatcaag
ctgtttaagaaattcacctcgaaagcaagc(全pviii)
tgataa
gctagcttgaggcatcaataaaacgaaaggctcagtcgaaagac
tgggcctttcattttatctgttgtttgtcggttaacgcttgtcgtcatcgtccttgtagtcttttttagcagaatctgcg
gctttcgcatcagcttccggctttgcatcagc(侧翼区域),(seqidno:57)
这一序列用以下引物产生:
pviii_fwd:agtgttttagtgtattctttcgcc(seqidno:58)
fkpa_r:gctgatgcaaagccgg(seqidno:59)
使用gatc(德国)的sanger测序服务对序列进行了分析。进行多达28轮(pcr循环)的突变,这在氨基酸水平上导致约25%的突变频率(在核苷酸水平上约10%),数据未示出。
如图2所示,将诱变的片段克隆回载体,所述载体包含torab7信号肽和flag标签。这一载体的torab7、flag、mng、接头和pviii组分的序列如图9所示。
文库的克隆:
克隆过程是直截了当的,并且我们严格遵循
完全的连接反应被转化为电感受态的ss320,以产生具有高度多样性的文库。文库的基于滴定的计算值为1.0e+06。然后,我们从ss320细胞和电穿孔的xl1-blue细胞(荧光噬菌体的标准包装菌株)制备dna。
按照标准包装方案将文库用高噬菌体包装,并通过facs筛选突变体文库成员。
facsariaiiu的门控策略
完整排序策略的概述在图8中示出。我们基于两个特征对文库进行门控:mneongreen性能(绿色;alexa488)和融合蛋白的展示水平(红色;apc)。(对于第二个特征,可以使用抗dykddddk-apc缀合的抗体,即荧光标记的抗flag标签抗体来定量表面结合的mneongreen-pviii融合蛋白的水平)。第一轮中的门控策略在两个通道的信号强度方面都是宽松的。包装分选的细胞以产生相应的噬菌体文库。然后,将抗
从图8所示的facs图谱可以看出,已经产生了在表面上具有增加的荧光团数量的文库成员和/或具有增加的mneongreen信号的克隆。参见从图8左侧facs图的左下方象限(其示出了分选前的结果)到右下方象限(高水平的表面flag、apc信号)、左上方象限(增加的mneongreen信号)或右上方象限(具有高水平的表面flag、apc信号和增加的mneongreen信号的克隆)的克隆的转移,参见图8的第2轮facs图谱(即在两轮分选之后)。
这样的结果表明,可以获得变体荧光噬菌体,所述变体荧光噬菌体具有与当使用亲本mneongreen序列时所观察到的荧光显著增加和改善的荧光,并且现在可以进一步表征单个克隆以便选择高度有利的突变体。
实例3–突变荧光团的表征
如上面所描述制备的文库可以筛选显示出改善的荧光性质的突变体。通过这些筛选鉴定的改善的克隆之一是如本文所述的所谓的f03突变体。从一种筛选中鉴定这一f03突变体,所述筛选含有在全长pviii中具有突变的文库。当测序时,f03突变体的pviii部分具有核苷酸序列:
gctgagggtgacgatcccgcaaaa
gcggcctttaactccctgcaagcctcagcgaccgaatatatcggttatgcgtgggc
gatg
gttgttgtcattataggcgccactatcggtatcaagctgtttaagaaattcacctcg
aaagcaagc(seqidno:60)
氨基酸序列是:
aegddpakaafnslqasateyigyawamvvviigatigiklfkkftskas
(seqidno:61)
与亲本克隆相比,f03克隆在mneongreen-pviii融合蛋白中仅含有单个氨基酸突变,这是在pviii蛋白的位置33处的v至i突变。对应的密码子在亲本克隆中是gtc,其在f03克隆中是ata。
将含有不同噬菌粒(在上述gb113融合系统的上下文中的mng-pviii野生型/亲本融合蛋白(即mngwt),或具有mng突变的pviii序列的f03克隆,mngf03)的xl1-blue细胞在37℃下在不存在葡萄糖的情况下培养16小时以允许融合蛋白的表达。细胞在pbs中洗涤3次。测量细胞的光密度,并基于光密度将所有样品调节至1的a600值。制备以1:2步骤的系列稀释。
结果在图10中示出。图10a示出了在表达原始荧光噬菌体(mngwt)的活xl-1blue细胞中直接测量的荧光强度与来自筛选的主要候选物(mngf03)的比较。图10b示出了通过监测噬菌体颗粒的荧光强度进行的相同比较,所述噬菌体颗粒已经使用如上面所描述的δ噬菌体按照标准方案包装在菌株中。样品浓度通过感染性滴定来确定,并且所有样品均被归一化为5,0e+12cfuamp/ml。在varioskanlux上以488ex/517em进行荧光测量。
可以看出,f03克隆显示出增加的细菌和噬菌体颗粒荧光,这比用亲本克隆所看到的荧光高大约2倍。
将mngf03-pviii融合体盒克隆到如上面所描述的phox-pix融合系统中以形成双重表达构建体。评估如上面所描述的表达phox-mngwt和phox-mngf03的细菌细胞的荧光强度(图11a),噬菌体颗粒也被评估(图11b)。
在图11a中,在包装过程期间取出细胞。测量来自起始培养物(含有葡萄糖)的荧光,以建立用于荧光测量(glu)的基线。在过夜包装(没有葡萄糖)后,测量相同细胞的荧光。基于光密度将所有样品均归一化至1的a600值。制备以1:2步骤的系列稀释。在图11b中,基于样品中存在的dna量来确定噬菌体浓度。将原始数据调整至最低值4,18e+12。
从图11a和b中可以看出,在phox系统中,mngwt产生良好的荧光,如在细菌细胞(图11a)和噬菌体颗粒(图11b)上测得的。图11a和b显示mngf03还在phox系统中产生荧光。图11b还显示,一旦对滴度进行调节,phox-mngf03噬菌体显示出比mngwt更好的每噬菌体荧光信号。
构建了进一步的双重展示载体,其中将h57抗小鼠tcrscfv与pix融合。
pgald9fn的构建
通过基因合成(gs)生成与pix融合的如huppa等人,2010(《自然》463:963-7)所描述的h57抗小鼠tcrscfv抗体序列,并且在兼容的ncoi/notire位点(huppa等人,2010,见上文;
h57-mngf03的构建
使用侧翼nhei将含有完整的mngf03-pviii盒的盒亚克隆到pgald9δlfn-h57噬菌粒中。通过eurofins的测序来确认正确的方向。
用于tcr表达的逆转录病毒载体的构建
将含有分别移植到鼠cα和cβ上的r12-c9tcr的vα和vβ的基因(straetemans,t.等人,2012)克隆到pmscv-neo(clontech)中,其中α和β通过p2a可切割肽偶联,在ecori和xhoi限制性位点上具有重组的mcs,基本上如(holst,j.等人“t细胞受体逆转录小鼠的产生(generationoft-cellreceptorretrogenicmice)”。《自然协议(nat.protocols)》1,406-417(2006)所描述的。
skw-3r12-c9细胞系的构建
将gp2-293细胞(clontech)与pmscv-neor12-c9和peco(clontech)共转染。将病毒上清液转导入skw-3细胞(德国,cls细胞系服务(clscelllineservices))中。将转导的skw-3r12-c9细胞池与peα-hcd3mab(bd生物科学)和mcβ特异性h57-a647mab(生命科技(lifetech))共染色,并在facsariaiiu(bd)上分离阳性转导子。
流式细胞术
将5×104skw-3r12-c9tcr转染子和未转染的skw-3细胞(阴性对照)的等分试样分配到v形96孔培养皿(康宁(costar))中。用0.5%w/vbsa/pbs(ph7.4)将总体积调节至250μl/孔。将板在室温下以300g/5分钟离心,并弃去上清液。加入在0.5%bsa/pbs中预先封闭的归一化滴度为5×1012cfuampr/ml的25μl/孔的噬菌体的等分试样(对照孔仅接受0.5%bsa/pbs)并将板在室温下孵育30分钟。通过以200μl/孔加入用0.5%bsa/pbs洗涤细胞,通过以300g/5分钟/rt离心沉淀细胞,并且弃去上清液。将在0.5%bsa/pbs中的apc缀合的鸡抗fd(norwegian抗体)添加到适当的孔中,随后在4℃下孵育30分钟。如上洗涤孔两次,并立即在accuric6(bd生物科学)上进行分析。使用flowjo(版本10.5)软件进行数据分析。
通过流式细胞术检测细胞表面表达的蛋白质
荧光噬菌体的强有力的应用将是用于facs的实时选择,不需要染色抗体,而仅依靠荧光团-pviii融合的固有荧光进行检测。为了评估这一点,我们使用流式细胞术测试了展示荧光噬菌体的h57scfv-pix与在人skw-3r12-c9t细胞杂交瘤细胞上表达的t细胞受体的特异性结合。为了比较,我们制备了展示没有荧光团-pviii融合体的噬菌体的h57scfv-pix,并进行了平行测试。
为了使用标准方法验证与表达tcrr12-c9的细胞的特异性结合,我们使用了apc缀合的抗fd抗体来检测结合的噬菌体。这还允许并排比较结合至tcr的荧光噬菌体和常规噬菌体(图12a),和均未结合至tcr阴性人类skw-3的荧光噬菌体和常规噬菌体(图12b)。当试图在不添加检测mab的情况下对细胞染色时,尽管当在溶液中时从荧光噬菌体scfvh57观察到了强荧光信号(h57-mngf03,图12d),但与对照相比,我们并未观察到荧光强度的增加(图12c)。因此,即使荧光噬菌体在溶液中显示出强荧光信号,但是我们也无法使用流式细胞术检测到它与表面结合抗原的结合。这意味着,尽管这一荧光噬菌体在许多测定(例如,如下所述的与facs测定具有许多相似性和组成部分的flisa)中是非常有用和有利的,但这一形式的荧光噬菌体尚未达到允许细胞的直接染色和facs分选所需的每颗粒亮度。
实例4-酶联免疫吸附测定(elisa)和荧光联免疫吸附测定(flisa)
材料和方法
tat-abc表达载体的构建
使用gensmart设计工具(金斯瑞有限公司(genscriptltd.))设计含有tatac/b基因的载体。所有构建体均由金斯瑞有限公司生产和质量控制。tatb序列来源为nc_000913.3:4022218-4022733(ncbi),并且tatac序列来源为#x73888.1(欧洲核苷酸档案(europeannucleotidearchive))。出于检测目的,将tatb与c端6xhis标签融合,并将tatc与c端flag标签融合。
过表达含有phox-mngf03噬菌粒的xl1-blue细胞的tat-ac/b的构建
使用ecm399电穿孔系统的标准规程,用1ng的每种载体转化电感受态的xl1-blue细胞。使xl1-blue细胞在含有30μg/ml氯霉素和50μg/ml博莱霉素(zeocin)的lb琼脂上生长,以选择具有两种表达载体的细胞。使用辣根过氧化物酶缀合的抗flag-m2(1:5000稀释度)和抗his(1:2000稀释度)mab在来自双重阳性克隆的裂解物中通过标准蛋白质印迹分析来确认tatc和tatb的表达。根据它们的表达水平选择3个细胞克隆。用携带含有phox-pix和mneongreen-f03-pviii融合体的噬菌粒的噬菌体转导来自所有三个细胞克隆的10ml。将10μl培养物在含有30μg/ml氯霉素、50μg/ml博莱霉素、500mm葡萄糖和100μg/ml氨苄青霉素的lb琼脂上孵育。基于它们的发荧光的能力测试所得的克隆。
在此实例中,在一些实验中,使用了xl-1宿主细胞,其被工程改造为过表达如上面所描述的tat转运蛋白,以及表达phox-mngf03噬菌粒,以便观察通过过表达tat转运蛋白是否可以改善荧光水平。
一些示例性结果在图14中示出,其中在不存在葡萄糖的情况下过夜培养细菌细胞显示出,与没有tat转运蛋白过表达的细菌细胞相比(xl1-blue;phox-mngf03),当tat转运蛋白被过表达时(xl1-bluetat;phox-mngf03),含有phox-mngf03噬菌粒的细菌细胞的荧光强度大幅增加。
另外,为了评估我们是否可以将在同一噬菌体颗粒上在与pviii不同的衣壳上展示功能性荧光团-pviii融合体和不同功能性融合体的能力结合,我们选择将描述良好的抗phoxscfv(marks等人,1992,《生物技术》,10:779-783)插入到表达噬菌粒的荧光团-pviii中作为pix融合体(这一抗phox噬菌粒也用于上述一些实验)。然后,我们制备了噬菌体,并在如上所述的经典的elisa以及flisa中测试了它们的phox结合。为了比较,我们制备了展示没有荧光团-pviii融合体的噬菌体的抗phoxscfv-pix,并进行了平行测试。
归一化的量的噬菌体颗粒的荧光强度在varioskanlux板读取器中测量(图13d),且显示出非常高的荧光强度,并且仅来自展示荧光团-pviii融合体的噬菌体。同样,当在噬菌体颗粒上进行评估时,f03克隆当与亲代(wt)克隆相比时显示出改善的每噬菌体颗粒荧光。
重要的是,当在标准elisa中测试相同噬菌体样品的scfv靶结合时,观察到特异性结合与荧光团-pviii融合体的存在与否无关(图13c)。因此,数据表明荧光团在pviii上的展示不影响双显示噬菌体上的pix融合体中由靶向部分介导的抗原结合。当使用flisa采用varioskanlux板读取器再次测试相同噬菌体样品的靶结合时,我们可以观察到来自展示mngwt-pviii(图13a,phox-mngwt)和mngf03-pviii融合蛋白(图13b)的噬菌体颗粒的荧光信号,特别是当使用高滴度的过表达tat的xl-1细菌宿主细胞(tat高)时。因此,数据表明这些噬菌体颗粒的固有荧光足够高以提供特异性检测在flisa中洗涤后保留在靶上的减少量的噬菌体的灵敏度,并且发现这些噬菌体颗粒在这样的测定中有利的用途,例如当结合至抗原时能够实现检测。重要的是,数据不可逆转地表明,荧光团-pviii融合体和scfv-pix融合体在同一噬菌体颗粒上的双重展示不影响scfv-pix融合体的表型。
序列表
<110>内克斯特伊拉股份公司
<120>载体构建体
<130>fsp1v200751zx
<150>gb1718294.0
<151>2017-11-03
<160>61
<170>patentin版本3.5
<210>1
<211>114
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>1
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgact114
<210>2
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>2
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>3
<211>705
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>3
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcacatc60
ttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaac120
gacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtgg180
atcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatg240
tctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcag300
ttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatc360
aaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaac420
tctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatc480
atctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgct540
cgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatg600
tacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatgg660
cagaaagctttcaccgacgttatgggtatggacgaactgtacaaa705
<210>4
<211>235
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>4
valserlysglyglugluaspasnmetalaserleuproalathrhis
151015
gluleuhisilepheglyserileasnglyvalasppheaspmetval
202530
glyglnglythrglyasnproasnaspglytyrglugluleuasnleu
354045
lysserthrlysglyaspleuglnpheserprotrpileleuvalpro
505560
hisileglytyrglyphehisglntyrleuprotyrproaspglymet
65707580
serpropheglnalaalametvalaspglyserglytyrglnvalhis
859095
argthrmetglnphegluaspglyalaserleuthrvalasntyrarg
100105110
tyrthrtyrgluglyserhisilelysglyglualaglnvallysgly
115120125
thrglypheproalaaspglyprovalmetthrasnserleuthrala
130135140
alaasptrpcysargserlyslysthrtyrproasnasplysthrile
145150155160
ileserthrphelystrpsertyrthrthrglyasnglylysargtyr
165170175
argserthralaargthrthrtyrthrphealalysprometalaala
180185190
asntyrleulysasnglnpromettyrvalphearglysthrgluleu
195200205
lyshisserlysthrgluleuasnphelysglutrpglnlysalaphe
210215220
thraspvalmetglymetaspgluleutyrlys
225230235
<210>5
<211>24
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>5
ggcggtggcagcggcggtggcagc24
<210>6
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>6
glyglyglyserglyglyglyser
15
<210>7
<211>150
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>7
gctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctcagcgaccgaa60
tatatcggttatgcgtgggcgatggttgttgtcattgtcggcgcaactatcggtatcaag120
ctgtttaagaaattcacctcgaaagcaagc150
<210>8
<211>50
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>8
alagluglyaspaspproalalysalaalapheasnserleuglnala
151015
seralathrglutyrileglytyralatrpalametvalvalvalile
202530
valglyalathrileglyilelysleuphelyslysphethrserlys
354045
alaser
50
<210>9
<211>24
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>9
gattacaaggatgacgatgacaag24
<210>10
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>10
asptyrlysaspaspaspasplys
15
<210>11
<211>1221
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>11
gctgaaactgttgaaagttgtttagcaaaaccccatacagaaaattcatttactaacgtc60
tggaaagacgacaaaactttagatcgttacgctaactatgagggctgtctgtggaatgct120
acaggcgttgtagtttgtactggtgacgaaactcagtgttacggtacatgggttcctatt180
gggcttgctatccctgaaaatgagggtggtggctctgagggtggcggttctgagggtggc240
ggttctgagggtggcggtactaaacctcctgagtacggtgatacacctattccgggctat300
acttatatcaaccctctcgacggcacttatccgcctggtactgagcaaaaccccgctaat360
cctaatccttctcttgaggagtctcagcctcttaatactttcatgtttcagaataatagg420
ttccgaaataggcagggggcattaactgtttatacgggcactgttactcaaggcactgac480
cccgttaaaacttattaccagtacactcctgtatcatcaaaagccatgtatgacgcttac540
tggaacggtaaattcagagactgcgctttccattctggctttaatgaggatccattcgtt600
tgtgaatatcaaggccaatcgtctgacctgcctcaacctcctgtcaatgctggcggcggc660
tctggtggtggttctggtggcggctctgagggtggtggctctgagggtggcggttctgag720
ggtggcggctctgagggaggcggttccggtggtggctctggttccggtgattttgattat780
gaaaagatggcaaacgctaataagggggctatgaccgaaaatgccgatgaaaacgcgcta840
cagtctgacgctaaaggcaaacttgattctgtcgctactgattacggtgctgctatcgat900
ggtttcattggtgacgtttccggccttgctaatggtaatggtgctactggtgattttgct960
ggctctaattcccaaatggctcaagtcggtgacggtgataattcacctttaatgaataat1020
ttccgtcaatatttaccttccctccctcaatcggttgaatgtcgcccttttgtctttggc1080
gctggtaaaccatatgaattttctattgattgtgacaaaataaacttattccgtggtgtc1140
tttgcgtttcttttatatgttgccacctttatgtatgtattttctacgtttgctaacata1200
ctgcgtaataaggagtcttaa1221
<210>12
<211>406
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>12
alagluthrvalglusercysleualalysprohisthrgluasnser
151015
phethrasnvaltrplysaspasplysthrleuaspargtyralaasn
202530
tyrgluglycysleutrpasnalathrglyvalvalvalcysthrgly
354045
aspgluthrglncystyrglythrtrpvalproileglyleualaile
505560
progluasngluglyglyglysergluglyglyglysergluglygly
65707580
glysergluglyglyglythrlysproproglutyrglyaspthrpro
859095
ileproglytyrthrtyrileasnproleuaspglythrtyrpropro
100105110
glythrgluglnasnproalaasnproasnproserleuglugluser
115120125
glnproleuasnthrphemetpheglnasnasnargpheargasnarg
130135140
glnglyalaleuthrvaltyrthrglythrvalthrglnglythrasp
145150155160
provallysthrtyrtyrglntyrthrprovalserserlysalamet
165170175
tyraspalatyrtrpasnglylyspheargaspcysalaphehisser
180185190
glypheasngluaspprophevalcysglutyrglnglyglnserser
195200205
aspleuproglnproprovalasnalaglyglyglyserglyglygly
210215220
serglyglyglysergluglyglyglysergluglyglyglyserglu
225230235240
glyglyglysergluglyglyglyserglyglyglyserglysergly
245250255
asppheasptyrglulysmetalaasnalaasnlysglyalametthr
260265270
gluasnalaaspgluasnalaleuglnseraspalalysglylysleu
275280285
aspservalalathrasptyrglyalaalaileaspglypheilegly
290295300
aspvalserglyleualaasnglyasnglyalathrglyasppheala
305310315320
glyserasnserglnmetalaglnvalglyaspglyaspasnserpro
325330335
leumetasnasnpheargglntyrleuproserleuproglnserval
340345350
glucysargprophevalpheglyalaglylysprotyrglupheser
355360365
ileaspcysasplysileasnleupheargglyvalphealapheleu
370375380
leutyrvalalathrphemettyrvalpheserthrphealaasnile
385390395400
leuargasnlysgluser
405
<210>13
<211>112
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>13
metprovalleuleuglyileproleuleuleuargpheleuglyphe
151015
leuleuvalthrleupheglytyrleuleuthrpheleulyslysgly
202530
pheglylysilealailealaileserleupheleualaleuileile
354045
glyleuasnserileleuvalglytyrleuseraspileseralagln
505560
leuproseraspphevalglnglyvalglnleuileleuproserasn
65707580
alaleuprocysphetyrvalileleuservallysalaalailephe
859095
ilepheaspvallysglnlysilevalsertyrleuasptrpasplys
100105110
<210>14
<211>33
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>14
metgluglnvalalaasppheaspthriletyrglnalametilegln
151015
ileservalvalleucysphealaleuglyileilealaglyglygln
202530
arg
<210>15
<211>32
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>15
metservalleuvaltyrserphealaserphevalleuglytrpcys
151015
leuargserglyilethrtyrphethrargleumetgluthrserser
202530
<210>16
<211>273
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>16
valserlysglyglugluaspasnmetalaserleuproalathrhis
151015
gluleuhisilepheglyserileasnglyvalasppheaspmetval
202530
glyglnglythrglyasnproasnaspglytyrglugluleuasnleu
354045
lysserthrlysglyaspleuglnpheserprotrpileleuvalpro
505560
hisileglytyrglyphehisglntyrleuprotyrproaspglymet
65707580
serpropheglnalaalametvalaspglyserglytyrglnvalhis
859095
argthrmetglnphegluaspglyalaserleuthrvalasntyrarg
100105110
tyrthrtyrgluglyserhisilelysglyglualaglnvallysgly
115120125
thrglypheproalaaspglyprovalmetthrasnserleuthrala
130135140
alaasptrpcysargserlyslysthrtyrproasnasplysthrile
145150155160
ileserthrphelystrpsertyrthrthrglyasnglylysargtyr
165170175
argserthralaargthrthrtyrthrphealalysprometalaala
180185190
asntyrleulysasnglnpromettyrvalphearglysthrgluleu
195200205
lyshisserlysthrgluleuasnphelysglutrpglnlysalaphe
210215220
thraspvalmetglymetaspgluleutyrlysglyglyglysergly
225230235240
glyglyseralagluglyaspaspproalalysalaalapheasnser
245250255
leuglnalaseralathrglutyrileglytyralatrpalametval
260265270
val
<210>17
<211>819
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>17
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcacatc60
ttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaac120
gacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtgg180
atcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatg240
tctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcag300
ttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatc360
aaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaac420
tctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatc480
atctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgct540
cgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatg600
tacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatgg660
cagaaagctttcaccgacgttatgggtatggacgaactgtacaaaggcggtggcagcggc720
ggtggcagcgctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctca780
gcgaccgaatatatcggttatgcgtgggcgatggttgtt819
<210>18
<211>293
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>18
valserlysglyglugluaspasnmetalaserleuproalathrhis
151015
gluleuhisilepheglyserileasnglyvalasppheaspmetval
202530
glyglnglythrglyasnproasnaspglytyrglugluleuasnleu
354045
lysserthrlysglyaspleuglnpheserprotrpileleuvalpro
505560
hisileglytyrglyphehisglntyrleuprotyrproaspglymet
65707580
serpropheglnalaalametvalaspglyserglytyrglnvalhis
859095
argthrmetglnphegluaspglyalaserleuthrvalasntyrarg
100105110
tyrthrtyrgluglyserhisilelysglyglualaglnvallysgly
115120125
thrglypheproalaaspglyprovalmetthrasnserleuthrala
130135140
alaasptrpcysargserlyslysthrtyrproasnasplysthrile
145150155160
ileserthrphelystrpsertyrthrthrglyasnglylysargtyr
165170175
argserthralaargthrthrtyrthrphealalysprometalaala
180185190
asntyrleulysasnglnpromettyrvalphearglysthrgluleu
195200205
lyshisserlysthrgluleuasnphelysglutrpglnlysalaphe
210215220
thraspvalmetglymetaspgluleutyrlysglyglyglysergly
225230235240
glyglyseralagluglyaspaspproalalysalaalapheasnser
245250255
leuglnalaseralathrglutyrileglytyralatrpalametval
260265270
valvalilevalglyalathrileglyilelysleuphelyslysphe
275280285
thrserlysalaser
290
<210>19
<211>879
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>19
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcacatc60
ttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaac120
gacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtgg180
atcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatg240
tctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcag300
ttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatc360
aaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaac420
tctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatc480
atctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgct540
cgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatg600
tacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatgg660
cagaaagctttcaccgacgttatgggtatggacgaactgtacaaaggcggtggcagcggc720
ggtggcagcgctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctca780
gcgaccgaatatatcggttatgcgtgggcgatggttgttgtcattgtcggcgcaactatc840
ggtatcaagctgtttaagaaattcacctcgaaagcaagc879
<210>20
<211>334
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>20
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathralaalaglyvalserlysglyglugluasp
354045
asnmetalaserleuproalathrhisgluleuhisilepheglyser
505560
ileasnglyvalasppheaspmetvalglyglnglythrglyasnpro
65707580
asnaspglytyrglugluleuasnleulysserthrlysglyaspleu
859095
glnpheserprotrpileleuvalprohisileglytyrglyphehis
100105110
glntyrleuprotyrproaspglymetserpropheglnalaalamet
115120125
valaspglyserglytyrglnvalhisargthrmetglnphegluasp
130135140
glyalaserleuthrvalasntyrargtyrthrtyrgluglyserhis
145150155160
ilelysglyglualaglnvallysglythrglypheproalaaspgly
165170175
provalmetthrasnserleuthralaalaasptrpcysargserlys
180185190
lysthrtyrproasnasplysthrileileserthrphelystrpser
195200205
tyrthrthrglyasnglylysargtyrargserthralaargthrthr
210215220
tyrthrphealalysprometalaalaasntyrleulysasnglnpro
225230235240
mettyrvalphearglysthrgluleulyshisserlysthrgluleu
245250255
asnphelysglutrpglnlysalaphethraspvalmetglymetasp
260265270
gluleutyrlysglyglyglyserglyglyglyseralagluglyasp
275280285
aspproalalysalaalapheasnserleuglnalaseralathrglu
290295300
tyrileglytyralatrpalametvalvalvalilevalglyalathr
305310315320
ileglyilelysleuphelyslysphethrserlysalaser
325330
<210>21
<211>1002
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>21
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgcggcg120
ggcgtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcac180
atcttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccg240
aacgacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccg300
tggatcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggt360
atgtctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatg420
cagttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcac480
atcaaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgacc540
aactctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaacc600
atcatctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctacc660
gctcgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccg720
atgtacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaa780
tggcagaaagctttcaccgacgttatgggtatggacgaactgtacaaaggcggtggcagc840
ggcggtggcagcgctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcc900
tcagcgaccgaatatatcggttatgcgtgggcgatggttgttgtcattgtcggcgcaact960
atcggtatcaagctgtttaagaaattcacctcgaaagcaagc1002
<210>22
<211>342
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>22
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathralaalaasptyrlysaspaspaspasplys
354045
glyvalserlysglyglugluaspasnmetalaserleuproalathr
505560
hisgluleuhisilepheglyserileasnglyvalasppheaspmet
65707580
valglyglnglythrglyasnproasnaspglytyrglugluleuasn
859095
leulysserthrlysglyaspleuglnpheserprotrpileleuval
100105110
prohisileglytyrglyphehisglntyrleuprotyrproaspgly
115120125
metserpropheglnalaalametvalaspglyserglytyrglnval
130135140
hisargthrmetglnphegluaspglyalaserleuthrvalasntyr
145150155160
argtyrthrtyrgluglyserhisilelysglyglualaglnvallys
165170175
glythrglypheproalaaspglyprovalmetthrasnserleuthr
180185190
alaalaasptrpcysargserlyslysthrtyrproasnasplysthr
195200205
ileileserthrphelystrpsertyrthrthrglyasnglylysarg
210215220
tyrargserthralaargthrthrtyrthrphealalysprometala
225230235240
alaasntyrleulysasnglnpromettyrvalphearglysthrglu
245250255
leulyshisserlysthrgluleuasnphelysglutrpglnlysala
260265270
phethraspvalmetglymetaspgluleutyrlysglyglyglyser
275280285
glyglyglyseralagluglyaspaspproalalysalaalapheasn
290295300
serleuglnalaseralathrglutyrileglytyralatrpalamet
305310315320
valvalvalilevalglyalathrileglyilelysleuphelyslys
325330335
phethrserlysalaser
340
<210>23
<211>1026
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>23
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgcggcg120
gattacaaggatgacgatgacaagggcgtttctaaaggtgaagaagacaacatggcttct180
ctgccggctacccacgaactgcacatcttcggttctatcaacggtgttgacttcgacatg240
gttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctacc300
aaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccac360
cagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttct420
ggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactac480
cgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttc540
ccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaa600
aaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggt660
aacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggct720
gctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactct780
aaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggac840
gaactgtacaaaggcggtggcagcggcggtggcagcgctgagggtgacgatcccgcaaaa900
gcggcctttaactccctgcaagcctcagcgaccgaatatatcggttatgcgtgggcgatg960
gttgttgtcattgtcggcgcaactatcggtatcaagctgtttaagaaattcacctcgaaa1020
gcaagc1026
<210>24
<211>90
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>24
gctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctcagcgaccgaa60
tatatcggttatgcgtgggcgatggttgtt90
<210>25
<211>30
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>25
alagluglyaspaspproalalysalaalapheasnserleuglnala
151015
seralathrglutyrileglytyralatrpalametvalval
202530
<210>26
<211>314
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>26
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathralaalaglyvalserlysglyglugluasp
354045
asnmetalaserleuproalathrhisgluleuhisilepheglyser
505560
ileasnglyvalasppheaspmetvalglyglnglythrglyasnpro
65707580
asnaspglytyrglugluleuasnleulysserthrlysglyaspleu
859095
glnpheserprotrpileleuvalprohisileglytyrglyphehis
100105110
glntyrleuprotyrproaspglymetserpropheglnalaalamet
115120125
valaspglyserglytyrglnvalhisargthrmetglnphegluasp
130135140
glyalaserleuthrvalasntyrargtyrthrtyrgluglyserhis
145150155160
ilelysglyglualaglnvallysglythrglypheproalaaspgly
165170175
provalmetthrasnserleuthralaalaasptrpcysargserlys
180185190
lysthrtyrproasnasplysthrileileserthrphelystrpser
195200205
tyrthrthrglyasnglylysargtyrargserthralaargthrthr
210215220
tyrthrphealalysprometalaalaasntyrleulysasnglnpro
225230235240
mettyrvalphearglysthrgluleulyshisserlysthrgluleu
245250255
asnphelysglutrpglnlysalaphethraspvalmetglymetasp
260265270
gluleutyrlysglyglyglyserglyglyglyseralagluglyasp
275280285
aspproalalysalaalapheasnserleuglnalaseralathrglu
290295300
tyrileglytyralatrpalametvalval
305310
<210>27
<211>942
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>27
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgcggcg120
ggcgtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcac180
atcttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccg240
aacgacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccg300
tggatcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggt360
atgtctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatg420
cagttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcac480
atcaaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgacc540
aactctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaacc600
atcatctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctacc660
gctcgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccg720
atgtacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaa780
tggcagaaagctttcaccgacgttatgggtatggacgaactgtacaaaggcggtggcagc840
ggcggtggcagcgctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcc900
tcagcgaccgaatatatcggttatgcgtgggcgatggttgtt942
<210>28
<211>322
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>28
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathralaalaasptyrlysaspaspaspasplys
354045
glyvalserlysglyglugluaspasnmetalaserleuproalathr
505560
hisgluleuhisilepheglyserileasnglyvalasppheaspmet
65707580
valglyglnglythrglyasnproasnaspglytyrglugluleuasn
859095
leulysserthrlysglyaspleuglnpheserprotrpileleuval
100105110
prohisileglytyrglyphehisglntyrleuprotyrproaspgly
115120125
metserpropheglnalaalametvalaspglyserglytyrglnval
130135140
hisargthrmetglnphegluaspglyalaserleuthrvalasntyr
145150155160
argtyrthrtyrgluglyserhisilelysglyglualaglnvallys
165170175
glythrglypheproalaaspglyprovalmetthrasnserleuthr
180185190
alaalaasptrpcysargserlyslysthrtyrproasnasplysthr
195200205
ileileserthrphelystrpsertyrthrthrglyasnglylysarg
210215220
tyrargserthralaargthrthrtyrthrphealalysprometala
225230235240
alaasntyrleulysasnglnpromettyrvalphearglysthrglu
245250255
leulyshisserlysthrgluleuasnphelysglutrpglnlysala
260265270
phethraspvalmetglymetaspgluleutyrlysglyglyglyser
275280285
glyglyglyseralagluglyaspaspproalalysalaalapheasn
290295300
serleuglnalaseralathrglutyrileglytyralatrpalamet
305310315320
valval
<210>29
<211>966
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>29
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgcggcg120
gattacaaggatgacgatgacaagggcgtttctaaaggtgaagaagacaacatggcttct180
ctgccggctacccacgaactgcacatcttcggttctatcaacggtgttgacttcgacatg240
gttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctacc300
aaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccac360
cagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttct420
ggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactac480
cgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttc540
ccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaa600
aaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggt660
aacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggct720
gctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactct780
aaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggac840
gaactgtacaaaggcggtggcagcggcggtggcagcgctgagggtgacgatcccgcaaaa900
gcggcctttaactccctgcaagcctcagcgaccgaatatatcggttatgcgtgggcgatg960
gttgtt966
<210>30
<211>6
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>30
aggaga6
<210>31
<211>25
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>31
ttaactttaagaaggagatatacat25
<210>32
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>32
metasnasnasnaspleupheglnalaserargargargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>33
<211>7
<212>prt
<213>人工序列
<220>
<223>人工序列
<220>
<221>misc_feature
<222>(1)..(1)
<223>ser或thr
<220>
<221>misc_feature
<222>(4)..(4)
<223>任何氨基酸
<400>33
xaaargargxaapheleulys
15
<210>34
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>34
thrserargglnargpheleuala
15
<210>35
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>35
metasnasnasnaspleupheglnthrserargargargleuleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>36
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>36
thrserargargargleuleuala
15
<210>37
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>37
metasnasnasnaspilepheglnalaserargargargpheleuala
151015
glnproglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>38
<211>13
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>38
ilepheglnalaserargargargpheleualaglnpro
1510
<210>39
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>39
metasnasnasngluleupheglnalaserargargargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>40
<211>12
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>40
gluleupheglnalaserargargargpheleuala
1510
<210>41
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>41
metasnasnasnaspleupheglnthrthrargargargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>42
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>42
thrthrargargargpheleuala
15
<210>43
<211>38
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>43
metasnasnasnaspserpheglnthrserargargargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathr
35
<210>44
<211>11
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>44
serpheglnthrserargargargpheleuala
1510
<210>45
<211>711
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>45
atggtgagcaagggcgaggaggataacatggcctctctcccagcgacacatgagttacac60
atctttggctccatcaacggtgtggactttgacatggtgggtcagggcaccggcaatcca120
aatgatggttatgaggagttaaacctgaagtccaccaagggtgacctccagttctccccc180
tggattctggtccctcatatcgggtatggcttccatcagtacctgccctaccctgacggg240
atgtcgcctttccaggccgccatggtagatggctccggataccaagtccatcgcacaatg300
cagtttgaagatggtgcctcccttactgttaactaccgctacacctacgagggaagccac360
atcaaaggagaggcccaggtgaaggggactggtttccctgctgacggtcctgtgatgacc420
aactcgctgaccgctgcggactggtgcaggtcgaagaagacttaccccaacgacaaaacc480
atcatcagtacctttaagtggagttacaccactggaaatggcaagcgctaccggagcact540
gcgcggaccacctacacctttgccaagccaatggcggctaactatctgaagaaccagccg600
atgtacgtgttccgtaagacggagctcaagcactccaagaccgagctcaacttcaaggag660
tggcaaaaggcctttaccgatgtgatgggcatggacgagctgtacaagtaa711
<210>46
<211>236
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>46
metvalserlysglyglugluaspasnmetalaserleuproalathr
151015
hisgluleuhisilepheglyserileasnglyvalasppheaspmet
202530
valglyglnglythrglyasnproasnaspglytyrglugluleuasn
354045
leulysserthrlysglyaspleuglnpheserprotrpileleuval
505560
prohisileglytyrglyphehisglntyrleuprotyrproaspgly
65707580
metserpropheglnalaalametvalaspglyserglytyrglnval
859095
hisargthrmetglnphegluaspglyalaserleuthrvalasntyr
100105110
argtyrthrtyrgluglyserhisilelysglyglualaglnvallys
115120125
glythrglypheproalaaspglyprovalmetthrasnserleuthr
130135140
alaalaasptrpcysargserlyslysthrtyrproasnasplysthr
145150155160
ileileserthrphelystrpsertyrthrthrglyasnglylysarg
165170175
tyrargserthralaargthrthrtyrthrphealalysprometala
180185190
alaasntyrleulysasnglnpromettyrvalphearglysthrglu
195200205
leulyshisserlysthrgluleuasnphelysglutrpglnlysala
210215220
phethraspvalmetglymetaspgluleutyrlys
225230235
<210>47
<211>4
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>47
gagg4
<210>48
<211>6
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>48
aggagg6
<210>49
<211>7
<212>rna
<213>人工序列
<220>
<223>人工序列
<400>49
aggaggu7
<210>50
<211>9
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>50
ttaacttta9
<210>51
<211>15
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>51
aaggagacagtcata15
<210>52
<211>17
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>52
metserglyleuasnaspilepheglualaglnlysileglutrphis
151015
glu
<210>53
<211>8
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>53
trpserhisproglnpheglulys
15
<210>54
<211>1032
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>54
atgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcggcggc60
ttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgcggcg120
gattacaaggatgacgatgacaagggcgtttctaaaggtgaagaagacaacatggcttct180
ctgccggctacccacgaactgcacatcttcggttctatcaacggtgttgacttcgacatg240
gttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatctacc300
aaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggtttccac360
cagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacggttct420
ggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaactac480
cgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccggtttc540
ccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttctaaa600
aaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccaccggt660
aacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgatggct720
gctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaacactct780
aaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtatggac840
gaactgtacaaaggcggtggcagcggcggtggcagcgctgagggtgacgatcccgcaaaa900
gcggcctttaactccctgcaagcctcagcgaccgaatatatcggttatgcgtgggcgatg960
gttgttgtcattgtcggcgcaactatcggtatcaagctgtttaagaaattcacctcgaaa1020
gcaagctgataa1032
<210>55
<211>342
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>55
metasnasnasnaspleupheglnthrserargglnargpheleuala
151015
glnleuglyglyleuthrvalalaglymetleuglyproserleuleu
202530
thrproargargalathralaalaasptyrlysaspaspaspasplys
354045
glyvalserlysglyglugluaspasnmetalaserleuproalathr
505560
hisgluleuhisilepheglyserileasnglyvalasppheaspmet
65707580
valglyglnglythrglyasnproasnaspglytyrglugluleuasn
859095
leulysserthrlysglyaspleuglnpheserprotrpileleuval
100105110
prohisileglytyrglyphehisglntyrleuprotyrproaspgly
115120125
metserpropheglnalaalametvalaspglyserglytyrglnval
130135140
hisargthrmetglnphegluaspglyalaserleuthrvalasntyr
145150155160
argtyrthrtyrgluglyserhisilelysglyglualaglnvallys
165170175
glythrglypheproalaaspglyprovalmetthrasnserleuthr
180185190
alaalaasptrpcysargserlyslysthrtyrproasnasplysthr
195200205
ileileserthrphelystrpsertyrthrthrglyasnglylysarg
210215220
tyrargserthralaargthrthrtyrthrphealalysprometala
225230235240
alaasntyrleulysasnglnpromettyrvalphearglysthrglu
245250255
leulyshisserlysthrgluleuasnphelysglutrpglnlysala
260265270
phethraspvalmetglymetaspgluleutyrlysglyglyglyser
275280285
glyglyglyseralagluglyaspaspproalalysalaalapheasn
290295300
serleuglnalaseralathrglutyrileglytyralatrpalamet
305310315320
valvalvalilevalglyalathrileglyilelysleuphelyslys
325330335
phethrserlysalaser
340
<210>56
<211>819
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>56
gtttctaaaggtgaagaagacaacatggcttctctgccggctacccacgaactgcacatc60
ttcggttctatcaacggtgttgacttcgacatggttggtcagggtaccggtaacccgaac120
gacggttacgaagaactgaacctgaaatctaccaaaggtgacctgcagttctctccgtgg180
atcttagttccgcacatcggttacggtttccaccagtacctgccgtacccggacggtatg240
tctccgttccaggctgctatggttgacggttctggttaccaggttcaccgtaccatgcag300
ttcgaagacggtgcttctctgaccgttaactaccgttacacctacgaaggttctcacatc360
aaaggtgaagctcaggttaaaggtaccggtttcccggctgacggtccggttatgaccaac420
tctctgaccgctgctgactggtgccgttctaaaaaaacctacccgaacgacaaaaccatc480
atctctaccttcaaatggtcttacaccaccggtaacggtaaacgttaccgttctaccgct540
cgtaccacctacaccttcgctaaaccgatggctgctaactacctgaaaaaccagccgatg600
tacgttttccgtaaaaccgaactgaaacactctaaaaccgaactgaacttcaaagaatgg660
cagaaagctttcaccgacgttatgggtatggacgaactgtacaaaggcggtggcagcggc720
ggtggcagcgctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctca780
gcgaccgaatatatcggttatgcgtgggcgatggttgtt819
<210>57
<211>1312
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>57
agtgttttagtgtattctttcgcctctttcgttttaggttggtgccttcgtagtggcatt60
acgtattttacccgtttaatggaaacttcctcatgataagctagcaagcaaggagacagt120
cataatgaacaataacgatctctttcagacatcacgtcagcgttttttggcacaactcgg180
cggcttaaccgtcgccgggatgctggggccgtcattgttaacgccgcgacgtgcgactgc240
ggcggattacaaggatgacgatgacaagggcgtttctaaaggtgaagaagacaacatggc300
ttctctgccggctacccacgaactgcacatcttcggttctatcaacggtgttgacttcga360
catggttggtcagggtaccggtaacccgaacgacggttacgaagaactgaacctgaaatc420
taccaaaggtgacctgcagttctctccgtggatcttagttccgcacatcggttacggttt480
ccaccagtacctgccgtacccggacggtatgtctccgttccaggctgctatggttgacgg540
ttctggttaccaggttcaccgtaccatgcagttcgaagacggtgcttctctgaccgttaa600
ctaccgttacacctacgaaggttctcacatcaaaggtgaagctcaggttaaaggtaccgg660
tttcccggctgacggtccggttatgaccaactctctgaccgctgctgactggtgccgttc720
taaaaaaacctacccgaacgacaaaaccatcatctctaccttcaaatggtcttacaccac780
cggtaacggtaaacgttaccgttctaccgctcgtaccacctacaccttcgctaaaccgat840
ggctgctaactacctgaaaaaccagccgatgtacgttttccgtaaaaccgaactgaaaca900
ctctaaaaccgaactgaacttcaaagaatggcagaaagctttcaccgacgttatgggtat960
ggacgaactgtacaaaggcggtggcagcggcggtggcagcgctgagggtgacgatcccgc1020
aaaagcggcctttaactccctgcaagcctcagcgaccgaatatatcggttatgcgtgggc1080
gatggttgttgtcattgtcggcgcaactatcggtatcaagctgtttaagaaattcacctc1140
gaaagcaagctgataagctagcttgaggcatcaataaaacgaaaggctcagtcgaaagac1200
tgggcctttcattttatctgttgtttgtcggttaacgcttgtcgtcatcgtccttgtagt1260
cttttttagcagaatctgcggctttcgcatcagcttccggctttgcatcagc1312
<210>58
<211>24
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>58
agtgttttagtgtattctttcgcc24
<210>59
<211>16
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>59
gctgatgcaaagccgg16
<210>60
<211>150
<212>dna
<213>人工序列
<220>
<223>人工序列
<400>60
gctgagggtgacgatcccgcaaaagcggcctttaactccctgcaagcctcagcgaccgaa60
tatatcggttatgcgtgggcgatggttgttgtcattataggcgccactatcggtatcaag120
ctgtttaagaaattcacctcgaaagcaagc150
<210>61
<211>50
<212>prt
<213>人工序列
<220>
<223>人工序列
<400>61
alagluglyaspaspproalalysalaalapheasnserleuglnala
151015
seralathrglutyrileglytyralatrpalametvalvalvalile
202530
ileglyalathrileglyilelysleuphelyslysphethrserlys
354045
alaser
50
- 还没有人留言评论。精彩留言会获得点赞!