一种提高水稻远缘杂交后代育性的方法与所用蛋白质与流程
本发明涉及生物技术领域中,一种提高水稻远缘杂交后代育性的方法与所用蛋白质。
背景技术:
水稻是世界上最重要的粮食作物之一,占世界50%比例的人口以稻米为主食。提高水稻单产是保障世界粮食安全的有力措施之一。水稻品种间狭窄的遗传多样性已成为制约水稻产量潜力进一步提高的瓶颈。野生稻是一个重要的有利基因库,携带抗病虫、抗逆、高产优质基因,通过远缘杂交利用野生稻的有利基因,是提升水稻育种水平的有效途径。然而,由于野生稻与栽培稻杂种及杂种后代存在不育或半不育现象,限制了野生稻优良基因的有效利用。
目前,有关水稻育性研究多集中于雄性不育,雌性不育研究较少,只有少数雌性不育相关基因通过图位克隆方法被鉴定出来,如:发生在亚种间杂种雌性不育基因s5、s7、hsa1,非洲栽培稻和亚洲栽培稻种间杂种雌性不育基因s1及通过突变体鉴定出的ptb基因。通过普通野生稻与亚洲栽培稻构建的渗入系鉴定出杂种弱势基因hwi1和hwi2,但还未见从普通野生稻和亚洲栽培稻杂种后代中分离出雌性不育基因的报导。
云南元江普通野生稻属于aa基因组。是我国迄今发现的海拔最高(780米)且隔离较好的普通野生稻,因气候生态环境独特,使它在中国栽培稻的起源演化研究上具有重要的地位。元江野生稻与籼稻杂种f1的育性为54.53%,说明元江野生稻与亚洲栽培稻之间存在杂种不育。
技术实现要素:
本发明所要解决的技术问题是如何提高水稻远缘杂交后代的育性。
为解决上述技术问题本发明首先提供了一种来源于元江野生稻的蛋白质(名称为esa1),esa1为如下a1)、a2)或a3):
a1)氨基酸序列是序列1的蛋白质;
a2)将序列表中序列1所示的氨基酸序列经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有相同功能的蛋白质;
a3)在a1)或a2)的n端或/和c端连接标签得到的融合蛋白质。
为了使a1)中的蛋白质便于纯化,可在由序列表中序列1所示的氨基酸序列组成的蛋白质的氨基末端或羧基末端连接上如下表所示的标签。
表:标签的序列
上述a2)中的esa1蛋白质,为与序列1所示蛋白质的氨基酸序列具有75%或75%以上同一性且具有相同功能的蛋白质。所述具有75%或75%以上同一性为具有75%、具有80%、具有85%、具有90%、具有95%、具有96%、具有97%、具有98%或具有99%的同一性。
上述a2)中的esa1蛋白质可人工合成,也可先合成其编码基因,再进行生物表达得到。
上述a2)中的esa1蛋白质的编码基因可通过将序列3的第83-1903位所示的dna序列中缺失一个或几个氨基酸残基的密码子,和/或进行一个或几个碱基对的错义突变,和/或在其5′端和/或3′端连上上表所示的标签的编码序列得到。其中,序列3的第83-1903位所示的dna分子编码序列1所示的esa1蛋白质。
本发明还提供了与esa1相关的生物材料,所述生物材料为下述b1)至b9)中的任一种:
b1)编码esa1的核酸分子;
b2)含有b1)所述核酸分子的表达盒;
b3)含有b1)所述核酸分子的重组载体、或含有b2)所述表达盒的重组载体;
b4)含有b1)所述核酸分子的重组微生物、或含有b2)所述表达盒的重组微生物、或含有b3)所述重组载体的重组微生物;
b5)含有b1)所述核酸分子的转基因植物细胞系、或含有b2)所述表达盒的转基因植物细胞系;
b6)含有b1)所述核酸分子的转基因植物组织、或含有b2)所述表达盒的转基因植物组织;
b7)含有b1)所述核酸分子的转基因植物器官、或含有b2)所述表达盒的转基因植物器官;
b8)抑制esa1编码基因表达的核酸分子;
b9)含有b8)所述核酸分子的表达盒、重组载体、重组微生物、转基因植物细胞系、转基因植物组织或转基因植物器官。
上述生物材料中,b1)所述核酸分子可为如下b11)或b12)或b13)或b14)或b15):
b11)编码序列是序列表中序列3的第83-1903位的cdna分子或dna分子;
b12)序列表中序列3所示的dna分子;
b13)序列表中序列2所示的dna分子;
b14)与b11)或b12)或b13)限定的核苷酸序列具有75%或75%以上同一性,且编码esa1的cdna分子或dna分子;
b15)在严格条件下与b11)或b12)或b13)或b14)限定的核苷酸序列杂交,且编码esa1的cdna分子或dna分子;
b8)所述核酸分子为序列表中序列2的第849-1152位和/或序列2的第849-1284位所示的dna分子。
其中,所述核酸分子可以是dna,如cdna、基因组dna或重组dna;所述核酸分子也可以是rna,如mrna或hnrna等。
本领域普通技术人员可以很容易地采用已知的方法,例如定向进化和点突变的方法,对本发明的编码esa1蛋白质的核苷酸序列进行突变。那些经过人工修饰的,具有与本发明分离得到的esa1蛋白质的核苷酸序列75%或者更高同一性的核苷酸,只要编码esa1蛋白质且具有esa1蛋白质功能,均是衍生于本发明的核苷酸序列并且等同于本发明的序列。
这里使用的术语“同一性”指与天然核酸序列的序列相似性。“同一性”包括与本发明的编码序列1所示的氨基酸序列组成的蛋白质的核苷酸序列具有75%或更高,或85%或更高,或90%或更高,或95%或更高同一性的核苷酸序列。同一性可以用肉眼或计算机软件进行评价。使用计算机软件,两个或多个序列之间的同一性可以用百分比(%)表示,其可以用来评价相关序列之间的同一性。
所述严格条件可为如下:50℃,在7%十二烷基硫酸钠(sds)、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,2×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.5×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在50℃,0.1×ssc,0.1%sds中漂洗;还可为:50℃,在7%sds、0.5mnapo4和1mmedta的混合溶液中杂交,在65℃,0.1×ssc,0.1%sds中漂洗;也可为:在6×ssc,0.5%sds的溶液中,在65℃下杂交,然后用2×ssc,0.1%sds和1×ssc,0.1%sds各洗膜一次;也可为:2×ssc,0.1%sds的溶液中,在68℃下杂交并洗膜2次,每次5min,又于0.5×ssc,0.1%sds的溶液中,在68℃下杂交并洗膜2次,每次15min;也可为:0.1×sspe(或0.1×ssc)、0.1%sds的溶液中,65℃条件下杂交并洗膜。
上述75%或75%以上同一性,可为80%、85%、90%或95%以上的同一性。
b2)所述的含有编码esa1蛋白质的核酸分子的表达盒(esa1基因表达盒),是指能够在宿主细胞中表达esa1蛋白质的dna,该dna不但可包括启动esa1基因转录的启动子,还可包括终止esa1基因转录的终止子。进一步,所述表达盒还可包括增强子序列。可用于本发明的启动子包括但不限于:组成型启动子,组织、器官和发育特异的启动子,和诱导型启动子。启动子的例子包括但不限于:花椰菜花叶病毒的组成型启动子35s;来自西红柿的创伤诱导型启动子,亮氨酸氨基肽酶("lap",chao等人(1999)plantphysiol120:979-992);来自烟草的化学诱导型启动子,发病机理相关1(pr1)(由水杨酸和bth(苯并噻二唑-7-硫代羟酸s-甲酯)诱导);西红柿蛋白酶抑制剂ii启动子(pin2)或lap启动子(均可用茉莉酮酸甲酯诱导);热休克启动子(美国专利5,187,267);四环素诱导型启动子(美国专利5,057,422);种子特异性启动子,如谷子种子特异性启动子pf128(cn101063139b(中国专利200710099169.7)),种子贮存蛋白质特异的启动子(例如,菜豆球蛋白、napin,oleosin和大豆betaconglycin的启动子(beachy等人(1985)emboj.4:3047-3053))。它们可单独使用或与其它的植物启动子结合使用。此处引用的所有参考文献均全文引用。合适的转录终止子包括但不限于:农杆菌胭脂碱合成酶终止子(nos终止子)、花椰菜花叶病毒camv35s终止子、tml终止子、豌豆rbcse9终止子和胭脂氨酸和章鱼氨酸合酶终止子(参见,例如:odell等人(i985)nature313:810;rosenberg等人(1987)gene,56:125;guerineau等人(1991)mol.gen.genet,262:141;proudfoot(1991)cell,64:671;sanfacon等人genesdev.,5:141;mogen等人(1990)plantcell,2:1261;munroe等人(1990)gene,91:151;ballad等人(1989)nucleicacidsres.17:7891;joshi等人(1987)nucleicacidres.,15:9627)。
可用现有的表达载体构建含有所述esa1基因表达盒的重组载体。所述植物表达载体包括双元农杆菌载体和可用于植物微弹轰击的载体等。如pahc25、pbin438、pcambia1302、pcambia2301、pcambia1301、pcambia1300、pbi121、pcambia1391-xa、psn1301或pcambia1391-xb(cambia公司)等。所述植物表达载体还可包含外源基因的3′端非翻译区域,即包含聚腺苷酸信号和任何其它参与mrna加工或基因表达的dna片段。所述聚腺苷酸信号可引导聚腺苷酸加入到mrna前体的3′端,如农杆菌冠瘿瘤诱导(ti)质粒基因(如胭脂碱合成酶基因nos)、植物基因(如大豆贮存蛋白基因)3′端转录的非翻译区均具有类似功能。使用本发明的基因构建植物表达载体时,还可使用增强子,包括翻译增强子或转录增强子,这些增强子区域可以是atg起始密码子或邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的正确翻译。所述翻译控制信号和起始密码子的来源是广泛的,可以是天然的,也可以是合成的。翻译起始区域可以来自转录起始区域或结构基因。为了便于对转基因植物细胞或植物进行鉴定及筛选,可对所用植物表达载体进行加工,如加入可在植物中表达的编码可产生颜色变化的酶或发光化合物的基因(gus基因、萤光素酶基因等)、抗生素的标记基因(如赋予对卡那霉素和相关抗生素抗性的nptii基因,赋予对除草剂膦丝菌素抗性的bar基因,赋予对抗生素潮霉素抗性的hph基因,和赋予对氨甲喋呤抗性的dhfr基因,赋予对草甘磷抗性的epsps基因)或是抗化学试剂标记基因等(如抗除莠剂基因)、提供代谢甘露糖能力的甘露糖-6-磷酸异构酶基因。从转基因植物的安全性考虑,可不加任何选择性标记基因,直接以逆境筛选转化植株。
所述载体可为质粒、黏粒、噬菌体或病毒载体。所述质粒具体可为ptck303。
b9)所述重组载体可为rnai-esa1,所述rnai-esa1为将ptck303的spei和saci识别序列间的dna片段替换为序列表中序列2的第849-1152位,并将bamhi和kpni识别序列间的dna片段替换为序列表中序列2的第849-1284位的反向互补序列得到的重组载体。
所述微生物可为酵母、细菌、藻或真菌。其中,细菌可为农杆菌,如根癌农杆菌eha105。
b9)所述重组微生物可为eha105-rnai-esa1,所述eha105-rnai-esa1为将所述rnai-esa1导入根癌农杆菌eha105中得到的重组菌。
所述转基因植物细胞系、转基因植物组织和转基因植物器官均不包括繁殖材料。
本发明还提供了提高水稻远缘杂交后代育性的方法,所述方法包括:降低受体水稻中esa1的含量,或抑制受体水稻中esa1的活性,得到与所述受体水稻相比育性增强的目的水稻,实现水稻育性的增强;所述受体水稻为水稻远缘杂交后代。
所述受体水稻含有esa1的编码基因。
所述降低受体水稻中esa1的含量可通过抑制所述受体水稻中esa1的编码基因的表达实现。
所述抑制所述受体水稻中esa1的编码基因的表达可通过向所述受体水稻中导入含有序列表中序列2的第849-1152位和序列2的第849-1284位的反向互补序列的dna片段(记为dna片段1)实现。
所述目的水稻中所述dna片段1能转录抑制esa1的编码基因表达的rna。
所述抑制所述受体水稻中esa1的编码基因的表达可通过向利用所述rnai-esa1转化所述受体水稻并筛选含有所述dna片段1的阳性转基因植株实现。所述阳性转基因植株中esa1的编码基因的表达降低。
所述目的水稻不仅包含esa1的编码基因降低的第一代植物,也包括其子代。所述目的水稻包括种子、愈伤组织、完整植株和细胞。
上述方法中,所述育性可为雌性生殖器官育性。
所述育性具体可体现在结实率上。
上述方法中,所述受体水稻可为野生稻与亚洲栽培稻杂交后代。
所述野生稻具体可为元江野生稻。所述亚洲栽培稻可为籼稻。所述籼稻进一步可为特青。
本发明还提供了培育杂交水稻的方法,所述方法包括:按照所述提高水稻远缘杂交后代育性的方法提高水稻远缘杂交后代育性,获得育性提高的水稻远缘杂交后代,将所述水稻远缘杂交后代作为亲本与其他水稻进行杂交,实现杂交水稻的培育。
所述其他水稻非所述水稻远缘杂交后代。
esa1或所述生物材料在调控水稻远缘杂交后代育性中的应用,也属于本发明的保护范围。
实验证明,抑制本发明中eas1编码基因的表达后,可提高水稻远缘杂交后代的结实率,提高水稻远缘杂交后代的育性,不仅有利于揭示雌性不育机理,同时对克服野生稻与栽培稻远缘杂交障碍,有效利用野生稻优良基因改良栽培稻具有重要意义。
附图说明
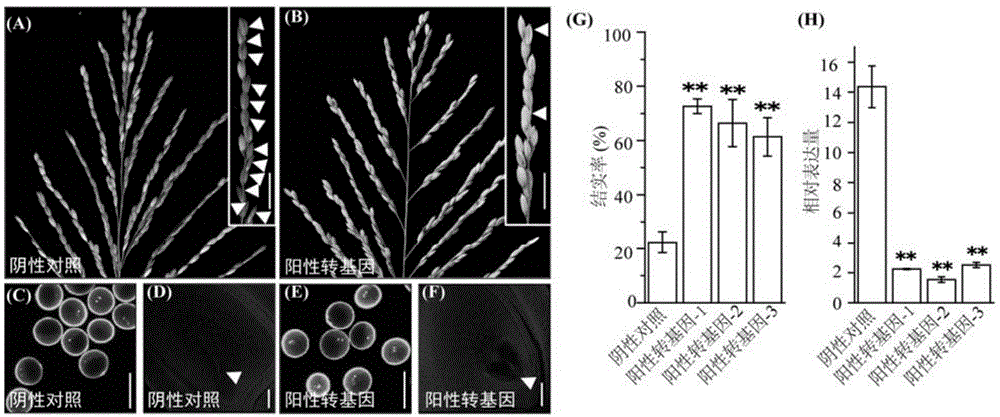
图1为esa1-rnai转基因植株表型及esa1基因相对表达量检测。a与b分别为阴性植株和阳性rnai-esa1转基因植株(图中阳性转基因、阳性转基因-1、阳性转基因-2和阳性转基因-3)的主茎穗结实图,右上角为枝梗结实情况放大图,白色箭头所指为空瘪籽粒。c与e分别为dapi染色阴性植株和阳性esa1-rnai转基因植株成熟花粉。d与f分别为阴性植株和阳性rnai-esa1转基因植株成熟胚囊,白色箭头所指为成熟胚囊。g为阴性植株和阳性esa1-rnai转基因植株的结实率比较。h为esa1在阴性植株和阳性esa1-rnai转基因植物中的相对表达水平。
图2为esa1基因在nil-qssr1植株中的相对表达量。p1-p20分别为nil-qssr1植株1-20cm穗长所对应的相对表达量。
具体实施方式
下面结合具体实施方式对本发明进行进一步的详细描述,给出的实施例仅为了阐明本发明,而不是为了限制本发明的范围。下述实施例中的实验方法,如无特殊说明,均为常规方法。下述实施例中所用的材料、试剂、仪器等,如无特殊说明,均可从商业途径得到。以下实施例中的定量试验,均设置三次重复实验,结果取平均值。下述实施例中,如无特殊说明,序列表中各核苷酸序列的第1位均为相应dna的5′末端核苷酸,末位均为相应dna的3′末端核苷酸。
下述实施例中的元江野生稻(yuanjiangcommonwildrice)和特青(teqing)均记载在文献(tanetal.,controlofakeytransitionfromprostratetoerectgrowthinricedomestication,naturegenetics,volume40,number11,november2008)中,公众可从申请人处获得,这些生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。
下述实施例中的ptck303(wangetal.,2004,apracticalvectorforefficientknockdownofgeneexpressioninrice(oryzasatival.),plantmolecularbiologyreporter22:409-417),公众可从申请人处获得,这些生物材料只为重复本发明的相关实验所用,不可作为其它用途使用。
实施例1、降低esa1编码基因的表达可以提高野生稻与栽培稻杂交后代的雌配子育性
本实施例发现了一个来源于元江野生稻的蛋白质,其名称为esa1,降低元江野生稻与特青杂交后代中该蛋白质编码基因的表达可以提高杂交后代的雌配子育性。esa1的氨基酸序列为序列表中序列1,在元江野生稻中,esa1基因的基因组序列为序列表中序列2,cds序列为序列表中序列3,序列3的第83-1903位编码序列1所示的esa1。
一、半不育渗入系yil42的制备
利用元江野生稻为供体亲本,籼型栽培稻特青为轮回亲本通过杂交、回交4代构建了一套含有127个系的渗入系群体,从该群体中选出一个结实率低的株系记为半不育渗入系yil42,yil42的结实率为20.67%,元江野生稻结实率为80.30%,特青结实率为87.73%。结实率=实粒数/总粒数×100%。
二、转基因植株的构建
1、降低esa1基因表达量的rnai载体的构建
dna片段的扩增:
以元江野生稻的cdna为模板,利用引物1和引物2组成的引物对进行pcr扩增,将得到的dna片段记为dna片段1;以元江野生稻的cdna为模板,利用引物3和引物4组成的引物对进行pcr扩增,将得到的dna片段记为dna片段2。所用引物序列如下:
引物1:5′-ggactagttcgtgtcatcgttgatcgtt-3′(下划线处为spei的识别序列);
引物2:5′-cgagctcagcactctcccagaatcacc-3′(下划线处为saci的识别序列);
引物3:5′-cgggatccccggtctcatccaatgctaa-3′(下划线处为bamhi的识别序列);
引物4:5′-ggggtacctcgtgtcatcgttgatcgtt-3′(下划线处为kpni的识别序列)。
将dna片段1利用spei和saci进行双酶切,将得到的dna片段与ptck303经过spei和saci双酶切得到的载体骨架相连,得到中间载体;将dna片段2利用bamhi和kpni进行双酶切,将得到的的dna片段与中间载体经bamhi和kpni双酶切得到的载体骨架相连,得到重组载体,序列正确的重组载体即为降低esa1基因表达量的rnai载体,记为rnai-esa1。rnai-esa1为将ptck303的spei和saci识别序列间的dna片段替换为序列表中序列2的第849-1152位,并将bamhi和kpni识别序列间的dna片段替换为序列表中序列2的第849-1284位的反向互补序列得到的重组载体。
2、转基因植株的构建
将步骤1得到的rnai-esa1导入根癌农杆菌eha105中,将得到的重组菌记为eha105-rnai-esa1,利用eha105-rnai-esa1转化yil42,操作步骤如下:
将去壳后的yil42种子去壳接种到nb培养基,培养5-7天,诱导得到愈伤组织。利用eha105-rnai-esa1侵染愈伤组织,然后在籼稻nb培养基中28℃共培养2-3天,洗去愈伤组织上的菌后将愈伤组织接种在含有200mg/l头孢菌素和200mg/l的特美汀且不含潮霉素的籼稻nb培养基上,28℃恢复培养4d。之后将恢复培养后的愈伤组织接种到筛选培养基(筛选培养基为含有头孢菌素、特美汀和潮霉素的籼稻nb培养基,筛选培养基中头孢菌素和特美汀的浓度均为200mg/l)上,筛选三轮,每轮15d,第一、二轮所用筛选培养基中潮霉素的浓度为30mg/l,第三轮所用筛选培养基中潮霉素的浓度为50mg/l;筛选培养结束后挑选淡黄色、松散、坚硬的颗粒状愈伤组织,即为抗性愈伤组织,将抗性愈伤组织接种到预分化培养基上暗培养7-15d;之后将愈伤组织转到含有分化培养基的三角瓶中,28℃培养(光照16h)30d,得到再生水稻幼芽。将再生的绿色水稻幼芽转移到生根培养基上,28℃培养(光照16h)30d,移栽土壤前炼苗7d,即得到rnai-esa1转基因植株。
其中,(1)nb基本培养基由溶质和溶剂组成,溶剂为水,溶质及浓度如下:
(2)籼稻nb培养基配方如下:4.1g/lnb基本培养基溶质(各溶质间的配比同nb基本培养基),300mg/l水解酪蛋白,2.878g/l脯氨酸,500mg/l谷氨酰胺,30g/l蔗糖,4.6g/l植物凝胶,2mg/l2,4-d,余量为水,ph为5.8。
(3)预分化培养基为向nb基本培养基中添加2mg/lnaa、1mg/l6-ba和5mg/laba得到的培养基,ph为5.8。
(4)分化培养基为向nb基本培养基中添加1mg/lnaa、2mg/l6-ba和1mg/lkt得到的培养基,ph为5.8。
(5)生根培养基为ms培养基配方如下:4.43g/lms(murashige&skoogbasalmediumwithvitamins),30g/l蔗糖,3g/l植物凝胶,ph为5.8。
按照上述方法,利用ptck303替换rnai-esa1构建转基因植株,作为空载体对照植株。
3、转基因植株的鉴定
提取rnai-esa1转基因植株8-13cm小穗的总rna,并反转录为cdna,利用引物对(5′-tggaacagaagaaaggcttg-3′(正向引物),5′-agccacatcctcaagatgaa-3′(反向引物))检测esa1基因表达水平,利用actin基因作为内参,内参引物为:5′-tacagtgtctggattggaggat-3′(正向引物)和5′-tcttggcttagcattcttgggt-3′(反向引物)。并利用yil42和空载体对照植株作为对照。
将与yil42和空载体对照植株相比esa1基因表达水平无显著变化的rnai-esa1转基因植株记为阴性植株,将与yil42和空载体对照植株相比esa1基因表达水平显著下降的rnai-esa1转基因植株记为阳性rnai-esa1转基因植株,选取11株t2代阳性rnai-esa1转基因植株记为rnai-esa1-1~rnai-esa1-11。
部分植株的esa1基因表达量的检测结果如图1中h所示,阳性rnai-esa1转基因植株中esa1基因表达水平均显著下降。
三、表型的鉴定
待测植株为:yil42、空载体对照植株、阴性植株、rnai-esa1-1~rnai-esa1-11。
1、结实率
在待测植株成熟期调查各水稻的结实率。
2、雄配子育性检测
1)碘-碘化钾染色
水稻材料开花前1-2d,从每个待测植株的主茎穗上取3朵部位一致的颖花,置于乙醇∶冰乙酸(3∶1)溶液中于4℃冰箱保存。镜检时用1%i2-ki染色、压片,10×20倍显微镜观察。
2)dapi染色
水稻材料开花前1-2d,将幼穗固定在乙醇∶冰乙酸(3∶1)溶液中于4℃冰箱保存。镜检时在载玻片上滴加一滴dapi染液(1ug/ml),将待测植株的花药置于dapi染液中,用解剖针夹破花药,释放出花粉粒,再用镊子去掉花药碎片。染色20分钟后,盖上盖玻片,荧光显微镜下观察并拍照。
3)花粉在柱头上的萌发情况观察
将待测植株分别在抽穗开花时用记号笔标记开花的时间,然后在30,60,90,120,150分钟后采取已经开过花的颖花,用faa固定液固定24小时后,依次放在乙醇浓度分别为70%、50%、30%的乙醇水溶液中复水,蒸馏水洗2-3次,然后放入10nnaoh溶液中浸泡5-8分钟(56℃),蒸馏水冲洗数次后用0.1%脱色苯胺蓝染色(可以过夜),最后在激光共聚焦显微镜下(leicasp2)观察花粉在柱头上的萌发情况和花粉管伸长情况并拍照。
4)扫描和透射电镜观察
扫描电镜制作流程:将待测植株花粉成熟时期的小穗置于2.5%的戊二醛中室温浸泡3h后,用0.1m的磷酸钠溶液(ph6.8)冲洗3次,每次15min,然后用2%oso4的溶液4℃固定过夜。固定后的样品同样采用0.1m的磷酸钠溶液(ph6.8)冲洗3次,每次15min,经过乙醇浓度分别为30%、50%、70%、80%、90%的乙醇水溶液分别脱水10~15分钟后,在乙醇-异戊基醋酸(v/v=1∶3)混合液中浸泡1h,co2临界点干燥,真空镀膜后,在日立s-3000n型扫描电子显微镜观察,照相。
透射电镜制作流程:收集待测植株花粉成熟时期的小穗立即固定于2.5%戊二醛固定液中,0.1mol/lpbs漂洗2次,30min/次。丙酮梯度脱水,3min/次(70%丙酮配制的饱和醋酸铀4℃过夜)。丙酮和epon812(1∶1)混合液37℃或常温1h,并在纯epon812中37℃过夜,epon812包埋剂包埋,37℃12h,45℃24h,60℃聚合24h。超薄切片厚度约60-70nm,醋酸双氧铀染色25min,柠檬酸铅染色15min后用jem-123o透射电子显微镜观察,照相。
3、雌配子检测:整体染色与透明技术
选取不同发育时期的小花,用faa固定液固定,观察前分离出子房,染色和观察的方法如下:以梯度乙醇水溶液(乙醇浓度分别为70%,50%,30%,15%)和蒸馏水复水,2%的硫酸铝钾媒染20分钟,10mg/l曙红(溶于4%蔗糖溶液)中染色10-12小时,2%的硫酸铝钾分色20分钟,水洗2-3次,再以梯度乙醇水溶液(乙醇浓度分别为30%,50%,70%,80%,90%)和无水乙醇脱水,无水乙醇重复3次。用v无水乙醇:v水杨酸甲酯=1:1的混合液过渡停留1-2小时,最后用纯的水杨酸甲酯透明1小时以上。透明后将子房放在滴有水杨酸甲酯的凹面载玻片上进行整体封片(指甲油封片)。在leicasp2激光扫描共聚焦显微镜下用543nm的波长的激光激发荧光进行成熟胚囊整体分层扫描观察并获取图像。
胚囊发育情况如图1中d、f所示,d图未形成正常胚囊(阴性植株),f图为正常发育的成熟胚囊(阳性rnai-esa1转基因植株)。
结果表明,与阴性植株(结实率为22.42%)相比,阳性rnai-esa1转基因植株雌配子育性明显恢复(结实率为50.85-74.55%),雄配子(即花粉)均表现正常,正常雌配子比例也得到提高。yil42、空载体对照植株与阴性植株的结实率无显著差异,雌雄配子表型也均无显著差异。
实施例2、esa1基因表达检测
待测植株为:近等基因系nil-qssr1,该近等基因系为利用yil42为供体亲本,籼型栽培稻特青为轮回亲本通过杂交、回交排除其他染色体渗入片段后获得的,该近等基因系含有esa1基因。
提取待测植株总rna,并反转录为cdna,利用引物(5′-tggaacagaagaaaggcttg-3′(正向引物),5′-agccacatcctcaagatgaa-3′(反向引物))检测esa1基因表达水平,利用actin基因作为内参,内参引物为:5′-tacagtgtctggattggaggat-3′(正向引物)和5′-tcttggcttagcattcttgggt-3′(反向引物)。
结果如图2所示。结果显示,p8-p13时期(即穗长为8-13cm时期),nil-qssr1穗部的相对表达量较高,该时期小穗发育处于减数分裂后。
<110>中国农业大学
<120>一种提高水稻远缘杂交后代育性的方法与所用蛋白质
<160>3
<170>patentinversion3.5
<210>1
<211>606
<212>prt
<213>水稻(oryzasativa)
<400>1
metgluasnglysertrpglnarghisileserileasnargglnval
151015
serserserservalthralaglnglyleuargalametvalalaser
202530
tyrtyralalysargcyssercysglyphepheleualalysleuleu
354045
leumetglyphealaleuvalglntyrseralaalavalalaserval
505560
valleualaalaleuargleuserlysglnasptyrvalaspproala
65707580
aspglnglyserserasphislysserilelysglyserleuasnleu
859095
phetyrglyleuvalleuvalglnglyalaseraspleuleualagln
100105110
alailephealavalalaaspileglnleuvalleulysilethrglu
115120125
alatyrglnleuglyproleuglylysglnmetvalasnhistyrmet
130135140
leuvalthrtyrleuargcysserglyglyasnvalargglualamet
145150155160
asnmetaspleuvalserphealametgluleuvalargserasnser
165170175
ilealaaspargleuvalglyvalargvalleuaspserileleuarg
180185190
valprolystyrargalaleualaleumetargleuargalaserala
195200205
aspthrvalglyglyvalvalsermetleuglyleuthrasnasnthr
210215220
argglugluvalasnthrargglyhisalaalaglyvalileleuglu
225230235240
leuserargaspleuleuleugluserpheproalametleuproile
245250255
valserserleuilevalalaalaaspasnserglyasnaspvalthr
260265270
valsermetgluphethrtrppheglyvallysileleuasnlysile
275280285
metaspasnproaspasncysasnlysvalalaaspalaaspglygln
290295300
valilealaserilevalasnleuthralavalthrglyaspasparg
305310315320
serleuserilevalserserseralavalargaspglugluileile
325330335
leuglualavalglnvalleuhislysleuvalseralaalaglyasp
340345350
serglyargvalleuargcyslysvalseraspasnvaltyrvalleu
355360365
argasnileserlysileleuglnhisproargserglnvallysleu
370375380
leuvalglualaileglyvalleualacysleualaleuaspgluthr
385390395400
glyargglugluilealaserserproglnileilearglysleuval
405410415
serpheleuvalproargserglnmetilesergluileseralaasp
420425430
argargglnleualalysproasnalaglualaleuvalmetleuala
435440445
metaspasnglnserilevaltrplysileglnglugluleulyspro
450455460
glnaspmetglnlysleuvalglumetleuseralaaspserthrgly
465470475480
phelysthrasnvalalalysleuleuglyileleuhisalaasnser
485490495
argalagluhisalahisleuglnlysthrileileasnthralaleu
500505510
proalaleuleulysalailelyssergluvalglulysleugluasp
515520525
provalleutyralaglygluhisalaasnasnpheglnglutrparg
530535540
thrlysglnglyalaleuleugluserphevalglyleuservalgln
545550555560
ilecysthrserilehisalaserasppheasnglualaleuargser
565570575
alaasnvalthrvaltyrmetvalmetglnlysleuarglysileleu
580585590
aspleutyrlysserproalaileglupheproglyilearg
595600605
<210>2
<211>2615
<212>dna
<213>水稻(oryzasativa)
<400>2
cagcagtgccgtggatccaagggatcagttcatcctcaatgctcctgaagtggtggcggc60
acagaccgaaagatttgaggctatggagaatggaagctggcaaagacacataagcatcaa120
ccggcaggtgtcgtcgtcgtctgtgacagcacaaggcttgagagccatggtcgcgtcata180
ctacgccaagcgctgcagctgcggcttcttcctcgccaagctcctgctgatgggcttcgc240
gctcgtgcagtactccgccgccgtcgcctccgtcgtcctcgccgcgctgcgcctgagcaa300
gcaagactatgttgatccggcggaccagggcagcagcgaccacaagagcatcaagggttc360
actcaacctcttctacggcctggtgctcgtacagggcgcctccgacttgctagcccaagc420
catcttcgccgtcgccgacattcagctcgtgctcaagatcacggaggcgtaccagctcgg480
ccccttggggaagcagatggtgaaccattacatgcttgtcacctacctgagatgctccgg540
tggtaacgtccgcgaagccatgaacatggacctggttagcttcgccatggagctggtgcg600
atccaactccatcgcggatcgcctcgtcggggtccgtgttcttgacagcatccttagagt660
gcccaagtacagagcgctggcgctcatgaggctccgagcttctgccgacacggtcggcgg720
cgtggtcagcatgctaggactgacgaacaacacccgggaggaagtgaataccagagggca780
tgctgcaggcgtcatcttggagctctctcgggaccttctccttgagagcttcccagcaat840
gctgccgatcgtgtcatcgttgatcgttgctgctgacaactctggcaacgatgtgactgt900
gagcatggagttcacctggttcggtgtgaaaatcctcaacaagatcatggacaatccgga960
caactgcaacaaggtcgcggatgctgatggccaggtgattgcaagcattgtgaacctcac1020
agctgttactggtgatgatcgcagcttgagcatagtctcatcatctgcagtcagagacga1080
agagatcattttggaggcagttcaggtgttgcacaagctagttagcgctgctggtgattc1140
tgggagagtgcttaggtgcaaagtctctgacaatgtctatgtactcaggaacattagcaa1200
gatactacaacaccctagaagccaagtaaagctacttgttgaagccattggagttcttgc1260
ttgtttagcattggatgagaccgggagggaagagattgcatcctctccacaaattattag1320
gaagcttgtctctttccttgttccaaggtcacagatgatttctgaaatttcagctgatag1380
aagacagctagcaaaacctaatgctgaagcactagttatgcttgcaatggataatcaaag1440
cattgtctggaagatccaagaagaactgaagcctcaagacatgcagaaacttgtcgagat1500
gctctctgctgactctactgggttcaaaacgaacgttgcaaaacttttgggcattttaca1560
tgccaactccagagcagagcatgctcatcttcagaagacaataatcaacactgcactacc1620
agcggtaatgtccggttattattttctaaaagttttatagtgtccttacagtgttaaatt1680
ctataattctaatgcttgtgctgtcatcatctacagctgctgaaagcaataaagtcagaa1740
gtggagaagctagaggatcctgtactgtatgctggagaacatgctaatgtaagaaagtta1800
tcctccaaatcacacatttctccaaaatcatatatcttttatcagttattaattctctct1860
ctaaatatattcttggtggtgtatataaacacagaatttccaagaatggaggaccaagca1920
gggtgcactactagagagctttgttgggctcagtgttcagatctgcacatccattcacgc1980
aagcgatttcaatgaagctctccggagtgccaatgttacagtgtacatggtcatgcagaa2040
gcttagaaagatcctagatttatacaagtcaccagccatcgagttcccagggataaggtg2100
agtcgcagtagaactaatcatttggatgaagcaatgcagtagccactgcaatgaagtttt2160
ctttcagtgtgaaatggataaggcgctgaaagaagtagctggaacagaagaaaggcttga2220
aatgtttaagatattctactatggtgttggaattgttaagcacagcgagcccatttcttc2280
ccttgtcaacttagctctcggtctctaagctatcactggtatacgctgaatgttctgttt2340
ctttattgcaaaagatgggaacagagatgtgcggccttcatcttgaggatgtggctaagt2400
caaagcttgaaggagaaaagctgaaggaactcgcaagaccttctgaagatgcagaataat2460
tttaattcttgtctagtttggtcatgttagttgttttggttaaacctttgtaggacaagt2520
ccttatcttttgcttgcttgcagtctgaacttgatatattcactcagttattttatttgt2580
aatgaagttctgtggcccaatctttatcaggcaaa2615
<210>3
<211>2417
<212>dna
<213>水稻(oryzasativa)
<400>3
cagcagtgccgtggatccaagggatcagttcatcctcaatgctcctgaagtggtggcggc60
acagaccgaaagatttgaggctatggagaatggaagctggcaaagacacataagcatcaa120
ccggcaggtgtcgtcgtcgtctgtgacagcacaaggcttgagagccatggtcgcgtcata180
ctacgccaagcgctgcagctgcggcttcttcctcgccaagctcctgctgatgggcttcgc240
gctcgtgcagtactccgccgccgtcgcctccgtcgtcctcgccgcgctgcgcctgagcaa300
gcaagactatgttgatccggcggaccagggcagcagcgaccacaagagcatcaagggttc360
actcaacctcttctacggcctggtgctcgtacagggcgcctccgacttgctagcccaagc420
catcttcgccgtcgccgacattcagctcgtgctcaagatcacggaggcgtaccagctcgg480
ccccttggggaagcagatggtgaaccattacatgcttgtcacctacctgagatgctccgg540
tggtaacgtccgcgaagccatgaacatggacctggttagcttcgccatggagctggtgcg600
atccaactccatcgcggatcgcctcgtcggggtccgtgttcttgacagcatccttagagt660
gcccaagtacagagcgctggcgctcatgaggctccgagcttctgccgacacggtcggcgg720
cgtggtcagcatgctaggactgacgaacaacacccgggaggaagtgaataccagagggca780
tgctgcaggcgtcatcttggagctctctcgggaccttctccttgagagcttcccagcaat840
gctgccgatcgtgtcatcgttgatcgttgctgctgacaactctggcaacgatgtgactgt900
gagcatggagttcacctggttcggtgtgaaaatcctcaacaagatcatggacaatccgga960
caactgcaacaaggtcgcggatgctgatggccaggtgattgcaagcattgtgaacctcac1020
agctgttactggtgatgatcgcagcttgagcatagtctcatcatctgcagtcagagacga1080
agagatcattttggaggcagttcaggtgttgcacaagctagttagcgctgctggtgattc1140
tgggagagtgcttaggtgcaaagtctctgacaatgtctatgtactcaggaacattagcaa1200
gatactacaacaccctagaagccaagtaaagctacttgttgaagccattggagttcttgc1260
ttgtttagcattggatgagaccgggagggaagagattgcatcctctccacaaattattag1320
gaagcttgtctctttccttgttccaaggtcacagatgatttctgaaatttcagctgatag1380
aagacagctagcaaaacctaatgctgaagcactagttatgcttgcaatggataatcaaag1440
cattgtctggaagatccaagaagaactgaagcctcaagacatgcagaaacttgtcgagat1500
gctctctgctgactctactgggttcaaaacgaacgttgcaaaacttttgggcattttaca1560
tgccaactccagagcagagcatgctcatcttcagaagacaataatcaacactgcactacc1620
agcgctgctgaaagcaataaagtcagaagtggagaagctagaggatcctgtactgtatgc1680
tggagaacatgctaataatttccaagaatggaggaccaagcagggtgcactactagagag1740
ctttgttgggctcagtgttcagatctgcacatccattcacgcaagcgatttcaatgaagc1800
tctccggagtgccaatgttacagtgtacatggtcatgcagaagcttagaaagatcctaga1860
tttatacaagtcaccagccatcgagttcccagggataaggtgagtcgcagtagaactaat1920
catttggatgaagcaatgcagtagccactgcaatgaagttttctttcagtgtgaaatgga1980
taaggcgctgaaagaagtagctggaacagaagaaaggcttgaaatgtttaagatattcta2040
ctatggtgttggaattgttaagcacagcgagcccatttcttcccttgtcaacttagctct2100
cggtctctaagctatcactggtatacgctgaatgttctgtttctttattgcaaaagatgg2160
gaacagagatgtgcggccttcatcttgaggatgtggctaagtcaaagcttgaaggagaaa2220
agctgaaggaactcgcaagaccttctgaagatgcagaataattttaattcttgtctagtt2280
tggtcatgttagttgttttggttaaacctttgtaggacaagtccttatcttttgcttgct2340
tgcagtctgaacttgatatattcactcagttattttatttgtaatgaagttctgtggccc2400
aatctttatcaggcaaa2417
- 还没有人留言评论。精彩留言会获得点赞!