莨菪醛还原酶及其应用的制作方法
本发明涉及生物技术领域,具体涉及莨菪醛还原酶,还涉及莨菪醛还原酶的应用。
背景技术:
托品烷类生物碱(tropanealkaloids,tas)是一类具有巨大医疗价值的抗胆碱药物,广泛用于麻醉、镇痛、止咳、平喘和抗晕动病,也用于控制帕金森病的僵直和震颤。临床上常用的是莨菪碱(hyoscyamine)和东莨菪碱(scopolamine),两者的市场需求均十分巨大,其中以东莨菪碱毒副作用更弱,药效更强,价格也更昂贵。目前tas都是从少数茄科tas资源植物中提取,包括颠茄(atropabelladonna)、曼陀罗(daturastramonium)和莨菪(hyoscyamusniger),颠茄是东莨菪碱和莨菪碱最主要的商业栽培药源,也是药典收录的tas药源植物。野生颠茄植株中莨菪碱质量分数为0.02%~0.17%(干重),东莨菪碱含量极低,仅为干重的0.01%~0.08%。因此,培育托品烷生物碱高产颠茄一直是该行业长期追求的目标。
大部分植物次生代谢产物其在天然植物中的含量极低,而使用化学合成的方法,工艺流程复杂、成本太高,并且还有许多植物次生代谢产物的生物合成途径不清晰,无法实现化学全合成。因此,研究人员开始探索其他的提高植物次生代谢产物含量的方法。例如,在植物中过量表达次生代谢生物合成途径关键酶基因,打破代谢产物合成的限速步骤,从而促进最终有用代谢产物在植物体内的积累,获得更有经济价值的材料。在颠茄中,过量表达h6h基因后,颠茄中的莨菪碱大量转化为更有价值的东莨菪碱,极大地提高了颠茄的经济价值。莨菪碱既是重要的抗胆碱药物也是东莨菪碱的直接前体。植物次生代谢工程的应用依赖于次生代谢产物生物合成途径的解析。因此,克隆莨菪碱生物合成基因,对于改良并提高颠茄中托品烷生物碱的含量具有重要的意义。
技术实现要素:
有鉴于此,本发明的目的之一在于提供一种莨菪醛还原酶;本发明的目的之二在于提供莨菪醛还原酶基因;本发明的目的之三在于提供含有所述莨菪醛还原酶基因的重组表达载体;本发明的目的之四在于提供含有所述莨菪醛还原酶基因的转基因细胞系或转基因重组菌;本发明的目的之五在于提供所述莨菪醛还原酶在体内或体外催化莨菪醛还原生成莨菪碱的应用;本发明的目的之六在于提供所述莨菪醛还原酶基因、所述的重组表达载体在不具有托品烷生物碱生物合成途径的原核生物或真核生物中重建莨菪碱合成途径中的应用;本发明之七在于提供所述莨菪醛还原酶、所述莨菪醛还原酶基因、所述的重组表达载体、所述的转基因细胞系或转基因重组菌在具有托品烷生物碱生物合成途径的生物中提高莨菪碱含量中的应用;本发明的目的之八在于提供一种提高托品烷生物碱合成植物中莨菪碱含量的方法。
为达到上述目的,本发明提供如下技术方案:
1、莨菪醛还原酶,具有下述氨基酸残基序列之一的蛋白质:
1)如seqidno.4所示的氨基酸残基序列;
2)将seqidno.4中的氨基酸残基经过一个或几个氨基酸残基的取代和/或缺失和/或添加且具有莨菪醛还原酶功能的氨基酸酸序列。
2、莨菪醛还原酶基因,所述莨菪醛还原酶基因具有下述核苷酸序列之一:
1)如seqidno.3所示的核苷酸序列;
2)编码如seqidno.3所示核苷酸序列的多核苷酸;
3)与1)或2)限定的核苷酸序列具有80%以上的同源性且编码具有所述莨菪醛还原酶功能的核苷酸序列;
4)与1)或2)所述的序列杂交的核苷酸序列。
3、含有所述莨菪醛还原酶基因的重组表达载体。
优选的,所述重组表达载体为将所述的莨菪醛还原酶基因插入原核或真核表达载体得到表达莨菪醛还原酶的重组表达载体。
更优选的,所述重组表达载体为将莨菪醛还原酶基因,如seqidno.3所示的核苷酸序列,经ecori和连入saci酶切位点连入pet28a载体而得。或将莨菪醛还原酶基因seqidno.3,如seqidno.3所示的核苷酸序列,替换改造pbi121的gus基因,其中改造pbi121为abpmt启动子替换pbi121的35s启动子。
3、含有所述莨菪醛还原酶基因的转基因细胞系或转基因重组菌。
优选的,所述细胞系为植物细胞系,可以为颠茄细胞系,也可以为具有托品烷生物碱生物合成途径的植物细胞系,也可以为不具有托品烷生物碱生物合成途径的植物细胞系。所述重组菌为bl21,或其他具有或不具有托品烷生物碱生物合成途径的细菌。
5、所述莨菪醛还原酶在体内或体外催化莨菪醛还原生成莨菪碱中的应用。
6、所述莨菪醛还原酶基因在不具有托品烷生物碱生物合成途径的原核生物或真核生物中重建莨菪碱合成途径中的应用。
所述的重组表达载体在不具有托品烷生物碱生物合成途径的原核生物或真核生物中重建莨菪碱合成途径中的应用。
7、所述莨菪醛还原酶在具有托品烷生物碱生物合成途径的生物中提高莨菪碱含量中的应用。
所述莨菪醛还原酶基因在具有托品烷生物碱生物合成途径的生物中提高莨菪碱含量中的应用。
所述的重组表达载体在具有托品烷生物碱生物合成途径的生物中提高莨菪碱含量中的应用。
所述的转基因细胞系或转基因重组菌在具有托品烷生物碱生物合成途径的生物中提高莨菪碱含量中的应用。
8、一种提高托品烷生物碱合成植物中莨菪碱含量的方法,将通过在具有托品烷生物碱生物合成途径的植物中过量表达所述的莨菪醛还原酶基因。
本发明的有益效果在于:本发明公开了莨菪醛还原酶,经研究发现莨菪醛还原酶的氨基酸序列如seqidno.4所示,其编码的核苷酸如seqidno.3所示,将莨菪醛还原酶进行原核表达后能够催化莨菪醛还原生成莨菪碱,将莨菪醛还原酶用于转化颠茄后能够提高颠茄细胞系中莨菪碱的含量,为提高颠茄中托品烷生物碱的含量具有重要的意义。
附图说明
为了使本发明的目的、技术方案和有益效果更加清楚,本发明提供如下附图进行说明:
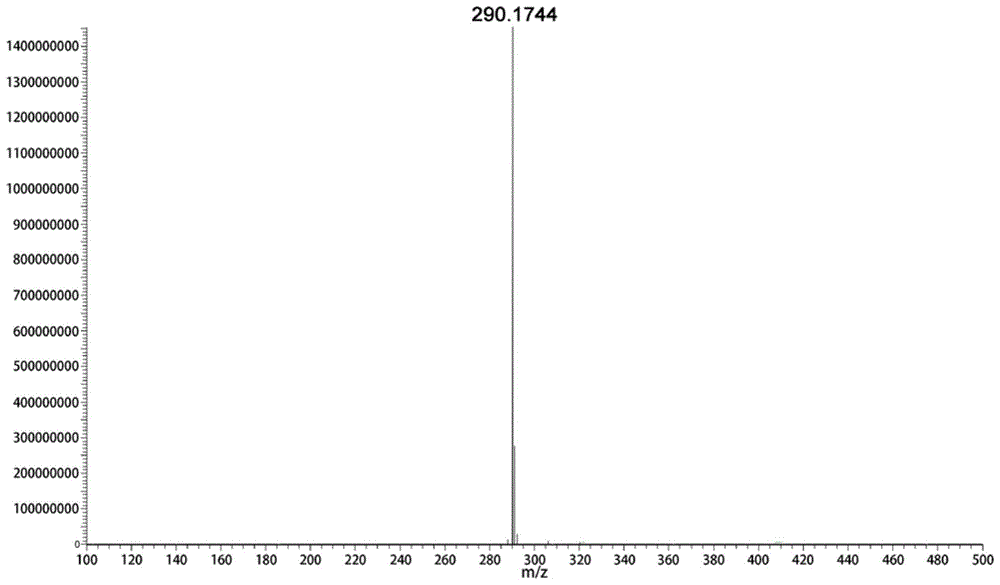
图1为har催化莨菪醛还原为莨菪碱结果图。
图2为转基因颠茄毛状根采用荧光定量检测har基因表达量。
图3为har超表达载体的转基因毛状根和对照组莨菪碱含量。
具体实施方式
下面对本发明的实施例作详细说明:本实施例在以本发明技术方案为前提下进行实施,给出了详细的实施方式和具体的操作过程,但本发明的保护范围不限于下述的实施例。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如sambrook等分子克隆:实验室手册(newyork:coldspringharborlaboratorypress,1989)中所述的条件,或按照制造厂商所建议的条件。
实施例1、莨菪醛还原酶(hyoscyaminealdehydereductasesynthase,har)基因的克隆
(1)颠茄须根总rna的提取
取适量颠茄须根组织,置于液氮中研碎,加入盛有裂解液的1.5mleppendorf(ep)离心管中,充分振荡后,按照tiangen试剂盒的说明书抽提总rna。用甲醛变性胶电泳鉴定总rna质量,在分光光度计上测定rna浓度。
(2)har基因的克隆
以所提取的总rna为模板,按照天根fastkingcdna第一链合成试剂盒说明书合成cdna;设计har基因特异性引物,具体引物如下:
har-f:5’-atggattcttctggtgtcctct-3’(seqidno.1);
har-r:5’-ttggttgctgctcaaacctag-3’(seqidno.2)。
通过pcr从总cdna中扩增har基因,并测序,测序结果获得har基因的核苷酸序列如seqidno.3,起始密码子为atg,终止密码子为taa;翻译出蛋白编码序列如seqidno.4所示。
实施例2、原核表达验证har基因的功能
(1)har的原核表达与蛋白纯化
har基因进行pcr扩增,在正向引物中引入酶切位点ecori,在反向引物中引入酶切位点saci。利用上述两个酶切位点将har编码区完整序列连接质粒pet28a,获得har原核表达载体pet28a-har。引物如下所示:
ecori-har-f:5’-cgcgaattcatggattcttctggtgtcctct-3’(seqidno.5);
xhoi-har-r:5’-cgcctcgagctaggtttgagcagcaaccaa-3’(seqidno.6)。
将构建好的pet28a-har质粒转化原核表达菌株bl21,用pcr筛选阳性克隆子,获得原核表达工程菌bl21-pet28a-har。取bl21-pet28a-har菌液100μl接种到含100mg/l卡那霉素的30mllb液体培养基中,37℃,200rpm过夜培养。再按1:50比例接种量分别接种到400mllb液体培养基中37℃,200rpm进行活化培养,培养至od600=0.6左右时加入iptg至终浓度1mm。在37℃,200rpm条件下继续培养6h。将收获的菌液8000rpm离心,去掉上清。再把菌体沉淀用磷酸盐缓冲液重悬,并在超声破碎后用ge公司的ni-nta填料纯化获得har蛋白。
(2)har酶活验证
har催化的莨菪醛还原反应体系如下:磷酸盐缓冲液(ph6.4),0.2mmnadph-4na,1mm莨菪醛,har蛋白30μg,37.5℃孵育1h,反应产物使用高分辨质谱进行鉴定。
高分辨质谱仪为brukerimpactiiq-tof,色谱柱为waters公司的symmetryc-18反相硅胶柱(3.5μm,100x2.1mm),流动相a为0.1%甲酸水溶液,b为乙腈,采用梯度方式洗脱,洗脱程序见表1:
表1、洗脱程序
柱温设定为40℃,流速0.15ml/min,进样量1μl,质谱检测器使用电喷雾离子源(esi),离子模式为正离子模式。检测结果表明,har催化的莨菪醛还原反应产物为莨菪碱,质荷比m/z为290.1744,保留时间为4.00min(图1)。
实施例3、过表达har提高颠茄莨菪碱含量
(1)har植物过表达载体的构建
为研究har基因对颠茄中托品烷类生物碱的影响,构建了中柱鞘特异性高表达载体pmtpromoter::har,原始质粒为pbi121。首先,利用酶切位点hindiii和xbai将原始质粒上pbi121的35s启动子替换为abpmt启动子;abpmt启动子以颠茄cdna为模板,seqidno.7和seqidno.8所示序列为引物进行pcr扩增,再利用酶切位点bamhi和saci将原始质粒上的gus基因替换为har基因,har基因以颠茄cdna为模板,seqidno.9和seqidno.10所示序列为引物进行pcr扩增,获得har植物过表达载体pmtpromoter::har。使用的引物序列如下:
hindiii-pmtpromoter-f:5’-cgcaagcttctgagttcggatctaggtca-3’(seqidno.7);
bamhi-pmtpromoter-r:5’-cgcggatccttcttcacttttggccttgct-3’(seqidno.8);
bamhi-har-f:5’-cgcggatccatggattcttctggtgtcctct-3’(seqidno.9);
saci-har-r:5’-cgcgagctcctaggtttgagcagcaaccaa-3’(seqidno.10)。
(2)发根农杆菌工程菌的获得
将pmtpromoter::bhar载体采用冻融法转入发根农杆菌(如c58c1),并进行pcr验证。结果表明,含har的植物双元过量表达载体已成功构建到发根农杆菌菌株中。
(3)转基因发根的获得
a.颠茄外植体准备
颠茄种子用75%乙醇浸泡1min,再用50%naclo浸泡20min,无菌水冲洗3-4次,用无菌吸水纸吸干表面水分,接种于无激素的1/2ms固体培养基中,25℃,16h/8h(光亮/黑暗)光照培养,即可获得颠茄无菌苗。该条件下培养2周左右后,剪取无菌苗叶片和下胚轴外植体用于转化。
b.农杆菌与外植体的共培养
将所述外植体,加入活化好的所述含har植物双元超表达载体的发根农杆菌工程菌的重悬液(ms+as100μmol/l)中,菌液与外植体充分接触5分钟,转到共培养培养基(ms+as100μmol/l)上,28℃暗培养2d。
c.抗性毛状根的筛选
将所述的共培养2d的颠茄外植体转入到筛选培养基(ms+kan100mg/l+cef500mg/l)上于25℃暗培养,每周继代培养一次,经过1-2次继代后即可获得kan抗性毛状根。将生长良好的毛状根剪下转入培养基(ms+cef200mg/l)上培养至完全无菌,从而获得kan抗性颠茄毛状根。
d.毛状根的基因组pcr和表达量检测
根据目的基因所在表达盒上游的35s启动子区域和har分别设计正向引物设计和反向引物对目的基因进行检测。结果表明,利用所设计的pcr特异引物,能扩增出特异dna片段。而以非转化颠茄毛状根基因组dna为模板时,没有扩增出任何片段。
将获得的转基因颠茄毛状根采用荧光定量检测,结果如图2所示,转基因毛状根的abhar基因表达量极显著提高。
e.毛状根莨菪碱提取与测定
毛状根在ms培养液中振荡培养30天后收获,材料于冷冻干燥机中冻干至恒重。磨成粉末,准确称取0.1g干燥至恒重的植物材料。加入10ml生物碱提取液(氯仿:甲醇:氨水=15:5:1),超声提取30min,于室温静置1h。提取液过滤去除植物残渣,滤液置于40℃干燥。用5ml氯仿和2ml0.5m的硫酸溶解上一步干燥物,充分乳化以使生物碱转移至水相,弃掉氯仿。将水相置于冰上,并用氨水(28%)调节水相至ph10.0。加入2ml氯仿以萃取生物碱,重复萃取两次,合并所有的氯仿,无水硫酸钠干燥去除水分,过滤,滤液置于40℃干燥。1ml液相色谱级甲醇溶解生物碱,0.22μm滤膜过滤,hplc测定生物碱含量,结果为三次重复的平均值,误差线表示标准差。统计分析用t-test检验。
hplc仪器配置:岛津lc-20ad二元泵系统,配备dug-20a在线脱气机,cto-20a柱温箱,spd-m20a全波长二极管阵列检测器,sil-20a自动进样器,色谱柱为岛津inertsustainc18色谱柱(5μm,4.6×250mm),使用岛津保护柱(5μm,4.0×10mm)。
生物碱分析色谱条件:流动相使用11%乙腈和89%水(20mm乙酸铵和0.1%甲酸,ph4.0),柱温箱为40℃,总流速1ml/min,检测波长为226nm。
结果如图3所示,在本发明中转abhar超表达载体的转基因毛状根显著提高了莨菪碱含量。在普通毛状根中莨菪碱含量为2.32mg/gdw时,同时期abhar过表达载体颠茄毛状根中莨菪碱的含量平均达到4.24-5.98mg/gdw,其含量是非转基因颠茄毛状根含量的1.83-2.58倍。
以上所述实施例仅是为充分说明本发明而所举的较佳的实施例,本发明的保护范围不限于此。本技术领域的技术人员在本发明基础上所作的等同替代或变换,均在本发明的保护范围之内。本发明的保护范围以权利要求书为准。
序列表
<110>西南大学
<120>莨菪醛还原酶及其应用
<160>10
<170>siposequencelisting1.0
<210>1
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>1
atggattcttctggtgtcctct22
<210>2
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>2
ttggttgctgctcaaacctag21
<210>3
<211>1038
<212>dna
<213>颠茄(atropabelladonna)
<400>3
atggattcttctggtgtcctctctccttttacattttcaagaagggcaactggtgaagaa60
gatgtgaggttgaaggtactttattgtgggatttgtcactcagatctgggttgcatcaag120
aatgaatggggctggtgttcataccctcttgtacctggccatgagattgttggcatagca180
acagaggtgggaagcaaagtgacaaaattcaaagttggagacagagtaggagtaggatgc240
atggttggatcctgtggcacttgccaaaactgcacacaaaatcaagaaagttactgtcct300
gaagttataatgacttgtgcttctgcttaccctgatggaacaccaacatatggaggattt360
tcaaatcaaatggtagcaaatgagaagtttgtaattcgaatcccaaattcacttccactt420
gatgctgctgcaccattgctctgtgctggaagtactgtgtatagtgctatgaagttttat480
ggattgtgtagtcaagggttgcacttgggagttgttggtttaggtggacttggtcatgtt540
gctgtgaaatttgcaaaggcatttgggatgaaagtgactgtgattagtacttcacttgga600
aaaaaagaagaagcaattaatcaacttggagctgattcattcttgatcaatactgataca660
gagcaaatgcagggtgccatggaggtaatggatggcattattgatacagtatctgcactt720
catccaattgaaccattgcttggtctactcaaatctcatcaagggaaactcatcattgtt780
ggattacccaataagcaacctgagctgccagtcttttccttgattaatgggaggaaaatg840
attggagggagtgctgttggaggagtgaaagagacacaagaaatgatagattttgcagca900
gagcataatatcactgcagacattgagattgtccccatggattatgtgaacacagccatg960
gaacgtcttgagaagggcgatgtcaaatttcgatttgtcattgatgttgaaaatacattg1020
gttgctgctcaaacctag1038
<210>4
<211>345
<212>prt
<213>颠茄(atropabelladonna)
<400>4
metaspserserglyvalleuserprophethrpheserargargala
151015
thrglyglugluaspvalargleulysvalleutyrcysglyilecys
202530
hisseraspleuglycysilelysasnglutrpglytrpcyssertyr
354045
proleuvalproglyhisgluilevalglyilealathrgluvalgly
505560
serlysvalthrlysphelysvalglyaspargvalglyvalglycys
65707580
metvalglysercysglythrcysglnasncysthrglnasnglnglu
859095
sertyrcysprogluvalilemetthrcysalaseralatyrproasp
100105110
glythrprothrtyrglyglypheserasnglnmetvalalaasnglu
115120125
lysphevalileargileproasnserleuproleuaspalaalaala
130135140
proleuleucysalaglyserthrvaltyrseralametlysphetyr
145150155160
glyleucysserglnglyleuhisleuglyvalvalglyleuglygly
165170175
leuglyhisvalalavallysphealalysalapheglymetlysval
180185190
thrvalileserthrserleuglylyslysgluglualaileasngln
195200205
leuglyalaaspserpheleuileasnthraspthrgluglnmetgln
210215220
glyalametgluvalmetaspglyileileaspthrvalseralaleu
225230235240
hisproilegluproleuleuglyleuleulysserhisglnglylys
245250255
leuileilevalglyleuproasnlysglnprogluleuprovalphe
260265270
serleuileasnglyarglysmetileglyglyseralavalglygly
275280285
vallysgluthrglnglumetileaspphealaalagluhisasnile
290295300
thralaaspilegluilevalprometasptyrvalasnthralamet
305310315320
gluargleuglulysglyaspvallyspheargphevalileaspval
325330335
gluasnthrleuvalalaalaglnthr
340345
<210>5
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>5
cgcgaattcatggattcttctggtgtcctct31
<210>6
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>6
cgcctcgagctaggtttgagcagcaaccaa30
<210>7
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>7
cgcaagcttctgagttcggatctaggtca29
<210>8
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>8
cgcggatccttcttcacttttggccttgct30
<210>9
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>9
cgcggatccatggattcttctggtgtcctct31
<210>10
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>10
cgcgagctcctaggtttgagcagcaaccaa30
- 还没有人留言评论。精彩留言会获得点赞!