抗稻曲病基因OsRFS2在水稻遗传改良中的应用的制作方法
本发明属于植物基因工程技术领域。具体涉及osrfs2基因的功能鉴定及在水稻抗稻曲病改良中的应用。本发明通过基因工程技术改变osrfs2基因的表达丰度,可以改良水稻对稻曲病的抗性。osrfs2基因可用于在培育抗稻曲病的水稻品种中的应用。
背景技术:
随着全世界人口的不断增加、耕地面积的不断减少和环境的不断被破坏,粮食安全危机凸显。作为全世界范围内最为重要的主粮—水稻,病害是限制其安全生产的重要因素之一。尤其近十几年来,逐年严重的稻曲病,不仅造成水稻减产,更产生对人畜有害的毒素,严重威胁粮食安全。稻曲病是由稻曲病菌(病原无性态ustilaginoideavirens,病原有性态villosiclavavirens)引起的水稻穗部病害。近十几年来,随着气候变化和我国高产优质杂交稻的大面积推广,以及水稻水肥水平的提高,稻曲病在我国各个水稻生产区大面积发生。生产上“红莲型”系列杂交稻对稻曲病全部表现为感病。近几年全国稻曲病年均发病面积超过1.5亿亩,稻曲病已经成为危害水稻安全生产的主要病害之一。稻曲病菌影响稻米成熟过程中的营养运输和正常发育,造成空秕率升高和千粒重下降,严重影响稻谷产量。稻曲病菌包裹谷粒,其产生的多种类型稻曲毒素污染稻米,人畜食用被稻曲毒素污染的稻米后,稻曲毒素可强烈抑制人畜微管蛋白组装和细胞骨架形成,严重抑制人畜细胞正常发育,对人畜健康造成极大威胁。文献报道食用稻曲毒素污染稻米的母鸡不产蛋,食用稻曲毒素污染稻米的母猪流产,稻曲毒素污染水体的水产动物发育畸形等。目前农业生产上对稻曲病的防治依赖于大量农药的喷施,而农药的喷施不仅增加人力、物力的投入,农药的残余更会造成对环境的不可修复污染。因此发掘水稻自身的抗稻曲病基因,改良水稻对稻曲病的抗病性,对培育优质抗病水稻新品种具有重要的实际生产意义。
目前针对稻曲病菌—水稻相互作用的研究相对比较少。针对稻曲病菌的研究,国内外少数几个实验室通过构建稻曲病菌突变体库,结合转录组分析等方法克隆了一些稻曲病菌的基因,研究发现它们在稻曲病菌的致病过程中发挥作用,敲除这些基因可以在一定程度上降低稻曲病菌的致病力。针对水稻抗稻曲病基因发掘的研究,通过对稻曲病菌侵染水稻幼穗的转录组分析,发现部分参与水稻的细胞磷酸化、细胞程序性死亡、细胞壁合成、脱落酸信号、钙离子调控、种子淀粉储藏、胚形成等相关基因的表达量在稻曲病菌侵染水稻幼穗前后发生了变化,但这些基因是否真正参与水稻对稻曲病菌的抗性,目前尚缺乏直接的遗传学证据。
对水稻不同种质资源进行稻曲病抗病性比较研究发现不同水稻品种对稻曲病存在抗病性的差异,暗示在部分水稻种质资源中存在尚未发掘的抗稻曲病基因。国内相关研究单位通过构建由抗稻曲病和感稻曲病水稻遗传材料组成的重组自交系群体或导入系群体,在水稻的多条染色体上鉴定到抗稻曲病数量性状位点。或通过全基因组关联分析发掘到基因组上序列变异与抗病性之间存在关联。但截至到目前,这些抗稻曲病数量性状位点中是否包含抗稻曲病基因,以及这些抗稻曲病基因能否通过遗传学手段改良水稻对稻曲病的抗病性不得而知。
本发明研究发现,水稻中的osrfs2基因正调控对稻曲病菌的抗性。过量表达osrfs2基因可以增强水稻对稻曲病菌的抗病性,而敲除osrfs2基因表达可以显著降低水稻对稻曲病菌的抗性。本发明对于重要粮食作物水稻在抗稻曲病的改良上具有重要的意义。
技术实现要素:
本发明的目的是从水稻中分离克隆一个osrfs2基因,旨在通过对水稻osrfs2基因的研究和应用,在水稻抗稻曲病的目标性状上改良水稻的抗性,为水稻遗传育种提供新的基因资源。本发明提供了osrfs2基因及其编码的蛋白质,对该基因进行功能及在水稻对稻曲病改良中的应用进行了生物功能验证。本发明分离的基因编码一个组蛋白变体,其生物学功能为调控水稻对稻曲病的抗性。
本发明分离和应用一个编码组蛋白变体的osrfs2基因的cdna片段,并对该片段的作用机理进行阐述。osrfs2基因具有如seqidno:1所示的核苷酸序列,其编码蛋白质序列如seqidno:2所示。
可以采用本发明中的预测基因序列的dna片段作探针,从bac文库中筛选得到本发明的基因。同样,也可以采用pcr技术,从栽培稻或者禾本科作物的基因组或者cdna中扩增得到本发明的基因。同样也可以采取化学合成的方法获得本发明的基因。
采用以上技术方案,可以分离得到包含osrfs2调控元件,如组成型启动子、诱导型启动子和器官特异性启动子融合构建基因表达载体。本发明的基因在构建到植物表达载体中时,也可使用增强子,这些增强子区域可以是atg起始密码子和邻接区域起始密码子等,但必需与编码序列的阅读框相同,以保证整个序列的翻译。
本发明涉及的osrfs2基因序列可用于在作物,特别是水稻抗病育种、转基因株系、基因新品种中的应用。
实现本发明的具体技术方案如下:
1、根据rt-pcr的结果,从水稻品种中花11中分离出包含osrfs2完整基因的cdna片段,其核苷酸序列如seqidno:1所示。具体的分离方法在实施例1中有详细描述。
2、将上述包含有osrfs2完整基因的cdna片段连接至pu1301载体上,构建得到osrfs2的过量表达载体,申请人将该表达载体命名为ubi:osrfs2oe。具体方法在实施例2中有详细描述。
3、设计osrfs2的crispr基因敲除靶点,构建osrfs2的crispr基因敲除载体。申请人将该表达载体命名为pyl-cas9-grna-osrfs2。具体方法在实施例3中有详细描述。
4、将上述包含osrfs2部分基因的cdna片段连接至pds1301载体上,构建得到osrfs2的抑制表达载体,申请人将该表达载体命名为osrfs2rnai。具体方法在实施例4中有详细描述。
5、利用成熟的农杆菌介导的转基因方法(linandzhang,optimisingthetissuecultureconditionsforhighefficiencytransformationofindicarice.plantcellrep,2005,23:540-548)将表达载体ubi:osrfs2oe、pyl-cas9-grna-osrfs2、osrfs2rnai转入水稻受体中花11中,获得遗传转化植株。具体方法在实施例5中有详细描述。
6、在t2代鉴定ubi:osrfs2oe和osrfs2rnai转基因植株的表达量以及对稻曲病菌的抗性,并对相关表型进行统计分析。具体方法在实施例6和实施例7中有详细描述。在t2代鉴定pyl-cas9-grna-osrfs1转基因植株的基因突变情况,并对相关表型进行统计分析。具体方法在实施例6和实施例7中有详细描述。
与现有技术相比,本发明的优点如下:
本发明描述了分离克隆包含有osrfs2基因完整编码区段的dna片段以及验证osrfs2基因功能的方法。本发明发现osrfs2基因可以控制水稻对稻曲病的抗性,将其过量表达可以增强水稻对稻曲病的抗性,将osrfs2基因抑制表达或者敲除可以降低水稻对稻曲病的抗性。本发明可以为水稻对稻曲病的抗性遗传改良提供一种新方法。
附图说明
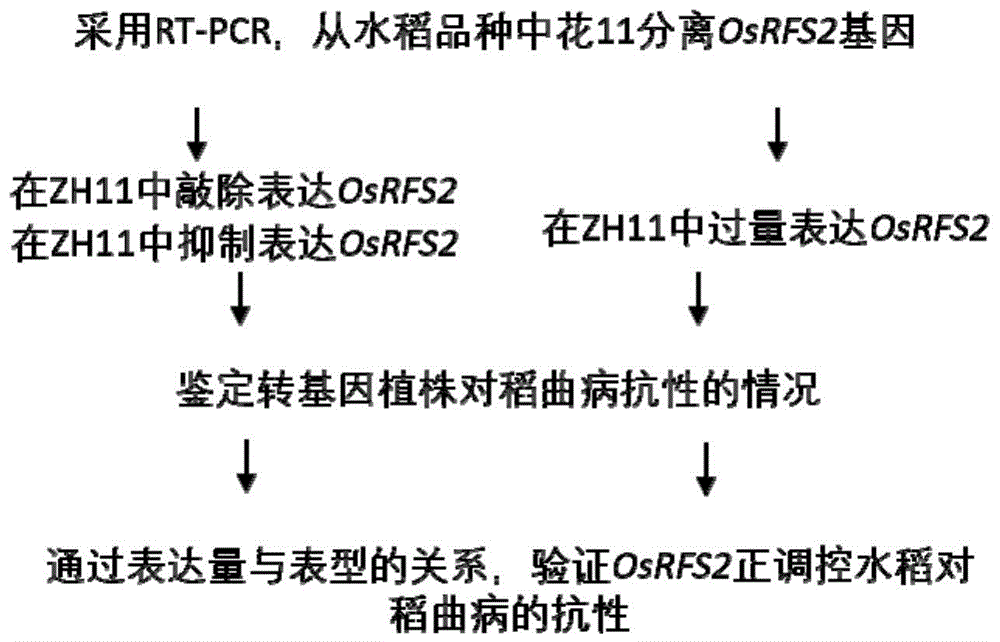
图1:本发明鉴定和分离克隆水稻抗稻曲病osrfs2基因以及验证osrfs2基因功能的流程图。
图2:本发明构建的表达载体ubi:osrfs2oe的构建过程示意图。附图标记说明:图2中的a图:用于构建表达载体ubi:osrfs2oe的空载体pu1301的结构示意图。图2中的b图:插入表达载体ubi:osrfs2oe的包含有osrfs2基因编码区的dna片段示意图。rb和lb分别表示t-dna的右边界和左边界,gus表示β-葡糖苷酸酶基因,hpt表示潮霉素磷酸转移酶基因,ubi表示玉米泛素ubiquitin基因的启动子,35s表示花椰菜花叶病毒的启动子,nos表示多聚腺苷酸化信号序列终止子,kpni和bamhi为限制性内切酶。图2中的c图:在水稻中花11背景中表达载体ubi:osrfs2oe获得转基因水稻中osrfs2基因表达量检测。
图3:crispr载体pyl-cas9-grna-osrfs2构建过程示意图。附图标记说明:图3中的a图:pyl-cas9-grna的载体图谱。p35:hpt表示花椰菜花叶病毒的35s启动子启动表达的潮霉素磷酸转移酶基因,pubi表示玉米泛素基因启动子,nls表示核定位信号序列,tnos表示根癌农杆菌终止子序列。图3中的b图:osrfs2基因crispr靶点t1和t2的grna的设计位点。图3中的c图:osrfs2基因敲除水稻株系与野生型相比缺失的脱氧核糖核苷酸数目及位置。图3中的d图:琼脂糖凝胶电泳检测osrfs2基因敲除水稻株系的pcr产物与野生型相比明显变小。
图4:抑制表达载体osrfs2rnai构建过程示意图。附图标记说明:图4中a图:插入表达载体osrfs2rnai的包含有osrfs2基因编码区的dna片段示意图。rb和lb分别表示t-dna的右边界和左边界,gus表示β-葡糖苷酸酶基因,hpt表示潮霉素磷酸转移酶基因,35s表示花椰菜花叶病毒的启动子,nos表示多聚腺苷酸化信号序列终止子,kpni、bamhi、saci和spei为限制性内切酶。图4中的b图:在水稻中花11背景中表达载体osrfs2rnai获得转基因水稻中osrfs2基因表达量检测。
图5:ubi:osrfs2oe、pyl-cas9-grna-osrfs2和osrfs2rnai的t2代转基因植株对稻曲病的抗病性分析。附图标记说明:图5中的a图:水稻中花11背景osrfs2基因超量表达家系5和9接种稻曲病菌后每穗稻曲球数。相比野生型(即非转基因,下同),携带ubi:osrfs2oe载体构建的转基因水稻接种稻曲病菌后每穗稻曲球数显著降低。图5中的b图:水稻中花11背景osrfs2基因超量表达家系5和9接种稻曲病菌后每穗稻曲球率。相比野生型,携带ubi:osrfs2oe载体构建的转基因水稻接种稻曲病菌后每穗稻曲球率显著降低。图5中的c图:水稻中花11背景osrfs2基因敲除家系1和2接种稻曲病菌后每穗稻曲球数。相比野生型,携带pyl-cas9-grna-osrfs2载体构建的转基因水稻接种稻曲病菌后每穗稻曲球数显著增加。图5中的d图:水稻中花11背景osrfs2基因敲除家系1和2接种稻曲病菌后每穗稻曲球率。相比野生型,携带pyl-cas9-grna-osrfs2载体构建的转基因水稻接种稻曲病菌后每穗稻曲球率显著增加。图5中的e图:水稻中花11背景osrfs2基因抑制表达家系2和4接种稻曲病菌后每穗稻曲球数。相比野生型,携带osrfs2rnai载体构建的转基因水稻接种稻曲病菌后每穗稻曲球数显著增加。图5中的f图:水稻中花11背景osrfs2基因抑制表达家系2和4接种稻曲病菌后每穗稻曲球率。相比野生型,携带osrfs2rnai载体构建的转基因水稻接种稻曲病菌后每穗稻曲球率显著增加。
具体实施方式
对序列表的说明:
序列表seqidno:1是本发明分离克隆的包含有osrfs2基因的核苷酸序列(序列长度为1-2810bp,其中:1-358bp是5'utr;359-475bp是第一个外显子(exton);476-2120bp是内含子(intron);2121-2417bp是第二个外显子(exton);2418-2810bp是3'utr)和对应的氨基酸序列。编码137个氨基酸残基。
根据以下的描述和这些实施例,本领域技术人员可以确定本发明的基本特征,并且在不偏离本发明精神和范围的情况下,对本发明做出各种改变和修改,以使其适用各种用途和条件。
下述实施例中所用方法如无特别说明均为本领域常规方法或试剂,具体步骤可参考:例如:《molecularcloning:alaboratorymanual》(sambrook,j.,russell,davidw.,molecularcloning:alaboratorymanual,3rdedition,2001,ny,coldspringharbor)或相关产品(实施本发明不限于此)。
实施例1:分离克隆包含有osrfs2基因完整编码区段的dna片段
为了得到osrfs2基因编码的完整dna片段,本发明采用invitrogen公司生产的trizol抽提试剂盒(具体操作步骤按照试剂盒提供的说明书操作),抽提水稻品种中花11(或称zh11)(来自中国农业科学院作物科学研究所)0.5mm长的幼穗的总rna。采用invitrogen公司生产的dnasei和反转录酶,对其进行反转录。经过反转录得到的cdna为模板,利用osrfs2基因(该基因的核苷酸序列如seqidno:1所示)的特异引物,rfs2oef(正向引物):5'-gaattcatggcgggaaaaggaggtaa-3'(下划线的序列为kpni识别位点);rfs2oer(反向引物):5'-ggatccctctttggaggacttgttga-3'(下划线的序列为bamhi识别位点);通过pcr的方法分离克隆osrfs2基因的编码区即cds序列(序列见seqidno:1),其蛋白质序列如seqidno:2所示。
实施例2:构建osrfs2基因的蛋白质编码区的过量表达载体
用kpni和bamhi双酶切实施例1获得的pcr产物,然后与经过kpni和bamhi双酶切的载体pu1301(该载体是公开报道的一种商业质粒:zhouetal.,over-expressionofaspartateaminotransferasegenesinriceresultedinalterednitrogenmetabolismandincreasedaminoacidcontentinseeds.theorapplgenet,2009,118:1381-1390;其基本骨架是澳大利亚cambia实验室的pcambia1301,通过加入玉米ubiquitin启动子实现对目标基因的表达调控,其结构如图2中的a图所示),利用t4dna连接酶(购自promega公司,具体用法与用量参考该公司产品的说明书)进行连接。连接产物通过热激化转的方法转入大肠杆菌tran-t1(购自北京全式金生物技术有限公司)中,加400μllb培养基复苏45min,涂于含50mg/l的卡那霉素的la培养基平板上,37℃温箱培养14-16h(la与lb配方参考:萨姆布鲁克,《分子克隆实验指南》第三版,科学出版社,北京,2002年)。挑取单克隆,扩大培养并抽提质粒,通过kpni和bamhi双酶切筛选阳性克隆,并将所得的表达载体命名为ubi:osrfs2oe。
实施例3:构建osrfs2基因敲除载体pyl-cas9-grna-osrfs2
针对水稻osrfs2基因设计基于crispr/cas9的sgrna序列,位于osrfs2基因的靠近5’端的外显子处,其核苷酸序列为t1:5’>acgattcgcgcccgatctgc<3’;t2:5’>gccaagtcggcggagaagga<3’(见图3中的b图)。将sgrnat1和t2通过重叠延伸pcr的方法分别插入到载体pyl-sgrna-osu3和pyl-sgrna-osu6a中,获得pcr片段osu3-t1-grna-polyt和osu6a-t2-grna-polyt。将以上pcr片段通过bsai酶切连接至pyl-cas9-grna载体中(见图3中的a图)。载体pyl-cas9-grna、pyl-sgrna-osu3和pyl-sgrna-osu6a由华南农业大学刘耀光教授惠赠(载体信息见已发表文章,maetal.,arobustcrispr/cas9systemforconvenient,high-efficiencymultiplexgenomeeditinginmonocotanddicotplants.molplant.2015,8:1274–1284)。具体操作步骤如下所述:
(1)osu3-t1-grna-polyt以及osu6a-t2-grna-polyt片段的构建
第一轮pcr以pyl-sgrna-osu3质粒为模板,用引物b1’和t1r扩增osu3启动子与osrfs2基因20bp的t1靶序列;同样以pyl-sgrna-osu3为模板,用引物t1f和b2扩增osrfs2基因t1靶序列与grna-polyt;第二轮pcr是以第一轮pcr产物为模板,用引物b1’和b2扩增得到osu3-t1-grna-polyt片段。
采用同样的方法获得osu6a-t2-grna-polyt的片段。第一轮pcr以pyl-sgrna-osu6a为模板,用引物b2’和t2r扩增osu6a启动子与osrfs2基因20bp的t2靶序列;同样以pyl-sgrna-osu6a为模板,用引物t2f和bl扩增osrfs2基因t2靶序列与grna-polyt;第二轮pcr是以第一轮pcr产物为模板,用引物b2’和bl扩增得到osu6a-t2-grna-polyt片段。该步骤使用到的引物序列信息为:
b1’:5'-ttcagaggtctctctcgcactggaatcggcagcaaagg-3'(下划线序列为bsai酶切位点);
b2:5'-agcgtgggtctcgtcagggtccatccactccaagctc-3'(下划线序列为bsai酶切位点);
b2’:5'-ttcagaggtctctctgacactggaatcggcagcaaagg-3'(下划线序列为bsai酶切位点);
bl:5'-agcgtgggtctcgaccgggtccatccactccaagctc-3'(下划线的序列为bsai酶切位点);
t1f:5'-acgtgtctccgggatgttgggttttagagctagaaatagc-3'(下划线序列为靶点t1);
t1r:5'-ccaacatcccggagacacgttgccacggatcatctgcaca-3'(下划线序列为靶点t1);
t2f:5'-gccccattgttcgcgccaaggttttagagctagaaatagc-3'(下划线序列为靶点t2);
t2r:5'-ccaagtcggcggagaaggacggcagccaagccagcacc-3'(下划线序列为靶点t2)。
(2)osrfs2的crispr突变载体pyl-cas9-grna-osrfs2的构建
利用限制性内切酶bsai,通过边酶切边连接的方法,将osu3-t1-grna-polyt和osu6-t2-grna-polyt的pcr片段连接到pyl-cas9-grna载体的bsai位点处中(参见图3中的a图)。
实施例4:构建osrfs2基因抑制载体osrfs2rnai
利用osrfs2基因的特异引物,rfs2dsf(正向引物):5'-gggactagtggtaccgacaggagcatccttagtt-3'(单下划线的序列为spei识别位点,双下划线的序列为kpni识别位点);rfs2dsr(反向引物):5'-ggggagctcggatccaggaacaagccgtagaaat-3'(单下划线的序列为saci识别位点,双下划线的序列为bamhi识别位点);通过pcr的方法获得osrfs2基因的一段特异序列的pcr产物,将pcr产物和载体pds1301(该载体是公开报道的一种质粒:chuetal.,promotermutationsofanessentialgeneforpollendevelopmentresultindiseaseresistanceinrice.gene&development,2006,20:1250-1255)同时用kpni和bamhi进行双酶切消化,利用t4dna连接酶(购自promega公司,具体用法与用量参考该公司产品的说明书)进行连接,得到中间载体pds1301-frag1。再将pcr产物和中间载体pds1301-frag1采用spei和saci进行双酶切消化,利用t4dna连接酶进行连接。连接产物通过热激化转的方法转入大肠杆菌tran-t1(购自北京全式金生物技术有限公司)中,加400μllb培养基复苏45min,涂于含50mg/l的卡那霉素的la培养基平板上,37℃温箱培养14-16h(la与lb配方参考:萨姆布鲁克,《分子克隆实验指南》第三版,科学出版社,北京,2002年)。挑取单克隆,扩大培养并抽提质粒,通过kpni和bamhi双酶切筛选阳性克隆,并将所得的表达载体命名为osrfs2rnai。
实施例5:转基因水稻的获得
将表达载体ubi:osrfs2oe(来自实施例2)、pyl-cas9-grna-osrfs2(来自实施例3)、osrfs2rnai(来自实施例4)通过农杆菌eha105介导的遗传转化方法转入水稻品种中花11(zh11)的愈伤组织,经过常规的预培养、侵染培养、共培养和筛选培养得到具有对潮霉素(用于筛选阳性的转基因愈伤的一种抗生素,购买自丹麦的罗氏制药有限公司)抗性的愈伤组织,再经过分化培养、生根培养,炼苗并移栽至大田,得到转基因植株。本发明的农杆菌遗传转化的方法和所用的试剂及配方是根据hiei等人的报道优化而来(hieietal.,efficienttransformationofrice(oryzasatival.)mediatedbyagrobacteriumandsequenceanalysisoftheboundariesofthet-dna.plantj.1994,6:271-282;linandzhang,optimisingthetissuecultureconditionsforhighefficiencytransformationofindicarice.plantcellrep.2005,23:540-548)(上述的优化方法是华中农业大学作物遗传改良国家重点实验室近20年来已经公开的专利申请文献所记载的方法,为公知方法)。
实施例6:osrfs2基因转基因水稻的鉴定
(1)osrfs2基因过量表达转基因植株的表达量检测
1)抽提来自ubi:osrfs2oe的转基因水稻分蘖期叶片的rna,反转录合成cdna。具体操作方法和过程同实施例1。
2)对得到的反转录产物用引物进行qrt-pcr的检测(引物组合为rfs2rt-f1和rfs2rt-r1),并且用水稻的持家actin基因(登陆号loc_os03g13170)的扩增结果作为内参(引物组合为actinrt-f和actinrt-r),序列如下所示:
rfs2rt-f1:5'-gtcggcggagaaggacaag-3'
rfs2rt-r1:5'-catttgcctgagtcctttgct-3'
actinrt-f:5'-aaccagctgaggcccaaga-3'
actinrt-r:5'-acgattgatttaaccagtccatga-3'
pcr反应总体积为20μl,包含cdna模板1ul,10×pcrbuffer2μl,10mmdntp2μl,10mm引物各0.2μl,rtaq酶0.2μl,加去离子水至20μl。pcr反应条件如下:①94℃4min;②94℃40s;③58℃30s;④72℃30s;⑤从步骤②至步骤④循环28次;⑥72℃7min;⑦4℃保存。pcr产物在2%(质量/体积)的tbe琼脂糖凝胶上电泳检测。
rt-pcr检测结果表明,表达载体ubi:osrfs2oe转基因阳性植株相对于野生型,osrfs2基因表达量明显上升,结果如图2c所示。表达载体osrfs2rnai转基因阳性植株相对于野生型,osrfs2基因表达量明显下降。结果如图4b所示。
(2)osrfs2基因敲除的转基因植株检测
选取靶点t1和t2的上下游序列设计引物,进行pcr扩增,对扩增片段进行测序,通过测序峰图判断靶位点t1和t2之间的dna区域是否发生碱基缺失。鉴定osrfs2基因缺失效果的引物序列如下:
rfs2e2f:5'-tctggcaagggcagtttggta-3'
rfs2e2r:5'-catacctacacaaatctaat-3'
用引物rfs2e2f对osrfs2基因敲除的转基因单株的pcr产物进行测序,鉴定osrfs2基因序列缺失情况。本发明成功获得osrfs2基因缺失的转基因植株osrfs2-1和osrfs2-2,其中转基因植株osrfs2-1缺失242个脱氧核糖核苷酸,转基因植株osrfs2-2缺失429个脱氧核糖核苷酸,结果见图3c和图3d。
实施例7:转基因水稻对抗稻曲病鉴定验证
为了分析转基因水稻植株对稻曲病的抗性,申请人将中花11背景的ubi:osrfs2oe、pyl-cas9-grna-osrfs2、osrfs2rnai各2个独立t2家系种子经过常规浸种、催芽后播种于秧田,20天后移栽至大田,种植密度即株行距为15*24厘米,种植地点为中国湖北省武汉市洪山区华中农业大学试验田,在一个有安全防护设施条件下按常规的水稻种植方法进行田间管理。在水稻分蘖期,申请人将水稻移栽在面包盒中,移至生长箱中。在水稻抽穗前一周,对osrfs2基因过量表达水稻家系5和家系9、敲除表达水稻家系osrfs2-1和osrfs2-2、抑制表达水稻家系2和家系4以及野生型中花11进行稻曲病菌接种试验。结果显示,本发明的osrfs2基因过量表达水稻家系5和家系9在孕穗期接种稻曲病菌后,与野生型中花11(非转基因,下同)相比,osrfs2基因过量表达水稻家系5和家系9转基因阳性单株每穗稻曲球数和每穗稻曲球率均显著性少于野生型对照(p<0.01),结果见图5a、图5b。以上结果表明,osrfs2基因超量表达水稻株系能增强水稻对稻曲病菌的抗性。而osrfs2基因敲除表达水稻家系osrfs2-1和osrfs2-2单株每穗稻曲球数和每穗稻曲球率均显著性高于野生型(非转基因)对照(p<0.01),结果见图5c、图5d。本发明的osrfs2基因抑制表达水稻家系2和家系4在孕穗期接种稻曲病菌后,与野生型中花11(非转基因,下同)相比,osrfs2基因抑制表达水稻家系2和家系4转基因阳性单株每穗稻曲球数和每穗稻曲球率均显著性高于野生型对照(p<0.01),结果见图5e、图5f。以上结果表明,osrfs2基因被敲除和抑制表达后的转基因水稻株系能降低水稻对稻曲病菌的抗性。
序列表
<110>华中农业大学
<120>抗稻曲病基因osrfs2在水稻遗传改良中的应用
<141>2020-06-29
<160>1
<170>siposequencelisting1.0
<210>1
<211>3102
<212>dna
<213>水稻(oryzasativa)
<220>
<221>gene
<222>(1)..(3102)
<220>
<221>3'utr
<222>(2625)..(3084)
<220>
<221>exon
<222>(771)..(2624)
<220>
<221>5'utr
<222>(1)..(770)
<400>1
caccacaccggagaggtgagtgagagtgagagagtgagagcagagaccaccaccaccgga60
gaggttagtgagagaggagtggtaatggtgaggcaacaagagtaggttccatttcatatc120
atcactaggatagcgtagtttgtaggctgcatctccatctccatcgccattgattcgcat180
tgcatccatcattttaggatgttctactagggttcttgatttttcttttggtttgttgtt240
ttgacgaatggaggtattgttgggattcgccgcctgctgctcgtcgtcgtcgtcgccgat300
gaggaggccgtgcgggctctgccccggcatgtccgatcgttcgtgatttgttttttctac360
atgttttagggcccatttgttcttgatcctattctttgattcttttgtactaagcattct420
aaggcgaagccacccattctttcctgcatatatacttacaaacacatagcccccatctga480
tctcacaaacattatttctctctctttttttctcagttttttctttgttgatttactgac540
caaattctttggaagaacaacaagatcatctggtttttatctgctcattcttttgtacat600
cgaatcatatacatttccattccaccaaagccttagccagataccacagagagagtgtga660
gagaaatcagagtgagaaacagaggaggaagaagaagaagaagacgaggaggaggaggag720
gaggagcaggaggaggaggaggtctcttcttggcacgtcgcgttccggcgagtgacgtgt780
ctccgggatgttggcgggttgctcgttctcgtcgtcgaggcatcagatgagcaccgcgca840
gcgtttcgacatcctcccctgcggcttctccaagcgcggcagccgcggcgacggcgccgc900
cccgcgggtcgccggcgacgccaggagcggcgccaccacctgctccttccggacgcaccc960
cgcgccgccggtcacccagtccgtgtcctggggcgccaagccggagcccggcggcaatgg1020
caatggcgcccaccgcgccgttaagcgggcgcatgacgaggacgcggtcgaggagtatgg1080
ccccattgttcgcgccaagcggacgcggatgggcggcgacggcgatgaggtatggttcca1140
tcaatccattgcagggacgatgcaagcgacggcggcgggagaaggagaggaggcggagga1200
ggagaaggtcttcttggtgccgagcgcggcggcgttcccgcacggcatggccgccgcggg1260
gccatcgctggccgcggccaagaaggaggagtacagcaagtcgccgtccgactcgtcgtc1320
ctcgtcgggcacggacggcggctcgtcggcgatgatgccgccgccgcagccgcccgagtt1380
cgacgcgaggaacggcgtgccggcgccggggcaggcggagcgggaggcgctggagctggt1440
gcgcgcgctcaccgcgtgcgccgactccctctccgccggcaaccacgaggccgccaacta1500
ctacctggcccggctcggcgagatggcctcgccggcggggcccacgccgatgcaccgcgt1560
ggccgcctacttcaccgaggcgctcgcgctccgcgtcgtgcgcatgtggccgcacatgtt1620
cgacatcggcccgccgcgggagctcaccgacgacgccttcggcggcggcgacgacgacgc1680
catggcgctgcggatactcaacgccatcacgcccatcccgaggttcctgcacttcacgct1740
caacgagcgcctcctccgcgagttcgaggggcacgagcgcgtccacgtcatcgacttcga1800
catcaagcaggggctccaatggccgggcttgctccagagcctggccgcgcgggcggtgcc1860
tccggcgcacgtgcggatcaccggagtcggcgagtcgaggcaggagctgcaggagacggg1920
agcgcggctggcgcgcgtcgccgccgcgctcggcctggcgttcgagttccacgccgtggt1980
cgaccggctcgaggacgtccgcctgtggatgctccacgtcaagcgcggcgagtgcgtggc2040
cgtgaactgcgtcctcgccatgcaccgcctgctccgcgacgacgccgcgctgaccgactt2100
cctggggctagcgcgcagcacgggcgccaccatcctcctcctcggcgagcacgagggcgg2160
cggcctcaactcggggaggtgggaggcgcggttcgcgcgcgcgctgcggtactacgccgc2220
ggcgttcgacgcggtggacgcggcggggctgccggaggcgagccccgcgagggccaaggc2280
ggaggagatgttcgcgcgggagatccgcaacgcggtggcgttcgagggccccgagcggtt2340
cgagcgccacgagagcttcgccgggtggcggcggcgcatggaggacggcggcgggttcaa2400
gaacgccggcatcggcgagcgcgaggcgatgcaggggcgcatgatcgcgaggatgttcgg2460
gccggacaagtacaccgtgcaggcgcacggcggcggcggcagcggcggcggcgaggcgct2520
cacgctccggtggctggaccagccgctgtacaccgtgacggcgtggacgccggcgggcga2580
cggcgcgggaggcagcaccgtgtcggcgtccacaacagcatcacattctcagcaaagcta2640
agctgacgatgaatggtgattaggtgaagagaaagaaagaacaaagcctttttttacagt2700
gcttcttttgttaatgatgattagttcatacagtatgacaattcttttatacattcagag2760
aaaagaaagaagaaagaaaggtgtagttttttgttttatagattgataggtggaaagatt2820
caattaaatcaaattcaattcaatttttagattgtaattctttataaatattcttttggc2880
tgttgagagagagtcccctgcaaaatgtagctgcatgtagaagaaagaaagcaaagaagc2940
agtagatagattagcaggggcagcatctctcacagtcactattagtgtctccggctgtta3000
ttatacaacattattattacaatcaaattctttcatcattcattctacatgtaatctctg3060
ttcagaatcagaatgaaatgaaacatgtgttatatttctcca3102
- 还没有人留言评论。精彩留言会获得点赞!