针对L1CAM的单域抗体及其衍生蛋白和应用的制作方法
针对l1cam的单域抗体及其衍生蛋白和应用
技术领域
1.本发明属于免疫学领域,涉及针对l1cam的单域抗体及其衍生蛋白和应用。
背景技术:
2.肿瘤的发生是一个多因素、多阶段的演进过程,近年来研究发现,l1cam(l1细胞黏附分子, l1
‑
cell adhesion molecule)在人类多种恶性肿瘤中表达增高,并与肿瘤的进展、转移、侵袭、预后密切相关,是肿瘤治疗的潜在靶点。分子靶向治疗相对于手术、放化疗三大传统治疗手段更具有“治本”的功效,具有能效高并选择性杀伤肿瘤细胞特点,减少对正常组织损伤。分子靶向相对其他治疗,带来的副作用较少,更有针对性的治疗肿瘤。
3.l1cam是
‑
种细胞黏附受体,属于神经细胞黏附分子,是免疫球蛋白超家族中的一员。l1cam 作为成熟期、非有丝分裂期的细胞黏附受体运行,除了在神经系统的发育及生理过程中起重要作用, l1cam还存在于许多肿瘤组织中,参与多种类型肿瘤的增殖、扩散、转移和侵袭。在结肠癌的研究中, l1cam呈现高表达。在70%的结肠癌中可以检测到l1cam,而在正常的结肠组织中无表达。当l1cam 引入缺乏l1cam的结肠癌细胞,这些细胞则变得具有侵袭、转移能力,在裸鼠体内的肝脏形成了大体转移灶,其分子机制为l1cam的胞外域裂解可以促进转移机制的形成。在乳腺癌中,l1cam的表达使细胞逃避凋亡,l1cam可以促进细胞运动,其机制不是依赖于erk,而是因为细胞之间的黏附连接丢失。在乳腺癌mcf7细胞系中,l1cam的过度表达导致细胞之间的黏附连接中断,细胞离散,增加了β
‑
catenin
‑
tcf的转录活动。但给予sirna基因抑制时,内源性的l1cam表达明显减少,mcf7 细胞运动减少,细胞聚集。在卵巢癌中,在各种组织类型中79%表达l1cam与结肠癌相似,l1cam 的表达与卵巢癌的不良预后和转移存在密切关系。l1cam可通过金属蛋白酶裂解形成ml1cam和 sl1cam。ml1cam高表达水平可抑制抑癌基因p53活化,并与能否行满意的卵巢癌肿瘤细胞减灭术密切相关。卵巢癌患者的血清和腹水细胞可检测到可溶性l1cam(sl1cam)。sl1cam已被证明是一个与临床病理类型及患者预后密切的诊断标记物,其高表达预示对化疗的敏感度降低及预后不良,在体外可以促进细胞迁移,侵袭和保护细胞凋亡。l1cam的过表达使得il
‑
1β的分泌增加并诱导nf
‑
kb 持续活化,因此肿瘤细胞裂解物水平可间接通过il
‑
1β水平测定。在肾细胞癌中,46%的透明细胞性肾细胞癌和28%的乳头状肾癌表达l1cam,正常肾脏通常不表达l1cam。在透明细胞癌中,l1cam 的表达与肿瘤转移机制相关,通过联合cyclind1表达的缺失的多因素分析,l1cam被定义为肿瘤发生转移的独立预后因素。在各种人体皮肤恶性黑色素瘤的组织学亚型中,l1cam的阳性表达率为42%,其阳性表达证明l1cam与转移性疾病相关,成为黑色素瘤患者评价预后的一个积极预测因子。在甲状腺癌中,koonsoonkim等通过免疫组化方法检测l1cam表达水平,发现在正常甲状腺上皮细胞及甲状腺分化癌中无表达。在甲状腺未分化肿瘤(atc)中存在明显的过表达。
4.li
‑
cam的作用机制除与细胞黏附有关外,还与信号传导通路有关,如图1。li
‑
cam与受体的结合能在胞内启动信号的传递并与其它受体的信号途径实现对接。在癌细胞中,l1cam可以结合到酪氨酸激酶受体(rtks)及整合蛋白,导致细胞外信号调节激酶(erk)活
化,从而促进细胞周期进程,诱导血管生成和通过激活磷酸肌醇3激酶/蛋白激酶b(pi3k/akt)信号通路或bcl
‑
2途径来诱导细胞凋亡。新的研究显示l1cam的下游信号除makps/erk外,还可通过两个旁路信号途径:“正向”信号通过调节性膜内蛋白水解和“反向”信号通过转录因子的核因子(nf)
‑
kb7。目前就l1cam的作用机制并没用统一的定论,但更多地倾向于wnt/β
‑
catenin信号通路。wnt信号通路的异常活化在各种肿瘤的增殖﹑侵袭和转移过程中起着至关重要的作用。正常成熟细胞中wnt信号通路处于抑制状态,大部分细胞质β
‑
catenin与e
‑
cadherin结合参与细胞黏附,从而维持细胞的稳定性。少量游离的β
‑
catenin 被gsk
‑
3p磷酸化而被降解,使细胞质β
‑
catenin保持在极低浓度,wnt/3
‑
catenin信号通路信号通过细胞质内游离的变异β
‑
catenin的过度聚积并进入细胞核,与tcf/lef结合而激活相应的靶基因如c
‑
myc 基因、cyclind1的转录过程,从而调控细胞的增殖与凋亡,参与肿瘤的发生。β
‑
catenin作为wnt信号通路的细胞黏附和信号转导分子,其水平除受β
‑
catenin基因突变影响外,还受β
‑
catenin的降解复合体,如l
‑
cam、apc、axin、gsk
‑
33、ck1,细胞黏附受体cd44、upar、mmps、细胞外短阵结构等基因影响,能够调节靶基因表达,使得晚期肿瘤细胞变得更具浸润性及转移性。
5.由于l1cam是存在于细胞表面,根据这个特性,从细胞外部对它进行治疗干预,通过加入针对 l1cam细胞外结构域进行干预的抗体,在细胞培养中能够有效地对肿瘤细胞起到抑制作用,将更大程度地杀伤肿瘤细胞。研究显示,在正常的乳腺上皮细胞中不表达l
‑
cam(大多数其他正常组织的上皮组织细胞也不存在表达),因此在培养基中(甚至在有机体中)加入抗l1cam抗体对细胞及组织无作用,而抗l1cam细胞外区域的抗体有效地抑制乳腺癌细胞的增殖。在人类结肠癌细胞系sw480和 hct116中,当抗l
‑
cam抗体加入到培养基中,细胞的侵袭及转移能力均下降。通过向表达l1cam 的卵巢癌细胞的裸鼠腹腔注射抗l1cam抗体可有效抑制肿瘤细胞的腹膜播散,因此,在裸鼠上证明,抗l1cam抗体的治疗可有效抑制肿瘤细胞增长。其他的研究已经证明l1cam抗体也能够直接应用于表达l1cam的癌细胞的化学治疗和放射治疗中。当放射性元素铜
‑
67与抗l1cam抗体的单克隆抗体结合,注入到种植有skov3ip的卵巢癌细胞的裸鼠腹膜中,可见正常组织的放射元素积累量较低,而肿瘤组织放射元素量水平高。这种治疗结果证实裸鼠体内skov3ip卵巢癌细胞本质上的受到抑制,并延长裸鼠的生存期。
6.综上所述,可以通过单克隆抗体的应用将l1cam作为分子靶向治疗的靶点。
技术实现要素:
7.为了克服上述缺陷,本发明的目的是提供针对l1cam的单域抗体及其衍生蛋白和应用,使用生物基因工程技术筛选出特异性针对l1cam单域抗体,这些抗体初步亲和力明显,并且具有阻断特定细胞释放细胞因子,通过原核表达即具有良好的结合活性,具有一定的成药性。
8.本发明的第一方面,提供了针对l1cam的单域抗体,所述的单域抗体由重链构成,重链包括重链cdr1、重链cdr2和重链cdr3;
9.所述的重链cdr1、重链cdr2和重链cdr3的氨基酸序列为下述(1)、(2)、(3)或者(4):
10.(1)seq id no:10所示的cdr1,seq id no:12所示的cdr2,seq id no:17所示的
cdr3;
11.(2)seq id no:10所示的cdr1,seq id no:13所示的cdr2,seq id no:17所示的cdr3;
12.(3)seq id no:11所示的cdr1,seq id no:14所示的cdr2,seq id no:16所示的cdr3;
13.(4)seq id no:9所示的cdr1,seq id no:15所示的cdr2,seq id no:16所示的cdr3。
14.即,所述的重链包括互补决定区cdr;所述互补决定区cdr包括重链cdr1、cdr2和cdr3的氨基酸序列。以上cdr序列(1)
‑
(4)依次与seq id no.1
‑
4对应。上述所有序列,可以替换为与该序列具有“至少80%同源性”的序列或仅一个或少数几个氨基酸替换的序列;优选为“至少85%同源性”,更优选为“至少90%同源性”,更优选为“至少95%同源性”,最优选为“至少98%同源性”。
15.在优选实施例中,所述的单域抗体的序列还包括框架区fr;所述框架区fr包括fr1、fr2、fr3 和fr4的氨基酸序列;
16.所述的单域抗体的框架区fr的序列为下述(a)、(b)、(c)或者(d);
17.(a)seq id no:20所示的fr1,seq id no:21所示的fr2,seq id no:24所示的fr3,seq id no:28 所示的fr4,或其变体,所述变体在所述fr中包含至多3个氨基酸的替换;
18.(b)seq id no:18所示的fr1,seq id no:21所示的fr2,seq id no:25所示的fr3,seq id no:28 所示的fr4,或其变体,所述变体在所述fr中包含至多3个氨基酸的替换;
19.(c)seq id no:19所示的fr1,seq id no:23所示的fr2,seq id no:26所示的fr3,seq id no:28 所示的fr4,或其变体,所述变体在所述fr中包含至多3个氨基酸的替换;
20.(d)seq id no:19所示的fr1,seq id no:22所示的fr2,seq id no:27所示的fr3,seq idno:28所示的fr4,或其变体,所述变体在所述fr中包含至多3个氨基酸的替换。
21.在一个实施方案中,所述针对l1cam的单域抗体与选自seq id no:1
‑
4的氨基酸序列具有至少 90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%序列同源性,并且能够特异性结合l1cam抗原。
22.在另一个优选实施例中,所述针对l1cam的单域抗体与选自seq id no:1
‑
4的氨基酸序列具有至少95%序列同源性,并且能够特异性结合l1cam抗原。
23.本发明的第二方面是提供针对l1cam的单域抗体,所述单域抗体分别如seq id no.1
‑
4所示,或者所述单域抗体与seq id no.1
‑
4的氨基酸序列具有至少95%序列同源性。
24.在一个实施方案中,编码所述针对l1cam的单域抗体的核酸分子与选自seq id no:5
‑
8的核苷酸序列具有至少90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、100%序列同源性,并且其编码的针对l1cam单域抗体能够特异性结合l1cam抗原。
25.优选的,所述单域抗体的编码序列分别如seq id no.5
‑
8所示,或者与seq id no.5
‑
8具有至少95%序列同源性。
26.本发明的第三方面是提供前述的针对l1cam的单域抗体的fc融合抗体。
27.本发明的第四个方面是提供编码前述的针对l1cam单域抗体的核苷酸分子,其核苷酸序列分别如 seq id no:5
‑
8所示,或者与seq id no.5
‑
8具有至少95%序列同源性。
28.本发明的第五个方面是提供一种表达载体,其包含编码前述的单域抗体或者前述
的fc融合抗体的核苷酸分子或者前述的核苷酸分子。
29.本发明的第六个方面是提供一种宿主细胞,其可以表达出前述的针对l1cam的单域抗体,或其包含前述的表达载体。
30.本发明还提供一种产生针对l1cam的单域抗体或其fc融合抗体的方法,包括步骤:(a)在适合产生单域抗体或其fc融合抗体的条件下,培养前述的宿主细胞,从而获得含所述针对l1cam单域抗体或其fc融合抗体的培养物;(b)从所述培养物中分离或回收所述的针对l1cam单域抗体或其fc融合抗体;以及(c)任选地,纯化和/或修饰步骤(b)中获得的针对l1cam单域抗体或其fc融合抗体。
31.本发明的第七个方面是提供一种药物组合物,所述药物组合物含有:(i)如前述的针对l1cam的单域抗体、或前述的针对l1cam的单域抗体的fc融合抗体;以及(ii)一种或几种药学上可接受的赋型剂。
32.本发明还提供前述的针对l1cam的单域抗体在制备抑制l1cam基因表达的药物或抗肿瘤药物中的用途。抑制l1cam基因表达的药物可适用于l1cam基因高表达的任何病症。优选的,所述的肿瘤包括但不限于结肠癌、乳腺癌、卵巢癌、甲状腺癌、肾细胞癌、黑色素瘤。
33.本发明还提供前述的针对l1cam的单域抗体、或前述的针对l1cam的单域抗体的fc融合抗体的用途,用于制备试剂、检测板或试剂盒;其中,所述试剂、检测板或试剂盒用于:检测样品中l1cam 蛋白的存在和/或含量。
34.所述的单域抗体为vhh,其仅包含抗体重链,不包含抗体轻链。
35.本发明使用生物基因工程技术筛选出特异性针对l1cam的单域抗体,这些抗体初步亲和力明显,并且具有阻断特定细胞释放细胞因子,通过原核表达即具有良好额结合活性,具有一定的成药性,这些单域抗体具有以下优势:
36.(1)这些单域抗体的表达体系选择灵活,既可以再原核系统中表达也可以在酵母细胞或哺乳动物细胞的真核系统中表达,且其在原核表达系统中的表达成本低,可降低后期生产成本。
37.(2)由于单域抗体为单结构域抗体,所以其抗体的多组合形式改造更为简单,通过基因工程的方式简单的串联即可以获得多价、多特异性的抗体,并且其免疫异质性很低,在不经过人源化改造的情况下也不会产生较强的免疫反应。
38.(3)正如多篇文献报道的那样,单域抗体的亲和力范围更为宽泛,在未进行亲和力成熟之前,其亲和力范围可以从nm级别至pm级别,为后期不同用途的抗体提供多个选择。
附图说明
39.图1人源重组l1cam蛋白sds
‑
page分析图;
40.图2vhh序列插入率分析,这里的vhh1
‑
30是从构建的针对l1cam单域抗体文库中随机挑取的不同克隆的pcr产物;
41.图3靶向l1cam淘选的文库富集情况;
42.图4l1cam靶点部分原核表达抗体sds
‑
page;
43.图5l1cam靶点抗体抗原结合活性。
具体实施方式
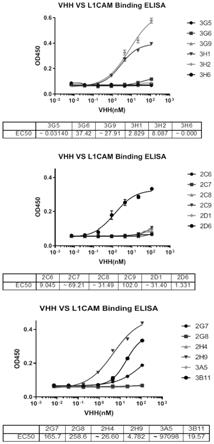
44.下面结合实施例对本发明做进一步的详细说明,以令本领域技术人员参照说明书文字能够据以实施。
45.单域抗体(sdab,也被研发者ablynx称为纳米抗体或vhh)是本领域技术人员公知的。单域抗体为其互补决定区是单域多肽的一部分的抗体。因此,单域抗体包含单个互补决定区(单个cdr1、单个 cdr2和单个cdr3)。单域抗体的实例为仅有重链的抗体(该抗体天然不包含轻链)、衍生自常规抗体的单域抗体和工程化抗体。
46.单域抗体可以衍生自任何物种,包括小鼠、人、骆驼、美洲驼、山羊、兔和牛。例如,天然存在的vhh分子可以衍生自骆驼科物种(例如骆驼、单峰骆驼、美洲驼和原驼)提供的抗体。像完整的抗体一样,单域抗体能够选择性地结合特定抗原。单域抗体可以仅含有免疫球蛋白链的可变结构域,该结构域具有cdr1、cdr2和cdr3以及框架区。
47.如本文所用,术语“序列同源性”表示两个(核苷酸或氨基酸)序列在比对中在相同位置处具有相同残基的程度,并且通常表示为百分数。优选地,同源性在被比较的序列的整体长度上确定。因此,具有完全相同序列的两个拷贝具有100%同源性。
48.本发明中,与本发明公开的cdr1
‑
3的序列同源性高的序列,也可以得到针对l1cam的纳米抗体。在一些实施例中,与(1)
‑
(4)中的序列具有“至少80%同源性”的序列,或者“至少85%同源性”、“至少90%同源性”、“至少95%同源性”、“至少98%同源性”的序列都可以实现发明目的(即衍生蛋白)。
49.在一些实施例中,与(1)
‑
(4)中的序列相比仅仅替换一个或少数几个氨基酸的序列,例如,包含1、2、3、4、5、6、7、8、9或10个保守氨基酸取代,也可以实现发明目的。实际上,在确定两个氨基酸序列之间的序列同源性程度或在确定单域抗体中的cdr1、cdr2和cdr3组合时,技术人员可以考虑所谓的“保守”氨基酸取代,在取代情况下,所述取代将优选为保守氨基酸取代,所述保守氨基酸,其通常可以被描述为氨基酸残基被具有相似化学结构的另一个氨基酸残基替代的氨基酸取代,并且该取代对多肽的功能、活性或其它生物学性质几乎没有或基本上没有影响。所述保守氨基酸取代在本领域是通用的,例如保守氨基酸取代是以下(a)
‑
(d)组内的一个或少数几个氨基酸被同一组内另一个或少数几个氨基酸所取代:(a)极性带负电残基及其不带电酰胺:asp、asn、glu、gln;(b)极性带正电残基:his、arg、lys;(c)芳香族残基:phe、trp、tyr;(d)脂肪族非极性或弱极性残基: ala、ser、thr、gly、pro、met、leu、ile、val、cys。特别优选的保守氨基酸取代如下:asp被glu 取代;asn被gln或his取代;glu被asp取代;gln被asn取代;his被asn或gln取代;arg被lys 取代;lys被arg、gln取代;phe被met、leu、tyr取代;trp被tyr取代;tyr被phe、trp取代; ala被gly或ser取代;ser被thr取代;thr被ser取代;gly被ala或pro取代;met被leu、tyr或 ile取代;leu被ile或val取代;ile被leu或val取代;val被ile或leu取代;cys被ser取代。另外,本领域技术人员知晓,单域抗体的创造性体现在cdr1
‑
3区,而框架区序列fr1
‑
4并非是不可改变的, fr1
‑
4的序列可以采取本发明公开的序列的保守序列变体。
50.本发明的优选宿主细胞为细菌细胞、真菌细胞或哺乳动物细胞。
51.本专利是通过基因工程技术制备目标蛋白以及目标蛋白的截短体形式,然后将获得的抗原蛋白免疫内蒙古阿拉善双峰驼,通过多次免疫后,获得骆驼的外周血淋巴细胞或者脾细胞,通过基因工程的方式,将驼源抗体可变区编码序列重组到噬菌体展示载体中,通
过噬菌体展示技术分别筛选出针对 l1cam的特异性抗体,并进一步检测其与抗原结合的能力以及在包括自身免疫疾病治疗中的应用。
52.现将上述技术方案拆分详解,以具体实施例的方式描述:
53.实施例1:人源l1cam重组胞外结构域蛋白的制备:
54.本专利中用到的人源重组胞外结构域蛋白为公司自己表达纯化获得,人源重组l1cam蛋白的表达载体设计方案具体如下:
55.(1)在ncbi中检索获得l1cam的编码序列,其收录号为nm_000425.4,该序列编码产生的氨基酸序列登录号为np_000416.1。
56.(2)分别通过tmhmm和smart网站对np_000416.1对应的氨基酸序列进行蛋白跨膜区和胞外端的分析。
57.(3)分析结果显示l1cam蛋白的胞外端为22~1120位氨基酸。
58.(4)利用基因合成的方式将编码l1cam蛋白的22~1120位氨基酸的核苷酸序列克隆到载体 pcdna3.4中。
59.(5)将构建好的载体进行sanger测序,比对原始序列,确认无误后,将该重组质粒进行批量抽提,去除内毒素,转染悬浮293f进行目的蛋白的表达、纯化,纯化后l1cam重组蛋白的sds
‑
page 分析结果如图1所示,纯化后蛋白纯度高达90%,满足动物免疫需求。
60.实施例2:针对l1cam蛋白的单域抗体文库的构建:
61.(1)将1mg实施例1中纯化获得的蛋白与等体积的弗氏完全佐剂混合,免疫一只内蒙古阿拉善双峰驼,每周免疫一次,共连续免疫7次,除首次免疫外,其余六次均是用600μg l1cam重组蛋白与弗氏不完全佐剂等体积混合进行动物免疫,该免疫过程是为了集中刺激骆驼使其产生针对l1cam 的抗体。
62.(2)动物免疫结束后,抽取骆驼外周血淋巴细胞100ml并提取细胞的rna;
63.(3)利用提取的总rna合成cdna,并通过套式pcr反应以cdna为模板扩增vhh(重链抗体可变区);
64.(4)利用限制性内切酶分别酶切pmecs载体和vhh片段,然后将酶切后的片段和载体链接;
65.(5)将连接后的片段点转化至感受态细胞tg1中,构建l1cam蛋白的噬菌体展示文库并测定库容,通过pcr检测文库中vhh片段的正确插入率,检测结果如图2,结果显示,从文库中随机挑选的30个菌落进行pcr扩增后,有28个克隆可以扩增出大小为600bp(预测大小)的条带,有2个克隆扩增出条带不正确,故正确插入率为28
÷
30
×
100%≈93.3%。
66.实施例3:针对l1cam蛋白的单域抗体筛选:
67.(1)取200μl重组tg1细胞至2
×
ty培养基中培养,期间加入40μl辅助噬菌体vcsm13侵染tg1细胞,并培养过夜以扩增噬菌体,次日利用peg/nacl沉淀噬菌体,离心收集扩增噬菌体;
68.(2)将稀释在100mm ph 8.3的nahco3中的l1cam蛋白500μg偶联在酶标板上,4℃放置过夜,同时设立阴性对照孔;
69.(3)第二天加入200μl的3%的脱脂乳,室温封闭2h;
70.(4)封闭结束后,加入100μl扩增后噬菌体文库(大约2
×
10
11
个噬菌体颗粒),室温作用1h;
71.(5)作用1小时后,用pbs+0.05%tween
‑
20洗5遍,以洗掉未结合的噬菌体;
72.(6)用终浓度为2.5mg/ml的胰蛋白酶将与l1cam蛋白特异性结合的噬菌体解离下,并感染处于对数生长期的大肠杆菌tg1细胞,37℃培养1h,产生并收集噬菌体用于下一轮的筛选,相同筛选过程重复1轮,逐步得到富集,当富集倍数达到10倍以上时,富集效果如图3所示,图3中,p/n=生物淘选中阳性孔洗脱下的噬菌体感染tg1细菌后生长的单克隆细菌数/阳性孔洗脱下的噬菌体感染 tg1细菌后生长的单克隆细菌数,该参数在富集发生后会逐渐增大;i/e=生物淘选中每轮加入阳性孔的噬菌体总量/生物淘选中每轮从阳性孔洗脱出的噬菌体总量,该参数在富集发生后会逐渐趋近于1。
73.实施例4:用噬菌体的酶联免疫方法(elisa)筛选针对l1cam的特异性阳性克隆:
74.(1)根据上述单域抗体筛选方法对l1cam蛋白进行3轮筛选,筛选结束后,针对重组l1cam 蛋白的噬菌体富集因子达到10以上,从筛选获得的阳性克隆中挑选400个单菌落分别接种于含 100μg/ml氨苄青霉素的tb培养基的96深孔板中,并设置空白对照,37℃培养至对数期后,加入终浓度为1mm的iptg,28℃培养过夜;
75.(2)利用渗透涨破法获得粗提抗体;将l1cam重组蛋白分别释至100mm ph 8.3的nahco3中并将100μg蛋白在酶标板中4℃包被过夜;
76.(3)将上述步骤中获得的抗体粗提液取100μl转移至加入抗原的elisa板上,室温孵育1h;
77.(4)用pbst洗去未结合的抗体,加入100μl经1:2000稀释后的mouse anti
‑
ha tag antibody (鼠抗ha抗体,thermo fisher),在室温孵育1h;
78.(5)用pbst洗去未结合的抗体,加入100ul经1:20000稀释后的anti
‑
rabbit hrp conjugate (山羊抗兔辣根过氧化物酶标记抗体,购自于thermo fisher),在室温孵育1h;
79.(6)用pbst洗去未结合的抗体,加入辣根过氧化物酶显色液,37℃下反应15min后,加入中止液,于酶标仪上450nm波长处,读取吸收值;
80.(7)当样品孔od值大于对照孔5倍以上时,判定为阳性克隆孔;
81.(8)将阳性克隆孔的菌转摇在含有100μg/μl氨苄青霉素的lb培养基中以便提取质粒并进行测序;
82.(9)根据序列比对软件vector nti分析各个克隆株的基因序列,把cdr1,cdr2,cdr3序列相同的株视为同一克隆株,而序列不同的株视为不同克隆株,最终获得特异性针对l1cam蛋白的单域抗体(seq id no.1
‑
4,以及未示出序列的单域抗体3g5、3g6、3g9、3h6、2c6、2c7、2c8、2c9、2d1、2g7、2g8、2h4、3a5、3b11)。其抗体的氨基酸序列为fr1
‑
cdr1
‑
fr2
‑
cdr2
‑
fr3
‑
cdr3
‑
fr4 结构,构成整个vhh。获得的单域抗体重组质粒可以在原核系统中进行表达,最终获得单域抗体蛋白。
83.4种单域抗体的cdr、fr序列如表1所示,4种单域抗体的氨基酸序列、核苷酸序列如表2所示。
[0084][0085]
表1 4种单域抗体的cdr、fr序列
[0086][0087]
表2 4种单域抗体的氨基酸序列、核苷酸序列
[0088]
实施例5:l1cam蛋白的特异性单域抗体在宿主菌大肠杆菌中的纯化及表达
[0089]
(1)将上述测序分析所获得不同克隆株的质粒(pmecs
‑
vhh)电转化到大肠杆菌hb2151中,并将其涂布在lb+amp+glucose即含有氨苄青霉素和葡萄糖的培养平板上,37℃培养过夜;
[0090]
(2)挑选单个菌落接种在5ml含有岸边青霉素的lb培养液中,37℃摇床培养过夜;
[0091]
(3)接种1ml的过夜培养菌种至330ml tb培养液中,37℃摇床培养,培养到od600nm值达到0.6
‑
0.9时,加入1m iptg,28℃摇床培养过夜;
[0092]
(4)离心,收集大肠杆菌,利用渗透胀破法,获得抗体粗提液;
[0093]
(5)通过镍柱亲和层析法纯化出抗体,纯化后的单域抗体,部分克隆表达结果如图4所示,图4中vhh1
‑
18分别对应3g5、3g6、3g9、3h1、3h2、3h6、2c6、2c7、2c8、2c9、2d1、2d6、 2g7、2g8、2h4、2h9、3a5、3b11,其中2d6、3h1、2h9、3h2的序列分别如seq id no:1
‑
4所示,其余单域抗体的序列未示出(为技术效果不够好或者无需在本发明申请中予以保护的单域抗体)。
[0094]
实施例6:l1cam蛋白的特异性单域抗体的fc融合抗体真核表达载体的构建
[0095]
(1)将实施例4中获得的目标序列亚克隆至真核表达载体中:将实施例4中筛选出
来的抗体经sanger测序得到其核苷酸序列;
[0096]
(2)通过序列合成的方式将密码子优化后的上述核苷酸序列(seq id no.5
‑
8)合成至本公司设计改造的载体rjk
‑
v4
‑
hfc中,该载体的改造方法如实施例10所述;
[0097]
(3)将公司构建好的重组真核表达载体转化至dh5α大肠杆菌中,培养进行质粒大提,去除内毒素;
[0098]
(4)将大提后的质粒再进行序列测序鉴定;
[0099]
(5)将确定无误后的重组载体准备后续真核细胞转染表达。
[0100]
实施例7:l1cam蛋白的特异性单域抗体的fc融合抗体在悬浮expicho
‑
s细胞中表达
[0101]
(1)转染前3天以2.5
×
105/ml细胞传代和扩大培养expicho
‑
s
tm
细胞,计算出的所需的细胞体积转移至装有新鲜的已预热的120ml(终体积)的expicho
tm
表达培养基的500ml摇瓶中;使细胞浓度达到约4
×
106‑6×
106活细胞/ml;
[0102]
(2)在转染前一天,将expicho
‑
s
tm
细胞稀释浓度至3.5
×
106活细胞/ml,使细胞过夜培养;
[0103]
(3)转染当天,测定细胞密度和活细胞百分比。转染之前细胞密度应达到约7
×
106‑
10
×
106活细胞/ml;
[0104]
(4)用预热至37℃新鲜的expicho
tm
表达培养基将细胞稀释至6
×
106个活细胞/ml。计算出的所需的细胞体积转移至装有新鲜的已预热的100ml(终体积)的expicho
tm
表达培养基的500ml 摇瓶中;
[0105]
(5)使轻轻颠倒混匀expifectamine
tm
cho试剂,用3.7ml optipro
tm
培养基稀释 expifectamine
tm
cho试剂,回荡或混匀;
[0106]
(6)用冷藏的4ml optipro
tm
培养基稀释质粒dna,回荡混匀;所述的质粒dna为实施例 6制得的h1cam蛋白的特异性单域抗体的fc融合抗体真核表达载体;
[0107]
(7)将expifectamine cho/质粒dna复合物室温孵育1
‑
5分钟,然后轻轻加入制备的细胞悬液中,加入过程中轻轻回荡摇瓶;
[0108]
(8)将细胞在37℃、8%co2、加湿的空气中震荡培养;
[0109]
(9)转染后第1天(18
‑
22小时后)添加600ul expifectamine
tm
cho enhancer和24ml expichofeed。
[0110]
(10)在转染后约8天(细胞活率低于70%)收集上清。
[0111]
实施例8:l1cam蛋白的特异性单域抗体的fc融合抗体在悬浮293f细胞中的表达
[0112]
重组单域抗体表达实验流程(以500ml摇瓶为例):
[0113]
(1)转染前3天以2.5
×
105/ml细胞传代和扩大培养293f细胞,计算出的所需的细胞体积转移至装有新鲜的已预热的120ml(终体积)的opm
‑
293 cd05 medium培养基的500ml摇瓶中。使细胞浓度达到约2
×
106‑3×
106活细胞/ml。
[0114]
(2)转染当天,测定细胞密度和活细胞百分比。转染之前细胞密度应达到约2
×
106‑3×
106活细胞/ml。
[0115]
(3)用预热的opm
‑
293 cd05 medium将细胞稀释至1
×
106个活细胞/ml。计算出所需的细胞体积转移至装有新鲜的已预热的100ml(终体积)的培养基的500ml摇瓶中。
[0116]
(4)用4ml opti
‑
mem培养基稀释pei(1mg/ml)试剂,回荡或吹打混匀;用4ml opt
‑
mem 培养基稀释质粒dna,回荡混匀,并用0.22um的滤头过滤。室温孵育5min。
[0117]
(5)将稀释的pei试剂加入稀释的dna中,颠倒混匀。将pei/质粒dna复合物室温孵育15
‑
20 分钟,然后轻轻加入制备的细胞悬液中,加入过程中轻轻回荡摇瓶。
[0118]
(6)将细胞在37℃、5%co2、120rpm震荡培养。
[0119]
(7)转染后第24h、72h添加5ml opm
‑
cho pff05补料。
[0120]
(8)在转染后约7天(细胞活率低于70%)收集上清。
[0121]
实施例9:人源fc重组单域抗体的纯化
[0122]
(1)将实施例7或8中获得蛋白表达上清用0.45μm的一次性滤头过滤除掉不可溶杂质;
[0123]
(2)将上述滤液使用蛋白纯化仪进行亲和层析纯化,利用人源fc与protein a结合的能力,使用偶联protein a的琼脂糖填料进行纯化;
[0124]
(3)将滤液通过1ml/分钟的流速流穿protein a预装柱,该步骤中滤液中的目标蛋白会与填料结合;
[0125]
(4)通过低盐和高盐缓冲液将柱上结合的杂质蛋白洗涤;
[0126]
(5)用低ph缓冲液将柱上结合的目标蛋白进行系统;
[0127]
(6)将洗脱液迅速加入ph9.0的tris
‑
hcl溶液,进行中和;
[0128]
(7)将上述中和后的蛋白溶液透析后,进行sds
‑
page分析,确定蛋白纯度在95%以上,且浓度在0.5mg/ml以上后,低温保存备用。
[0129]
实施例10:纳米抗体真核表达载体rjk
‑
v4
‑
hfc的构建
[0130]
所提及纳米抗体通用的目标载体rjk
‑
v4
‑
hfc,为本公司在invitrogen商业化载体pcdna3.4(载体资料链接:
[0131]
https://assets.thermofisher.com/tfs
‑
assets/lsg/manuals/pcdna3_4_topo_ta_cloning_kit_man.pdf)的基础上融合了人源igg的重链编码序列(ncbi accession no.:ab776838.1)中的fc区段后改造而来的,即该载体包含了igg重链的铰链区(hinge)ch2和ch3区。具体改造方案如下:
[0132]
(1)选取pcdna3.4上的限制性酶切位点xbai和agei;
[0133]
(2)在fc片段编码序列的5’端和3’端通过重叠pcr的方式分别引入多克隆位点(mcs, multiple cloning site)和6
×
his标签;
[0134]
(3)使用分别带有xbai和agei酶切位点的一对引物通过pcr的方式将上述片段扩增;
[0135]
(4)使用限制性内切酶xbai和agei分别酶切pcdna3.4和(3)中的重组dna片段;
[0136]
(5)将酶切后的载体和插入片段在t4连接酶的作用下连接,然后将连接产物转化至大肠杆菌,扩增,测序核实,获得重组质粒。
[0137]
实施例11:l1cam蛋白的特异性单域抗体的结合量效曲线测定
[0138]
(1)包被50μl 1μg/ml l1cam,4℃过夜。
[0139]
(2)洗板;加入200μl5%牛奶,37℃封闭1h。
[0140]
(3)将vhh稀释至2μg/ml,然后5倍梯度稀释抗体共8个浓度梯度,这里的vhh即实施例5中经原核表达制得的l1cam蛋白的特异性单域抗体。
[0141]
(4)洗板;加入50μl经步骤(3)稀释的单域抗体,两复孔,37℃孵育1h。
[0142]
(5)洗板;加入50μl鼠抗ha标签hrp二抗,37℃孵育30min。
[0143]
(6)洗板(多洗几次);加入50μl预先恢复常温的tmb,避光常温反应15min。
[0144]
(7)加入50μl终止液(1n hcl),酶标仪读数保存。
[0145]
(8)绘制曲线,计算ec50,如图5所示和表3所示,可见本发明的针对l1cam的单域抗体 2d6、3h1、2h9、3h2对l1cam蛋白的结合效力和特异性优异。
[0146] 3g53g63g93h13h23h6ec5o~0.0314037.42~27.912.8298.087~0.000
[0147] 2c62c72c82c92d12d6ec509.045~69.21~31.49102.0~31.401.331
[0148] 2g72g82h42h93a53b11ec50165.7258.6~26.604.782~9709819.57
[0149]
表3各单域抗体的ec50值
[0150]
尽管本发明的实施方案已公开如上,但其并不仅仅限于说明书和实施方式中所列运用,它完全可以被适用于各种适合本发明的领域,对于熟悉本领域的人员而言,可容易地实现另外的修改,因此在不背离权利要求及等同范围所限定的一般概念下,本发明并不限于特定的细节。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1