一种双顺反子翻译偶联表达载体及其应用
1.本发明涉及基因工程生物技术领域,具体涉及的是一种分子伴侣与贻贝蛋白构建成双顺反子翻译偶联表达载体并用其进行协同表达提高可溶性表达水平的方法。
背景技术:
2.海洋贻贝在各沿海国家均有分布,其足端能够分泌足丝,使贻贝能够在高强度的冲击下和潮湿水环境中仍然牢固地附着在物体表面。在足丝中发挥主要粘附作用的被认为是贻贝足蛋白(mussel foot proteins,mfps)。贻贝足蛋白强大的粘附力使贻贝在水中具有持久的粘附强度,以及在几乎所有类型的有机和无机材料上同时保持柔韧性和弹性。贻贝蛋白优异的性能使其在蛋白质组成、粘附机制的推测以及仿贻贝足蛋白粘附材料方面受到了很多关注。
3.为了进一步研究贻贝蛋白,必须获取一定量的蛋白。采用直接提取的办法,提取1g贻贝足蛋白(mfp-1和mfp-2混合物)需要数万个贻贝,高昂的成本限制了贻贝的研究。采用基因工程法使贻贝蛋白进行异源表达是现在研究的热门方向。mfp-1是第一个被鉴定的蛋白,尝试对其重组生产。mfp-1部分十肽重复序列(6-20)在大肠杆菌中高效表达,6个十肽重复以包涵体形式表达,20个十肽重复以可溶和不可溶形式表达。mfp-3富含dopa,为关键粘附蛋白,拥有30-35种不同的变体。来源于m.galloprovincialis的mgfp-3a和mgfp-5的cdna序列先后被鉴定,根据大肠杆菌的密码子使用偏好对密码子进行优化后,重组mgfp-3a的产量从0.8mg/l提高到47mg/l(一半表达为包涵体);mgfp-5的产量从2.6mg/l提高到50mg/l。本课题组在之前的研究中实现了m.galloprovincialis中的另一种变体3b(mgfp-3b)在大肠杆菌中的重组表达,简单的纯化步骤后产量为~51mg/l。但mgfp-3b与mgfp-3a表达相似,仍有一半的蛋白为包涵体蛋白。
4.大肠杆菌是基因工程领域被最常用的宿主,培养简单,菌体生产快,容易操作,因此被广泛应用于表达重组异源蛋白。但是使用重组dna技术在大肠杆菌表达目的蛋白时,由于缺少用于蛋白折叠的关键元件,导致折叠效率较低,重组蛋白容易聚集生成没有生物活性的包涵体,需要经过复杂的变性复性过程才能使蛋白重新折叠变成功能蛋白。同时异源表达生产目的蛋白的表达量也很低。目前已经有较多方法来解决包涵体表达的问题。与标签蛋白融合表达,与分子伴侣共表达,更换启动子或者rbs序列,降低表达温度等均被尝试用来解决上述问题。
技术实现要素:
5.为了克服现有技术的不足,本发明提供一种双顺反子翻译偶联表达载体,进一步提出了一种共表达贻贝蛋白mgfp-3b与分子伴侣sumo和/或trxa的基因工程菌的构建方法。
6.本发明还要解决的技术问题在于提供一种促进贻贝蛋白mgfp-3b可溶性表达的发酵培养基和发酵工艺。
7.综上所述,本研究旨在探寻一种高效表达可溶性贻贝蛋白的方法。在大肠杆菌中
共表达分子伴侣可以协助外源基因的表达,在贻贝蛋白的研究中还未有利用分子伴侣提高可溶性表达的方法。
8.本发明中,m.galloprovincialis中的mfp-3变体b(mgfp-3b)在e.coli中重组表达,分子量仅有12kda。通过基因工程手段,将分子伴侣sumo和/或trxa基因与贻贝蛋白mgfp-3b在表达载体pet-28a(+)上构建成双顺反子翻译偶联表达载体并转入e.coli bl21(de3)中进行表达,实现了60-70%目的蛋白以可溶性形式;表达通过不断优化发酵培养基和发酵条件,可溶性贻贝蛋白产量可达200-300mg/l。
9.为了实现上述目的,本发明提供了一种双顺反子翻译偶联表达载体,所述表达载体在pet-28a(+)质粒上插入分子伴侣、新的核糖体结合位点和目的蛋白基因,构建成双顺反子翻译偶联表达载体,其中,分子伴侣基因位于插入的新的核糖体结合位点上游,贻贝蛋白基因位于插入的新的核糖体结合位点下游,所述分子伴侣为smuo和trxa中的任意一种或两种的组合。
10.在一个具体的实施方式中,所述目的蛋白基因为贻贝蛋白mgfp-3b,其核苷酸序列如seq id no:1所示。
11.本发明进一步提出了上述的双顺反子翻译偶联表达载体的构建方法,包括如下步骤:
12.(1)对贻贝蛋白mgfp-3b、分子伴侣sumo、trxa和sumo-trxa基因编码区的克隆,其中在分子伴侣的下游引物和贻贝蛋白的上游引物均引入一个额外的核糖体结合位点;当贻贝蛋白与分子伴侣片段连接时,新引入的核糖体结合位点充当连接肽避免两者直接融合表达,同时双顺反子结构也避免了融合表达而带来的下游分子伴侣酶切操作,其中,新引入的核糖体结合位点独立于原质粒中的核糖体结合位点且与原质粒中的核糖体结合位点序列不同。
13.(2)贻贝蛋白mgfp-3b分别与分子伴侣sumo、trxa和sumo-trxa进行连接并转入质粒pet-28a(+)中构建成表达载体pet-28a(+)-sumo-mgfp-3b、pet-28a(+)-trxa-mgfp-3b和pet-28a(+)-sumo-trxa-mgfp-3b。
14.其中,所述编码基因sumo的核苷酸序列如序列表3所示,其氨基酸序列如序列表4所示;所述编码基因trxa的核苷酸序列如序列表5所示,其氨基酸序列如序列表6所示。
15.本发明进一步提出了一种重组基因工程菌株,所述重组基因工程菌株通过将上述双顺反子翻译偶联表达载体转入表达宿主e.coli bl21(de3)中得到。
16.本发明进一步提出了上述双顺反子翻译偶联表达载体或重组基因工程菌株在制备可溶性贻贝蛋白上的应用。
17.具体地,本发明提出了一种可溶性贻贝蛋白的制备方法,该方法为摇瓶发酵制备贻贝蛋白的方法,具体包括如下步骤:
18.(1)将所述重组基因工程菌株经平板活化后接种至装有液体培养基的摇瓶中,35-40℃,180-220rpm培养8-12h用作种子液;
19.(2)将步骤(1)中的种子液按1-10%的接种量接种至装有液体培养基的挡板摇瓶中,35-40℃,180-220rpm培养,在od
600
达到2~3时加入诱导剂,诱导剂终浓度0.1-1mm,继续培养6-10h结束。
20.本发明还提供了一种发酵罐发酵制备贻贝蛋白的方法,包括如下步骤:
21.(1)将所述重组基因工程菌株经平板活化后接种至装有液体培养基的摇瓶中,35-40℃,180-220rpm培养8-12h用作一级种子液;
22.(2)将步骤(1)中的一级种子液按1-10%的接种量接种至装有液体培养基的挡板摇瓶中,35-40℃,180-220rpm培养至od
600
达到5~6用作二级种子液;
23.(3)二级种子液按1-10%的接种量接种至发酵罐中进行发酵培养,发酵罐中装有发酵罐体积的40%-60%的初始发酵培养基,初始条件为ph 7.0
±
0.5,发酵过程中用浓度10-25%的氨水自动调节ph,温度35-40℃,通过调节转速和通气,使得溶氧保持在20-40%;
24.(4)当初始碳源耗尽后,ph和do会持续上升,当ph高于7.5,溶氧超过40%时,此时加入补料培养基,维持菌体生长,在发酵过程中每隔两小时取样测定od
600
,当od
600
在50-70时,开始加入诱导剂iptg进行诱导表达,诱导剂终浓度0.1-1mm,菌体缓慢生长或者产量不再提升时,则诱导结束。
25.具体地,如果ph高于7.5,则补料速度提高至30ml/h左右;如果ph低于6.5,则降低补料速度至20ml/h左右。维持菌体生长,在发酵过程中每隔两小时取样测定od
600
,当od
600
在50-70时,开始加入诱导剂iptg进行诱导表达,诱导剂终浓度0.1-1mm,菌体缓慢生长或者产量不再提升时,则诱导结束。
26.其中,所述液体培养基配方为:10-20g/l蛋白胨、5-10g/l酵母粉、10-20g/l氯化钠和25-50μg/ml硫酸卡那霉素。
27.所述初始发酵培养基配方为:酵母粉10-40g/l、蛋白胨10-40g/l、葡萄糖5-20g/l、硫酸铵5-10g/l、磷酸二氢钾1-5g/l、无水硫酸镁0.5-5g/l和5-50μg/ml硫酸卡那霉素;所述补料培养基配方为:甘油400-600g/l和酵母粉100-400g/l。
28.所述补料培养基的添加方式为一次性补料、分批补料或者流加补料中的任意一种。
29.有益效果:本发明建立了一个更加简便高效地表达具有生物活性的可溶性贻贝蛋白的方法,避免了包涵体贻贝蛋白需要经过繁琐的、低效的复性工艺的过程。现有技术研究中使用单一分子伴侣较多,有时候效果并不显著。本研究考察单一分子伴侣sumo和trxa与分子伴侣组合sumo-trxa对可溶性表达水平的影响。sumo与trxa均是小分子促溶标签,避免了分子伴侣过大导致目的蛋白表达减弱的情况,以单独插入的核糖体结合位点作为连接肽,使上游的分子伴侣与下游的目的蛋白协同作用,实现了贻贝蛋白60-70%的可溶性表达。分子伴侣sumo和trxa分别提高了12.52%和10.99%的可溶性表达水平;而分子伴侣组合sumo-trxa提高了18.07%。尽管有大量借助分子伴侣提高可溶性表达的研究已经开展,但大多是采用融合表达来生产目的蛋白。在后续的纯化工作中需要切除分子伴侣以免影响目的蛋白的功能。而切除分子伴侣所需的蛋白酶不仅价格昂贵而且特异性不高,导致切除效率较低。除此之外,过多的纯化步骤会使目的蛋白的回收率降低。本研究构建的双顺反子偶联偶联表达载体,在保证分子伴侣有效发挥作用的同时,避免了融合表达带来的下游分子伴侣酶切操作,简化了工艺,降低了成本。本发明的提供的发酵培养基及发酵工艺,使得贻贝蛋白mgfp-3b的od
600
达到120-150,可溶性贻贝蛋白产量可达200-300mg/l,相比于简单的摇瓶发酵,生物量和产量均提高了数倍。本发明制备的贻贝蛋白可在医药、器械等领域得到广泛应用。
附图说明
30.下面结合附图和具体实施方式对本发明做更进一步的具体说明,本发明的上述和/或其他方面的优点将会变得更加清楚。
31.图1为双顺反子翻译偶联表达载体构建框架图;
32.图2为贻贝蛋白mgfp-3b与分子伴侣sumo、trxa基因编码区的克隆;m:dna标准分子量;1:实施例1步骤1中mgfp-3b的pcr扩增产物,250bp下方;2:实施例2和实施例3步骤1中mgfp-3b的pcr扩增产物,250bp下方;3:实施例1步骤2中sumo的pcr扩增产物,300bp左右;4:实施例3步骤4中sumo的pcr扩增产物,300bp左右;5:实施例2步骤2中trxa的pcr扩增产物,300bp上方;6:实施例3步骤2中trxa的pcr扩增产物,300bp上方;
33.图3为贻贝蛋白与分子伴侣的连接:m:dna标准分子量;1:sumo-trxa-mgfp-3b的pcr扩增产物,900bp下方;2:sumo-mgfp-3b的pcr扩增产物,550bp左右;3:trxa-mgfp-3b的pcr扩增产物,600bp下方;
34.图4为共表达菌株的sds-page电泳分析;m:蛋白质标准分子量;
35.sumo-mgfp-3b表示mgfp-3b与sumo共表达;trxa-mgfp-3b表示mgfp-3b与trxa共表达;sumo-trxa-mgfp-3b表示与mgfp-3b与sumo-trxa共表达;s为可溶性表达,is为不溶性包涵体表达。mgfp-3b的分子量约为12kda左右;
36.图5为共表达菌株贻贝蛋白的表达量分析,soluble fraction为可溶性表达产量,insoluble fraction为包涵体表达产量;
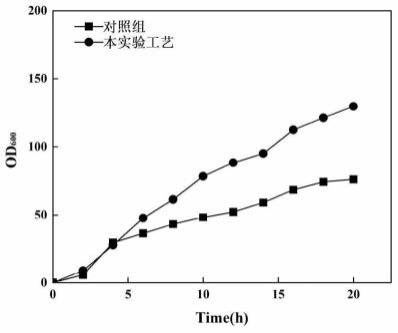
37.图6为使用本实验所述的发酵培养基及发酵工艺发酵贻贝蛋白mgfp-3b与对照组发酵贻贝蛋白mgfp-3b的生长曲线图。
具体实施方式
38.下述实施例中,总蛋白浓度采用bradford蛋白浓度测定试剂盒进行检测所得;贻贝蛋白占总蛋白比例,是采用gel-pro analyzer(media cybernatics,version 4.0.0.4)软件进行灰度扫描计算所得;贻贝蛋白产量是用目标蛋白占比与总蛋白浓度相乘所得;可溶性贻贝蛋白占总贻贝蛋白比例,是采用gel-pro analyzer(media cybernatics,version4.0.0.4)软件进行灰度扫描计算所得。可溶性贻贝蛋白产量是用可溶性蛋白占比与总贻贝蛋白蛋白浓度相乘所得。
39.实施例1生产可溶性贻贝蛋白mgfp-3b的基因工程菌的构建。
40.1.sumo-mgfp-3b基因片段的获得
41.(1)根据mgfp-3b的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,其中上游引物中引入新的核糖体结合位点(斜体表示),
42.命名为rbs2,设计的引物为:
43.上游引物3b-f1:
[0044]5’‑
ggtggtggtaaggagggctaatgaacaacatcagcgttgc-3’[0045]
下游引物3b-r:
[0046]5’‑
tggtggtggtggtgctcgagatagttatatttacgacgac-3’[0047]
pcr反应体系为25μl:mgfp-3b基因组模板dna2μl,上下游引物各2μl,2
×
phanta max master mix 12.5μl,ddh2o 6.5μl。2
×
phanta max master mix购自诺唯赞(中国南
京)。pcr反应条件为94℃预变性5min,然后进行94℃30s,60℃30s,72℃15s,共30个循环,最后72℃延伸10min。pcr产物经1%琼脂糖凝胶,结果表明,在250bp下面有特异性条带,切胶回收mgfp-3b基因片段。
[0048]
(2)根据sumo的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,其中下游引物中引入新的核糖体结合位点(斜体表示),设计的引物为:
[0049]
上游引物sumo-f:
[0050]5’‑
gaaggagatataccatgggcatgagcgatagtgaagttaa-3’[0051]
下游引物sumo-r1:
[0052]5’‑
gttcattagccctccttaccaccaccaatctgttcacgat-3’[0053]
pcr反应体系及条件同步骤1(1)。pcr产物经1%琼脂糖凝胶,结果表明,在300bp处有特异性条带,切胶回收sumo。
[0054]
(3)sumo与mgfp-3b的基因片段通过pcr进行连接,上游引物sumo-f,下游引物3b-r。pcr反应体系同步骤1(1)。pcr反应条件为94℃预变性5min,然后进行94℃30s,60℃30s,72℃20s,共30个循环,最后72℃延伸10min。pcr产物经1%琼脂糖凝胶,结果表明,在550bp处有特异性条带,切胶回收得到sumo-mgfp-3b基因片段。
[0055]
2.trxa-mgfp-3b基因片段的获得
[0056]
(1)根据mgfp-3b的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,其中上游引物中引入新的核糖体结合位点(斜体),命名为rbs2,设计的引物为:
[0057]
上游引物3b-f2:
[0058]5’‑
ctggccggtaaggagggctaatgaacaacatcagcgttgc-3’[0059]
下游引物3b-r:
[0060]5’‑
tggtggtggtggtgctcgagatagttatatttacgacgac-3’[0061]
pcr反应体系及条件同步骤1(1)。pcr产物经1%琼脂糖凝胶,结果表明,在250bp下面有特异性条带,切胶回收mgfp-3b基因片段。
[0062]
(2)根据trxa的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,其中下游引物中引入新的核糖体结合位点rbs2,设计的引物为:
[0063]
上游引物trxa-f1:
[0064]5’‑
gaaggagatataccatgggcatgagcgataaaatcatcca-3’[0065]
下游引物trxa-r:
[0066]5’‑
gttcattagccctccttaccggccagattggcatccagaa-3’[0067]
pcr反应体系及条件同步骤1(1)。pcr产物经1%琼脂糖凝胶,结果表明,在300bp上方有特异性条带,切胶回收trxa。
[0068]
(3)trxa与mgfp-3b的基因片段通过pcr进行连接,上游引物trxa-f1,下游引物3b-r。pcr反应体系及条件同步骤1(3)。pcr产物经1%琼脂糖凝胶,结果表明,在600bp下方有特异性条带,切胶回收得到trxa-mgfp-3b基因片段。
[0069]
3.sumo-trxa-mgfp-3b基因片段的获得
[0070]
(1)mgfp-3b基因片段的获得同步骤2(1)。
[0071]
(2)根据trxa的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,其中下游引物中引入新的核糖体结合位点rbs2,设计的引物为:
[0072]
上游引物trxa-f2:
[0073]5’‑
atcgtgaacagattggtggtatgagcgataaaatcatcca-3’[0074]
下游引物trxa-r:
[0075]5’‑
gttcattagccctccttaccggccagattggcatccagaa-3’[0076]
pcr反应体系及条件同步骤1(1)。pcr产物经1%琼脂糖凝胶,结果表明,在300bp上方有特异性条带,切胶回收trxa。
[0077]
(3)trxa与mgfp-3b的基因片段通过pcr进行连接,上游引物trxa-f2,下游引物3b-r。pcr反应体系及条件同步骤1(3)。pcr产物经1%琼脂糖凝胶,结果表明,在600bp下方有特异性条带,切胶回收得到trxa-mgfp-3b基因片段。
[0078]
(4)根据sumo的编码区序列设计上下游引物,上游自起始密码子处设计,下游引物至终止密码子处设计,设计的引物为:
[0079]
上游引物sumo-f:
[0080]5’‑
gaaggagatataccatgggcatgagcgatagtgaagttaa-3’[0081]
下游引物sumo-r2:
[0082]5’‑
tggatgattttatcgctcataccaccaatctgttcacgat-3’[0083]
pcr反应体系及条件同步骤1(1)。pcr产物经1%琼脂糖凝胶,结果表明,在300bp处有特异性条带,切胶回收sumo。
[0084]
(5)smuo与trxa-mgfp-3b的基因片段通过pcr进行连接,上游引物sumo-f,下游引物3b-r。pcr反应体系同步骤1(3)。pcr反应条件为94℃预变性5min,然后进行94℃30s,60℃30s,72℃25s,共30个循环,最后72℃延伸10min。pcr产物经1%琼脂糖凝胶,结果表明,在900bp下方有特异性条带,切胶回收得到smuo-trxa-mgfp-3b基因片段。
[0085]
4.双顺反子翻译偶联表达载体的构建
[0086]
(1)少量提取pet-28a空质粒。pcr回收产物(sumo-mgfp-3b、trxa-mgfp-3b、sumo-trxa-mgfp-3b)与质粒pet-28a经限制性内切酶nco i和xho i双酶切,酶切产物经1%琼脂糖凝胶纯化后回收备用。
[0087]
(2)酶切产物sumo-mgfp-3b、trxa-mgfp-3b、sumo-trxa-mgfp-3b与线性化的pet-28a质粒连接。反应体系为:5μl线性化载体,3μl插入片段,2μl exnase ii,4μl 5
×
ce ii buffer,6μl ddh2o。exnase ii与5
×
ce ii buffer均购自诺唯赞(中国南京)。20μl反应体系冰浴反应30分钟后,加入100μl大肠杆菌dh5α感受态菌液中继续冰浴30分钟。42℃热激60~90秒,加5倍体积lb液体培养基(无抗性)恢复培养1小时,之后取适当体积涂平板(具有卡那抗性),于37℃培养12小时。筛选单克隆,培养并提取质粒测序,最终得到相应的双顺反子表达载体pet-28a(+)-sumo-mgfp-3b、pet-28a(+)-trxa-mgfp-3b、pet-28a(+)-sumo-trxa-mgfp-3b。重组质粒转入大肠杆菌bl21(de3)中进行表达。
[0088]
实施例2摇瓶发酵制备可溶性贻贝蛋白。
[0089]
将实施例1中构建成功的三株重组基因工程菌sumo-mgfp-3b、trxa-mgfp-3b、sumo-trxa-mgfp-3b在平板划线活化。挑取平板上长出的单菌落,接种于装有5ml液体培养基的摇瓶中在37℃,200rpm振荡过夜培养10h。按10%的接种量转接于装有200ml液体培养
基的1l挡板摇瓶中,在37℃,200rpm培养至od600为2~3,添加iptg至终浓度为1mm/l,在37℃,200rpm继续培养6h。培养结束后取菌液进行离心破碎,然后采用sds-page电泳确定重组蛋白的表达。表达结果如下表1所示。
[0090]
表1
[0091][0092]
在实施例2中,本研究构建的sumo-mgfp-3b、trxa-mgfp-3b、sumo-trxa-mgfp-3b诱导后生成的mgfp-3b在sds-page凝胶上的表观分子量约12kda,与单独表达的mgfp-3b相对分子质量一致,表明贻贝蛋白成功实现异源表达。单独表达的mgfp-3b中仅有43.8%的目的蛋白以可溶性的形式存在;而与分子伴侣sumo、trxa和sumo-trxa进行共表达时,目的蛋白的可溶性比例均有所提高,分别达到了56.32%、54.79%和61.87%。相比于sumo提高了12.52%和trxa提高了10.99%,sumo-trxa的提升效果最好,提高了18.07%,证明本发明所选的分子伴侣确实能有效提高目的蛋白的可溶性,而且分子伴侣组合效果要远优于单一分子伴侣。
[0093]
实施例3发酵罐发酵制备可溶性贻贝蛋白。
[0094]
(1)所述生产可溶性贻贝蛋白的基因工程菌sumo-trxa-mgfp-3b经平板活化后接种至装有5ml液体培养基的摇瓶中,37℃,200rpm培养10h用作一级种子液。
[0095]
(2)将步骤(1)中的一级种子液按10%的接种量接种至装有200ml液体培养基的挡板摇瓶中,37℃,200rpm培养至od
600
达到5~6用作二级种子液。
[0096]
(3)二级种子液按10%的接种量接种至5l发酵罐中进行发酵培养。发酵罐中装有2l初始发酵培养基,初始条件为ph 7.0
±
0.5,发酵过程中用浓度25%的氨水自动调节ph,温度37℃,通过调节转速和通气,使得溶氧保持在20%。
[0097]
(4)当初始碳源耗尽后,ph和do会持续上升,高于设定值。此时加入补料培养基,维持菌体生长。补料速度维持在20-30ml/h,根据ph来调整补料速度。如果ph高于7.5,则补料速度提高至30ml/h左右;如果ph低于6.5,则降低补料速度至20ml/h左右。在发酵过程中每隔两小时取样测定od
600
,当od
600
在60时,开始加入诱导剂iptg进行诱导表达,诱导剂终浓度0.2mm。此批次诱导持续12h。
[0098]
(5)发酵液进行离心收集菌体,破碎菌体收集上清,用于后续纯化。
[0099]
(6)破碎液的上清和沉淀进行sds-page电泳,考察发酵表达水平,以及可溶性蛋白表达量。
[0100]
其中,液体培养基配方为:液体培养基配方为:10g/l蛋白胨、5g/l酵母粉、10g/l氯
化钠和25μg/ml硫酸卡那霉素。
[0101]
初始发酵培养基配方为:酵母粉20g/l、蛋白胨10g/l、葡萄糖15g/l、硫酸铵5g/l、磷酸二氢钾2g/l、无水硫酸镁1g/l和25μg/ml硫酸卡那霉素;补料培养基配方为:甘油400g/l和酵母粉200g/l。
[0102]
实施例4发酵罐发酵制备可溶性贻贝蛋白(作为对照例)。
[0103]
(1)(2)(3)同实施例2。
[0104]
(4)当初始碳源耗尽后,ph和do会持续上升,高于设定值。此时加入补料培养基,维持菌体生长。补料速度维持在20-30ml/h,根据ph来调整补料速度。如果ph高于7.5,则补料速度提高至30ml/h左右;如果ph低于6.5,则降低补料速度至20ml/h左右。在发酵过程中每隔两小时取样测定od
600
,当od
600
在40时,开始加入诱导剂iptg进行诱导表达,诱导剂终浓度0.2mm。此批次诱导持续12h。
[0105]
步骤4所涉及的补料培养基配方为:葡萄糖600g/l。
[0106]
(5)发酵液离心收集菌体,破碎菌体收集上清,用于后续纯化。
[0107]
(6)破碎液的上清和沉淀进行sds-page电泳,考察发酵表达水平,以及可溶性蛋白表达量。
[0108]
实验结果如下表所示:
[0109][0110]
其中:最高od600为整个发酵过程中的最高生物量。
[0111]
从上面的结果可以看出:本发明实施例3及实施例4由于采用了本发明的发酵培养基配方与工艺,实现了60-70%目标蛋白的可溶性表达。实施例3可溶性表达量达到224.35mg/l;实施例4目标蛋白的可溶性表达量明显低于实施例3。
[0112]
本发明提供了一种促进重组贻贝蛋白mgfp-3b可溶性表达的思路及方法,具体实现该技术方案的方法和途径很多,以上所述仅是本发明的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也应视为本发明的保护范围。本实施例中未明确的各组成部分均可用现有技术加以实现。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1