一组烟草NtDTRG基因序列及其在开发防干旱烟草品种中的应用的制作方法
一组烟草ntdtrg基因序列及其在开发防干旱烟草品种中的应用
技术领域
1.本发明涉及烟草基因工程研究技术领域,具体涉及一组烟草ntdtrg基因序列及其在开发防干旱烟草品种中的应用。
背景技术:
2.干旱相关基因dtrg(drought tolerance related gene,dtrg)在植物生长发育及抗逆调控过程中发挥着重要作用。烟草中管控抗干旱能力的就是ntdtrg基因,其核苷酸序列长度为990bp,具体如seq id no.1所示:
3.atggctataaaaatgcttctttttctatgcatttccgccttattaagtacttcctctttatcccagcgccattcgccgctaaatactgctttttaccgcaaatcatgcccaagatttgagcagataatggaagaaaccactacaaacaaacagatcacttctcctactaccgccgccgccaccctccgcctcttcttccacgactgcttcgtcggcggctgtgacgcctccgtacttgtctcctccactcccttcaacaaagccgagcgtgacgccgaaatcaacctttccctccccggcgatggcttcgacgtggtcgtacgcgccaagaccgcgcttgaactttcctgtccaggtatagtctcttgctccgacattctcgccgtcgctgcccgaaaccttgtcgtccaaactggcggcccgttttatccagttaaattgggccgtaaagattcttttgtatcgaaagcctcattagtagaaggaaatctgccccggcccacaatgccgatggatcaaatcattaagattttcgaatccagaggattttcgattcaagaaatggtagcattatctggagctcacacaattggattttcccactgtaaagagttcaactcggatctttataactacaacaaaacttcacaagctgatccttcgtataatccgaggtttgctcaagctttgaaagatgcttgtagtaattcccagaaagatccaacattatcagtgttcaacgatataatgactcctaacaaattcgataatatgtattatcagaacttgcctaagggtttgggtttattgtcttcagaccgtggtctgttttcagatccgaggacgagaattcacgttgaggaatatgttagagatcagaatgcattttttaaggcgtttgcttcagcaatgcagaagcttagtgaacatggtgttaaaattggtaaacatggtgagatcaggcacaggtgtgacgctttcaacaattga。
4.而ntdtrg基因所编码的蛋白,包括329个氨基酸残基,具体序列为:
5.maikmllflcisallstsslsqrhsplntafyrkscprfeqimeetttnkqitspttaaatlrlffhdcfvggcdasvlvsstpfnkaerdaeinlslpgdgfdvvvraktalelscpgivscsdilavaarnlvvqtggpfypvklgrkdsfvskaslvegnlprptmpmdqiikifesrgfsiqemvalsgahtigfshckefnsdlynynktsqadpsynprfaqalkdacsnsqkdptlsvfndimtpnkfdnmyyqnlpkglgllssdrglfsdprtrihveeyvrdqnaffkafasamqklsehgvkigkhgeirhrcdafnn。
6.为了进一步提高烟草的防干旱能力,对ntdtrg基因的研究是很有必要的,而现有技术针对ntdtrg基因调控研究还比较少,因此,如何通过ntdtrg基因调控获得防干旱烟草,是未来烟草基因研究的重点之一。
技术实现要素:
7.针对现有技术中的上述问题,本发明提供一组烟草ntdtrg基因序列及其在开发防干旱烟草品种中的应用,以填补现有烟草防旱技术的空白,提供更多的防旱烟草品种。
8.本发明采用的技术方案如下:
9.一组烟草ntdtrg基因序列,在如seq id no.1所示烟草ntdtrg基因序列的294~316片段进行人工干预突变,突变后的三种294~316片段序列分别如seq id no.2~4所示。
10.所述的烟草ntdtrg基因序列在开发防干旱烟草品种中的应用。
11.综上所述,相比于现有技术,本发明具有如下优点及益效果:
12.1.本发明获得的烟草ntdtrg基因序列,在烟草抗干旱方面具有显著的效果;
13.2.本发明提供了烟草基因防旱技术的新可能,填补了相应技术领域的空白。
附图说明
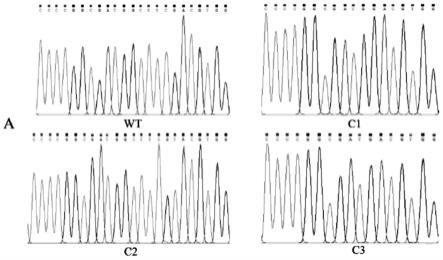
14.图1为烟草ntdtrg突变体检测峰图;
15.图2为过表达植株pcr检测电泳图;
16.图3为过表达烟草与k326中ntdtrg基因的相对表达量柱状图;
17.图4为western blot检测ntdtrg过表达植株与k326结果图,其中,左图为烟草总蛋白提取sds-page图,右图为western blot检测图;
18.图5为烟草ntdtrg基因表达量分析对比图;
19.图6为ntdtrg基因的不同组织表达模式分析结果柱状图;
20.图7为peg-6000处理下ntdtrg基因表达模式分析结果柱状图;
21.图8为peg-6000处理下烟草种子萌发试验实物图;
22.图9为peg-6000处理下烟草种子萌发率柱状图;
23.图10为peg-6000处理下抗旱及aba信号相关基因表达分析结果柱状图;
24.图11为aba处理下烟草种子萌发试验实物图;
25.图12为aba处理下烟草种子萌发率柱状图;
26.图13为aba处理下抗旱及aba信号相关基因表达分析结果柱状图;
27.图14为干旱胁迫下烟草叶片表型分析试验实物图;
28.图15为干旱胁迫下烟草存活率的统计柱状图;
29.图16为干旱胁迫下烟草光合指标测定结果柱状图;
30.图17为干旱胁迫下烟草spad值测定结果柱状图;
31.图18为干旱胁迫下烟草抗氧化酶活性测定结果柱状图;
32.图19为干旱胁迫下烟草渗透调节物质及丙二醛含量测定结果柱状图;
33.图20为干旱胁迫下烟草过氧化氢含量测定结果柱状图;
34.图21为干旱胁迫下烟草的根冠干鲜重测定结果柱状图;
35.图22为干旱胁迫下烟草叶片相对含水量的变化柱状图;
36.图23为抗旱及aba信号相关基因表达模式分析结果柱状图;
37.图24为离体叶片失水表型鉴定实物图;
38.图25为离体叶片失水率的变化折线图;
39.图26为干旱胁迫下气孔开度表型分析图;
40.图27为干旱胁迫下气孔数目及开度分析柱状图;
41.图28为nbt组织染色试验实物图;
42.图29为田间烟草表型实物图;
43.图30为田间烟草光合指标及spad测定结果柱状图;
44.图31为田间烟草生物量测定结果柱状图。
具体实施方式
45.为了使本发明的目的、技术方案及优点更加清楚明白,以下结合实施例对本发明进行进一步详细说明。应当理解,此处所描述的具体实施例仅用以解释本发明,并不用于限定本发明,即所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。
46.实施例1
47.本实施例提供的烟苗分别为k326(野生型,wt)、三个ntdtrg基因突变体(c1、c2、c3)、三个ntdtrg基因过表达体(oe-1、oe-2、oe-3)三组烟苗。
48.其中,三个ntdtrg基因突变体由k326(含ntdtrg基因)的烟草通过基因编辑技术而获得,c1、c2、c3是在如seq id no.1所示烟草ntdtrg基因序列的294~316片段进行人工干预突变:
49.原始含ntdtrg基因294~316片段wt:ccccggcgatggcttcgacgtgg;
50.c1该片段变为:ccccggcgatggacgtgg;
51.c2该片段变为:ccccggcgatggcttcgtacgtgg;
52.c3该片段变为:ccccggcgatgacgtgg。
53.1、先分别通过dna、rna水平对三组烟苗进行鉴定,其中通过dna水平鉴定证实三个ntdtrg基因突变体,通过rna水平鉴定证实三个ntdtrg基因过表达体。
54.采用漂浮育苗法进行供试烟苗的培养,待烟苗长至3叶1心时,选取长势健壮的烟苗转移至黑色小盆中,每个小盆中均移栽4株烟苗,最后将其将置于温度为25℃,16h光/8h暗的生长室中培养。
55.dna水平鉴定:
56.(1)首先使用试剂盒分别提取三组材料的基因组dna;
57.(2)以烟草基因组dna为模板,按照以下体系(表1)和pcr程序扩增目的片段。反应体系为(10μl):
58.表1 pcr反应体系
59.反应体系成分反应体积上游引物0.5μl下游引物0.5μldna模板1μl2
×
taqpcr master mix5μlddh2o3μl
60.反应程序如表2:
61.表2 pcr反应程序
[0062][0063]
引物如表3:
[0064]
表3引物序列
[0065]
引物名称引物序列seq id nontdtrg-f5'-gcatttccgccttattaagtactt-3'5ntdtrg-r5'-cgaaggatcagcttgtgaagtt-3'6kn-f5'-atggtctttgcctgctctgac-3'7kn-r5'-ttaagttcatacggaaagaga-3'8
[0066]
(3)pcr产物送至公司(上海生工生物工程有限公司)测序,利用chromas软件对测序结果进行分析。
[0067]
如图1所示,通过dna测序结果表明,仅c1、c2、c3在靶位点处共检测到3种引起开放阅读框改变的突变方式,分别为缺失5个碱基、增加1个t碱基、缺失6个碱基,与人工突变片段吻合,证明c1、c2、c3确系三个ntdtrg基因突变体。
[0068]
rna水平鉴定:
[0069]
(1)取未处理及自然干旱处理14d时的烟叶,液氮速冻后置于-80℃备用,采用trizol法提取rna。
[0070]
(2)将所有直接接触样品的实验物品用液氮预冷。
[0071]
(3)将烟叶置于研钵中多次研磨至粉末状。
[0072]
(4)将0.1g粉末转移至1.5ml离心管中,加入1ml trizol后迅速斡旋混匀,冰上静置5min。
[0073]
(5)12000rpm,4℃离心10min,取上清至新的1.5ml离心管中。
[0074]
(6)加入200μl氯仿,剧烈震荡混匀,冰上静置5min。
[0075]
(7)12000rpm,4℃离心15min,取最上层无色水样层至新的1.5ml离心管中。
[0076]
(8)加入等体积异丙醇,上下颠倒混匀,冰上静置20min。
[0077]
(9)12000rpm,4℃离心8min,弃上清。
[0078]
(10)加入1ml 75%酒精,上下颠倒3次。
[0079]
(11)12000rpm,4℃离心3min,弃上清。
[0080]
(12)超净台中晾干沉淀后,加入30-100μl无菌无酶超纯水溶解沉淀,-80℃保存备用。
[0081]
cdna第一链合成:使用takara公司的1st strand cdna synthesis kit试剂盒合成cdna,方法参照说明书。
[0082]
目的片段扩增:以烟草cdna为模板,按照以下体系(如表4所示)和pcr程序扩增目的片段,表中上、下游引物序列如表3所示。反应体系为(10μl):
[0083]
表4 pcr反应体系
[0084]
反应体系成分反应体积上游引物0.5μl下游引物0.5μlcdna模板1μl2
×
taqpcr master mix5μlddh2o3μl
[0085]
反应程序如表5:
[0086]
表5 pcr反应程序
[0087][0088]
实时荧光定量pcr:反应体系如表6:
[0089]
表6 pcr反应体系
[0090]
反应体系成分反应体积上游引物0.3μl下游引物0.3μlcdna模板1μl2
×
sybr green5μlddh2o3.4μl
[0091]
qrt-pcr表达检测:实时荧光定量pcr的引物序列,采用premier5.0设计,如表7所示:
[0092]
表7引物序列
[0093]
引物名称引物序列seq id nontdtrg-qf5'-ggtttgggtttattgtctt-3'9ntdtrg-qr5'-acctgtgcctgatctcac-3'1018s-f5'-cctacgctctgtatacattagc-3'1118s-r5'-gtgttgagtcaaattaagcccc-3'12
[0094]
在定量仪cfx96
tm
real-time system中进行扩增检测,运行程序如表8:
[0095]
表8扩增检测运行程序
[0096][0097]
结果分析:利用bio-rad cfx manager软件,根据2
‑△△
ct
法进行结算分析。
[0098]
本实施例分别以转化烟草、野生型(k326)、ddh2o为pcr扩增模板,kn引物进行pcr鉴定。如图2所示,oe-1、oe-2、oe-3三个转化植株中均扩增出一条大小500bp左右的条带,与卡那基因(539bp)的序列长度基本一致,以野生型和ddh2o为模板的体系中无条带,可初步证实三个转化植株为ntdtrg基因过表达株系。
[0099]
为了进一步证明上述鉴定结果,便分别在rna和蛋白水平上利用实时荧光定量pcr(图3)及western blot(图4)进行材料中ntdtrg基因的表达量分析。如图3所示,oe-1、oe-2、oe-3三个过表达株系中ntdtrg基因的表达量均显著高于野生型。然后提取烟草总蛋白进行凝胶电泳检测,如图4所示,每个泳道中都有清晰的蛋白条带,表明该烟草总蛋白可以用于后续实验,如免疫沉淀、western blot等。使用ntdtrg蛋白抗体进行western blot检测,结果表明三个oe株系中的ntdtrg蛋白表达量均比野生型高。该部分结果证实了oe-1、oe-2、oe-3确为ntdtrg基因过表达株系。
[0100]
为了分别检测突变体、野生型及过表达中ntdtrg基因在转录水平上的变化,使用半定量rt-pcr方法进行其表达水平的分析。由图5可得,ntdtrg基因在三个过表达株系中的表达量高于野生型,表达量高低顺序依次为:oe-1》oe-2》oe-3》wt。然而,3个突变体株系中ntdtrg基因几乎不表达。
[0101]
2、ntdtrg基因的表达模式分析
[0102]
组织表达模式分析:分别提取野生型根、茎、叶、花组织的rna,进行qrt-pcr定量分析。
[0103]
胁迫诱导表达模式分析:使用10%peg-6000处理野生型材料,分别在0h、3h、6h、12h时间段取样,液氮速冻,存于-80℃,进行qrt-pcr定量分析。
[0104]
通过实时荧光定量pcr检测ntdtrg基因分别在烟草根、茎、叶和花不同组织中的表达情况(图6)。结果如图6所示,该基因在根、茎、叶及花组织中均有表达,而根和叶中的表达量最高,其次为茎,花中的表达较少。该结果表明了,ntdtrg基因具有不同的组织表达特异性。
[0105]
为了确认ntdtrg基因是否响应干旱胁迫诱导,在野生型材料中进行了该基因的表达模式分析(图7)。使用10%peg-6000模拟干旱胁迫,处理烟草幼苗12h后检测ntdtrg的基因表达,发现相比0h,分别在处理3h、6h、12h时,ntdtrg基因的表达呈显著下调趋势。该结果表明了,ntdtrg基因表达会受到干旱胁迫抑制。
[0106]
3、peg及aba处理下烟草种子萌发试验
[0107]
处理方法:(1)本试验采用peg-6000作为渗透剂,模拟干旱胁迫,配制胁迫溶液,质量体积比分别为5%、10%、15%。选取两份材料中成熟度、大小均一致的烟草种子进行萌发试验,0.5%次氯酸钠消毒,超纯水清洗三次,每种材料分别30粒,均匀摆放在培养皿中,培养皿经灭菌处理,并铺放两层滤纸(灭菌)作为发芽床,分为处理组和对照组,使用移液枪分
别注入处理溶液或超纯水,每组试验均设三次重复。放置在光照培养箱内,温度设置为25℃。(2)aba处理:浓度分别为0μm、0.5μm、1μm aba。其他方法同上。
[0108]
测定方法:从播种第3天起,每天观察记录种子萌发情况。以胚根露出种皮为发芽标准,记录每天发芽数,并且拍照观察表型。14天后统计发芽率:
[0109]
发芽率(%)=种子发芽数/供试种子总数
×
100%
[0110]
对同一个处理不同材料之间的发芽率进行差异显著性检验,以分析对比材料之间的差异。
[0111]
peg-6000作为一种大分子物质,无法进入细胞且对细胞没有毒性,但其亲水性强,加入水中后可降低水势,致使植物根系无法吸收比细胞水势还低的水分,造成对细胞的渗透胁迫,故常被用于模拟干旱。分别利用5%、10%、15%peg-6000模拟干旱胁迫处理突变体、野生型及过表达种子,且以h2o处理为对照,观察种子萌发表型及其萌发率(图8、图9)。结果发现,随着peg-6000处理浓度的增加,突变体、野生型及过表达种子的萌发率均呈下降趋势。5%peg-6000处理下,三组材料的萌发率间无差异;10%peg-6000处理下,三个突变体种子的萌发率比野生型和过表达都高,c1萌发率最高,c2和c3其次。15% peg-6000处理下,三个过表达种子均未萌发,但突变体和野生型种子有少数萌发,突变体c3萌发率最高,c1和c2其次。以上结果说明了ntdtrg突变体对peg胁迫的敏感性降低,同时初步推断ntdtrg基因可能在烟草抗旱方面起负调控作用。
[0112]
4、抗旱及aba信号相关基因表达分析
[0113]
分别利用10% peg-6000和1μm aba溶液处理1月龄三组材料的幼苗,在处理0h和6h时进行取样,液氮速冻后,储存于-80℃冰箱,进行抗旱相关基因的qrt-pcr分析,其中所用引物序列如表9:
[0114]
表9引物序列
[0115]
引物名称引物序列seq id nontdreb3-qf5'-acaggggagtgaggaagagga-3'13ntdreb3-qr5'-gcagaagggaaagtgccaag-3'14nterd10c-qf5'-aagaaggttgaagagggtagc-3'15nterd10c-qr5'-tcctctcctatttcttcctcc-3'16ntlea5-qf5'-ttgaatctggggttttggtt-3'17ntlea5-qr5'-ggaagcattgacgagctagg-3'18ntabi3-qf5'-gagtatcagaccatggaatctgc-3'19ntabi3-qf5'-ttccatcgcggagaattg-3'20ntabi5-qf5'-cgcaaaaggcgactaacaa-3'21ntabi5-qr5'-acacatcaagggcaactcaa-3'22ntnced1-qf5'-aagaatggctccgcaagtta-3'23ntnced1-qr5'-gcctagcaattccagagtgg-3'2418s-f5'-cctacgctctgtatacattagc-3'1118s-r5'-gtgttgagtcaaattaagcccc-3'12
[0116]
利用10%peg-6000溶液水培处理烟苗,分别在处理0h和6h时取样,使用实时荧光定量(qrt-pcr)方法检测应激相关基因,包括ntdreb3(脱水应答转录因子)、nterd10c(脱水
诱导早期应答基因)、ntlea5的表达水平,还有aba信号相关基因ntnced1、ntabi3、ntabi5(图10)。结果发现,干旱处理前,6个抗旱及aba信号相关基因表达水平均较低,ntdreb3、ntabi3、ntabi5比nterd10c和ntlea5表达更低。当干旱处理6h后,突变体、野生型及过表达植株均能诱导胁迫相关基因的表达显著上调,但突变体植株中6个基因的表达量明显高于野生型和过表达植株。研究结果表明,ntdtrg基因突变体植株抗旱性的增强伴随着逆境相关基因表达的增加。
[0117]
aba对种子萌发与休眠具有核心调节作用,aba可以促进种子休眠,所以进一步探究了外源aba对突变体、野生型及过表达种子萌发的影响。分别利用0μm、0.5μm、1μm aba对三组材料的种子进行处理,观察表型并统计萌发率(图11、图12)。结果发现,外源aba处理下,三组材料的种子萌发速度都有所减慢,且相比于野生型和过表达,突变体种子的萌发速率更慢。0.5μm及1μmaba处理下突变体种子的萌发率都是显著低于野生型及过表达。
[0118]
利用1μm aba和10%peg溶液水培处理烟苗,分别在处理0h和6h时取样,使用实时荧光定量(qrt-pcr)方法检测应激相关基因(ntdreb3、nterd10c、ntlea、ntnced1、ntabi3、ntabi5)的表达水平(图13)。处理前,6个胁迫相关基因表达水平均较低。在aba处理6h后,突变体、野生型及过表达植株中的胁迫相关基因表达均上调以此来响应aba胁迫,但突变体植株中应激基因的表达丰度增加量显著高于野生型及过表达,c2中ntdreb3、nterd10c和ntabi5基因表达量为最高,c1中ntlea5基因表达最高。研究结果表明,ntdtrg基因突变体植株对aba敏感性增强的同时也伴随着逆境相关基因表达量的增加。
[0119]
5、室内自然干旱试验
[0120]
供试苗的培养:采用漂浮育苗方式进行育苗,一个月后移栽至高9cm,宽7cm的小盆中培养,并且每个小盆中土重均为0.3kg。随机挑选每个株系长势一致的2月龄幼苗各18株(干旱处理9株,对照9株)进行试验。
[0121]
盆栽苗自然干旱处理方法:小盆中每次浇水均定量为150ml,最后一次浇水量为150ml水,50ml营养液。在预试验中,当土壤水分饱和时,随即开始干旱处理,处理开始后便每隔两天测定一次土壤含水量,控制土壤干旱程度。根据预试验测定结果,确定正式实验中,在干旱14d(土壤含水量为13.5%-14.2%)时,进行取样和指标测定;在干旱30d时(土壤含水量为8.7%-9.2%),停止干旱。
[0122]
当植物受到一定程度的干旱胁迫后,因持续缺水,会使植物叶片渐渐失水最终萎焉,严重时甚至死亡。自然干旱处理下突变体、野生型及过表达烟草叶片的表型鉴定如图(图14,图中上中下三幅图分别表示干旱0d、干旱30d以及复水5d)所示,在正常供水条件下,不同烟草材料之间无表型差异。当干旱30d时,三组材料的底部老叶均已明显变黄,野生型和过表达材料的部分老叶已死亡,且新叶也严重萎焉并失绿。然而,突变体材料的新叶部分仅轻度萎焉,几乎未变黄。复水5d后,统计不同材料的存活率(图15),结果表明,三个突变体烟苗全部存活,比野生型和过表达存活数目均多。
[0123]
光合指标及spad值的测定:分别在处理0d,14d和30d后复水5d,进行光合指标的测定。选择使用li-6400光合仪进行光合指标的测定,每次的测定时间选在上午10:00-12:00。每株烟苗所测定的烟叶为中心叶下部的第3-4片。测定时使用仪器自带的光源,设置光源强度为800μmol
·
m-2
s-1
。测定指标分别为净光合速率(pn)、气孔导度(gs)及蒸腾速率(tr)。分别在处理0d和14d时,使用spad-502叶绿素仪测定spad值。
[0124]
净光合速率(pn)的高低能够衡量植物有机物质的累计速度快慢,在逆境研究中,净光合速率越高说明植物能更好的适应当前环境,对逆境的耐受能力更强。另外,气孔导度(gs)和蒸腾速率(tr)是衡量植物叶片保水能力的主要指标之一。分别测定了干旱0d、14d及复水5d的pn、gs和tr三个光合指标(图16)。干旱0d时,三组材料之间的净光合速率均无差异;处理14d时,三个突变体材料的净光合速率是最高的,且在干旱复水后仍然是最高的。干旱0d时,三组材料的气孔导度无差异,而处理14d后均显著降低,突变体材料的降低幅度最大,且为最小。干旱0d时,野生型的蒸腾速率是最高的,c1其次,其他材料间无显著差异;干旱14时,所有材料的蒸腾速率均减慢,其中突变体相较野生型和过表达更慢。以上实验结果说明,干旱胁迫会显著降低烟草的pn、gs和tr,以此减小体内水分流失的速度,确保叶片有较强的保水能力。突变体c1较c2与c3具有更低的气孔导度和蒸腾速率,说明其保水性更强;且c1和c2的净光合速率更快,说明在同一干旱胁迫下,c1和c2相比c3可累计较多的生物有机质,适应能力就强。
[0125]
叶绿体是植物进行光合作用的重要细胞器,而存在于叶绿体内的叶绿素是光反应所必需的催化剂,其含量的多少是判断植物光合能力强弱的一项重要指标。spad值便可反映出叶绿素的相对含量。如图所示(图17),干旱0d时,突变体、野生型及过表达材料间的spad值无显著性差异,但在干旱14d后,spad值均变小,且突变体c1和c2为最高,c3其次。结果表明,水分匮乏时,叶肉细胞中的叶绿素合成就会受阻。
[0126]
抗氧化酶活性的测定:超氧化物歧化酶(sod)采用氮蓝四唑光化学还原法;过氧化物酶(pod)愈创木酚法、过氧化氢酶(cat)动力学扫描法。
[0127]
超氧化物歧化酶(sod)、过氧化物酶(pod)和过氧化氢酶(cat)均为植物活性氧清除机制中的重要抗氧化酶,在植物抗逆过程中有重要作用,其酶活越强说明植物的抗氧化能力就越强。如图所示(图18),干旱胁迫下,突变体、野生型和过表达材料的三个抗氧化酶活性均显著增强。突变体中的sod、pod、cat酶活均明显强于野生型和过表达,其中突变体c1的三个酶活均为最高,c2的pod和cat酶活也是最高的。该结果表明,干旱胁迫会引起烟草抗氧化酶活性的变化,从而激活植物酶清除系统及时响应逆境胁迫。
[0128]
渗透调节物质含量的测定:脯氨酸、可溶性蛋白含量的测定;
[0129]
过氧化氢及丙二醛含量的测定。
[0130]
脯氨酸(pro)和可溶性蛋白(sp)为植物体内的两大渗透调节物质,干旱胁迫下,它们可平衡原生质与外界环境之间的渗透势,以维持细胞正常生长。由图可得(图19),正常生长下突变体、野生型及过表达材料的脯氨酸及可溶性蛋白含量间无差异,但在干旱复水后两个物质含量均有了显著差异。突变体中的脯氨酸及可溶性蛋白含量均高于野生型和过表达。突变体c2的脯氨酸含量最高,c1和c3之间无差异;c3的可溶性蛋白含量最高,c1和c2之间无差异。
[0131]
丙二醛(mda)含量的多少是一项衡量植物细胞膜受损程度及膜脂过氧化强弱的重要生理指标。分别测定三组材料正常生长及干旱复水后这两个阶段的mda含量(图19),研究发现对照中不同材料的mda含量并无显著差异,而干旱复水后发生了明显变化,三个过表达材料中的mda含量是最多的,其次为野生型,突变体材料中较少,c2是最少的。上述结果说明,经干旱胁迫后,突变体材料的细胞膜受损较轻微,膜脂过氧化反应弱。
[0132]
过氧化氢(h2o2)为活性氧成分中的一种,过量的h2o2会使蛋白质和脂质发生氧化,
植物细胞受损。干旱胁迫下突变体、野生型及过表达三组材料中h2o2含量的变化表明(图20),在正常生长条件下,所有材料中的h2o2含量均无显著差异。而干旱14d后,不同材料中的h2o2含量都呈上升趋势,但在突变体中的含量显著低于野生型与过表达。以上结果说明,相比野生型与过表达材料,突变体中因水分胁迫引起所产生的ros积累量较少,细胞被氧化程度更轻,表型出较强的耐旱性。
[0133]
生物量的测定:自然干旱,处理30d后复水5d时,将处理烟苗整株取样,用清水清洗干净,并擦干表面水分,根冠分别用电子天平称量鲜重,并用直尺测量根长,烟叶以备下述实验使用。
[0134]
叶片相对含水量的测定:将不同植株的叶片采下后立即称取鲜重(fw),然后将叶片浸于蒸馏水中,4℃下暗处放置24h吸水饱和,取出,吸水纸擦干表面水分,称量叶片饱和重(tw),最后将叶片放置在80℃烘箱烘48h,称取干重(dw),相对含水量计算公式=(fw-dw)/(tw-dw)
×
100%。
[0135]
分别选取自然干旱处理30d复水5d后,突变体、野生型及过表达烟草的整株苗,清水冲洗并擦干水分后测量其根冠鲜重。接着,在80℃烘箱烘干至恒重后,再测定根冠干重(图21)。研究发现,干旱胁迫后,三组材料的根冠鲜重及干重间均有显著差异。干旱复水后,突变体的根冠鲜重显著高于野生型和过表达高,c3的根冠鲜重值最大;同样,突变体的根冠干重也明显比野生型和过表达高。结果表明了,相比野生型和过表达,干旱胁迫对突变体植物生长的影响较小。
[0136]
相对含水量(rwc)是一种直观地衡量植物叶片含水多少的重要指标,既能够局部反映出植物耐旱性的强弱,又能体现出缺水对植物影响的大小。干旱胁迫下突变体、野生型及过表达烟草叶片相对含水量变化如图所示(图22),干旱14d相较于干旱0d时,所有材料的相对含水量均显著降低,但突变体材料的rwc明显高于野生型和过表达材料。结果表明,干旱胁迫下,突变体材料的保水能力强于野生型和过表达。
[0137]
抗旱相关marker基因表达分析:在自然干旱处理过程中,分别选取干旱0d、14d时突变体、野生型及过表达材料的样品,液氮速冻后-80℃保存。提取rna,利用qrt-pcr方法,检测抗旱相关基因(ntdreb3、nterd10c、ntlea5、ntnced1、ntabi3、ntabi5)的表达(图23)。干旱14d时,6个应激基因表达均上调,但突变体株系上调幅度最大,且与野生型和过表达株系之间具有显著性差异。突变体c2中ntdreb3和nterd10c的表达丰度最高,其次为c1和c3;c1中ntabi5的表达最高,其次为c2和c3;c1中ntlea5的表达最高,其次为c2和c1。结果证明了,ntdtrg基因突变体抗旱性强,与抗旱相关基因的表达量增加密切相关。
[0138]
6、离体叶片失水率分析
[0139]
随机挑选突变体、过表达和wt长势一致的幼苗各9株,进行叶片失水率的测定,在当地时间早上10点每株都取从中心叶往下数第3片叶,每个株系共取9片叶,每片叶按编号,按顺序立即用电子天平称重,然后在常温(25℃)的自然条件下正常放置,每隔1h用电子天平称1次重,连续12h,其中每个株系随机选3片叶连续在0h、6h、12h照相。
[0140]
干旱是植物可利用水分流失的重要影响因素,而叶片失水率是判断其是否抗旱的一项重要生理指标。离体叶片失水表型如图所示(图24),当离体叶片放置12h小时之后,观察可得突变体株系叶片的萎焉程度比野生型和过表达均轻。离体叶片失水率进一步体现出(图25),相比野生型和过表达株系,突变体株系叶片的失水速度较为缓慢。
[0141]
7、气孔开度分析
[0142]
将30d龄幼苗置于mes缓冲液(10mm mes-koh,ph 6.15,10mm kcl,50μm cacl2)中进行peg诱导的气孔关闭试验,在22℃下光照2h使气孔完全开放。将幼苗转移到含有10%peg的mes缓冲液中2~3h,在olympus bx53下拍摄子叶。气孔孔径的测定方法为imagej软件。
[0143]
为了研究烟草叶片蒸腾失水是否与气孔调控相关,观察了10% peg-6000胁迫处理下不同材料的气孔开闭变化(图26),及其气孔数目和开度大小(图27)。突变体株系的叶片气孔较对照关闭程度大,而野生型和过表达较小,说明了干旱处理时,突变体叶片因气孔相对关闭,所以水分流失较慢。
[0144]
8、nbt组织染色试验
[0145]
随机挑选每个株系长势一致的烟苗各3株,每个株系共取3片叶,为3个重复,每株取中心叶下部的第3片叶,將每个株系的离体叶片浸泡在10%peg中,放置于16h光照/8h黑暗的恒温光照培养箱(25℃),处理24h。随后将叶片完全浸泡在0.5mg/mlnbt染色液中,抽真空30min,25℃避光保存8h。弃去染色液,在乙醇:乳酸:甘油(3:1:1)固定液中沸水煮5min,直至叶绿素全部脱去,冷却,加入无水乙醇,置于4℃冰箱内保存一段时间后观察。
[0146]
超氧阴离子自由基(o
2-)是活性氧中的一种含氧自由基,它可与nbt发生组织化学反应,将nbt还原成不溶于水的蓝色甲腙,定位于o
2-的产生部位。利用nbt染色对烟草叶片进行组织化学定位检测,通过观察叶片颜色深浅便可检测o
2-的积累量(图28)。使用10% peg-6000溶液处理烟草叶片后,发现三个过表达株系叶片变蓝程度最深,野生型次之,而突变体株系叶片颜色较浅,c1株系的叶片颜色是最浅的。上述结果说明了,突变体植株相比野生型和过表达更能降低烟草体内o
2-的含量。
[0147]
9、田间表型观察及生理指标测定
[0148]
试验点基本情况:试验在四川省广元市剑阁县进行。试点属中亚热带西部半湿润气候区,气候温和,光热资源丰富,无霜期为250d左右,年降雨量为1211.7mm,降雨量90%以上集中在6-10月。试验地为空闲地。
[0149]
试验设计:育苗方式为漂浮育苗。三组材料均分别种植两行,株距为50cm,行距为1.2m。于4月30号进行移栽,根据当地烟农习惯进行常规水肥管理。
[0150]
测定指标与方法:
[0151]
(1)表型观察:移栽30d后,田间观察表型并及时拍照记录;
[0152]
(2)形态指标的测定:主要指标为株高、有效叶片数、最大叶长、最大叶宽,具体方法参考《烟草农艺性状调查方法(yc/t 142-2010)》;
[0153]
(3)光合指标的测定:具体指标为净光合速率(pn)、气孔导度(gs)及蒸腾速率(tr),spad。
[0154]
在四川剑阁烟区进行田间表型的鉴定,所观察到的表型如图所示(图29)。当烟苗移栽至田间一个月内,除了移栽时的定根水,再无其他水源灌溉,发现相比野生型,三个ntdtrg突变体株系的生长状况良好且长势整齐一致;而过表达株系的生长状况就明显较差,最重要的是三个株系oe-1、oe-2、oe-3竟全都出现早花现象。田间表型说明了ntdtrg突变体株系确实比过表达株系的耐旱性强。
[0155]
农艺性状测定如下表所示(表10),结果发现,三个突变体株系的株高和有效叶片
数均是显著高于野生型,而三个过表达株系明显比野生型低;同时,突变体株系的最大叶长和叶宽也比野生型和过表达株系大,且具有显著性差异。结果表明了,过表达株系的早花确实使其停止了营养生长。
[0156]
表10农艺性状测定
[0157][0158]
叶绿素具有吸收、传递、转换功能的作用。干旱胁迫会对叶绿素的合成、分解代谢产生很大影响,使得烟草叶绿素合成降低,从而直接影响光合效率,导致烟株不能正常生长发育。田间验证试验中(图30),三个突变体株系的净光合速率与spad值均比野生型和过表达高,其蒸腾速率和气孔导度呈相反趋势。结果说明了,若在水分胁迫下,突变体株系的净光合速率较快,并且因其气孔导度和蒸腾速率较小,所以相较野生型和过表达株系,失水速度就较慢。
[0159]
田间烟草生物量(干重)的测定结果,如图31所示,发现突变体株系根和冠的生物量比野生型大,且比过表达株系更大。结果说明了,ntdtrg突变体株系比过表达株系更适合在当地烟区种植。
[0160]
以上所述实施例仅表达了本技术的具体实施方式,其描述较为具体和详细,但并不能因此而理解为对本技术保护范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本技术技术方案构思的前提下,还可以做出若干变形和改进,这些都属于本技术的保护范围。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1