用于确定同时结合的基于细胞的FRET测定法的制作方法
背景技术:
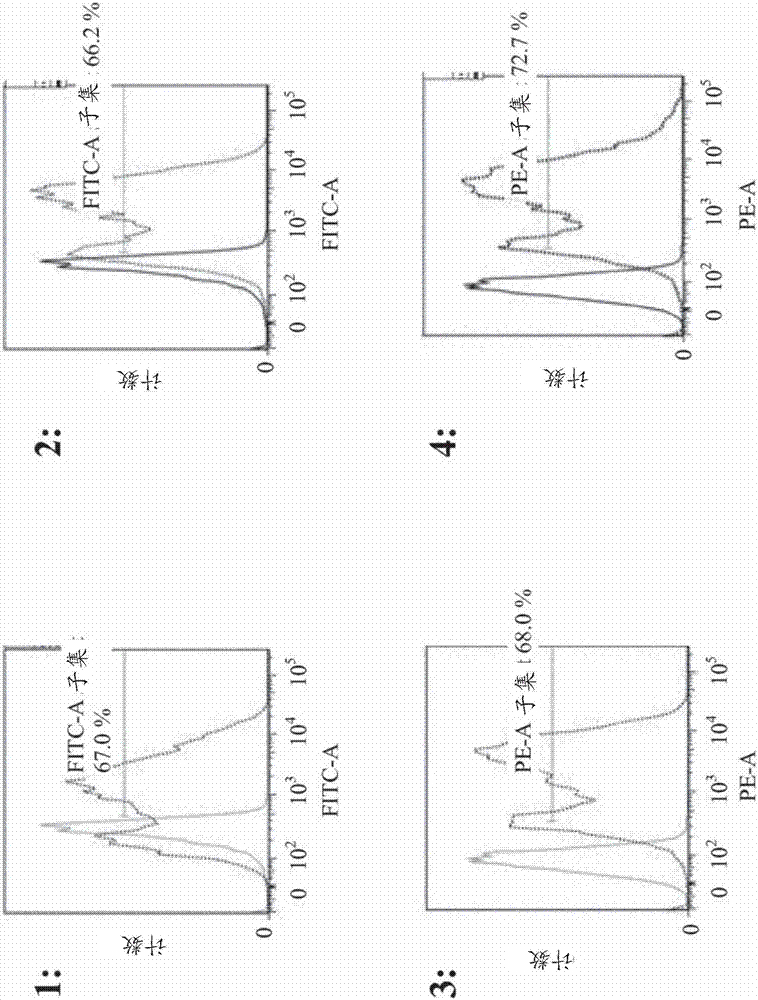
:抗体功能结合的测定是在药物开发中进行的常规测试。荧光共振能量转移(FRET)是在50多年前首次描述的物理现象。FRET基于从供体分子到接受体分子的无辐射能量转移。因此不需要两种分子的直接接触,即它是依赖于距离的能量转移。因此,FRET可以用来研究分子间的相互作用。在US2015/024410中,涉及用于测定抗体与存在于测量培养基中的细胞膜或完整细胞中表达的Fc受体的结合的体外方法,其包括以下步骤:(i)用一对FRET配偶体的第一成员直接或间接标记所述Fc受体,或者将预先直接用一对FRET配偶体的第一成员标记Fc受体的细胞膜或完整细胞引入培养基;(ii)将直接或间接地用所述一对FRET配偶体的第二成员标记的所述抗体添加到测量培养基上;(iii)测量FRET信号,由抗体与Fc受体结合表示FRET信号的存在。在US2010/190266中报道了测量抗体与抗原或抗原表位以及Fc受体或其片段两者的结合活性的方法,包括:将抗体与用一组能够进行荧光共振能量转移的供体和接受体中的一个成员标记的抗原或抗原表位以及用所述一组供体和接受体的另一个成员标记的Fc受体或其片段混合的步骤;用具有能够激发供体的波长的光照射所得混合物的步骤;以及测量所述混合物的荧光水平的步骤。在US2006/165685中报道了双特异性抗Erb-B抗体及其在肿瘤治疗中的用途。在US2015/0204847中报道了用于检测活细胞中蛋白质结构变化的高通量、高精度方法。在US2015/0090936中报道了大斯托克斯位移染料。在US2015/0044690中报道了FRET测量装置和FRET测量方法。技术实现要素:本文报道了测定双特异性抗体与第一和第二抗原/靶同时结合的方法,包括以下步骤:a)将表达细胞膜结合的带FRET-供体标签的第一抗原/靶和带FRET-接受体标签的第二抗原/靶与双特异性抗体一起孵育,和b)通过测定从FRET-供体向FRET-接受体的能量转移来测定双特异性抗体的同时结合。在一个实施方案中,细胞膜结合的FRET-供体和细胞膜结合的FRET-接受体以N-至C-端方向包含:H2N-抗原/靶-FRET-供体/接受体-跨膜结构域-COOH,或H2N-FRET-供体/接受体-抗原/靶-跨膜结构域-COOH,或H2N-FRET-供体/接受体-跨膜结构域-抗原/靶-COOH。在一个实施方案中,FRET-供体和/或接受体分别是标签和FRET-供体-荧光染料或FRET-接受体-荧光染料的缀合物。在一个实施方案中,缀合物是非共价缀合物,并且FRET-供体-荧光染料和/或FRET-接受体-荧光染料是荧光染料缀合的抗体。在一个实施方案中,缀合物是共价缀合物,标签是酶并且FRET-供体-荧光染料和/或FRET-接受体-荧光染料是荧光染料标记的自杀酶底物。在一个实施方案中,i)FRET-供体或FRET-接受体是SNAP-标签和苄基鸟嘌呤的共价复合物,并且ii)FRET-接受体或FRET-供体是CLIP-标签和O2-苄基胞嘧啶的共价复合物,其中FRET-供体和FRET-接受体是不同的标签。在一个实施方案中,双特异性抗体包含特异性结合第一抗原/靶的一个或两个结合位点和特异性结合第二抗原/靶的一个或两个结合位点。本文报道的一个方面是用于确定双特异性抗体的以较高KD值特异性结合其抗原/靶的结合位点的结合相互作用(强度)的方法,其包含特异性结合第一抗原/靶的第一结合位点和特异性结合第二抗原/靶的第二结合位点,由此两个结合位点以结合亲和力至少10倍差异结合其各自抗原/靶,包括以下步骤:a)将表达细胞膜结合的带FRET-供体标签的第一抗原/靶和带FRET-接受体标签的第二抗原/靶与双特异性抗体一起孵育,并通过测定来自FRET-供体到FRET-接受体的能量转移来测定双特异性抗体的结合,b)在增加浓度的包含具有较低KD值的双特异性抗体的结合位点的单特异性抗体存在下重复步骤a),和c)通过测定从FRET-供体向FRET-接受体的能量转移减少时的单特异性抗体的浓度来测定结合相互作用(强度)。在一个实施方案中,单特异性抗体是二价的。发明详细描述杵臼二聚化模块及其在抗体工程中的用途描述于CarterP.RidgwayJ.B.B.;PrestaL.G.:Immunotechnology,第2卷,第1期,1996年2月,第73-73(1)页。关于人免疫球蛋白轻链和重链的核苷酸序列的一般信息在以下文献中给出:Kabat,EA等人,SequencesofProteinsofImmunologicalInterest,第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991)。如本文所用,重链和轻链的所有恒定区和结构域的氨基酸位置根据Kabat等人,SequencesofProteinsofImmunologicalInterest,第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991)中描述的Kabat编号系统来编号,在此被称为“根据Kabat编号”。具体而言,Kabat等人,SequencesofProteinsofImmunologicalInterest,第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991)的Kabat编号系统(参见第647-660页)用于κ和λ同种型的轻链恒定结构域CL和KabatEU索引编号系统(参见第661-723页)用于恒定重链结构域(CH1,铰链,CH2和CH3,在这种情况下其在本文中通过参考“根据KabatEU索引编号”而澄清)。I.定义如本文所用,术语“抗原”表示抗体结合位点所结合的化合物/部分。抗原可以是可以产生抗体的任何部分,即靶,例如肽、多肽、蛋白质、半抗原或小分子。如本文所用,术语“FRET”和“荧光共振能量转移”表示在两个生色团之间发生的非辐射能量转移过程。术语“生色团”表示包括吸收某些波长的光并与另一生色团的这种区域相互作用以便可用于FRET的区域的部分。适用于FRET测定的生色团是本领域技术人员已知的并且容易获得。在一个实施方案中,生色团可以是供体(也称为FRET-供体)。FRET-供体表示这样的分子,其将吸收能量然后随着时间重新发射至少一部分能量。在一个实施方案中,生色团可以是接受体(也称为FRET-接受体)。FRET-接受体表示将接受来自FRET-供体的无辐射能量,由此降低供体的发射强度和激发态寿命的分子。因此,只要FRET-供体和FRET-接受体在物理上位置足够接近(最通常在2.5-12nm内),则两个FRET-部分一起发挥作用,并且在以合适的波长激发时,FRET-供体向FRET-接受体转移精确量的能量(与FRET-供体-FRET-接受体距离的负六次方成正比)。通过观察FRET-供体荧光强度或寿命的减少或者在某些情况下还有FRET-接受体作为荧光再次发射的能量,可以特异性且定量检测该过程。“多特异性抗体”表示对相同抗原或两种不同抗原上的至少两个不同表位具有结合特异性的抗体。多特异性抗体可以制备为全长抗体或抗体片段(例如F(ab')2双特异性抗体)或其组合(例如全长抗体加上另外的scFv或Fab片段)。具有两个、三个或更多(例如四个)功能性抗原结合位点的工程化抗体也已有报道(参见例如US2002/0004587A1)。为了本文的目的的"接受体人框架"是包含如下文定义的源自人免疫球蛋白框架或人共有框架的轻链可变结构域(VL)框架或重链可变结构域(VH)框架的氨基酸序列的框架。"源自"人免疫球蛋白框架或人共有框架的接受体人框架可以包含其相同氨基酸序列,或它可以含有氨基酸序列变化。在一些实施方案中,氨基酸变化的数量为10或更少、9或更少、8或更少、7或更少、6或更少、5或更少、4或更少、3或更少或2或更少。在某些实施方案中,VL接受体人框架与VL人免疫球蛋白框架序列或人共有框架序列在序列上相同。“亲和力”指分子(例如抗体)的单个结合部位与其结合配偶体(例如抗原)之间非共价相互作用的总和的强度。除非另有说明,本文所用的“结合亲和力”指内在的结合亲和力,其反映结合对(例如抗体和抗原)的成员之间的1:1相互作用。分子X对其配偶体Y的亲和力一般可通过解离常数(kd)来表示。亲和力可以通过本领域公知的方法来测量,例如表面等离子体共振,并且包括本文中所述的那些。“亲和力成熟的”抗体指与不具有这类改变的亲本抗体相比在一个或多个高变区(HVR)中具有一个或多个改变的抗体,这类改变导致抗体对其一个或多个抗原的亲和力的改善。术语“抗体”在本文中以最广泛的含义使用并涵盖多种抗体结构,包括但不限于单克隆抗体、多克隆抗体,和多特异性抗体(例如双特异性抗体),只要它们显示希望的抗原结合活性。“抗体片段”指除完整抗体以外的包含完整抗体的部分的分子,其结合该完整抗体所结合的抗原。抗体片段的实例包括但不限于Fv、Fab、Fab'、Fab’-SH、F(ab')2;双抗体;线性抗体;单链抗体分子(例如scFv);及从抗体片段形成的多特异性抗体。术语“嵌合”抗体指这样的抗体,其中部分重链和/或轻链源自特定来源或物种,而重链和/或轻链的其余部分源自不同的来源或物种。抗体的“种类”指的是其重链具有的恒定结构域或恒定区的类型。有五大类抗体:IgA,IgD,IgE,IgG和IgM,其中有些可进一步分为亚类(同种型),例如IgG1,IgG2,IgG3,IgG4,IgA1和IgA2。对应于免疫球蛋白不同种类的重链恒定结构域分别被称为α,δ,ε,γ和μ。IgG抗体Fc区受体(FcγR)的交联触发各种各样的效应器功能,包括吞噬作用、抗体依赖性细胞毒作用和炎性介质的释放,以及免疫复合物清除和抗体产生的调节。在人类中,已经表征了三类FcγR,它们是:-FcγRI(CD64)以高亲和力结合单体IgG并在巨噬细胞、单核细胞、嗜中性粒细胞和嗜酸性粒细胞上表达。在Fc区IgG中氨基酸残基E233-G236,P238,D265,N297,A327和P329(根据Kabat的EU索引编号)的至少一个中的修饰降低了与FcγRI的结合。在233-236位被取代成IgG1和IgG4的IgG2残基使与FcγRI的结合降低了103倍,并消除了对抗体致敏的红血细胞的人单核细胞应答(Armor,KL等人,Eur.J.Immunol.29(1999)2613-2624),-FcγRII(CD32)以中等至低亲和力结合复合IgG并被广泛表达。该受体可以分为两个亚型FcγRIIA和FcγRIIB。FcγRIIA在涉及杀死的许多细胞(例如巨噬细胞、单核细胞、嗜中性粒细胞)中发现并似乎能够激活杀死过程。FcγRIIB似乎在抑制过程中发挥作用,并在B细胞、巨噬细胞和肥大细胞以及嗜酸性粒细胞上发现。在B细胞上,它似乎起到进一步抑制免疫球蛋白产生和同种型转换成例如IgE类的作用。在巨噬细胞上,FcγRIIB起到抑制通过FcγRIIA的吞噬作用的作用。在嗜酸性粒细胞和肥大细胞上,B型可能有助于抑制通过IgE与其单独的受体结合的这些细胞的活化。发现对FcγRIIA的结合降低,例如对于包含IgGFc-区的抗体,所述Fc-区在氨基酸残基E233-G236,P238,D265,N297,A327,P329,D270,Q295,A327,R292和K414(根据Kabat的EU索引编号)中的至少一个处具有突变,-FcγRIII(CD16)以中等至低亲和力结合IgG并存在两种类型。FcγRIIIA存在于NK细胞、巨噬细胞、嗜酸性粒细胞以及一些单核细胞和T细胞上并介导ADCC。FcγRIIIB在嗜中性粒细胞上高度表达。发现与FcγRIIIA的降低的结合,例如,对于包含IgGFc-区的抗体,所述Fc区在氨基酸残基E233-G236,P238,D265,N297,A327,P329,D270,Q295,A327,S239,E269,E293,Y296,V303,A327,K338和D376(根据Kabat的EU索引编号)中至少一个处具有突变。人IgG1上的结合位点对于Fc受体的作图、上述提到的突变位点和用于测量与FcγRI和FcγRIIA的结合的方法描述于Shields,R.L.等人,J.Biol.Chem.276(2001)6591-6604。术语"全长抗体"、"完整抗体"和"全抗体"在本文中互换地用于表示这样的抗体:其具有与天然抗体结构基本上类似的结构或具有含有如本文中定义的Fc区的重链。“全长抗体”是包含抗原结合可变区以及轻链恒定结构域(CL)和重链恒定结构域CH1,CH2和CH3的抗体。恒定结构域可以是天然序列恒定结构域(例如人类天然序列恒定结构域)或其氨基酸序列变体。更详细地,全长抗体包含两条抗体轻链(各自包含轻链可变结构域和轻链恒定结构域)和两条抗体重链(各自包含重链可变结构域,铰链区和重链恒定结构域CH1,CH2和CH3)。C-端氨基酸残基K或GK可以在全长抗体的两条抗体重链中彼此独立存在或不相互独立。“人源化”抗体是指包含来自非人HVR氨基酸残基和来自人FR氨基酸残基的嵌合抗体。在某些实施方案中,人源化抗体将基本上包含至少一个,通常两个可变结构域的全部,其中HVR(例如CDR)的全部或基本上全部对应于非人类抗体的那些,和全部或基本上全部对应于人抗体的FR的FR。人源化抗体任选地可以包含来自人抗体的抗体恒定区的至少一部分。抗体(例如,非人抗体)的“人源化形式”,是指已经历人源化的抗体。“分离的抗体”指已经与其天然环境的组分分开的抗体。在一些实施方案中,抗体纯化至大于95%或99%的纯度,如通过例如电泳(例如,SDS-PAGE、等电聚焦(IEF)、毛细管电泳)或层析(例如,离子交换或反相HPLC)测定的。关于用于评估抗体纯度的方法的综述,参见例如Flatman等人,J.Chromatogr.B848(2007)79-87。在用于本文时,术语“单克隆抗体”指从一群基本上同质的抗体获得的抗体,即构成群体的各个抗体是相同的和/或结合相同表位,除了例如含有天然存在的突变或在单克隆抗体制备物的生成期间发生的可能的变体抗体外,此类变体一般以极小量存在。与通常包含针对不同决定簇(表位)的不同抗体的多克隆抗体制备物不同,单克隆抗体制备物的每种单克隆抗体针对抗原上的单一决定簇。如此,修饰语“单克隆”指示抗体自一群基本上同质的抗体获得的特性,而不应解释为要求通过任何特定方法来生成抗体。例如,可以通过多种技术来生成要依照本发明使用的单克隆抗体,包括但不限于杂交瘤方法、重组DNA方法、噬菌体展示方法、和利用含有所有或部分人免疫球蛋白基因座的转基因动物的方法,本文中描述了用于生成单克隆抗体的此类方法和其它例示性方法。“裸抗体”指未与异源模块(例如细胞毒性部分)或放射性标记物缀合的抗体。裸抗体可以存在于药物配制剂中。“天然抗体”指具有不同结构的天然存在的免疫球蛋白分子。例如,天然IgG抗体是约150,000道尔顿的异四聚糖蛋白,由二硫化物键合的两条相同轻链和两条相同重链构成。从N至C端,每条重链具有一个可变区(VH),又称作可变重链结构域或重链可变结构域,接着是三个恒定结构域(CH1、CH2、和CH3),其中铰链区位于第一和第二恒定结构域之间。类似地,从N至C端,每条轻链具有一个可变区(VL),又称作可变轻链结构域或轻链可变结构域,接着是一个恒定轻(CL)结构域。根据其恒定结构域氨基酸序列,抗体轻链可归入两种类型中的一种,称作κ和λ。II.本文报道的方法荧光共振能量转移(FRET)是50多年前首次描述的物理现象。FRET是基于从供体分子到接受体分子的无辐射能量转移。因此不需要两个分子的直接接触,即它是依赖于距离的能量转移。因此,FRET可以用来研究分子间的相互作用。FRET-供体是最初吸收提供的能量的生色团。FRET-接受体是能量随后从FRET-供体转移到的生色团。这种相互作用在没有将吸收的能量转化为热能的情况下发生,并且不需要直接的分子相互作用(例如通过碰撞)。从FRET-供体到FRET-接受体的能量转移导致FRET-接受体能量状态的增加。最后,这导致FRET-接受体发射辐射。为了使FRET在FRET-供体和FRET-接受体特别接近的地方发生,FRET-接受体的激发波长必须在FRET-供体的荧光发射光谱内。基于FRET的能量转移通常可以通过FRET-接受体荧光的出现或通过FRET-供体荧光的减弱来检测。FRET-供体总是荧光分子。对于FRET检测,存在许多不同的方式。由于FRET影响FRET-供体的荧光以及FRET-接受体的荧光改变,所以可以产生两个信号的比率,其与绝对浓度无关。基于荧光的光谱学为生物学测定提供了卓越的灵敏度,因为待测荧光的波长与激发和背景荧光的波长不同。用于生物测定的基于荧光的光谱方法的一个缺点是可检测信号强度的可变性(Gribbon等人,DrugDiscov.Today8(2003)1035;Gakamsky等人,Anal.Biochem.409(2011)89;Thorne等人,Curr.Opin.Chem.Biol.14(2010)315)。由于时间分辨信号在很大程度上与强度变化无关(Moger等人,Screening11(2006)765),所以建立了时间分辨荧光测量以改善测定性能。荧光寿命用于诸如矩分析(Isenberg等人,Biophys.J.9(1969)1337),多指数拟合(Knutson等人,Chem.Phys.Lett.102(1983)501;Beechem等人,Num.Comput.Methods210(1992)37)或相量分析(Jameson等人,Methods59(2013)278)的方法中。它用于基于生物荧光的测定中,如共振能量转移(参见例如Lakowicz,PrinciplesofFluorescenceSpectroscopy,第3版。(Springer,NewYork,2006))。已经使用该方法测量了生物相互作用(Jameson等人,Methods59(2013)278;Dong等人,Biochem.Biophys.Res.Commun.449(2014)196;Lebakken等人,J.Biomol.Screening(2007)828;Maltman等人,Chem.Commun.46(2010)6929;Paterson等人,Anal.Biochem.402(2010)54)。采用荧光寿命测定的方法包括时间相关单光子计数(TCSPC),(Becker,TheBHTCSPCHandbook,第5版。(Becker&Hickl,Berlin,2012)),直接波形记录(DWR)(Muretta等人,Rev.Sci.Instrum.81(2010)103101)。本文报道的方法基于多特异性抗体同时结合其抗原中的两种时,荧光共振能量转移(FRET)的改变(增加或出现)。抗原呈现为在真核细胞外细胞膜上具有荧光团的融合蛋白,其中一种抗原与FRET-供体荧光团融合,并且相应的另一种抗原与FRET-接受体荧光团融合。本文报道了测定多特异性抗体与第一和第二抗原同时结合的方法,包括以下步骤:a)将表达细胞膜结合的带FRET-供体标签的/FRET-供体融合的第一抗原和带FRET-接受体标签的/FRET-接受体融合的第二抗原的细胞与多特异性抗体一起孵育,和b)通过测定从FRET-供体到FRET-接受体的能量转移来测定多特异性抗体的同时结合。本文报道的一个方面是用于测定双特异性抗体的方法,其包含特异性结合第一抗原的第一结合位点和特异性结合第二抗原的第二结合位点,由此两个结合位点以结合亲和力至少10差异与其各自抗原结合,以较高KD值特异性结合其抗原的结合位点的结合相互作用(强度),包括以下步骤:a)将表达细胞膜结合的带FRET-供体标签的/FRET-供体-融合的第一抗原和带FRET-接受体标签的/FRET-接受体融合的第二抗原的细胞与双特异性抗体一起孵育,并通过测定从FRET-供体到FRET-接受体的能量转移来测定双特异性抗体的结合,b)在增加浓度的包含具有较低KD值的双特异性抗体的结合位点的单特异性(二价)抗体存在下重复步骤a),和c)通过测定从FRET-供体向FRET-接受体的能量转移减少时的单特异性(二价)抗体的浓度来测定结合相互作用(强度)。本文报道的一个方面是通过测定第一生色团和第二生色团之间的荧光共振能量转移(FRET)中的改变来测定多特异性抗体与第一抗原和第二抗原的同时结合的方法,包括以下步骤:-提供在细胞外表面/膜上包含第一抗原和第二抗原的基因工程细胞,其中第一抗原与第一生色团融合,并且其中第二抗原与第二生色团融合;-使细胞与多特异性抗体接触以形成混合物;和-测量第一生色团、第二生色团或其组合的荧光寿命,其中多特异性抗体存在时的荧光寿命与不存在多特异性抗体时的荧光寿命之间的差异表明多特异性抗体同时结合第一抗原和第二抗原。在一个实施方案中,细胞膜结合的FRET-供体和细胞膜结合的FRET-接受体以N-至C-端方向包含:H2N-抗原-FRET-供体/接受体-跨膜结构域-COOH。在一个实施方案中,FRET-供体和/或接受体是标签分别和FRET-供体-荧光染料或FRET-接受体-荧光染料的缀合物。在一个实施方案中,缀合物是非共价缀合物,并且FRET-供体-荧光染料和/或FRET-接受体-荧光染料是荧光染料缀合的抗体。在一个实施方案中,缀合物是共价缀合物,标签是酶并且FRET-供体-荧光染料和/或FRET-接受体-荧光染料是荧光染料标记的自杀酶底物。在所有方面和实施方案的一个实施方案中,FRET是TR-FRET(时间分辨荧光共振能量转移)。在所有方面和实施方案的一个实施方案中,第一生色团是FRET-供体,并且第二生色团是FRET-接受体。FRET-供体和FRET-接受体是根据它们进行FRET的适宜性而选择的。在所有方面和实施方案的一个实施方案中,生色团是与荧光染料缀合的氨基酸。合适的生色团的实例包括但不限于荧光蛋白,包括绿色荧光蛋白、红色荧光蛋白、黄色荧光蛋白和蓝色荧光蛋白。绿色荧光蛋白和红色荧光蛋白可以用作FRET-供体-FRET-接受体对,蓝色荧光蛋白和黄色荧光蛋白可以用作FRET-供体-FRET-接受体对。不同形式的绿色荧光蛋白、红色荧光蛋白、黄色荧光蛋白和蓝色荧光蛋白以及这些蛋白的类似物的氨基酸序列对于本领域技术人员是已知的并且容易获得。其他生色团包括荧光染料,例如当蛋白质存在于细胞上时可以附着到蛋白质上的荧光染料。这种染料的实例在本领域中是已知的并且常规使用。实例包括与半胱氨酸反应的染料,所述半胱氨酸带有反应性碘乙酰胺、马来酰亚胺或硫代磺酸盐基团。其他实例包括蛋白质标记试剂FLASH-EDT2(一种可以与结构域CCXXCC(Invitrogen)反应的染料)和SNAP-标签(一种自我标记的蛋白质标签(NewEnglandBiolabs))。可以与通过修饰的tRNA掺入靶蛋白的非天然氨基酸特异性反应的其他荧光染料是可用的。任何适当选择的两种生色团可以在本文报道的方法中用作FRET-供体-FRET-接受体对,条件是由FRET-供体发射的能量(发射光谱)与由FRET-接受体吸收的能量(激发光谱)重叠,例如在两个生色团之间发生能量转移过程(FRET)。符合这种重叠的FRET-供体和FRET-接受体被称为FRET-供体-FRET-接受体对。在一个实施方案中,可以在较长波长激发的FRET-供体优于在较短波长激发的FRET-供体。此外,具有更长FLT(超过3纳秒(ns))的探针将优于具有更短FLT的探针。在所有方面和实施方案的一个实施方案中,FRET-供体是具有与FRET-接受体的激发波长重叠的发射波长的荧光染料。FRET-供体以及FRET-接受体可以在N-端、C-端或内部位置插入或融合至多肽,即相应的抗原。在一个实施方案中,i)FRET-供体或FRET-接受体是SNAP-标签和苄基鸟嘌呤的共价复合物,并且ii)FRET-接受体或FRET-供体是CLIP-标签和O2-苄基胞嘧啶共价复合物,其中FRET-供体和FRET-接受体是不同的标签。在所有方面和实施方案的一个实施方案中,通过混合细胞和多特异性抗体来启动FRET。在本文报道的所有方面和实施方案的一个实施方案中,FRET-供体和FRET-接受体包含荧光染料可以附着的异源肽结构域。在一个实施方案中,本文报道的方法包括使在其表面上呈递抗原的细胞与适当的蛋白质标记试剂接触以用荧光染料标记抗原的步骤。在所有方面和实施方案的一个实施方案中,FRET-供体和FRET-接受体各自是酶和荧光团的共价缀合物。在所有方面和实施方案的一个实施方案中,多特异性抗体存在时的荧光寿命与不存在多特异性抗体时的荧光寿命之间的差异是FRET-供体的荧光寿命的减少。在所有方面和实施方案的一个实施方案中,多特异性抗体存在时的荧光寿命与不存在多特异性抗体时的荧光寿命之间的差异是FRET-接受体的荧光寿命的增加。在所有方面和实施方案的一个实施方案中,多特异性抗体是双特异性抗体。在本文报道的所有方面和实施方案的一个实施方案中,第一抗原不同于第二抗原。在本文报道的所有方面的一个实施方案中,第一抗原和第二抗原是相同的,并且双特异性抗体结合第一抗原上的第一表位和第一抗原上的第二表位,其中第一表位不与第二表位重叠。在所有方面和实施方案的一个实施方案中,通过测定FRET-供体的发射信号与FRET-接受体的发射信号的比率或FRET-接受体的发射信号与FRET-供体的发射信号的比率来测定FRET。在所有方面和实施方案的一个实施方案中,用于测定的发射波长带宽小于激发波长带宽。在所有方面的一个实施方案中,细胞是真核细胞。在一个实施方案中,细胞是HEK细胞或CHO细胞。在本文报道的方法的一个实施方案中,在单个发射波长处测定FRET。在某些实施方案中,FRET荧光寿命在两个或更多个波长下测定。在某些实施方案中,本文报道的方法在具有多个孔的板的孔中进行,所述板例如多孔板或多域多孔板。III.多特异性抗体本方法的抗体是多特异性抗体。更详细地说,本方法的抗体是多特异性抗体,例如双特异性抗体。多特异性抗体是对至少两个不同靶具有结合特异性的单克隆抗体。在某些实施方案中,结合特异性之一针对第一抗原/靶,另一个针对任何其他抗原/靶。在某些实施方案中,双特异性抗体可以结合相同抗原/靶的两个不同表位。双特异性抗体可以制备为全长抗体或抗体片段。用于制备多特异性抗体的技术包括但不限于具有不同特异性的两个免疫球蛋白重链-轻链对的重组共表达(见Milstein,C.和Cuello,A.C.,Nature305(1983)537-540,WO93/08829,和Traunecker,A.等人,EMBOJ.10(1991)3655-3659),和“杵臼(knob-in-hole)”工程化(见,例如,US5,731,168)。多特异性抗体还可以通过如下制备:用于制备抗体Fc-异二聚体分子的工程化静电转向效果(WO2009/089004);交联两个或更多个抗体或片段(参见,例如,US4,676,980,和Brennan,M.等人,Science229(1985)81-83);使用亮氨酸拉链产生双特异性抗体(见,例如,Kostelny,S.A.等人,J.Immunol.148(1992)1547-1553;使用“双抗体(diabody)”技术制备双特异性抗体片段(见,例如,Holliger,P.等人,Proc.Natl.Acad.Sci.USA90(1993)6444-6448);和使用单链Fv(sFv)二聚体(见,例如Gruber,M等人,J.Immunol.152(1994)5368-5374);和制备三特异性抗体,如,例如,Tutt,A.等人,J.Immunol.147(1991)60-69)中所述。具有三个或更多个功能性抗原结合位点的工程化抗体,包括“章鱼抗体”也包括在本文中(参见,例如US2006/0025576)。本文中的抗体或片段还包括“双作用Fab”或“DAF”,其包含结合[[PRO]]以及另一不同抗原的抗原结合位点(参见,例如US2008/0069820)。本文中的抗体或片段还包括在以下所述的多特异性抗体:WO2009/080251,WO2009/080252,WO2009/080253,WO2009/080254,WO2010/112193,WO2010/115589,WO2010/136172,WO2010/145792和WO2010/145793。如本方法中的抗体至少是双特异性抗体。双特异性抗体是多特异性抗体组的成员。双特异性抗体包含至少两个结合位点,每个结合位点通过结合不同抗原或同一抗原上的不同表位的抗体重链可变结构域(VH)和抗体轻链可变结构域(VL)对形成。这种双特异性抗体是1+1形式。其他双特异性抗体形式为2+1形式(包含第一抗原或表位的两个结合位点和第二抗原或表位的一个结合位点)或2+2形式(包含第一抗原或表位的两个结合位点和第二抗原或表位的两个结合位点)。在所有方面的一个实施方案中,本方法的抗体包含(所有位置根据Kabat的EU索引)i)任选地具有突变P329G,L234A和L235A的人IgG1亚类的同源二聚体Fc区,或ii)任选具有突变P329G,S228P和L235E的人IgG4亚类的同源二聚体Fc-区,或iii)任选具有突变P329G,L234A,L235A,I253A,H310A和H435A,或任选地具有突变P329G,L234A,L235A,H310A,H433A和Y436A的人IgG1亚类的同源二聚体Fc-区,或iv)异源二聚体Fc区,其中a)一个Fc-区多肽包含突变T366W,另一个Fc-区多肽包含突变T366S,L368A和Y407V,或b)一个Fc-区多肽包含突变T366W和Y349C,另一个Fc-区多肽包含突变T366S,L368A,Y407V和S354C,或c)一个Fc-区多肽包含突变T366W和S354C,并且另一个Fc-区多肽包含突变T366S,L368A,Y407V和Y349C,或v)人IgG1亚类的异二聚体Fc区,其两个Fc-区多肽包含突变P329G,L234A和L235A,并且a)一个Fc-区多肽包含突变T366W,并且另一个Fc-区多肽包含突变T366S,L368A和Y407V,或b)一个Fc-区多肽包含突变T366W和Y349C,并且另一个Fc-区多肽包含突变T366S,L368A,Y407V和S354C,或c)一个Fc-区多肽包含突变T366W和S354C,并且另一个Fc-区多肽包含突变T366S,L368A,Y407V和Y349C,或vi)人IgG4亚类的异二聚体Fc区,其两个Fc区多肽包含突变P329G,S228P和L235E,并且a)一个Fc-区多肽包含突变T366W,并且另一个Fc-区多肽包含突变T366S,L368A和Y407V,或b)一个Fc-区多肽包含突变T366W和Y349C,并且另一个Fc-区多肽包含突变T366S,L368A,Y407V和S354C,或c)一个Fc-区多肽包含突变T366W和S354C,并且另一个Fc-区多肽包含突变T366S,L368A,Y407V和Y349C,或vii)i)、ii)和iii)之一与vi)、v)和vi)之一的组合。在一个实施方案中,多特异性抗体是二价、双特异性抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中第二轻链和第二重链的可变结构域VL和VH相互取代。a)中的抗体不包含如b)所报告的修饰,a)中的重链和轻链是分离的链。在b)的抗体中在轻链内可变轻链结构域VL被所述抗体的可变重链结构域VH取代,和在重链内可变重链结构域VH被所述抗体的可变轻链结构域VL取代。在一个实施方案中i)在a)的第一轻链的恒定结构域CL中,第124位氨基酸(根据Kabat编号)被带正电的氨基酸取代,和其中在a)的第一重链的恒定结构域CH1中,第147位氨基酸或第213位氨基酸(根据KabatEU索引编号)被带负电的氨基酸取代,或者ii)在b)的第二轻链的恒定结构域CL中,第124位氨基酸(根据Kabat编号)被带正电的氨基酸取代,和其中在b)的第二重链的恒定结构域CH1中,第147位氨基酸或第213位氨基酸(根据KabatEU索引编号)被带负电的氨基酸取代。在一个实施方案中i)在a)的第一轻链的恒定结构域CL中,第124位氨基酸独立地被赖氨酸(K)、精氨酸(R)或组氨酸(H)取代(根据Kabat编号)(在一个优选的实施方案中,独立地被赖氨酸(K)或精氨酸(R)取代),和其中在a)的第一重链的恒定结构域CH1中,第147位氨基酸或第213位氨基酸独立地被谷氨酸(E)或天冬氨酸(D)取代(根据KabatEU索引编号),或者ii)在b)的第二轻链的恒定结构域CL中,第124位氨基酸独立地被赖氨酸(K)、精氨酸(R)或组氨酸(H)取代(根据Kabat编号)(在一个优选的实施方案中,独立地被赖氨酸(K)或精氨酸(R)取代),和其中在b)的第二重链的恒定结构域CH1中,第147位氨基酸或第213位氨基酸独立地被谷氨酸(E)或天冬氨酸(D)取代(根据KabatEU索引编号)。在一个实施方案中,在第二重链的恒定结构域CL中,第124位和123位氨基酸被K取代(根据KabatEU索引编号)。在第二重链的恒定结构域CL的一个实施方案中,第123位的氨基酸被R取代,第124位的氨基酸被K取代(根据KabatEU索引编号)。在一个实施方案中,在第二轻链的恒定结构域CH1中,第147位和213位氨基酸被E取代(根据KabatEU索引编号)。在一个实施方案中,在第一轻链的恒定结构域CL中,第124位和123位的氨基酸被K取代,和在第一重链的恒定结构域CH1中,第147位和213位的氨基酸被E取代(根据KabatEU索引编号)。在一个实施方案中,在第一轻链的恒定结构域CL中,第123位的氨基酸被R取代并且第124位的氨基酸被K取代,并且在第一重链的恒定结构域CH1中,第147和213位的氨基酸都被E取代(根据KabatEU索引编号)。在一个实施方案中,在第二重链的恒定结构域CL中,第124位和123位的氨基酸被K取代,和其中在第二轻链的恒定结构域CH1中,第147位和213位氨基酸被E取代,和在第一轻链的可变结构域VL中,第38位的氨基酸被K取代,在第一重链的可变结构域VH中,第39位的氨基酸被E取代,在第二重链的可变结构域VL中,第38位的氨基酸被K取代,和在第二轻链的可变结构域VH中,第39位的氨基酸被E取代(根据KabatEU索引编号)。在一个实施方案中,多特异性抗体是二价、双特异性抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中第二轻链和第二重链的可变结构域VL和VH相互取代,和其中第二轻链和第二重链的恒定结构域CL和CH1相互取代。a)中的抗体不包含如b)所报告的修饰,a)中的重链和轻链是分离的链。在b)的抗体中在轻链内可变轻链结构域VL被所述抗体的可变重链结构域VH取代,恒定轻链结构域CL被所述抗体的恒定重链结构域CH1取代;和在重链内可变重链结构域VH被所述抗体的可变轻链结构域VL取代,恒定重链结构域CH1被所述抗体的恒定轻链结构域CL取代。在一个实施方案中,多特异性抗体是二价、双特异性抗体,其包含a)特异性结合第一抗原的抗体的第一轻链和第一重链,和b)特异性结合第二抗原的抗体的第二轻链和第二重链,其中第二轻链和第二重链的恒定结构域CL和CH1相互取代。a)中的抗体不包含如b)所报告的修饰,a)中的重链和轻链是分离的链。在b)的抗体中在轻链内恒定轻链结构域CL被所述抗体的恒定重链结构域CH1取代;和在重链内恒定重链结构域CH1被所述抗体的恒定轻链结构域CL取代。在一个实施方案中,多特异性抗体是一种多特异性抗体,其包含a)特异性结合第一抗原的全长抗体,其由两条抗体重链和两条抗体轻链组成,和b)特异性结合一至四个其他抗原(即第二和/或第三和/或第四和/或第五抗原,优选特异性结合一个其他抗原,即第二抗原)的一、二、三或四个单链Fab片段,其中在b)中所述的单链Fab片段通过在所述全长抗体的重链或轻链的C-或N-端的肽接头融合至a)中的所述全长抗体。在一个实施方案中,结合第二抗原的一个或两个相同的单链Fab片段通过在全长抗体的重链或轻链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,结合第二抗原的一个或两个相同的单链Fab片段通过在全长抗体的重链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,结合第二抗原的一个或两个相同的单链Fab片段通过在全长抗体的轻链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,结合第二抗原的两个相同的单链Fab片段通过在全长抗体的每条重链或轻链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,结合第二抗原的两个相同的单链Fab片段通过在全长抗体的每条重链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,结合第二抗原的两个相同的单链Fab片段通过在全长抗体的每条轻链的C-端的肽接头融合至所述全长抗体。在一个实施方案中,多特异性抗体是三价、双特异性抗体,其包含a)特异性结合第一抗原的全长抗体,其由两条抗体重链和两条抗体轻链组成,b)第一多肽,其由以下组成ba)抗体重链可变结构域(VH),或者bb)抗体重链可变结构域(VH)和抗体恒定结构域1(CH1),其中所述第一多肽通过所述全长抗体的两条重链之一的C-端的肽接头与其VH结构域的N-端融合,c)第二多肽由以下组成ca)抗体轻链可变结构域(VL),或者cb)抗体轻链可变结构域(VL)和抗体轻链恒定结构域(CL),其中所述第二多肽通过所述全长抗体的两条重链的另一条的C-端的肽接头与VL结构域的N-端融合,和其中第一多肽的抗体重链可变结构域(VH)和第二多肽的抗体轻链可变结构域(VL)一起形成特异性结合第二抗原的抗原结合位点。在一个实施方案中,b)中多肽的抗体重链可变结构域(VH)和c)中多肽的抗体轻链可变结构域(VL)通过在以下位置之间引入二硫键的链间二硫键桥相连接和稳定:i)重链可变结构域第44位至轻链可变结构域第100位,或ii)重链可变结构域第105位至轻链可变结构域第43位,或iii)重链可变结构域第101位至轻链可变结构域第100位(通常根据KabatEU索引编号)。引入非天然二硫键桥用于稳定的技术描述在例如WO94/029350,Rajagopal,V.,等人,Prot.Eng.(1997)1453-1459;Kobayashi,H.,等人,Nucl.Med.Biol.25(1998)387-393;和Schmidt,M.,等人,Oncogene18(1999)1711-1721。在一个实施方案中b)和c)中多肽的可变结构域之间的任选二硫键在重链可变结构域第44位和轻链可变结构域第100位之间。在一个实施方案中b)和c)中多肽的可变结构域之间的任选二硫键在重链可变结构域第105位和轻链可变结构域第43位之间(通常根据Kabat编号)。在一个实施方案中,优选三价、双特异性抗体在单链Fab片段的可变结构域VH和VL之间无所述任选的二硫键稳定结构。在一个实施方案中,多特异性抗体是一种三特异性或四特异性抗体,其包含a)特异性结合第一抗原的全长抗体的第一轻链和第一重链,和b)特异性结合第二抗原的全长抗体的第二(经修饰)轻链和第二(经修饰)重链,其中可变结构域VL和VH相互取代,和/或其中恒定结构域CL和CH1相互取代,和c)其中特异性结合一个或两个其他抗原(即第三和/或第四抗原)的一至四个抗原结合肽通过肽接头融合至a)和/或b)的轻链或重链的C-或N-端。a)中的抗体不包含如b)中所报告的修饰,和a)中的重链和轻链是分离的链。在一个实施方案中,三特异性或四特异性抗体包含c)中的一个或两个特异性结合一个或两个其他抗原的抗原结合肽。在一个实施方案中,抗原结合肽选自scFv片段和scFab片段。在一个实施方案中,抗原结合肽是scFv片段。在一个实施方案中,抗原结合肽是scFab片段。在一个实施方案中,抗原结合肽融合至a)和/或b)的重链的C-端。在一个实施方案中,三特异性或四特异性抗体包含c)中的一个或两个特异性结合一个其他抗原的抗原结合肽。在一个实施方案中,三特异性或四特异性抗体包含c)中的特异性结合第三抗原的两个相同的抗原结合肽。在一个优选的实施方案中,这样的两个相同的抗原结合肽都通过相同的肽接头融合至a)和b)的重链的C-端。在一个优选的实施方案中,两个相同的抗原结合肽是scFv片段或scFab片段。在一个实施方案中,三特异性或四特异性抗体包含c)中的特异性结合第三和第四抗原的两个抗原结合肽。在一个实施方案中,所述两个抗原结合肽都通过相同的肽连接子融合至a)和b)的重链的C-端。在一个优选的实施方案中,所述两个抗原结合肽是scFv片段或scFab片段。在一个实施方案中,多特异性抗体是双特异性、四价抗体,其包含a)特异性结合第一抗原的抗体的两条轻链和两个重链(和包含两个Fab片段),b)特异性结合第二抗原的抗体的两个额外的Fab片段,其中所述额外的Fab片段都通过肽接头融合至a)的重链的C-或N端,和其中,在Fab片段中进行了以下的修饰i)在a)的两个Fab片段中,或在b)的两个Fab片段中,可变结构域VL和VH相互取代,和/或恒定结构域CL和CH1相互取代,或ii)在a)的两个Fab片段中可变结构域VL和VH相互取代,和恒定结构域CL和CH1相互取代,和在b)的两个Fab片段中可变结构域VL和VH相互取代,或恒定结构域CL和CH1相互取代,或iii)在a)的两个Fab片段中可变结构域VL和VH相互取代,或恒定结构域CL和CH1相互取代,和在b)的两个Fab片段中可变结构域VL和VH相互取代,和恒定结构域CL和CH1相互取代,或ⅳ)在a)的两个Fab片段中可变结构域VL和VH相互取代,和在b)的两个Fab片段中恒定结构域CL和CH1相互取代,或v)在a)的两个Fab片段中恒定结构域CL和CH1相互取代,和在b)的两个Fab片段中可变结构域VL和VH相互取代。在一个实施方案中,所述额外的Fab片段都通过肽接头融合至a)的重链的C-端,或融合至a)的重链的N-端。在一个实施方案中,所述额外的Fab片段都通过肽接头融合至a)的重链的C-端。在一个实施方案中,所述额外的Fab片段都通过肽连接子融合至a)的重链的N-端。在一个实施方案中,在Fab片段中进行以下的修饰:i)在a)的两个Fab片段中,或b)的两个Fab片段中,可变结构域VL和VH相互替换,和/或恒定结构域CL和CH1相互替换。在一个实施方案中,在Fab片段中进行以下的修饰:i)在a)的两个Fab片段中,可变结构域VL和VH相互替换,和/或恒定结构域CL和CH1相互替换。在一个实施方案中,在Fab片段中进行以下的修饰:i)在a)的两个Fab片段中恒定结构域CL和CH1相互替换。在一个实施方案中,在Fab片段中进行以下的修饰:i)在b)的两个Fab片段中可变结构域VL和VH相互替换,和/或恒定结构域CL和CH1相互替换。在一个实施方案中,在Fab片段中进行以下的修饰:i)在b)的两个Fab片段中恒定结构域CL和CH1相互替换。在一个实施方案中,多特异性抗体是一种双特异性、四价抗体,其包含:a)特异性结合第一抗原的第一抗体的(经修饰)的重链,其包含第一VH-CH1结构域对,其中所述第一抗体的第二VH-CH1结构域对的N-端通过肽接头融合至所述重链的C-端,b)所述a)的第一抗体的两条轻链,c)特异性结合第二抗原的第二抗体的(经修饰)的重链,其包含第一VH-CL结构域对,其中所述第二抗体的第二VH-CH1结构域对的N-端通过肽接头融合至所述重链的C-端,和d)所述c)的第二抗体的两条(经修饰)的轻链,每条包含CL-CH1结构域对。在一个实施方案中,多特异性抗体是双特异性抗体,其包含a)特异性结合第一抗原的第一全长抗体的重链和轻链,和b)特异性结合第二抗原的第二全长抗体的重链和轻链,其中重链的N-端通过肽接头连接到轻链的C-端。a)中的抗体不包含如b)所报道的修改,且重链和轻链是分离的链。在一个实施方案中,多特异性抗体是双特异性抗体,其包含a)特异性结合第一抗原的全长抗体,其由两条抗体重链和两条抗体轻链组成,和b)特异性结合第二抗原的Fv片段,其包含VH2结构域和VL2结构域,其中两个结构域通过二硫键桥相互连接,其中只有VH2结构域或VL2结构域通过肽接头融合至特异性结合第一抗原的全长抗体的重链或轻链。在双特异性(抗体)中a)中的重链和轻链是分离的链。在一个实施方案中,VH2结构域或VL2结构域的另一个不通过肽接头融合至特异性结合第一抗原的全长抗体的重链或轻链。在如本文所报告的所有方面,第一轻链包含VL结构域和CL结构域,第一重链包含VH结构域、CH1结构域、铰链区、CH2结构域和CH3结构域。在一个实施方案中,多特异性抗体是双特异性三价抗体,其包含a)特异性结合第一抗原的两个Fab片段,b)一个CrossFab片段,其特异性结合其中CH1和CL结构域彼此交换的第二抗原,c)包含第一Fc区重链和第二Fc区重链的一个Fc区,其中两个Fab片段的CH1结构域的C-端连接至重链Fc-区多肽的N-端,并且其中CrossFab片段的CL结构域的C-端连接至Fab片段之一的VH结构域的N-端。在一个实施方案中,多特异性抗体是双特异性三价抗体,其包含a)特异性结合第一抗原的两个Fab片段,b)一个CrossFab片段,其特异性结合其中CH1和CL结构域彼此交换的第二抗原,c)包含第一Fc区重链和第二Fc区重链的一个Fc区,其中所述第一Fab片段的CH1结构域的C-端连接至所述重链Fc-区多肽之一的N-端,并且所述CrossFab片段的CL-结构域的C-端连接至所述另一个重链Fc-区多肽的N-端,并且其中第二Fab片段的CH1结构域的C-端连接至第一Fab片段的VH结构域的N-端或与CrossFab片段的VH结构域的N-端连接。在一个实施方案中,多特异性抗体是双特异性抗体,其包含a)特异性结合第一抗原并由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合第二抗原的Fab片段,其包含VH2结构域和VL2结构域,其包含重链片段和轻链片段,其中在轻链片段内可变轻链结构域VL2被所述抗体的可变重链结构域VH2置换,和在重链片段内可变重链结构域VH2被所述抗体的可变轻链结构域VL2置换,其中重链Fab片段插入全长抗体重链之一的CH1结构域和全长抗体的相应Fc-区之间,并且轻链Fab片段的N-端与全长抗体的轻链的C-端缀合,所述轻链与已插入重链Fab片段的全长抗体的重链配对。在一个实施方案中,多特异性抗体是双特异性抗体,其包含a)特异性结合第一抗原并由两条抗体重链和两条抗体轻链组成的全长抗体,和b)特异性结合第二抗原的Fab片段,其包含VH2结构域和VL2结构域,其包含重链片段和轻链片段,其中在轻链片段内可变轻链结构域VL2被所述抗体的可变重链结构域VH2置换,和在重链片段内可变重链结构域VH2被所述抗体的可变轻链结构域VL2置换其中Fab片段的重链片段的C-端与全长抗体的一条重链的N-端缀合,并且Fab片段的轻链片段的C-端缀合至全长抗体的轻链的N-端,该轻链与Fab片段的重链片段所缀合的全长抗体的重链配对。在所有方面的一个实施方案中,如本文报告的抗体是多特异性抗体,其需要至少两条重链多肽的异源二聚化。已经描述了一些为支持异源二聚化的CH3修饰的方法,例如在WO96/27011,WO98/050431,EP1870459,WO2007/110205,WO2007/147901,WO2009/089004,WO2010/129304,WO2011/90754,WO2011/143545,WO2012/058768,WO2013/157954,WO2013/096291中。通常,在本领域已知的方法中,第一重链的CH3结构域和第二重链的CH3结构域均以互补的方式工程化,使得含有一个工程化的CH3结构域的重链不再与另一相同结构的重链同源二聚化(例如CH3工程化的第一重链不再与另一CH3工程化的第一重链同源二聚化;和CH3工程化的第二重链不再与另一CH3工程化的第二重链同源二聚化)。从而包含一个工程化的CH3结构域的重链被强制与另一包含以互补方式工程化的CH3结构域的重链异源二聚化。对于这个实施方案,第一重链的CH3结构域和第二重链的CH3结构域通过氨基酸取代以互补方式工程化,使得第一重链和第二重链被强制异源二聚化,而第一重链和第二重链可以不再同源二聚化(例如由于位阻的原因)。用于支持本领域中已知的重链异源二聚化的不同方法(以上引用和包括的),预期为提供如本文报道的多特异性抗体中使用的不同替代,其包括源自特异性结合第一抗原的第一抗体的“非交叉Fab区”,和源自特异性结合第二抗原的第二抗体的“交叉Fab区”,组合上述的特定氨基酸取代。如本文所报告的多特异性抗体的CH3结构域可通过“杵臼”技术改变,该技术和一些实例详细描述于例如WO96/027011,Ridgway,J.B.,等人,ProteinEng.9(1996)617-621;和Merchant,A.M.,等人,Nat.Biotechnol.16(1998)677-681。在该方法中改变两个CH3结构域的相互作用表面以增加含有这两种CH3结构域的两条重链的异源二聚化。(两条重链的)两个CH3结构域的每个可以是“杵”,而另一个是“臼”。二硫键桥的引入进一步稳定异源二聚体(Merchant,A.M.,等人,NatureBiotech.16(1998)677-681;Atwell,S.,等人,J.Mol.Biol.270(1997)26-35),以及增加了产量。在一个优选的实施方案中,多特异性抗体包含在“杵链”的CH3结构域中的T366W突变和在“臼链”的CH3结构域中的突变T366S,L368A,Y407V(根据KabatEU索引编号)。也可使用CH3结构域之间的额外的链间二硫键桥(Merchant,A.M.,等人,NatureBiotechnol.16(1998)677-681),例如通过将Y349C突变引入“杵链”的CH3结构域和将E356C突变或S354C突变引入“臼链”的CH3结构域。因此,在另一个优选的实施方案中,多特异性抗体包含在两个CH3结构域之一中的突变Y349C和T366W和在两个CH3结构域的另一个中的突变E356C,T366S,L368A和Y407V,或多特异性抗体包含在两个CH3结构域之一中的突变Y349C和T366W和在两个CH3结构域的另一个中的突变S354C,T366S,L368A和Y407V(在一个CH3结构域中的额外Y349C突变和在另一个CH3结构域中的额外E356C或S354C突变形成链间二硫键桥)(根据KabatEU索引编号)。但是还可以替代地或额外地使用由EP1870459描述的其他杵-臼技术。在一个实施方案中多特异性抗体包含在“杵链”的CH3结构域中的突变R409D和K370E和在“臼链”的CH3结构域中的突变D399K和E357K(根据KabatEU索引编号)。在一个实施方案中多特异性抗体包含在“杵链”的CH3结构域中的突变T366W和在“臼链”的CH3结构域中的突变T366S,L368A和Y407V,以及额外地在“杵链”的CH3结构域中的突变R409D和K370E和在“臼链”的CH3结构域中的突变D399K和E357K(根据KabatEU索引编号)。在一个实施方案中,多特异性抗体包含在两个CH3结构域之一中的突变Y349C和T366W和在两个CH3结构域的另一个中的突变S354C,T366S,L368A和Y407V,或多特异性抗体包含在两个CH3结构域之一中的突变Y349C和T366W和在两个CH3结构域的另一个中的突变S354C,T366S,L368A和Y407V和额外地在“杵链”的CH3结构域中的突变R409D和K370E和在“臼链”的CH3结构域中的突变D399K和E357K(根据KabatEU索引编号)。除了“杵-臼技术”,用于修饰多特异性抗体重链的CH3结构域以增强异源二聚化的其他技术在本领域中是公知的。这些技术,尤其是在WO96/27011,WO98/050431,EP1870459,WO2007/110205,WO2007/147901,WO2009/089004,WO2010/129304,WO2011/90754,WO2011/143545,WO2012/058768,WO2013/157954和WO2013/096291中描述的那些,在此预期为“杵-臼技术”的替代与多特异性抗体组合。在一个实施方案中,在多特异性抗体中,在EP1870459中描述的方法被用于支持多特异性抗体的第一重链和第二重链的异源二聚化。这种方法基于将具有相反电荷的带电氨基酸引入第一和第二重链二者之间的CH3/CH3-结构域界面中的特定氨基酸位置。因此,在本实施方案中,在多特异性抗体的三级结构中第一重链的CH3结构域和第二重链的CH3结构域形成位于各抗体CH3结构域之间的界面,其中第一重链的CH3结构域的氨基酸序列和第二重链的CH3结构域的各自的氨基酸序列包括在抗体的三级结构中位于所述界面的一组氨基酸,其中从位于一条重链的CH3结构域的界面中的氨基酸组中,将第一氨基酸用带正电的氨基酸取代,和从位于另一条重链的CH3结构域的界面中的氨基酸组中,将第二氨基酸用带负电的氨基酸取代。根据本实施方案的多特异性抗体在本文也称为“CH3(+/-)-工程化的多特异性抗体”(其中缩写“+/-”表示在各CH3结构域中引入的带相反电荷的氨基酸)。在CH3(+/-)-工程化的多特异性抗体的一个实施方案中,带正电的氨基酸选自K、R和H,带负电的氨基酸选自E或D。在CH3(+/-)-工程化的多特异性抗体的一个实施方案中,带正电的氨基酸选自K和R,带负电的氨基酸选自E或D。在CH3(+/-)-工程化的多特异性抗体的一个实施方案中,带正电的氨基酸是K,带负电的氨基酸选自是E。在CH3(+/-)-工程化的多特异性抗体的一个实施方案中,在一条重链的CH3结构域中第409位的氨基酸R被D取代和第位的氨基酸K被E取代,在另一条重链的CH3结构域中第399位的氨基酸D被K取代和第357位的氨基酸E被K取代(根据KabatEU索引编号)。在一个实施方案中,使用WO2013/157953中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在一个实施方案中,在一条重链的CH3结构域中第366位的氨基酸T被K取代,在另一条重链的CH3结构域中第351位的氨基酸L被D取代(根据KabatEU索引编号)。在另一实施方案中,在一条重链的CH3结构域中第366位的氨基酸T被K取代,第351位的氨基酸L被K取代,在另一条重链的CH3结构域中第351位的氨基酸L被D取代(根据KabatEU索引编号)。在另一实施方案中,在一条重链的CH3结构域中第366位的氨基酸T被K取代和第351位的氨基酸L被K取代,在另一条重链的CH3结构域中第351位的氨基酸L被D取代(根据KabatEU索引编号)。另外以下取代中的至少一个包括在另一条重链的CH3结构域中:第349位氨基酸Y被E取代,第349位氨基酸Y被D取代和第368位氨基酸L被E取代(根据KabatEU索引编号)。在一个实施方案中,第368位氨基酸L被E取代(根据KabatEU索引编号)。在一个实施方案中,使用WO2012/058768中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在一个实施方案中,在一条重链的CH3结构域中第351位的氨基酸L被Y取代和第407位的氨基酸Y被A取代,在另一条重链的CH3结构域中第366位的氨基酸T被A取代和第409位的氨基酸K被F取代(根据KabatEU索引编号)。在另一个实施方案中,除了以上所述的取代,在一条重链的CH3结构域中至少一个以下位置的氨基酸被取代:第411位(原来为T)、第399位(原来为D)、第400位(原来为S)、第405位(原来为F)、第390位(原来为N)和第392位(原来为K)(根据KabatEU索引编号)。优选的取代为:-由选自N,R,Q,K,D,E和W的氨基酸取代第411位的氨基酸T(根据KabatEU索引编号),-由选自R,W,Y和K的氨基酸取代第399位的氨基酸D(根据KabatEU索引编号),-由选自E,D,R和K的氨基酸取代第400位的氨基酸S(根据KabatEU索引编号),-由选自I,M,T,S,V和W的氨基酸取代第405位的氨基酸F(根据Kabat的EU索引编号);-由选自R,K和D的氨基酸取代第390位的氨基酸N(根据KabatEU索引编号);和-由选自V,M,R,L,F和E的氨基酸取代第392位的氨基酸K(根据KabatEU索引编号)。在多特异性抗体的另一个实施方案中(根据WO2012/058768工程化),在一条重链的CH3结构域中第351位的氨基酸L被Y取代和第407位的氨基酸Y被A取代,在另一条重链的CH3结构域中第366位的氨基酸T被V取代和第409位的氨基酸K被F取代(根据KabatEU索引编号)。在多特异性抗体的另一实施方案中,在一条重链的CH3结构域中第407位的氨基酸Y被A取代,在另一条重链的CH3结构域中第366位的氨基酸T被A取代和第409位的氨基酸K被F取代(根据KabatEU索引编号)。在上述实施方案中,在另一条重链的CH3结构域中第392位的氨基酸K被E取代,第411位的氨基酸T被E取代,第399位的氨基酸D被R取代和第400位的氨基酸S被R取代(根据KabatEU索引编号)。在一个实施方案中,使用WO2011/143545中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在一个实施方案中,在两条重链的CH3结构域中的氨基酸修饰在第368和/或409位引入(根据KabatEU索引编号)。在一个实施方案中,使用WO2011/090762中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。WO2011/090762涉及根据“杵臼”(KiH)技术的氨基酸修饰。在一个实施方案中,在一条重链的CH3结构域中第366位的氨基酸T被W取代,在另一条重链的CH3结构域中第407位的氨基酸Y被A取代(根据KabatEU索引编号)。在另一个实施方案中,在一条重链的CH3结构域中第366位的氨基酸T被Y取代,在另一条重链的CH3结构域中第407位的氨基酸Y被T取代(根据KabatEU索引编号)。在一个实施方案中,使用在WO2011/090762中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化,所述多特异性抗体为IgG2同种型的。在一个实施方案中,使用WO2009/089004中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在一个实施方案中,在一条重链的CH3结构域中第392位的氨基酸K或N被带负电的氨基酸取代(在一个实施方案中被E或D取代,在一个优选的实施方案中被D取代),在另一条重链的CH3结构域中第399位的氨基酸D第356位的氨基酸E或D或第357位的氨基酸E被带正电的氨基酸取代(在一个实施方案中被K或R取代,在一个优选实施方案中被K取代,在一个优选的实施方案中第399或356位的氨基酸被K取代)(根据KabatEU索引编号)。在一个进一步的实施方案中,除了上述取代,在一条重链的CH3结构域中第409位的氨基酸K或R被带负电的氨基酸取代(在一个实施方案中被E或D取代,在一个优选的实施方案中被D取代)(根据KabatEU索引编号)。在一个更进一步的实施方案中,除了或替代上述取代,在一条重链的CH3结构域中第439位的氨基酸K和/或第370位的氨基酸K彼此独立地被带负电的氨基酸取代(在一个实施方案中被E或D取代,在一个优选的实施方案中被D取代)(根据KabatEU索引编号)。在一个实施方案中,使用WO2007/147901中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在一个实施方案中,在一条重链的CH3结构域中第253位的氨基酸K被E取代、第282位的氨基酸D被K取代和第322位的氨基酸K被D取代,在另一条重链的CH3结构域中第239位的氨基酸D被K取代、第240位的氨基酸E被K取代和第292位的氨基酸K被D取代(根据KabatEU索引编号)。在一个实施方案中,使用WO2007/110205中描述的方法支持多特异性抗体的第一重链和第二重链的异源二聚化。在如本文所报告的所有方面和实施方案的一个实施方案中,多特异性抗体是双特异性抗体或三特异性抗体。在一个优选实施方案中,多特异性抗体是双特异性抗体。在如本文所报告的所有方面的一个实施方案中,所述抗体是二价或三价抗体。在一个实施方案中,抗体是二价抗体。在如本文所报告的所有方面的一个实施方案中,多特异性抗体具有IgG型抗体的恒定结构域结构。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于人亚类IgG1,或者属于具有突变L234A和L235A的人亚类IgG1。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于人亚类IgG2。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于人亚类IgG3。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于人亚类IgG4,或属于具有额外的突变S228P的人亚类IgG4。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于人亚类IgG1或人亚类IgG4。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于具有突变L234A和L235A的人亚类IgG1(根据KabatEU索引编号)。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于具有突变L234A、L235A和P329G的人亚类IgG1(根据KabatEU索引编号)。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于具有突变S228P和L235E的人亚类IgG4(根据KabatEU索引编号)。在如本文所报告的所有方面的另一实施方案中,多特异性抗体属于具有突变S228P、L235E和P329G的人亚类IgG4(根据KabatEU索引编号)。如本文报道的抗体在一个实施方案中属于带突变PVA236,L234A/L235A和/或GLPSS331(根据Kabat的EU索引编号)的人亚类IgG1或属于亚类IgG4。在进一步的实施方案中,抗体属于任何IgG类,在一个实施方案中属于IgG3或IgG4亚类,其含有E233,L234,L235,G236,D270,N297,E318,K320,K322,A327,A330,P331和/或P329(根据Kabat的EU索引编号)中至少一个突变。另外在一个实施方案中,抗体属于IgG4亚类并且含有突变S228P或突变S228P和L235E(Angal,S.等人,Mol.Immunol.30(1993)105-108)(根据Kabat的EU索引编号)。如本文报道的抗体重链的C-端可以是具有氨基酸残基PGK的完整C-端结尾。重链的C-端可以是其中一个或两个C-端氨基酸残基已经被去除的缩短的C-端。在一个优选的实施方案中,重链的C-端是缩短的C-端结尾PG。在如本文报告的全部方面的一个实施方案中,包含了包括如本文具体说明的C-端CH3结构域的重链的抗体包含C-端甘氨酸-赖氨酸二肽(G446和K447,根据KabatEU索引编号)。在如本文报告的全部方面的一个实施方案中,包含了包括如本文具体说明的C-端CH3结构域的重链的抗体包含C-端甘氨酸残基(G446,根据KabatEU索引编号)。提供以下实施例,序列和附图以帮助理解本发明,其真实范围在所附权利要求中阐述。应该理解,可以在不背离本发明精神的情况下对所述程序进行修改。附图说明图1:在双特异性抗体与呈递双特异性抗体的抗原/靶的重组细胞同时结合之后,本文报道的用于FRET-供体和-接受体之间FRET诱导的FRET测定的图。图2:PD-1或Tim3表达细胞(单次转染)的FACS结果:1:PD1-CLIP;2:PD1-SNAP;3:Tim3-CLIP;4:Tim3-SNAP。图3:PD-1和Tim3表达细胞(双重转染)(下行的点图)的FACS结果;各个同种型对照的染色显示在上行点图中。图4:在PD1和Tim3表达细胞上处理/结合不同双特异性抗PD1/Tim3抗体时FRET的诱导:使用BMGPherastar读数器在665/620nm处测量时间分辨荧光(绘示的是FRET信号的平均值+/-SD[比率665/620nm*10,000],n=3)A:1+1形式双特异性抗体(抗体PD1-Tim3-0389和PD1-Tim3-0168)和2+2形式双特异性抗体(抗体PD1-Tim3-0358和PD1-Tim3-0359);B:双特异性抗体PD1-Tim3-0476和PD1-Tim3-0477。图5:在PD1和Tim3表达细胞上处理/结合双特异性抗PD1/Tim3抗体后FRET的诱导:使用BMGPherastar读数器在665/620nm处测量时间分辨荧光(绘示的是FRET信号的平均值+/-SD[比率665/620nm*10,000],n=3);测试1+1形式双特异性抗体PD1-Tim3-0389和单特异性二价抗PD1抗体——仅双特异性抗体诱导FRET。图6:单独或作为混合物的单和双特异性抗体的FRET测定:菱形:单独的双特异性抗体PD1-Tim3-0168→FRET信号可见;方块:双特异性PD1-Tim3-0168和单特异性抗PD1抗体的混合物→没有可见的FRET信号;三角形:单独的单特异性、二价抗PD1单克隆抗体→没有可见的FRET信号。图7:单独或作为混合物的单和双特异性抗体的FRET测定:菱形:单独的双特异性抗体PD1-Tim3-0168→FRET信号可见;方块:双特异性PD1-Tim3-0168和单特异性抗Tim3抗体的混合物→没有可见的FRET信号。图8:使用双特异性抗体和不同浓度的单特异性抗PD1抗体的FRET测定→依赖于抗PD1抗体浓度和Tim3相互作用强度的FRET信号;测试了两种不同的双特异性抗PD1/Tim3抗体:PD1-Tim3-0168(黑色)和PD1-Tim3-0389(灰色)。材料和一般方法关于人免疫球蛋白轻链和重链的核苷酸序列的一般信息在以下给出:Kabat,E.A.,等人,SequencesofProteinsofImmunologicalInterest,第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991)。对抗体链的氨基酸的编号和提及根据Kabat编号(Kabat,E.A.,等人,SequencesofProteinsofImmunologicalInterest,第5版,PublicHealthService,NationalInstitutesofHealth,Bethesda,MD(1991))。重组DNA技术使用标准方法操作DNA,如Sambrook,J.等人,MolecularCloning:Alaboratorymanual;ColdSpringHarborLaboratoryPress,ColdSpringHarbor,NewYork,1989所述。分子生物学试剂根据制造商的说明使用。基因合成从化学合成生成的寡核苷酸制备期望的基因区段。通过退火和连接寡核苷酸(包括PCR扩增)装配长的基因区段,其侧翼有单数的(singular)限制性内切酶切割位点,和随后经由指定的限制性位点。亚克隆基因片段的DNA序列通过DNA测序证实。基因合成片段根据给定的说明在Geneart(Regensburg,Germany)定制。DNA序列的确定通过MediGenomixGmbH(Martinsried,Germany)或SequiServeGmbH(Vaterstetten,Germany)进行双链测序来确定DNA序列。DNA和蛋白质序列分析和序列数据管理GCG(GeneticsComputerGroup,Madison,Wisconsin)软件包版本10.2和Infomax'sVectorNT1高级套件8.0版用于序列创建、汇谱、分析、注释和说明。表达载体对于所描述双特异性抗体的表达,可以应用用于瞬时表达细胞(例如在HEK293中)的表达质粒,其或者基于具有或不具有CMV-内含子A启动子的cDNA组构(organization)、或者基于具有CMV启动子的基因组组构。除了抗体表达盒,载体包含:-复制起点,其允许该质粒在大肠杆菌中复制,和-β-内酰胺酶基因,其在大肠杆菌中赋予氨苄青霉素抗性。抗体基因的转录单位由以下元件组成:-在5'端的独特限制性位点(一个或多个),-来自人巨细胞病毒的即时早期增强子和启动子,-在cDNA组构的情况下内含子A序列,-源自人抗体基因的5'-非翻译区,-免疫球蛋白重链信号序列,-编码相应的抗体链的核酸,其或者作为cDNA或具有基因组外显子-内含子组构,-3'非翻译区,其具有聚腺苷酸化信号序列,和-在3'端的独特限制性位点(一个或多个)。通过PCR和/或基因合成生成编码抗体链的融合基因,通过已知的重组方法和技术通过连接相应的核酸区段(例如使用各自载体中的独特的限制性位点)来装配。通过DNA测序验证亚克隆的核酸序列。对于瞬时转染,通过质粒制备从转化的大肠杆菌培养物(NucleobondAX,Macherey-Nagel)制备大量质粒。对于所有构建体,在第一CH3结构域中使用具有典型的杵(T366W)取代以及第二CH3结构域中相应的臼取代(T366S,L368A和Y407V)(以及另外两个引入的半胱氨酸残基S354C/Y349'C)(包含在上述各个相应的重链(HC)序列中)的杵臼二聚化技术残基。细胞培养技术使用标准细胞培养技术,如CurrentProtocolsinCellBiology(2000),Bonifacino,J.S.,Dasso,M.,Harford,J.B.,Lippincott-Schwartz,J.andYamada,K.M.(编辑),JohnWiley&Sons,Inc.中描述的。在HEK293系统中瞬时转染双特异性抗体通过瞬时表达产生。因此,根据制造商的说明使用HEK293系统(Invitrogen)转染各个质粒。简言之,用相应的表达质粒和293fectinTM或fectin(Invitrogen)的混合物转染在摇瓶中的悬浮液中或者在搅拌发酵罐中的无血清FreeStyleTM293表达培养基(Invitrogen)中生长的HEK293细胞(Invitrogen)。对于2升摇瓶(Corning),HEK293细胞以600mL中1.0*106细胞/mL的密度接种,并在120rpm、8%CO2中孵育。第二天,将细胞以约1.5*106细胞/mL的细胞密度与约42mLA)和B)的混合物转染,其中A)20mLOpti-MEM培养基(Invitrogen),其包含600μg总质粒DNA(1μg/mL)和B)20mLOpti-MEM培养基,其补充了1.2mL293fectin或fectin(2μl/mL)。根据葡萄糖消耗在发酵过程中加入葡萄糖溶液。5-10天后收获含有分泌抗体的上清液,从上清中直接纯化抗体或将上清液冷冻保存。对于1+1CrossMab的1:1:1:1或对于2+2CrossMab的1:1:1的相对质粒比用于LC,HC,交叉LC和交叉HC质粒的共转染。蛋白质测定通过测定280nm处的光密度(OD)、利用摩尔消光系数确定纯化抗体和衍生物的蛋白质浓度,所述摩尔消光系数根据Pace,等人,ProteinScience,4(1995)2411-1423,基于氨基酸序列计算。上清中抗体浓度测定在细胞培养物上清液中的抗体和衍生物的浓度通过使用蛋白A琼脂糖珠(RocheDiagnosticsGmbH,Mannheim,Germany)的免疫沉淀估计。因此,60μL蛋白A琼脂糖珠在TBS-NP40(50mMTris缓冲液,pH7.5,补充有150mMNaCl和1%Nonidet-P40)中洗涤三次。随后,将1-15mL细胞培养上清液施加到在TBS-NP40中预平衡的蛋白A琼脂糖珠上。在室温下孵育1小时后,将珠在Ultrafree-MC-过滤柱(Amicon)上用0.5mLTBS-NP40洗涤一次,用0.5ml2×磷酸盐缓冲盐水(2xPBS,RocheDiagnosticsGmbH,Mannheim,Germany)洗涤两次,用0.5ml100mM柠檬酸钠pH5.0短暂地洗涤四次。通过添加35μlLDS样品缓冲液(Invitrogen)洗脱结合的抗体。分别将样品的一半与样品还原剂组合或保持未还原,在70℃下加热10分钟。因此,5-30μl施加到4-12%Bis-TrisSDS-PAGE(Invitrogen)(对于非还原SDS-PAGE,使用MOPS缓冲液,对于还原SDS-PAGE,使用MES缓冲液和抗氧化运行缓冲液添加剂(Invitrogen)),并用考马斯亮蓝染色。通过亲和HPLC色谱法定量测定细胞培养物上清液中的抗体的浓度。简言之,在AgilentHPLC1100系统上,将含有结合于蛋白A的抗体的细胞培养物上清液施加到200mMKH2PO4、100mM柠檬酸钠,pH7.4中的AppliedBiosystemsPorosA/20柱上,用200mMNaCl、100mM柠檬酸,pH2.5洗脱。洗脱的抗体通过UV吸光度和峰面积积分进行定量。纯化的标准IgG1抗体作为标准。可替代地,通过Sandwich-IgG-ELISA测定细胞培养物上清液中抗体和衍生物的浓度。简言之,用100μL/孔的生物素化抗人IgG捕获分子F(ab')2<h-Fcγ>BI(Dianova)0.1μg/mL,在室温下放置1小时或备选地在4℃过夜包被StreptaWell高结合链霉亲和素A-96孔微量滴定板(RocheDiagnosticsGmbH,Mannheim,Germany),随后用200μL/孔PBS、0.05%Tween(PBST,Sigma)洗涤三次。之后,在PBS(Sigma)中系列稀释的各含有抗体的细胞培养物上清液100μL/孔加入到孔中,并在室温下在摇床上孵育1-2小时。用200μL/孔PBST洗涤孔三次,用100μlF(ab‘)2<hFcγ>POD(Dianova),0.1μg/mL作为检测抗体在室温在摇床上通过孵育放置1-2小时,检测结合的抗体。用200μL/孔PBST三次洗涤来移除未结合的检测抗体,并通过加入100μLABTS/孔检测结合的检测抗体,之后孵育。在Tecan氟分光光度计上在405nm测量波长下进行吸光度的测定(参考波长492nm)。制备抗体纯化参照标准流程从过滤的细胞培养物上清液中纯化蛋白。简言之,将抗体施加到蛋白ASepharose柱(GEHealthcare)上,并用PBS洗涤。抗体洗脱液在pH2.8获得,随后立即中和。通过尺寸排阻色谱法(Superdex200,GEHealthcare)在PBS或在20mM组氨酸、包含150mMNaCl的缓冲液(pH6.0)中从单体抗体分离聚集的蛋白质。合并单体抗体级分,(如需要)用例如MILLIPOREAmiconUltra(30MWCO)离心浓缩机浓缩,冷冻储存在-20℃或-80℃。提供部分样品用于随后的蛋白分析和分析表征,例如通过SDS-PAGE、尺寸排阻层析(SEC)或质谱。SDS-PAGE根据制造商的指示使用Pre-Cast凝胶系统(Invitrogen)。特别是,使用10%或4-12%Bis-TrisPre-Cast凝胶(pH6.4)和(还原凝胶,具有抗氧运行缓冲液添加剂)或MOPS(非还原凝胶)运行缓冲液。CE-SDS使用微流体Labchip技术(PerkinElmer,USA)通过CE-SDS分析纯度和抗体完整性。因此,根据制造商的说明使用HTProteinExpress试剂盒制备5μl抗体溶液用于CE-SDS分析,并使用HT蛋白表达芯片在LabChipGXII系统上分析。使用LabChipGX软件分析数据。分析型尺寸排阻色谱法通过HPLC色谱法进行用于确定抗体聚集和寡聚状态的尺寸排阻色谱法(SEC)。简言之,将蛋白A纯化的抗体施加到Dionex系统(ThermoFischerScientific)的300mMNaCl、50mMKH2PO4/K2HPO4缓冲液(pH7.5)中的TosohTSKgelG3000SW柱上,或DionexHPLC系统的2×PBS中的Superdex200柱(GEHealthcare)上。通过UV吸光度和峰面积积分定量洗脱的抗体。BioRad凝胶过滤标准151-1901作为标准。质谱本部分描述双特异性抗体的特征,重点在于它们的正确组装。预期的一级结构通过脱糖基化完整抗体的电喷雾电离质谱(ESI-MS)以及脱糖基化/限制性LysC消化抗体的特殊情况中进行分析。在37℃将抗体在磷酸盐或Tris缓冲液中以1mg/ml的蛋白质浓度用N-糖苷酶F脱糖基化高达17h。用100μg在Tris缓冲液(pH8)中的脱糖基化抗体分别在室温下进行120小时或在37℃进行40min的限制性LysC(RocheDiagnosticsGmbH,Mannheim,Germany)消化。在质谱法之前,样品通过HPLC在SephadexG25柱(GEHealthcare)上脱盐。在装有TriVersaNanoMate源(Advion)的maXis4GUHR-QTOFMS系统(BrukerDaltonik)上通过ESI-MS确定总质量。实施例1与PD1和Tim3结合的多特异性抗体的产生和表达如常规方法部分所述通过经典分子生物学技术产生结合人PD1和人Tim3的多特异性(双特异性)抗体,并如上所述在HEK293细胞中瞬时表达。在一个结合臂(1+1双特异性抗体形式)中具有VH/VL结构域交换/替换的双特异性抗体(CrossMAbVH-VL)所使用的双特异性1+1抗体形式也描述于WO2009/080252中。双特异性抗体使用含有编码下表所示氨基酸序列的核酸的表达质粒来表达。表:双特异性抗体(1+1CrossMAbVH-VL)的轻链(LC)和重链(HC)的氨基酸序列。1+1抗体HC1HC2LC1LC2PD1-Tim3-0389SEQIDNO:01SEQIDNO:02SEQIDNO:03SEQIDNO:04PD1-Tim3-0168SEQIDNO:05SEQIDNO:06SEQIDNO:07SEQIDNO:08PD1-Tim3-0476SEQIDNO:09SEQIDNO:10SEQIDNO:11SEQIDNO:12PD1-Tim3-0477SEQIDNO:13SEQIDNO:14SEQIDNO:15SEQIDNO:16PD1-Tim3-0166SEQIDNO:17SEQIDNO:18SEQIDNO:19SEQIDNO:20对于所有构建体,使用杵臼异二聚化技术,具有在第一CH3结构域中典型的杵(T366W)取代以及第二CH3结构域中的相应臼置换(T366S,L368A和Y410V)(以及另外两个引入的半胱氨酸残基S354C/Y349C)(包含在上面描述的各个相应的重链(HC)序列中)。在两个结合臂(2+2双特异性抗体形式)中具有VH/VL结构域交换/替换(2+2CrossMAbVH-VL)的双特异性抗体所使用的多特异性2+2抗体形式也描述于WO2010/145792中。双特异性抗体使用含有编码下表所示氨基酸序列的核酸的表达质粒来表达。表:双特异性抗体(2+2CrossMAbVH-VL)的轻链(LC)和重链(HC)的氨基酸序列。2+2抗体HCLC1LC2PD1-Tim3-0358SEQIDNO:21SEQIDNO:22SEQIDNO:23PD1-Tim3-0359SEQIDNO:24SEQIDNO:25SEQIDNO:26PD1-Tim3-0321SEQIDNO:27SEQIDNO:28SEQIDNO:29实施例2与PD1和Tim3结合的多特异性抗体的纯化和表征通过制备型蛋白A亲和色谱和制备型尺寸排阻色谱的组合从上清液中纯化上述表达的双特异性抗体。所有双特异性抗体可以以良好的收率生产并且稳定。所得产物通过质谱法表征身份以及表征诸如纯度(通过SDS-PAGE)、单体含量和稳定性等分析性质。质谱预期的一级结构通过脱糖基化的完整双特异性抗体和脱糖基化/纤溶酶消化的或替代地脱糖基化/限制性LysC消化的双特异性抗体的电喷雾电离质谱(ESI-MS)分析。在磷酸盐或Tris缓冲液中,双特异性抗体在37℃下以1mg/ml的蛋白质浓度用N-糖苷酶F脱糖基化高达17h。用100μg脱糖基化的双特异性抗体在Tris缓冲液(pH8)中分别进行在室温下120小时或在37℃40min的纤溶酶或限制性LysC(RocheDiagnosticsGmbH,Mannheim,Germany)消化。在质谱法之前,样品通过HPLC在SephadexG25柱(GEHealthcare)上脱盐。在装有TriVersaNanoMate源(Advion)的maXis4GUHR-QTOFMS系统(BrukerDaltonik)上通过ESI-MS确定总质量。实施例3用于双特异性抗体与重组细胞同时结合的FRET测定法概述在该实施例中,描述了基于细胞的TR-FRET测定来测定双特异性抗体形式与存在于一个细胞的外膜(表面)上的两种不同靶(例如受体)的同时结合。技术已被选为典型的FRET方法,但任何其他FRET方法都可以同样选择。它是经典TR-FRET(时间分辨荧光共振能量转移)和SNAP-标签技术(例如NewEnglandBiolabs,CISBIO)的组合,其允许存在于细胞表面的抗原被荧光供体或接受体染料标记。利用该测定形式,可以证明双特异性抗体同时结合表达双特异性抗体的两种抗原的细胞,在这种情况下是PD1和Tim3接受体。抗原呈现为由给定接受体的细胞外结构域(ECD)和标签组成的重组融合蛋白,可以向其添加荧光染料。在抗PD1/Tim3双特异性抗体的存在下,该抗体可以结合两种接受体,从而间接地结合FRET-供体和FRET-接受体,这是在同时结合两种接受体以及同样地FRET-供体和FRET-接受体将接近以允许能量转移(FRET)时(参见图1)。使用细胞内方法和两种以细胞外方式工作的双特异性抗体,导致无FRET诱导。膜渗透染料(在这种方法中必须使用)的背景非常高(数据未示出)。重组PD1+Tim3+HEK细胞的产生使用标准方法产生DNA,如Sambrook等人,Molecularcloning:Alaboratorymanual;ColdSpringHarborLaboratoryPress,ColdSpringHarbor,NewYork,1989。为了克隆PD1和Tim3变体,将SNAP或CLIP插入到接受体的相应跨膜区域的近侧,除了7个氨基酸残基以外,移除细胞质结构域,并用Flag标签替换。瞬时转染HEK293细胞在30ml培养体积中用转染试剂293free(Novagen)和还原血清培养基(LifeTechnologies)一次使用两种质粒翊15μg总量的DNA共转染。简而言之,如下别处所述,用下列编码分别由PD1或Tim3ECD和SNAP或CLIP标签组成的融合蛋白的质粒瞬时转染HEK293细胞:质粒:a)PD1-SNAP(5'至3'方向):-编码人PD1-细胞外结构域的核酸,包括信号肽(SEQIDNO:30的残基1-170),-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码来自pSNAP-标签(T7)2(无N-末端甲硫氨酸残基)的SNAP的核酸,所述标签是O6-烷基鸟嘌呤-DNA-烷基转移酶(hAGT)的人基因的突变形式。(与野生型hAGT相比,SNAP-标签蛋白含有突变C26A,K125A,A127T,R128A,G131K,G132T,M134L,R135S,C150S,N157G,S159E,并且在G182后截短(SEQIDNO:32),-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码人PD1-跨膜和细胞质结构域(SEQIDNO:30的残基171-191)的核酸,-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码Flag标签(DYKDDDDK;SEQIDNO:33)的核酸。b)Tim3-CLIP(5'至3'方向):-编码人Tim3胞外结构域(SEQIDNO:34的残基1-202)的核酸,所述结构域包括信号肽-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码来自pCLIPf的CLIP的核酸(无N-末端甲硫氨酸残基),所述CLIP是O6-烷基鸟嘌呤-DNA-烷基转移酶(hAGT)的人基因的突变形式(SEQIDNO:35),-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码人Tim3跨膜和胞质结构域(SEQIDNO:34的残基203-230)的核酸-编码GGGGS间隔区(SEQIDNO:31)的核酸,-编码Flag标签(DYKDDDDK;SEQIDNO:33)的核酸。还构建并表达了PD1-CLIP和Tim3-SNAP的组合,并且导致只有低但仍然可检测的FRET信号(数据未显示)。转染后,将细胞在摇瓶中孵育直至FACS(转染后24-48小时)或FRET实验(48小时后)的最终使用。在瞬时转染的HEK293细胞上的PD1和Tim3表达确认(通过FACS)在瞬时转染HEK293细胞24-48小时后,分析细胞的PD1和Tim3表达:通常,将1-3x105个单或双转染的细胞以10μg/ml在冰上染色30min,用PBS/2%FCS洗涤两次,并在FacsCantoII上使用以下进行分析-FITC缀合的抗人PD1抗体(Biolegend,目录号329904,克隆EH12.2H7)或藻红蛋白(PE)缀合的抗人PD-1抗体(R&DSystems,目录号FAB1086P),和/或-藻红蛋白(PE)缀合的抗人Tim3抗体(R&Dsystems,目录号FAB2365P,克隆344823)。单转染细胞的FACS结果如图2所示,双转染细胞的结果如图3所示。用抗PD1/Tim3双特异性抗体描述细胞标记和FRET测定将转染的细胞沉淀并以1×106细胞/ml的密度重悬于缓冲液(Cisbio)中。此后,在37℃下在缓冲液(Cisbio)中用100nMSNAP-Lumi4-Tb(Cisbio)和100nMCLIP-Red(Cisbio)对细胞染色1小时。洗涤并重悬于包含2%FCS(v/v)的PBS缓冲液中后,将约50,000个细胞(以50μl体积)接种到96孔平底白色平板(Costar)中,对照抗体(例如二价,单特异性抗体或同种型对照)或双特异性抗体以0.001-10nM的终浓度添加到细胞中。在4℃或室温孵育1小时后,使用BMGPherastar读数器或TecanInfiniteM1000Pro,使用由相应的供应商提供标准设置,以665nm和620nm的发射信号的比率(665/620nm比)测量时间分辨荧光。示例性标准设置如下:-模式荧光强度顶-激发波长340nm-发射波长620/665nm-激发带宽20nm-发射带宽10nm-增益232最佳(100%)-闪光次数100-闪光频率100Hz-积分时间500μs-滞后时间60μs-固定时间(SettleTime)0ms任选地,将SNAP-Lumi4-Tb和100nMCLIP-Red标记的细胞储存在80℃或液氮中并新鲜解冻以进行FRET实验。或者,在一些情况下,经由山羊抗人Fc区抗体(20nM终浓度)交联肠胃外,单价或二价单特异性单克隆抗体是可能的。为了显示通过双特异性抗体的同时结合诱导的FRET反应的特异性,仅添加一种特异性的单克隆IgG用于竞争。结果如上所述处理表达PD1和Tim3的HEK细胞,以通过与滴定量的不同双特异性抗体(0.12nM-10nM)孵育,在同时受体结合时测量FRET信号。所有测试的双特异性抗体以剂量依赖性方式在PD1/Tim3表达细胞中诱导FRET信号。所有测试的双特异性抗体形式(1+1,2+2和2+1(数据未示出))是可比较的。在图4A中示出了用1+1形式双特异性抗体(抗体PD1-Tim3-0389和PD1-Tim3-0168)和2+2形式双特异性抗体(抗体PD1-Tim3-0358和PD1-Tim3-0359)获得的结果的比较。还评估了两种其他双特异性抗体PD1-Tim3-0476和PD1-Tim3-0477在处理后它们在细胞中诱导FRET的能力。如图4B所示,两种双特异性抗体在PD1+Tim3+HEK细胞中诱导显著的FRET信号,强调以功能性方式同时结合。同样如图5所示,如果仅添加单特异性二价抗体,则FRET测定不起作用,而对于双特异性抗体(抗体PD1-Tim3-0389),可以观察到FRET信号。为了显示通过双特异性抗体的同时结合诱导的FRET信号的特异性,仅添加一种特异性的单克隆IgG用于竞争。因此,用100nMSNAP-Lumi4-Tb和100nMCLIP-Red标记如前所述的表达PD1和Tim3的HEK细胞。洗涤后,将标记的细胞与指定浓度0.01nM,0.05nM,0.1nM,0.5nM,1nM,5nM,10nM和40nM的双特异性抗PD1/Tim3抗体PD1-Tim3-0168在4℃下一起孵育一小时,然后使用BMGPherastar读数器(图6,菱形)在665/620nm处测量时间分辨荧光。检测到FRET信号。为了突出双特异性抗体处理后FRET信号的特异性,单独测试单特异性二价抗-PD1单克隆抗体(图6,三角形)。没有检测到FRET信号。如果存在单特异性二价抗PD1单克隆抗体和双特异性抗PD1/Tim3抗体(80nM单特异性抗体浓度),则检测不到FRET信号(图6,正方形)。同样如果添加二价单特异性抗Tim3抗体则不能获得FRET信号(见图7)。实施例4用于测定结合强度的FRET测定法为了区分双特异性抗体的两个结合位点的贡献,如下所述进行竞争实验。为了区分PD1结合位点和Tim3结合位点对整体相互作用的贡献,使用了特殊设置。PD1结合位点比Tim3结合位点具有更高的亲和力。不同的双特异性抗体在标准条件下显示出类似的FRET信号强度。当PD1相互作用(即以较高亲和力/较低KD值的相互作用)被阻断或至少减少时,可以看到差异。因此,将不同浓度(80nM,20nM,5nM,1.25nM和0.31nM)的单特异性二价抗-PD1抗体添加到在Tim3结合位点不同的双特异性抗PD1/Tim3抗体(抗体PD1-Tim3-0168和PD1-Tim3-0389)。在完全PD1中和时,几乎没有测量到FRET信号,而在较低的抗PD1抗体浓度下,FRET信号恢复并且对抗体PD1-Tim3-0168观察到更强的信号,其中Tim3结合位点对Tim3具有更高的亲和力(参见图8)。当前第1页1 2 3
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1