一种冻害芦苇草青贮方法与流程
本发明涉及青贮技术领域,特别涉及一种接种外源lab优化冻害芦苇草青贮的方法。
背景技术:
青藏高原,大约70%的是高海拔、寒冷的牧场,由于天然、极端、不稳定的气候和自然环境,很少有农作物适合种植。其中一种替代作物是芦苇草,例如虉草(phalarisarundinaceal.),这是一种高产的冷季草种,比该地区的燕麦和其他当地草更具生产力。由于恶劣的天气条件和放牧的管理不善,当环境温度低于0℃并且持续4到5个小时,芦苇草的茎和叶就会发生冻害,导致其营养物质迅速减少。霜冻损坏的芦苇草(reedcanarygrass,rcg)通常可用作当地动物饲料成分,尤其适用于冬季和早春的牦牛。日本,加拿大和美国的许多研究表明,rcg在用于厌氧消化池生产甲烷之前可以作为青贮饲料保存。然而,这些研究是在夏季rcg的最佳生长季节进行的,很少有关于霜冻死亡后收获的rcg的青贮发酵研究。
实际上,由于青贮的操作程序不当或者其他因素,使得青贮饲料产品的可储存性和稳定性达到要求仍然是一项艰巨任务。附生外源菌不足、有益原生细菌(初级lab)数量减少、冻害后样品上腐败生物(如酵母和霉菌)数量较多等,均可引起青贮饲料发酵不良。因此,研究外源菌种的种类和添加量,是提高青贮期间的发酵速度及有氧稳定性的重要途径。
技术实现要素:
本发明的主要目的是针对目前青藏高原冻害芦苇草利用率低、青贮过程和产品质量不稳定的问题,提出一种冻害芦苇草青贮工艺,通过接种外源lab,优化冻害芦苇草青贮发酵过程,降低成本、提高青藏高原天然芦苇草利用率。同时,研究在青海、青藏高原等地区无外源或有外源条件下芦苇草青贮的特征和细菌群落等指标,提高青贮期间的发酵速度、有氧稳定性、产品适口性及存放周期。最后本发明还提供了一种该方法制备的青贮饲料,其品质稳定,营养成分含量高,适口性好,存放周期长。
为了实现上述发明目的,本发明采用的技术方案如下:
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab,比率为90-120cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的环境下青贮160-200天。
所述步骤(3)接种的外源lab是植物乳杆菌chikuso-1或l.buchneri的一种或两种。
植物乳杆菌被广泛地应用在其他品种原料青贮饲料发酵中,它可以迅速增加乳酸的含量以降低青贮饲料的ph值。一些研究表明,霜冻后收获的青贮材料表现出低水平的lab和大量的酵母和霉菌。而另一方面,频繁的霜冻可以破坏植物细胞并释放可利用的底物从而引起植物微生物组迅速繁殖。而专性的异型发酵lab如l.buchneri,可以在厌氧条件下缓慢地将乳酸转化为乙酸和丙酸。青贮饲料最终发酵产品受微生物种群影响,并且有氧暴露等情况的后续补救调整措施,也可以通过研究微生物种群来获得启示。
作为优选,所述步骤(3)是将步骤(2)得到的新鲜材料接种外源lab,比率为106cfu/g,彻底混合,得到青贮材料。
经过反复试验和数据统计分析,以每克新鲜材料106cfu的比率,将青贮材料接种外源lab可以取得较好的发酵结果。
所述步骤(4)是在环境温度低于20℃的9月至次年3月青贮180天。
发酵过程要做一定监控,避免低含量的有益菌种数和高含量的不良微生物(包括好氧细菌,酵母和霉菌)分布在植物上。
本发明充分利用当地(青藏高原等我国西部高原)自然环境发酵,由于乳杆菌,小球菌,weissellaspp的高分布,加之本发明所述工艺,rcg青贮饲料得以发酵良好。
一种所述冻害芦苇草青贮方法制备的青贮饲料。
后续对于鲜料原料、rcg原料及青贮产品的检测显示,较鲜料原料,rcg原料的水溶性碳水化合物(wsc)含量降低,而青贮后中性洗涤纤维(ndf)含量增加,这是由于在厌氧条件下植物本身存在的和外源微生物将大量可利用的底物转化为代谢产物(主要是有机酸,如乳酸和乙酸),有效提高所得青贮饲料中有机酸的总含量,降低氨态氮的含量,从而有利于改善青贮饲料的适口性,提高营养价值。单宁的存在被认为是有利的组分,因为它们通过抑制植物和微生物酶和/或通过与蛋白质形成复合物来保护饲料蛋白质免于降解,同样降低氨态氮的含量。青贮后剩余的总单宁持续相对较高的水平。然而,与对照相比,外源lab接种后青贮饲料的总单宁含量显著下降(p<0.05)。从理论上讲,粗蛋白(cp)含量(tn×6.25)不直接受发酵影响(氨氮包括在tn中),但随着青贮饲料中气体和废水流失的增加而呈线性下降(ferreira等,15al。,2013;santos等,2014)。本发明中,青贮发酵后霜冻rcg的cp含量下降。接种lb增加(p<0.05)乙酸和丙酸含量,降低(p<0.05)丁酸和氨氮含量,增强了整个作物中粗蛋白(cp)和醚提取物(ee)的保存。所以接种外源lab可以增强青贮饲料中营养成分的保存,还能显著降低乙醇的含量,抑制青贮饲料中真菌的生长,提高其有氧稳定性,从而有利于延长青贮饲料的保存期限。
在外源lab对青贮饲料ph和发酵产物的影响方面,先前的研究表明,与正常青贮饲料相比,霜冻损坏的青贮饲料表现出较差的发酵特性,乳酸含量降低,ph值增加,丁酸和氨氮含量增加(mohammadzadeh等,2014)。外源lab接种的主要目的是通过消耗wsc产生大量的乳酸以达到增强青贮ph降低的效果。对于外源lab接种的青贮饲料观察到的最低ph值(3.93,平均值)表明这些微生物对糖的利用效率更高。青贮饲料中乙酸的推荐含量为10-30g/kgdm。高水平的乙酸(>30-40g/kgdm)可能导致低dm回收率和能量(kung和shaver,2001;kung等,2018)。然而,太低的乙酸浓度会在暴露于空气时产生不稳定的青贮饲料状态。在本发明中,对照和lp接种的青贮饲料中的乙酸含量(4.8至7.3g/kgdm)表明维持有氧稳定性的可能性较差。然而,与对照相比,lb的接种确实增加了(p<0.05)乙酸和丙酸的浓度,并且降低了青贮饲料中丁酸,乙醇和氨-n的水平(p<0.05)。霜冻损坏的rcg青贮饲料的ph值(3.88-4.16)和乳酸(34.9-40.3g/kgdm)和乙酸(4.8-14.3g/kgdm)浓度相对于(contreras-govea)的数据较低等人,2009)他们报道了在美国从茎秆伸长到早期穗收获的正常青贮rcg的ph值正常为4.58-4.87,乳酸为45.8-63.4g/kdm和乙酸为13.2-28.1g/kgdm。(kungetal.,2018)综述并得出结论,当青贮饲料干物质为25-35%时,发酵良好的草料青贮饲料具有无法检测到的丁酸、丙酸,含量小于1g/kgdm和氨氮的含量为80-120g/kgdm等的特征。青贮饲料中丙酸(1.0-4.7g/kgdm)、丁酸(4.6-8.0g/kgdm)、氨氮(104.2-127.0g/kgtn)含量较高,青贮饲料发酵效果较差。因此,实验室外源接种对冻害rcg青贮发酵的改善作用有限。
在外源lab对青贮平板培养的微生物种群的影响方面,微生物种群与青贮饲料发酵之间存在很强的相关性(mcdonald等,1991)。与新鲜牧草相比,lab的数量增加了74.60-82.93%,而青贮饲料的好氧菌和酵母菌的数量在180天的青贮期后减少了19.30-65.22%。与对照相比,lb的接种增加了青贮饲料中的lab数量。类似的结果来自(da11silva等,2018)。牧草种类的相关效应和布氏乳杆菌的接种率不同会引起结果的不同。以前的研究表明,高水平的乙酸可以抑制青贮饲料中酵母的活性,因为它能够以非解离形式穿透细胞膜,随后释放细胞质中的h+,然后破坏细胞核中的dna结构(romero等人,2017;tabacco等,2011)。然而,在这项研究中,基于培养的技术的高酵母数(3.2-4.6cfu/gfm)仍在所有青贮中。在其他青贮中观察到类似的酵母计数(zhou等人,2016;19xing等人,2009)。这可能是因为(1)相对较低的乙酸(<20g/kg20dm,青贮水分约为57%)不能抑制某些酵母物种的活性,特别是对于乳酸同化酵母;(2)当青贮饲料中存在挥发性脂肪酸(特别是乙酸和丙酸)时,可以抑制生产酵母孢子;(3)然后是某些酵母,如醋酸杆菌属(acetobacterspp),可能在青贮期间由于l.buchneri产生乙酸而刺激和繁殖。与青贮饲料相比,叶青贮具有较低的lab数量和较高数量的酵母和霉菌,表明暴露恶化的可能性很高。(3)然后是某些酵母,如醋酸杆菌属(acetobacterspp),可能在青贮期间由于l.buchneri产生乙酸而刺激和繁殖。与青贮茎秆饲料相比,叶青贮具有较低的lab数量和较高数量的酵母和霉菌,这表明暴露恶化的可能性很高。
在外源lab对青贮细菌群落组成的影响方面,覆盖率>0.99表明采样时的大多数细菌群落组成已被深深捕获。在新鲜牧草中观察到相对较高的细菌otu(72至117),丰富度(chao1,70至119;ace,76至129)和多样性(shannon,3.69至5.51;simpson,0.86至0.96),并且这些在整个作物中青贮后减少35.10%至64.10%,32.77%至64.71%,44.96%至64.34%,31.50%至55.17%及7.29%至36.46%。(guan等人,2018;li等人,2019)报道了类似的结果,但是在其他报告中没有报道(ni等人,2017;zhao等人,2017)。这可以通过青贮期间ph降低的速率和程度来解释。与对照相比,外源lab的接种均降低了整个作物和茎青贮中ace和chao1的细菌丰富度指数(p<0.05)。与对照相比,在用lab处理的玉米青贮饲料中也观察到较低的细菌多样性(ogunade等,2018)。特别是lb的接种减少了(p<0.05)细菌otu和青贮的shannon和simpson的多样性指数。这与乙酸和丙酸的即时生产相吻合,它们可以有效地破坏细菌的dna。总体上,青贮叶的丰富度指数和多样性指数最高,但可以通过接种外源lab减少了这种情况。属水平上的细菌相对丰富度图显示,massilia是主要的微生物,其次是冻害后新鲜草料中的乳酸杆菌。青贮后细菌群落组成发生了很大变化。乳杆菌属(60.97-90.50%),pedioccocus(1.57-16.99%)和梭菌(2.10-5.46%)在青贮中占优势,相对丰富度分别为60.97-90.50%,1.57-16.99%和2.10-5.46%。在我们之前关于中国南方一年生黑麦草,三叶草及其混合物青贮饲料的研究中也曾报道了高丰富度的乳杆菌(lietal。,2019)。在外源lab接种的青贮饲料和对照青贮饲料之间没有观察到乳杆菌的相对丰富度的差异。来自(ogunade等,2018)的研究表明,接种植物乳杆菌可以减少小球菌和weissella的丰度。(romero等,2017)报道,接种l.buchneri增强了燕麦青贮饲料中weissella的存在。梭状芽孢杆菌的主要分布表明青贮饲料发酵效果差,丁酸水平高。接种外源lab(特别是lb)可以降低梭菌的相对丰富度。这可能是因为梭状芽孢杆菌对高渗透压和低ph值的耐受性较差,而乳酸的生产相对较快。此外,在青贮早期,梭菌孢子可以被l.buchneri产生的乙酸和丙酸损坏.与其他青贮饲料相比,叶青贮饲料表现出较高(p<0.05)的pedioccous,clostridium,caproiciproducens和bacillus的相对丰富度,但乳杆菌的相对丰度较低(p<0.05)。通过nmds图能清楚地鉴定青贮饲料中不同细菌群落的组成。新鲜饲草中的样品与青贮饲料中的样品显著分离。这一观察证实发酵对青贮饲料细菌群落结构有显著影响。接种lb后,对照青贮饲料中全株、茎叶的细菌群落组成发生了变化,这种变化非常明显,说明接种l.buchneri对细菌群落组成的变化起着重要作用。
在青贮饲料细菌群落组成与发酵变量的关系方面,分析表明,青贮细菌α多样性指数与氨氮浓度呈正相关(观察到的物种,r=0.686,p=0.041;shannon,r=0.711,p=0.032;chao1,r=0.669,p=0.049)和乙醇(观察到的物种,r=0.767,p=0.016;shannon,r=0.750,p=0.002;chao1,r=0.750,p=0.002)。典型相关分析(cca)图也用于探索属种水平的细菌群落组成与青贮发酵变量之间的相关性。细菌群落组成的61.90%和24.23%的变异可分别用x和y轴解释。氨氮,乙醇和丁酸之间存在正相关,与乙酸和丙酸呈负相关。大多数属与青贮发酵变量显著相关。例如,leuconostoc,caproiciproducens和clostridium属与丁酸,乙醇和氨-n的浓度正相关。我们推测lab延迟乳酸发酵,刺激了丁酸和乙醇的连续生产以及青贮饲料中粗蛋白的降解。这些证实了异型发酵细菌如布氏乳杆菌对于重建霜冻损伤的rcg青贮饲料的细菌群落至关重要。较低的青贮温度会对发酵过程产生负面影响。一些青贮饲料添加剂在较低温度下可能在功能上受损,因为青贮饲料接种剂中的菌株通常是在与较暖气候相关的温度下选择的(weinberg和muck,1996)。本发明中使用的外源lab也来自热带或温带地区。温度为15℃可能是布氏乳杆菌生长的下限,但其他异源性属如明串珠菌和威斯塞拉在青贮期间主导了lab群体(zhouetal。,2016)。此外,在<15℃的低青贮温度下延长贮存期可导致青贮饲料中布氏乳杆菌的无法解释的消失(zhou等,2016)。在本研究中,青贮初期的白天平均温度在15℃以上,这可以加快接种布氏乳杆菌的青贮饲料中乙酸和丙酸的生产。但是,在接下来的青贮期,温度<15℃时,布氏乳杆菌作用的过程可能会停止。青藏高原的研究表明,10℃贮藏的青贮饲料成功地启动了乳酸发酵(lietal.,2019;wangetal.,2017),由于一些本土lab是嗜冷菌,在低温条件下具有较强的发酵潜力(zhangetal.,2017)。因此,外源lab接种并没有显著增加乳酸菌的相对丰富度。我们推测冻害rcg植物上的本地lab(尤其是乳酸菌)主导了青贮饲料的发酵模式,贮藏期优选180天。今后青贮饲料在低温发酵过程中也应监测类似情况,通过对元基因组、元转录组和元蛋白质组的测序,更好地了解某些物种在微生物群落中的功能作用。
与现有技术相比,本发明的有益效果是:
发明人通过提供一种冻害芦苇草青贮方法,通过接种外源lab,减少(p<0.05)细菌α多样性和细菌群落组成移位(p<0.05),但没有改变(p>0.05)乳杆菌作为青贮饲料中优势属的相对丰度,特别是接种lb(植物乳杆菌l.buchneri)增加乙酸和丙酸含量,降低丁酸和氨氮含量,提高青贮期间的发酵速度、有氧稳定性、产品适口性及存放周期,整体优化冻害芦苇草青贮发酵过程,适宜在西部高原地区大规模生产,其成本低廉,大大提高青藏高原天然芦苇草利用率。同时,本发明还提供了一种该方法制备的青贮饲料,其品质稳定,营养保存较好,适口性好,存放周期长。
附图说明
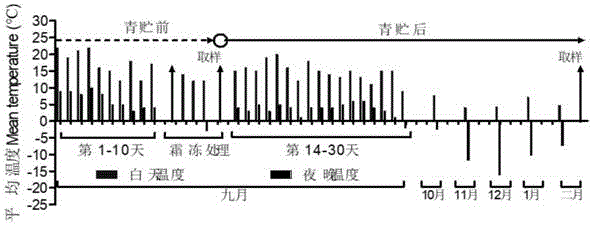
图1是本发明实施例1-6中青贮前后环境温度示意图;
图2是本发明检测例3中在属水平上的细菌相对丰富度图;
图3是本发明检测例3中鲜样、空白和加入外源lab青贮的rcg乳酸菌属图;
图4是本发明检测例3中鲜样、空白和加入外源lab青贮的rcg片球菌属图;
图5是本发明检测例3中鲜样、空白和加入外源lab青贮的rcg魏斯特菌属图;
图6是本发明检测例3中鲜样、空白和加入外源lab青贮的rcg丁酸菌图;
图7是本发明检测例3中鲜样、空白和加入外源lab青贮的rcg的群落组成分析图;
图8是本发明检测例3中鲜样、空白和加入外源lab青贮的ph和发酵产物影响下的细菌群落组成图;
图9是本发明检测例4中在属水平上的细菌相对丰富度图;
图10是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg乳酸菌属图;
图11是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg片球菌属图;
图12是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg魏斯特菌属图;
图13是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg丁酸菌图;
图14是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg的群落组成分析图;
图15是本发明检测例4中鲜样、空白和加入外源lab青贮的rcg的ph和发酵产物影响下的细菌群落组成图;
图16是本发明检测例5中在属水平上的细菌相对丰富度图;
图17是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg乳酸菌属图;
图18是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg片球菌属图;
图19是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg魏斯特菌属图;
图20是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg丁酸菌图;
图21是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg的群落组成分析图;
图22是本发明检测例5中鲜样、空白和加入外源lab青贮的rcg的ph和发酵产物影响下的细菌群落组成图;
图23是本发明检测例6中在属水平上的细菌相对丰富度图;
图24是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg乳酸菌属图;
图25是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg片球菌属图;
图26是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg魏斯特菌属图;
图27是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg丁酸菌图;
图28是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg的群落组成分析图;
图29是本发明检测例6中鲜样、空白和加入外源lab青贮的rcg的ph和发酵产物影响下的细菌群落组成图。
具体实施方式
下面结合具体实施方式对本发明的上述发明内容作进一步的详细描述。
但不应将此理解为本发明上述主题的范围仅限于下述实施例。在不脱离本发明上述技术思想情况下,根据本领域普通技术知识和惯用手段,做出各种替换和变更,均应包括在本发明的范围内。
实施例1
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌chikuso-1,比率为120cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的环境下青贮160天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料a-1。
实施例2
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌l.buchneri,比率为120cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的环境下青贮160天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料a-2。
实施例3
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌chikuso-1,比率为90cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的环境下青贮200天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料b-1。
实施例4
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌l.buchneri,比率为90cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的环境下青贮200天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料b-2。
实施例5
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌chikuso-1和l.buchneri各一半,比率为100cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的9月至次年3月青贮180天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料c。
实施例6
本实施例为冻害芦苇草青贮方法实施例,原料品种为虉草。
一种冻害芦苇草青贮方法,包括以下步骤:
(1)9月份连续三个夜晚环境温度低于0℃时,在第四天早上立即手工采摘无种子芦苇草,得到霜冻损坏的原料草料;
(2)将原料草料分成叶片和叶鞘l、茎s及整个作物w,将l、s和w分别切成0.5-1.0cm的段,混合均匀;
(3)将步骤(2)得到的新鲜材料接种外源lab植物乳杆菌chikuso-1和l.buchneri各一半,比率为106cfu/g,彻底混合,得到青贮材料;
(4)将步骤(3)得到的青贮材料按约1.0kg/份分份,放入聚酯袋中,抽真空,然后在环境温度低于20℃的9月至次年3月青贮180天。
环境温度条件如图1所示。
得到以上冻害芦苇草青贮方法制备的青贮饲料d。
检测例1
对青贮之前的新鲜牧草(rcg)和青贮之后的青贮饲料进行取样,将每个叶鞘l、茎s及整个作物w样品再分别分成四个相等的子样品,分别用于分析化学组成、ph和氨-n(g/kg总氮)和发酵产物、微生物种群及细菌多样性,最后进行统计分析:
(1)化学分析
将样品在105℃下干燥至恒重,然后通过0.60mm筛研磨以进行化学组成分析。通过凯氏定氮法(aoac,1990)测定粗蛋白,并通过kjeldaln×6.25的公式计算。中性洗涤纤维(ndf)通过(vansoest等,1991)的方法使用ankom2000纤维分析仪(ankomtechnology,fairport,ny)测定。wsc由mcdonald&henderson(1964)的方法决定。使用炸弹量热计(parr630,parrinstrumentco.,moline,illinois,usa)测量醚提取物(ee)。通过(jones和kinghorn,2006)的方法从乙醇提取物中提取总生物碱。
(2)ph、氨-n(g/kg总氮)和发酵产品的分析
在stomacher搅拌器中将20g样品与180ml超纯水混合3分钟。通过ph计测定滤液的ph。将约10ml的滤液离心(4500×g,15分钟,4℃),并使用高效液相色谱法和气相色谱法分析上清液中的乳酸,乙酸,丙酸,丁酸和乙醇。由(li等人,2017)描述。氨氮(nh3-n)通过(broderick和kang,1980)的方法测定。
(3)微生物种群分析
使用(cai8etal。,1999)的方法测定样品上的微生物群体。将10克每种样品放入无菌玻璃瓶中,悬浮在90ml无菌水中,并在实验室混合器(lb20es,shanghaiprimescienceco.,ltd,shanghai,china)中均化2小时。进行10-3至10-5的系列稀释。外源菌种(如有)的数量计数mrs琼脂(gcm188,landbridgetechnologyco.,ltd,beijing,china)在厌氧条件下于37℃温育48小时(厌氧箱;teherhardanaerobox,anx-1;hirosawaltd,tokyo,日本)。在营养琼脂(cm107,landbridgetechnologyco.,ltd,beijing,china)上计数需氧细菌,在30℃下孵育24小时。用1.5mg/l四环素(cm173,landbridgetechnologyco.,ltd,beijing,china)在麦芽提取物琼脂上计数霉菌和酵母,在28℃下孵育48小时。通过菌落外观和细胞形态观察将酵母与霉菌区分开来。
(4)描述样品细菌生态学
使用下一代测序方法描述新鲜样品及制备的青贮饲料细菌生态学。
(5)统计分析
对于3(馏分)×3(添加剂)因子排列,对样品数据进行双向anova。通过tukey的学生化范围测试确定平均值之间的显著差异,其中p<0.05被指定为显著。使用spss19.0程序(spssinc.,chicago,il,usa)进行这些分析。
检测例2
本检测例为检测例1中步骤“(4)描述样品细菌生态学”的具体方法,为下一代测序方法,通过该方法测定从样品中提取的细菌dna。
使用
检测例3
本检测例为按照检测例1和2中所述方法检测鲜样及实施例1和2中青贮饲料a的检测例。
(1)化学分析
未处理过(鲜样)的rcg的化学和微生物组成:表1-1
青贮过的rcg的化学和微生物组成:表1-2
(2)ph、氨-n(g/kg总氮)和发酵产品的分析:表1-3
(3)微生物种群分析:表1-4
(4)下一代测序方法描述样品细菌生态学
(5)统计分析
结果见图2-图8。
检测例4
本检测例为按照检测例1和2中所述方法检测鲜样及实施例3和4中青贮饲料b的检测例。
(1)化学分析
未处理过(鲜样)的rcg的化学和微生物组成:表2-1
青贮过的rcg的化学和微生物组成:表2-2
(2)ph、氨-n(g/kg总氮)和发酵产品的分析:表2-3
(3)微生物种群分析:表2-4
(4)下一代测序方法描述样品细菌生态学
(5)统计分析
结果见图9-图15。
检测例5
本检测例为按照检测例1和2中所述方法检测鲜样及实施例5中青贮饲料c的检测例。
(1)化学分析
未处理过(鲜样)的rcg的化学和微生物组成:表3-1
青贮过的rcg的化学和微生物组成:表3-2
(2)ph、氨-n(g/kg总氮)和发酵产品的分析:表3-3
(3)微生物种群分析:表3-4
(4)下一代测序方法描述样品细菌生态学
(5)统计分析
结果见图16-图22。
检测例6
本检测例为按照检测例1和2中所述方法检测鲜样及实施例6中青贮饲料d的检测例。
(1)化学分析
rcg在冻害前后的化学和微生物组成:表4-1
青贮过的rcg的化学和微生物组成:表4-2
(2)ph、氨-n(g/kg总氮)和发酵产品的分析:表4-3
(3)微生物种群分析:表4-4
(4)下一代测序方法描述样品细菌生态学
(5)统计分析
结果见图23-图29。
综上,本发明提供了一种冻害芦苇草青贮方法,通过接种外源lab,减少(p<0.05)细菌α多样性和细菌群落组成移位(p<0.05),但没有改变(p>0.05)乳杆菌作为青贮饲料中优势属的相对丰度,特别是接种lb(植物乳杆菌l.buchneri)增加乙酸和丙酸含量,降低丁酸和氨氮含量,提高青贮期间的发酵速度、有氧稳定性、产品适口性及存放周期,整体优化冻害芦苇草青贮发酵过程,适宜在西部高原地区大规模生产,其成本低廉,大大提高青藏高原天然芦苇草利用率。同时,本发明还提供了一种该方法制备的青贮饲料,其品质稳定,营养保存较好,适口性好,存放周期长。
- 还没有人留言评论。精彩留言会获得点赞!