一种非人哺乳动物焦虑症或其相关疾病动物模型的建立方法及其用途与流程
本发明涉及生物技术领域,具体地,涉及一种非人哺乳动物焦虑症或其相关疾病动物模型的建立方法及其用途。
背景技术:
焦虑症,又称为焦虑性神经症,是神经症这一大类疾病中最常见的一种,以焦虑情绪为主要特征,其核心症状为担忧。主要包括广泛性焦虑、急性焦虑发作、恐怖症、创伤后应激障碍、急性应激障碍、强迫障碍。主要表现为无明确客观对象的紧张担心、坐立不安、植物神经症状(心悸、手抖、出汗、尿频等)。
世界卫生组织(WHO)统计表明,从1990年至2003年,遭受抑郁症或焦虑症痛苦的人数从4.16亿增至6.15亿,几乎达到了全球人口的10%。并且,精神障碍疾病占到了非致命疾病的30%。据估算,在发生紧急情况的人员中,每5名中就有1名患抑郁症或焦虑症。全球每年需要负担约1万亿美元来治疗抑郁症和焦虑症。
由于焦虑症的发病机制非常复杂,至今焦虑症的致病机理尚未研究清楚,缺乏安全有效的治疗手段,因此深入探讨其发病机理是治疗焦虑症的重要基础。小鼠疾病模型在研究人类疾病的致病机理和药物筛选中起到了非常重要的作用。在探究人类的认知功能、神经退行性疾病、神经精神疾病等方面,小鼠模型具有巨大的优势。
基因敲除是上世纪80年代发展起来的一项复杂的分子生物学技术,其基于小鼠胚胎干细胞DNA同源重组原理,也称为“基因打靶”技术。其后利用这项技术构建了数千种基因突变小鼠,这些基因工程小鼠不仅为生物科学和医学的研究带来了突破,也在新药研发中发挥了至关重要的作用。
目前已有的小鼠焦虑症模型的建立方法主要包括两类:一类是基于非条件化的行为模型,根据其特点又可分为探究行为模型和社会行为模型。然而,这类模型基本仅限于躯体应激反应,缺乏有效的病因学监测手段。另一类焦虑模型是基于条件反射的模型,主要包括饮水冲突模型、条件性电击模型等。然而,这类模型包含有电击这类物理应激的成分,使得心理应激中躯体应激的比例增大是其主要缺陷。
因此,本领域迫切需要开发一种可以较好地模拟临床上原发性的焦虑症,个体差异较小,其遗传背景高度一致,可以作为研究焦虑症的发病机理以及新药筛选的有力工具的动物模型,并将构建的动物模型用于研究焦虑症的发病机理以及筛选或鉴定治疗焦虑症的物质。
技术实现要素:
本发明的目的在于提供一种可以较好地模拟临床上原发性的焦虑症,个体差异较小,其遗传背景高度一致,可以作为研究焦虑症的发病机理以及新药筛选的有力工具的动物模型,并将构建的动物模型用于研究焦虑症的发病机理以及筛选或鉴定治疗焦虑症的物质。
本发明的第一方面提供了一种非人哺乳动物的焦虑症或其相关疾病动物模型的制备方法,包括以下步骤:
(a)提供非人哺乳动物的细胞,将所述细胞中的Glce基因失活,得到Glce基因失活的非人哺乳动物细胞;
(b)利用步骤(a)中得到的Glce基因失活的细胞,制备得到Glce基因失活的焦虑症或其相关疾病的动物模型。
在另一优选例中,在步骤(a)中,还包括如下步骤:
(a1)利用DNA同源重组技术,将所述Glce基因中的外显子3至外显子5中的一个或多个外显子剔除或中断,并用筛选标记替换,得到Glce基因失活的非人哺乳动物细胞。
在另一优选例中,在步骤(b)中,还包括如下步骤:
(b1)利用步骤(a)中得到的Glce基因失活的非人哺乳动物细胞制备得到嵌合非人哺乳动物;
(b2)将步骤(b1)中得到的嵌合非人哺乳动物和正常野生型非人哺乳动物交配繁育,在后代中筛选获得Glce基因失活的杂合子非人哺乳动物;
(b3)通过将步骤(b2)中得到的杂合子非人哺乳动物相互交配获得Glce基因失活的纯合子非人哺乳动物,从而得到Glce基因失活的非人哺乳动物的动物模型。
在另一优选例中,在步骤(b3)中,还包括步骤(b4):将Glce基因失活的纯合子非人哺乳动物与同一物种的神经元特异性敲除工具非人哺乳动物进行杂交,从而获得神经元特异性的Glce基因失活的非人哺乳动物的动物模型。
在另一优选例中,所述将Glce基因失活包括基因剔除、基因中断或基因插入。
在另一优选例中,所述基因失活包括Glce基因不表达,或表达没有活性的Glce蛋白。
在另一优选例中,所述Glce基因失活是通过缺失或敲除Glce的外显子3而失活。
在另一优选例中,所述的Glce基因失活是神经元特异性的Glce基因失活。
在另一优选例中,所述非人哺乳动物为啮齿动物或灵长目动物,较佳地包括小鼠、大鼠、兔和/或猴。
在另一优选例中,所述非人哺乳动物是小鼠,并且在步骤(b4)中把Glce Loxp/Loxp小鼠与工具鼠NSE(neuron-specific enolase)-Cre交配,即得到在神经元细胞特异性Glce基因敲除小鼠简称cKO小鼠(即神经元特异性Glce失活小鼠)。
在另一优选例中,所述筛选标记选自下组:抗性基因、荧光蛋白基因、或其组合。
在另一优选例中,所述筛选标记包括neo基因。
在另一优选例中,所述步骤(b)中得到的Glce基因失活的非人哺乳动物的动物模型中,与野生型对照动物相比,具有选自下组的一个或多个特征:
(t1)焦虑样的行为增加;
(t2)焦虑程度增加;
(t3)旷场活动水平减少;
(t4)探索新异环境的欲望降低;
(t5)抑郁样行为增加;
(t6)抑郁程度增加;
(t7)恐惧样行为增加;
(t8)恐惧程度增加;
(t9)认知障碍增加;
(t10)中风的发生率增加;
(t11)形态的肥胖程度增加;
(t12)肥胖症状的发生率增加;
(t13)脂肪组织湿重增加。
在另一优选例中,所述焦虑样行为选自下组:在旷场实验中探索中央区域的时间减少,表现出探索新异环境的欲望降低、在高架十字迷宫实验中探索开放臂的时间减少,停留在封闭臂的时间增加,表现出焦虑样行为、或其组合。
在另一优选例中,所述旷场活动水平选自下组旷场活动的路程、活动时间、活动速度、或其组合。
在另一优选例中,所述抑郁样行为选自下组:在旷场实验中探索中央区域的时间减少,表现出探索新异环境的欲望降低、在强迫游泳实验中的不动时间增加,表现出行为绝望、或其组合。
在另一优选例中,所述脂肪组织湿重选自下组:内脏脂肪组织湿重、皮下脂肪组织湿重、或其组合。
在另一优选例中,所述内脏脂肪组织湿重选自下组:双侧性腺周围脂肪组织湿重、双侧肾周脂肪组织湿重、或其组合。
在另一优选例中,所述皮下脂肪组织湿重包括双侧腹股沟脂肪组织湿重。
本发明第二方面提供了一种本发明第一方面所述方法制备的非人哺乳动物模型的用途,将该模型用作研究焦虑症或其相关疾病的动物模型。
在另一优选例中,所述焦虑症或其相关疾病选自下组:广泛性焦虑、急性焦虑发作、恐怖症、创伤后应激障碍、急性应激障碍、强迫障碍、或其组合。
在另一优选例中,所述慢性焦虑症包括广泛性焦虑。
在另一优选例中,所述急性焦虑症包括惊恐障碍。
本发明第三方面提供了一种本发明第一方面所述方法制备的非人哺乳动物模型的用途,用于筛选或鉴定可减轻或治疗焦虑症或其相关疾病的物质(治疗剂)。
在另一优选例中,所述焦虑症或其相关疾病选自下组:广泛性焦虑、急性焦虑发作、恐怖症、创伤后应激障碍、急性应激障碍、强迫障碍、或其组合。
在另一优选例中,所述慢性焦虑症包括广泛性焦虑。
在另一优选例中,所述急性焦虑症包括惊恐障碍。
本发明第四方面提供了一种筛选或鉴定治疗或缓解焦虑症或其相关疾病的潜在治疗剂的方法,包括以下步骤:
(a)在测试组中,在测试化合物的存在下,将测试化合物施用于本发明第一方面所述方法制备的非人哺乳动物模型,对所述动物模型的行为进行行为学评价;并且在不施用所述测试化合物且其他条件相同的对照组中,对所述动物模型的行为进行行为学评价;
(b)对测试组和对照组动物模型的行为进行比较,其中,与对照组相比,如果施用了测试化合物的动物模型中表征焦虑行为得到改善,则表明该测试化合物可作为焦虑症的潜在治疗剂。
在另一优选例中,所述的行为学分析包括:自主活动、旷场实验、和/或高架十字迷宫实验。
在另一优选例中,所述的方法是非诊断性和非治疗性的。
在另一优选例中,所述改善是在统计学上具有显著性意义的改善。
在另一优选例中,所述方法包括步骤(c),将步骤(b)筛选或鉴定的潜在治疗剂施用于本发明第一方面所述方法制备的非人哺乳动物模型,从而测定其对所述动物模型焦虑症或其相关疾病的严重程度的影响。
本发明第五方面提供了一种非人哺乳动物模型,用本发明第一方面所述方法制备。
在另一优选例中,对于Glce基因失活而言,所述的非人哺乳动物模型是杂合的或纯合的。
在另一优选例中,所述的Glce基因失活是神经元特异性的Glce基因失活。
本发明第六方面提供了一种细胞的用途,所述细胞中的葡萄糖醛酸C5异构酶(Glucuronyl C5-epimerase,Glce)基因失活或下调,用于制备构建非人哺乳动物的焦虑症或其相关疾病动物模型的生物制剂。
在另一优选例中,所述生物制剂为液态制剂。
本发明第七方面提供了一种Glce基因或其蛋白的失活剂的用途,用于制备构建非人哺乳动物的焦虑症或其相关疾病动物模型的制剂。
在另一优选例中,所述失活剂包括抑制剂。
在另一优选例中,所述失活剂选自下组:抗体、小分子化合物、核酸、或其组合。
应理解,在本发明范围内中,本发明的上述各技术特征和在下文(如实施例)中具体描述的各技术特征之间都可以互相组合,从而构成新的或优选的技术方案。限于篇幅,在此不再一一累述。
附图说明
图1显示了Glce基因突变序列设计策略。
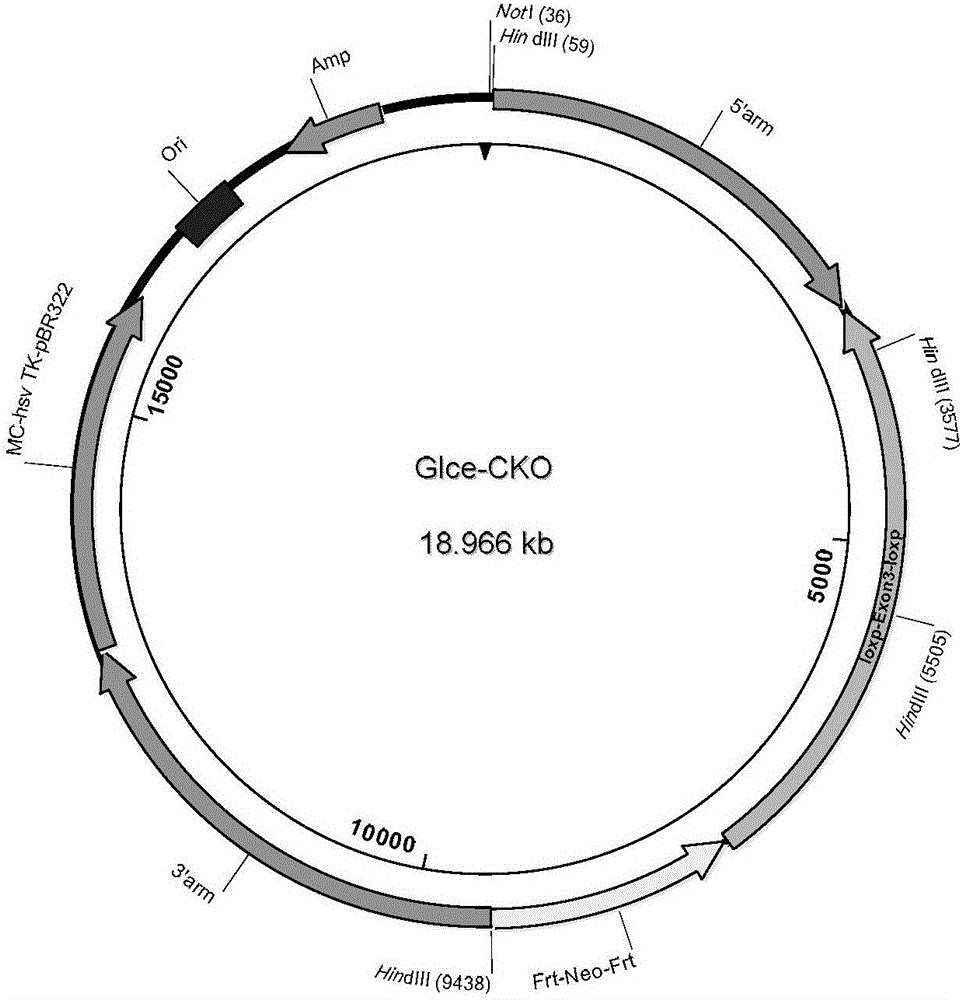
图2显示了Glce基因剔除打靶载体质粒图谱。
图3显示了Glce基因剔除打靶载体质粒DNA的酶切鉴定结果。
图4显示了ES克隆基因组DNA的5’端PCR产物的电泳结果。
图5显示了ES克隆基因组DNA的3’端PCR产物的电泳结果。
图6显示了Glce转基因小鼠(Loxp/+)序列突变位点PCR结果。
图7显示了Glce突变纯合子(Loxp/Loxp Cre)、突变杂合子(Loxp/+Cre)和Wild type(+/+Cre)的序列突变位点PCR结果。
图8显示了Glce突变纯合子小鼠的自发活动路程、时间及速度。
图9显示了Glce突变纯合子小鼠在旷场中央区域的活动路程、时间及速度。
图10显示了Glce突变纯合子小鼠在高架十字迷宫实验中分别探究开放臂、中央区域和封闭臂的时间。
图11显示了Glce突变纯合子小鼠的自发活动路程、时间及速度。
图12显示了Glce突变纯合子小鼠在旷场实验中的活动总路程、时间及速度。
图13显示了Glce突变纯合子小鼠在旷场中央区域的活动路程、时间及速度。
图14显示了Glce突变纯合子小鼠在旷场周边区域的活动路程、时间及速度。
图15显示了Glce突变纯合子小鼠在强迫游泳实验中的不动时间。
图16显示了Glce突变纯合子小鼠与C57小鼠的形态对比。其中,左图:上为C57雌性小鼠,下为Glce突变纯合子雌性小鼠;右图:上为C57雄性小鼠,下为Glce突变纯合子雄性小鼠。
图17显示了Glce突变纯合子小鼠与C57小鼠的体重对比情况。
图18显示了Glce突变纯合子小鼠与C57小鼠的体脂湿重对比情况。
图19显示了Glce突变纯合子小鼠与C57小鼠的双侧性腺周围脂肪组织湿重对比情况。
图20显示了Glce突变纯合子小鼠与C57小鼠的双侧肾周脂肪组织湿重对比情况。
图21显示了Glce突变纯合子小鼠与C57小鼠的内脏脂肪组织湿重对比情况。
图22显示了Glce突变纯合子小鼠与C57小鼠的双侧腹股沟脂肪组织湿重对比情况。
图23显示了中风行为学评价实验。
图24显示了TTC染色检测中风小鼠脑组织梗塞情况。
具体实施方式
本发明人经过广泛而深入的研究,建立了一种遗传稳定、表型稳定的焦虑症或其相关疾病模型,它是Glce基因被剔除或失活的小鼠或其他非人哺乳动物。本发明的动物模型是一种有效的焦虑症或其相关疾病动物模型,可用于研究焦虑症或其相关疾病(如广泛性焦虑),并可以用于特定药物的筛选和测试试验。在此基础上完成了本发明。
此外,本发明还意外地发现,剔除或失活Glce基因所得到的动物模型还可以同时用于研究中风、肥胖、抑郁等疾病。在此基础上完成了本发明。
Glce基因
硫酸乙酰肝素是在细胞表面和细胞质基质中广泛存在的一种多糖,作为一个带负电的线性大分子,许多细胞因子、生长因子、趋化因子和白介素能与之特异性地结合,进而在胚胎发育、细胞生长、炎症反应、凝血、肿瘤转移和病毒侵染等生理过程中发挥作用1。
葡萄糖醛酸C5异构酶(Glce)是硫酸乙酰肝素蛋白聚糖糖链合成过程中的一个关键酶,它能将糖链中的D-葡萄糖醛酸异构为L-艾杜糖醛酸2,大大提高了硫酸乙酰肝素的复杂度,并为糖链提供了更多的柔性,L-艾杜糖醛酸是硫酸乙酰肝素识别众多蛋白分子的一个必不可缺的位点3。
在对21例正常乳腺组织和74例乳腺肿瘤组织的研究发现,82%-84%的人乳腺肿瘤组织中Glce在mRNA水平和蛋白水平的表达是显著下调或者完全丧失的4。此外,在乳腺癌和肺癌细胞系中过表达Glce能抑制小细胞肺癌和乳腺癌细胞的增殖,这提示我们Glce可能是一个潜在的抑癌基因5,6。
Glce基因位于小鼠基因组9号染色体上,全长618(EnsemblGene ID:ENSMUSG00000032252,Genebank登录号:93683)。Glce基因组序列包括4个内含子和5个外显子,Glce基因表达蛋白有3个转录本,转录本1具有3个外显子、2个内含子,转录本2具有5个外显子、4个内含子,转录本3具有2个外显子、1个内含子。这些序列信息可参见文献或EnsemblGene、Genebank等公共数据库。
人类等其他物种的Glce基因也可参见文献或EnsemblGene、Genebank等公共数据库。
应理解,术语“Glce”还包括各种天然存在的Glce基因的变异形式。代表性的例子包括:因密码子的简并性而编码与野生型相同的Glce蛋白的核苷酸序列,编码野生型Glce蛋白的保守性变异多肽的核苷酸序列。此外,对于小鼠之外的其他哺乳动物时,该术语指Glce基因在该哺乳动物中的同系物。例如对于人而言,该术语指人的Glce(已知小鼠Glce基因与人类Glce基因的cDNA同源度为91.4%,氨基酸序列的同源度为97.4%)。
Glce基因或其蛋白的失活剂
在本发明中,所述Glce的失活剂包括全部失活或部分失活。
本发明的Glce蛋白的失活剂包括(a)抑制剂,所述抑制剂的例子包括(但并不限于):小分子化合物、抗体、反义核酸、miRNA、siRNA、或其组合;(b)Glce基因的敲除剂。
焦虑症或其相关疾病
焦虑症,又称为焦虑性神经症,是神经症这一大类疾病中最常见的一种,以焦虑情绪为主要特征,其核心症状为担忧。包括广泛性焦虑、急性焦虑发作、恐怖症、创伤后应激障碍、急性应激障碍、强迫障碍等。主要表现为无明确客观对象的紧张担心、坐立不安、植物神经症状(心悸、手抖、出汗、尿频等)。
广泛性焦虑的特征主要有:(1)情绪症状。主要表现为在没有明显诱因的情况下,患者经常出现与现实情境不符的过分担心、紧张害怕,患者感觉自己一直处于一种紧张不安、提心吊胆,恐惧、害怕、忧虑的内心体验中。(2)植物神经症状。主要表现为头晕、胸闷、心慌、呼吸急促、口干、尿频、尿急、出汗、震颤等躯体方面的症状。(3)运动性不安。主要表现为坐立不安,坐卧不宁,烦躁,很难静下心来。
惊恐障碍的特征主要有:(1)濒死感或失控感。在正常的日常生活中患者几乎跟正常人一样,而一旦发作时(有的有特定触发情境),患者突然出现极度恐惧的心理,体验到濒死感或失控感。
(2)植物神经系统症状。同时出现如胸闷、心慌、呼吸困难、出汗、全身发抖等。
(3)一般持续几分钟到数小时。发作开始突然,发作时意识清楚。
(4)极易误诊。发作时患者往往拨打“120”急救电话,去看心内科的急诊。尽管患者看上去症状很重,但是相关检查结果大多正常,因此往往诊断不明确。
恐怖症(包括社交恐怖、场所恐怖、特定的恐怖)的特征主要有:恐怖症的核心表现和急性焦虑发作一样都是惊恐发作。不同点在于恐怖症的焦虑发作是由某些特定的场所或者情境引起,患者不处于这些特定场所或情境时不会引起焦虑。
在本发明中,焦虑症或其相关疾病包括(但不限于):广泛性焦虑、急性焦虑发作、恐怖症、创伤后应激障碍、急性应激障碍、和/或强迫障碍。
基因失活
对于功能未知基因的研究可采用许多方法,例如使有待研究的基因失活,分析所得的遗传修饰的表型变化,进而获得该基因的功能信息。这一研究方法的另一优点是可以将基因功能和疾病进行关联,从而在获得基因功能的同时也能获得该基因作为潜在药物或者药物靶点所能治疗的疾病信息和疾病动物模型。基因失活的方法可通过基因剔除、基因中断或基因插入的方式来完成。其中,基因剔除技术是研究人类基因在整体中的功能的非常强有力的手段。
动物模型
在本发明中,提供了一种非常有效的焦虑症或其相关疾病的非人哺乳动物模型。
在本发明中,非人哺乳动物的例子包括(但并不限于):小鼠、大鼠、兔、猴等,更佳地是大鼠和小鼠。
如本文所用,术语“Glce基因失活”包括一个或两个Glce基因被失活的情况,即包括Glce基因杂合地和纯合地失活。例如,Glce基因失活的小鼠可以是杂合或纯合的小鼠。
在本发明中,可基因剔除或转入外源基因(或片段)而使Glce基因失活等方法制备Glce基因失活的非人哺乳动物(如小鼠)。在本领域中,通过基因剔除或转入外源基因而使靶基因失活的技术是已知的,这些常规技术都可用于本发明。
在本发明的另一优选例中,Glce基因的失活是通过基因剔除实现的。
在本发明的另一优选例中,Glce基因的失活是通过Glce基因中插入外源基因(或片段)而实现的。
在本发明的一具体实例中,可构建一含有外源插入片段的构建物,该构建物含有与靶基因(Glce)的插入位点的两侧的侧翼序列同源的同源臂,从而可以通过同源重组高频地将外源插入片段(或基因)插入至Glce基因组序列(尤其是外显子区域),造成小鼠Glce基因的移码、提前终止、或敲除,从而导致Glce缺失或失活。
用本发明方法获得的纯合或杂合的小鼠可育,发育正常。失活的Glce基因可以孟德尔规律遗传给后代小鼠。
在一优选例中,本发明提供了一种缺失Glce基因的纯合小鼠模型动物。
候选药物或治疗剂
在本发明中,还提供了一种利用本发明的动物模型,筛选治疗焦虑症或其相关疾病的候选药物或治疗剂的方法。
在本发明中,候选药物或治疗剂是指已知具有某种药理学活性或正在被检测的可能具有某种药理学活性的物质,包括但不限于核酸、蛋白、糖类、化学合成的小分子或大分子化合物、细胞等。候选药物或治疗剂的给药方式可以是口服、静脉注射、腹腔注射、皮下注射、椎管给药或直接脑内注射。
本发明的主要优点包括:
(1)本发明可以较好地模拟临床上原发性的焦虑症;
(2)个体差异较小,其遗传背景高度一致;
(3)可以作为研究焦虑症的发病机理以及新药筛选的有力工具。
(5)本发明的焦虑症模型遗传稳定、表型稳定。
(6)用本发明方法获得的纯合或杂合的动物模型可育。转基因杂合小鼠具有生殖能力,失活的Glce基因可以孟德尔规律遗传给后代小鼠。
(7)本发明的焦虑症或其相关疾病动物模型表现出焦虑症状,因此可以广泛用于焦虑症相关疾病的药物筛选和测试。
(8)本发明首次揭示了剔除或失活Glce基因所得到的动物模型还可以同时用于研究中风、肥胖、抑郁等疾病。
下面结合具体实施例,进一步阐述本发明。应理解,这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件,例如Sambrook等人,分子克隆:实验室手册(New York:Cold Spring Harbor Laboratory Press,1989)中所述的条件,或按照制造厂商所建议的条件。除非另外说明,否则百分比和份数是重量百分比和重量份数。
如无特别说明,实施例所用的材料均为市售产品。
实施例1建立Glce基因在神经元特异条件敲除的小鼠动物模型。
首先构建Glce突变基因序列(图1)。如图1所示,设计Glce基因剔除打靶载体序列,将Loxp/Loxp等位基因插入到Glce基因3号外显子两侧,在3’端插入neo基因,5’端臂3125bp,3’端臂3718bp。
图2为Glce基因剔除打靶载体质粒图谱。1.获得目的基因(Glce)的同源片段,将此DNA片段克隆到质粒载体中;2.从重组质粒中切除目的基因的大部分同源DNA序列,只留部分序列在线性质粒载体的两端;3.将neo基因克隆到带有目的基因同源顺序的线性质粒中,使之位于残留目的基因同源顺序的中间;4.在目的基因同源顺序的外侧线性化重组质粒载体,将hsv-tk基因克隆到此线性载体中。基因位点如表1所示。
表1
注:基因位置标注号根据“10kb Up and Down of Glce gene”。
图3为Glce基因剔除打靶载体质粒DNA的酶切鉴定,使用1Kb DNA ladder。对打靶载体进行线性化:100μg Glce-CKO质粒DNA用NotI(酶用量:150U)线性化,酶切体系为150μl,37℃消化过夜,等体积酚氯仿、氯仿处理后,无水乙醇沉淀,100μl无菌PBS重悬备用。ES细胞打靶:ES细胞SCR012来源于129SV/EV品系雄性小鼠(购自中科院上海实验动物中心)的胚胎干细胞,线性化DNA量:35μg,电转仪型号:Bio-Rad Gene Pulser(Cat.No.165-2105),电穿孔条件:电压240v,电容500μF,实际通电时间10.5ms,实际电压256v,克隆筛选条件:300μg/ml G418和2μM GanC筛选8天。挑取抗性克隆和提供DNA样本共96份。
阳性ES细胞基因组鉴定方法:
1.5’arm PCR鉴定
P1引物位于5’arm外,P2引物位于neo重组区域,距离臂外引物8.2kb。
P1和P2引物序列:
P1:GGCATTTGCACTCACATACACAACCCA(gene site:15824-15850)(SEQ ID NO.:1)
P2:GTGCCACTCCCACTGTCCTTTCC(SEQ ID NO.:2)PCR反应体系(ul):
PCR反应条件:
PCR仪:Eppendorf AG 22331Hamburg
试剂:TaKaRa La Taq宝生物工程(大连)有限公司(Cat:DRR002B)
分子量Marker:MBI GeneRuler 1kb DNA Ladder(晶美生物,Cat:SM0311)
2.3’arm PCR鉴定
P4引物位于3’arm外,P3引物位于neo重组区域,距离臂外引物4.7kb。
P3和P3引物序列:
P4:GAGAGGCTTGGAGGCGGTGCTGATCTT(gene site:29603-29629)(SEQ ID NO.:3)
P3:GATATACTATGCCGATGATTAATTGTC(SEQ ID NO.:4)PCR反应体系(ul):
PCR反应条件:
PCR仪:Eppendorf AG 22331Hamburg
试剂:TaKaRa La Taq宝生物工程(大连)有限公司(Cat:DRR002B)
分子量Marker:MBI GeneRuler 1kb DNA Ladder(晶美生物,Cat:SM0311)
ES细胞克隆鉴定结果PCR鉴定药物抗性ES细胞克隆96个,其中19个ES克隆发生双臂同源重组。PCR产物经DNA序列测定进一步证实。图4显示了ES克隆基因组DNA的5’端PCR产物的电泳结果。图5显示了ES克隆基因组DNA的3’端PCR产物的电泳结果。
阳性ES克隆囊胚注射:
显微注射用囊胚来源:C57BL/6J小鼠(购自上海斯莱克实验动物有限公司)超数排卵,自然受孕体内发育至囊胚阶段。注射60枚胚胎,注射后移植入3只假孕小鼠受体子宫,受体为C57BL/6J(♂)与CBA(♀)(购自上海斯莱克实验动物有限公司)的杂交一代。从出生的小鼠中挑选嵌合率大于50%的小鼠饲养至成年,与C57BL/6J雌性小鼠进行交配,后代灰色小鼠经提取尾基因组DNA进行PCR鉴定(鉴定策略同上),结果如图6所示,获得双臂阳性F1代小鼠(Loxp/+)。
将F1代小鼠饲养至成年,与NSE-Cre工具鼠(购自上海斯莱克实验动物有限公司)杂交,获得F2代Loxp/+Cre小鼠。将F2代小鼠饲养至成年,F2代内雌雄相互交配,按照孟德尔规律遗传,获得F3代突变纯合子(Loxp/Loxp Cre):突变杂合子(Loxp/+Cre):Wild type(+/+Cre)的比例约为1:2:1。经提取尾基因组DNA进行PCR鉴定(鉴定策略同上),结果如图7所示。将突变纯合子小鼠(Loxp/Loxp Cre)用于后续动物行为学实验。
实施例2Glce突变纯合子小鼠的行为学分析
将携带Cre重组酶的Glce基因突变纯合子小鼠(以下简称Glce突变纯合子小鼠)置于清洁饲养环境饲养。在小鼠青年期(>3月龄)通过一系列动物行为学实验包括自主活动、旷场实验、高架十字迷宫等实验评价小鼠动物模型的焦虑症样症状。
2.1自主活动
将小鼠放置于黑暗实验箱内(110mm X 110mm X 330mm),测试小鼠的自发活动5min。通过红外摄像拍摄小鼠的活动情况。使用上海吉量软件科技有限公司的视频跟踪软件和分析软件对小鼠活动轨迹以及活动时间进行分析。
研究发现,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠的自发活动的总路程、活动时间以及活动速度并没有显著差异(图8)。
结果表明,青年期Glce突变纯合子小鼠天生的习性与C57BL/6正常小鼠相比较并没有显著差异。
2.2旷场实验
将小鼠放置于明亮空旷实验箱内(500mm X 500mm X 590mm),测试小鼠的探索活动5min。通过红外摄像拍摄小鼠的活动情况。使用上海吉量软件科技有限公司的视频跟踪软件和分析软件对小鼠活动轨迹以及活动时间进行分析。
研究发现,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在旷场中央区域活动的路程、活动时间以及活动速度均显著减小(图9)。
结果表明,青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较探索新异环境的活动显著减少,并产生更强烈的焦虑情绪。
2.3高架十字迷宫实验
将小鼠放置于开放臂单臂长30cm、宽6cm、封闭臂单臂长30cm、宽6cm、高14.5cm的高架十字迷宫中,离地面高度约50cm,通过摄像拍摄小鼠5min内在高架十字迷宫中的活动情况。对小鼠的活动进行分析发现,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在高架十字迷宫实验中探究开放臂的时间显著减少,停留在封闭臂的时间显著增加,在中央区域停留的时间没有显著差异(图10)。
结果表明,青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较,焦虑程度明显增加。雄性青年期Glce突变纯合子小鼠与雌性青年期Glce突变纯合子小鼠相比较焦虑情绪更明显。除此之外,产生一定程度焦虑的青年期Glce突变纯合子小鼠同时也产生一定程度的抑郁。
实施例3Glce突变纯合子小鼠的抑郁行为学分析
3.1自主活动
将携带Cre重组酶的Glce基因突变纯合子小鼠放置于黑暗实验箱内(110mm X 110mm X 330mm),测试小鼠的自发活动5min。通过红外摄像拍摄小鼠的活动情况。使用上海吉量软件科技有限公司的视频跟踪软件和分析软件对小鼠活动轨迹以及活动时间进行分析。
研究显示,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠的自发活动的总路程、活动时间以及活动速度并没有显著差异(图11)。
结果表明,青年期Glce突变纯合子小鼠天生的习性与C57BL/6正常小鼠相比较并没有显著差异。
3.2旷场实验
将小鼠放置于明亮空旷实验箱内(500mm X 500mm X 590mm),测试小鼠的探索活动5min。通过红外摄像拍摄小鼠的活动情况。使用上海吉量软件科技有限公司的视频跟踪软件和分析软件对小鼠活动轨迹以及活动时间进行分析。
结果显示,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在旷场实验中活动的总路程、活动时间以及活动速度均显著减小(图12)。结果表明,青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较活动显著减少。
与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在旷场中央区域活动的路程、活动时间以及活动速度均显著减小(图13),结果表明,青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较探索新异环境的活动显著减少。
与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在旷场周边区域活动的路程、活动时间以及活动速度均显著减小(图14),结果表明青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较活动显著减少。
总的来说,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠的活动显著减少,并且探索新异环境的活动显著减少,表明青年期Glce突变纯合子小鼠有一定程度的抑郁。
3.3强迫游泳
将小鼠放置于直径12cm、高度25cm的水缸中(水温21-22℃),小鼠被强迫在水温较低的水中游泳,测试小鼠的活动6min,记录小鼠在后4min中的不动时间。通过摄像拍摄小鼠的活动情况。对小鼠活动时间进行分析。研究发现,与青年期C57BL/6正常小鼠相比较,青年期Glce突变纯合子小鼠在强迫游泳实验中的不动时间显著增加(图15)。
结果表明,青年期Glce突变纯合子小鼠与C57BL/6正常小鼠相比较有一定程度的抑郁。
实施例4Glce突变纯合子小鼠的形态学分析
Glce突变纯合子小鼠的出生比例符合孟德尔定律,通过检测小鼠的体重,结果如图16所示。结果表明,与成年C57BL/6正常小鼠相比较,成年Glce神经元特异敲除小鼠的形态更加肥胖。
实施例5Glce突变纯合子小鼠的体重分析
将Glce突变纯合子小鼠置于电子天平上称重,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的体重差异。
研究发现(图17),与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠肥胖症状的发生率更高,并且体重与成年C57BL/6正常小鼠相比较具有统计学显著差异。
结果显示,以超过成年C57BL/6正常小鼠体重平均值的20%以上的标准认为是肥胖,雌性成年C57BL/6正常小鼠的体重平均为28.7g,雌性成年Glce突变纯合子小鼠的体重平均为34.6g,雌性成年Glce突变纯合子小鼠肥胖症状的发生率约为50%,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的体重平均为32.2g,雄性成年Glce突变纯合子小鼠的体重平均为39.5g,雄性成年Glce突变纯合子小鼠肥胖症状的发生率约为66%,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的体重平均为30.7g,成年Glce突变纯合子小鼠(包含雌雄)的体重平均为37.8g,成年Glce突变纯合子小鼠肥胖症状的发生率总计约为60%,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠肥胖症状的发生率更高。
实施例6Glce突变纯合子小鼠的体脂分析
将Glce突变纯合子小鼠解剖后取脂肪组织包括双侧性腺周围脂肪组织、双侧肾周脂肪组织、双侧腹股沟脂肪组织置于电子天平上称重,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的脂肪组织湿重差异。
研究发现(图18),与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的体脂湿重增加,具有统计学显著差异。
结果显示,雌性成年C57BL/6正常小鼠的体脂湿重平均为0.94g,雌性成年Glce突变纯合子小鼠的体脂湿重平均为8.50g,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的体脂湿重平均为2.98g,雄性成年Glce突变纯合子小鼠的体脂湿重平均为7.52g,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的体脂湿重平均为2.21g,成年Glce突变纯合子小鼠(包含雌雄)的体脂湿重平均为8.01g,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的体脂湿重增加。
实施例7Glce突变纯合子小鼠的脂肪组织分析
7.1Glce突变纯合子小鼠的双侧性腺周围脂肪组织分析
将Glce突变纯合子小鼠解剖后取双侧性腺周围脂肪组织置于电子天平上称重,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的双侧性腺周围脂肪组织湿重差异。
与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧性腺周围脂肪组织湿重显著增加(图19)。
结果显示,雌性成年C57BL/6正常小鼠的双侧性腺周围脂肪组织湿重平均为0.54g,雌性成年Glce突变纯合子小鼠的双侧性腺周围脂肪组织湿重平均为3.10g,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的双侧性腺周围脂肪组织湿重平均为1.66g,雄性成年Glce突变纯合子小鼠的双侧性腺周围脂肪组织湿重平均为2.85g,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的双侧性腺周围脂肪组织湿重平均为1.24g,成年Glce突变纯合子小鼠(包含雌雄)的双侧性腺周围脂肪组织湿重平均为2.98g,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧性腺周围脂肪组织湿重显著增加。
7.2Glce突变纯合子小鼠的双侧肾周脂肪组织分析
将Glce突变纯合子小鼠解剖后取双侧肾周脂肪组织置于电子天平上称重,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的双侧肾周脂肪组织湿重差异。
与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧肾周脂肪组织湿重显著增加(图20)。
结果显示,雌性成年C57BL/6正常小鼠的双侧肾周脂肪组织湿重平均为0.36g,雌性成年Glce突变纯合子小鼠的双侧肾周脂肪组织湿重平均为2.06g,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的双侧肾周脂肪组织湿重平均为1.18g,雄性成年Glce突变纯合子小鼠的双侧肾周脂肪组织湿重平均为1.61g,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的双侧肾周脂肪组织湿重平均为0.87g,成年Glce突变纯合子小鼠(包含雌雄)的双侧肾周脂肪组织湿重平均为1.83g,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧肾周脂肪组织湿重显著增加。
7.3Glce突变纯合子小鼠的内脏脂肪组织分析
将Glce突变纯合子小鼠解剖后取双侧性腺周围脂肪组织和双侧肾周脂肪组织置于电子天平上称重,内脏脂肪组织湿重总重为双侧性腺周围脂肪组织和双侧肾周脂肪组织湿重之和,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的内脏脂肪组织湿重差异。
与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的内脏脂肪组织湿重显著增加(图21)。
结果显示,雌性成年C57BL/6正常小鼠的内脏脂肪组织湿重平均为0.90g,雌性成年Glce突变纯合子小鼠的内脏脂肪组织湿重平均为5.16g,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的内脏脂肪组织湿重平均为2.84g,雄性成年Glce突变纯合子小鼠的内脏脂肪组织湿重平均为4.45g,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的内脏脂肪组织湿重平均为2.11g,成年Glce突变纯合子小鼠(包含雌雄)的内脏脂肪组织湿重平均为4.81g,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的内脏脂肪组织湿重显著增加。
7.4Glce突变纯合子小鼠的皮下脂肪组织(如双侧腹股沟脂肪组织)分析
将Glce突变纯合子小鼠解剖后取双侧腹股沟脂肪组织置于电子天平上称重,皮下脂肪组织湿重为双侧腹股沟脂肪组织湿重之和,研究成年Glce突变纯合子小鼠与成年C57BL/6正常小鼠的皮下脂肪组织湿重差异。
与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧腹股沟脂肪组织(代表皮下脂肪组织)湿重显著增加(图22)。
结果显示,雌性成年C57BL/6正常小鼠的双侧腹股沟脂肪组织湿重平均为0.04g,雌性成年Glce突变纯合子小鼠的双侧腹股沟脂肪组织湿重平均为3.34g,与雌性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);雄性成年C57BL/6正常小鼠的双侧腹股沟脂肪组织湿重平均为0.13g,雄性成年Glce突变纯合子小鼠的双侧腹股沟脂肪组织湿重平均为3.07g,与雄性成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01);成年C57BL/6正常小鼠(包含雌雄)的双侧腹股沟脂肪组织湿重平均为0.10g,成年Glce突变纯合子小鼠(包含雌雄)的双侧腹股沟脂肪组织湿重平均为3.20g,与成年C57BL/6正常小鼠相比较具有统计学显著差异(P<0.01)。
上述结果充分表明,与成年C57BL/6正常小鼠相比较,成年Glce突变纯合子小鼠的双侧腹股沟脂肪组织(代表皮下脂肪组织)湿重显著增加。
实施例8Glce突变纯合子小鼠的中风行为学分析
将带Cre重组酶的Glce基因突变纯合子小鼠置于清洁饲养环境饲养。该小鼠动物模型在进入老年期(约1.5年)发生中风样症状。通过一系列中风行为学实验评价小鼠动物模型的中风样症状及严重程度,实验方法参考2001年发表于Stroke上的评价大鼠中风症状严重程度的一系列系统的行为学实验(Chen J,et al.Stroke.2001),包括运动测试、感觉测试、平衡能力测试、本体的反射测试、异常的活动能力测试等,具体为平放测试、提尾测试、视觉触觉测试、本体觉测试、平衡木测试、耳廓反射、睑闭反射、惊跳反射、抽搐、痉挛、肌张力障碍等行为测试,对小鼠的行为进行评分。
Glce基因突变纯合子小鼠组40只有10只在老年时期发生中风,中风发生率为25%;对照组C57BL/6小鼠组40只小鼠有2只在老年时期发生中风,中风发生率为5%,结果表明,Glce基因突变纯合子小鼠与C57BL/6小鼠相比较更易发生中风。
8.1运动测试。
(1)平放测试:将小鼠放置于平地上,观察小鼠是否能够正常行走,正常行走得分0分;是否能够直行,不能直行得分1分,否则得分0分;是否围绕损伤侧转圈,围绕损伤侧转圈得分1分,否则得分0分;是否向损伤侧倾倒,向损伤侧倾倒得分1分,否则得分0分。
(2)提尾测试:提起小鼠的尾巴,观察小鼠肢体活动的情况,是否前肢向内弯曲、爪子抓紧,若小鼠前肢向内弯曲、爪子抓紧得分1分,否则得分0分;是否后肢向内弯曲、爪子抓紧,若小鼠后肢向内弯曲、爪子抓紧得分1分,否则得分0分;是否30秒内头部抬起、与垂直轴角度大于10度,若小鼠30秒内头部抬起、与垂直轴角度大于10度得分1分,否则得分0分。
8.2感觉测试。
(1)视觉触觉测试:把握住小鼠身体,让小鼠前肢能够自由活动,使小鼠面向台子边缘或者笼子边缘,将小鼠快速靠近台子边缘或者笼子边缘,观察小鼠前肢是否能够快速向前伸、并且及时张开爪子准确抓住台子边缘或者笼子边缘。若小鼠前肢不能快速向前伸、并且及时张开爪子准确抓住台子边缘或者笼子边缘得分1分,否则得分0分。
(2)本体觉测试:把握住小鼠身体,让小鼠后肢能够自由活动,将小鼠前肢放置于台子边缘或者笼子边缘、后肢悬空,使用镊子夹小鼠后肢大腿肌肉,观察小鼠后肢是否能够快速回缩。若小鼠后肢不能快速回缩得分1分,否则得分0分。
8.3平衡能力测试。
将小鼠放置于平衡木一端,观察小鼠在平衡木上自由活动的情况,主要包括以下7种情况:(1)是否能够保持身体平衡、在平衡木上自由行走,若小鼠能够保持身体平衡、在平衡木上自由行走得分0分。
(2)是否抓住平衡木边缘,若小鼠抓住平衡木边缘得分1分。
(3)抱住平衡木,但是有一后肢掉落,若小鼠抱住平衡木,但是有一后肢掉落得分2分。
(4)抱住平衡木,但是有两后肢掉落,或者在平衡木上旋转,并且在平衡木上的时间大于60秒,若小鼠抱住平衡木,但是有两后肢掉落,或者在平衡木上旋转,并且在平衡木上的时间大于60秒得分3分。
(5)尝试在平衡木上保持平衡但是最终掉落,在平衡木上的时间大于40秒,若小鼠尝试在平衡木上保持平衡但是最终掉落,在平衡木上的时间大于40秒得分4分。
(6)尝试在平衡木上保持平衡但是最终掉落,在平衡木上的时间大于20秒,若小鼠尝试在平衡木上保持平衡但是最终掉落,在平衡木上的时间大于20秒得分5分。
(7)没有尝试在平衡木上保持平衡或者抱紧平衡木的欲望,在平衡木上的时间小于20秒掉落,若小鼠没有尝试在平衡木上保持平衡或者抱紧平衡木的欲望,在平衡木上的时间小于20秒掉落得分6分。
8.4本体的反射测试及异常的活动能力测试。
(1)耳廓反射:使用棉签刺激小鼠耳道,观察小鼠是否有甩头反应,若有说明小鼠具有耳廓反射,若无说明小鼠没有耳廓反射、得分1分。
(2)睑闭反射:使用棉签刺激小鼠虹膜,观察小鼠是否有闭上眼睑的反应,若有说明小鼠具有睑闭反射,若无说明小鼠没有睑闭反射、得分1分。
(3)惊跳反射:制造一个大的噪声,比如水瓶掉落,观察小鼠是否有受惊吓跳起来的反应,若有说明小鼠具有惊跳反射,若无说明小鼠没有惊跳反射、得分1分。
(4)观察小鼠是否出现抽搐、痉挛、肌张力障碍等现象。若小鼠出现抽搐、痉挛、肌张力障碍等现象,得分1分。
计算每一项相加的总得分,总得分1-6分为轻度损伤,总得分7-12分为中度损伤,总得分13-18分为重度损伤。
例如,图23中的小鼠在平放测试中不能直行得分1分,围绕损伤侧转圈得分1分,向损伤侧倾倒得分1分;在提尾测试中前肢向内弯曲、爪子抓紧得分1分,后肢向内弯曲、爪子抓紧得分1分,30秒内头部抬起、与垂直轴角度大于10度得分1分;在视觉触觉测试中前肢能快速向前伸、并且及时张开爪子准确抓住台子边缘或者笼子边缘得分0分;在本体觉测试中后肢能快速回缩得分0分;在平衡木测试中小鼠尝试在平衡木上保持平衡但是最终掉落,在平衡木上的时间大于40秒得分4分;小鼠具有耳廓反射得分0分;具有睑闭反射得分0分;具有惊跳反射得分0分;小鼠出现抽搐得分1分。该小鼠的总得分为:1+1+1+1+1+1+0+0+4+0+0+0+1=11分,评价为中度中风。
实施例9Glce突变纯合子小鼠的脑组织梗塞分析
通过TTC(2,3,5—氯化三苯基四氮唑)染色方法检测中风小鼠脑组织的梗塞情况。操作步骤是:麻醉后直接取脑,-20度冰箱中速冻5-10分钟左右,便于切片。切片:每隔1mm切一片。将切片置于TTC中,常规浓度为2%。用锡箔纸盖住后,放入37度温箱15-30min,不时翻动脑片,使脑片均匀接触到染色液。然后使用4%多聚甲醛固定30min。拍照。
TTC是脂溶性光敏感复合物,可用来染色检测哺乳动物组织的缺血梗塞。它是呼吸链中吡啶-核苷结构酶系统的质子受体,与正常组织中的脱氢酶反应而呈红色,而缺血组织内脱氢酶活性下降,不能反应,故不会产生变化呈苍白。
结果如图24所示,结果显示,经过TTC染色之后,中风小鼠的正常脑组织呈红色,而梗塞的脑组织呈苍白。观察到嗅球、前额叶、胼胝体、海马组织、纹状体、杏仁核、下丘脑、颞叶、小脑、脑桥、延髓等均有梗塞现象。
实施例10用治疗焦虑症(如广泛性焦虑)的药物验证药物筛选平台
在本实施例中,给实施例1构建的模型动物小鼠注射当前临床治疗中焦虑症的药物帕罗西汀或枸杞多糖,随即对模型动物小鼠的焦虑严重程度进行评估。
结果表明,药物帕罗西汀或枸杞多糖可显著降低模型动物小鼠的焦虑症的严重程度,具体地,可显著降低模型动物小鼠的焦虑症的发生率。
实施例11利用治疗焦虑症或其相关疾病(如广泛性焦虑)的药物筛选平台筛选候选药物
在本实施例中,计划通过给实施例1构建的模型动物小鼠注射焦虑症或其相关疾病的治疗药物,随即对模型动物小鼠的焦虑症状的严重程度进行评估。
通过与给安慰剂的模型动物小鼠比较焦虑症状的严重程度的差异,能够显著降低焦虑症的发生率的候选药物,即为该焦虑症或其相关疾病的潜在治疗药物。
其它焦虑症或其相关疾病也可以参照上述方法,对模型动物小鼠的焦虑症状的严重程度进行评估,从而筛选候选药物。
在本发明提及的所有文献都在本申请中引用作为参考,就如同每一篇文献被单独引用作为参考那样。此外应理解,在阅读了本发明的上述讲授内容之后,本领域技术人员可以对本发明作各种改动或修改,这些等价形式同样落于本申请所附权利要求书所限定的范围。
参考文献:
1.Kreuger J,Spillmann D,Li JP,Lindahl U.Interactions between heparan sulfate and proteins:the concept of specificity.The Journal of cell biology2006;174:323-7.
2.Li JP,Gong F,Hagner-McWhirter A,Forsberg E,Abrink M,Kisilevsky R,Zhang X,Lindahl U.Targeted disruption of a murine glucuronyl C5-epimerase gene results in heparan sulfate lacking L-iduronic acid and in neonatal lethality.The Journal of biological chemistry 2003;278:28363-6.
3.Jia J,Maccarana M,Zhang X,Bespalov M,Lindahl U,Li JP.Lack of L-iduronic acid in heparan sulfate affects interaction with growth factors and cell signaling.The Journal of biological chemistry 2009;284:15942-50.
4.Grigorieva E,Eshchenko T,Rykova VI,Chernakov A,Zabarovsky E,Sidorov SV.Decreased expression of human D-glucuronyl C5-epimerase in breast cancer.International Journal of cancer 2008;122:1172-6.
5.Prudnikova TY,Mostovich LA,Domanitskaya NV,Pavlova TV,Kashuba VI,Zabarovsky ER,Grigorieva EV.Antiproliferative effect of D-glucuronyl C5-epimerase in human breast cancer cells.Cancer Cell international 2010;10:27.
6.Grigorieva EV,Prudnikova TY,Domanitskaya NV,Mostovich LA,Pavlova TV,Kashuba VI,Zabarovsky ER.D-glucuronyl C5-epimerase suppresses small-cell lung cancer cell proliferation in vitro and tumour growth in vivo.British Journal of cancer 2011;105:74-82.
- 还没有人留言评论。精彩留言会获得点赞!