一种调控高山被孢霉发酵产花生四烯酸的制备方法与流程
本发明涉及花生四烯酸,尤其是涉及一种调控高山被孢霉发酵产花生四烯酸的制备方法。
背景技术:
花生四烯酸(Arachidonic acid,ARA)即全顺式-5,8,11,14-二十碳四烯酸,分子式为C20H32O2(Dedyukhina E G.et al.2011),为ω-6高级多不饱和脂肪酸,具有促进婴幼儿脑部发育、调节神经传导、降低胆固醇等多种重要的生理功能,在膳食营养、医药、化妆品等多领域有广泛应用(Njie-Mbye Y F.et al.2013)。ARA的传统来源是从动物肝脏、深海鱼油、月见草种子等提取而来(Kusumoto A.et al.2007)。但由于动物组织中ARA含量有限且有异味,植物生长周期漫长,难以满足市场需求,利用微生物发酵法生产ARA具有周期短、不受地域限制等优点,成为当前研究的热点(Wu W.et al.2015)。
常见的可以合成ARA的微生物主要包括部分细菌、酵母菌、微藻和多数丝状真菌(Dyal S D.et al.2005)。工业上常用的菌种主要集中在藻状菌纲:被孢霉属,根霉属,耳霉属,毛霉属及枝霉属等。其中,高山被孢霉脂质积累可达菌体干重的40%左右,ARA含量亦可达到PUFAs的50%以上,成为工业生产的主要菌株(Li X.et al.2015)。高山被孢霉作为生产使用的微生物已经通过美国农业部的安全性评估,是迄今为止生产长链多不饱和脂肪酸的微生物中少数具有正式安全性评估(GRAS)的菌种之一(Streekstra H.et al.2015;Nisha A.et al.2009)。
目前,关于微生物法生产ARA的研究主要集中在筛选优良菌株、ARA的生物合成、培养基及培养条件的优化和工业化大规模生产工艺的探索上。除菌株本身对微生物代谢产物的影响外,培养基的组成对其生长和产物积累也有很大的影响,如最佳碳源、氮源、碳氮比、辅因子和前体的添加等。当前,已有大量关于碳氮源等基础营养成份的研究,而辅因子和前体物添加的研究成果较少,亦有大量分歧的研究结果。由于高山被孢霉发酵生产ARA的代谢过程较为复杂,成产过程的代谢调控也成为难点之一。
高山被孢霉发酵生产ARA的油脂合成途径倾向于认为是FAS途径(Ward O P.et al.2005)。在ARA的生物合成代谢过程中中,多不饱和脂肪酸的合成本质是碳链的不断延长和氧化脱氢的酶促反应(NADPH为还原力)的共同作用。首先由乙酰-CoA在脂肪酸合酶复合体的酶促作用下,历经乙酰基转移、丙二酸单酰基转移、缩合、还原、脱水、再还原等循环反应,生成C16:0;接着由C16:0经延长酶加碳,经脂肪酸去饱和酶脱氢引入双键(NADPH为电子供体,分子氧为电子受体),合成一系列多不饱和脂肪酸,ARA作为ω-6系列的终端不饱和脂肪酸由此产生。研究认为,M.alpina菌体内,用于脂肪酸合成特别是多不饱和脂肪酸脱氢过程的NADPH主要由苹果酸在苹果酸酶的脱羧作用下产生丙酮酸的过程产生的,可见苹果酸酶的活性在脂肪酸合成过程中至关重要(Wynn J P.et al.2001)。Kendrick(Kendrick A.et al.1992)等的研究认为苹果酸能刺激苹果酸酶的活性;Lan(Lan W Z.et al.2002)等研究结果分析MSG刺激了磷酸戊糖途径(PPP),提高了葡萄糖-6-磷酸脱氢酶(G6PDH)活力,产生更多的NADPH,为油脂的合成提供了还原力,促进了ARA的合成。本发明基于此代谢路径及前人的研究成果,通过苹果酸和MSG的添加对其发酵过程进行调控,以期提高发酵产能。
技术实现要素:
本发明的第一目的在于提供高山被孢霉(Mortierella alpina)LU166。
本发明的第二目的在于提供一种调控高山被孢霉发酵产花生四烯酸的制备方法。
所述高山被孢霉(Mortierella alpina)LU166,已于2016年07月05日保藏于中国微生物菌种保藏管理委员会普通微生物中心,地址:北京市朝阳区北辰西路1号院3号中国科学院微生物研究所,邮编:100101,保藏中心登记入册编号:CGMCC No.12764。
所述调控高山被孢霉发酵产花生四烯酸的制备方法,包括以下步骤:
1)将高山被孢霉(Mortierella alpina)LU166经过平板培养基培养和种子培养基活化,得种子液;
2)将步骤1)得到的种子液接种于发酵培养基中,加入有机酸,待发酵稳定菌丝体合成后再加入有机酸和氮源,促进油脂大量积累和花生四烯酸的转化,得花生四烯酸(ARA)。
在步骤1)中,所述平板培养基的组成可为:马铃薯200g/L,葡萄糖20g/L,琼脂20g/L;所述种子培养基的组成可为:葡萄糖30g/L,酵母粉4g/L,KH2PO4 2g/L,MgSO4·7H2O 0.3g/L,pH调至6.0。
在步骤2)中,所述发酵培养基的组成可为:葡萄糖50g/L,酵母粉8g/L,玉米浆3g/L,KH2PO4 2g/L,谷氨酸1g/L,MgSO4·7H2O 0.1g/L,ZnSO4·7H2O 0.1g/L,CaCO3 0.05g/L,1000×微量元素1mL;pH调至6.0;1000×微量元素的组成可为:MnCl2·4H2O 0.8g/L,H3BO30.5g/L,FeCl3·6H2O 0.2g/L,NiSO4·6H2O 0.05g/L,CoCl2·6H2O 0.005g/L,CuSO4·5H2O0.002g/L,放置4℃保存;所述发酵稳定菌丝体合成的培养时间可为120h;
所述有机酸可采用苹果酸,所述苹果酸的质量百分浓度可为0.025%。
所述氮源可采用有机氮源,所述有机氮源可采用NaNO3、KNO3、谷氨酸钠(MSG)等中的一种,所述谷氨酸钠(MSG)的质量百分浓度可为0.05%。
本发明调控原理如下:初期添加苹果酸主要提高了生物量,对ARA合成有一定的抑制,而初期和稳定期复合添加苹果酸除了进一步提高了生物量,对油脂和ARA的合成也有一定的促进作用,尤其是促进了油脂向ARA的转化,使得ARA占油脂的比例明显提高。对比只在实验初期添加苹果酸和初期添加苹果酸复合稳定期添加MSG的生物量提高率可以发现MSG的添加对生物量的提高作用不明显,而从油脂和ARA的产量变化可以看到,稳定期MSG的添加对两者的合成均有明显的促进作用,尤其是对总油的提高作用非常显著。在以上添加策略基础上,本发明最终确定的复合添加策略(在发酵初期添加苹果酸,复合稳定期添加苹果酸和MSG)的发酵效果最好,该策略下生物量、油脂和ARA产量得到了最大幅度的增加,产量分别达到24.5g/L、12.4g/L和6.2g/L,相比对照组分别提高了21.21%、50.67%和43.47%。
附图说明
图1为发酵初期添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵细胞干重、总油及ARA产量的影响。
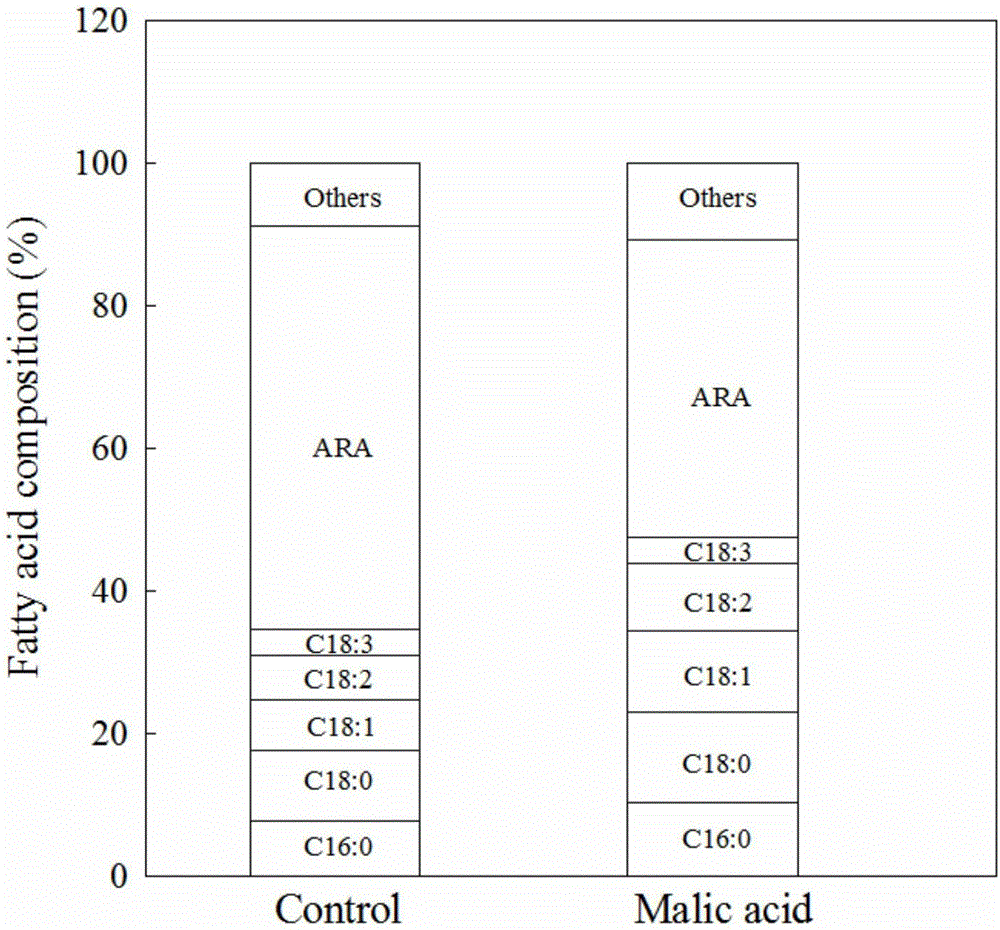
图2为发酵初期添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵总油油脂成分的影响。
图3为发酵初期和稳定期两次添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵细胞干重、总油及ARA产量的影响。
图4为发酵初期和稳定期两次添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵油脂、ARA含量的影响。
图5为初期添加苹果酸复合稳定期添加不同氮源对高山被孢霉(Mortierella alpina)LU166发酵细胞干重、总油及ARA产量的影响。
图6为初期添加苹果酸复合稳定期添加不同氮源对高山被孢霉(Mortierella alpina)LU166发酵油脂、ARA含量的影响。
图7为流加苹果酸复合添加MSG对高山被孢霉(Mortierella alpina)LU166发酵细胞干重、总油及ARA产量的影响。
图8为流加苹果酸复合添加MSG对高山被孢霉(Mortierella alpina)LU166发酵油脂、ARA含量的影响。
具体实施方式
以下实施例用于进一步说明本发明。
实施例1.实验初期添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵的影响
高山被孢霉(Mortierella alpina)LU166经PDA平板活化后,制成孢子悬浮液,接于250mL摇瓶中培养,载液量为50mL,150rpm,28℃,活化2天,转二级种子,将活化后的菌株接入250mL摇瓶培养,载液量为50mL,在基础发酵培养基中添加0.025%苹果酸,28℃,150rpm培养。结果如图1和2所示。
由图1可见,在在发酵初始阶段添加苹果酸促进了生物量和总油脂产量,但在一定程度上抑制了ARA的产量。结合图2分析可见,与对照组相比,实验组中ARA占总油的比例较低,而相对应地C16:0、C18:1和C18:2的占比较高,这说明苹果酸的添加虽然促进了FAS途径合成油脂,但并没有提高饱和脂肪酸和单不饱和脂肪酸向下游多不饱和脂肪酸ARA的有效转化。
实施例2.实验初期和稳定期复合添加苹果酸对高山被孢霉(Mortierella alpina)LU166发酵的影响
考虑到初期添加苹果酸主要用于促进了细胞和总油的积累,在稳定期没有足够的苹果酸作为反应底物刺激苹果酸酶的活性提供足够还原力NADPH,因此,实验考虑在初期添加苹果酸的基础上,在稳定期再添加0.025%的苹果酸,以期提高ARA的转化率。结果如图3和4所示。
由图3和4可见,在稳定期再次添加苹果酸不仅提高了菌体生物量和油脂产量,ARA的产量和含量也得到了一定的提升,虽然总油含量有稍微降低,但ARA占细胞的比重和ARA占总油的比重均有所上升。可见在稳定期适当添加苹果酸有利于饱和脂肪酸和单不饱和脂肪酸向ARA的转化,这可能是由于稳定期苹果酸的添加再次激活了苹果酸酶的活性,“转氢酶循环”通量增加,生成的NADPH此时可有效参与到脂肪酸去饱和的过程,促进了ARA的转化。
实施例3.实验初期添加苹果酸和稳定期添加氮源对高山被孢霉(Mortierella alpina)LU166发酵的影响
少量氮源的添加能够激活苹果酸酶的活性,解除NADPH的限制,促进ARA的转化。本实验在实验初期添加苹果酸的基础上在细胞稳定期复合添加三种不同的氮源NaNO3、KNO3和谷氨酸钠(MSG)(添加浓度皆为0.05%),考察其对高山被孢霉(Mortierella alpina)LU166中ARA的转化影响。结果如图5和6所示。
由图5和6可以看出,在培养初期添加苹果酸的基础上稳定期复合添加氮源不仅提高了生物量、总油脂和ARA的产量也在一定程度上提高了总油和ARA在细胞中的含量在添加的不同氮源中,MSG对促进ARA的合成效果最为明显,NaNO3次之,而KNO3效果最低。MSG效果最佳,可能是因为MSG作为有机氮源,转化为NH4+的效率更高,NH4+作为氮源被有效吸收,激发了苹果酸酶的活性,并且MSG可以被有效转化为α-酮戊二酸,促进TCA循环生成乙酰-CoA,为脂肪酸合成提供更多前体物。
实施例4.实验初期添加苹果酸和稳定期复合添加苹果酸及氮源对高山被孢霉(Mortierella alpina)LU166发酵的影响
在实施例2和3的基础上,实验进一步考虑在实验初期添加苹果酸,在稳定期复合添加苹果酸和MSG,考察对高山被孢霉(Mortierella alpina)LU166的发酵影响,以期进一步提高苹果酸酶脱羧反应的通量,提供更多的NADPH还原力,从而提高ARA的合成。结果如图7和8所示。
由图7可见,在稳定期复合添加苹果酸和MSG进一步提高了ARA的产量。结合图8看出,实验组中ARA占总油脂的比例没有提高,可见ARA产量的提高来源于油脂产量的提高。对比以上策略,实施例4的添加策略下获得最佳发酵效果,生物量、油脂和ARA产量得到了最大幅度的增加,产量分别达到24.5g/L、12.4g/L和6.2g/L,相比对照组分别提高了21.21%、50.67%和43.47%。
- 还没有人留言评论。精彩留言会获得点赞!