用于人乳头状瘤病毒的疫苗及其使用方法与流程
本申请是申请日为2011年10月12日的题为“用于人乳头状瘤病毒的疫苗及其使用方法”的中国专利申请号201180074173.2的分案申请。
本发明涉及改进的人乳头状瘤病毒(hpv)疫苗、用于诱导免疫应答的改进的方法、以及用于针对hpv来预防性和/或治疗性免疫个体的改进的方法。
背景技术:
乳头状瘤病毒是小dna病毒,其包含至多7个早期基因和2个晚期基因。通常,乳头状瘤病毒早期基因表示为e1-e7,并且乳头状瘤病毒晚期基因表示为l1和l2。若干个物种的动物可被乳头状瘤病毒家族的成员感染。
人乳头状瘤病毒(hpv)感染是常见的,并且可发生性传播。基于dna序列的同源性,hpv已经分成56种或更多种类型。16和18型hpv(其引起上皮异常增生和其它病变)通常和下列情况有关:癌症风险增加,特别是子宫颈、阴道、阴门以及肛管的原位和侵入性癌瘤。在全世界将近88%的子宫颈癌是hpv亚型16、18、45、31、33、52以及58所造成的结果。此外,各种研究已揭示hpv6和hpv11存在于大多数发生的复发性呼吸道乳头状瘤病中。尽管已知了hpv6和hpv11与生殖器疣的相关性并且在小百分比的子宫颈癌病例中发现了它们,但已经在2.6%至5.2%的所有情况的低级子宫颈病变中发现了hpv6和hpv11。此外,hpv6和hpv11现已发现与大约20%的现被视为子宫颈癌先兆的低级鳞状上皮内病变,包括2级和3级子宫颈上皮内瘤形成和轻度子宫颈发育不良有关。更多的研究已揭示了两种血清型与各种形式的耳鼻喉科疾病,包括生殖器疣、复发性呼吸道乳头状瘤病、肺癌、扁桃体癌、喉癌、以及头颈部本来良性肿瘤的其它恶性转化和发育不良的相关性。此研究的发现结果揭示了使用共有序列dna疫苗来对抗hpv6和hpv11的光明前景。
dna疫苗相对于更加传统的疫苗接种法具有许多概念上的优点,如活减毒病毒和重组蛋白基疫苗。dna疫苗安全、稳定、容易生产并且在临床试验的人中具有良好耐受性,这表明质粒整合的迹象微乎其微[martin,t.等,plasmiddnamalariavaccine:thepotentialforgenomicintegrationafterintramuscularinjection.humgenether,1999.10(5):第759-68页;nichols,w.w.等,potentialdnavaccineintegrationintohostcellgenome.annnyacadsci,1995.772:第30-9页]。另外,dna疫苗非常适于反复施用,因为疫苗的效果实际上不受到对于载体预先存在的抗体滴度的影响[chattergoon,m.、j.boyer以及d.b.weiner,geneticimmunization:anewerainvaccinesandimmunetherapeutics.fasebj,1997.11(10):第753-63页]。然而,在移至较大动物时,临床采用dna疫苗的一个主要障碍是平台免疫原性的降低[liu,m.a.和j.b.ulmer,humanclinicaltrialsofplasmiddnavaccines.advgenet,2005.55:第25-40页]。dna疫苗免疫原工程化中的近期技术进展如密码子优化、rna优化和添加免疫球蛋白前导序列已经提高了dna疫苗的表达和免疫原性[andre,s.等,increasedimmuneresponseelicitedbydnavaccinationwithasyntheticgp120sequencewithoptimizedcodonusage.jvirol,1998.72(2):第1497-503页;deml,l.等,multipleeffectsofcodonusageoptimizationonexpressionandimmunogenicityofdnacandidatevaccinesencodingthehumanimmunodeficiencyvirustype1gagprotein.jvirol,2001.75(22):第10991-1001页;laddy,d.j.等,immunogenicityofnovelconsensus-baseddnavaccinesagainstavianinfluenza.vaccine,2007.25(16):第2984-9页;frelin,l.等,codonoptimizationandmrnaamplificationeffectivelyenhancestheimmunogenicityofthehepatitiscvirusnonstructural3/4agene.genether,2004.11(6):第522-33页],以及质粒递送系统方面的近期开发的技术如电穿孔[hirao,l.a.等,intradermal/subcutaneousimmunizationbyelectroporationimprovesplasmidvaccinedeliveryandpotencyinpigsandrhesusmacaques.vaccine,2008.26(3):第440-8页;luckay,a.等,effectofplasmiddnavaccinedesignandinvivoelectroporationontheresultingvaccine-specificimmuneresponsesinrhesusmacaques.jvirol,2007.81(10):第5257-69页;ahlen,g.等,invivoelectroporationenhancestheimmunogenicityofhepatitiscvirusnonstructural3/4adnabyincreasedlocaldnauptake,proteinexpression,inflammation,andinfiltrationofcd3+tcells.jimmunol,2007.179(7):第4741-53页]。另外,研究表明相比于单独的天然抗原,共有免疫原的使用能够增加细胞免疫应答的宽度[yan.,j.等,enhancedcellularimmuneresponseselicitedbyanengineeredhiv-1subtypebconsensus-basedenvelopednavaccine.molther,2007.15(2):第411-21页;rolland,m.等,reconstructionandfunctionofancestralcenter-of-treehumanimmunodeficiencyvirustype1proteins.jvirol,2007.81(16):第8507-14]。
仍存在对于改进的疫苗和预防与治疗hpv感染、尤其是引起子宫颈癌和/或肺癌、扁桃体癌以及喉癌的感染的方法的需要。
技术实现要素:
发明概述
本发明的方面包括核酸分子,所述核酸分子包含编码hpv抗原的选自由以下各项组成的组的核苷酸序列:seqidno:1、seqidno:3、seqidno:5、seqidno:7;其具有90%同源性的片段;及其组合。在一些方面,所述核酸分子包括包含选自由以下各项组成的组的核苷酸序列的核酸分子:编码seqidno:2、seqidno:4、seqidno:6或seqidno:8的核苷酸序列;编码与seqidno:2、seqidno:4、seqidno:6或seqidno:8具有至少90%同源性的氨基酸片段的核苷酸序列;编码seqidno:2、seqidno:4、seqidno:6或seqidno:8的核苷酸序列的片段;以及编码与seqidno:2、seqidno:4、seqidno:6或seqidno:8具有至少90%同源性的氨基酸片段的核苷酸序列的片段。
在一些方面,存在包含本文所述的核酸分子的药物组合物。
在其它方面,提供在个体中诱导针对hpv的免疫应答的方法,所述方法包括向所述个体施用包含本文所述的核酸分子的组合物。
本发明还包括包含本文所述的核酸分子的重组疫苗。所述重组疫苗包括重组牛痘疫苗、活减毒疫苗以及dna质粒。
本发明的一些方面包括包含选自由以下各项组成的组的氨基酸序列的蛋白质:seqidno:2、seqidno:4、seqidno:6或seqidno:8;以及与seqidno:2、seqidno:4、seqidno:6或seqidno:8具有至少90%同源性的序列的片段。
还提供在个体中诱导针对hpv的免疫应答的方法,所述方法包括向所述个体施用本文提供的重组疫苗。
在一些方面,提供包含以下各项的药物组合物:编码hpv抗原的核苷酸序列seqidno:1和seqidno:3;或其片段;或其具有90%同源性的片段。还存在在个体中诱导针对hpv亚型6或11的免疫应答的方法,所述方法包括向所述个体施用前述药物组合物。
在一些方面,提供包含以下各项的药物组合物:编码hpv抗原的核苷酸序列seqidno:5和seqidno:7;或其片段;或其具有90%同源性的片段。还存在在个体中诱导针对hpv亚型33或58的免疫应答的方法,所述方法包括向所述个体施用前述药物组合物。
附图说明
图1.基于e6和e7比对的相邻接合评估的系统发生树(分别为a和b)。星号指示每个树上的共有序列的位置。
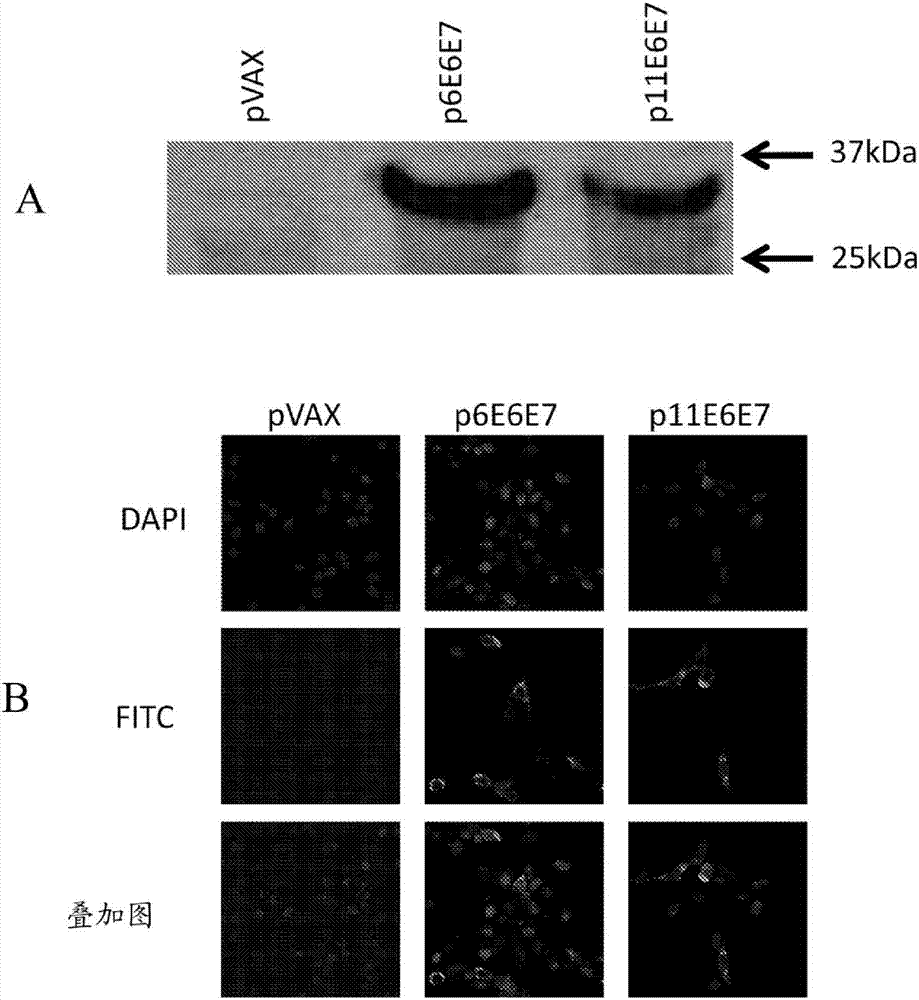
图2.p6e6e7和p11e6e7的体内表达。从裂解的转染293-t细胞分离基因产物,穿过sds-page凝胶,并使用放射自显影术进行检测。hpv6和hpv11e6/e7蛋白皆各自为大约32kda(a)。人横纹肌肉瘤(rd)细胞也用p6e6e7和p11e6e7转染并且随后在免疫荧光染色之后固定。fitc荧光确定了p6e6e7和p11e6e7的表达(b)。dapi荧光确定了hoescht染色之后的核定位。
图3.ifn-γelispot测定示出p6e6e7(a)和p11e6e7(b)在c57bl/6小鼠中诱导稳固的细胞介导的应答。使用在三次两周一次的免疫之后从各对应的组(每组5只小鼠)的小鼠中分离的脾细胞进行测定。每次免疫包括每种构建体20μg。组合组中的小鼠以每种构建体20μg接受p6e6e7和p11e6e7两者,每次免疫共计40μgdna。经由im注射,随后进行电穿孔来施用dna(*指示p<0.0001;
图4.使用个别肽进行另外的ifn-γelispot测定以表征优势肽。从疫苗接种的小鼠和阴性对照中分离的脾细胞用涵盖整个hpv6e6/e7融合蛋白(a)或hpv11e6/e7融合蛋白(b)的重叠肽刺激。
图5a和图5b.由细胞内细胞因子染色表征的抗原特异性t细胞的细胞因子产生。将从接种了p6e6e7(图5a)和p11e6e7(图5b)的小鼠中分离的脾细胞用r10生长培养基、pma或共有基因肽刺激4小时,随后进行表面标记和细胞内染色。以上点图示出产生ifn-γ、il-2和tnf-α的总cd4+或cd8+细胞的减去了背景的百分比的差异。因为阴性对照组的平均值是零,所以带有星号的点的p值不能被确定。
具体实施方式
如本文所使用的,措词"严格杂交条件"或"严格条件"是指这样的条件:在该条件下核酸分子将杂交另一核酸分子,但不杂交其它序列。严格条件是序列依赖性的并且在不同情况下将是不同的。较长序列在较高温度下特异性杂交。通常,对于在限定离子强度和ph下的特定序列,严格条件被选择为低于热熔点(tm)约5℃。tm是这样的温度(在限定的离子强度、ph和核酸浓度下):在该温度下平衡时,与靶序列互补的探针中的50%与靶序列杂交。由于靶序列通常过量存在,因此在tm下平衡时50%的探针被占据。通常,严格条件是这样的条件:其中在ph7.0至8.3下盐浓度低于约1.0m钠离子,通常约0.01至1.0m钠离子(或其它盐),并且对于较短探针、引物或寡核苷酸(例如10至50个核苷酸)温度为至少约30℃,且对于较长探针、引物或寡核苷酸温度为至少约60℃。严格条件还可通过添加去稳定剂如甲酰胺来实现。
核苷酸和氨基酸的序列同源性可使用fasta、blast和gappedblast(altschul等,nuc.acidsres.,1997,25,3389,其以引用的方式整体并入本文)以及paup*4.0b10软件(d.l.swofford,sinauerassociates,massachusetts)来测定。“相似性百分比”使用paup*4.0b10软件(d.l.swofford,sinauerassociates,massachusetts)来计算。共有序列的平均相似性是相较于系统发生树中的所有序列来计算。
简言之,blast算法(表示基本局部比对搜索工具(basiclocalalignmentsearchtool))适用于确定序列相似性(altschul等,j.mol.biol.,1990,215,403-410,其以引用的方式整体并入本文)。进行blast分析的软件可通过国家生物技术信息中心(nationalcenterforbiotechnologyinformation)公开获得(http://www.ncbi.nlm.nih.gov/)。该算法包括首先通过鉴定在查询序列中长度w的短字串,该字串与数据库序列中相同长度的字串比对时匹配或者满足某些正数值阈值分值t,从而来鉴定高分序列配对(hsp)。t被称为邻近字串分值阈值(altschul等,同上)。这些初始邻近字串命中作为用于起始搜索以发现含有它们的hsp的种子。只要累积序列比对分值可以增加,那么字串命中则在两个方向上沿每个序列延伸。字串命中在每个方向上的延伸当遇到下述情况时终止:1)累积序列比对分值从其获得的最大值下降了数量x;2)累积分值为零或零以下,这是由于一个或多个负分值残基比对的累积所致;或者3)到达任一序列的末端。blast算法参数w、t和x决定所述比对的灵敏度和速度。blast程序使用如下默认值:字长(w)为11,blosum62评分矩阵(参见henikoff等,proc.natl.acad.sci.usa,1992,89,10915-10919,其以引用的方式整体并入本文)比对(b)为50,期望值(e)为10,m=5,n=4,以及两条链都比较。blast算法(karlin等,proc.natl.acad.sci.usa,1993,90,5873-5787,其以引用的方式整体并入本文)和gappedblast进行两个序列之间相似性的统计学分析。blast算法提供的一种相似性量度是最小和概率(p(n)),它提供两个核苷酸序列可能偶然发生匹配的概率的指示。例如,如果一个核酸与另一核酸比较中的最小和概率小于约1、优选小于约0.1、更优选小于0.01、并且最优选小于约0.001,则认为该测试核酸与另一核酸相似。
如本文所使用的,术语“基因构建体"是指包含编码蛋白质的核苷酸序列的dna或rna分子。编码序列包括可操作地连接至调控元件(包括能够在核酸分子所施用的个体的细胞内指导表达的启动子和聚腺苷酸化信号)的起始和终止信号。
如本文所使用的,术语"可表达的形式"是指含有可操作地连接至编码蛋白质的编码序列的必需调控元件的基因构建体,以使得当存在于个体的细胞中时,编码序列将被表达。
公开了改进的疫苗,所述疫苗源自用以增强由免疫原诱导的细胞免疫应答的多阶段战略。产生修饰的共有序列。还公开了基因修饰,包括密码子优化、rna优化以及高效免疫球蛋白前导序列添加。新的构建体已经被设计来引发比对应的密码子优化的免疫原更强和更宽的细胞免疫应答。
改进的hpv疫苗是基于蛋白质和编码具有使它们作为可用以诱导出抗-hpv的免疫原特别有效的表位的蛋白质的基因构建体。因此,疫苗可诱导治疗或预防性免疫应答。在一些实施方案中,递送免疫原的方式是dna疫苗、重组疫苗、蛋白质亚单位疫苗、包含免疫原的组合物、减毒疫苗或灭活疫苗。在一些实施方案中,疫苗包含选自由以下各项组成的组的组合:一种或多种dna疫苗、一种或多种重组疫苗、一种或多种蛋白质亚单位疫苗、一种或多种包含免疫原的组合物、一种或多种减毒疫苗以及一种或多种灭活疫苗。
根据一些实施方案,疫苗被递送至个体以调节个体的免疫系统的活性并且由此增强针对hpv的免疫应答。当编码蛋白质的核酸分子被个体细胞所摄取时,核苷酸序列在细胞中表达并且蛋白质由此被递送至个体。提供:在核酸分子如质粒(作为重组疫苗的一部分和作为减毒疫苗的一部分,作为分离的蛋白质或载体的蛋白质部分)上递送蛋白质的编码序列的方法。
提供针对hpv预防性和/或治疗性免疫个体的组合物和方法。
用于递送包含编码免疫原的核苷酸序列的核酸分子组合物可操作地连接至调控元件。组合物可包含:编码免疫原的质粒、包含编码免疫原的核苷酸序列的重组疫苗、编码本发明的蛋白质和/或包含本发明的蛋白质的活减毒病原体、包含本发明的蛋白质的灭活病原体、或包含本发明的蛋白质的组合物如脂质体或亚单位疫苗。本发明还涉及包含组合物的可注射的药物组合物。
seqidno:1包含编码hpv6e6和e7蛋白的共有免疫原的核苷酸序列。seqidno:1包含连接至seqidno:1的5’末端处的核苷酸序列的ige前导序列seqidno:9。seqidno:2包含hpv6e6和e7蛋白的共有免疫原的氨基酸序列。seqidno:2包含位于共有免疫原序列的n-末端处的ige前导序列seqidno:10。ige前导序列是seqidno:10,并且可由seqidno:9编码。
在一些实施方案中,疫苗包含seqidno:2或编码seqidno:2的核酸分子。
在一些实施方案中,seqidno:1的片段可包含786个或更多个核苷酸;在一些实施方案中,可包含830个或更多个核苷酸;在一些实施方案中,可包含856个或更多个核苷酸;并且在一些实施方案中,可包含865个或更多个核苷酸。在一些实施方案中,seqidno:1的片段如本文所列出的那些片段可进一步包含ige前导序列的编码序列。在一些实施方案中,seqidno:1的片段不包含ige前导序列的编码序列。
在一些实施方案中,seqidno:2的片段可包含252个或更多个氨基酸;在一些实施方案中,可包含266个或更多个氨基酸;在一些实施方案中,可包含275个或更多个氨基酸;并且在一些实施方案中,可包含278个或更多个氨基酸。
seqidno:3包含编码hpv11e6和e7蛋白的共有免疫原的核苷酸序列。seqidno:3包含连接至seqidno:3的5’末端处的核苷酸序列的ige前导序列seqidno:9。seqidno:4包含hpv11e6和e7蛋白的共有免疫原的氨基酸序列。seqidno:4包含位于共有免疫原序列的n-末端处的ige前导序列seqidno:10。ige前导序列是seqidno:10,并且可由seqidno:9编码。
在一些实施方案中,疫苗包含seqidno:4或编码seqidno:4的核酸分子。
在一些实施方案中,seqidno:3的片段可包含786个或更多个核苷酸;在一些实施方案中,可包含830个或更多个核苷酸;在一些实施方案中,可包含856个或更多个核苷酸;并且在一些实施方案中,可包含865个或更多个核苷酸。在一些实施方案中,seqidno:3的片段如本文所列出的那些片段可进一步包含ige前导序列的编码序列。在一些实施方案中,seqidno:3的片段不包含ige前导序列的编码序列。
在一些实施方案中,seqidno:4的片段可包含252个或更多个氨基酸;在一些实施方案中,可包含266个或更多个氨基酸;在一些实施方案中,可包含275个或更多个氨基酸;并且在一些实施方案中,可包含278个或更多个氨基酸。
seqidno:5包含编码hpv33e6和e7蛋白的共有免疫原的核苷酸序列。seqidno:5包含连接至seqidno:5的5’末端处的核苷酸序列的ige前导序列seqidno:9。seqidno:6包含hpv33e6和e7蛋白的共有免疫原的氨基酸序列。seqidno:6包含位于共有免疫原序列的n-末端处的ige前导序列seqidno:10。ige前导序列是seqidno:10,并且可由seqidno:9编码。
在一些实施方案中,疫苗包含seqidno:6或编码seqidno:6的核酸分子。
在一些实施方案中,seqidno:5的片段可包含746个或更多个核苷酸;在一些实施方案中,可包含787个或更多个核苷酸;在一些实施方案中,可包含812个或更多个核苷酸;并且在一些实施方案中,可包含820个或更多个核苷酸。在一些实施方案中,seqidno:5的片段如本文所列出的那些片段可进一步包含ige前导序列的编码序列。在一些实施方案中,seqidno:5的片段不包含ige前导序列的编码序列。
在一些实施方案中,seqidno:6的片段可包含235个或更多个氨基酸;在一些实施方案中,可包含248个或更多个氨基酸;在一些实施方案中,可包含256个或更多个氨基酸;并且在一些实施方案中,可包含259个或更多个氨基酸。
seqidno:7包含编码hpv58e6和e7蛋白的共有免疫原的核苷酸序列。seqidno:7包含连接至seqidno:7的5’末端处的核苷酸序列的ige前导序列seqidno:9。seqidno:8包含hpv58e6和e7蛋白的共有免疫原的氨基酸序列。seqidno:8包含位于共有免疫原序列的n-末端处的ige前导序列seqidno:10。ige前导序列是seqidno:10,并且可由seqidno:9编码。
在一些实施方案中,疫苗包含seqidno:8或编码seqidno:8的核酸分子。
在一些实施方案中,seqidno:7的片段可包含752个或更多个核苷酸;在一些实施方案中,可包含794个或更多个核苷酸;在一些实施方案中,可包含819个或更多个核苷酸;并且在一些实施方案中,可包含827个或更多个核苷酸。在一些实施方案中,seqidno:7的片段如本文所列出的那些片段可进一步包含ige前导序列的编码序列。在一些实施方案中,seqidno:7的片段不包含ige前导序列的编码序列。
在一些实施方案中,seqidno:8的片段可包含235个或更多个氨基酸;在一些实施方案中,可包含249个或更多个氨基酸;在一些实施方案中,可包含257个或更多个氨基酸;并且在一些实施方案中,可包含260个或更多个氨基酸。
根据一些实施方案,在个体中诱导针对免疫原的免疫应答的方法包括向所述个体施用选自由以下各项组成的组的共有免疫原的氨基酸序列:hpv6e6和e7、hpv11e6和e7、hpv33e6和e7、hpv58e6和e7、其功能片段、及其可表达的编码序列。一些实施方案包括分离的核酸分子,其编码选自由以下各项组成的组的共有免疫原的氨基酸序列:hpv6e6和e7、hpv11e6和e7、hpv33e6和e7、hpv58e6和e7、及其片段。一些实施方案包括重组疫苗,其编码选自由以下各项组成的组的共有免疫原的氨基酸序列:hpv6e6和e7、hpv11e6和e7、hpv33e6和e7、hpv58e6和e7、及其片段。一些实施方案包括亚单位疫苗,其包含选自由以下各项组成的组的共有免疫原的氨基酸序列:hpv6e6和e7、hpv11e6和e7、hpv33e6和e7、hpv58e6和e7、及其片段。一些实施方案包括活减毒疫苗和/或灭活疫苗,其包含选自由以下各项组成的组的共有免疫原的氨基酸序列:hpv6e6和e7、hpv11e6和e7、hpv33e6和e7、以及hpv58e6和e7。
一方面,本文的疫苗是引发针对主要发现与多种形式的头颈部癌和其它形式的耳鼻喉科疾病有关的hpv亚型的免疫应答的那些疫苗,尤其所述疫苗包含hpv6e6和e7以及hpv11e6和e7,并且优选包含两者。
另一方面,本文的疫苗是引发针对主要发现与美国之外的患者并且更具体来说亚洲的患者的多种形式的子宫颈癌有关的hpv亚型的免疫应答的那些疫苗,尤其所述疫苗包含hpv33e6和e7以及hpv58e6和e7,并且优选包含两者。
改进的疫苗包含蛋白质和编码具有使它们作为可用于诱导抗-hpv免疫应答的免疫原特别有效的表位的蛋白质的基因构建体。因此,可提供疫苗以诱导治疗或预防性免疫应答。在一些实施方案中,递送免疫原的方式是dna疫苗、重组疫苗、蛋白质亚单位疫苗、包含免疫原的组合物、减毒疫苗或灭活疫苗。在一些实施方案中,疫苗包含选自由以下各项组成的组的组合:一种或多种dna疫苗、一种或多种重组疫苗、一种或多种蛋白质亚单位疫苗、一种或多种包含免疫原的组合物、一种或多种减毒疫苗以及一种或多种灭活疫苗。
本发明的方面提供在核酸分子如质粒(作为重组疫苗的一部分和作为减毒疫苗的一部分,作为分离的蛋白质或载体的蛋白质部分)上递送蛋白质的编码序列的方法。
根据本发明的一些方面,提供预防性和/或治疗性免疫个体的组合物和方法。
dna疫苗描述于美国专利号5,593,972、5,739,118、5,817,637、5,830,876、5,962,428、5,981,505、5,580,859、5,703,055、5,676,594以及本文中引用的优先权申请中,其均以引用的方式并入本文。除了这些申请中所述的递送方案之外,递送dna的替代性方法还描述于美国专利号4,945,050和5,036,006中,其都以引用的方式并入本文。
本发明涉及改进的减毒活疫苗、改进的灭活疫苗、和改进的使用重组载体以递送编码抗原的外源基因的疫苗、以及亚单位疫苗和糖蛋白疫苗。减毒活疫苗、使用重组载体以递送外源抗原的那些疫苗、亚单位疫苗以及糖蛋白疫苗的实例描述于美国专利号:4,510,245;4,797,368;4,722,848;4,790,987;4,920,209;5,017,487;5,077,044;5,110,587;5,112,749;5,174,993;5,223,424;5,225,336;5,240,703;5,242,829;5,294,441;5,294,548;5,310,668;5,387,744;5,389,368;5,424,065;5,451,499;5,453,364;5,462,734;5,470,734;5,474,935;5,482,713;5,591,439;5,643,579;5,650,309;5,698,202;5,955,088;6,034,298;6,042,836;6,156,319以及6,589,529中,其均以引用的方式并入本文。
当被细胞摄取时,基因构建体可保持存在于细胞中作为功能性染色体外分子和/或整合到细胞的染色体dna中。dna可引入细胞中,在细胞中其保持为呈一个或多个质粒形式的单独的遗传材料。或者,可整合到染色体中的线性dna可引入到细胞中。当将dna引入细胞中时,可以添加促进dna整合到染色体中的试剂。适用于促进整合的dna序列还可包括在dna分子中。或者,可对细胞施用rna。还涵盖提供呈线性微型染色体形式的基因构建体,包括着丝粒、端粒以及复制起点。基因构建体可在减毒活微生物或在细胞中存活的重组微生物载体中保持为遗传材料的一部分。基因构建体可以是重组病毒疫苗的基因组的一部分,其中遗传材料整合到细胞的染色体中或保持在染色体外。基因构建体包括核酸分子的基因表达所必需的调控元件。元件包括:启动子、起始密码子、终止密码子以及聚腺苷酸化信号。另外,通常需要增强子用于编码靶蛋白或免疫调节蛋白的序列的基因表达。这些元件需要可操作地连接至编码所需蛋白质的序列,并且调控元件需要在其所施用的个体中是可操作的。
起始密码子和终止密码子通常被认为是编码所需蛋白质的核苷酸序列的一部分。然而,这些元件需要在基因构建体所施用的个体中有功能。起始密码子和终止密码子必须与编码序列同框。
所使用的启动子和聚腺苷酸化信号必须在个体的细胞内有功能。
适用于实施本发明,尤其在制造人用基因疫苗中实施本发明的启动子的实例包括但不限于:来自猿猴病毒40(sv40)的启动子、小鼠乳腺肿瘤病毒(mmtv)启动子、人类免疫缺陷病毒(mv)如biv长末端重复(ltr)启动子、莫洛尼氏病毒、alv、巨细胞病毒(cmv)如cmv即刻早期启动子、eb病毒(ebv)、劳氏肉瘤病毒(rsv)以及来自人基因如人肌动蛋白、人肌球蛋白、人血红蛋白、人肌肉肌酸和人金属硫蛋白的启动子。
适用于实施本发明,尤其在制造人用基因疫苗中实施本发明的的聚腺苷酸化信号的实例包括但不限于:sv40聚腺苷酸化信号和ltr聚腺苷酸化信号。具体地说,使用在pcep4质粒(invitrogen,sandiegoca)中的sv40聚腺苷酸化信号,其称为sv40聚腺苷酸化信号。
除了dna表达所需要的调控元件之外,其它元件也可包含在dna分子中。这类另外的元件包括增强子。增强子可选自下组,所述组包括但不限于:人肌动蛋白、人肌球蛋白、人血红蛋白、人肌肉肌酸和病毒增强子,如来自cmv、rsv和ebv的那些增强子。
可提供具有哺乳动物复制起点的基因构建体以便将构建体维持在染色体外并且在细胞中产生构建体的多个拷贝。来自invitrogen(sandiego,ca)的质粒pvax1、pcep4和prep4含有eb病毒复制起点和核抗原ebna-1编码区,该编码区产生高拷贝的附加型复制而没有整合。
在涉及免疫应用的一些优选实施方案中,递送核酸分子,所述核酸分子包含编码本发明的蛋白质的核苷酸序列,并且另外包含进一步增强针对这类靶蛋白的免疫应答的蛋白质的基因。这类基因的实例是:编码其它细胞因子和淋巴因子的那些基因,如α-干扰素、γ-干扰素、血小板源生长因子(pdgf)、tnfα、tnfβ、gm-csf、上皮生长因子(egf)、il-1、il-2、il-4、il-5、il-6、il-10、il-12、il-18、mhc、cd80、cd86和il-15(包括信号序列缺失并且任选地包含来自ige的信号肽的il-15)。可能有用的其它基因包括编码下列的那些基因:mcp-1、mip-lα、mip-1p、il-8、rantes、l-选择蛋白、p-选择蛋白、e-选择蛋白、cd34、glycam-1、madcam-1、lfa-1、vla-1、mac-1、pl50.95、pecam、icam-1、icam-2、icam-3、cd2、lfa-3、m-csf、g-csf、il-4、il-18的突变形式、cd40、cd40l、血管生长因子、il-7、神经生长因子、血管内皮生长因子、fas、tnf受体、flt、apo-1、p55、wsl-1、dr3、tramp、apo-3、air、lard、ngrf、dr4、dr5、killer、trail-r2、trick2、dr6、半胱天冬酶ice、fos、c-jun、sp-1、ap-1、ap-2、p38、p65rel、myd88、irak、traf6、ikb、无活性nik、sapk、sap-1、jnk、干扰素应答基因、nfkb、bax、trail、trailrec、trailrecdrc5、trail-r3、trail-r4、rank、rankligand、ox40、ox40ligand、nkg2d、mica、micb、nkg2a、nkg2b、nkg2c、nkg2e、nkg2f、tap1、tap2及其功能片段。
可添加另外的元件,如果出于任何原因希望消除接受基因构建体的细胞,则所述元件充当用于细胞破坏的靶。可表达形式的疱疹病毒胸苷激酶(tk)基因可包含在基因构建体中。药物更昔洛韦可施用至个体并且所述药物将引起任何产生tk的细胞的选择性杀死,由此提供选择性破坏具有基因构建体的细胞的方式。
为了最大化蛋白质制备,可选择调控序列,所述调控序列非常适用于构建体所施用的细胞中的基因表达。而且,可选择在细胞中最高效地转录的密码子。本领域技术人员可制备在细胞中具有功能的dna构建体。
在一些实施方案中,可提供基因构建体,其中本文所述的蛋白质的编码序列连接至ige信号肽。在一些实施方案中,本文所述的蛋白质连接至ige信号肽。
在使用蛋白质的一些实施方案中,例如使用熟知的技术,本领域技术人员可使用熟知的技术来制备和分离本发明的蛋白质。在使用蛋白质的一些实施方案中,例如,本领域技术人员可使用熟知的技术来将编码本发明的蛋白质的dna分子插入在熟知表达系统中使用的市售表达载体中。例如,市售质粒pse420(invitrogen,sandiego,calif.)可用于在大肠杆菌中制造蛋白质。市售质粒pyes2(invitrogen,sandiego,calif.)可例如用于在酵母的酿酒酵母菌株中的制备。市售maxbactm完整杆状病毒表达系统(invitrogen,sandiego,calif.)可例如用于在昆虫细胞中的制备。市售质粒pcdnai或pcdna3(invitrogen,sandiego,calif.)可例如用于在哺乳动物细胞如中国仓鼠卵巢细胞中的制备。本领域技术人员可使用这些商业表达载体和系统或其它载体和系统来通过常规技术和可容易获得的起始材料制备蛋白质。(例如参见sambrook等,molecularcloningalaboratorymanual,第二版coldspringharborpress(1989),其以引用的方式并入本文)。因此,所需蛋白质可在原核和真核系统中制备,从而产生一系列加工形式的蛋白质。
本领域技术人员可使用其它市售表达载体和系统或使用熟知方法和可容易获得的起始材料来制备载体。含有需要的控制序列(如启动子和聚腺苷酸化信号、并且优选增强子)的表达系统对于多种宿主来说是容易获得的和本领域已知的。例如参见sambrook等,molecularcloningalaboratorymanual,第二版coldspringharborpress(1989)。基因构建体包括可操作地连接至启动子的蛋白质编码序列,所述启动子在构建体所转染的细胞系中具有功能。组成型启动子的实例包括来自巨细胞病毒或sv40的启动子。可诱导启动子的实例包括小鼠乳腺白血病病毒或金属硫蛋白启动子。本领域技术人员可由可容易获得的起始材料容易地制备适用于使用编码本发明的蛋白质的dna转染细胞的基因构建体。使用包含编码蛋白质的dna的表达载体来转化相容性宿主,然后培养所述宿主并保持在其中发生外源dna表达的条件下。
制备的蛋白质通过使细胞裂解来从培养物中回收或者从对于本领域技术人员来说适当和已知的培养基中回收。本领域技术人员可使用熟知技术分离使用这类表达系统制备的蛋白质。使用如上所述特异性结合特定的蛋白质的抗体从天然来源纯化蛋白质的方法可同样应用于通过重组dna方法制备的蛋白质的纯化。
除了通过重组技术来制备蛋白质,还可以使用自动肽合成器来制备分离的基本上纯的蛋白质。这类技术是本领域技术人员熟知的,并且适用于具有dna-编码的蛋白质制备中未提供的取代的衍生物。
核酸分子可使用若干种熟知的技术中的任一种来递送,所述技术包括dna注射(也称为dna疫苗接种)、重组载体,如重组腺病毒、重组腺病毒相关病毒和重组牛痘。
施用途径包括但不限于肌内、鼻内、腹膜内、皮内、皮下、静脉内、动脉内、眼内和口服以及局部、透皮、通过吸入或栓剂施用、或对粘膜组织施用,如通过灌洗阴道、直肠、尿道、口腔和舌下组织施用。优选的施用途径包括肌内、腹膜内、皮内和皮下注射。基因构建体可通过这样的方式来施用,所述方式包括但不限于电穿孔方法和装置、传统注射器、无针注射装置或"微弹轰击基因枪法"。
优选用于促进dna疫苗的递送的电穿孔装置和电穿孔方法的实例包括下列专利中描述者:美国专利号7,245,963(draghia-akli等)、美国专利公布2005/0052630(由smith等递交),其全部内容以引用的方式并入本文。还优选的是,于共同待决和共同拥有的美国专利申请序列号11/874072中提供的用于促进dna疫苗的递送的电穿孔装置和电穿孔方法,所述美国专利申请提交于2007年10月17日,并且在35usc119(e)下要求下列申请的优先权:2006年10月17日提交的美国临时申请序列号60/852,149和2007年10月10日提交的美国临时申请序列号60/978,982,其均以引用的方式整体并入本文。
以下是使用电穿孔技术的实施方案的实例,并且在上述专利参考文献中更详细地讨论:电穿孔装置可被配置为递送产生类似于用户输入的预设电流的恒定电流的能量脉冲至哺乳动物的期望组织。电穿孔装置包括电穿孔部件和电极组件或手柄组件。电穿孔部件可包括且并有电穿孔装置的多种元件中的一种或多种,包括:控制器、电流波形产生器、阻抗试验器、波形记录仪、输入元件、状态报告元件、通信端口、记忆部件、电源和电源开关。电穿孔部件可充当电穿孔装置的一个元件,并且其它元件是与所述电穿孔部件通信的单独元件(或部件)。在一些实施方案中,电穿孔部件可充当电穿孔装置的多于一个元件,其可与电穿孔装置的独立于所述电穿孔部件的其它元件通信。使用电穿孔技术递送改进的hpv疫苗不受到现存作为一个机电或机械装置的部分的电穿孔装置的元件的限制,因为所述元件可充当与彼此通信的一个装置或单独元件。电穿孔部件能够递送在期望组织中产生恒定电流的能量脉冲,并且包括反馈机制。电极组件包括在空间布置中具有多个电极的电极阵列,其中电极组件从电穿孔部件接收能量脉冲,并且通过电极将所述能量脉冲递送至期望组织。多个电极中的至少一个在能量脉冲的过程中是中性的,并且测量期望组织中的电阻和将电阻通知电穿孔部件。反馈机制可接收所测量到的电阻,并且可调整由电穿孔部件递送的能量脉冲以保持恒定电流。
在一些实施方案中,多个电极可以分散型式来递送能量脉冲。在一些实施方案中,多个电极可以分散型式通过控制电极在编程序列下递送能量脉冲,并且编程序列由用户输入至电穿孔部件。在一些实施方案中,编程序列包含依序递送的多个脉冲,其中多个脉冲中的各脉冲通过至少两个有效电极(其中有一个中性电极测量电阻)来递送,并且其中多个脉冲的后续脉冲通过至少两个有效电极(其中有一个中性电极测量电阻)中的不同的那个来递送。
在一些实施方案中,反馈机制通过硬件或软件来执行。优选地,反馈机制通过模拟闭环电路来执行。优选地,该反馈每隔50μs、20μs、10μs或1μs发生,但优选是实时反馈或瞬时的(即,如通过用于测定应答时间的可用技术所测定,其基本上是瞬时的)。在一些实施方案中,中性电极测量期望组织中的电阻并将电阻通知反馈机制,并且反馈机制回应于电阻并调整能量脉冲以使恒定电流维持在类似于预设电流的数值下。在一些实施方案中,反馈机制在能量脉冲递送的过程中连续和瞬时地维持恒定电流。
在一些实施方案中,核酸分子是结合多核苷酸功能增强子或基因疫苗促进剂的施用而递送至细胞。多核苷酸功能增强子描述于美国序列号5,593,972、5,962,428和1994年1月26日提交的国际申请序列号pct/us94/00899,其均以引用的方式并入本文。基因疫苗促进剂描述于1994年4月1日提交的美国序列号021,579中,其以引用的方式并入本文。结合核酸分子施用的辅剂可作为与核酸分子的混合物来施用,或者在核酸分子施用的同时、之前或之后单独施用。另外,可用作转染剂和/或复制剂和/或炎症剂并且可与gvf共同施用的其它药剂包括:生长因子、细胞因子和淋巴因子,如a-干扰素、γ-干扰素、gm-csf、血小板源生长因子(pdgf)、tnf、表皮生长因子(egf)、il-1、il-2、il-4、il-6、il-10、il-12和il-15以及成纤维细胞生长因子、表面活性剂如免疫刺激性复合物(iscoms)、弗氏不完全佐剂、lps类似物(包括单磷酰基脂质a(wl))、胞壁肽、醌类似物和囊泡如三十碳六烯和角鲨烯,并且透明质酸也可与基因构建体结合而使用施用。在一些实施方案中,免疫调节蛋白可用作gvf。在一些实施方案中,联合提供核酸分子与plg以增强递送/摄取。
根据本发明的药物组合物包含约1纳克至约2000微克dna。在一些优选实施方案中,根据本发明的药物组合物包含约5纳克至约1000微克dna。在一些优选实施方案中,药物组合物含有约10纳克至约800微克dna。在一些优选实施方案中,药物组合物含有约0.1至约500微克dna。在一些优选实施方案中,药物组合物含有约1至约350微克dna。在一些优选实施方案中,药物组合物含有约25至约250微克dna。在一些优选实施方案中,药物组合物含有约100至约200微克dna。
根据本发明的药物组合物根据所使用的施用方式来进行配制。在药物组合物是可注射的药物组合物的情况下,它们是无菌、无热原质和无颗粒的。优选使用等渗制剂。通常,用于等渗性的添加剂可包括氯化钠、右旋糖、甘露糖醇、山梨糖醇和乳糖。在一些情况下,等渗溶液如磷酸盐缓冲盐水是优选的。稳定剂包括明胶和白蛋白。在一些实施方案中,将血管收缩剂添加至制剂中。
根据本发明的一些实施方案,提供诱导免疫应答的方法。疫苗可以是基于蛋白质的、活减毒疫苗、细胞疫苗、重组疫苗或核酸或dna疫苗。在一些实施方案中,在个体中诱导针对免疫原的免疫应答的方法(包括诱导粘膜免疫应答的方法)包括向个体施用下列中的一种或多种:ctack蛋白、teck蛋白、mec蛋白及其功能片段、或其可表达的编码序列,与编码本发明的蛋白质的分离的核酸分子、和/或编码本发明的蛋白质的重组疫苗、和/或包含本发明的蛋白质的亚单位疫苗、和/或活减毒疫苗和/或灭活疫苗的组合。ctack蛋白、teck蛋白、mec蛋白及其功能片段中的一种或多种可在下列物质施用之前、同时或之后施用:编码免疫原的分离的核酸分子、和/或编码免疫原的重组疫苗、和/或包含免疫原的亚单位疫苗、和/或活减毒疫苗和/或灭活疫苗。在一些实施方案中,向个体施用编码选自由以下各项组成的组的一种或多种蛋白质的分离的核酸分子:ctack、teck、mec及其功能片段。
在下列实施例中进一步说明了本发明。应理解,此实施例尽管指出了本发明的实施方案,但是其仅以说明的方式给出。根据上述讨论和此实施例,本领域技术人员可确定本发明的实质性特性,并且在不偏离本发明的精神和范围的情况下,可对本发明进行各种改变和修改以使其适应各种用途和条件。因此,除了本文中示出和描述的修改之外,根据先前的描述,本发明的各种修改对于本领域技术人员来说将是显而易见的。这类修改也意在属于所附权利要求书的范围内。
本公开通篇中引用的美国专利、美国申请和引用文献各自均以引用的方式整体并入本文。
实施例
本发明在下列实施例中进行进一步限定。应理解,这些实施例尽管指出本发明的优选实施方案,但是其仅以说明的方式给出。根据上述讨论和这些实施例,本领域技术人员可确定本发明的实质性特性,并且在不偏离本发明的精神和范围的情况下,可对本发明进行各种改变和修改,以使其适应各种用途和条件。因此,除了本文中示出和描述的那些修改之外,根据先前的描述,本发明的各种修改对于本领域技术人员来说将是显而易见的。这类修改也意在属于所附权利要求书的范围内。
实施例1
hpv6和hpv11e6/e7疫苗设计和表达
hpv6和11e6/e7共有序列基融合免疫原的构建
从基因库收集6或11型hpve6和e7基因序列,并且在进行多重比对之后获得共有e6和e7核苷酸序列。hpv6e6或e7蛋白质的共有序列分别从98或20个序列产生,而hpv11e6或e7蛋白质的共有序列分别从76或13个序列产生。在系统发生研究中应用的多重比对程序包括clustalx(版本2.0)的应用。如图1中的a和图1中的b所示,在其e6和e7蛋白质属于相同类型的hpv病毒株中存在约0-2%的序列分歧。然而,在hpv6和11之间,e6蛋白质中的基因距离高达19.3%,而e7蛋白质中的基因距离高达16.3%。基于这些来自系统发生分析的结果,我们开发出两种类型特异性的e6/e7共有dna疫苗。
在产生共有e6/e7融合序列之后进行若干修饰(图1中的c)。将高效的前导序列同框融合在起始密码子上游以促进表达。还如先前所述进行密码子和rna优化(j.yan等,cellularimmunityinducedbyanovelhpv18dnavaccineencodingane6/e7fusionconsensusproteininmiceandrhesusmacaques,vaccine.26(2008)5210-5215;和j.yan等,inductionofantitumorimmunityinvivofollowingdeliveryofanovelhpv-16dnavaccineencodingane6/e7fusionantigen,vaccine.27(2009)431-440)。内切蛋白水解裂解位点引入在e6和e7蛋白质之间以用于恰当蛋白折叠和更好的ctl加工。合成的工程化6e6e7基因和11e6e7基因的长度均为840bp。将序列验证的合成基因分别亚克隆至pvax表达载体的bamhi和xhoi位点用于进一步研究。通过翻译共有核苷酸序列获得共有氨基酸序列。
在获得hpv6和11共有e6和e7序列之后,如先前所述进行密码子优化和rna优化(j.yan等,vaccine.26(2008)5210-5215;和j.yan等,induction,vaccine.27(2009)431-440)。合成编码6或11型hpv共有e6/e7融合蛋白(6e6e7或11e6e7)的融合基因并进行序列验证。合成的6e6e7或11e6e7用bamhi和xhoi消化,克隆至表达载体pvax(invitrogen)中受巨细胞病毒即刻早期启动子控制,并且将这些构建体称为p6e6e7或p11e6e7。
293-t细胞在6孔板中培养,并且使用fugene6转染试剂(rocheappliedscience,indianapolis,in)用pvax、p6e6e7或p11e6e7转染。转染之后两天,使用改良的ripa细胞裂解缓冲液使细胞裂解并且收集细胞裂解物。使用抗-ha单克隆抗体进行蛋白质印迹分析(sigma-aldrich,st.louis,mo)并且使用ecltm蛋白质印迹分析系统(amersham,piscataway,nj)使用辣根过氧化物酶偶联的山羊抗-小鼠igg(sigma-aldrich,stlouis,mo)进行观测。
使用人横纹肌肉瘤细胞(rd细胞)进行间接免疫荧光测定以验证p6e6e7和p11e6e7的表达。在室玻片中培养的rd细胞使用turbofectin8.0(origene,rockville,md)用pvax、p6e6e7或p11e6e7转染。然后,细胞用pfa固定并用0.1%triton-x的pbs溶液进行渗透。用一级抗体和二级抗体对细胞进行1-2小时的孵育,另外在孵育之间用补充有甘氨酸和bsa的pbs洗涤。所使用的一级抗体和二级抗体分别是单克隆小鼠抗-ha(sigma-aldrich,st.louis,mo)和fitc-偶联的抗小鼠igg(abcam,cambridge,ma)。还进行hoechst染色以鉴定细胞核并定位rd细胞。在完成全部孵育之后,将细胞置于装有fluoromount-g(southernbiotech,birmingham,al)的载玻片上。样品用共聚焦显微镜(cdbmicroscopycore,universityofpennsylvaniacellanddevelopmentalbiology,philadelphia,pa)观察并成像。
实施例2
小鼠和处理组疫苗接种
在此实验中使用6至8周龄之间的c57bl/6雌性小鼠。小鼠是从杰克逊实验室(barharbor,me)获得。小鼠由宾夕法尼亚大学(philadelphia,pa)的universitylaboratoryanimalresources在遵守国立卫生研究院和宾夕法尼亚大学实验动物管理和使用委员会(iacuc)的政策下圈养和维持。将用于这些实验的小鼠分成四组用于免疫。小鼠用p6e6e7、p11e6e7或这两种构建体进行免疫并且pvax组用作阴性对照。
dna疫苗接种和电穿孔
每个小鼠以14天的间隔接受三个剂量的20μg各dna质粒。接受p6e6e7和p11e6e7两者的组中的小鼠接受20μg的两种质粒,每次疫苗接种共计40μgdna。dna构建体经由肌内注射右四头肌来施用,随后通过cellectratm电穿孔仪(inoviopharmaceuticals,bluebell,pa)产生方波脉冲。电穿孔仪被配置用来在0.2amps下以52ms/脉冲、间隔1秒延迟递送两个电脉冲。如先前所述进行电穿孔程序[12、13]。
ifn-γelispot测定
处理组和对照组的小鼠都在第三次免疫1周后处死。从每个小鼠收集脾脏并将其转移至具有10%fbs和抗生素的rpmi-1640培养基(r10)中。使用均质袋(stomacher)(sewardlaboratorysystems,bohemia,ny)将脾脏粉碎并随后转移通过细胞滤网并悬浮在ack裂解缓冲液中。在去除红细胞之后,分离脾细胞并悬浮于r10培养基中。高蛋白ip96孔multiscreentm板(millipore,bedford,ma)用单克隆小鼠ifn-γ捕捉抗体(r&dsystems,minneapolis,mn)预涂覆并在4℃下孵育过夜。在用1xpbs洗涤三次之后,在环境温度下将板用1%bsa和5%蔗糖的1×pbs溶液阻断2小时。对r10培养基中的分离的脾细胞进行计数并且以每孔2×105个细胞一式三份地添加至孔中。涵盖hpv6和hpv11的共有e6/e7序列的两组肽在dmso(genscriptusa,piscataway,nj)中复原。所述肽包含15个氨基酸的序列,其中8个残基与每个连续肽重叠。将hpv6和hpv11肽以在dmso中2μg/ml的浓度各自分成两个池—一个池是e6且另一个池是e7。保留用于阳性对照和阴性对照的孔分别接收刀豆素a(concavalina)(sigma-aldrich,st.louis,mo)和r10培养基代替肽。随后将板放置在5%co2氛围孵育器中。在37℃下孵育24小时之后,用1×pbs洗涤孔。将生物素化的抗小鼠ifn-γ检测抗体(r&dsystems,minneapolis,mn)添加至每个孔中并且接着在4℃下孵育过夜。随后洗涤板并按照r&dsystems提供的显色方案使用抗生蛋白链菌素-ap和bcip/nbtplus(r&dsystems,minneapolis,mn)进行处理。将孔风干过夜,并且通过装有
考虑到ifn-γelispot测定的灵敏度和说明t细胞活性的能力,使用它们来测定响应于hpv6或11e6和e7肽刺激的抗原特异性ifn-γ分泌细胞的数目。
如图3中的a和图3中的b所示,接种p6e6e7的小鼠的sfu/106脾细胞的平均数是1442.8,而接种p11e6e7的小鼠的sfu/106的平均数是2845,这都显著大于阴性对照组。因此,p6e6e7和p11e6e7都对在小鼠中引发稳固的类型特异性e6和e7特异性免疫应答有效。
有趣的是,接种p6e6e7或p11e6e7也诱导交叉反应性细胞免疫应答。在p11e6e7免疫的小鼠中针对hpv6e6/e7肽的sfu/106个脾细胞的加和频率是552.8sfu/106个脾细胞,而p6e6e7免疫的小鼠中的hpv11e6/e7特异性免疫应答是888.3sfu/106个脾细胞。hpv6或11的e6蛋白共享约80%的同一性,而hpv6或11的e7蛋白共享约84%的同一性。在hpv6或11e6和e7抗原之间可能存在一些共享的免疫表位。从ifn-γelispot测定中观察到的交叉反应性表明在hpv6和11e6和e7抗原之间存在共享的免疫表位。
也使来自接受p6e6e7和p11e6e7两者的小鼠(组合组)的脾细胞经受以上elispot测定,以便检验当一起接种这两种构建体时是否存在任何免疫干扰(图3中的a和图3中的b)。组合组针对hpv6e6和e7肽展现出平均1670sfu/106个脾细胞(σ=55.7,p<0.001)。同一组的脾细胞针对hpv11e6和e7肽产生2010sfu/106个脾细胞(σ=247.8,p=0.002)。数据表明并行接种所述两种构建体可针对hpv6和hpv11引发统计上显著的e6和e7特异性细胞应答,并且这些应答互不干扰。
表位作图
进行表位作图研究以确定肽池内的优势表位,以便确定e6/e7共有抗原内的免疫优势肽(图4中的a和图4中的b)。所述研究与先前提及的ifn-γelispot测定类似进行。使用个别的肽而非池来刺激脾细胞。
用于此单肽分析中的每个肽代表hpv6或hpv11的e6和e7抗原的部分重叠的片段。作图数据表明肽7(taeiysyaykqlkvl)seqidno:11是hpv6e6和e7免疫原的优势表位(图4中的a)。taeiysyaykqlkvlseqidno:11包含8、9、10-mer氨基酸表位,所述表位经nihbimas提供的hla-结合预测软件验证是h2-kb限制型表位。为了进一步描述hpv11e6和e7特异性t细胞免疫应答(图4中的b)。还用hpv11e6和e7的重叠片段进行了肽分析。肽作图显示hpv11e6和e7抗原的优势表位是肽7(taeiyayayknlkvv)seqidno:12和27(hcyeqledssedevd)seqidno:13。与hpv6表位作图测定中的肽7一样,bimashla-结合预测软件确定taeiyayayknlkvvseqidno:12是h2-kb限制型表位。另一个hla-结合肽数据库,由nihniaid提供的免疫表位数据库和分析资源(immuneepitopedatabaseandanalysisresource)确定hcyeqledssedevdseqidno:13是h2-kb限制型表位。通过此表位作图研究鉴定了三种免疫亚优势肽:第6号(fcknalttaeiysya)seqidno:14、第9号(lfrggypyaacaccl)seqidno:15以及第13号(yagyattveeetkqd)seqidno:16。
细胞内细胞因子染色
根据ifn-γelispot测定所描绘的高免疫应答,进行细胞内细胞因子染色测定以提供对p6e6e7和p11e6e7所诱导的细胞应答的更全面的概观。自疫苗接种小鼠组和未处理小鼠组分离脾细胞并在37℃下5%co2环境中用涵盖hpv6和hpv11的e6和e7区域的肽刺激4小时。通过将细胞分别放置在佛波醇12-肉豆蔻酸酯13-乙酸酯(pma)和r10细胞培养基中在测定中使用阳性和阴性对照。在孵育之后,细胞首先用vivid染料(invitrogen,carlsbad,ca)染色以区分活细胞和死细胞,接着所有的细胞用下列表面标记抗体染色:apc-cy7仓鼠抗小鼠cd3e、percp-cy5.5大鼠抗小鼠cd4和apc大鼠抗小鼠cd8a(bdbiosciences,sandiego,ca)。细胞随后使用cytofix/cytoperm试剂盒(bdbiosciences,sandiego,ca)固定。在按照制造商方案固定之后,细胞用下列细胞内标记抗体染色:alexafluor700大鼠抗小鼠ifn-γ、pe-cy7大鼠抗小鼠tnf克隆和pe大鼠抗小鼠il-2(bdbiosciences,sandiego,ca)。染色之后,细胞用含2%多聚甲醛的pbs溶液固定。制备的细胞使用装有bdfacsdiva软件的lsrii流式细胞仪(bdbiosciences,sanjose,ca)来获取。所获取的数据使用最新版本的flowjo软件(treestar,ashland,or)进行分析。使用下列门序列来分离cd4+和cd8+事件:来自fsc-a对fsc-h的单线态、来自fsc-a对ssc-a的所有脾细胞、来自vivid染料(pacificblue)对ssc-a的活细胞、来自cd3(apc-cy7)对ssc-a的cd3+细胞以及来自cd4(percp-cy5.5:阳性–cd4+,阴性–cd8+)对ssc-a的cd4+或cd8+。后两个群体分别针对alexafluor700、pe-cy7和pe进行门控以观察il-2、ifn-γ和tnf-α产生的变化。
对细胞进行门控以使得细胞内细胞因子染色数据可通过cd4+和cd8+细胞应答进行区分。当用hpv6e6和e7特异性肽刺激时,接种p6e6e7的小鼠分别展现出0.163%(σ=0.09)、0.003%(σ=0.07)和0.188%(σ=0.20)的产生ifn-γ、tnf-α和il-2的总cd4+细胞的平均值(图5a)。同一组的小鼠分别具有3.323%(σ=1.39)、0.838%(σ=0.32)和1.172%(σ=1.81)的产生ifn-γ、tnf-α和il-2的总cd8+细胞的平均值。在用hpv11e6和e7抗原孵育之后使用接种p11e6e7的小鼠的脾细胞收集相同的细胞内细胞因子数据(图5b)。在p11e6e7接种小鼠的所有cd4+细胞当中,平均为0.051%(σ=0.04)产生ifn-γ、0.068%(σ=0.09)产生tnf-α,并且0.026%(σ=0.037)产生il-2。另外,在p11e6e7接种小鼠的所有cd8+细胞当中,产生ifn-γ、tnf-α和il-2的平均值分别为4.52%(σ=2.53)、2.08%(σ=1.56)和0.21%(σ=0.22)。除了少数组之外,处理组与它们相应未处理组相比,产生细胞因子的细胞的百分比具有统计学显著性,置信水平为89%或更高。观察了cd4+细胞与cd8+t细胞的细胞因子产生的量值的比较,可得到结论:由p6e6e7和p11e6e7引发的免疫应答高度趋向于驱动cd8+淋巴细胞,所述免疫应答在所有模型中与它们的细胞清除率相关联。
统计分析
进行学生t检验以分析此研究中所产生的所有定量数据的统计学显著性。除非另外指出,否则p值被计算来确定不同置信水平下的统计学显著性。
此研究已显示出有力的证据证明dna疫苗能够实现在使用其它流行且传统的疫苗平台的实验中发现的免疫原性水平。在其它类似的e6和e7特异性hpvdna疫苗研究中测量的高水平的细胞应答与表明预防性和治疗性抗瘤功效的数据相关联。大多数hpv相关性研究和疾病挑战模型聚焦于子宫颈癌。考虑到hpv6和hpv11不像其它hpv血清型那样与子宫颈癌相关,p6e6e7和p11e6e7的治疗功效不能完全使用常规hpv疾病挑战模型来评估。可预见在可获得适当的疾病挑战模型时确定p6e6e7和p11e6e7的保护和治疗潜力。因此,人们只能够通过观察由elispot测定和细胞内细胞因子染色定量的ifn-γ和细胞因子产生的高水平来推断p6e6e7和p11e6e7的免疫原性功效。然而,发现这类水平的量值明显较高并且显示出两种构建体将引发高水平的细胞免疫应答的前景。由于hpv相关性恶性肿瘤通常继发于多种血清型的并发感染,所以急需进一步研究组合靶向不同病毒血清型的疫苗的可行性。保证研究t细胞动力学和疫苗竞争的未来研究进一步表征两种质粒的并行疫苗接种的效果。
细胞内细胞因子染色显示接种p6e6e7和p11e6e7能够引发显著百分比的产生ifn-γ、tnf-α和il-2的t细胞。鉴于ifn-γ当前已知的在免疫系统中的功能,在历史上ifn-γ已被用作细胞免疫应答的指标。这些重要作用中的一些包括调节和刺激先天性和适应性免疫的能力。此外,广泛公认原则上ifn-γ的产生者是t细胞,使得ifn-γ产生成为测量给定疫苗抗原暴露后的细胞免疫原性的可接受的模式。tnf-α是免疫系统的调控中所涉及的另一种细胞因子。tnf-α已知的诱导细胞凋亡和调控肿瘤增殖的能力使其成为在表征疫苗接种后免疫应答时重要的考虑参数。考虑到hpv6和hpv11的潜在肿瘤增殖特性,tnf-α产生可能更受到关注。il-2是另一种信号传导分子,已观察到它在t细胞在免疫系统中的增殖和分化中起到中枢作用。结果是,il-2经常与其它细胞因子一起检查以获得对特定免疫应答的量值和品质的进一步的总体认识。鉴于上文,接种p6e6e7之后产生ifn-γ、tnf-α和il-2的cd4+和cd8+细胞的显著百分比表明疫苗成功诱导有效的免疫应答。除分泌il-2的cd4+细胞之外,相同趋势适用于从接种p11e6e7的小鼠中分离的细胞。此外,值得注意的是,观察到cd8+细胞在疫苗接种小鼠中驱动强烈的免疫应答,这是在评估两种质粒的抗瘤功效中重要的特征。
序列表
<110>宾夕法尼亚大学理事会(thetrusteesoftheuniversityofpennsylvania)
大卫·b·韦纳
严健
<120>用于人乳头状瘤病毒的疫苗及其使用方法
<130>upvg0025wo2
<160>16
<170>patentin版本3.5
<210>1
<211>873
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>cds
<223>hpv6e6e7dna序列
<400>1
ggtaccggatccgccaccatggactggacctggattctgttcctggtggccgctgccaca60
cgggtgcacagcgaaagcgccaatgccagcaccagcgctacaaccatcgaccagctgtgc120
aagaccttcaacctgagcatgcacaccctgcagatcaactgcgtgttctgcaagaacgcc180
ctgaccaccgccgagatctacagctacgcctacaagcagctgaaggtgctgttcagaggc240
ggctacccctatgccgcctgcgcctgctgcctggaatttcacggcaagatcaaccagtac300
cggcacttcgactacgccggctacgccaccaccgtggaagaggaaacaaagcaggacatc360
ctggacgtgctgatccggtgctacctgtgccacaagcccctgtgcgaggtggaaaaagtg420
aagcacatcctgaccaaggcccggttcatcaagctgaactgcacctggaagggccggtgc480
ctgcactgctggaccacctgtatggaagatatgctgcccagaggccggaagcggcggagc540
catggcagacacgtgaccctgaaggacatcgtgctggacctgcagccccccgatcctgtg600
ggcctgcactgttacgagcagctggtggacagcagcgaggacgaggtggacgaagtggac660
ggccaggacagccagcccctgaagcagcacttccagatcgtgacctgctgctgcggctgc720
gacagcaacgtgcggctggtggtgcagtgcaccgagacagacatcagagaggtccagcag780
ctcctgctgggcaccctgaacatcgtgtgccccatctgcgcccccaagacctacccttac840
gacgtgcccgactacgcctgatgactcgagctc873
<210>2
<211>280
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv6e6e7蛋白质序列
<400>2
metasptrpthrtrpileleupheleuvalalaalaalathrargval
151015
hissergluseralaasnalaserthrseralathrthrileaspgln
202530
leucyslysthrpheasnleusermethisthrleuglnileasncys
354045
valphecyslysasnalaleuthrthralagluiletyrsertyrala
505560
tyrlysglnleulysvalleupheargglyglytyrprotyralaala
65707580
cysalacyscysleugluphehisglylysileasnglntyrarghis
859095
pheasptyralaglytyralathrthrvalgluglugluthrlysgln
100105110
aspileleuaspvalleuileargcystyrleucyshislysproleu
115120125
cysgluvalglulysvallyshisileleuthrlysalaargpheile
130135140
lysleuasncysthrtrplysglyargcysleuhiscystrpthrthr
145150155160
cysmetgluaspmetleuproargglyarglysargargserhisgly
165170175
arghisvalthrleulysaspilevalleuaspleuglnproproasp
180185190
provalglyleuhiscystyrgluglnleuvalaspsersergluasp
195200205
gluvalaspgluvalaspglyglnaspserglnproleulysglnhis
210215220
pheglnilevalthrcyscyscysglycysaspserasnvalargleu
225230235240
valvalglncysthrgluthraspilearggluvalglnglnleuleu
245250255
leuglythrleuasnilevalcysproilecysalaprolysthrtyr
260265270
protyraspvalproasptyrala
275280
<210>3
<211>873
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>cds
<223>hpv11e6e7dna序列
<400>3
ggtaccggatccgccaccatggactggacctggattctgttcctggtggccgctgccaca60
agagtgcacagcgagagcaaggatgccagcaccagcgccaccagcatcgaccagctgtgc120
aagaccttcaacctgagcctgcacaccctgcagatccagtgcgtgttctgccggaacgcc180
ctgaccaccgccgagatctacgcctacgcctacaagaacctgaaggtcgtgtggcgggac240
aacttccccttcgccgcctgcgcctgctgcctggaactgcagggcaagatcaaccagtac300
cggcacttcaactacgccgcttacgcccccaccgtggaagaggaaacaaacgaggacatc360
ctgaaggtgctgatccggtgctacctgtgccacaagcccctgtgcgagatcgagaagctg420
aagcacatcctgggcaaggcccggttcatcaagctgaacaaccagtggaagggccggtgc480
ctgcactgctggaccacctgtatggaagatctgctgcccagaggccggaagcggagaagc540
cacggcagactggtcaccctgaaggacatcgtgctggacctgcagccccccgatcctgtg600
ggcctgcactgttacgagcagctggaagatagcagcgaggacgaggtggacaaagtggac660
aagcaggacgcccagcccctgacccagcactaccagatcctgacctgctgctgcggctgc720
gacagcaacgtgcggctggtggtggaatgcaccgacggcgacatccggcagctccaggat780
ctgctgctgggcaccctgaacatcgtgtgccccatctgcgcccccaagccctacccctac840
gacgtgcccgactacgcctgatgactcgagctc873
<210>4
<211>280
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv11e6e7蛋白质序列
<400>4
metasptrpthrtrpileleupheleuvalalaalaalathrargval
151015
hissergluserlysaspalaserthrseralathrserileaspgln
202530
leucyslysthrpheasnleuserleuhisthrleuglnileglncys
354045
valphecysargasnalaleuthrthralagluiletyralatyrala
505560
tyrlysasnleulysvalvaltrpargaspasnpheprophealaala
65707580
cysalacyscysleugluleuglnglylysileasnglntyrarghis
859095
pheasntyralaalatyralaprothrvalgluglugluthrasnglu
100105110
aspileleulysvalleuileargcystyrleucyshislysproleu
115120125
cysgluileglulysleulyshisileleuglylysalaargpheile
130135140
lysleuasnasnglntrplysglyargcysleuhiscystrpthrthr
145150155160
cysmetgluaspleuleuproargglyarglysargargserhisgly
165170175
argleuvalthrleulysaspilevalleuaspleuglnproproasp
180185190
provalglyleuhiscystyrgluglnleugluaspsersergluasp
195200205
gluvalasplysvalasplysglnaspalaglnproleuthrglnhis
210215220
tyrglnileleuthrcyscyscysglycysaspserasnvalargleu
225230235240
valvalglucysthraspglyaspileargglnleuglnaspleuleu
245250255
leuglythrleuasnilevalcysproilecysalaprolysprotyr
260265270
protyraspvalproasptyrala
275280
<210>5
<211>828
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>cds
<223>hpv33e6e7dna序列
<400>5
ggtaccgtcgacggatccgccaccatggactggacctggattctgttcctggtggccgct60
gctacccgggtgcacagcttccaggacaccgaggaaagcggccggaccctgcacgatctg120
tgccaggccctggaaaccaccatccacaacatcgagctgcagtgcgtggaatgcaagaag180
cccctgcagcggagcgaggtgtacgacgccgacctgaccgtggtgtacagagagggcaac240
cccttcggcatctgcaagctgtgcctgcggttcctgagcaagatcagcgagtaccggcac300
tacaactacagcgtgtacggcaacaccctggaacagaccgtgaagaagcctctgaacgag360
atcctgatccggtgcatcatctgccagcggcccctgaagcggcacgtggacctgaacaag420
cggttccacaatatcagcggcagatgggccggcagatgcgccgcctgttggagaagccgg480
cggagagagacagccctgcggggcagaaagcggcggagcagaggccacaagcccaccctg540
aaagaatacgtgctggacctgtaccccgagcccaccgacctgtacggctacggccagctg600
agcgacagcagcgacgaggacgagggcctggacagacctgatggacaggcccagcccgcc660
accgccgactactacatcgtgacctgctgccacacctgtaacaccaccgtgcggctgtgc720
gtgaacagcaccgccagcgacctgcggacaatccagcagctcctgatgggcaccgtgaac780
atcgtgtgccctacctgcgcccagctgtgactcgagacgcgtgagctc828
<210>6
<211>261
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv33e6e7蛋白质序列
<400>6
metasptrpthrtrpileleupheleuvalalaalaalathrargval
151015
hisserpheglnaspthrglugluserglyargthrleuhisaspleu
202530
cysglnalaleugluthrthrilehisasnilegluleuglncysval
354045
glucyslyslysproleuglnargsergluvaltyraspalaaspleu
505560
thrvalvaltyrarggluglyasnpropheglyilecyslysleucys
65707580
leuargpheleuserlysileserglutyrarghistyrasntyrser
859095
valtyrglyasnthrleugluglnthrvallyslysproleuasnglu
100105110
ileleuileargcysileilecysglnargproleulysarghisval
115120125
aspleuasnlysargphehisasnileserglyargtrpalaglyarg
130135140
cysalaalacystrpargserargargarggluthralaleuarggly
145150155160
arglysargargserargglyhislysprothrleulysglutyrval
165170175
leuaspleutyrprogluprothraspleutyrglytyrglyglnleu
180185190
seraspserseraspgluaspgluglyleuaspargproaspglygln
195200205
alaglnproalathralaasptyrtyrilevalthrcyscyshisthr
210215220
cysasnthrthrvalargleucysvalasnserthralaseraspleu
225230235240
argthrileglnglnleuleumetglythrvalasnilevalcyspro
245250255
thrcysalaglnleu
260
<210>7
<211>835
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>cds
<223>hpv58e6e7dna序列
<400>7
ggtaccgtttaaacggatccgccaccatggactggacctggattctgttcctggtggccg60
ctgctacccgggtgcacagcttccaggatgccgaggaaagcggccggaccctgcacgatc120
tgtgccaggccctggaaaccagcgtgcacgagatcgagctgaagtgcgtggaatgcaaga180
aaaccctgcagcggagcgaggtgtacgacgccgacctgcggatcgtgtaccgggacggca240
accccttcgccgtgtgcaaagtgtgcctgcggctgctgagcaagatcagcgagtaccggc300
actacaactacagcctgtacggcgacaccctggaacagaccctgaagaagtgcctgaacg360
agatcctgatccggtgcatcatctgccagcggcccctgaagcggcacgtggacctgaaca420
agcggttccacaacatcagcggccggtggaccggcagatgcgccgtgtgttggaggccca480
ggcggagacagacccaggtgcgcggcagaaagcggcggagcagaggcaacaaccccaccc540
tgagagagtacatcctggacctgcaccccgagcccaccgacctgtttggctacggccagc600
tgtgcgacagcagcgacgaggacgagatcggcctggacggccctgatggacaggcccagc660
ccgccaccgccaactactacatcgtgacctgctgctacacctgtggcaccaccgtgcggc720
tgtgcatcaacagcaccaccaccgacgtgcgcaccctgcagcagctcctgatgggcacct780
gtaccatcgtgtgccccagctgcgcccagcagtgactcgaggcggccgcgagctc835
<210>8
<211>262
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv58e6e7蛋白质序列
<400>8
metasptrpthrtrpileleupheleuvalalaalaalathrargval
151015
hisserpheglnaspalaglugluserglyargthrleuhisaspleu
202530
cysglnalaleugluthrservalhisgluilegluleulyscysval
354045
glucyslyslysthrleuglnargsergluvaltyraspalaaspleu
505560
argilevaltyrargaspglyasnprophealavalcyslysvalcys
65707580
leuargleuleuserlysileserglutyrarghistyrasntyrser
859095
leutyrglyaspthrleugluglnthrleulyslyscysleuasnglu
100105110
ileleuileargcysileilecysglnargproleulysarghisval
115120125
aspleuasnlysargphehisasnileserglyargtrpthrglyarg
130135140
cysalavalcystrpargproargargargglnthrglnvalarggly
145150155160
arglysargargserargglyasnasnprothrleuargglutyrile
165170175
leuaspleuhisprogluprothraspleupheglytyrglyglnleu
180185190
cysaspserseraspgluaspgluileglyleuaspglyproaspgly
195200205
glnalaglnproalathralaasntyrtyrilevalthrcyscystyr
210215220
thrcysglythrthrvalargleucysileasnserthrthrthrasp
225230235240
valargthrleuglnglnleuleumetglythrcysthrilevalcys
245250255
prosercysalaglngln
260
<210>9
<211>54
<212>dna
<213>人工序列(artificialsequence)
<220>
<221>cds
<223>ige前导dna序列
<400>9
atggactggacctggatcctgttcctggtggccgctgccacacgggtgcacagc54
<210>10
<211>18
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>ige前导蛋白质
<400>10
metasptrpthrtrpileleupheleuvalalaalaalathrargval
151015
hisser
<210>11
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv6肽7
<400>11
thralagluiletyrsertyralatyrlysglnleulysvalleu
151015
<210>12
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv11肽7
<400>12
thralagluiletyralatyralatyrlysasnleulysvalval
151015
<210>13
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv11肽27
<400>13
hiscystyrgluglnleugluaspsersergluaspgluvalasp
151015
<210>14
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv亚优势肽6
<400>14
phecyslysasnalaleuthrthralagluiletyrsertyrala
151015
<210>15
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv亚优势肽9
<400>15
leupheargglyglytyrprotyralaalacysalacyscysleu
151015
<210>16
<211>15
<212>prt
<213>人工序列(artificialsequence)
<220>
<221>chain
<223>hpv亚优势肽13
<400>16
tyralaglytyralathrthrvalgluglugluthrlysglnasp
151015
- 还没有人留言评论。精彩留言会获得点赞!