一种肽核酸介导的蛋白精准自组装方法与流程
本发明属于生物技术领域,具体涉及一种肽核酸介导的蛋白精准自组装方法。
背景技术:
肽核酸(peptidenucleicacid,pna)作为组装模板。肽核酸是由nielsen等人于1991年发明的类似于dna和rna的人工合成高分子聚合物(nielsen,p.e.;egholm,m.;berg,r.h.;buchardt,o.science1991,254,1497-1500)。它主要存在以下特点:(1)超强结合力。肽核酸可以跟dna(或rna)在watson-crick氢键以及hoogsteen氢键的驱动下形成很好的碱基配对相互作用。此外,由于pna/dna之间的碱基错配比dna/dna之间的更加不稳定,再加上pna电中性的特点无法与互补的dna链之间产生电荷排斥作用,因此pna与dna(或rna)之间的结合能力要强于dna(或rna)本身。(2)超强稳定性。由于人工合成的主链骨架结构无法被体内的各种蛋白水解酶以及和酸水解酶所识别,因此无论是pna本身,还是pna参与结合的任何双链结构都不会被微生物体内的酶所降解。
目前针对蛋白的自组装主要基于以下平台:(1)骨架蛋白。利用蛋白-蛋白间的相互作用,通过构建融合骨架蛋白的方法,使目标蛋白在骨架蛋白存在的情况下进行组装。但该方法存在相互作用较差、无法较好地控制蛋白间的比例以及骨架蛋白分子量太大等弊端;(2)纳米粒子。利用蛋白-纳米粒子间的相互作用,是目标蛋白在纳米粒子表面进行自组装。但是该方法存在无法精确控制蛋白间的比例、无法控制蛋白组装时的构型、潜在生物毒性等弊端;(3)dsdna分子。利用蛋白-dsdna间的相互作用,使目标蛋白在dsdna上形成自组装。该方法存在选择性低、无法控制组装比例、无法实现精确自组装等弊端。
技术实现要素:
本发明要解决的技术问题是:为了克服蛋白在自组装时无法精确控制、特异性差等问题,限制其在多酶体系、生物传感器、蛋白结构调控等生物领域中的普及使用。为解决上述问题,本发明提供一种肽核酸介导的蛋白精准自组装方法:
(1)制备含活性多肽标签的融合蛋白,
具体为,在蛋白的n端或c端融合了长度为21个天然氨基酸的多肽序列;
(2)制备含靶向pna序列的非天然多肽探针,
具体为,在n端或c端连有长度为10个碱基pna序列的非天然多肽(多肽长度为21个氨基酸);
(3)将步骤(1)中得到的含活性多肽标签的融合蛋白与步骤(2)中得到的含靶向pna序列的非天然多肽探针进行共价键结合,
这里的共价键结合具体是指的:融合蛋白上的半胱氨酸与多肽探针上的非天然x氨基酸间的共价键结合,常规多肽之间形成共价键时可能是识别某一位点,换言之标记位点是随机的,结合能力不强,而本专利中识别位点仅为融合蛋白上的21个氨基酸中固定的半胱氨酸,具有相当高的反应识别特异性;
(4)将步骤(3)共价键结合的产物在ssdna序列上进行可控自组装,
ssdna序列是指包含一段连接臂的长度为26个碱基的单链dna序列,其作为结合模板。
本发明提供的蛋白精准自组装的方式合成简单,组装精准,特异性高,生物兼容性好。通过多肽序列将目标蛋白上标记一段特异的pna序列,并通过pna与dna模板高效、精准的结合,促成了多个目标蛋白的精准组装,从而为多酶体系、生物传感器等生物技术领域提供了普适的组装平台。
附图说明
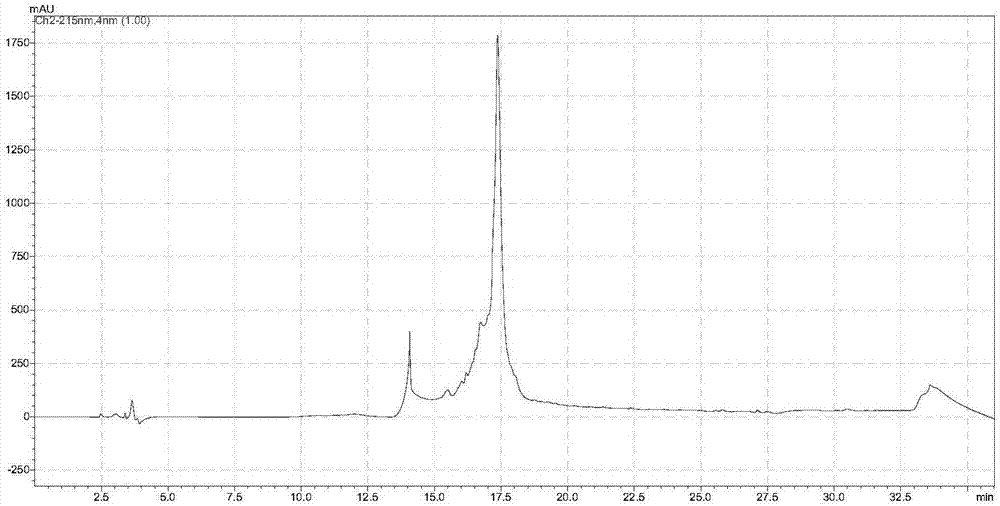
图1是含有pna序列的非天然多肽(pna-p1)的hplc表征图。
图2是含有pna序列的非天然多肽(pna-p2)的hplc表征图。
图3是含有活性多肽标签的融合蛋白(egfp-cce)与pna-p1以及pna-p2反应后的sds-page表征图。其中第一泳道为蛋白marker,第二泳道为天然的egfp-cce,第三泳道为egfp-cce与pna-p1共价结合后的复合物egfp-pna1,第四泳道为egfp-cce与pna-p2共价结合后的复合物egfp-pna2。
图4是含有活性多肽标签的融合蛋白(mcherry-cce与pna-p2)反应后的sds-page表征图。其中第一泳道为蛋白marker,第二泳道为天然的mcherry-cce,第三泳道为cherry-cce与pna-p2共价结合后的复合物mcherry-pna2。
图5是egfp-pna1与ssdna模板在不同化学计量数下的自组装效果图。随着蛋白量/模板量的增加,产物转化为两个egfp-p1组装后的复合物。
图6是egpf-pna1与ssdna模板在不同化学计量数下自组装的机理图。随着蛋白量/模板量的增加,预测了不同产物的存在形式。
图7是egfp-pna1和mcherry-pna2在ssdna模板的作用下形成1:1精准自组装的效果图。
图8是本专利中,肽核酸介导的蛋白精准自组装的结合机理,其中(1)含活性多肽标签的融合蛋白、(2)含靶向pna序列的非天然多肽探针、(3)融合蛋白上的活性多肽标签与非天然多肽的共价键结合、(4)ssdna序列。
具体实施方式
实施例1
载体构建:
(1)将两段单链dna序列
5’-caaatctgaagagtc-ttatgaatgtgctgccttagagaaggaagttgcagcgttagagaaggaagttgctgcattagagaagtaga-3’和
5’-agcttctacttctctaatgcagcaacttccttc-tctaacgctgcaacttccttctctaaggcagcacattcataagactcttcagatttgagct-3’进行褪火,并使用pcr纯化试剂盒进行产物纯化;
(2)将pet28m-egfp与pet28m-mcherry质粒使用saci和hindiii进行双酶切,并利用dna胶回收试剂盒对电泳产物进行回收;
(3)将上述(1)和(2)的产物混合,在t4dna连接酶作用下进行酶连反应;
(4)将(3)的产物进行转化、克隆筛选及测序获得质粒pet28m-egfp-cce及pet28m-mcherry-cce。
蛋白表达及纯化:
(1)将上述构建的质粒pet28m-egfp-cce及pet28m-mcherry-cce转化到大肠杆菌bl21感受态细胞中,并挑单克隆至5mllb培养基中过夜培养;
(2)将(1)中产物转移至500mllb溶液中并在37度培养,待od600到达0.6-0.8时使用iptg进行诱导,随后16度培养过夜;
(3)将(2)中产物离心收集,并使用超声破壁,再次离心后将蛋白上清液与ni-nta纯化柱孵育,并使用咪唑溶液进行梯度洗脱;
(4)将(3)中产物进行sds-page分析,收集目标蛋白并进行溶液置换,得到产物egfp-cce以及mcherry-cce(如附图3,4)。
实施例2
本发明所述的方法采用常规的固相fmoc法,即固相树脂上被fmoc保护的单体氨基酸(或肽核酸)去保护后露出氨基,通过缩合反应与溶液中氨基酸(或肽核酸)的羧基形成肽键,从而将氨基酸(或肽核酸)连接到树脂上,使肽链从c端向n端延伸:
树脂与连接分子:固相fmoc法选择的树脂为rink
单体:合成所用单体为经化学修饰的α-氨基酸(或肽核酸);
反应步骤:
第一步,将第一个氨基酸共价连接到树脂上
加入适当的缩合剂如hbtu和hobt,使被保护氨基酸羧基端与树脂形成共脂以完成氨基酸的固定;
第二步,去保护
采用碱性溶剂20%哌啶去除氨基上的fmoc,暴露出氨基;
第三步,激活和交联
采用活化剂hbtu和hobt活化下一个氨基上的羧基,与树脂上的氨基交联,形成肽键;
第四步,重复第二步和第三步,反复循环添加单体氨基酸(或肽核酸),直到合成完成;
第五步,涉及非天然氨基酸合成时,需先选择性脱去dap氨基酸侧链氨基保护基,并使其在edc与hobt的作用下与氯代乙酸进行缩合;
合成后处理:
(1)洗脱和脱保护:用脱保护剂三氟乙酸(tfa)将肽链从树枝上洗脱下来,并脱除保护基。
(2)hplc分析纯化,冻干保存。
利用上述方法合成的两条非天然多肽序列为
cck-pna-1:gactcacatc-kxalkekvaalkekvaalke-nh2(其中x为(2s)-2-amino-3-[(2-chloroacetyl)amino]propanoicacid);
cck-pna-2:ttaggcatca-kxalkekvaalkekvaalke-nh2(其中x为(2s)-2-amino-3-[(2-chloroacetyl)amino]propanoicacid)。
实施例3
本发明的融合蛋白与多肽探针的共价结合是基于融合蛋白上的半胱氨酸与多肽探针上的非天然x氨基酸间的自发反应:
将实施例1得到的融合蛋白使用bca试剂盒进行浓度测定,并将融合蛋白溶解成100μm浓度;
将实施例2得到的多肽探针通过a260的紫外吸收测定浓度,并将多肽探针溶解成500μm浓度;
配置200μl反应体系如下:上述融合蛋白终浓度50μm,上述多肽探针终浓度100μm,20mmhepes,150mmnacl,0.5mmtcep,室温下过夜反应,并使用sds-page检测反应效率(如图3,4);
反应后处理:
反应后的溶液通过排阻色谱柱进行分离,将目标流出液收集并浓缩,最终储存浓度为20μm。
融合蛋白与多肽探针的共价交联产物egfp-pna-1与ssdna模板td(序列为5’-ctgagtgtagtactgtgactgagtgtag-3’,含有两个pna-1的反向互补结合位点)在不同比例时的结合效果。由图5可见,当egfp-pna-1与td的比例为1:10时,溶液中主要成分为结合了一个egfp-pna-1蛋白的td与游离的td;当egfp-pna-1与td的比例为1:1时,溶液中主要成分为结合了一个egfp-pna-1蛋白的td;当egfp-pna-1与td的比例为2.5:1时,溶液中的主要成分为结合了两个egfp-pna-1的td。更精确的浓度滴定与溶液中主成分推测的关系如图6所示。
融合蛋白与多肽探针的共价交联产物egfp-pna-1、mcherry-pna-2与ssdna模板tb(序列为5’-ctgagtgtagtactgtgaaatccgtagt-3’,各含有一个pna-1以及pna-2的反向互补结合位点)结合效果。由图7可见,当egfp-pna-1、mcherry-pna-2以及tb在溶液中进行孵育后,形成了egfp-pna-1与mcherry-pna-2在tb模板上1:1的精确自组装。
- 还没有人留言评论。精彩留言会获得点赞!