雷公藤红素在诱导癌细胞自噬上的应用及其与替莫唑胺的联合用药的制作方法
本发明涉及细胞自噬领域,更具体地,涉及雷公藤红素在诱导癌细胞自噬上的作用。
背景技术:
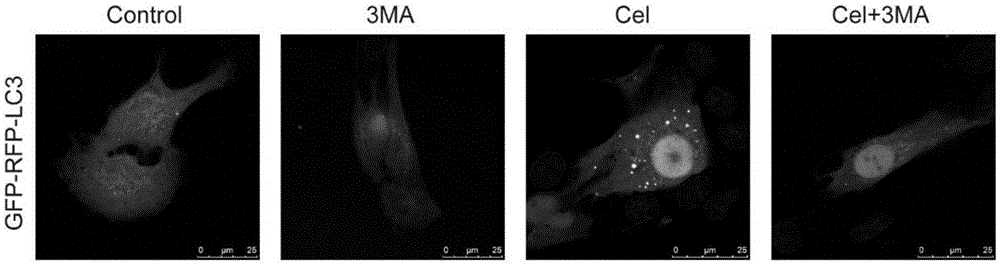
:脑胶质瘤(glioma)是来源于神经上皮的肿瘤,是威胁人类健康的最常见的原发性恶性颅内肿瘤,同时也是侵袭性最强、进展迅速、易复发的恶性肿瘤之一。目前临床上多采用外科手术为主,放化疗为辅的综合治疗方法。替莫唑胺是目前唯一具有明确疗效的化疗药物,但是,其耐药性限制了广泛的应用。因此,迫切需要研究新的化疗药物。雷公藤红素(celastrol)是从雷公藤根茎中提取的一种三萜类化合物,是治疗类风湿性关节炎的雷公藤片和雷公藤多甙片的有效成分。据报道,它是一种具有广泛生物活性的天然化合物,如抗氧化,抗癌,抗炎和杀精作用等。目前的研究表明,雷公藤红素在治疗某些炎症性疾病和肿瘤方面有很多潜在的优势,许多相应的分子靶点已被报道。2007年,cell杂志将雷公藤红素,雷公藤甲素,辣椒素,姜黄素和青蒿素列为最有可能发展成现代药物的五种天然化合物。先前的研究表明,雷公藤红素在多种恶性肿瘤中有明显的抗肿瘤活性,包括肺癌,骨肉瘤,肝癌和前列腺癌。然而,雷公藤红素对脑胶质瘤的作用鲜有报道。细胞凋亡(apoptosis),又称为i型程序性细胞死亡(pcd),是化疗药物作用于各种癌症过程中细胞死亡的主要模式。细胞凋亡的发生通常具有典型的形态变化,包括细胞收缩,核凝结和碎裂以及凋亡小体的形成等。事实上,细胞凋亡的发生比典型的形态学改变更早。细胞自噬(autophagy)是一种进化上保守的细胞内自我消化过程,通过溶酶体依赖性机制维持细胞内稳态,也被称为ii型非凋亡性pcd(也称为自噬性细胞死亡)。自噬的顺利进行有赖于自噬体的形成和自噬体和自噬溶酶体的降解。自噬和肿瘤之间的关系是复杂的,自噬在肿瘤发展的不同阶段以及个体细胞与癌症组织之间的作用是不同的。自噬在肿瘤发生和转移中是一把双刃剑,它可以在某些细胞环境中抑制人类癌症的生长,而在有些环境中,它也可以促进肿瘤的细胞存活。最近的研究表明,已知的诱导细胞凋亡的大量抗肿瘤药物也会引起细胞自噬的发生。另外,但是,至今为止,国内外还未有报道雷公藤红素可以增加胶质瘤细胞对化疗的敏感性。本研究旨在探索雷公藤红素是否可以增加胶质瘤细胞对tmz的化疗敏感性并探索其潜在的机制。技术实现要素:本发明提供一种可促进脑胶质瘤细胞自噬体形成的药物:雷公藤红素。该药物通过影响ros/jnk和akt/mtor信号通路相关蛋白的表达变化,从而引起脑胶质瘤细胞自噬的发生。为实现上述目的,雷公藤红素在制备诱导癌细胞自噬上的药物中的应用。所述药物对癌细胞自噬体形成的诱导作用是通过增强jnk和p38蛋白的磷酸化水平,抑制p-akt和p-mtor蛋白的表达发挥作用的。所述癌细胞为脑胶质瘤细胞。所述癌细胞自噬过程中的诱导因素为增强癌细胞中p-jnk蛋白的表达,抑制p-akt和p-mtor蛋白的表达。根据需求再提供一种雷公藤红素在制备p-jnk或p-p38蛋白激活剂,p-akt或p-mtor抑制剂中的应用。根据需求再提供一种雷公藤红素在p-jnk、p-p38、p-akt、或p-mtor蛋白的研究制剂上的应用根据需求再提供一种雷公藤红素在研究p-jnk、p-p38、p-akt、或p-mtor蛋白,以及它们的底物蛋白的生物学功能上的应用。根据需求再提供一种雷公藤红素在制备自噬启动剂或/和诱导剂上的应用。根据需求,本发明还提供雷公藤红素与化疗药物协同制备治疗肿瘤的药物应用,所述的肿瘤为脑胶质瘤,所述的化疗药物为替莫唑胺或贝伐单抗。进一步地,本发明还提供了一种药物组合物,其用于治疗脑胶质瘤,所述组合物由公藤红素与替莫唑胺组成,所述雷公藤红素与替莫唑胺的重量比为1:1-16;优选为1:1-4。进一步地,本发明还提供了另一种药物组合物,所述雷公藤红素与贝伐单抗的重量比为1:1-16;优选为1:4-16本发明的优点在于,1、本发明公开了雷公藤红素在诱导癌细胞自噬方面的作用,通过诱导自噬作用,帮助癌细胞回归其正常状态,抑制凋亡的发生。雷公藤红素和凋亡抑制剂合用可以促进癌细胞的死亡。2、雷公藤红素可以通过促进ros/jnk和抑制akt/mtor信号通路以实现其对自噬的作用。具体来说,雷公藤红素可以促进ros的产生进而活化jnk蛋白,促进ros/jnk信号通路,同时可以抑制p-akt和p-mtor蛋白的表达,负作用于akt/mtor信号通路,进一步影响自噬。3、作为一种药物,雷公藤红素可以靶向性地作用于癌细胞,有效控制癌细胞,而对正常细胞无影响。4、作为协同药物,雷公藤红素可以提高肿瘤细胞系对于化疗药物的敏感性,所述的肿瘤细胞为脑胶质瘤,并与化疗药物在治疗脑胶质瘤方面产生了明显的协同作用,所述的化疗药物优选为替莫唑胺或贝伐单抗。附图说明图1:u251细胞瞬时转染gfp-mcherry-lc3质粒,向培养液中加入1.5um雷公藤红素或自噬抑制剂3-ma或两者混合物后,共聚焦显微镜观察自噬体的形成。图2:不同浓度雷公藤红素作用于脑胶质瘤细胞系u251,u87-mg和c624h或3um的雷公藤红素作用于三种细胞系不同时间后,自噬相关蛋白lc3ⅱ,p62和beclin-1变化的westernblotting结果。图3:u251细胞提前加入25um的自噬抑制剂cq(提高溶酶体的ph值,抑制自噬的降解过程)2h,再用3um雷公藤红素处理24h后,自噬相关指标lc3ⅱ,p62和beclin-1变化的westernblotting结果。图4:自噬抑制剂和雷公藤红素合用后,cck8检测的细胞活性的变化。图5:雷公藤红素和自噬抑制剂合用后,流式细胞术监测的细胞凋亡的变化。图6:雷公藤红素和自噬抑制剂合用后,自噬和凋亡相关蛋白lc3ⅱ,p62,beclin-1,cleaved-parp,cleaved-caspase3,8和9变化的westernblotting结果.图7:用nac(ros抑制剂)或sp(jnk抑制剂)(图8(1))提前处理三种细胞系2h,再用3um的雷公藤红素作用于细胞24h后,用荧光显微镜和流式细胞术检测ros的产生。图8:不同浓度雷公藤红素作用于u251细胞24h和3um的雷公藤红素作用于细胞不同时间后,细胞中jnk,p38,akt,mtor蛋白及其磷酸化蛋白含量变化的westernblotting结果。图9:雷公藤红素和ros抑制剂nac或jnk抑制剂sp合用后,自噬相关蛋白lc3ⅱ和p62蛋白变化的westernblotting结果。图10:雷公藤红素和akt抑制剂mk或mtor抑制剂ra合用后,自噬相关蛋白lc3ⅱ和p62蛋白变化的westernblotting结果。图11:雷公藤红素在小鼠体内的诱导自噬的相关蛋白的westernblotting结果。图12:雷公藤红素对脑胶质瘤细胞的作用机制图。具体实施方式以下结合附图和具体实施例,进一步阐述本发明。这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中未注明具体条件的实验方法,通常按照常规条件或按照制造厂商所建议的条件。除非另行定义,文中所使用的所有专业与科学用语与本领域熟练人员所熟悉的意义相同。此外,任何与所记载内容相似或均等的方法及材料皆可应用于本发明方法中。下述实施例中所使用的实验方法如无特殊说明,均为常规方法。所用的材料、试剂等,如无特殊说明,均可从商业途径得到。实施例1共聚焦显微镜检测雷公藤红素对脑胶质瘤细胞自噬的诱导作用一、脑胶质瘤细胞株体外培养本实验所用细胞为人脑胶质瘤细胞u251,u87-mg和大鼠脑胶质细胞c6,使用添加10%胎牛血清(fetalbovineserum,fbs,gibco)的mem培养基,在37℃,5%co2培养箱中进行细胞培养,待细胞生长至占培养皿底部70~80%时,使用0.1%胰酶一edta消化并传代。二、雷公藤红素诱导脑胶质瘤细胞自噬的检测实验方法:本实验使用共聚焦显微镜对雷公藤红素诱导的脑胶质瘤细胞的自噬进行检测。雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时稀释为3um。用脂质体lip20002.0ul将0.8uggfp-mcherry-lc3双荧光质粒瞬时转染入u251细胞中,用于标记及追踪lc3以及自噬流的变化。其中gfp是酸敏感型gfp蛋白,而mrfp是稳定的荧光表达基团,不受外界影响。由于自噬小体进入第二阶段后,与溶酶体进行融合,形成自噬溶酶体。自噬溶酶体由于溶酶体内部的酸性环境,可以导致ph下降,gfp淬灭,因此,gfp的减弱可指示自噬溶酶体形成的顺利程度,gfp越少,则从自噬小体到自噬溶酶体阶段流通得越顺畅。反之,自噬小体和溶酶体融合被抑制,自噬溶酶体进程受阻。mrfp是一直稳定表达的,因而可以通过gfp与mrfp的亮点比例来评价自噬流进程。转染24h后,用3-methyladenine(3-ma)预处理细胞2h,再用3um的雷公藤红素处理细胞24h,至于共聚焦显微镜下观察并拍照。实验结果:通过共聚焦显微镜拍摄到的结果可以看出,雷公藤红素可有效诱导细胞自噬体的形成,红色荧光和绿色荧光的亮点均增多。当雷公藤红素和3-ma合用时,这种效应减弱。(图1)说明,雷公藤红素可有效促进自噬体的产生。实施例2lc3蛋白标记法检测雷公藤红素对脑胶质瘤细胞自噬的诱导作用:lc3是细胞自噬形成的一个标志蛋白,具有lc3i和lc3ii两种形式。无自噬发生时,该蛋白再胞质中以lc3i形式存在,自噬形成时,胞浆型lc3(即lc3-i)会酶解掉一小段多肽,转变为(自噬体)膜型(即lc3-ii),lc3-ii/i比值的大小可估计自噬水平的高低。beclin-1是细胞发生自噬时的一个起始蛋白,p62是自噬溶酶体降解自噬底物的一个关键性蛋白。实验方法:雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时分别稀释为0.3,1,3,10um。将细胞消化,离心并重悬浮于相应培养液中,以2.5x105/孔的密度种植于6孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,将细胞分为三组,第一组移除原培养基,加入新鲜配置的,含有0.3,1,3,10um雷公藤红素的培养基中继续培养;第二组移除培养基后加入新鲜配置的,含有3um的培养基中继续培养,分别于培养的6,12,24,36h终止培养。第三组移除培养基后用25um的chloroquinediphosphatesalt(cq)(改变溶酶体的ph值,抑制自噬的溶酶体降解过程)预处理2h后,再用3um的雷公藤红素处理细胞24h。细胞终止培养后立即提取细胞总蛋白,westernblotting法检测上述自噬相关蛋白的表达量的变化。实验结果:westernblotting结果显示,在三种细胞系中,beclin-1和lc3ii的表达了随着雷公藤作用浓度的增高和时间的延长而有升高趋势。同时,p62的表达量也会随着雷公藤红素作用浓度的增高和时间的延长而生高(图2)。当使用雷公藤红素和cq合用后,lc3ii的表达了较之单用cq增加的更加明显。(图3)说明雷公藤红素可以引起脑胶质瘤自噬的发生,可能增加自噬体形成的同时抑制了溶酶体的降解。实施例3雷公藤红素引起的自噬对脑胶质瘤细胞生长的影响实验方法:雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时稀释为3um。将细胞消化,离心并重悬浮于相应培养液中,以2500/孔的密度种植于96孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,分别用3mm的3-ma和25um的cq预处理细胞2h后,用3um的雷公藤红素处理细胞24h,用cck8法检测细胞的存活率。实验结果:结果显示,当使用自噬抑制剂3-ma或cq后,细胞的存活率明显下降。说明雷公藤红素引起的细胞自噬可能起着细胞促存活的作用。(图4)实施例4雷公藤引起的自噬与凋亡的关系。实验方法:雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时分别稀释为0.3,1,3,10um。将细胞消化,离心并重悬浮于相应培养液中,以2.5x105/孔的密度种植于6孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,将细胞分为二组,第一组移除原培养基,加入新鲜配置的,含有0.3,1,3,10um雷公藤红素的培养基中继续培养;第二组,用3mm的3-ma预处理细胞2h后,用3um的雷公藤红素处理细胞24h。细胞终止培养后,第一组收集细胞,用流式细胞仪检测细胞自噬的变化。第二组立即提取细胞总蛋白,westernblotting法检测自噬相关蛋白和凋亡相关蛋白的表达量的变化。实验结果:流式检测的结果显示,当使用3-ma抑制自噬后,细胞的凋亡率明显增加;(图5)westernblotting结果显示,使用3-ma抑制自噬后,凋亡相关蛋白的表达量升高(图6)。综上结果表明,雷公藤红素引起的自噬可以抑制凋亡。实施例5ros/jnk和akt/mtor信号通路的检测实验方法:雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时分别稀释为0.3,1,3,10um。将细胞消化,离心并重悬浮于相应培养液中,以2.5x105/孔的密度种植于6孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,将细胞分为三组,第一组移除原培养基,加入一定浓度的新鲜配置的nac,sp或与3um的雷公藤红素的混合物继续培养;第二组移除培养基后加入新鲜配置的,含有0.3,1,3,10um的雷公藤红素培养24h,或移除培养基后加入新鲜配置的含有3um的培养基,分别继续培养3,6,12,24h后终止培养。细胞终止培养后,第一组收集细胞,分别用显微镜和流式细胞术检测ros的产生。第二组用westernblotting检测相关通路jnk,akt和mtor指标表达的变化。实验结果:荧光显微镜和流式细胞术的结果显示雷公藤红素可以促进ros的产生,nac(ros抑制剂)可以抑制雷公藤红素的这种效果(图7)。当雷公藤红素和sp(jnk抑制剂)时,ros的产生增多的更明显(图8(1))。表明雷公藤红素可以促进ros的产生,而jnk是ros的下游分子。westernblotting法结果显示,随着雷公藤作用浓度的增高和时间的延长,p38,jnk的磷酸化水平升高,而akt和mtor的磷酸化水平则被抑制(图8(2))。这些结果表明雷公藤红素可能激活了ros/jnk信号通路,抑制了akt/mtor信号通路。实验方法:雷公藤红素溶解于dmso中配成1mm浓缩液,-20℃保存,使用时稀释为3um。将细胞消化,离心并重悬浮于相应培养液中,以2.5x105/孔的密度种植于6孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,移除原培养基,加入一定浓度的新鲜配置的nac,sp,mk(akt抑制剂)或ra(mtor抑制剂)或与3um的雷公藤红素的混合物继续培养。细胞终止培养后,用westernblotting检测自噬相关指标lc3ⅱ和p62表达的变化。实验结果:nac和sp可以抑制雷公藤红素引起的lc3ⅱ表达的增高(图9),mk和ra则有相反的效应(图10)。进一步表明,雷公藤红素是通过激活ros/jnk信号通路,抑制akt/mtor信号通路引起胶质瘤细胞的自噬的。实施例6雷公藤红素引起脑胶质瘤细胞自噬及相关通路的体内验证实验方法:将3.5x105u251细胞通过立体定位仪注入balb/c裸鼠的尾状核,造成裸鼠的脑胶质瘤模型。7天模型成功后,分别腹腔注射大(4mg/kg)中(2mg/kg)小(1mg/kg)剂量的雷公藤红素。隔天一次,共注射7次。给药后第14天取材,提取组织的总蛋白,用westernblotting检测自噬蛋白lc3ⅱ的表达和相关通路蛋白jnk,akt和mtodr蛋白的磷酸化水平(图11)。实验结果:结果显示,lc3ⅱ的表达量随着药物剂量的升高而升高,jnk的磷酸化水平升高,akt和mtor的磷酸化水平降低。表明雷公藤红素可以引起脑胶质瘤细胞的自噬,主要是通过活化jnk信号通路,抑制akt/mtor信号通路实现的。与体外实验结果相符。实施例7雷公藤红素联合用药考察基于实施例1-6的发现,雷公藤红素具有明显的p-jnk或p-p38蛋白激活剂和p-akt或p-mtor蛋白抑制剂的作用;进一步地,雷公藤红素对人胶质瘤细胞系u251和u87均具有明显的诱导凋亡作用;因此我们进一步地考察了雷公藤红素与本领域已知的胶质瘤化疗药物的联合用药。实验方法:将u251细胞消化,离心并重悬浮于相应培养液中,以1x105/孔的密度种植于96孔板中,置于37℃,5%co2培养箱中过夜培养。第二天,将细胞分组,第一组移除原培养基,加入新鲜配置的作为对照组,其他各组添加含有不同浓度药物的培养基继续培养24h。各组给药浓度如下:雷公藤红素(ug/ml)替莫唑胺(ug/ml)对照组00组120组201组302组408组5032组621组722组828组9232细胞终止培养后,收集细胞,采用mtt法测定各组细胞活性并计算抑制率(抑制率=(对照组od值-给药组od值)/对照组od值×100%)。协同作用计算方法:采用金正均q值法进行判定(参见非专利文献:戴体俊.合并用药的定量分析[j].中国药理学通报,1998(5):479-480.),q值由以下的公式求得:q=pa+b/(pa+pb-pa×pb)。式中pa、pb和pa+b分别为a药组、b药组和两药联用组治疗率。q<1说明两药合用后产生拮抗作用;q>1,说明两药合用后产生协同作用,q=1说明两药合用后产生相加作用。实验结果:抑制率(%)q值组155.3±4.1-组22.4±0.9-组38.7±2.6-组417.4±3.3-组546.3±4.5-组659.2±3.91.05组778.4±4.21.32组889.3±4.41.41组991.7±3.81.21从上述结果可以看出,雷公藤红素与替莫唑胺比例在1:0.5时两者仅是表现简单的相加作用,而比例在1:1-16时表现为明显协同抑制作用,其中以1:1和1:4效果最佳。提示雷公藤红素与替莫唑胺联合用药在治疗脑胶质瘤方面产生了明显的协同作用,最佳比例在1:1-4之间。类似的结果在u87-mg细胞系上,我们同样观察到了类似的结果,即雷公藤红素与替莫唑在比例1:1和1:4产生了最佳的协同作用。另外,我们采用同样的方法在u251细胞系上考察了雷公藤红素与氯乙基亚硝基脲、依托泊苷、长春新碱或贝伐单抗的联合用药效果,具体结果如下:当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1