核酸的检测的制造方法与工艺
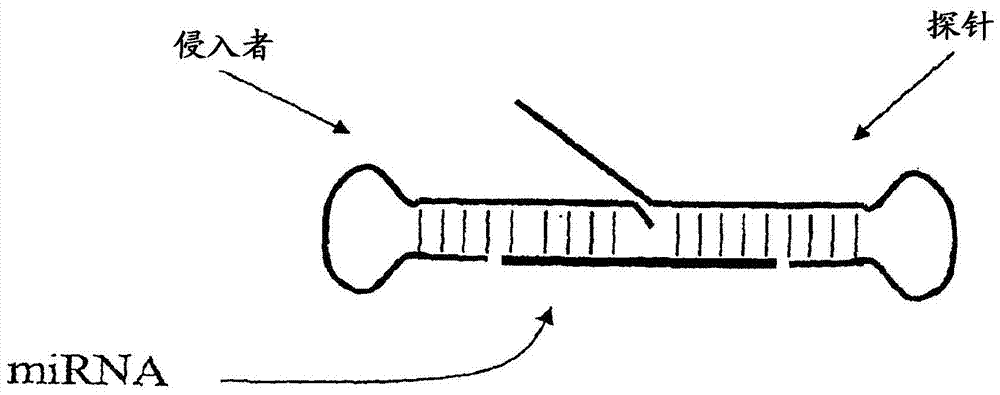
核酸的检测本申请是申请日为2007年6月1日、申请号为200780027007.0、发明名称为“核酸的检测”的发明专利申请的分案申请。技术领域本发明涉及用于检测和表征核酸分子(例如,RNA(例如,小RNA例如微RNA(miRNA)和小干扰RNA(siRNA))和其他短核酸分子)的组合物和方法。更具体而言,本发明涉及用于检测和定量RNA表达的方法。本发明进一步提供了miRNA和siRNA的突变体(例如,缺失突变体)和变体的检测。发明背景微RNA(miRNA)是新的一类非编码RNA,其在无脊椎动物和脊椎动物基因组中编码为短的反向重复(Ambros,(2001)Cell107,823-826;Moss(2002)Curr.Biol.12,R138-R140)。miRNA是靶mRNA翻译和稳定性的调节剂,虽然大多数靶mRNA仍未鉴定。miRNA通过结合至在3'非翻译区(UTRs)中的反义互补位点而以序列特异性的方式来控制靶mRNA的翻译(Ambros,同上;Moss,同上;Lagos-Quintana等人,(2001)Science294,853-858;Lau等人,(2001)Science294,858-862;Lee等人,(2001)Science294,862-864)。miRNA还可以通过其他机制来抑制基因表达(参见,例如Pillai等人,Science309,1573-1576(2005);Humphreys等人,ProcNatlAcadSciUSA102,16961-16966(2005))。在无脊椎动物和脊椎动物之间,几种miRNA,例如let-7RNA、miR-1、miR-34、miR-60和miR-87是高度保守的,这暗示它们可以识别具有可能保守的功能的多个位点和/或多个靶(Lagos-Quintana等人,同上;Lau等人,同上;Lee等人,同上;Pasquinelli等人,(2000)Nature408:86)。小时序RNA(stRNA)lin-4和let-7代表了一亚类通过在秀丽隐杆线虫(Caenorhabditiselegans)中的遗传分析而鉴定出的miRNA,它们在发育上受到调控并且它们本身控制发育进程,例如神经元重新布线(neuronalrewiring)的时机、持久幼虫的形成、阴门的形成以及皮下细胞的终末分化。miRNA通常从60至70个核苷酸的折回RNA前体结构中切出,所述折回RNA前体结构有时在miRNA前体表达开始时(Grishok等人,(2001)Cell106,23-34;Hutvagner等人(2001)Science93,834-838;Ketting等人,(2001)GenesDev.15,2654-2659)或在非常丰富的miRNA的表达过程中被检测出(Lagos-Quintana等人,同上;Lau等人,同上;Lee等人,同上)。一般地,发夹结构前体分子的仅一条链被切除并积累,这可能是由于其受到相关蛋白质的保护而免于RNA降解。这些假定的蛋白质可能介导了翻译抑制。miRNA前体加工反应需要DicerRNaseIII和Argonaute家族成员(Grishok等人,同上;Hutvagner等人,同上;Ketting等人,同上)。除了它们对于基因表达的影响外,小RNA(例如,具有18-25个核苷酸的siRNA或miRNA)还可以用于治疗和药物发现领域(例如,药品靶或药剂)。因此,在某些情况下,重要的是大概知道在细胞中每种miRNA存在的量(例如,在治疗前、在治疗过程中或在治疗后)。此外,miRNA基因的缺失和下调与癌症(例如,B细胞慢性淋巴细胞白血病(CLL))相关,从而在本领域中产生了对于能够检测和表征miRNA表达的需要(参见,例如Calin等人,ProcNatlAcadSciUSA,99,15524-15529(2002))。在某些情况下,也重要的是,比较miRNA在不同组织类型中的水平,或者比较在施加刺激(例如化学或物理干预)之前和之后miRNA的水平。由于相关的siRNA和miRNA可能在细胞中以少量存在,所以希望检测方法是灵敏的和特异的。此外,对于某些应用而言,可能有利的是,鉴定出适用于高通量筛选的方法,例如均相(homogeneous)方法,多重(multiplexed)方法,或适用于高度平行的自动化操作和有限的温度变化的方法。虽然miRNA在基因表达的调节中起着重要作用,但是缺少用于检测和定量miRNA表达的有效技术。用于定量miRNA的方法基于凝胶电泳。通过Northern印迹法或者通过放射性的RNA酶抗性双链体的存在来检测miRNA。Northern印迹法和芯片杂交方法具有相对较低的分析灵敏度(Krichevsky等人2003),因此需要微克量的RNA来用于分析;此外,小RNA向过滤器的转移可引起与定量的再现性有关的问题,并且通常不能修正成高通量。而且,基于RNA酶抗性的检测方法需要高放射性的探针。此外,仅仅基于探针杂交的测定法可能不提供在序列方面密切相关的同种型之间的足够的区分。备选的方法包括克隆miRNA,然后对插入片段进行测序。尽管该方法可适用于区分在密切相关的miRNA种类之间的单碱基差异,但是这种方法是耗时且费力的。与miRNA相似,小干扰RNA(siRNA)是通过称为RNA干扰(RNAi)的应答而参与细胞防御(例如,针对病毒RNA)的小RNA分子(Cullen,B.R.,NatureImmunology,3:597-599(2002))。通过作为针对细胞中存在双链RNA的RNAi应答的一部分的Dicer酶和RNA诱导的沉默复合物(RNA-inducedsilencingcomplex,RISC)蛋白质复合体的作用,产生出一类siRNA(Khvorova,A.等人,Cell115:209-216(2003))。另一类siRNA是合成的,并且包括通常21-23nt的短双链体,其具有特征性的二核苷酸突出端(Elbashir,S.M.等人,EMBOJ.20:6877-6888(2001)),其通过所引入的载体的转染或表达而被直接引入到细胞中(Paul,C.P.等人,NatureBiotechnology20:505-508(2002),美国专利申请公开号2003/0148519A1,通过提及而以其整体合并入本文以用于所有目的)。在某些情况下,siRNA似乎坚持作为确定的序列,这使得它们在功能和组成方面与miRNA相似(Elbashir,S.M.等人,同上)。需要用于检测和表征(例如定量)miRNA和siRNA水平的有效且精确的方法。发明概述本发明涉及用于检测和表征核酸分子(例如,RNA(例如,小RNA例如微RNA(miRNA)和小干扰RNA(siRNA))和其他短核酸分子)的组合物和方法。更具体而言,本发明涉及改进的用于检测和表征(例如定量)RNA表达的方法。例如,本发明提供了一种方法,其包括:使至少一种核酸(例如,其包含不与干扰RNA互补的序列)与干扰RNA靶杂交从而产生检测结构,并检测该检测结构。在一些实施方案中,干扰RNA靶为miRNA。在其他实施方案中,干扰RNA靶为siRNA。在一些实施方案中,siRNA是双链的。在一些实施方案中,所述检测结构包括侵入性切割结构(invasivecleavagestructure)。例如,在一些实施方案中,所述核酸包含第一种和第二种寡核苷酸,它们被构造成与miRNA相组合地形成侵入性切割结构。在一些实施方案中,所述核酸包含第一种寡核苷酸,该寡核苷酸被构造成与所述miRNA相组合地形成侵入性切割结构。在一些实施方案中,所述第一种寡核苷酸包含5'部分和3'部分,其中所述3'部分被构造成与所述靶序列杂交,和其中所述5'部分被构造成不与所述靶序列杂交。在使用第二种寡核苷酸的一些实施方案中,所述第二种寡核苷酸包含5'部分和3'部分,其中所述5'部分被构造成与所述靶序列杂交,和其中所述3'部分被构造成不与所述靶序列杂交。在一些实施方案中,所述检测步骤包括使用INVADER测定法。在一些实施方案中,所述检测结构包含环状寡核苷酸,其与所述小RNA杂交从而产生环状检测结构。在一些实施方案中,所述检测步骤包括使用滚环复制测定法。在一些实施方案中,所述检测结构包含具有通过聚合酶进行延伸(例如,模板依赖性的延伸)的游离3'-OH基团的核酸分子,并且直接或间接检测经延伸的序列。在一些实施方案中,所述检测步骤包括使用检测测定法,包括但不限于:测序测定法、聚合酶链式反应测定法、杂交测定法、使用与突变互补的探针的杂交测定法、微阵列测定法、珠阵列测定法、引物延伸测定法、酶错配切割测定法、分支杂交测定法(branchedhybridizationassays)、NASBA测定法、分子信标测定法、循环探针测定法、连接酶链式反应测定法、侵入性切割结构测定法、ARMS测定法以及夹心式杂交测定法。在一些优选的实施方案中,所述检测步骤是在细胞裂解物中进行的。在一些实施方案中,本发明的方法包括检测第二种核酸靶。在一些优选的实施方案中,所述第二种核酸靶为RNA。在一些特别优选的实施方案中,所述第二种核酸靶为U6RNA或GAPDHmRNA。在一些实施方案中,用于形成检测结构的核酸包括模板,其具有一个或多个与小RNA充分互补的位点,从而使得所述RNA与所述模板杂交并且在延伸反应中被延伸。在一些实施方案中,所述延伸反应为聚合酶链式反应,其中一种或多种RNA在所述聚合酶链式反应中用作引物。在一些此类实施方案中,单种类型的RNA与所述模板上的两个位置结合,从而提供了聚合酶链式反应引物。在其他实施方案中,两种或更多种RNA被用作引物。在此类实施方案中,扩增产物的检测表明样品中存在所述两种或更多种RNA(即,miRNA多重(multiplex)测定法)。在连接酶链式反应中,可以采用类似的方法,其中miRNA用作连接的寡核苷酸。在一些实施方案中,将RNA用作模板,以用于通过跨越该RNA模板的至少一部分的引物延伸来对检测复合物进行修饰。在一些实施方案中,所述方法包括检测多种miRNA。在一些此类实施方案中,所述多种miRNA包括同一miRNA的多态性。在其他实施方案中,所述多种miRNA包括不同的miRNA(例如,Let-7、miR-1、miR-1d、miR-135、miR-15、miR-16、miR-124a或miR125b)。本发明还提供了用于实施上述方法中的任一种方法的试剂盒。例如,在一些实施方案中,本发明提供了包含核酸的试剂盒,所述核酸被构造成当与RNA靶序列杂交时形成检测结构。在一些实施方案中,所述试剂盒被构造成用于检测miRNA。在一些优选的实施方案中,所述试剂盒被构造成用于检测Let-7、miR-1、miR-135、miR-15、miR-16、miR-1b、miR-124a或miR125bmiRNA。在一些优选的实施方案中,所述试剂盒被构造成用于共同检测第二种RNA靶以及miRNA靶。本发明还提供了用于检测miRNA靶的方法,其包括下列步骤:提供(i)miRNA靶,(ii)第一种未标记的寡核苷酸,(iii)第二种未标记的寡核苷酸,(iv)逆转录酶,(v)聚合酶,和(vi)探针寡核苷酸;在使得检测结构能够形成的条件下温育(i)至(vi);以及检测所述检测结构。在一些实施方案中,所述第一种未标记的寡核苷酸包含与所述miRNA靶互补的第一区域以及与所述miRNA靶不互补的第二区域。在一些实施方案中,所述第二种未标记的寡核苷酸包含与所述miRNA靶的第二区域互补的第一区域以及与所述miRNA靶的第二区域不互补的第二区域。在一些实施方案中,所述检测包括:形成侵入性切割结构,切割所述侵入性切割结构,并检测所述侵入性切割结构的切割。在一些实施方案中,所述温育进一步包括使用能够切割检测结构但不具有聚合酶活性的酶进行温育。在一些实施方案中,所述酶为5'核酸酶,而在一些实施方案中,所述酶包括FEN-1核酸酶。在一些实施方案中,切割侵入性切割结构的过程在45℃至60℃的温度下进行。在一些实施方案中,切割侵入性切割结构的过程在大约50℃的温度下进行。在一些实施方案中,所述第一种未标记的寡核苷酸用作用于反转录的引物。在一些实施方案中,所述第一种未标记的寡核苷酸在侵入性切割反应中用作INVADER寡核苷酸。在一些实施方案中,(i)至(vi)存在于同一个反应容器内。在一些实施方案中,所述方法进一步包括提供(vii)第二种探针寡核苷酸。在一些实施方案中,所述第一种未标记的寡核苷酸和所述逆转录酶反转录miRNA靶。在一些实施方案中,在聚合酶链式反应中通过所述第一种未标记的寡核苷酸和所述第二种未标记的寡核苷酸和所述DNA聚合酶来扩增经反转录的miRNA靶(即,miRNAcDNA靶)。在一些实施方案中,该经扩增的、经反转录的miRNA靶在所述探针寡核苷酸存在下形成检测结构。在一些实施方案中,所述第一种未标记的寡核苷酸包含这样的核酸序列,即使得在所述寡核苷酸和所述miRNA靶之间形成大约6-7个碱基对的双链体。本发明能够检测以非常少的拷贝数存在的miRNA。例如,在一些实施方案中,检测了样品中少于200个拷贝的miRNA。在一些实施方案中,检测了样品中少于100个拷贝的miRNA。在一些实施方案中,所述第二种未标记的寡核苷酸包含这样的核酸序列,即使得在所述寡核苷酸和所述miRNA靶之间形成大约9个碱基对的双链体。在一些实施方案中,所述探针寡核苷酸包含这样的核酸序列,即使得在所述寡核苷酸和所述miRNA靶或所述miRNA靶的经扩增的拷贝之间形成大约8-10个碱基对的双链体。在一些实施方案中,所述第一种未标记的寡核苷酸探针的第二区域包含第一部分和第二部分,其中所述第一部分和所述第二部分可以彼此杂交。在一些实施方案中,当所述第一部分和所述第二部分彼此杂交时,在所述第一种未标记的寡核苷酸探针中形成发夹结构。在一些实施方案中,所述第二种未标记的寡核苷酸探针的第二区域包含第一部分和第二部分,其中所述第一部分和所述第二部分可以彼此杂交。在一些实施方案中,当所述第一部分和所述第二部分彼此杂交时,在所述第二种未标记的寡核苷酸探针中形成发夹结构。在一些实施方案中,所述方法进一步包括提供(vii)与所述第一种未标记的寡核苷酸探针的区域互补的寡核苷酸。在一些实施方案中,所述方法进一步包括提供(vii)与所述第二种未标记的寡核苷酸探针的区域互补的寡核苷酸。在一些实施方案中,在大约50℃的温度下切割侵入性切割结构使得能够以高保真度来区分靶序列,虽然可以基于序列、缓冲液成分等来选择其他温度以达到最佳性能。在一些实施方案中,所述靶序列包括单一种类的变体miRNA。在一些实施方案中,增加探针寡核苷酸的浓度会增加检测miRNA靶的灵敏度。在一些实施方案中,检测了两种或更多种miRNA。在一些实施方案中,所述检测包括使用经标记的探针。在一些实施方案中,所述经标记的探针是经荧光标记的。在一些实施方案中,所述经标记的探针被构造成用于FRET检测。在一些实施方案中,所述经标记的探针当未在双链体中杂交时具有第一构象,和当在双链体中杂交时具有第二构象。在一些实施方案中,当在双链体中杂交时,所述经标记的探针展现出增强的荧光。本发明并不受到所检测的miRNA的限制。事实上,可以使用本发明的组合物和方法来检测各种miRNA,包括但不限于Let-7、miR-1、miR-135、miR-15、miR-16、miR125b、miR-1d和miR124a。本发明还提供了试剂盒,其包含下列中的一种或多种:第一种未标记的寡核苷酸,其包含与miRNA靶互补的第一区域以及与所述miRNA靶不互补的第二区域;第二种未标记的寡核苷酸,其包含与所述miRNA靶的第二区域互补的第一区域以及与所述miRNA靶的所述第二区域不互补的第二区域;逆转录酶;DNA聚合酶;探针寡核苷酸;以及能够切割检测结构的酶。在一些实施方案中,所述检测结构包括侵入性切割结构。在一些实施方案中,所述试剂盒被构造成用于检测miRNA靶以及至少一种其他RNA靶。在一些实施方案中,所述试剂盒被构造成用于检测细胞裂解物中的miRNA靶序列。附图说明图1显示了用于在单个反应中检测两个不同的等位基因(例如,区别在于单个核苷酸)的INVADER(侵入者)寡核苷酸、探针寡核苷酸和FRET盒的示意图。图2显示了在本发明的一些实施方案中使用的示例性检测结构。图3显示了在本发明的一些实施方案中使用的第二种示例性检测结构。图4显示了在本发明的一些实施方案中使用的第三种示例性检测结构。图5显示了本发明所使用的示例性寡核苷酸。靶miRNA的碱基为小写字体并带有下划线。探针或INVADER寡核苷酸中的DNA残基为规则字体。小写字体表示2'-O-甲基残基。图6显示了关于let-7的温度最佳化实验的结果。图7显示了关于let-7的温度最佳化实验的结果。图8显示了关于let-7的检测极限(LOD)实验的结果。图9显示了使用let-7miRNA的交叉反应性实验的结果。图10显示了关于miR-1的LOD实验的结果。图11显示了使用let-7miRNA的CLEAVASE酶IX和XII比较的结果。图12显示了来自各种生物的U6RNA序列的部分序列比对。图13显示了关于mir-135的温度最佳化实验的结果。图14显示了关于mir-135的LOD实验的结果。图15包含了关于检测细胞裂解物中的let-7所获得的平均计数的图形表示。图16显示了在使用和未使用RNA酶A处理的细胞裂解物中miRNA和mRNA的结果。图17显示了侵入性切割测定法的结果,其比较了包括全长ARRESTOR(阻止者)寡核苷酸的效果与包括缩短的ARRESTOR寡核苷酸的效果。图18显示了温度最佳化实验的结果,其使用在图16中所描述的测定法设计。图19显示了温度最佳化实验的结果,其使用10聚体探针和12聚体INVADER寡核苷酸设计。图20显示了比较两种备选的寡核苷酸设计的LOD的实验结果。图21显示了结果,其比较了用2’-脱氧残基替代探针和INVADER寡核苷酸中的一些或全部2’-O-甲基残基的效果。图22显示了用于检测miR-124a的侵入性切割测定法的结果。图23显示了用于在从20种不同组织类型中分离得到的总RNA中检测五种不同的miRNA种类的实验结果。图24显示了实验结果,所述实验测试了改变探针和寡核苷酸长度对于miRNA检测的影响。图25显示了用于检测siRNA的示例性的侵入性切割寡核苷酸设计。小写字母表示2’-O-甲基。图26显示了具有相同的let-7a杂交区域、但具有不同的5'-翼片(flap)“臂”序列的两种探针。图27显示了图表,其描绘了对于分别以1μM和10nM(混合物vii)以及1μM和4nM(混合物ix)进行混合的探针1544-82-01和2343-25-01而言,拷贝数对净信号的图。图28显示了具有相同的U6RNA杂交区域、但具有不同的5'-翼片“臂”序列的两种寡核苷酸探针。图29显示了图表,其描绘了对于分别以1μM和4nM混合物进行混合的探针1796-53-01和2343-30-01而言,拷贝数对净信号的图。图30显示了图表,其描绘了对于双重U6和Let-7a检测而言,拷贝数对净信号的图。图31显示了在用于检测和表征与癌症相关的miRNA的本发明开发过程中所产生的各种寡核苷酸。图32描述了用于使用包括反转录反应、聚合酶链式反应和侵入性切割测定法反应的测定法来检测miRNA的总体设计。图33显示了净信号对Let-7a拷贝数/反应的图。图34显示了作为温度的函数的净信号。图35显示了作为let-7a拷贝数的函数进行作图的let-7a反应的原始信号(rawsignal)。图36显示了作为let-7aRNA拷贝数的函数进行作图的let-7a堆叠者(stacker)和发夹测定法的原始信号。图37显示了作为let-7a拷贝数的函数的通过一步和两步let-7a测定法所产生的净信号。图38显示了作为let-7a拷贝数的函数的通过使用探针1716-94-1、1717-94-10或1716-94-11的let-7a测定法所产生的净信号。图39显示了作为let-7a拷贝数的函数的通过使用不同比率的探针1716-94-1和1717-94-11的let-7a测定法所产生的净信号。图40显示了作为let-7a、let-7c、let-7e或let-7f拷贝数的函数的通过let-7a测定法所产生的净信号。图41显示了根据实施例19(M)中所列出的指导方针而设计的、用于检测与癌症相关的miRNA而产生的寡核苷酸。图42显示了具有变化长度的初级探针(primaryprobe)和PCR引物杂交区域的用于let-7a和miR-16的另外设计。图43提供了表格,其列出了在实施例19中使用的所有寡核苷酸,分别为SEQIDNO:144-285。定义为了帮助理解本发明,下面定义了许多术语和短语:如本文所使用的,术语“miRNA”是指微RNA。如本文所使用的,术语“miRNA靶序列”是指待检测(例如在其他核酸存在下)的miRNA。在一些实施方案中,miRNA靶序列为miRNA的变体。如本文所使用的,术语“RNA检测结构”和“检测结构”是指通过使核酸(例如寡核苷酸)与RNA靶(例如miRNA或siRNA)杂交而形成的结构。在一些实施方案中,所述核酸为单种核酸(例如,其一个或多个小区域与miRNA具有同源性的较大核酸)。在其他实施方案中,所述核酸包括两种核酸(例如,它们与miRNA杂交从而形成发夹(例如单个或两个发夹)结构)。在优选的实施方案中,miRNA检测结构能够采用已知的核酸检测方法(包括但不限于本文所公开的那些)来检测。在一些实施方案中,在杂交步骤之后,进一步修饰RNA检测结构。例如,在一些实施方案中,所述检测结构的一个或多个组分提供了用于通过核酸聚合酶进行延伸的模板。在其他实施方案中,所述检测结构的一个或多个组分与连接酶相接触,并被连接至另外的核酸。术语“siRNA”是指短干扰RNA。在一些实施方案中,siRNA包含双链体或双链区域,其中所述双链区域的每条链的长度为大约18至25个核苷酸;所述双链区域的长度可以短至16个碱基和长至29个碱基对,其中该长度由反义链决定。通常,siRNA在每条链的3'末端包含大约2至4个未配对的核苷酸。在无脊椎动物和脊椎动物中引发RNA干扰方面,以及在植物中在转录后基因沉默过程中引发序列特异性RNA降解方面,siRNA似乎起着关键的中间物的作用。siRNA的双链体或双链区域中的至少一条链与靶RNA分子基本上同源或者基本上互补。与靶RNA分子互补的链为“反义”链;与靶RNA分子同源的链为“正义”链,并且该链还与siRNA反义链互补。双链区域中的一条链不必具有相反链的确切长度,因此,一条链可以比相反的互补链少至少一个核苷酸,从而导致在相反链中形成“泡”或至少一个未匹配的碱基。双链区域的一条链不必与相反链精确互补;因此,所述链(优选为正义链)可以具有至少一个错配的碱基对。siRNA还可以包含另外的序列;此类序列的非限定性实例包括,连接序列,或环,其连接了双链体区域的两条链。这种形式的siRNA可以被称为“si-样RNA”、“短发夹siRNA”(其中短涉及siRNA的双链体区域)或“发夹siRNA”。在siRNA中存在的另外序列的另外非限定性实例包括茎和其他折叠结构。所述另外的序列可以具有或不具有已知的功能;此类功能的非限定性实例包括增加siRNA分子的稳定性或提供细胞目的信号(cellulardestinationsignal)。如本文所使用的,术语“受试者”和“患者”是指任何生物,包括植物、微生物和动物(例如,哺乳动物例如狗、猫、家畜和人)。如本文所使用的,术语“INVADER测定法试剂”或“侵入性切割测定法试剂”是指一种或多种用于检测靶序列的试剂,所述试剂包括能够在靶序列存在下形成侵入性切割结构的寡核苷酸。在一些实施方案中,INVADER测定法试剂进一步包括用于检测侵入性切割结构的存在的试剂(例如切割试剂)。在一些实施方案中,所述寡核苷酸包括第一种寡核苷酸和第二种寡核苷酸,所述第一种寡核苷酸包含与靶核酸的第一区域互补的5'部分,和所述第二种寡核苷酸包含3'部分和5'部分,其中所述5'部分与位于所述第一部分下游并与之邻接的靶核酸的第二区域互补。在一些实施方案中,所述第二种寡核苷酸的3'部分包含与靶核酸不互补的3'末端核苷酸。在优选的实施方案中,所述第二种寡核苷酸的3'部分由与靶核苷酸不互补的单个核苷酸组成。在一些实施方案中,所述第一种寡核苷酸和第二种寡核苷酸彼此共价偶联(例如,通过连接体)。在一些实施方案中,INVADER测定法试剂进一步包括固体支持物。例如,在一些实施方案中,测定法试剂的所述一种或多种寡核苷酸(例如,第一种和/或第二种寡核苷酸,不论桥接还是非桥接)被附着在固体支持物上。在一些实施方案中,INVADER测定法试剂进一步包括缓冲液。在优选的实施方案中,所述缓冲液包含二价阳离子(例如Mn2+和/或Mg2+离子)源。共同构成INVADER测定法试剂的独个成分(例如,寡核苷酸、酶、缓冲剂、靶核酸)被称为“INVADER测定法试剂组分”。在一些实施方案中,INVADER测定法试剂进一步包括第三种寡核苷酸,其与位于所述第一种靶核酸的第一部分上游的靶核酸的第三部分互补。在其他实施方案中,INVADER测定法试剂进一步包括靶核酸。在一些实施方案中,INVADER测定法试剂进一步包括第二种靶核酸。在其他实施方案中,INVADER测定法试剂进一步包括第三种寡核苷酸,该寡核苷酸包含与所述第二种靶核酸的第一区域互补的5'部分。在一些具体的实施方案中,所述第三种寡核苷酸的3'部分与所述第二种靶核酸共价连接。在其他具体的实施方案中,所述第二种靶核酸进一步包含5'部分,其中所述第二种靶核酸的所述5'部分为第三种寡核苷酸。在其他实施方案中,INVADER测定法试剂进一步包括ARRESTOR分子(例如ARRESTOR寡核苷酸)。包括2'O-甲基化的ARRESTOR寡核苷酸(其与每种探针的靶特异性区域完全碱基配对,并且与该探针的5'-翼片区域部分碱基配对)会隔离未切割的探针并防止在次级反应(secondaryreaction)中X结构的形成,如Eis等人,NatureBiotechnology,19:673-676(2001)所述,该文献通过提及而以其整体合并入本文以用于所有目的。在一些优选的实施方案中,INVADER测定法试剂进一步包括用于检测核酸切割产物的试剂。在一些实施方案中,在INVADER测定法试剂中的一种或多种寡核苷酸包含标记物。在一些优选的实施方案中,所述第一种寡核苷酸包含标记物。在其他优选的实施方案中,所述第三种寡核苷酸包含标记物。在特别优选的实施方案中,所述试剂包括用能够产生荧光共振能量转移(FRET)效应的部分进行标记的第一种和/或第三种寡核苷酸。在一些实施方案中,可以以预调配(predispensed)形式提供一种或多种INVADER测定法试剂(即,进行预测量以在该操作过程的步骤中使用,而无需再测量或再调配)。在一些实施方案中,将所选的INVADER测定法试剂组分混合和预调配在一起。在优选的实施方案中,将经预调配的测定法试剂组分进行预调配,并提供在反应容器(包括但不限于反应管或孔(例如微量滴定板中的孔))中。在特别优选的实施方案中,经预调配的INVADER测定法试剂组分在反应容器中干燥(例如,烘干或冻干)。在一些实施方案中,INVADER测定法试剂以试剂盒的形式提供。如本文所使用的,术语“试剂盒”是指用于递送材料的任何递送系统。在反应测定法的情况下,这样的递送系统包括允许从一个地方至另一个地方贮存、运输或递送反应试剂(例如,在合适的容器中的寡核苷酸、酶等)和/或支持材料(例如,缓冲液、施行该测定法的书面说明书等)的系统。例如,试剂盒包含一个或多个装有相关反应试剂和/或支持材料的封围物(例如,盒子)。如本文所使用的,术语“分部式试剂盒(fragmentedkit)”是指包含两个或更多个分开的容器的递送系统,所述容器各装有全部试剂盒组分的亚部分(subportion)。可一起或分开地将所述容器递送至想要的接受者。例如,第一个容器可装有用于测定法的酶,而第二个容器装有寡核苷酸。术语“分部式试剂盒”意在涵盖装有在联邦食品、药品和化妆品法案(FederalFood,Drug,andCosmeticAct)的条款520(e)的管制之下的分析物特异性试剂(Analytespecificreagents,ASR’s)的试剂盒,但并不局限于此。事实上,在术语“分部式试剂盒”中包括任何包含两个或更多个分开的容器的递送系统,所述容器各装有全部试剂盒组分的亚部分。相反地,“组合式试剂盒(combinedkit)”是指在单个容器中(例如,在装有各想要的组分的单个盒子中)装有反应测定法的所有组分的递送系统。术语“试剂盒”包括分部式试剂盒和组合式试剂盒。在一些实施方案中,本发明提供了包含一种或多种对于实施本发明所必需的组分的INVADER测定法试剂盒。例如,本发明提供了用于贮存或递送对于实施INVADER测定法所必需的酶和/或反应组分的试剂盒。该试剂盒可包含对于测定法所必需的或所希望的任何和所有组分,包括但不限于试剂本身、缓冲液、对照试剂(例如,组织样品、正和负对照靶寡核苷酸等)、固体支持物、标签、文字和/或图示的说明书和产品信息、抑制剂、标记和/或检测试剂、包装环境控制物(例如,冰、干燥剂等),等等。在一些实施方案中,试剂盒提供所需组分的亚组,其中期望用户可提供其余的组分。在一些实施方案中,试剂盒包含两个或更多个分开的容器,其中每个容器装有待递送的组分的亚组。例如,第一个容器(例如,盒子)可包含酶(例如,在合适的贮存缓冲液和容器中的结构特异性切割酶),而第二个盒子可包含寡核苷酸(例如,INVADER寡核苷酸、探针寡核苷酸、对照靶寡核苷酸等)。如本文所使用的,术语“标记物”是指可用于提供可检测的(优选地可定量的)效果且可附着至核酸或蛋白质的任何原子或分子。标记物包括但不限于:染料;放射性标记物例如32P;结合部分例如生物素;半抗原例如洋地黄毒苷;发光的、发磷光的或发荧光的部分;质量标签;以及荧光染料,其是单独的或者与可通过荧光共振能量转移(FRET)抑制或移动发射光谱的部分相组合。标记物可提供能够通过荧光、放射性、比色法、重量分析法、X射线衍射或吸收、磁力、酶活性、质量特征或受质量影响的行为(例如,MALDI飞行时间质谱法;荧光偏振)等进行检测的信号。标记物可以是带电荷的部分(正或负电荷),或者备选地,可以是电中性的。标记物可包括或由核酸或蛋白质序列组成,只要包含该标记物的序列是可检测的。如本文所使用的,当涉及信号时,术语“不同的”是指可以相互区分开的信号,例如,通过光谱的性质例如荧光发射波长、颜色、吸光度、质量、大小、荧光偏振性质、电荷等,或者通过与另一部分(例如,与化学试剂、酶、抗体等)相互作用的能力。如本文所使用的,术语“互补”或“互补性”用于涉及通过碱基配对法则建立联系的多核苷酸(即,核苷酸例如寡核苷酸或靶核酸的序列)。例如,序列“5'-A-G-T-3'”与序列“3'-T-C-A-5'”互补。互补性可以是“部分的”,其中核酸的碱基中只有一些按照碱基配对法则相匹配。或者,核酸之间可存在“完全”或“全部”互补性。核酸链之间的互补性程度对于核酸链之间的杂交效率和强度具有重大影响。这在扩增反应以及依赖于核酸之间的结合的检测方法中是特别重要的。上述术语中的任何一个还可以用于涉及独个核苷酸,特别是在多核苷酸的情况下。例如,寡核苷酸内的特定核苷酸可以就其与另一核酸链内的核苷酸具有互补性或缺乏互补性进行标注,与所述寡核苷酸的其余部分与所述核酸链之间的互补性相比对或相比较。术语“同源性”和“同源的”是指同一性的程度。可以存在部分同源性或完全同源性。部分同源的序列是与另一序列的同一性小于100%的序列。如本文所使用的,术语“杂交”用于涉及互补核酸的配对。杂交和杂交强度(即,核酸之间结合的强度)受到例如下列因素的影响:核酸之间的互补程度,所涉及的条件的严格性,和所形成的杂交物的Tm。“杂交”方法涉及使一个核酸与另一个互补的核酸(即,具有互补的核苷酸序列的核酸)退火。两个包含互补序列的核酸聚合物彼此发现并通过碱基配对相互作用进行退火的能力是得到充分认识的现象。Marmur和Lane,Proc.Natl.Acad.Sci.USA46:453(1960)以及Doty等人,Proc.Natl.Acad.Sci.USA46:461(1960)对“杂交”过程进行了最初的观察,在这之后该过程被改进成现代生物学的必要工具。如本文所使用的,核酸序列的互补物是指这样的寡核苷酸,所述寡核苷酸当以使得一个序列的5'末端与另一序列的3'末端配对的方式与所述核酸序列进行比对时,处于“抗平行联系”之中。在本发明的核酸中可以包括通常在天然核酸中无法找到的某些碱基,包括例如肌苷和7-脱氮鸟嘌呤。互补性不必是完全的;稳定的双链体可以包含错配的碱基对或未匹配的碱基。核酸技术领域中的技术人员可以通过考虑许多变量来凭经验确定双链体的稳定性,所述变量包括例如寡核苷酸的长度、寡核苷酸的碱基组成和序列、离子强度以及错配的碱基对的发生率。如本文所使用的,术语“Tm”用于表示“解链温度”。解链温度是一群双链核酸分子有一半解离成单链时所处的温度。用于计算核酸的Tm的几种公式在本领域内是熟知的。如由标准参考文献所指出的,当核酸在1MNaCl的水溶液中时,可通过公式:Tm=81.5+0.41(%G+C)来简单地估算Tm值(参见,例如Anderson和Young,QuantitativeFilterHybridization,inNucleicAcidHybridization(1985))。其他文献(例如Allawi和SantaLucia,Biochemistry36:10581-94(1997))包括考虑结构和环境以及序列特征来计算Tm的更复杂的计算方法。术语“基因”是指这样的DNA序列,所述DNA序列包含对于产生具有非编码功能的RNA(例如核糖体RNA或转移RNA)、多肽或前体所必需的控制和编码序列。所述RNA或多肽可以由全长编码序列或者由编码序列的任何部分来编码,只要所需的活性或功能得到保留。术语“野生型”是指这样的基因或基因产物,所述基因或基因产物具有当从天然发生的来源分离时的该基因或基因产物的特征。野生型基因是在群体中最常观察到的从而被专断地设定为该基因的“正常”或“野生型”形式的基因。相反地,术语“经修饰的”、“突变体”或“多态的”是指当与野生型基因或基因产物相比较时,展示出在序列和/或功能特性方面的修饰(即,改变的特征)的基因或基因产物。要指出的是,可分离天然发生的突变体;通过当与野生型基因或基因产物相比较时,它们具有改变的特征这样的事实来鉴定这些突变体。如本文所使用的,术语“重组DNA载体”是指包含所需异源序列的DNA序列。例如,虽然该术语不限于使用所表达的序列或编码表达产物的序列,但是在一些实施方案中,所述异源序列为这样的编码序列和合适的DNA序列,所述编码序列和合适的DNA序列对于在宿主生物体中复制编码序列或者在特定宿主生物体中表达有效连接的编码序列而言是必需的。对于在原核生物中进行表达而言所必需的DNA序列包括启动子、可任选地操纵子序列、核糖体结合位点以及可能地其他序列。已知,真核细胞使用了启动子、聚腺苷酸化信号和增强子。如本文所使用的,术语“寡核苷酸”被定义为包含2个或更多个脱氧核糖核苷酸或核糖核苷酸、优选至少5个核苷酸、更优选至少大约10-15个核苷酸、和更优选至少大约15-30个核苷酸的分子。确切大小将取决于许多因素,这些因素反过来取决于该寡核苷酸的最终的功能或用途。可以以任何方式产生寡核苷酸,所述方式包括化学合成、DNA复制、反转录、PCR或其组合。因为以使得一个单核苷酸戊糖环的5'磷酸通过磷酸二酯键在一个方向上附着至其相邻者的3'氧这样的方式使单核苷酸进行反应从而制备寡核苷酸,所以如果其5'磷酸不连接至单核苷酸戊糖环的3'氧,则寡核苷酸的末端被称为“5'末端”,并且如果其3'氧不连接至随后的单核苷酸戊糖环的5'磷酸,则被称为“3'末端”。如本文所使用的,核酸序列,即使位于更大的寡核苷酸的内部,也可被认为具有5'和3'末端。当以5'至3'的方向沿着核酸链移动时,如果第一区域的3'末端在第二区域的5'末端前面,则沿着核酸链的第一区域被称为在另一区域的上游。当两个不同的非重叠的寡核苷酸与相同的线性互补核酸序列的不同区域退火,并且一个寡核苷酸的3'末端指向另一个寡核苷酸的5'末端时,可以将前者称为“上游”寡核苷酸,和将后者称为“下游”寡核苷酸。类似地,当两个重叠的寡核苷酸与相同的线性互补核酸序列杂交,并且以使得第一个寡核苷酸的5'末端位于第二个寡核苷酸的5'末端的上游而且第一个寡核苷酸的3'末端位于第二个寡核苷酸的3'末端的上游这样的方式对第一个寡核苷酸进行定位时,可以将第一个寡核苷酸称为“上游”寡核苷酸,和可以将第二个寡核苷酸称为“下游”寡核苷酸。术语“引物”是指这样的寡核苷酸,当被置于起始引物延伸的条件下时,所述寡核苷酸能够用作合成的起始点。寡核苷酸“引物”可以天然地发生,如在经纯化的限制性消化物中,或者可以合成产生。选择引物,以使其“基本上”与模板的特异性序列的链互补。引物必须具有足够的互补性,以便与模板链杂交,从而发生引物的延伸。引物序列不必反映模板的精确序列。例如,非互补的核苷酸片段可以附着至引物的5'末端,而引物序列的其余部分与该链基本上互补。可以将非互补的碱基或更长的序列散置在引物中,条件是引物序列与模板序列具有足够的互补性,以便进行杂交并由此形成模板引物复合物以用于合成引物的延伸产物。如本文所使用的,术语“切割结构”是指通过以下过程形成的结构:使至少一种探针寡核苷酸与靶核酸相互作用,从而形成包含双链体的结构,所得到的结构是可以通过切割手段(包括但不限于酶)进行切割的。切割结构是用于通过切割手段进行特异性切割的底物,这与作为用于通过诸如磷酸二酯酶的试剂进行非特异性切割的底物的核酸分子不同,所述诸如磷酸二酯酶的试剂切割核酸分子而不管二级结构(即,无需形成双链体结构)。如本文所使用的,术语“切割手段”或“切割试剂”是指能够对切割结构进行切割的任何手段,包括但不限于酶。“结构特异性核酸酶”或“结构特异性酶”是识别核酸分子中的特异性二级结构并切割这些结构的酶。本发明的切割手段响应于切割结构的形成而切割核酸分子;切割手段并不必需在切割结构内的任何特定位置处切割所述切割结构。切割手段可以包括由各种来源提供的核酸酶活性,所述来源包括CLEAVASE酶(ThirdWaveTechnologies,Madison,WI)、FEN-1核酸内切酶(包括RAD2和XPG蛋白,以及FEN-1核酸内切酶,其源自古细菌)、TaqDNA聚合酶和大肠杆菌(E.coli)DNA聚合酶I。切割手段可以包括具有5'核酸酶活性的酶(例如,TaqDNA聚合酶(DNAP)、大肠杆菌DNA聚合酶I)。切割手段还可以包括具有5'核酸酶活性但缺乏合成活性的经修饰的DNA聚合酶。适于在本发明的方法和试剂盒中使用的切割手段的实例在美国专利号5,614,402、5,795,763和5,843,669以及PCT申请号WO98/23774、WO02/070755A2和WO0190337A2中提供,所述文献中的每一个都通过提及而以其整体合并入本文。当涉及酶(例如5'核酸酶)进行使用时,术语“热稳定的”表明,该酶在升高的温度下,即在大约55℃或更高的温度(例如,包括但不限于60℃、65℃、70℃、75℃、80℃、85℃或90℃)下是有功能的或有活性的(即,可以实施催化)。如本文所使用的,术语“切割产物”是指通过切割手段与切割结构的反应(即,用切割手段处理切割结构)而产生的产物。术语“非靶切割产物”是指并非来源于靶核酸的切割反应的产物。如上文所述,在本发明的一些方法中,切割结构的切割通常在探针寡核苷酸内发生。通过这种靶核酸依赖性切割所产生的探针寡核苷酸的片段为“非靶切割产物”。在INVADER测定法反应的情况下,术语“探针寡核苷酸”是指在INVADER寡核苷酸存在或不存在下与靶核酸相互作用从而形成切割结构的寡核苷酸。当与靶核酸进行退火时,探针寡核苷酸与靶形成切割结构,并且在探针寡核苷酸内发生切割。术语“INVADER寡核苷酸”是指在探针和靶核酸之间的杂交区域附近的位置处与靶核酸杂交的寡核苷酸,其中INVADER寡核苷酸包含与在探针和靶之间的杂交区域相重叠的部分(例如,化学部分,或核苷酸,无论其是否与该靶互补)。在一些实施方案中,INVADER寡核苷酸在其3'末端包含与位于探针寡核苷酸的5'末端的序列基本上相同的序列。术语“ARRESTOR分子”是指被加入或包括在侵入性切割反应中以停止一种或多种反应组分参与到随后的作用或反应中的试剂。这可以通过下列方式来进行:隔离一些反应组分或使其失活(例如,通过与核酸组分结合或与其发生碱基配对,或者通过与蛋白质组分结合)。术语“ARRESTOR寡核苷酸”是指被包括在侵入性切割反应中以停止或阻止任何反应(例如第一反应和/或任何随后的反应或作用;不希望将ARRESTOR寡核苷酸局限于任何特定的反应或反应步骤)的一个或多个方面的寡核苷酸。这可以通过下列方式来进行:隔离一些反应组分(例如,通过与另一核酸发生碱基配对,或者与蛋白质组分结合)。但是,不希望将该术语仅局限于其中反应组分被隔离的情况。如本文所使用的,术语“盒”是指这样的寡核苷酸或寡核苷酸的组合,其被构造成在INVADER测定法中响应于探针寡核苷酸的切割而产生可检测的信号。在优选的实施方案中,所述盒与来自探针寡核苷酸的切割过程的非靶切割产物杂交,从而形成第二个侵入性切割结构,这样所述盒随后可以被切割。在本发明的一些优选的实施方案中,次级切割反应(secondarycleavagereaction)包括使用FRET盒。这样的分子提供了次级靶(次级反应靶(SecondaryReactionTarget)或SRT)以及FRET标记的可切割序列,从而允许进行相续的侵入性切割反应的均相检测(即,在反应后无需进行产物分离或其他操作)。其他优选的实施方案使用了次级反应系统,其中FRET探针和合成靶以分开的寡核苷酸形式提供。来自初级反应的经切割的5’-翼片(flap)在次级反应中用作侵入性寡核苷酸,其中它们与合适的次级反应靶(SRT)结合。在一些实施方案中,所述盒为包含发夹部分(即,这样的区域,在所述区域中所述盒寡核苷酸的一个部分在反应条件下与同一个寡核苷酸的第二个部分杂交从而形成双链体)的单个寡核苷酸。在其他实施方案中,盒包含至少两个寡核苷酸,所述寡核苷酸包含可以在反应条件下形成双链体的互补部分。在优选的实施方案中,所述盒包含标记物。在特别优选的实施方案中,所述盒包含产生荧光共振能量转移(FRET)效应的经标记的部分。当涉及核酸底物进行使用时,术语“基本上单链的”表示,底物分子主要以核酸的单链的形式存在,这不同于双链底物,其以核酸的两条链的形式存在,这两条链通过链内碱基配对作用而保持在一起。如本文所使用的,短语“未扩增的寡核苷酸检测测定法(non-amplifiedoligonucleotidedetectionassay)”是指这样的检测测定法,该检测测定法被构造成用于检测未经扩增(例如通过PCR)的特定靶序列(例如,miRNA、SNP、重复序列等)存在与否,而不形成该靶序列的拷贝。“未扩增的寡核苷酸检测测定法”可以例如放大用于指示特定靶序列或者靶序列内的多态性存在与否的信号,只要该靶序列没有被拷贝。如本文所使用的,短语“非扩增性寡核苷酸检测测定法(non-amplifyingoligonucleotidedetectionassay)”是指这样的检测测定法,该检测测定法被构造成用于检测靶序列(例如,miRNA、SNP、重复序列等)存在与否,而不形成该靶序列的拷贝。“非扩增性寡核苷酸检测测定法”可以例如放大用于指示特定靶序列或者靶序列中的多态性存在与否的信号,只要该靶序列没有被拷贝。如本文所使用的,术语“序列变异(sequencevariation)”是指两个核酸之间在核酸序列方面的差异。例如,野生型结构基因与该野生型结构基因的突变体形式可以通过存在单个碱基置换和/或一个或多个核苷酸的缺失或插入而在序列方面发生变化。据信,这两种形式的结构基因在序列方面彼此不同。可以存在结构基因的第二种突变体形式。据信,该第二种突变体形式在序列方面不同于野生型基因和该基因的第一种突变体形式。如本文所使用的,术语“释出”是指通过例如5'核酸酶的作用从更大的核酸片段(例如寡核苷酸)中释放出核酸片段,从而使得所释放的片段不再与所述寡核苷酸的其余部分共价连接。如本文所使用的,术语“Km”是指酶的米-曼(Michaelis-Menten)常数,其被定义为在酶催化的反应中在给定的酶达到其最大速度的一半时特异底物的浓度。如本文所使用的,术语“核苷酸类似物”是指经修饰的或非天然存在的核苷酸,其包括但不限于:具有改变的堆叠相互作用的类似物,例如7-脱氮嘌呤(即,7-脱氮-dATP和7-脱氮-dGTP);具有备选的氢键键合构型的碱基类似物(例如,Iso-C和Iso-G,以及在属于S.Benner的美国专利号6,001,983中所描述的其他非标准碱基对,该文献通过提及而合并入本文);非氢键键合类似物(例如非极性的芳香族核苷酸类似物,如2,4-二氟甲苯,其在B.A.Schweitzer和E.T.Kool,J.Org.Chem.,1994,59,7238-7242,B.A.Schweitzer和E.T.Kool,J.Am.Chem.Soc.,1995,117,1863-1872中有所描述,所述文献中的每一个均通过提及而合并入本文);“通用(universal)”碱基,例如5-硝基吲哚和3-硝基吡咯;以及通用嘌呤和嘧啶(例如,分别为“K”和“P”核苷酸;P.Kong等人,NucleicAcidsRes.,1989,17,10373-10383,P.Kong等人,NucleicAcidsRes.,1992,20,5149-5152)。核苷酸类似物包括在糖部分具有修饰的核苷酸,例如二脱氧核苷酸和2'-O-甲基核苷酸。核苷酸类似物包括脱氧核糖核苷酸以及核糖核苷酸的修饰形式。术语“多态基因座”是在种群中存在的基因座,其在该种群的成员之间显示出变异(例如,最普遍的等位基因具有小于0.95的频率)。相反地,“单态基因座”是在种群的成员之间具有很少或没有可见的变异的基因座位(通常为这样的基因座,在该基因座处,在该种群的基因库中,最普遍的等位基因超过0.95的频率)。如本文所使用的,术语“微生物”是指太小以致不能用肉眼观察到的生物体,其包括但不限于细菌、病毒、原生动物、真菌和纤毛虫。术语“微生物基因序列”是指来源于微生物的基因序列。术语“细菌”是指任何细菌物种,包括真细菌和古细菌物种。术语“病毒”是指不能自主复制的(即,复制过程需要利用宿主细胞的机构)、专性的、超显微的细胞内寄生物。在本说明书和权利要求书中,术语“样品”以其最广泛的意义进行使用。一方面其表示包括样本或培养物(例如,微生物培养物)。另一方面,其表示包括生物和环境样品。样品可以包括合成来源的样本。生物样品可以为:动物,包括人,流体,固体(例如粪便)或组织,以及液体和固体食物和饲料产品和配料例如奶制品、蔬菜、肉和肉的副产品,和废物。生物样品可以获自所有的各科家畜以及未驯化的或野生的动物(包括但不限于,诸如有蹄类动物、熊、鱼、兔类动物、啮齿动物等的动物)。环境样品包括:环境材料,例如表面物质、土壤、水和工业样品,以及从食物和奶制品加工设备、装置、器材、器具、一次性和非一次性制品中获得的样品。这些实例不应被解释为限制了可用于本发明的样品类型。术语“靶核酸的来源”是指含有核酸(RNA(例如miRNA)或DNA)的任何样品。靶核酸的特别优选的来源为生物样品,包括但不限于血液、唾液、脑脊液、胸膜液、乳、淋巴、痰和精液。如果一种寡核苷酸以比另一种寡核苷酸(或靶核酸序列)更高的摩尔浓度存在,则认为这种寡核苷酸相对于另一种寡核苷酸(或靶核酸序列)而言“过量”存在。当寡核苷酸例如探针寡核苷酸在切割反应中相对于互补靶核酸序列的浓度而言过量存在时,则该反应可以用于指示所存在的靶核酸的量。通常,当过量存在时,探针寡核苷酸将以至少100倍摩尔过量存在;当靶核酸序列以大约10飞摩尔或更少的量存在时,所使用的每种探针寡核苷酸的量通常为至少1皮摩尔。“怀疑包含”第一种和第二种靶核酸的样品,可以包含这两种靶核酸分子之一,或者包含这两种靶核酸分子,或者这两种靶核酸分子都不包含。术语“反应物”在本文中以其最广泛的意义进行使用。反应物可以包括例如酶反应物、化学反应物或光(例如紫外光,已知波长特别短的紫外光可以打断寡核苷酸链)。能够与寡核苷酸反应从而缩短(例如切割)或延长该寡核苷酸的任何试剂都被涵盖在术语“反应物”之内。如本文所使用的,术语“纯化的”或“纯化”是指从样品中除去污染物。例如,在一些实施方案中,在细菌宿主细胞中表达重组CLEAVASE核酸酶,并通过除去宿主细胞蛋白质而纯化出所述核酸酶;由此增加了这些重组核酸酶在所述样品中的百分比。如本文所使用的,当涉及蛋白质(如在“给定的蛋白的部分”中)时,术语“部分”是指该蛋白质的片段。所述片段可在大小上在从4个氨基酸残基至整个氨基酸序列减去一个氨基酸的范围内变化(例如,4、5、6、……、n-1)。如本文所使用的,术语“核酸序列”是指寡核苷酸、核苷酸或多核苷酸以及它们的片段或部分,以及是指基因组或合成来源的DNA或RNA,它们可以是单链或双链的,并且代表了正义或反义链。类似地,如本文所使用的,“氨基酸序列”是指肽或蛋白质序列。如本文所使用的,术语“纯化的”或“基本上纯化的“是指这样的分子(核酸或氨基酸序列),它们被从其天然环境中取出、分离或分开,并且至少60%地不含、优选地75%地不含和最优选地90%地不含天然地与其相联合的其他组分。因此,“分离的多核苷酸”或“分离的寡核苷酸”是基本上纯化的多核苷酸。如本文所使用的,术语“连续的核酸链”是指具有连续的、共价连接的主链结构,但没有切口或其他中断的核酸链。无论是碱基配对的、单链的还是错配的,每个核苷酸的碱基部分的布置并不是在连续链的定义中的要素。连续链的主链不限于在天然存在的、未修饰的核酸中发现的核糖-磷酸或脱氧核糖-磷酸组成。本发明的核酸可以包含在主链结构中的修饰,包括但不限于硫代磷酸酯残基、膦酸酯残基、2'取代的核糖残基(例如,2'-O-甲基核糖)以及含有备选的糖(例如阿拉伯糖)的残基。如本文所使用的,术语“连续的双链体”是指这样的双链核酸的区域,在该区域中在双链体内的碱基对进程没有中断(即,在连续的双链体的区域范围内,沿着该双链体的碱基对没有被扭曲而提供了缺口、凸起或错配)。如本文所使用的,该术语仅是指双链体内的碱基对排列,而不涉及核酸链的主链部分中的连续性。具有未间断的碱基配对,但在一条或两条链中具有切口的双链体核酸也在“连续的双链体”的定义范围之内。术语“双链体”是指核酸的状态,在该状态下,一条链上的核苷酸的碱基部分通过与排列在第二条链上的其互补碱基发生氢键键合而被结合。形成双链体形式的条件反映了核酸的碱基的状态。借助于碱基配对,核酸链还经常呈现出双螺旋的三级结构,该三级结构具有大沟和小沟。螺旋形式的呈现在变成双链体的行为中是固有的。术语“模板”是指这样的核酸链,在该核酸链上,通过模板依赖性核酸聚合酶的活性从核苷三磷酸形成互补的拷贝。在双链体内,模板链按照惯例被描绘和描述为“底部”链。类似地,非模板链常常被描绘和描述为“顶部”链。发明详述本发明涉及用于检测和表征核酸分子(例如,RNA(例如,小RNA例如微RNA(miRNA)和小干扰RNA(siRNA))和其他短核酸分子)的组合物和方法。本发明提供了用于检测、表征和定量miRNA表达的方法。在一些实施方案中,本发明提供了用于检测miRNA表达的方法,该方法包括将核酸加入到miRNA中以帮助检测。然后,采用任何合适的方法来检测所得到的“miRNA检测结构”,所述方法包括但不限于在本文中公开的那些。尽管以下描述集中于miRNA的检测和定量,但是应该理解,本发明还可用于其他短核酸分子(例如,长度小于例如50、40、30或20个核苷酸的DNA和RNA)。下文使用miRNA作为例子来举例说明各种实施方案。但是,应该理解,所述方法可应用于其他小核酸分子。I.miRNA检测结构的形成在一些实施方案中,本发明提供了产生miRNA检测结构以帮助检测miRNA的方法。miRNA的大小是小的(大约21个核苷酸(例如大约18-25个核苷酸)),因此miRNA难以采用标准化的杂交方法来进行检测。在一些实施方案中,本发明的方法包括将核酸分子加入到miRNA(例如,通过杂交、延伸或连接)中以产生检测结构。然后,可以采用任何合适的方法来检测这样的检测结构。在一个具体的实施方案中,产生图2中所示的检测结构以用于检测miRNA。在该实施方案中,两个寡核苷酸与miRNA退火,从而形成双环或“哑铃”状结构。通过延伸具有寡核苷酸双链区域的miRNA的末端,该哑铃结构形成更大区域的双链核酸。在一些实施方案中,将miRNA的每个末端延伸2至5个核苷酸。在一些实施方案中,所述寡核苷酸的末端包含不与miRNA杂交的额外的核酸序列。在一些实施方案中,这些额外的序列形成侵入性切割结构(例如,INVADER测定法侵入性切割结构)。在一些实施方案中,侵入性切割结构通过INVADER测定法来进行检测(参见,例如以下描述)。例如,在一些实施方案中,将在实施例18中所描述的寡核苷酸(参见,例如图31)用于检测与癌症相关的miRNA(例如,其形成可以通过INVADER测定法来检测的侵入性切割结构)。在其他实施方案中,产生图3中所示的检测结构以用于检测miRNA。在该实施方案中,一个寡核苷酸与miRNA退火,从而形成弓形结构。miRNA使寡核苷酸的末端靠在一起,其效率比当miRNA不存在时更高。在一些实施方案中,寡核苷酸的末端包含额外的序列,该序列延伸到miRNA外,并且不与miRNA杂交。在一些实施方案中,这些额外的序列形成侵入性切割结构(例如,INVADER测定法侵入性切割结构)。在一些实施方案中,侵入性切割结构通过INVADER测定法来进行检测(参见,例如以下描述)。在其他实施方案中,在切割INVADER测定法侵入性切割结构之后,使所得到的末端进行连接,从而形成环状结构。在其他实施方案中,一个寡核苷酸与miRNA杂交,使得该寡核苷酸的末端紧密接近(例如,与miRNA的邻近核苷酸杂交),然后进行连接。在其他实施方案中,产生图24和25中所示的检测结构。在该实施方案中,延伸探针或INVADER寡核苷酸,以产生单个发夹环或“半哑铃”结构。例如,在一些实施方案中,将在实施例18中所描述的寡核苷酸(参见,例如图31)用于检测与癌症相关的miRNA(例如,其形成可以通过INVADER测定法来检测的侵入性切割结构)。在一些实施方案中,所述寡核苷酸的末端包含不与miRNA杂交的额外的核酸序列(参见,例如实施例19G和19H)。在一些实施方案中,这些额外的序列形成侵入性切割结构(例如,INVADER测定法侵入性切割结构)。在一些实施方案中,侵入性切割结构通过INVADER测定法来进行检测(参见,例如以下描述)。在其他实施方案中,这些额外的序列与被加入到反应混合物中的另外的寡核苷酸是互补的,从而稳定切割结构,例如INVADER测定法侵入性切割结构(图4)。在一些实施方案中,使用滚环复制测定法来检测如上文所述产生的环状结构(参见,例如以下关于滚环复制的描述)。在其他实施方案中,由带有与miRNA具有同源性的短区域的长寡核苷酸(例如,多于50、100、1000个或更多个核苷酸)产生检测结构。使一个或多个miRNA与所述寡核苷酸杂交,从而产生检测结构。在一些实施方案中,这些检测结构通过延伸(例如,通过连接或聚合反应,如RT-PCR)miRNA来进行检测。在一些实施方案中,这些检测结构进一步通过与缀合至固体支持物(例如,微球体,或者其他表面或结构)的寡核苷酸杂交来进行检测。在一些实施方案中,延伸非miRNA组分,或者将其连接至另一核酸,并直接或间接进行检测。在一些实施方案中,用于形成检测结构的寡核苷酸包含一个或多个核苷酸类似物。例如,在一些实施方案中,使用了2'-O-甲基核苷酸。本发明不限于特定的机制。事实上,理解所述机制不是实施本发明所必需的。但是,考虑了2'-O-甲基碱基的存在增加了经杂交的检测结构的稳定性并为进一步的检测方法提供帮助。II.核酸(例如,干扰RNA)的检测在一些实施方案中,本发明提供了用于检测miRNA的方法。本发明不限于具体的检测测定法。可以使用任何合适的方法,包括但不限于本文所公开的那些。在本发明一些优选的实施方案中,miRNA检测方法是定量的。本发明不限于特定的机制。事实上,理解所述机制不是实施本发明所必需的。但是,考虑了特定miRNA在体内的水平与来自其同族(cognate)基因的基因表达水平有关。因此,本发明提供了使miRNA与特定基因(例如,参与疾病状态(例如癌症)或代谢的基因)的基因表达相关联的方法。例如,在一些实施方案中,本发明的方法用于确定异常(例如,高或低)水平的特定miRNA的存在(例如,与癌症有关的miRNA表达(参见,例如Calin等人,ProcNatlAcadSciUSA,99,15524-15529(2002)),例如通过使用实施例18和图31中所描述的寡核苷酸),或者确定对于miRNA表达的干预(例如,药物)的效果。在其他实施方案中,检测异源miRNA(例如,来自表达载体、转基因构建体、转染等)以表征miRNA表达系统的效能。在一些实施方案中,本发明提供了用于检测特定miRNA(例如,miRNA,如mir-1或mir-135)的方法。在其他实施方案中,使用本发明的方法来区别特定miRNA中的变体(例如,多态性或突变)。在其他实施方案中,本发明提供了裂解待就miRNA的存在进行测试的细胞。A.INVADER测定法在一些实施方案中,采用INVADER测定法来检测miRNA。在一些实施方案中,INVADER测定法包括:形成核酸切割结构,该结构依赖于靶核酸的存在;以及切割所述核酸切割结构以释放独特的切割产物。例如,利用5'核酸酶活性来切割靶依赖性切割结构,并且所得到的切割产物或者所述切割结构的切割指示了样品中特定靶核酸序列的存在。当核酸或寡核苷酸的一条或两条(或更多条)链都与靶核酸杂交从而使得它们形成重叠的侵入性切割结构(如下文所述)时,可以发生侵入性切割。通过切割试剂(例如,5'核酸酶)与上游寡核苷酸的相互作用,可以使得切割试剂以产生独特片段的方式在内部位点处切割下游寡核苷酸。这样的实施方案被称为INVADER测定法(ThirdWaveTechnologies),并且描述于美国专利号5,846,717、5,985,557、5,994,069、6,001,567、6,090,543、6,348,314和6,458,535,WO97/27214,WO98/42873,以及Lyamichev等人,Nat.Biotech.,17:292(1999),Hall等人,PNAS,USA,97:8272(2000)中,每篇文献通过提及而以其整体合并入本文以用于所有目的)。INVADER测定法通过用结构特异性酶对特异性结构进行酶切来检测探针与靶的杂交(参见,INVADER测定法,ThirdWaveTechnologies;参见例如美国专利号5,846,717;6,090,543;6,001,567;5,985,557;5,994,069;6,090,543;6,348,314;6,458,535;美国专利申请号20030186238(系列号10/084839);20030104378A1(系列号09/864636);Lyamichev等人,Nat.Biotech.,17:292(1999);Hall等人,PNAS,USA,97:8272(2000);WO97/27214;和WO98/42873,所述文献中的每一个都通过提及而以其整体合并入本文以用于所有目的)。INVADER测定法通过下列方式来检测特异性的DNA和RNA序列:使用结构特异性酶(例如,FEN核酸内切酶)来切割通过重叠的寡核苷酸探针的杂交而形成的复合物(参见,例如图1)。升高的温度以及过量的所述探针之一使得对于每种靶序列能够存在多个待切割的探针而无需温度循环。在一些实施方案中,这些被切割的探针随后指导第二中经标记的探针的切割。可以使用被内部染料猝灭的荧光素对次级探针(secondaryprobe)寡核苷酸的5’-末端进行标记。在切割之后,可以使用标准荧光平板读取器来检测去猝灭的经荧光素标记的产物。INVADER测定法检测在未扩增的以及扩增的(参见,例如实施例19,图32)RNA和DNA(包括基因组DNA)中的特异性序列、突变和SNP。在图1中示意性地显示的实施方案中,INVADER测定法采用了两个级联步骤(初级(primary)和次级(secondary)反应),这两个反应步骤都产生并随后扩增靶特异性的信号。为了方便起见,在下面的讨论中,将等位基因描述为野生型(WT)和突变体(MT),尽管该术语并不应用于所有的基因变异。在初级反应中(图1,图版A),WT初级探针和INVADER寡核苷酸以串联方式与靶核酸杂交,从而形成重叠结构。未配对的“翼片”被包括在WT初级探针的5'末端上。结构特异性酶(例如CLEAVASE酶,ThirdWaveTechnologies)识别该重叠区,并切掉未配对的翼片,从而将翼片作为靶特异性产物释放出来。在次级反应中,这种切割的产物用作在WT荧光共振能量转移(WT-FRET)探针上的INVADER寡核苷酸,从而再次形成被结构特异性酶所识别的结构(图版A)。当在单个FRET探针上的两种染料通过切割而分开时(如在图1中由箭头所示),在上述背景荧光之上产生了可检测的荧光信号。因此,该次级结构的切割导致信号增强,由此表明了WT等位基因(或者突变体基因,如果所述测定法被构造成对于突变体等位基因而产生可检测的信号)的存在。在一些实施方案中,对于待检测的每个等位基因或基因座,提供了具有不同标记物(例如,可通过发射或激发波长的差异来分辨的,或者可通过时间分辨荧光检测(time-resolvedfluorescencedetection)来分辨的)的FRET探针,从而使得可以在单个反应中检测不同的等位基因或基因座。在这样的实施方案中,初级探针组和不同的FRET探针可以组合在在单个测定法中,从而允许比较同一个样品中来自每个等位基因或基因座的信号。如果初级探针寡核苷酸和靶核苷酸序列在切割位点处不完全匹配(例如,如使用MT初级探针和WT靶,图1,图版B),则不形成重叠的结构,并且切割受到抑制。与非重叠的结构相比较,所使用的结构特异性酶(例如,CLEAVASEVIII酶,ThirdWaveTechnologies)更高效地切割重叠的结构(例如,至少340倍),从而允许极好地区分等位基因。探针无需温度循环地进行周转,从而产生许多信号/靶(即,线性信号扩增)。类似地,每个靶特异性产物可以使得能够对许多FRET探针进行切割。初级INVADER测定法反应针对所检测的靶DNA(或RNA)。在第一个侵入性切割中,靶DNA为限制性组分,这是因为INVADER和初级探针是以摩尔过量来提供的。在第二个侵入性切割中,释放的翼片为限制性的。当顺次进行这两个切割反应时,来自该复合反应的荧光信号相对于靶DNA的量而言线性地积累。在某些实施方案中,使用经可接近位点设计的寡核苷酸和/或桥接寡核苷酸来施行INVADER测定法或其他核苷酸检测测定法。此类方法、操作程序和组合物描述于美国专利号6,194,149、6,358,691、6355,437,美国专利申请系列号09/882,945,以及PCT申请WO9850403和WO0198537中,所有这些文献均通过提及而明确地以其整体合并入本文。在一些优选的实施方案中,将样品(例如,核酸序列(例如,干扰RNA(例如,miRNA或siRNA)))暴露于所述寡核苷酸和所述试剂包括将样品在以下条件下将样品暴露于所述寡核苷酸和所述试剂:如果所述样品中存在靶序列,则在所述靶序列和所述寡核苷酸之间形成侵入性切割结构,其中所述侵入性切割结构被所述切割试剂切割,从而形成切割产物。在一些实施方案中,所述靶序列(例如,miRNA)包含第一区域和第二区域,其中所述第二区域位于所述第一区域的下游并与之邻接;并且所述寡核苷酸包括第一种寡核苷酸和第二种寡核苷酸,其中所述第一种寡核苷酸的至少一部分与所述靶序列的所述第一部分完全互补,并且其中所述第二种寡核苷酸包含3'部分和5'部分,其中所述5'部分与所述靶核酸的所述第二部分完全互补。在一些优选的实施方案中,将样品暴露于所述寡核苷酸和所述试剂包括将样品在以下条件下将样品暴露于所述寡核苷酸和所述试剂:如果所述样品中存在靶序列,则在所述靶序列和所述寡核苷酸之间形成侵入性切割结构,其中所述侵入性切割结构被所述切割试剂切割,从而形成切割产物。在一些特别优选的实施方案中,所述靶序列包含第一区域和第二区域,其中所述第二区域位于所述第一区域的下游并与之邻接;并且所述寡核苷酸包括第一种寡核苷酸和第二种寡核苷酸,其中所述第一种寡核苷酸的至少一部分与所述靶序列的所述第一部分完全互补,并且其中所述第二种寡核苷酸包含3'部分和5'部分,其中所述5'部分与所述靶核酸的所述第二部分完全互补。在某些实施方案中,本发明提供了用于使用INVADER检测试剂(例如,初级探针、INVADER探针和FRET盒)来分析汇集的样品(例如,汇集的血液样品或汇集的细胞裂解物)的试剂盒。在优选的实施方案中,所述试剂盒进一步包含关于如何施行INVADER检测测定法的说明书,并且在一些实施方案中,所述试剂盒进一步包含关于如何将INVADER检测测定法应用于来自许多个体的汇集的样品或者应用于来自单个受试者的许多细胞(例如,来自活组织检查样品)的“汇集的”样品的说明书。本发明进一步提供了测定法,其中在多轮与寡核苷酸探针杂交并切割探针的过程中靶核酸被再使用或再利用,而无需使用温度循环(即,用于靶核酸链的周期的变性)或核酸合成(即,用于靶或探针核酸链的基于聚合的置换)。当在靶链上探针被连续地替代(例如,通过探针-探针置换,或者通过在探针/靶的缔合和解离之间的平衡,或者通过包括这些机制的组合(Reynaldo等人,J.Mol.Biol.97:511-520(2000)))的条件下运行切割反应时,多个探针可以与同一个靶杂交,从而允许多次切割并产生多个切割产物。INVADER测定法反应在INVADERDNA测定法的优选的实施方案中,两种寡核苷酸(有区分能力的初级探针和INVADER寡核苷酸)以串联方式与靶DNA杂交,从而形成重叠结构。初级探针的5'-末端包含不与靶DNA杂交的5'-翼片(图1)。结合的INVADER寡核苷酸的3'-核苷酸与初级探针重叠,但不必与靶DNA杂交(参见,例如实施例15和16)。CLEAVASE酶识别该重叠结构并切掉初级探针的未配对的5'-翼片,从而将其作为靶特异性产物释放出来。初级探针被设计成具有与反应温度接近的解链温度。因此,在等温测定法条件下,以过量方式提供的初级探针在靶DNA上循环。这使得对于每个靶DNA可以进行多轮的初级探针切割,并且扩增被释放的5'-翼片的数目。在次级反应中,酶个被释放的5'-翼片可以在荧光共振能量转移(FRET)盒上用作INVADER寡核苷酸,从而形成另一个被CLEAVASE酶识别和切割的重叠结构(图1)。当FRET盒被切割时,荧光团(F)与猝灭剂(Q)被分开,从而产生可检测的荧光信号。类似于初始反应,被释放的5'-翼片和FRET盒进行循环,从而导致扩增的荧光信号。初始反应和次级反应在同一个孔中同时进行。双重形式的INVADERDNA测定法使得能够在单个孔中同时检测两种DNA序列(参见,例如实施例17和19(L))。最通常地,这涉及检测特定多态性的两种变体(例如,在miRNA中)。所述双重形式使用两种不同的有区分能力的初级探针,每种探针具有独特的5'-翼片;和两种不同的FRET盒,每种FRET盒具有在光谱上不同的荧光团。通过设计,被释放的5'-翼片将只与它们各自的FRET盒结合,从而产生靶特异性信号。在一些实施方案中,本发明提供了试剂盒,其包含对于实施本发明而言所必需的组分中的一种或多种。例如,本发明提供了用于贮存或递送对于实施切割测定法(例如,INVADER测定法)而言所必需的本发明的酶和/或反应组分的试剂盒。通过举例的方式而无意于将本发明的试剂盒限定于任何特定的配置或组分组合,以下部分描述了用于实施本发明的试剂盒的一个实施方案:在一些实施方案中,本发明的试剂盒提供了以下试剂:CLEAVASE酶初级探针OligosDNA反应缓冲液1INVADEROligoFRET盒1(例如,F)FRET盒2(例如,R)突变体DNA对照野生型DNA对照"非靶"空白对照在其他实施方案中,本发明的试剂盒被构造成用于直接检测RNA。这些试剂盒可以提供以下试剂:CLEAVASE酶初级探针寡核苷酸DNA反应缓冲液1INVADEROligoFRET探针1(例如,F)FRET探针2(例如,R)次级反应靶1次级反应靶2ARRESTOR寡核苷酸1ARRESTOR寡核苷酸2突变体DNA对照野生型DNA对照"非靶"空白对照必须额外考虑,在单个反应中,由于寡核苷酸的特定组合而导致的不希望的效应。一个此类效应是靶依赖性地产生背景信号。在INVADER测定法中,某些寡核苷酸与其他寡核苷酸相组合可能会在不存在待检测的特定靶的情况下产生信号。将这些寡核苷酸组合分开成不同的汇集物可以用于减轻这种效应。类似地,某些寡核苷酸组合可以人为地压制来自所希望的靶的信号产生。再次地,将这些寡核苷酸组合分开成不同的汇集物可以用于减轻这种效应。采用本文所提供的关于反应设计和最佳化的指导方针(参见,例如试验部分),对探针组(例如,寡核苷酸和/或它们的序列)的设计进行调整以适合于在miRNA检测测定法中使用。例如,在一些实施方案中,降低反应温度(例如,至50-60℃),以解决较小的杂交区域。在一些实施方案中,本发明的试剂盒提供了由用户提供的为了实施本发明方法的额外组分(例如,试剂、供给品和/或设备)的列表。例如,并且无意于将这些额外组分的列表限定于任何特定的组分,此类列表的一个实施方案包括以下方面:·澄清的CHILLOUT-14液体蜡(MJResearch)或者不含RNA酶的光学级矿物油(Sigma,Cat.No.M-5904)·96孔聚丙烯微量培养板(MJResearch,Cat.No.MSP-9601)·无菌的1.5-ml或2.0-ml微量离心管·无菌的不含DNA酶/RNA酶的一次性气溶胶屏障移液器枪头(aerosolbarrierpipettip)·多道移液器(0.5-10μl,2.5-20μl)·热循环仪或其他热源(例如,实验室烘箱或加热器)·其他实验室设备(试管架、微量移液器、多道吸移器、微量离心机、漩涡混合器)·荧光微量培养板读取器,优选的微量培养板读取器为上方读取的,并且装备有光滤波器,该滤波器具有以下特征:激发(波长/带宽)发射(波长/带宽)485nm/20nm530nm/25nm560nm/20nm620nm/40nm在一些实施方案中,本发明的试剂盒提供了由用户提供的有助于实施本发明方法的可选组分(例如,试剂、供给品和/或设备)的列表。例如,并且无意于将这些可选组分的列表限定于任何特定的组分,此类列表的一个实施方案包括以下方面:·无菌的8联管(8-tubestrip)或微量培养板(可选的)·一次性塑料槽(可选的)·平板密封带(可选的)。在一些实施方案中,本发明的试剂盒提供了由用户提供的有助于实施本发明方法的所需成分的列表(对此,多种备选形式是可接受的(例如,样品制备试剂盒))。例如,并且无意于将这些可选组分的列表限定于任何特定的组分,此类列表的一个实施方案包括以下方面:·QIAGENQIAAMPBloodKit·GentraSystemsPUREGENEKit·GentraSystemsGENERATIONProducts。在试剂盒的一些实施方案中,提供了详细的操作方案。在优选的实施方案中,提供了关于INVADER测定法反应的装配(例如,制剂以及用于制备反应混合物的优选操作程序)的操作方案。在特别优选的实施方案中,关于反应混合物的装配的操作方案包括计算或绘图辅助,以减小在实施本发明方法中的错误风险(例如,表格,以有助于计算对于多个反应而言所需的试剂的体积;以及平板布局指导,从而帮助构造多孔测定法平板以包含许多测定法反应)。在一些实施方案中,提供了补充文件资料,例如用于辅助操作程序(例如,用于制备额外的试剂,或用于制备在本发明的方法中使用的样品)的操作方案。在优选的实施方案中,补充文件资料包括指导和注意事项列表,它们被提供用于有助于未经训练的或没有经验的用户能够成功地使用所述方法和试剂盒。在特别优选的实施方案中,补充文件资料包括发现并解决问题的指导,例如描述了用户可能会遇到的可能的问题、并且提供了旨在帮助用户解决或避免此类问题的建议性的解决方案或改正方法的指导。在优选的实施方案中,将样品稀释至相当于10-μl添加/反应的浓度。100-ng样品的浓度应该为15ng/μl。B.滚环复制(RollingCircleReplication)在其他实施方案中,利用滚环复制方法(AmershamBiosciences,Piscataway,NJ)来检测miRNA检测结构(参见,例如美国专利号6,344,329;6,143,495;6,316,229;6,210,884;6,183,960和6,235,502,所述文献中的每一个都通过提及而以其整体合并入本文)。在一些实施方案中,采用滚环复制来检测由与miRNA退火的单个寡核苷酸的末端的退火而产生的环状miRNA检测结构。在一些实施方案中,所述寡核苷酸的末端与没有重叠的miRNA杂交。这种寡核苷酸可以在miRNA存在或不存在下进行连接。但是,在miRNA存在下,连接反应更有效。在此类实施方案中,将在一定时间内检测到的环状分子的水平与缺少miRNA的对照反应相比较。在其他实施方案中,所述寡核苷酸的末端与具有重叠末端的miRNA杂交,从而形成侵入性切割结构。此类结构在连接之前被切割,由此提高了产生环状检测结构的特异性。滚环扩增(rollingcircleamplification,RCA)涉及环状单链DNA分子的复制。在RCA中,滚环复制引物与环状核酸分子杂交,然后使用链置换DNA聚合酶(strand-displacingDNApolymerase)来进行核酸分子的滚环复制。在单个反应循环中,在滚环复制过程中发生扩增。滚环复制导致形成大的DNA分子,其包含串联重复的核酸序列。这种DNA分子被称为串联序列DNA(tandemsequenceDNA,TS-DNA)。在一些实施方案中,采用连接介导的滚环扩增(ligation-mediatedrollingcircleamplification,LM-RCA),其涉及在复制之前的连接操作。在所述连接操作中,探针与其同族靶核酸序列(如果存在的话)杂交,然后经杂交的探针的末端进行连接,从而形成共价闭合的单链核酸。在连接后,滚环复制引物与探针分子杂交,然后使用链置换DNA聚合酶来进行环状分子的滚环复制。一般地,LM-RCA方法包括:将开放的环状探针与靶样品相混合,导致形成探针-靶样品混合物;并在促进开放的环状探针与靶序列杂交的条件下温育所述探针-靶样品混合物;将连接酶与所探针-靶样品混合物相混合;导致形成连接混合物,并在促进开放的环状探针进行连接从而形成扩增靶环(amplificationtargetcircle,ATC)的条件下温育所述连接混合物;将滚环复制引物(rollingcirclereplicationprimer,RCRP)与所述连接混合物相混合;导致形成引物-ATC混合物,并在促进扩增靶环与滚环复制引物之间的杂交的条件下温育所述引物-ATC混合物;将DNA聚合酶与所述引物-ATC混合物相混合;导致形成聚合酶-ATC混合物,并在促进扩增靶环复制的条件下温育所述聚合酶-ATC混合物,其中扩增靶环的复制导致形成串联序列DNA(TS-DNA)。C.另外的检测方法本发明不限于INVADER测定法或滚环测定法检测。可以使用任何允许检测miRNA检测结构的方法。下文描述了可在本发明的方法中使用的示例性的、非限定性的检测测定法。1.杂交测定法在本发明的一些实施方案中,采用杂交测定法来检测检测结构。在杂交测定法中,基于来自样品的DNA与互补DNA分子(例如,寡核苷酸探针)杂交的能力来确定给定的核酸序列的存在与否。使用了各种关于杂交和检测的技术的各种杂交测定法是可用的。下面提供了测定法的选择的描述。a.使用“DNA芯片”测定法进行的杂交的检测在本发明的一些实施方案中,使用DNA芯片杂交测定法来检测序列。在该测定法中,将一系列寡核苷酸探针固定于固体支持物上。将寡核苷酸探针设计成对于给定的靶序列(例如,检测复合物的组分)是独特的。将目的样品与DNA“芯片”接触并检测杂交。在一些实施方案中,所述DNA芯片测定法是GeneChip(Affymetrix,SantaClara,CA;参见,例如美国专利号6,045,996、5,925,525和5,858,659;所述文献中的每一个都通过提及而合并入本文)测定法。GeneChip技术使用小型化、高密度的固定在“芯片”上的寡核苷酸探针阵列。通过Affymetrix的光引导化学合成(light-directedchemicalsynthesis)方法来制备探针阵列,该方法将固相化学合成与在半导体工业中所使用的光刻蚀制造技术(photolithographicfabricationtechniques)相结合。通过使用一系列光刻蚀掩膜来确定芯片暴露位点,然后进行特异的化学合成步骤,该方法构建出高密度的寡核苷酸阵列,其中每个探针位于阵列中预先确定的位置中。在大的玻璃薄片上同时合成多个探针阵列。然后将薄片切成小片,并将独个的探针阵列包装入注塑成型的塑料吸纳盒(cartridge)中,该吸纳盒保护探针阵列免受环境的破坏和用作杂交室。分离待分析的核酸,通过PCR进行扩增,并用荧光报告基团进行标记。然后使用全自动洗涤工作站(fluidicsstation),将经标记的DNA与该阵列一起进行温育。然后将阵列插入扫描仪,在扫描仪中检测杂交图式。作为从已掺入靶的荧光报告基团发射出的光,收集杂交数据,所述靶已被结合至该探针阵列。完全匹配靶的探针通常产生比具有错配的探针更强的信号。因为阵列上每个探针的序列和位置是已知的,所以通过互补性,可确定施加于探针阵列上的靶核酸的身份。在其他实施方案中,使用包含电子捕获探针(electronicallycapturedprobes)(Nanogen,SanDiego,CA)的DNA微芯片(参见,例如美国专利号6,017,696、6,068,818和6,051,380;所述文献中的每一个都通过提及而合并入本文)。通过使用微电子学,Nanogen的技术使得带电分子能够在其半导体微芯片上主动运动和浓集至指定的测试位点和从指定的测试位点移开。将对于给定的靶序列来说独特的DNA捕获探针通过电子作用置于或“送”至微芯片上的特定位点。因为DNA具有强负电荷,所以其可通过电子作用移动至正电荷区域。首先,用正电荷电子地激活微芯片上的测试位点或成排的测试位点。然后,将含有DNA探针的溶液导至微芯片上。带负电荷的探针快速地移动至带正电荷的位点,在那里它们浓集并化学地结合至微芯片上的位点。然后洗涤该微芯片并加入另一种不同DNA探针的溶液直至完成经特异性结合的DNA探针的阵列。然后通过确定哪些DNA捕获探针与靶序列杂交,而就靶序列的存在来分析受试样品。电子电荷也用于将靶分子移动和浓集至微芯片上的一个或多个测试位点。样品DNA在每个测试位点处的电子浓集促进了样品DNA与互补的捕获探针的快速杂交(杂交可在数分钟内发生)。为了从每个位点除...
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1