一种植物瞬时表达植物DNA错配识别的核酸内切酶的方法与流程
本发明涉及一种植物瞬时表达植物DNA错配识别的核酸内切酶的方法,属于生物技术领域。
背景技术:
植物DNA错配识别的核酸内切酶(Recognizing DNA Mismatch Specific endonucleases in plants)是一种单链、双链都特异识别的特异性核酸酶(Pimkin et al,2007),mung bean nuclease、SP1、S1、P1、ZEN1、ZEN2、CEL I、CEL II、M1、BENI、T7Endonuclease I、Endo V或T4Endonuclease VII等具有同源性(图7和图8)的核酸内切酶都属于植物DNA错配识别的核酸内切酶(Till et al,2004;Pimkin et al,2007)。传统理论认为该酶的催化活性只需要Mg2+和Zn2+(Till et al,2004),它可以识别八种错配形式的碱基(mismatches)分别为AA、AG、AC、GG、GT、CC、CT和TT(Oleykowski et al,1999;Oleykowski et al,1998),因此被广泛应用于突变检测领域,如植物TILLING技术平台中(Colbert et al,2001;Till et al,2006)。它可以特异性切割错配碱基(Yang et al,1999)。通过酶切扩增的目标检测片段,并经琼脂糖或毛细管电泳分离检测(Jiang et al,2013),一般可方便获得突变位置和大小。因此,它是一种比较方便和经济的检测未知和已知突变位点的手段(Yeung et al,2005)。
目前CELI酶主要提取途径是从芹菜中提取,主要提取方法是硫酸铵沉淀法,得到的粗提酶液一般可直接用于检测(Till et al,2006)。如果需要得到纯度更高的酶,需要经过刀豆体蛋白A-琼脂糖柱吸附(Pimkin et al,2007),用alpha-甘露糖苷洗脱,但是得率较低(Yang et al,1999)。因此,大部分TILLING突变体检测实验是用粗提酶来检测的,该粗提酶的主要成分即为CELI酶。
由于CELI酶为糖基化修饰酶,故不能在原核细胞中表达(Yang et al,1999)。因此,Maxim Pimkin等人用昆虫细胞系Sf9细胞系得到重组的CEL I酶(Pimkin et al,2007),但是该方法成本较高、周期较长,实验条件苛刻。此外,目前商业化的CEL I和CEL II都需要依赖进口,由于冷链运输价格昂贵,导致其未广泛在国内使用。
早在上世纪八十年代人们就开始在植物中表达重组蛋白,并且广泛地应用在医学制药领域(Buyel et al,2014)。植物细胞不仅仅能够表达一般外源蛋白,当然还包括医用抗体等产品(Rodr guez et al,2004)。因此转基因植物表达外源蛋白是一种不可或缺的方法,相比其他动物,如昆虫细胞,它既经济而且可以大规模种植来用于生产。传统的重组外源蛋白方法主要依赖于稳定的转基因植株,它不仅耗时并且受资源、空间的限制。相比而言,植物瞬时表达法是一种在几天内非常快速产生大量外源活性蛋白 的方法,并且能够迅速满足商业化产品的生产要求(Sainsbury et al,2014)。一般来说这种系统表达的蛋白量的高低,主要受mRNA密码子选择及GC含量、表达载体的框架结构、寄主细胞、培养条件等影响(Ullrich et al,2015)。植物瞬时表达能够在农杆菌浸染后2-4天内迅速表达至最高水平(Lacroix et al,2013)。而稳定的表达体系需要T-DNA整合至目标的基因组,在浸染农杆菌10-14天后,才有选择性地提高表达水平(Janssen et al,1990)。间接的证据表明瞬时表达中的外源T-DNA拷贝在表达外源蛋白时占主要的作用,而非整合至植物基因组中再发挥作用(Lacroix et al,2013;et al,2015)。
烟草作为植物界的“小白鼠”,被作为反应器的主要物种(Lacroix et al,2013),主要是由于烟草的叶片量多,非常适合表达外源蛋白。
技术实现要素:
本发明的一个目的是提供一种植物DNA错配识别的核酸内切酶(Recognizing DNA Mismatch Specific endonucleases in plants)的制备方法。
本发明提供的植物DNA错配识别的核酸内切酶的制备方法包括如下步骤:在烟草叶片中表达植物DNA错配识别的核酸内切酶,再从表达植物DNA错配识别的核酸内切酶的烟草叶片中提取植物DNA错配识别的核酸内切酶。
上述方法中,所述在烟草叶片中表达植物DNA错配识别的核酸内切酶,再从表达植物DNA错配识别的核酸内切酶的烟草叶片中提取植物DNA错配识别的核酸内切酶的方法包括如下步骤:将所述植物DNA错配识别的核酸内切酶的编码基因导入农杆菌,得到重组菌;再用所述重组菌侵染烟草外植体,得到侵染后烟草外植体;再从所述侵染后烟草外植体中提取所述植物DNA错配识别的核酸内切酶,得到植物DNA错配识别的核酸内切酶。
上述方法中,所述植物DNA错配识别的核酸内切酶的编码基因是通过重组表达载体导入所述农杆菌的。
上述方法中,所述侵染的方法为将所述重组菌注射到所述烟草叶片下表皮。
上述方法中,所述重组表达载体为将所述植物DNA错配识别的核酸内切酶的编码基因插入表达载体中得到的载体。
上述方法中,所述外植体为非子叶叶片;所述目的农杆菌为农杆菌EH105。
上述方法中,所述植物DNA错配识别的核酸内切酶为mung bean nuclease、SP1、S1、P1、ZEN1、ZEN2、CEL I、CEL II、M1、BENI、T7Endonuclease I、Endo V或T4Endonuclease VII;所述植物DNA错配识别的核酸内切酶具体为CELI。
上述方法中,所述CELI酶的编码基因的核苷酸序列为序列表中序列3的第65-958位。
上述方法中,所述表达载体为pCambia 2300-GFP(N)或pCambia 2300-GFP(C)。
本发明的另一个目的是提供由上述方法制备得到的植物DNA错配识别的核酸内切酶。
上述植物DNA错配识别的核酸内切酶在切割含有错配位点DNA中的应用也属于本发明的保护范围。
本发明采用烟草瞬时表达系统,高效表达外源CEL I酶。通过实验证明:本发明的表达系统从注射农杆菌到纯化蛋白仅需3-5天时间,大大节约时间和成本,并且酶活并没有降低。
附图说明
图1为重组载体pCambia2300-CELI-His的示意图。
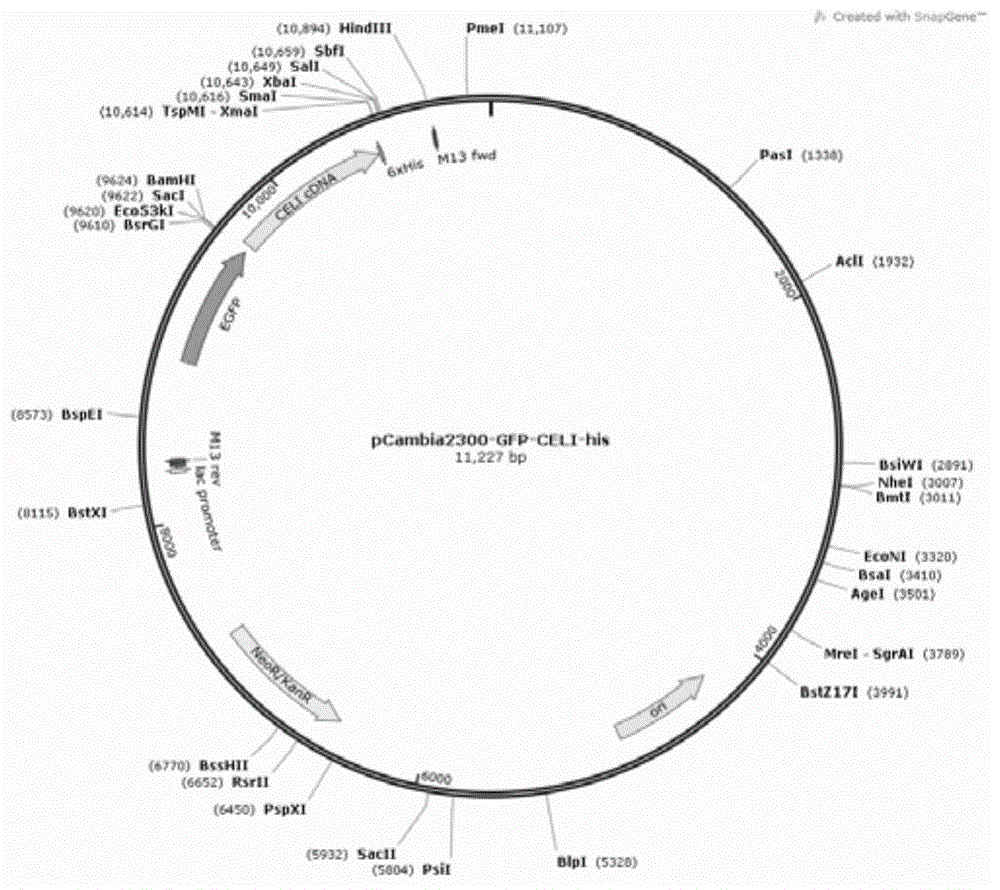
图2为重组载体pCambia2300-GFP-CELI-His的示意图。
图3为烟草注射农杆菌后,重组载体pCambia2300-CELI-His的表达情况。左侧为含有空载体的重组菌;右侧为含有重组载体pCambia2300-GFP-CELI-His的重组菌。
图4为重组CELI-GFP的SDS-PAGE图。
图5为CELI-His注射农杆菌后,不同天数表达情况。
图6为重组CELI-GFP和CELI-His(外源CELI)与内源CELI比较。泳道1为内源CELI对野生型DNA的酶切结果;泳道2为重组CELI-GFP对突变体DNA的酶切结果;泳道3为CELI-His对突变体DNA的酶切结果;泳道4为内源CELI对突变体DNA的酶切结果。
图7为部分植物DNA错配识别的核酸内切酶的同源性分析。图7A为部分植物DNA错配识别的核酸内切酶的同源性分析谱图;图7B为部分植物DNA错配识别的核酸内切酶的同源性分析(Bradley J.Till,2004;Maxim Pimkin,2007)。
图8为为部分植物DNA错配识别的核酸内切酶的同源性分析(Bradley J.Till,2004;Maxim Pimkin,2007)。
具体实施方式
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。
pCambia 2300-GFP(N)和pCambia 2300-GFP(C)是北京鼎国昌盛生物技术有限责任公司的产品,产品目录号为MCV036。
农杆菌EH105是上海迈其生物科技有限公司的产品,产品目录号为CH5002B。
实施例1、植物瞬时表达CELI酶的方法及效果
一、植物瞬时表达载体的构建
1、提取新鲜的芹菜叶片的RNA,反转录得到cDNA。反转录引物如下所示(下划线为His-tag):正向引物:5’-TAAAAAAACCTATAAATACGGATCTCGAGATGACGC-3’;反向引物:5’-TCAGTGGTGGTGGTGGTGGTGTTCTTCTGCCAAAGAATGATCTG-3’。
2、以步骤1获得的cDNA为模板,采用正向引物F和反向引物R进行PCR扩增,得到PCR扩增产物。引物序列如下(下划线的序列为酶切位点):正向引物:5’-GGATCCTATTTTACTGTTTTCGTAACAGTTTTGTAATAAAAAAACCTATAAATA-3’;反向引物:5’-TCTAGATTCAGCAGAGTCGGTACAGATCTCCCGGGGGTACCGAATTCAGTGGTGG-3’。
3、用限制性内切酶BamH I和Xba I对PCR扩增产物进行双酶切,回收并纯化大小为1025bp的PCR扩增产物(即为CELI的编码基因);用限制性内切酶BamH I和Xba I对pCambia 2300-GFP(N)和pCambia 2300-GFP(C)进行双酶切,回收大小为11,227bp的骨架载体1和大小为11,236bp的骨架载体2;将大小为1,025bp的PCR扩增产物分别与大小为11,227bp的骨架载体1和大小为11,236bp的骨架载体2连接,分别得到重组载体pCambia 2300-CELI-His(图1)和重组载体pCambia2300-GFP-CELI-His(图2)。
对重组载体pCambia 2300-CELI-His进行测序验证:重组表达载体pCambia 2300-CELI-His为将pCambia 2300-GFP(N)的BamH I和Xba I位点间的DNA替换为序列3中第65-958位核苷酸所示的CELI酶的编码基因,保持pCambia 2300载体的其他序列不变得到的重组载体,表达GFP-CELI。
对重组载体pCambia2300-GFP-CELI-His进行测序验证:重组表达载体pCambia2300-GFP-CELI-His为将pCambia 2300-GFP(C)载体的BamH I和Xba I位点间的DNA替换为序列3中第65-958位核苷酸所示的CELI酶的编码基因,保持pCambia 2300-GFP(C)载体的其他序列不变得到的重组载体,表达CELI-His。
PCR扩增产物双酶切反应(总体积20ul):Cutesmart buffer(NEB)2.0ul;KpnI 0.5ul;XbaI 0.5ul;PCR扩增产物(200ng/ul)5ul;H2O 12ul。37℃反应1-2h。
pCambia 2300-GFP(N)或pCambia 2300-GFP(C)的酶切反应(总体积20ul):Cutesmart buffer(NEB)2.0ul;KpnI 0.5ul;XbaI 0.5ul;pCambia 2300-GFP(N)或pCambia 2300-GFP(C)(200ng/ul)5ul;H2O 12ul。37℃反应1-2h。
连接反应体系(总体积20ul):10倍ligation buffer 2.0ul;PCR扩增产物双酶切(胶回收)2.0ul;载体双酶切(胶回收)4.0ul;Ligase 1.0ul;H2O 11ul。16℃过夜。
二、植物瞬时表达CEL I酶
1、农杆菌感受态细胞的制备
将农杆菌EH105接种到5ml的50mg/ml含利福平的LB培养基中,过夜培养,得到菌液;将5ml的菌液全部加入250ml的含利福平的LB培养基中;28℃,200rpm培养至OD600为0.6-0.8;冰浴10min,5000rpm,4℃,离心10min,弃上清,收集菌体;等体积10%甘油悬浮,收集菌体;1/2体积10%甘油悬浮,收集菌体;1/2体积10%甘油悬浮,收集菌体;50ml超纯水配制的甘油悬浮,收集菌体;1ml超纯水配制的10%甘油悬浮,得到农杆菌感受态细胞;100ml分装,保存于-70℃待用。
2、重组菌的获得
将40ml的农杆菌感受态细胞分别和重组载体pCambia 2300-CELI-His、重组载体pCambia2300-GFP-CELI-His和pCambia 2300-GFP(N)载体(对照)混合;冰浴30分钟;转入预冷的电击杯,电击,分别得到含有重组载体pCambia 2300-CELI-His的重组菌、含有重组载体pCambia2300-GFP-CELI-His的重组菌和含有pCambia 2300-GFP(N)载体的重组菌;将含有重组载体pCambia 2300-CELI-His的重组菌、含有重组载体pCambia2300-GFP-CELI-His的重组菌和含有pCambia 2300-GFP(N)载体的重组菌分别加入1ml的LB培养基,28℃培养3h;浓缩至150ml涂布于含利福平50mg/ml,卡纳50mg/ml的LB培养基。对含有重组载体pCambia 2300-CELI-His的重组菌、含有重组载体pCambia2300-GFP-CELI-His的重组菌和含有pCambia 2300-GFP(N)载体的重组菌进行菌液PCR鉴定。
PCR反应体系(总体积20ul):H2O 16.5ul;10×PCR buffer(含20mmol/L MgCl2)2.0ul;dNTPs mix(10mmol/L each)0.5ul;P1(10mmol/L)0.5ul;P2(10mmol/L)0.5ul;农杆菌菌液0.5ul;Tag(2.5U/ml)0.5ul。
PCR反应程序:94℃预变性4min;94℃变性30s,56℃退火30s,72℃延伸30s,30个循环;最后72℃延伸5min。
3、烟草瞬时体系
(1)挑取含有重组载体pCambia 2300-CELI-His的重组菌、含有重组载体pCambia2300-GFP-CELI-His的重组菌和含有pCambia 2300-GFP(N)载体的重组菌的单克隆于5ml含有100ug/ml潮霉素和50ug/ml卡纳的LB液体培养中,28~30℃震荡过夜培养。以含有pCambia 2300载体的重组菌为对照。
(2)分别将1ml培养后的含有重组载体pCambia 2300-CELI-His-GFP的重组菌、含有重组载体pCambia2300-GFP-CELI-His的重组菌和含有pCambia 2300-GFP(N)载体的重组菌转接到25ml含有100ug/ml潮霉素、50ug/ml卡纳和乙酰丁香酮的LB液体培养基中,检测过夜培养的菌液OD600的值。
(3)5000g,离心15分钟,分别收集菌体,并用重悬液(10mM MgCl2、10mM 2-(N-吗啉代)乙磺酸、100uM乙酰丁香酮)重悬菌体至OD600为0.4,分别得到侵染液1、侵染液2和侵染液3。
(4)将侵染液1、侵染液2和侵染液3室温放置2~3h后分别装入5ml注射器内,用拇指按压注射器反板将侵染液从叶片下表皮注射到烟草(本式烟草,Nicotiana benthamiana)叶片内(勿使用子叶)。
(5)注射后将烟草培养2~5天,用荧光显微镜观察荧光情况,发现CEL I酶在侵染液1注射得到的烟草的细胞间质中表达(图3),并且在4天时表达量最高(图5)。
三、CEL I酶的提取及纯化
1、取侵染液1和侵染液2注射得到的烟草叶片各30-40片(含CELI酶),分别用液氮磨碎成粉末加至100ml的50mM Tris-Hcl(pH 7.7)、0.2M KCl、100um PMSF的缓冲液中,混匀。
2、分别用纱布过滤残渣,得到滤液;将滤液9000g离心力离心45min,弃沉淀,分别收集上清;向上清中加入硫酸铵至饱和度25%(每100ml接入10.6g硫酸铵),分十次加入,并缓慢搅拌,分别得到溶液1和溶液2。
3、分别将溶液1和溶液2在9000g离心力的条件下离心45min,弃沉淀,分别收集上清;向上清中加硫酸铵至饱和度80%(每100ml加入39g硫酸铵),分十次加入,并缓慢搅拌,分别得到溶液3和溶液4。
5、分别将溶液3和溶液4在9000g离心力的条件下离心45min,弃上清,分别收集沉淀;用2ml 20mM Tris-Hcl(pH7.7)和0.2M KCl溶解沉淀,离心,弃去不溶物,分别得到粗酶液1和粗酶液2。
6、将粗酶液1和粗酶液2经Ni柱吸附,用300mM咪唑洗脱,分别得到纯化后的酶液1和酶液2。
7、将纯化后的酶液1和酶液2分别经SDS-PAGE胶分离,酶液1在75kDa有明显条带(GFP-CELI),酶液2在43kDa有明显条带(CELI-His)(图4)。
实施例2、CEL I酶的功能验证
用含有错配位点的待酶切DNA样品测试上述实施例1获得的外源CEL I酶(CEL I-His和GFP-CEL I)的活性,以内源CEL I酶作对照。
一、内源CEL I酶的制备
1、取10kg芹菜,放入榨汁机中,缓慢榨汁后,避免温度过热,在汁液中加入预冷匀浆缓冲液(含100mM Tris-HCl(pH均为7.7)和100uM PMSF),得到混合液。
2、将所述混合液4℃下14000rpm离心10min,弃沉淀,取上清液;向上清液中 缓慢加入硫酸铵(每100ml上清液加25g硫酸铵),分10次加入(每100ml体积加入14.4g硫酸胺),待完全溶解后,低速搅拌30min;14000rpm离心45min,弃沉淀,取上清液。
3、向上清液中缓慢加入硫酸铵至(每100ml上清液加80g硫酸铵),分十次加入(每100ml体积加入39.0g固体硫酸铵)。低速搅拌30min后,14000rpm离心3min。弃上清,收集沉淀。
4、用1/10体积的缓冲液(0.1M Tris-HC1、100uM PMSF;pH为7.7)溶解步骤3中得到的沉淀,1000rpm离心20min,弃沉淀,取上清。
5、把透析袋剪成适当长度的小段。用1mmol/L EDTA(pH 8.0)中将透析袋煮沸5min。用超纯水清洗透析袋。将透析袋放入10倍蛋白质溶液以上体积的缓冲液(0.1M Tris-HC1、100uM PMSF;pH为7.7)中,透析过夜,透析4-5次至透析液无颜色为止。
6、取40ml透析过的酶液经ConA-Sapharose-4B柱,用缓冲液(0.1M Tris-HC1、100uM PMSF;pH为7.7)冲洗,经alpha-甘露糖苷洗脱后,经缓冲液(0.1M Tris-HC1、100uM PMSF;pH为7.7)透析过夜,得到纯化后的CELI酶,经过SDS-PAGE检测,其大小为43kDa,即为内源CELI酶。
二、待酶切DNA样品的制备
1、DNA的合成
人工合成序列表中序列1所示的DNA分子和序列表中序列2所示的DNA分子,将其按2:1比例(质量比)混合,得到待PCR扩增的DNA(浓度为1ng/ul)。
序列2为将序列1的第544位核苷酸由G突变为A后得到的DNA分子。
2、PCR扩增
以步骤1获得的待PCR扩增的DNA为模板,采用F和R引物对所述模板进行PCR扩增,扩增至45个热循环,然后进行99℃变性10min,降温至70℃,每个循环降低0.3℃,69个循环结束,得到PCR扩增产物,即待酶切DNA样品。待扩增完毕后,用1%琼脂糖凝胶检测,检测条带是否特异,条带亮度是否达到要求。若条带较暗或不特异,需要重新扩增。引物序列如下:
F:5’-ATGCACTTAACAGAGGAAATCTTG-3’;
R:5’-TTACTTAATACTTTTCTTGTAGCCTTTA-3’。
对PCR扩增产物进行测序。测序结果表明:PCR扩增产物含有序列表中序列1所示的DNA分子和序列表中序列1的第544位核苷酸由G突变为A的异源双链DNA分子。
PCR扩增体系(10ul):10倍PCR缓冲液1.0ul、dNTP 0.8ul、Forward primer(10um)0.16ul、Reverse primer(10um)0.16ul、DNA模板1.0ul、聚合酶0.1ul、H20 6.78ul;
PCR扩增热循环:95℃2min;94℃10s,60℃30s,72℃1-3min;72℃5min,12℃hold。
异源错配双链形成:99℃10min;70℃20s,-0.3℃per cycle,70cycles;12℃10min。
3、PCR产物纯化
将步骤2获得的PCR扩增产物进行纯化,得到待酶切DNA样品,纯化的具体步骤如下:
(1)用在1倍体积的PCR扩增产物中加入2倍体积的Binding Buffer,翻转充分混匀(对小于200bp的PCR扩增产物,加入5倍体积的Binding Buffer)得到混合液。
(2)将所述混合液加入DNA纯化柱内(如果溶液体积>700μl,分次转移溶液);室温放置1-2min或更长时间。
(3)13,000g离心1min,弃废液(目的DNA长度≤500bp时,离心结束后将收集管中的溶液再次转入DNA纯化柱中,离心)。
(4)在DNA纯化柱中加入650ul Wash Buffer,13,000g离心30秒,弃废液,将DNA纯化柱放回收集管。
(5)重复步骤(4)。
(6)13,000g离心3min,以去除柱中残余乙醇。
(7)将纯化柱放入新的离心管中,向柱中加入在60℃预热的30-50μl Elution Buffer或ddH2O,室温放置1-2min或更长时间。
三、酶切反应
1、酶切缓冲液的配制
酶切缓冲液组成:10倍digestion buffer(含有Mg2+):2ug/ml BSA、0.02%Triton X-100(vol/vol)、0.1M KCl、0.1M HEPES(pH 7.5)、100mM MgCl2;
2、酶切反应
取2ul的待酶切DNA样品作为酶切反应底物,用上述酶切缓冲液进行酶切反应,得到酶切产物。
外源CEL I的酶切反应体系(6ul):10倍酶切缓冲液0.6ul、外源CELI(0.3mg/ml)0.2ul、PCR扩增产物2.0ul、纯水3.2ul。
内源CEL I的酶切反应体系(6ul):10倍酶切缓冲液0.6ul、内源CELI(0.3mg/ml)0.2ul、PCR扩增产物2.0ul、纯水3.2ul。
上述酶切反应条件均为45℃温育30min。
若酶切产物含有791bp、544bp和247bp的酶切片段,则待酶切DNA样品含有错 配位点(AC);若酶切产物仅含有791bp的酶切片段,则待酶切DNA样品不含有错配位点(AC)。
3、毛细管电泳检测酶切效果
酶切反应结束后立即加入2ul的0.225M的EDTA终止反应(15ul体系应加入3ul EDTA),再补入91ul超纯水稀释样品,可直接上机(FS 96,美国AATI公司)进行毛细管电泳,毛细管电泳条件如表1所示。
表1、毛细管电泳条件
结果如图6所示:本发明获得的外源CEL I-His与内源CEL I酶效果相当,酶切产物均分离得到大小为791bp、544bp和247bp的酶切片段,与待酶切DNA样品中的错配位点(AC)一致,说明本发明获得的外源CEL I酶与内源CEL I酶的活性无差异。
- 还没有人留言评论。精彩留言会获得点赞!