用于转移对亚洲大豆锈病抗性的多核苷酸和方法与流程
相关专利申请的交叉引用本申请要求2015年5月11日提交的美国临时申请62/159,718的权益,其全部内容以引用方式并入本文。序列表的并入本申请包含2015年5月11日经由efs-web以ascii格式提交的序列表,其包含文件名36446_0235u1_sl,大小为47,607字节,于2015年5月11日创建,包含八个序列,并且全文以引用方式并入本文。本公开涉及可用于增强豆类植物,并且更具体地大豆植物中的病原体抗性的组合物和方法,其通过向所述植物提供一种或多种基因进行,所述基因与对亚洲大豆锈病(asr)的致病原的抗性相关。本公开还涉及能够增强豆类对asr的抗性的多核苷酸,和使用这些多核苷酸序列制备抗asr的转基因豆类植物的方法。
背景技术:
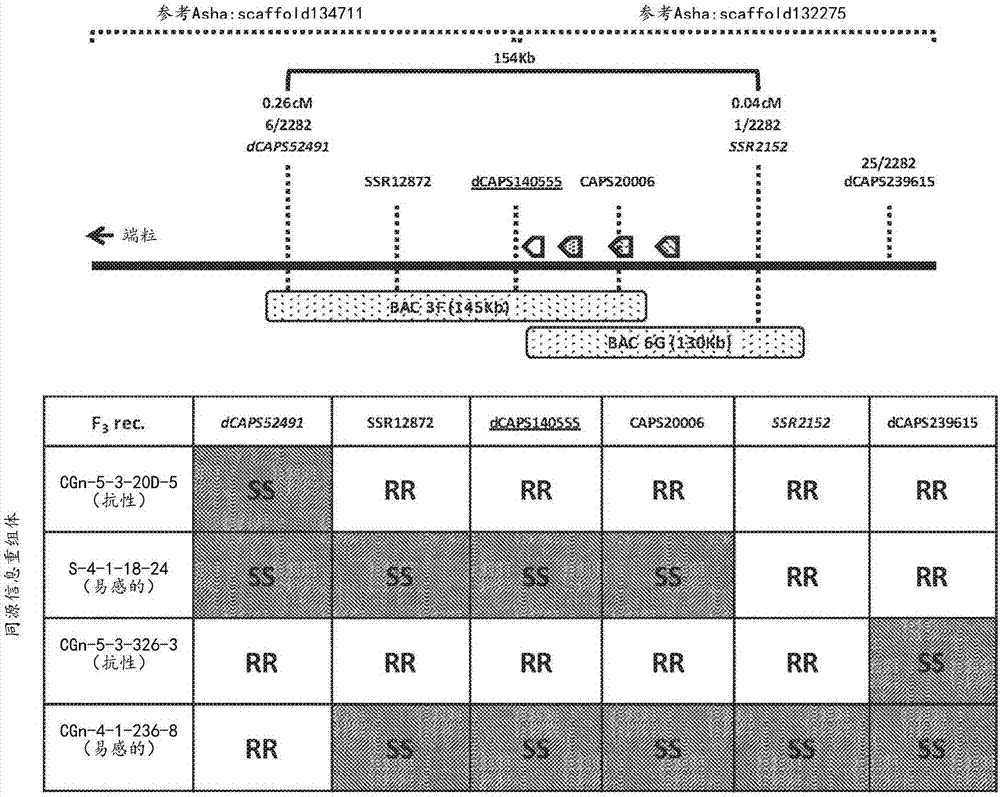
:大豆(glycinemax)是一种主要工业用作物,也是最重要的蛋白质源作物之一,并且被许多公共健康组织(包括美国糖尿病协会、美国心脏协会和美国癌症协会)认为是用于预防疾病和优化健康的关键食品组。亚洲大豆锈病(asr)是影响大豆的主要作物病害并且可不利地影响生长和产量。其由专性活体寄生真菌大豆锈菌(phakopsorapachyrhizi)形成,并且在较少程度上,由密切相关的真菌蟥层锈菌(phakopsorameibomiae)形成。该病害可导致在10-90%范围内的产量损失。技术实现要素:本公开涉及用于鉴定来自大豆物种的锈病抗性基因并将那些基因转化到豆类作物或豆类作物物种,诸如大豆中,以生成抗asr的植物的组合物和方法。本发明公开了分离的多核苷酸,其包含编码豆类来源的nb-lrr多肽的核苷酸序列,所述多肽与本文所公开的大豆序列具有至少90%的氨基酸序列同一性。在一个方面,当与易感植物和/或未转化的植物相比时,用多核苷酸转化的植物显示出对亚洲大豆锈病的增强的抗性。本发明还公开了重组dna构建体,其包含本文所述的多核苷酸。本文公开了可用的多肽,其可包括seqidno:1-8的多核苷酸或序列以及它们的变体,由其组成、或由其编码。本文公开了一种在豆类作物物种(例如,大豆)中赋予抗病性的方法,所述方法包括用异源豆类来源的nb-lrr基因转化豆类作物物种,所述基因赋予对豆类作物物种病害(例如,asr)的抗病性。本文公开了用重组dna构建体稳定转化的转基因豆类作物植物。在一个方面,所述重组dna构建体包含本文所公开的多核苷酸,所述多核苷酸编码能够对植物病害,如asr赋予抗性的一种或多种豆类来源的nb-lrr抗性基因。在一个方面,所述多核苷酸包含能够赋予对植物病害的抗性的一种或多种非豆类来源的nb-lrr抗性基因和/或非nb-lrr抗性基因。本文所述的多核苷酸还可包含抗性基因的任何组合。所述转基因豆类作物植物可包含一种或多种农学性状。还设想由此类转基因豆类作物植物获得种子。另外,本公开的特征还在于稳定转化的转基因豆类作物植物,其包含豆类来源的nb-lrr多核苷酸,所述多核苷酸与本文所述的序列具有至少90%序列同一性。本文公开了鉴定赋予对植物病害(例如,asr)的抗性的一种或多种抗性基因的方法。本文公开了产生asr抗性植物(例如,大豆物种)的方法。在一个方面,所述方法包括用豆类来源的nb-lrr抗性基因转化植物细胞。所述方法还可包括由所述转化的植物细胞再生转化的植物。在一个方面,所述方法包括使所述转化的植物生长,使得豆类来源的nb-lrr抗性基因的表达产生显示出对asr病害的增强的抗性的转化的植物。本文公开了产生豆类植物的方法,所述豆类植物为来自与包含本文所述的豆类来源的nb-lrr抗性基因的豆类植物杂交的子代。本文公开了测定豆类植物对植物病害(例如,asr)的抗病性的方法。在一个方面,所述方法包括使豆类植物的一部分暴露于植物病原体(例如,大豆锈菌);测量暴露于植物病原体的豆类植物上的植物病害症状;以及将所述植物病害症状与用于抗病性的参考标准进行比较。本文公开了增强植物对asr病害的抗性的方法。在一个方面,所述方法包括通过在抗asr的育种程序中将豆类来源的nb-lrr抗性基因基因掺入种质(例如,大豆作物物种)中来赋予对asr病原体(例如大豆锈菌)的抗性。所述方法的特征在于豆类来源的nb-lrr抗性基因,其编码nb-lrr多肽。在一个方面,nb-lrr多肽包含氨基酸序列,所述氨基酸序列与本文所公开的豆类来源的nb-lrr多肽具有至少90%同源性。本文所述的方法的特征还在于用所述多肽转化的植物,当与易感植物相比时,所述植物显示出对asr的增强的抗性。附图说明图1示出木豆(cajanuscajan)中的ccrpp1基因区域与来自大豆染色体12和9的基因组区域具有同线性。g119-99中紧密连接到ccrppl的标记dcaps140555定位成接近大豆中的单个同线nb-lrr基因(glyma12g01420)(表示为“*”)。类似地,位于木豆基因中的caps20006标记定位在同源大豆基因glyma12g01420中。图2a-2b示出经鉴定的ccrpp1基因座(图2a)和四个nb-lrr旁系同源基因(图2b)的物理和高分辨率基因图谱区间。品种g119-99中的ccrpp1基因组区域包含四个nb-lrr旁系同源基因(图2a)。r基因区域缩小至涵盖标记dcaps52491和ssr2152的154kb区域。其是在筛选1141种分离的f2植物后获得的最具信息的重组物(2282个配子)。功能增益区间由标记dcaps52491和dcaps239615描绘出,并且功能丧失区间由标记dcaps52491和ssr2152描绘出。bac3f携带三个旁系同源基因(-1、-2和-3)并且bac6g携带四个旁系同源基因(-1至-4)(图2b)。最密切相关的标记dcaps140555由包含iq钙调蛋白结合基序的基因设计。具体实施方式作物病害导致严重的作物管理问题并且有时可导致所有作物歉收。亚洲大豆锈病是世界大豆生产的威胁并且目前通过使用叶面杀真菌剂来解决。商业植物品系中稳定且可靠的遗传抗性是与大豆作物产量相关的重要特征,并且目前不能获得对由大豆锈菌形成的亚洲大豆锈病具有完全抗性的商业上种植的大豆品种。asr的致病原,大豆锈菌和蟥层锈菌感染宽范围的豆科植物的叶组织(分别为17个属中至少31种;slaminko等人,(2008)plantdis.,92:797-771;和19个属中至少42种;frederick等人,(2002)mycology,92:217-227)。总之,其它属中的另外152种已被描述为是大豆锈菌的潜在宿主(bonde等人,(2008)plantdis.,92:30-38;goellner等人,(2010)molecularplantpathology,11:169-177;ono等人,(1992)mycol.res.,96(10):825-850;和slaminko等人,(2008)plantdis.,92:797-771)。当前,杀真菌剂应用是缓解asr的唯一可用方法。目前,不能获得对大豆锈菌具有完全抗性的商业种植的大豆(glycinemax)品种。大豆中对大豆锈菌的抗性很少;usda对整个美国大豆种质资源进行了评价并且发现少于5%具有对大豆锈菌的抗性或部分抗性。另外,这些大豆品种中可用的基因仅提供分离株特异性的抗性;因此,这些资源不能在田间条件下,诸如存在多个种族时,提供耐久的抗性。考虑到asr是大豆生产的主要威胁,有利的是鉴定抗性基因的源并将这些转基因的基因并入豆类种质诸如大豆中以增强防护。为鉴定新型抗性基因,对若干非大豆豆类物种进行筛选以获得对大豆锈菌具有抗性的变型。鉴定若干豆类中的显性抗性基因并确认其是抗性(r)基因的良好表征类型,核苷酸结合结构域富含亮氨酸重复序列(nb-lrr)基因家族的成员。当例如,作为单个基因、多个基因,或以多基因盒的形式转移到大豆时,这些抗性基因可以经由异源表达提供对大豆锈菌的抗性。虽然已经尝试了植物物种之间的抗性基因的转移,但在很大程度上,其是不成功的或与适应性障碍相关(tai等人,(1999)pnas,96:14153-14158;ren等人,(1997)euphytica,93:353-360;day等人,(2005)plantcell17:1292-1305;banerjee等人,(2001)genetics158:439-450;tian等人,(2003),nature423:74-77;和frost等人,(2004)mpmi,17:224-232)。并且,虽然在一些植物中已经实现了抗性的成功转移(haltermand.等人,(2008)plantdisease92:339-343(土豆);fostersj等人,(2009)molplantmicrobeinteract22:589-600(土豆);brunners等人,(2012)plantbiotechnologyjournal10:398-409(小麦);和horvathdm等人,(2012)plosone7:e42036(西红柿)),但先前并未实现通过r基因转移在大豆中获得对大豆锈菌的抗性。植物可以通过各种细胞机制来进行自我保护。当前,应当理解植物免疫系统由细胞外(通常被称为第一层级免疫)和细胞内(通常被称为第二层级免疫)的受体组成。两组受体均可以检测病原体并对病原体进行应答。第一层级对病原体的主要元件进行应答,从而导致病原体相关分子模式(pamp)触发的免疫性的激活。成功的病原体通过分泌被称为“效应蛋白”或“效应子”的分子来克服pamp-触发的免疫性,所述分子位于植物质外体或吸收在植物细胞中。效应子调控宿主细胞功能以抑制宿主免疫应答,以便有助于建立感染或以其它方式增强病原体的生长条件,例如通过确保营养物质的可用性。在一些情况下,植物已经进化出第二层级免疫性,其中r基因产物识别特异性效应子的活性,从而导致效应子触发的免疫。植物基因组中的抗性(r)基因产生识别特异性病原体效应子的r蛋白。最大类的r基因编码包含核苷酸结合(nb)结构域和富含亮氨酸重复序列(lrr)结构域的蛋白质,其被命名为“nb-lrr”并保护细胞内环境。第二类r基因编码由偶联到受体样结构域的细胞外膜锚定的lrr结构域组成并被称为受体样蛋白(rlp)的蛋白质。rlp类通常与受体样激酶偶联并监控细胞的外部。r蛋白通过病原体效应子的直接结合或病原体效应子对宿主细胞靶标的间接作用来传递对病原体的抗病性。因为宿主与病原体之间的“军备竞赛(armsrace)”,病原体效应子可具有无毒性或毒性效应。效应子的毒性活性与操纵正常宿主细胞功能或由病原体抑制宿主免疫应答以便建立成功的感染相关联。在无毒性的情况下,通过相应植物r蛋白进行识别激活了宿主免疫或防御反应,从而导致程序性细胞死亡和对病原体的抗性。依赖于病原体效应子和植物抗性基因之间的一对一对应(“基因对基因之间的关系”)的主要基因抗性已被广泛用于育种方法中。然而,此类基于引入单个r基因的抗性通常是种族特异性的并且容易被病原体avr基因中的单个突变克服,这是由于多样性选择的结果以避免被宿主识别。因此,此类定性抗性的耐久性是令人担忧的。已经尝试引入新型抗微生物/抗真菌基因或修饰转基因植物中的内源性防御相关基因的表达。然而,在许多情况下,效果仅是部分的或短期的并且可能以牺牲植物产量和/或活力为代价。因此,有效使用r基因仍然是工程化抗性的最有效方式之一。另外,虽然单独的r基因可被病原体快速克服,但若干r基因同时成功基因掺入可提供对病原体分离株的耐久的种族独立性抗性。例如,使用基因叠加,将受关注的两种或更多种基因组合到单个植物中的方法,可能是提供抗病性的有效策略。r基因的成功基因叠加的示例是20世纪70年代将cf-9抗性基因座基因掺入西红柿中,这有效阻止了由番茄芽枝病菌(cladosporiumfulvum)引起的番茄叶霉问题。然而,使用经典育种以在作物中形成多个r基因的有效“叠加”通常受r基因的显性性质以及r基因在作物如大豆中的可用性受到阻碍。本文所公开的核酸和多肽可用于对植物(例如豆类作物物种)赋予或增强或增加真菌抗性的方法。本文所公开的方法和组合物可包含以下多肽和多核苷酸序列:seqidno:1ccrpp1基因,来自木豆(多核苷酸序列)(nb-lrr-2)seqidno:2:ccrpp1(多肽序列)(nb-lrr-2)seqidno:3:nb-lrr-1(多核苷酸序列)seqidno:4:nb-lrr-1(多肽序列)seqidno:5:nb-lrr-3(多核苷酸序列)seqidno:6:nb-lrr-3(多肽序列)seqidno:7nb-lrr-4(多核苷酸序列)seqidno:8:nb-lrr-4(多肽序列)本文所公开的组合物和方法可用于保护植物免受真菌病原体侵害。宿主和病原体之间的相互作用可以“免疫性”到“部分抗性”到“易感性”的连续体来描述。术语“免疫性”或“免疫”在本文中用于表示不存在任何宏观可见的病害症状。术语“部分抗性”在本文中用于表示不具有或具有有限孢子形成的宏观可见病变的存在和/或任何病害症状的范围或程度的减少和/或进展延缓,并且可以例如表现为病变数减少或具有孢子形成减少的病变。如本文所用,对asr的术语“易感性”或短语“缺乏抗性”是指在孢子形成水平等于或高于参考标准(诸如例如品种williams或peking)中观察到的孢子形成水平的情况下,出现病变。术语“抗性”在本文中用于表示在植物中不存在由植物病原体所致的一种或多种病害症状或所述病害症状减少。抗性可以是指当与对病害易感的植物或不包含减少一种或多种病害症状的有效抗性基因(例如nb-lrr基因)的植物相比时,病害症状,例如病变数、脱叶、和相关的产量损失降低、最小化或减少。另外,抗性可包括病原体(例如,真菌)增殖的预防或延缓。广义上来讲,术语“抗性”包括如上文所定义的免疫性和部分抗性。“植物病原体”或“真菌病原体”在本文中可用于表示例如层锈菌属(phakopsora)(包括大豆锈菌和蟥层锈菌物种)的真菌病原体。已知这些物种引起植物中的asr。植物病害或豆类作物物种病害例如可以为asr。可进行本公开的方法以例如提供大豆对专性活体寄生真菌大豆锈菌(asr的主要致病原)或对蟥层锈菌的增强的抗性。例如,对真菌病原体的增加或增强的抗性可与易感植物,诸如例如williams或peking的反应进行比较。抗性可以是变化的并且与植物或植物部分(例如,叶)上观察到的病害症状(例如,病变)的比例(即,百分比)相关。可以给出免疫性、抗性和易感性的数值分数或值。例如,抗性的数值分数表示植物对植物病害(例如,asr)表现出的抗性程度。数值分数还可用于在例如所关注的植物(例如,转基因豆类作物植物)与易感植物(例如williams或peking)或参考标准之间进行抗性程度的比较。本文所公开的抗性的方法和组合物涉及从豆类物种中分离一种或多种抗性基因,并且随后经由同源或异源表达将这些抗性基因中的一种或多种转移到另一种植物,例如大豆中,以提供对层锈菌属的抗性。术语“抗病性基因”或“抗性基因”在本文中用于表示编码能够增强或改善植物中的防御或免疫系统应答的蛋白质或多肽的基因。本公开的一个方面包括将功能性r基因转移到性相容或不相容物种以产生抗病性。本文所述的多肽和r基因(例如,nb-lrr多肽和nb-lrr基因)可单独使用或与其它r基因叠加使用或与非r基因(包括非-nb-lrr抗性基因)叠加使用以提供对asr的抗性。因此,本公开的转基因方法可单独使用或与其它策略组合使用以在植物中产生或赋予asr抗性。其它可用的策略包括但不限于阻断效应子的功能活性,抑制病原体或病原体因子(例如真菌)摄入宿主细胞(例如植物细胞)中和/或常规的抗性育种。本公开的方法可提供或增强植物的抗性,使得病害如asr的致病原不能再繁殖。术语“增强”是指改善、增加、扩增、繁殖、升高和/或提升,从而减少一种或多种病害症状。因此,当与对层锈菌属易感或耐受的植物相比时,植物(例如,大豆)表现出对病害(例如,asr)的增加的抗性。在一个方面,本文所述的方法可减少豆类植物病害(例如,asr)的一种或多种症状(即,病害症状)。一种方法可包括使转基因豆类作物植物(例如,大豆)暴露于豆类植物病害,从而导致具有对植物病害的增强的抗性的转基因豆类作物植物。在一些方面,转基因豆类作物植物包含一种或多种豆类来源的nb-lrr多核苷酸。一种或多种豆类来源的nb-lrr多核苷酸可与如本文所公开的序列具有至少90%序列同一性。术语“植物”在本文中用于包括任何植物、组织或器官(例如,植物部分)。植物部分包括但不限于可被培养成整个植株的细胞、茎、根、花、胚珠、雄蕊、种子、叶。植物细胞是植物的细胞,其直接从种子或植物中获取,或者来源于由植物获取的细胞的培养。再生植物的子代、变体和突变体在本公开的范围内,前提条件是这些部分包含被引入的多核苷酸。在一个方面,植物、植物部分或植物细胞来源于植物,所述植物包括但不限于苜蓿、三叶草、豌豆、菜豆、兵豆、羽扇豆、牧豆树、角豆树、大豆、花生和罗望子。在一个方面,植物为豆类。在一个方面,nb-lrr多肽、nb-lrr多核苷酸、和/或nb-lrr抗性基因(或nb-lrr基因)来源于豆类。豆类的示例包括但不限于菜豆属(例如,法国菜豆、四季豆、菜豆(phaseolusvulgaris)、棉豆(phaseoluslunatus)、宽叶菜豆(phaseolusacutifolius)、红花菜豆(phaseoluscoccineus));大豆属(例如,野大豆(glycinesoja)、大豆(glycinemax(l.)));豌豆(pisum)(例如,脱荚豌豆(有时被称为光皮豌豆或圆粒豌豆;菜豌豆(pisumsativum));皱粒豌豆(pisumsativum)、糖荚豌豆(pisumsativum),其也被称为糖荚豌豆(snowpea)、食荚豌豆(edible-poddedpea)或嫩豌豆(pisumgranda));花生(arachishypogaea)、三叶草(三叶草属(trifoliumspp.))、苜蓿(medicago)、葛藤(puerarialobata)、常见的苜蓿、苜蓿(紫花苜蓿(medicagosativa))、鹰嘴豆(鹰嘴豆属(cicer))、兵豆(lensculinaris)、羽扇豆(lupinus);野豌豆(vicia)、蚕豆、蚕豆(viciafaba)、连理草(山黧豆属(lathyrus))(例如家山黧豆(lathyrussativus)、玫红山黧豆(lathyrustuberosus));豇豆属(例如,乌头叶菜豆(vignaaconitifolia)、赤豆(vignaangularis)、黑吉豆(vignamungo)、绿豆(vignaradiata)、班巴拉花生(vignasubterrane)、赤小豆(vignaumbellata)、野豇豆(vignavexillata)、豇豆(vignaunguiculata)(也称为长豇豆、豇豆));木豆(cajanuscajan)、硬皮豆属(macrotyloma)(例如,落花生(macrotylomageocarpum)、马蚕豆(macrotylomauniflorum);四棱豆(psophocarpustetragonolobus)、非洲豆薯(sphenostylisstenocarpa)、埃及黑豆、鹊豆(lablabpurpureus)、豆薯(pachyrhizuserosus)、瓜尔豆(cyamopsistetragonolobus);和/或刀豆属(例如,矮刀豆(canavaliaensiformis))、刀豆(canavaliagladiata)。本文所述的组合物和方法可导致农学上期望的品系或品种。农学特性或性状包括但不限于,除草剂耐受性、增加产量、病虫害防治、野草防治、害虫防治、病原体抗病性(例如,真菌、病毒、细菌)、高蛋白产量、发芽和幼苗生长控制、增强营养、环境胁迫抗性、增加可消化性、雄性不育、开花时间、或转化技术性状,诸如细胞周期调节和/或基因靶向。本公开提供了用于筛选或测定对植物病害具有抗性、免疫性或易感性的豆类植物的方法。植物对具体病原体的抗性、免疫性或易感性的测定是本领域技术人员已知的。用于筛选或测定对植物病害具有抗性、免疫性或易感性的豆类植物的方法包括使植物细胞、组织或器官(例如叶)暴露于病原体(例如,大豆锈菌),并且然后测定和/或测量暴露的植物中对由病原体形成的植物病害(例如,asr)的抗性、免疫性和/或易感性的程度。所述方法还可包括测量暴露于植物病原体的植物上的任何可观察到的植物病害症状,并且然后将所述植物病害症状与参考标准进行比较以确定抗病性的程度或范围。使植物细胞、组织或器官暴露于病原体的方法在本领域中是已知的。测量、比较和测定对由病原体所致的病害诸如例如asr的抗性、免疫性和/或易感性(例如,植物病害症状)的水平的方法在本领域中也是熟知的。还可评估暴露的植物以分离与植物对特定病原体或病害的抗性、免疫性或易感性相关、有关和/或赋予所述抗性、免疫性或易感性的多核苷酸、氨基酸序列和/或遗传标记。进一步评估包括但不限于从暴露的植物分离多核苷酸、核酸或氨基酸序列,进行分离的多核苷酸或核酸的测定,例如,以检测与一种或多种农学特性或性状相关的一种或多种生物学或分子标记,所述农学特性或性状包括但不限于抗性、免疫性和/或易感性。由此类方法收集的信息可用于例如育种程序中。在本公开中,“核酸”是指单链或双链形式的脱氧核糖核苷酸或核糖核苷酸聚合物,并且除非另外限制,否则涵盖在以下方面具有天然核苷酸的基本性质的已知类似物(例如如肽核酸):其以与天然存在的核苷酸相似的方式杂交至单链核酸。术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,是指氨基酸残基的聚合物。这些术语适用于其中一个或多个氨基酸残基为相应的天然氨基酸的人工化学类似物的氨基酸聚合物,并且适用于天然氨基酸聚合物。本公开的多肽可以由本文所公开的核酸产生,或者通过使用标准的分子生物学技术产生。例如,本公开的截短蛋白可通过在适当的宿主细胞中表达该实施方案的重组核酸,或者另选地通过体外方法的组合,诸如蛋白酶消化和纯化产生。术语“编码”在本文中用于表示核酸包含所需信息,其通过使用密码子以指导核苷酸序列(例如,豆类序列)翻译成特定蛋白质来指定。编码蛋白质的核酸可以包含在所述核酸的翻译区内的非翻译序列(例如内含子)或可以缺少这种居间的非翻译序列(例如,如在cdna中)。本公开的方面涵盖分离的或实质上纯化的多核苷酸或蛋白质组合物。“分离的”或“纯化的”多核苷酸或蛋白质或其生物活性部分,实质上或基本上不含通常在该多核苷酸或蛋白质的天然环境中存在的、与该多核苷酸或蛋白质相伴随或相互作用的成分。因此,分离的或纯化的多核苷酸或蛋白质,当通过重组技术(例如pcr扩增)生产时实质上不含其它细胞物质或培养基,或者当通过化学法合成时实质上不含化学前体或其它化学品。最佳地,“分离的”多核苷酸不含在衍生该多核苷酸的生物体的基因组dna中天然位于该多核苷酸的旁侧的序列(即,位于该多核苷酸的5′和3′末端的序列)(例如,蛋白质编码序列)。例如,在本公开的一些实施方案中,分离的多核苷酸可含有少于约5kb、约4kb、约3kb、约2kb、约1kb、约0.5kb或约0.1kb的在衍生该多核苷酸的细胞的基因组dna中天然位于该多核苷酸的旁侧的核苷酸序列。基本上不含细胞物质的蛋白质包括具有小于约30%、约20%、约10%、约5%或约1%(以干重计)的污染性蛋白质的蛋白质制剂。当实施方案的蛋白质或其生物活性部分用重组法产生时,最佳地,培养基具有少于约30%、约20%、约10%、约5%或约1%(以干重计)的化学前体或非受关注蛋白的化学品。与编码的核苷酸序列和蛋白质相关的片段和变体在本公开的范围内。“片段”是指核苷酸序列的一部分或由其编码的氨基酸序列以及蛋白质的一部分。核苷酸序列的片段可编码蛋白质片段,所述蛋白质片段保留天然蛋白质的生物学活性并具有赋予对植物的抗性(即,抗真菌性)的能力。作为另一者选择,可用作杂交探针的核苷酸序列片段不一定编码保持生物活性的片段蛋白质。因此,核苷酸序列的片段可在至少约15个核苷酸、约50个核苷酸、约100个核苷酸并且至多编码本公开的多肽的全长核苷酸序列的范围内。编码本公开的多肽的生物活性部分的核苷酸序列的片段可编码至少约15、约25、约30、约40、或45、约50个连续氨基酸或者至多该实施方案的全长多肽中存在的氨基酸的总数(例如对于由seqidno:1编码的肽而言,925个氨基酸)。可用作杂交探针或pcr引物的核苷酸序列的片段通常不必编码蛋白质的生物活性部分。当涉及指定的多核苷酸时,术语“全长序列”是指具有天然序列的整个核酸序列。“天然序列”在本文中用于表示内源性序列,即存在于生物体基因组中的非工程化序列。因此,本公开的核苷酸序列的片段可编码多肽的生物活性部分,或者它可以是可采用下文所公开的方法用作杂交探针或pcr引物的片段。赋予抗性的多肽的生物活性部分可通过分离实施方案的核苷酸序列之一的一部分,表达所述蛋白质的编码部分并评估蛋白质的编码部分在植物中赋予或增强真菌抗性的能力来制备。属于该实施方案的核苷酸序列的片段的核酸分子包含至少约15、约20、约50、约75、约100、或约150个核苷酸或者至多本文所公开的全长核苷酸序列中存在的核苷酸的数目(例如对于seqidno:1为2,778个核苷酸)。术语“变体”表示基本上类似的序列。对于多核苷酸,变体包含在天然多核苷酸中的一个或多个位点处的一个或多个核苷酸的缺失和/或添加,和/或在天然多核苷酸中的一个或多个位点处的一个或多个核苷酸的置换。如本文所用,“天然”多核苷酸或多肽分别包含天然存在的核苷酸序列或氨基酸序列。本领域技术人员可认识到该实施方案的核酸变体将被构造成使得开放阅读框得以保持。对于多核苷酸,保守变体包括那些这样的序列,其由于遗传密码的简并性而编码该实施方案的多肽中的一者的氨基酸序列。天然等位基因变体(如这些)可用公知的分子生物学技术进行鉴定,例如用下文所述的聚合酶链反应(pcr)和杂交技术来鉴定。变体多核苷酸还包括通过合成来源的多核苷酸,诸如那些例如通过使用定点诱变产生但仍编码该实施方案的蛋白质的那些多核苷酸。通常,如通过本领域熟知的序列比对程序所测定的,本公开的特定多核苷酸的变体可与该特定多核苷酸具有至少约40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或更高的序列同一性。该实施方案的特定多核苷酸(即参考多核苷酸)的变体还可通过比较变体多核苷酸所编码的多肽与该参考多核苷酸所编码的多肽之间的序列同一性百分比来进行评价。任何两条多肽之间的序列同一性百分比可用本领域已知的序列比对程序来计算。在任何给定的本公开多核苷酸对是通过比较它们编码的两个多肽所共享序列同一性百分比进行评价的情况中,其中两个被编码的多肽之间的序列同一性百分比为至少约40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或更高的序列同一性。“变体蛋白质”是指通过在天然蛋白质中的一个或多个位点处缺失或添加一个或多个氨基酸和/或在天然蛋白质的一个或多个位点处置换一个或多个氨基酸而从该天然蛋白质衍生的蛋白质。本公开的一些方面所涵盖的变体蛋白是具有生物活性的,也就是说它们继续拥有天然蛋白质的所需生物活性,即如本文所述的赋予或增强植物抗性(即,植物真菌病原体抗性)的能力。此类变体可例如由遗传多态性或由人类操纵而得到。如通过本领域已知的序列比对程序所测定的,本实施方案的天然蛋白质的生物活性变体可与该天然蛋白质的氨基酸序列具有至少约40%、约45%、约50%、约55%、约60%、约65%、约70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或更高的序列同一性。本公开的蛋白质的生物活性变体与该蛋白可相差少至约1-15个氨基酸残基,少至约1-10个(如约6-10个)、少至约5个、少至4个、3个、2个或甚至1个氨基酸残基。本文所公开的蛋白质可例如通过包括氨基酸置换、缺失、截短和插入来进行变更。此类操纵的方法在本领域中是已知的。例如,抗性蛋白的氨基酸序列变体和片段可通过在dna中作出突变来制备。诱变和多核苷酸变更的方法是本领域已知的。变体多核苷酸和蛋白质还涵盖由诱变和重组程序,包括但不限于诸如dna改组的程序获得的序列和蛋白质。可从一群包含具有实质序列同一性并可在体外或体内进行同源重组的序列区的相关序列多核苷酸产生重组多核苷酸的文库。例如,使用该方法,可使编码受关注结构域的序列基序在本公开的蛋白质基因和其它已知的蛋白质基因之间发生改组,以获得编码具有改善的受关注性质(诸如增加的赋予或增强植物对真菌病原体的抗性的能力)的蛋白质的新基因。这种dna改组的策略是本领域已知的。本文所述的多核苷酸可用来从其它生物,特别是其它植物分离相应的序列。以此方式,诸如pcr或杂交的方法可用来基于这类序列与本文所述序列的序列同源性来鉴定此类序列。本公开涵盖基于其与本文示出的整个序列或与其变体和片段的序列同一性而分离的序列。这类序列包括为所公开的序列的直系同源物的序列。术语“直系同源物”是指衍生自共同祖先基因并且由于物种形成而存在于不同物种中的基因。存在于不同物种中的基因,当它们的核苷酸序列和/或它们的编码蛋白质序列共享至少约60%、约70%、约75%、约80%、约85%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%或更大的序列同一性时,被认为是直系同源物。直系同源物的功能在各种物种中常常是高度保守的。因此,本公开涵盖编码赋予或增强真菌植物病原体抗性的蛋白质并且与本文所公开的序列杂交,或与其变体或片段杂交的分离的多核苷酸。在pcr方法中,可设计寡核苷酸引物用于pcr反应,以从提取自任何受关注生物体的cdna或基因组dna扩增相应的dna序列。设计pcr引物和pcr克隆的方法是本领域已知的,在以下文献中公开:sambrook等人,(1989)molecularcloning:alaboratorymanual(第2版,coldspringharborlaboratorypress,plainview,n.y.)。已知的pcr方法包括但不限于利用成对引物、巢式引物、单一特异性引物、简并引物、基因特异性引物、载体特异性引物、部分错配引物等的方法。在杂交技术中,将已知的多核苷酸的全部或一部分用作探针,该探针与来自所选生物体的一组克隆的基因组dna片段或cdna片段(即基因组或cdna文库)中存在的其它对应多核苷酸选择性杂交。该杂交探针可以是基因组dna片段、cdna片段、rna片段或其它寡核苷酸,并且可用可检测基团如32p或任何其它可检测标记物标记。因此,例如,杂交用探针可通过对基于该实施方案的多核苷酸的合成寡核苷酸进行标记来制备。制备杂交用探针和构建cdna文库和基因组文库的方法在本领域中是已知的。各种程序均可用于检查dna、rna或蛋白质的特定序列的存在或缺失。这些包括,例如,southern印迹、northern印迹、western印迹和elisa分析。这些技术在本领域中是熟知的。本公开的组合物和方法可用于调节植物中一种或多种蛋白质的含量。术语“调节”在本文中用于表示遗传学上改变的(即,转化的)植物中的蛋白质含量相对于来自相应的未转化的植物(即,未根据本公开的方法在遗传学上改变的植物)的蛋白质含量的增加或减小。如本文所用,术语“抑制”、“减少”等是指靶基因产物的表达或功能的任何减少,包括表达或功能的任何相对减少至多并包括完全消除靶基因产物的表达或功能。术语“增加”、“增强”等在本文中用于表示相比于易感植物,靶基因(例如,r基因)产物的表达、功能或活性的任何促进或增益或上升,从而提供对一种或多种病原体(例如,层锈菌属)或病害(例如,asr)的增加的抗性。另外,如本文所用,术语“引起”或“增加”可表示相对于缺乏本公开的靶基因或蛋白质的细胞或植物,靶基因产物的更高表达,使得所述水平增加10%或更多,50%或更多或100%。如本文所用,术语“表达”是指由此产生多核苷酸的生物合成或过程,包括所述基因产物的转录和/或翻译。例如,本公开的多核苷酸可由dna模板转录(诸如转录成mrna或其它rna转录物)和/或转录的mrna随后由此翻译成多肽或蛋白质的过程。术语“基因产物”可以指例如转录物和编码的多肽。基因产物(即,受关注的基因产物)的表达或功能的抑制(或增加)可处于任两种植物之间进行比较的环境中,例如遗传学上更改的植物中的基因产物的表达或功能相对于相应的但易感的野生型植物或其它易感植物中的该基因产物的表达或功能。野生型植物中的基因产物的表达水平可以不存在。例如,“野生型”植物可以是不表达外源nb-lrr核酸或外源nb-lrr蛋白的植物、植物细胞或植物部分。另选地,靶基因产物的表达或功能的抑制(或增加)可处于在同一植物内或植物之间的植物细胞、细胞器、器官、组织或植物部分之间进行比较的环境中,并且包括在同一植物或植物之间的发育或时间阶段之间的比较。任何在转录或翻译水平上减量调节靶基因产物的表达,或者减量调节靶基因产物的功能活性的方法或组合物可用于实现靶基因产物的表达或功能的抑制。类似地,任何在转录或翻译水平上诱导或增量调节靶基因产物的表达,或者增加或活化或增量调节靶基因产物的功能活性的方法或组合物可用于实现靶基因或蛋白质的增加的表达或功能。用于抑制或增强基因表达的方法是本领域熟知的。本公开的基因和多核苷酸包括天然存在的序列以及突变体或更改的形式。本文所公开的蛋白质还涵盖天然存在的蛋白质以及它们的变型、片段和修饰形式。此类变体和片段将继续具有赋予或增强植物真菌病原体抗性的所需能力。在一个方面,将在编码它们的变体或片段的dna中所作的突变一般不将序列设置在阅读框外,并且最佳地将不产生互补区域,该互补区域可能产生二级mrna结构。本公开的一种或多种基因可表达为转基因以便产生抗asr的植物。使用本文所述或本领域技术人员已知的不同启动子将使得基因的表达在不同情况下调节(即,启动子可基于期望的结果来选择)。例如,可能需要在特定组织系统或器官(例如,叶)中的较高的表达水平以增强抗性。可以转基因形式插入整个基因(例如,天然启动子和编码序列两者),从而允许与其它性状如抗虫性或抗除草剂性的快速组合。在本公开的一些方面中,核酸序列可与受关注的多核苷酸序列的任何组合叠加,以便形成具有期望的表型的植物。该叠加可通过dna构建体内的基因的组合,或通过将具有转基因的一种或多种植物与包含期望组合的另一种植物品系杂交来实现。例如,本公开的多核苷酸或其片段可与本公开的任何其它多核苷酸或其它基因叠加。所产生的组合还可包括所关注的多核苷酸中的任何一者的多个拷贝。本公开的多核苷酸还可与任何其它基因或基因的组合叠加以产生具有多种期望的性状组合的植物,所述性状包括但不限于动物饲料所期望的性状,诸如高油基因、平衡氨基酸、增加的消化率、抗虫性、抗病性或除草剂抗性、无毒性和抗病性基因、农学性状(例如,雄性不育、开花时间)和/或转化技术性状(例如,细胞周期调节或基因靶向)。这些叠加的组合可通过任何方法来产生,包括但不限于通过任何常规或已知方法或遗传转化来对植物进行杂交育种。如果性状是通过遗传转化植株来堆叠,则目的多核苷酸序列可在任何时间以任何顺序进行组合。例如,可将包含一种或多种期望性状的转基因植物用作靶标,通过后续的转化引入更多性状。在共转化方案中可将性状与受关注多核苷酸同时引入,所述多核苷酸由转化盒的任何组合提供。例如,如果将要引入两条序列,则可将该两条序列包含在单独的转化盒中(反式)或包含在同一转化盒中(顺式)。可通过相同启动子或不同启动子驱动所述序列表达。在某些情况下,期望引入转化盒,所述转化盒可抑制受关注的多核苷酸的表达。这可以与其它抑制盒或过度表达盒的任何组合进行组合以在植物中生成期望的性状组合。本公开的特征是包括将多核苷酸引入植物中的方法。如本文所用,术语“引入”是指对植物提供例如多核苷酸。在本公开的一些方面,多核苷酸可以使得序列进入植物细胞内部,包括其潜在地插入植物的基因组中这样的方式存在。本公开的方法不依赖于将序列引入植物中的具体方法,只要多核苷酸进入植物的至少一个细胞的内部即可。将多核苷酸引入植物的方法是本领域已知的,其包括并且不限于稳定转化方法、瞬时转化方法和病毒介导的方法。术语“转化”在本文中用于表示例如核酸片段转移至宿主生物体的基因组中,从而得到遗传上稳定的遗传性。含有转化的核酸片段的宿主生物体称作“转基因”生物体。术语“宿主细胞”是指其中发生重组dna构建体的转化的细胞并且可包括酵母细胞、细菌细胞和/或植物细胞。植物转化的方法的示例包括农杆菌属(agrobacterium)-介导的转化和粒子轰击,其然后可用于通过本领域技术人员已知的方法使转化的植物再生。可将多核苷酸瞬时或稳定地引入宿主细胞中,并且可以维持未整合,例如以质粒形式。“稳定转化”或“稳定地转化”是指被引入植物中的核苷酸构建体整合进植物的基因组中,并能够由其后代继承。如本文所用,“瞬时转化”意指将多核苷酸引入植物中但它没有整合到植物的基因组中,或者将多肽引入植物中。转化方法以及将多核苷酸序列引入植物中的方法,可取决于要进行转化的植物或植物细胞的类型,即单子叶植物或双子叶植物。将多肽或多核苷酸引入植物细胞中的合适方法包括但不限于微注射、电穿孔、直接基因转移、lec1转化和弹道粒子加速。随着更新的方法变得可用,其也可施用于本公开,因为转化或转染的方法并不是关键。可根据常规方式将已转化的细胞培育成植株。然后可使这些植株生长,并用同一转化株系或不同株系授粉,再鉴定出具有所需表型特征的组成型表达的子代。可培育两代或更多代植株,以确保所需表型特征的表达得到稳定保持和遗传。然后收获种子,以确保已实现所需表型特征的表达。在本公开的一些方面,将具有核苷酸构建体或表达盒的转化的种子或转基因种子稳定地并入其基因组中。在一个方面,本公开涵盖包含本文所公开的多核苷酸序列的种子,其可发育成或可用于使一种或多种植物发育,与例如野生型品种的植物种子比,所述植物具有对病原体(例如真菌)或由病原体形成的感染的增强的抗性。在一个方面,本公开的特征在于来自转基因豆类作物植物的种子,其中所述种子包含本文所公开的多核苷酸。本公开可用于转化任何植物物种,包括但不限于单子叶植物和双子叶植物。受关注的植物的示例包括但不限于玉米(zeamays)、芸苔属物种(brassicaspp.)(例如甘蓝型油菜(brassicanapus)、芜菁(brassicarapa)、芥菜(brassicajuncea)),特别是可用作种子油来源的那些芸苔属物种,苜蓿(medicagosativa)、水稻(oryzasativa)、黑麦(secalecereale)、高粱(sorghumbicolor,sorghumvulgare)、粟(例如珍珠粟(pennisetumglaucum)、黄米(panicummiliaceum)、谷子(setariaitalica)、龙爪稷(eleusinecoracana))、向日葵(helianthusannuus)、红花(carthamustinctorius)、小麦(triticumaestivum)、大豆(glycinemax)、烟草(nicotianatabacum)、马铃薯(solanumtuberosum)、花生(arachishypogaea)、棉花(海岛棉(gossypiumbarbadense)、陆地棉(gossypiumhirsutum))、甘薯(ipomoeabatatus)、木薯(manihotesculenta)、咖啡(coffeaspp.)、椰子(cocosnucifera)、菠萝(ananascomosus)、柑橘(citrusspp.)、可可(theobromacacao)、茶(camelliasinensis)、香蕉(musaspp.)、鳄梨(perseaamericana)、无花果(ficuscasica)、番石榴(psidiumguajava)、芒果(mangiferaindica)、橄榄(oleaeuropaea)、木瓜(caricapapaya)、腰果(anacardiumoccidentale)、澳洲坚果(macadamiaintegrifolia)、杏树(prunusamygdalus)、糖用甜菜(betavulgaris)、甘蔗(saccharumspp.)、燕麦、大麦、蔬菜、观赏植物和针叶树。在一个方面,受关注的植物包括豆类作物物种,其包括但不限于苜蓿(medicagosativa);三叶草或车轴草(三叶草属物种);豌豆,包括(豌豆(pisumsatinum))、木豆(gajanuscajan)、豇豆(vignaunguiculata)和山黧豆属(lathyrusspp.);菜豆(fabaceae或leguminosae);兵豆(lensculinaris);羽扇豆(lupinusspp.);牧豆树(prosopisspp.);长豆角(ceratoniasiliqua)、大豆(glycinemax)、花生(arachishypogaea)或罗望子(tamarindusindica)。术语“豆类物种”和“豆类作物物种”在本文中用于是指植物,并且可以例如为受关注的植物。在一些方面,豆类物种或豆类作物物种是植物、植物部分或植物细胞。术语“转基因”在本文中用于是指植物,包括来源于植物的任何部分,诸如细胞、组织或器官,其中通过基因工程方法,诸如农杆菌属转化,将外源性核酸(例如,包括一种或多种核酸的重组构建体、载体或表达盒)整合进基因组中。通过进行基因技术方法,将外源性核酸稳定整合进染色体中,使得下一代也可以是转基因的。如本文所用,“转基因”还涵盖生物处理,其包括植物杂交和/或天然重组。在一个方面,构建体或载体或表达盒不存在于初始植物的基因组中或存在于转基因植物的基因组中,但不在其初始植物的基因组的天然基因座处。本文所公开的组合物可通过基因掺入的方法来生成或维持。当所述方法重复两次或更多次时,基因掺入有时被称为“回交”。在基因掺入或回交时,“供体”亲本是指具有待渗入的所需基因或基因座的亲本植物。“受体”亲本(使用一次或多次)或“轮回”亲本(使用两次或更多次)是指基因或基因座被基因掺入到其中的亲本植物。初始杂交产生f1代;术语“bc1”则是指轮回亲本的第二次使用,并且“bc2”是指轮回亲本的第三次使用,等等。因此,本公开的一个方面是增强植物对植物病害诸如asr的抗性的方法。所述方法可包括通过在育种程序(即,抗asr的育种程序)中将豆类来源的nb-lrr抗性基因基因掺入种质中来赋予对病原体,例如导致asr的病原体的抗性。术语“种质”在本文中用于表示来自个体(例如植物)、一组个体(例如,植物种系、品种或家族)的遗传物质,或源自品系、品种、物种或培养物的克隆体。种质可以为生物体或细胞的部分,或可以从生物体或细胞分离。种质提供具有特定分子构成的遗传物质,所述特定分子构成提供有机体或细胞培养物的一些或全部遗传性质的物理基础。在本公开的上下文中,种质包括新植物可由此生成的细胞、种子或组织,或者可被培育成整株植物的植物部分诸如叶、茎、花粉、或细胞。本公开的方面包括用于鉴定作为抗性源的种质(包括但不限于以下属中一种或多种的种质)的方法:大豆属、豇豆属和扁豆属。如本文所述,豆类来源的nb-lrr型抗性基因赋予对大豆锈菌的不同反应。先前对大豆锈菌的六个澳洲种族的研究可例如基于其对野生型大豆的若干物种的相应相容性或不相容性来辨别(burdon和marshall(1981)journalofecology,69:381-390;burdon和marshall(1981)plantdisease,65:44-45;burdon和speer(1984)euphytica,33:891-896;burdon(1987)ocecologia,73(2):257-267;burdon(1988)theor.appl.genet.,75:923-928;以及jarosz和burdon(1990)heredity,64:347-353)。因此,在一个方面,豆类作物物种或豆类来源的基因源自大豆属。大豆属物种的示例包括但不限于沙生大豆(glycinearenaria)、银毛大豆(glycineargyrea)、弯裂片大豆(glycinecyrtoloba)、灰毛大豆(glycinecanescens)、澎湖大豆(glycineclandestine)、弯荚大豆(glycinecurvata)、镰荚大豆(glycinefalcata)、宽叶大豆(glycinelatifolia)、小叶大豆(glycinemicrophylla)、glycinepescadrensis、glycinestenophita、glycinesyndetica、野大豆(glycinesoja)、烟豆(glycinetabacina)和短绒野大豆(glycinetomentella)。其它属,诸如豇豆属和扁豆属也显示出对大豆锈菌的不同反应。因此,在一个方面,豆类作物物种或豆类来源的基因源自豇豆属。豇豆属为包含约100个种的泛热带属。其是再分成以下亚属的分类群:豇豆属(vigna)、haydonia、plectotropis(非洲)、ceratotropis(亚洲)、sigmoidotropis和lasiopron。所述属包括经济型相关物种,诸如vignaunguiculata(l.)walp(豇豆)、vignaradiata(l.)wilczek(绿豆)、vignaangularis(赤豆)ohwi和ohashi(赤小豆)、vignamungo(l.)hepper(黑吉豆)、和vignaumbellata(赤小豆)ohwi和ohashi(饭豆)。豇豆中的四种亚种是公知的:dekindtiana,培育的亚种的野生近缘种;眉豆(cylindrica),培育的乌豇豆;长豇豆(sesquipedalis),培育的长豇豆;和豇豆(unguiculata),培育的黑眼豆。豇豆属物种豇豆(unguiculata)进一步分成以下品种群:豇豆,其作为豆类种植;biflora或cilindrica(乌豇豆),主要用作饲料;长豇豆(yardlong或豇豆),其作为蔬菜种植;textilis,其培养为长花序梗的纤维;以及melanophthalmus(黑眼豆)。已经在田间和温室条件下记录了若干豇豆属物种,包括绿豆(vignaradata)、黑吉豆(vignamungo)和豇豆(vignaunguiculata)对大豆锈菌的易感性。在一个方面,豆类作物物种或豆类来源的基因源自扁豆属。扁豆(lablabpurpureus(l.)sweet)(也被称为dolichosbenghalensisjacq.、dolichoslablabl.、dolichospurpureusl.、lablabnigermedikus、lablabpurpurea(l.)sweet、lablabvulgaris(l.)savi、vignaaristatapiper)是原产于亚洲和非洲的豆类物种(verdcourt(1971)floraoftropicaleastafrica,第696-699页,crownagents,london,uk;和duke等人,(1981),handbookoflegumesofworldeconomicimportance,第102-106页,plenumpress,newyork,usa和london,uk)(pengelly和maass,(2001)gen.resour.cropev.48:261-272)。其通常被称为鹊豆、扁豆、紫扁豆、蚕豆、埃及豆、穷人豆(poorman′sbean)、香翅豆(tongabean)(英国),和至少20个附加的俗名。其在非洲、亚洲和加勒比地区作为豆类作物或作为绿色蔬菜种植(duke等人,(1981)handbookoflegumesofworldeconomicimportance,第102-106页,plenumpress,newyork,usa和london,uk);以及pengelly和maass,(2001)gen.resour.cropev.48:261-272)。已经报道了鹊豆(lablabpurpureus)作为大豆锈菌的可选的宿主(pérez-hernández,(2007)alternativehostsofphakopsorapachyrhiziintheamericas:ananalysisoftheirroleintheepidemiologyofasiansoybeanrustinthecontinentalu.s.m.sc.thesis.iowastateuniversity.ames,iowa.u.s.a.;vakili(1981)plantdis.65:817-819;以及poonpolgul和surin,(1980)soybeanrustnewsletter,3:30-31)。在一个方面,豆类作物物种或豆类来源的基因源自鹰嘴豆属、木豆属、苜蓿属、菜豆属、豌豆属、葛属或三叶草属。鹰嘴豆属物种的示例包括但不限于,鹰嘴豆、cicerechinospermum、cicerreticulatum和cicerpinnatifidum。木豆属物种的示例包括但不限于木豆。苜蓿属物种的示例包括但不限藜苜蓿(medicagotruncatula)和紫花苜蓿(medicagosativa)。菜豆属的示例包括但不限于,菜豆(phaseolusvulgaris)、棉豆(phaseoluslunatus)、宽叶菜豆(phaseolusacutifolius)或红花菜豆(phaseoluscoccineus)。豌豆属物种的示例包括但不限于pisumabyssinicum、菜豌豆(pisumsativum)、pisumelatius、pisumfulvum、pisumtranscaucasium或pisumhumile。葛属物种的示例包括但不限于葛根。三叶草属物种的示例包括但不限于黄车轴草(trifoliumaureum)和trifoliumoccidentale。本公开还可包括本文所述的序列,其可在受关注的植物中表达的表达盒或dna构建体中提供。在一个方面,该表达盒可包括与本文所公开的序列可操作地连接的5′和3′异源调控序列。术语“可操作地连接”在本文中用于表示以允许核酸表达的方式(即,当将载体引入宿主植物细胞中时,在宿主植物细胞中)将待表达的核酸连接至调控序列,包括启动子、终止子、增强子和/或其它表达控制元件(例如,聚腺苷酸化信号)。此类调控序列是本领域中所熟知的并且包括在多种宿主细胞中直接组成型表达核苷酸序列的那些,和在特定宿主细胞中或在特定条件下直接表达核苷酸序列的那些。载体的设计可取决于例如待转化的宿主细胞的类型,或期望的核酸表达水平。所述表达盒可含有一个或多个待共转化到植物中的附加基因。并且,可在多个表达盒上提供任何附加基因。本公开的表达盒可包括多个限制位点用于插入该核苷酸序列以便处于调节区的转录调节之下。该表达盒还可含有选择性标记基因。表达盒还可在5′-3′转录方向上包括在植物中起作用的转录和翻译起始区、本公开的dna序列、以及转录和翻译终止区。转录起始区,启动子,相对于植物宿主,可以是天然的或类似的,或是外来或异源的。另外,该启动子可以是天然序列或另选地是合成序列。术语“异源的”是指转录初始区不存在于所述转录初始区被引入其中的天然植物中。如本文所用,嵌合基因包含与转录起始区可操作连接的编码序列,该转录起始区对于该编码序列是异源的。启动子的示例包括但不限于,花椰菜花叶病毒35s和大豆泛素6。虽然可优选使用异源启动子来表达序列,但可以使用同源启动子或天然启动子序列。此类构建体将改变宿主细胞(即,植物或植物细胞)中的表达水平。因此,改变宿主细胞(即,植物或植物细胞)的表型。终止区可以天然具有转录起始区,天然具有可操作地连接的受关注的dna序列,或源自另一个源。易得的终止区可获自根瘤农杆菌(agrobacteriumtumefaciens)的ti质粒,如章鱼碱合酶和胭脂碱合酶终止区。在一个方面,内源性或源基因抗性直系同源物可通过同源或非同源重组方法,诸如例如通过基因组编辑来更改。当与其未修饰的序列相比时,此类更改是指核苷酸序列具有至少一种修饰,并且包括例如:(i)置换至少一个核苷酸,(ii)缺失至少一个核苷酸,(iii)插入至少一个核苷酸,或者(iv)(i)-(iii)的任何组合。在一些实施方案中,可使用基因组编辑技术将本发明所公开的nb-lrr多核苷酸组合物引入植物的基因组中,或者可以使用基因组编辑技术来编辑先前引入植物基因组中的nb-lrr多核苷酸。基因组编辑可使用任何可用的基因编辑方法来实现。例如,基因编辑可通过将多核苷酸修饰模板(有时被称为基因修复寡核苷酸)引入宿主细胞中来实现,其中所述多核苷酸修饰模板包括对宿主细胞基因组内的基因的靶向修饰。多核苷酸修饰模板可以为单链或双链。例如,参见美国专利公布2013/0019349。在一些实施方案中,基因编辑可通过在期望更改附近的基因组中的限定位置处引入双链断裂(dsb)来进行。dsb可使用任何可用的dsb诱导剂来诱导,其包括但不限于talen、归巢核酸内切酶、锌指核酸酶、cas9-grna体系(基于细菌crispr-cas9体系)等。在一些实施方案中,dsb的引入可以与多核苷酸修饰模板的引入组合。用于编辑基因序列的方法可包括将dsb与多核苷酸修饰模板组合并且一般来讲还包括:1)向宿主细胞提供dsb-诱导剂、或编码dsb诱导剂的核酸,其中所述dsb诱导剂识别染色体序列中的靶序列,并且因此能够在基因序列中诱导dsb;和2)一种或多种多核苷酸修饰模板,其包含与待编辑的核苷酸序列相比的一个或多个核苷酸更改。多核苷酸修饰模板还可包含旁接一个或多个核苷酸更改的核苷酸序列,其中所述旁侧序列与旁接dsb的染色体区基本上同源。使用dsb诱导剂诸如cas9-grna络合物的基因组编辑技术是本领域中已知的(参见,例如,2014年8月20日提交的美国专利申请14/463,687,2014年8月20日提交的pct专利申请pct/us14/51781,和2014年8月13日提交的美国专利申请62/036,652;这些专利均以引用方式并入本文)。向导多核苷酸/cas内切核酸酶体系也是本领域中已知的(参见,例如,2014年8月20日提交的美国专利申请14/463,691,其以引用的方式并入本文)。向导rna/cas内切核酸酶体系的附加使用描述于2014年8月20日提交的美国专利申请14/463,687和14/463691中,并且包括但不限于修饰或置换受关注的核苷酸序列(例如,调控元件),插入受关注的多核苷酸、基因敲除、基因敲入、接合位点的修饰和/或引入另选的接合位点、修饰编码受关注的蛋白质的核苷酸序列、氨基酸和/或蛋白质融合、和通过将反向重复序列表达到受关注的基因中使基因沉默。根据需要,可优化一种或多种基因以增加在转化的植物中的表达。换句话讲,可使用植物优选的密码子来合成基因以改善表达。用于合成植物优选的基因的方法是本领域已知的。已知有另外的序列修饰能增强细胞宿主中的基因表达。这些序列修饰包括消除以下序列:编码假多腺苷酸化信号、外显子-内含子剪接位点信号、转座子样重复序列,以及其它此类得到充分表征的可能对基因表达有害的序列。可将序列的g-c含量调整到给定的细胞宿主的平均水平,该平均水平通过参考在宿主细胞中表达的已知基因计算得到。当可能时,修饰该序列以避免可预见的发夹二级mrna结构。表达盒可以在表达盒构建体中额外含有5′前导序列。此类前导序列可以起到增强翻译的作用。翻译前导序列是本领域已知的,并且包括:小核糖核酸病毒前导序列,例如emcv前导序列(脑心肌炎5′非编码区);马铃薯y病毒组前导序列,例如tev前导序列(烟草蚀纹病毒),和人免疫球蛋白重链结合蛋白(bip);来自苜蓿花叶病毒的外壳蛋白mrna(amvrna4)的非翻译前导序列;烟草花叶病毒前导序列(tmv);以及玉蜀黍枯黄斑点病毒前导序列(mcmv)(lommel等人,(1991)virology81:382385)。也可利用其它已知的用于增强翻译的方法,诸如内含子。可在制备表达盒时操纵各种dna片段,以确保dna序列处于正确方向,并且视情况而定,在适当的阅读框中。为此,可以使用衔接子或接头来连接dna片段。另选地,可使用其它操纵来提供便利的限制位点、移除多余的dna、或移除限制位点。出于这个目的,可能涉及体外诱变、引物修复、限制、退火、再置换(例如转换和颠换)。一般来讲,表达盒可包含用于选择转化的细胞的选择性标记基因。选择性标记基因用于选择转化的细胞或组织。标记基因包括编码抗生素抗性的基因,诸如那些编码新霉素磷酸转移酶ii(neo)和潮霉素磷酸转移酶(hpt)的基因,以及赋予对除草化合物的抗性的基因,所述除草化合物为诸如草铵膦、溴草腈、咪唑啉酮和2,4-二氯苯氧基乙酸(2,4-d)。以上选择性标记基因的列表并不意指是限制性的。任何选择性标记基因均可用于本公开。为了在植物或植物细胞中表达本公开的靶基因和/或蛋白质(例如一个或多个nb-lrr基因和/或一个或多个r蛋白),本文所述的方法包括用(例如,如本文所公开的)编码靶标r蛋白的多核苷酸转化植物或植物细胞。本文所述的多核苷酸可以可操作地连接至启动子,所述启动子驱动植物细胞中的表达。本领域中已知的任何启动子均可用于本公开的方法中,其包括但不限于组成型启动子、病原体诱导型启动子、伤口诱导型启动子、组织优选的启动子和化学调节启动子。启动子的选择可取决于转化的植物中期望的表达时间和位置,以及本领域技术人员已知的其它因素。可种植或培育转化的细胞或植物以形成一种植物,所述植物包含被引入例如编码r蛋白的细胞或植物中的多核苷酸中的一种或多种。多种启动子可用于实施本公开。可根据期望的结果来选择启动子。即,可将核酸与组成型启动子、组织优选的启动子、或其它启动子进行组合,以在受关注的宿主细胞中表达。此类组成型启动子包括(例如)rsyn7启动子的核心启动子和wo99/43838和美国专利6,072,050中公开的其他组成型启动子;芯camv35s启动子;水稻肌动蛋白;泛素;pemu;mas;als等。其它组成型启动子包括例如以下美国专利中描述的那些启动子:5,608,149、5,608,144、5,604,121、5,569,597、5,466,785、5,399,680、5,268,463、5,608,142、和6,177,611,其在本领域中是已知的,并且可设想用于本公开中。一般来讲,从诱导型启动子,特别是从病原体诱导型启动子表达基因可能是有利的。这种启动子包括来自发病相关蛋白(pr蛋白)的那些,所述蛋白在病原体感染后诱导,例如pr蛋白、sar蛋白、β-1,3-葡聚糖酶、壳多糖酶等等。在病原体感染位点处或附近局部表达的启动子值得关注。另外,因为病原体可通过伤口或昆虫病变进入植物中,可将伤口诱导型启动子用于本公开的构建体中。此类伤口诱导型启动子包括马铃薯蛋白酶抑制剂(pinii)基因、wun1和wun2、win1和win2、系统素(systemin)、wip1、mpi基因等。可将化学调控启动子用于通过施用外源化学调控剂来调节基因在植物中的表达。取决于目标,启动子可以是化学诱导型启动子,其中施用化学物质诱导了基因表达,或者是化学阻遏型启动子,其中施用化学物质抑制了基因表达。化学诱导型启动子是本领域已知的,并且包括但不限于玉蜀黍in2-2启动子(其通过苯磺酰胺除草剂安全剂激活)、玉蜀黍gst启动子(其通过用作前突生除草剂的疏水性亲电化合物激活)和烟草pr-1a启动子(其通过水杨酸激活)。受关注的其它化学调节的启动子包括甾类化合物响应性启动子(例如,糖皮质素诱导型启动子、和四环素诱导型启动子和四环素阻遏型启动子)。组织优选的启动子可用于靶基因或蛋白质(例如,编码豆类来源的nb-lrr多肽的多核苷酸序列)在特定植物组织内的靶向增强的表达。此类组织优选的启动子包括但不限于叶优选的启动子、根优选的启动子、种子优选的启动子和茎优选的启动子。组织优选的启动子包括yamamoto等人,(1997)plantj.12(2):255-265(yamamoto等人,1997年,《植物杂志》,第12卷,第2期,第255-265页);kawamata等人,(1997)plantcellphysiol.38(7):792-803;hansen等人,(1997)mol.gengenet.254(3):337-343;russell等人,(1997)transgenicres.6(2):157-168;rinehart等人,(1996)plantphysiol.112(3):1331-1341;vancamp等人,(1996)plantphysiol.112(2):525-535;canevascini等人,(1996)plantphysiol.112(2):513-524;yamamoto等人,(1994)plantcellphysiol.35(5):773-778;lam,(1994)resultsprobl.celldiffer.20:181-196;orozco等人,(1993)plantmolbiol.23(6):1129-1138;matsuoka等人,(1993)procnatl.acad.sci.usa90(20):9586-9590;以及guevara-garcia等人,(1993)plantj.4(3):495-505。此类启动子可被修饰。叶特异性启动子是本领域已知的。参见例如,yamamoto等人,(1997)plantj.12(2):255-265;kwon等人,(1994)plantphysiol.105:357-67;yamamoto等人,(1994)plantcellphysiol.35(5):773-778;gotor等人,(1993)plantj.3:509-18;orozco等人,(1993)plantmol.biol.23(6):1129-1138;以及matsuoka等人,(1993)proc.natl.acad.sci.usa90(20):9586-9590。“种子优选的”启动子包括“种子特异性”启动子(那些启动子在种子发育期间活跃,诸如种子贮藏蛋白的启动子)和“种子萌发”启动子(那些启动子在种子萌发期间活跃)。此类种子优选的启动子包括但不限于cim1(细胞分裂素诱导信息)、cz19b1(玉蜀黍19kda玉米醇溶蛋白)、mi1ps(肌醇-1-磷酸合酶)和cela(纤维素合酶)(参见wo00/11177,其以引用的方式并入本文)。γ-玉米醇溶蛋白是优选的胚乳特异性启动子。球蛋白-1(glob-1)是优选的胚特异性启动子。对于双子叶植物而言,种子特异性启动子包括但不限于菜豆β-菜豆蛋白基因启动子、油菜籽蛋白(napin)基因启动子、β-伴球蛋白基因启动子、大豆凝集素基因启动子、十字花科蛋白基因启动子等。对于单子叶植物,种子特异性启动子包括但不限于玉蜀黍15kda玉米醇溶蛋白、22kda玉米醇溶蛋白、27kda玉米醇溶蛋白、g-玉米醇溶蛋白、waxy、shrunken1、shrunken2、球蛋白1等。还可参见wo00/12733,其中公开了来自end1和end2基因的种子偏好启动子;该专利以引用方式并入本文。本公开的多核苷酸的表达可涉及使用完整的天然r基因,其中所述表达由同源5′上游启动子序列驱动。另选地,表达可使用与由宿主豆类中表达的异源nb-lrr抗病性基因提供的5′转录对照序列组装的构建体产生。本领域技术人员将能够鉴定编码nb-lrr蛋白质的基因,以评价其表达水平,并且选择可用于表达受关注的r基因的优选的启动子序列。使用同源或异源nb-lrr启动子序列提供调节蛋白质表达的选择,以避免或最小化与不适当或不期望的表达和植物防御活化相关的任何潜在的不期望的结果。特异性大豆启动子包括但不限于大豆泛素(subi-1)、延长因子1a和用于组成型表达的s-腺苷甲硫氨酸合酶和rpp4、rpg1-b以及包含在基因模型中的启动子,例如本领域技术人员已知用于更紧密地调节由nb-lrr基因启动子提供的表达的glyma启动子。本公开还包括用于本文所述的测定的试剂盒。多肽序列和多核苷酸还可作为试剂盒的组成与说明书一起包装,用于完成本文所公开的测定。本公开的试剂盒可包括本文所述的多肽和/或多核苷酸和适宜的说明书(书面和/或作为音频、视觉或视听材料提供)的任何组合。在一个实施方案中,试剂盒涉及用于鉴定抗asr的r基因(例如,nb-lrr基因)或r蛋白的dna检测试剂盒。提供利用本文所公开的序列中的任一种来鉴定植物中的转基因事件(例如,ccrpp1)的抗asr功效的试剂盒。例如,试剂盒可包括特定的探针,所述特定的探针具有的序列对应于以下序列或与以下序列互补,所述序列与转基因事件的特定区域具有介于80%和100%之间的序列同一性。试剂盒可包括进行测定或检测方法所需的任何试剂和材料。实施例实施例1:ccrpp1的定位和克隆筛选对大豆锈菌具有易感性的变型的非大豆物种的种质。筛选最初得自国际半干旱热带研究中心(icrisat)并引入巴西的巴西木豆(cajanuscajan)的品种,其已经针对期望的农学性状进行选择(godoy等人,(2005)rev.bras.zootec;34:7-19;和provazi等人,(2007)rev.bras.zootec;36:328-334)。木豆是二倍体豆类,其基因组大小为约830mbp(varshney等人,(2012)nat.biotechnol.,30:83-89),是自花授粉的,并且具有介于2-3个月的种子间生命周期。这些品种对大豆锈菌的不同反应允许鉴定抗性和易感品种(noriega,(2007)deplantashospedeirasedegenesdiferencialmenteexpressosnasoja-phakopsorapachyrhizi.m.sc.thesis.universidadefederaldebrazil)。将来自抗性品种的植物与易感品种的那些植物杂交,并且将所得的f1植物自花授粉。筛选所得f2子代的抗性和易感性。选择基因型g119-99(抗性基因ccrpp1的源)、g59-95、g146-97、g108-99、g127-97和g184-97的ccrpp(抗大豆锈菌的木豆抗性)基因用于进一步表征。测量若干木豆品种针对大豆锈菌分离株ppufv02的不同反应。反应的范围从抗性、具有斑点的部分抗性、红棕色病变但无孢子形成、红棕色病变且具有孢子形成到易感且具有“棕褐色”病变并在小叶轴侧上具有丰富的孢子形成。该基因型g119-99的抗性在f2群体中分离成3∶1指示其受单一显性基因控制。g108-99是例外,并且在f2中显示出15∶1分离比率,从而指示两个r基因可提供抗性。木豆中的ccrpp1遗传区域。为了将ccrpp1基因座的遗传定位,从来自杂交g119-99×g48-95的50种易感和50种抗性木豆f2植物体系中提取mrna。smarttm试剂盒和smartiv有专利权的寡核苷酸(clontech)用于第一链cdna合成。使用双重特异性核酸酶方法将由抗性和易感性体系产生的cdna归一化,以便有助于鉴定并分析稀有转录物并增强cdna文库的基因发现率(zhulidov等人,(2004)nucleicacidsres.32:e37;和zhulidov等人,(2005)bioorgkhim.31:186-94)。然后用sfii消化归一化的cdna并且通过chromaspin-1000柱进行粒度分级以选择超过1kb的转录物片段。随后使用covariss20剪切抗性和易感的本体cdna以获得200bp的平均片段尺寸。这些经剪切的cdna用于形成文库,所述文库用illuminaga2序列分析仪测序。在连接至ccrpp1基因座的易感和抗性cdna本体之间鉴定若干单核苷酸多态性。这些多态性用于形成精细作图的标记。标记dcaps140555连接至2282个被分析配子中的ccrpp1基因座(1141种f2植物)。因为在形成作图数据时没有可用的参照木豆基因组,所以进行了木豆与蒺藜苜蓿和大豆的同线性分析。为此,使用blast算法将来自遗传上连接至ccrpp1的七个标记物的序列与蒺藜苜蓿基因组(蒺藜苜蓿测序资源-mt3.0)和大豆基因组(phytozomev5.0)进行比对。大豆是一种古老的异源四倍体,并且染色体9和12是同源的。有意思的是,木豆中鉴定出的七个连接的标记显示出与大豆基因组中的两个区域的强同线性;一个在染色体12上,并且一个在染色体9上。还鉴定出了蒺藜苜蓿的染色体4上的同源区。为鉴定标记开发的附加多态性和插入缺失,将从含有经鉴定的同源区的染色体12上的端粒区起始的2mb区用作参照。该参照用于比对来自木豆抗性亲本品种g119-99读数的转录illumina数据,所述读数与由50个f2品系的抗性本体产生的读数组合,并与得自易感本体的读数进行比对。使用该方法,获得五种多态性和插入缺失序列,这提供ccrpp1的精细作图的额外标记(dcaps3978、ssr10581、dcaps52491、ssr2152、dcaps239615;图1)。这些标记序列中的大部分位于大豆的染色体12上的1.5mb区域内。值得注意的是,dcaps140555标记(目前连接至木豆中的ccrppl基因)距大豆的经典cc-nb-lrr基因(glyma12g01420)6kb定位并且位于由dcaps52491和ssr2152标记序列界定的106kb遗传区间中(图1)。来自参照木豆品种“asha”的bac克隆体的测序揭示了ccrpp1基因座包含来自nb-lrr基因家族的一个或多个基因。为此,使用源自标记dcaps140555并鉴定为97kb的bac克隆体的探针从经测序参照品种“asha”筛选bac文库(varshney等人,(2009)nat.biotechnol.,30:83-89)。将该克隆体剪切以获得使用sanger测序法进行测序的5kb的平均片段尺寸。读数随后使用phred、phrap和consed数据分析来组装。该bac的序列注释揭示存在四个高度同源的nb-lrr基因旁系同源物、三个全长基因和一个截短基因。高分辨率图谱揭示ccrpp1位于标记dcaps52491和ssr2152之间并且非常紧密地连接至dcaps140555。使用具有ssr、cap和dcap标记的多于1114种单独的f2植物,将ccrpp1基因座作图到g119-99中的染色体5。含有ccrpp1的区域缩小至涵盖dcaps52491(6个重组体/2282个配子)和ssr2152(1个重组体/2282个配子)标记的<154kb区域(图2a)。将这些标记紧密连接至ccrpp1,其中dcaps52491和ssr2152分别距r基因座0.26cm和0.04cm(图2a)。观察到侧接标记dcaps52491和ssr12872,两个功能增益重组体,并且观察到侧接标记caps20006和ssr2152,1个功能丧失重组体。因此,功能增益区间由标记dcaps52491和dcaps239615来界定,功能丧失区间由标记dcaps52491和ssr2152来界定(图2a),这证实ccrpp1基因座位于含有nb-lrr基因的区域中。实施例2:来自g119-99的ccrpp1基因座包含nb-lrr家族的四个成员为了确定g119-99中的ccrpp1基因座的基因组织形式,使用分别源自标记dcaps140555和ssr2152的两个探针产生并筛选g119-99bac文库。鉴定出两个阳性bac克隆体(3f和6g),其一起跨越介于dcaps52491和ssr2152之间的整个区间(图2b)。使用限制性酶hindiii经由dna指纹图谱测试包含bac3f(145kb)和bac6g(130kb)的多个克隆体的完整性。此外,这些克隆体的bac尺寸通过经noti消化的dna的脉冲场凝胶电泳来验证。利用pacbio和illuminamiseq对通过上述质量控制的每个3f和6g的一个克隆体进行测序,以使得每个bac序列能够快速且准确的组装(koren等人(2012)naturebiotechnology30:693-970)。将两个bac序列组装成一个205,344kb的大重叠群(图2b)。鉴定该重叠群中的四个nb-lrr候选基因序列;bac3f携带3个nb-lrr基因序列,nb-lrr-1、nb-lrr-2和nb-lrr-3(分别为seqidno:3、1和5),并且bac6g携带4个nb-lrr基因序列,nb-lrr-1至nb-lrr-4(seqidno:3、1、5和7)。使用来自非微生物攻击型g119-99基因型的转录物illumina数据,仅与trinity组装重头测序,发现nb-lrr-2的全长转录物。southern印迹分析示出g119-99中的ccrpp1基因座包含nb-lrr基因家族的四个成员,从而证实了bac基因注释。实施例3:用木豆nb-lrr-2基因(seqidno:1)转化大豆植物转化构建体经设计用于提供nb-lrr-2(seqidno:1)在大豆中的高水平组成型表达。包含nb-lrr-2编码区的2775bpsfii片段在5′端处连接至1948bp大豆泛素启动子+内含子i片段,并且在3′端上连接至888bp拟南芥属泛素终止子片段。整个启动子编码区终止子盒位于基植物表达载体中的attr1和attr2重组位点之间。除了上述元件,该载体还包含用于细菌选择的潮霉素抗性基因和作为植物选择性标记的除草剂抗性大豆als基因。将最终的nb-lrr-2植物表达载体电穿孔到大肠杆菌(escherichiacoli)中。然后选择转化体,并且通过标准微量制备方法分离pdna。转化体的特征在于微量制备的dna的诊断限制性酶消化。选择包含消化带的预期模式的阳性克隆体,并且将分离的dna提交用于基因枪转化。大豆的基因枪转化通过使用bioradbiolisticpds1000/he仪器和质粒或片段dna的粒子枪轰击法(美国专利号4,945,050)生成转基因大豆品系。使用以下原液和培养基进行大豆植物的转化和再生。原液:硫酸盐100x原液(37.0gmgso4.7h2o,1.69gmnso4.h2o,0.86gznso4.7h2o,0.0025gcuso4.5h2o);卤化物100x原液(30.0gcacl2.2h2o,0.083gki,0.0025gcocl2.6h2o);p、b、mo100x原液(18.5gkh2po4,0.62gh3bo3,0.025gna2moo4.2h2o);feedta100x原液(3.724gna2edta,2.784gfeso4.7h2o);2,4d原液(10mg/ml)和b5维生素,1000x原液(100.0g肌醇、1.0g烟酸、1.0g吡哆素hcl和10g硫胺素hcl。培养基(每升):sb199固体培养基(1包ms盐(gibco/brl;目录号11117-066)、1mlb5维生素1000x原液、30g蔗糖、4ml2,4-d(40mg/l终浓度),ph7.0,2g结冷胶);sb1固体培养基(1包ms盐(gibco/brl;目录号11117-066)、1mlb5维生素1000x原液、31.5g葡萄糖、2ml2,4-d(20mg/l终浓度),ph5.7,8gtc琼脂);sb196(上述原液1-4各10ml、1mlb5维生素原液、0.463g(nh4)2so4、2.83gkno3、1ml2,4-d原液、1g天冬酰胺、10g蔗糖,ph5.7);sb71-4(甘博格(gamborg)b5盐,20g蔗糖、5gtc琼脂,ph5.7);sb103(1pk.murashige&skoog盐混合物、1mlb5维生素原液、750mgmgcl2六水合物、60g麦芽糖、2g结冷胶tm,ph5.7);和sb166(补充有5g/l活性炭的sb103)。大豆胚芽发生悬浮培养物的引发。在种植45-55天后从可用的大豆植株挑取带有未成熟种子的豆荚,剥壳并放入无菌绛红色盒子中。将大豆种子在含有1滴ivorytm皂的5%溶液(即,95ml的高压灭菌蒸馏水加5ml和1滴皂,充分混合)中振摇15分钟,对它们进行灭菌。用2l无菌蒸馏水清洗种子,将小于3mm的种子放在单独的显微镜载玻片上。切开种子的小的一端,将子叶挤出种皮。将子叶转移到含有sb199培养基的平板上(每个平板25-30个子叶)2周,然后转移到sb1上2-4周。用纤维带包住平板。这个时间后,切取次生胚并放入sb196液体培养基中7天。培养条件:将大豆胚发生悬浮培养物在100-150rpm、26℃的回转摇床上保持在50ml液体培养基sb196中,光周期为16:8小时白天/黑夜,光强度为80-100μe/m2/s。每7-14天通过将最多1/2硬币大小数量的组织(组织块聚团在一起)接种到50ml新鲜液体sb196,进行培养物继代培养。用于轰击的dna的制备。在粒子枪轰击程序中,可能使用纯化的整个质粒dna或仅包含一个或多个受关注的重组dna表达盒的dna片段。对于每十七次轰击转化,制备85μl悬浮液,其中含有每个dna质粒的每碱基对1至90微微克(pg)的质粒dna。按照如下方式将dna质粒或片段共沉淀到金粒子上。将悬浮液中的dna添加到50μl的10-60mg/ml0.6μm金粒子悬浮液中,然后与50μlcacl2(2.5m)和20μl亚精胺(0.1m)混合。将混合物涡旋5秒,在微量离心机中短暂离心5秒,然后移除上清液。然后用150μl的100%乙醇洗涤dna包被的粒子一次,涡旋并再次在微量离心机中短暂离心,然后重悬于85μl的无水乙醇。然后将五微升的dna包被的金粒子加载至每个巨载体盘上。组织制备和用dna轰击。将大约100mg的两周龄悬浮培养物置于空的60mm×15mm培养皿中,用移液管从组织移除残余液体。将组织距离阻滞屏(retainingscreen)大约3.5英寸放置,将每个平板的组织轰击一次。将膜破裂压力设定为650psi,并将室抽至28英寸汞柱的真空。轰击后,将每个平板中的组织分至两个培养瓶,放回进液体培养基中,并如上所述进行培养。转化胚的选择与植物再生。轰击后,将每个经轰击的平板中的组织分放至sb196液体培养维持培养基的两个培养瓶(单位平板的经轰击的组织)。轰击后七天,用补充有100ng/ml选择剂的新鲜sb196培养维持培养基(选择培养基)更换每个培养瓶中的液体培养基。对于转化的大豆细胞的选择,所用选择剂为磺酰脲(su)化合物,其化学名为2-氯-n-((4-甲氧基-6-甲基-1,3,5-三嗪-2-基)氨基羰基)苯磺酰胺(其它常用名为dpx-w4189和氯磺隆)。氯磺隆为dupont磺酰脲类除草剂,中的活性成分。两周一次更换包含su的选择培养基并持续8周。8周选择期之后,观察到绿色转化组织的岛状物从未转化的坏死的胚发生簇长出来。将这些推定转基因事件分离并保持在含有100ng/mlsu的sb196液体培养基中另外5周,并且每1-2周更换一次培养基,从而产生新的无性繁殖的转化胚发生悬浮培养物。胚与su接触总共约13周。然后将悬浮培养物继代培养,并作为未成熟胚簇维持,并且还通过使单独体细胞胚成熟和发芽而再生成完整植株。在成熟培养基上四周(在sb166上1周,接着在sb103上3周)后,体细胞胚变得适于萌发。然后将其从成熟培养基中移除,并且在空培养皿中干燥并持续最多七天。将干燥的胚种植到sb71-4培养基中,在培养基中允许它们在与如上所述相同的光照和温度条件下萌发。将萌发的胚转移到盆栽基质上并生长至成熟以产生种子。实施例4:测试转基因植物的抗asr功效通过将植物表达构建体转化到大豆中,之后用大豆锈菌接种转基因植物,并且对植物病害症状进行评分来测试nb-lrr-2基因的抗asr功效。从大豆转化实验回收总共3个转基因事件,并且通过qpcr确认包含nb-lrr-2基因(seqidno:1)。3个事件全部均另外由rt-pcr示出以表达nb-lrr-2转录物的诊断性543bp片段。对初级转化体进行初步测试,以评估nb-lrr-2转基因对asr感染的效应。为此,用1×105个孢子/ml的大豆锈菌对t0植物材料进行喷雾接种。温育来自对照源植物和t0转基因植物的接种物质,并在接种后12天对病害症状进行评分。目视评价植物中病变的存在,并且用显微镜法评价以检测夏孢子堆(uredinia)的存在。在来自表示两个独立事件(5.1和7.1)的三种植物的叶上没有观察到孢子形成,所述事件被证实表达全长nb-lrr-2。有意思的是,一个转基因事件(6.1)展示对asr的完全易感性,并且包含棕褐色重度孢子形成病变。进一步的分析显示,在这种特定转化体中,插入的dna的完整性受损,导致合成截短的转录物,从而排除nb-lrr-2在该植物中的表达。因为nb-lrr-2能够在事件5.1和7.1中提供针对大豆锈菌的抗性,因此就针对大豆锈菌1的木豆抗性而言,将nb-lrr-2更名为ccrpp1,并且对事件5.1和事件7.1两者均在t1植物中推进用于进一步测试。对ccrpp1抗大豆锈菌的功效进行t1转基因测试。在生长室条件下种植并使来自所选t1事件的种子生长17天,直至vc。对植物进行qpcr取样以确定转基因拷贝数并用大豆锈菌孢子的悬浮液进行接种。利用从易感品种中收集的夏孢子进行接种,并在-80℃下储存。从储存物中取回后,使孢子悬浮于0.01%tween20的水溶液中,在40℃下热休克5分钟,并且充分混合;然后用血球计将孢子浓度调节至2×104sp/ml。用夏孢子悬浮液对植物进行喷雾接种,并在黑暗中在100%相对湿度下温育24小时,然后转移到生长室(22℃,70%rh,16hr光周期),使其在所述生长室中生长并发展出症状并持续15-29天。新生长物定期离体切除以便在实验期间保持单叶。为了评价ccrpp1的效果,将植物定性评级为抗性(r;没有病变)、部分抗性(pr;红棕色(rb,低度孢子形成病变)和易感的(s;棕褐色,高度孢子形成病变);并且通过离体切除并扫描叶,之后测定病变技术来定量评级。在接种后15天对大多数空样品进行评分,然而在接种后第29天对半合子和纯合植物进行评分。为了确定基因的效应,将转基因植物与来自相同事件的空植物进行比较。asr感染测定结果总结于表1中。这些结果示出纯合样品中的ccrpp1提供对asr的抗性。很少发现病变;当对所有纯合植物进行平均时,存在每叶面积单位(cm2)病变数的>99%减少。半合子植物显示出部分抗性,红棕色病变,并且示出每cm2病变数的55-70%减少。空植物包含棕褐色高孢子形成病变,通常具有对病原体的易感反应。这些asr感染测定结果示出当从宿主豆类(木豆至大豆植株)转基因地转移时,ccrpp1基因能够提供对大豆锈菌的抗性。表1:测得的携带ccrpp1的两个事件的性状。将接合性用作转基因拷贝数(空=0,半合子=1,纯合=2);r=抗性,pr=部分抗性,s=易感性;avglc/cm2=每单位面积(cm2)的平均病变计数。事件接合性n反应病变类型平均(lc/cm2)5.1纯合10r抗性0.01半合子26prrb,低度孢子形成3.79空16s棕褐色8.927.1纯合27r抗性0.01半合子48prrb,低度孢子形成2.83空30s棕褐色9.5实施例5:鉴定木豆品种g108-99中的asr抗性基因利用分离株ppufv01筛选来自群体cg8-1(g48-95×g108-99)的两百九十二种f2植物。在接种后,将266种植物分类为抗性并且24种分类为易感的。这种观察的分离比率表明两种独立显性基因座的存在。使用侧接ccrpp1基因座的标记101581和239615,选择对ccrpp1基因座处的易感等位基因纯合的53种抗性植物。将这些选择的f2植物自交以获得f3种子。在多个f2:3家族中观察到与ccrpp1基因座无关的抗性分离,从而确认在品种g108-99中存在新的抗性基因座。该品种用illuminahiseq2000(20x覆盖)进行测序,并且该数据用于鉴定g108-99和易感亲本品种g48-95(先前测序的)之间的84535种单核苷酸多态性(snp)。使用sequenomiplex平台的snp基因分型鉴定与g108-99中的新型抗性相关联的区域。在本公开的范围内的是,这些抗性植物用作用于鉴定赋予对大豆锈菌的抗性的r基因的源。将有症状的品种用于产生在相应抗性品种中作图并克隆赋予对大豆锈菌抗性的基因所需的分离种群。实施例6:鉴定作为豇豆属中抗性源的种质用单疱分离株ppufv02筛选得自不同源的总共89种豇豆属品种。最初,筛选得自巴西育种程序的55种豇豆品种(表2)。利用单疱分离株ppufv02攻击这些品种允许鉴定三种抗性品种和若干形成病害症状的品种。显著的例外为vu32,其也在成熟叶中发展成病害症状。在若干实验中,品种vu3、vu7和vu21一致示出对大豆锈菌的抗性。得自usda-grin的豇豆的16个附加品种的筛选揭示品种vun_00002、vun_00008、vun_00094、vun_00095和vun_00145的植株示出抗性,然而vun_00001和vun_00135示出缺乏抗性(表3)。接着,筛选得自auspgris的各种豇豆属物种的18个品种。后一组包括豇豆的七个品种,vignadalzelliana和长圆叶豇豆(vignaoblongifolia)的各两个品种、和以下的各一个品种:长虹豆(vignaparkeri)、vignafilicaulisvar.filicaulis、vignakirkii、长叶豇豆(vignaluteola)、绿豆(vignaradiata)、三裂叶豇豆(vignatrilobata)以及豇豆属种。品种arg88(长叶豇豆)、atf2361、atf2364(长圆叶豇豆)、atf2073(豇豆属)、ajp004长虹豆和cpi121683(豇豆)形成强病害症状。相比之下,品种atf2783(vignadalzelliana)、atf2363(豇豆)和ni456(豇豆亚种mensensis)对病害具有抗性。在本公开范围内的是,品种vu3、vu7、vu21、vun_00002、vun_00008、vun_00094、vun_00095、vun_00145、atf2783、atf2363和ni456用作用于鉴定赋予对大豆锈菌的抗性的r基因的源。有症状的品种vu32、vun_00001、vun_00135、arg88、atf2361、atf2364、atf2073、ajp004和cpi121683可用于形成在相应抗性品种中作图并克隆赋予对大豆锈菌的抗性的基因所需的分离种群。f1种群由豇豆属品种与对比表型杂交获得。来自vun_00135×vun_00094和vu32×vu21杂交的f2种群以3∶1比率分离,指示类似于木豆抗性,豇豆中的抗性由显性抗性基因座赋予。表2:来自巴西的豇豆品种中对大豆锈菌ppufv02具有抗性的源。根据病变尺寸和受影响的叶面积,使用从0(抗性)到4(缺乏抗性)范围内的分数,对病害症状进行评分。表3:来自usda-grin的豇豆品种中对大豆锈菌ppufv02具有抗性的源。根据病变尺寸和受影响的叶面积,使用从0-3(不同的抗性等级)到4(缺乏抗性)范围内的分数,对病害症状进行评分。对三叶型和子叶分别进行评分。实施例7:鉴定作为扁豆属中抗性源的种质利用单疱分离株ppufv02(表4)筛选得自auspgris的鹊豆(lablabpurpureus)的五十三种品种。以5×104孔/ml,用具有0.01%tween-80的水中的悬浮液接种具有两个三叶型叶的植物。将接种的植物在完全黑暗中在潮湿室中保持24h,然后转移到温室中。根据病变尺寸和受影响的叶面积,使用从0(抗性)到4(缺乏抗性)范围内的分数,对症状进行评分。对三叶型和子叶分别进行评分。鉴定出两个品种(ibs059和ibs837),其示出对该真菌分离株的抗性,然而其它品种全部均形成病害症状。本公开设想使用这两个品种作为对大豆锈菌具有抗性的源。有症状的品种代表使用基于作图的克隆来分解单独的nb-lrr基因的重要工具,并且克隆的基因可如大豆中的转基因那样提供有效的田间抗性。获得源自ibs059和ibs837与若干有症状的品种(包括,tamely、cpi51565、cpi52508、ibs879、cpi40167、tamelyearly、cpi18662、rjw5117、cpi36903、corbranca和若干其它品种)之间的杂交的f1种群,以便作图并克隆相应的抗性基因。为了作图并克隆相应的抗性基因,并且基于开花时间和种子产生,源自×ibs837和tamely×ibs059杂交的f2种群是下一组实验的关注点。分离分析指示这两个f2种群中的抗性表现出针对ppufv02的3:1分离,从而指示对大豆锈菌的抗性由显性基因座赋予。本公开设想作图并克隆在这些基因座处赋予大豆抗性的功能性基因。表4:来自auspgris的鹊豆品种中对大豆锈菌ppufv02具有抗性的源。根据病变尺寸和受影响的叶面积,使用从0-3(抗性)到4(缺乏抗性)范围内的分数,对病害症状进行评分。对三叶型和子叶分别进行评分。*austrcf参考号未发现。实施例8:鉴定作为菜豆属中抗性源的种质测试phaseolusvulgaris(菜豆)的使用以鉴定对大豆锈菌具有抗性的源。菜豆已经被描述为田间条件下大豆锈菌的宿主(dupreez等人,(2005)plantdis.89:206;和lynch等人,(2006)plantdis.,7:970)。此外,报道了16个菜豆栽培品种对大豆锈菌的不同反应,以及栽培品种-分离株相互作用的严重度和孢子形成(miles等人,(2007)plantdis.,91:698-704)。作为豆类作物,其独特之处在于具有两个平行的驯化事件,一个在中美洲,一个在安第斯山脉(bitocchi等人,(2013)mesoamericaandtheandes.newphytologist197:300-313)。结果,野生原始品种跨越较大的地理区域并由两个不同的基因库组成(kwak和gepts,(2009)theoreticalandappliedgenetics118.5:979-992)。使用与豇豆相同的评分表(参见,表3)筛选来自巴西的菜豆属的十三个品种(表5),并且鉴定其对ppufv02的不同反应。可产生抗性表型分离的来自对比基因型之间杂交的群体,并用于遗传研究和基因作图。本公开设想鉴定菜豆属中的nb-lrr型抗性基因在大豆中抗大豆锈菌的功效。表5:来自巴西的菜豆品种对大豆锈菌分离株ppufv02的反应实施例9:鉴定作为豌豆属中抗性源的种质使用巴西单疱分离株(ppufv-02)对来自usda/grin的豌豆(pisumsativum)群的品种进行大豆锈菌筛选(表6)。有意思的是,在接种大豆锈菌时,在接种后21天在72种测试的豌豆品种中观察到不同的反应。选择部分抗性的两个品种(pi271118和pi220189)用于进一步研究。为此,进行显微镜法和fitc-wga染色,之后进行荧光显微镜法以监测病原体随时间推移的生长。这些研究示出,虽然病原体能够成菌落到一定程度,但然后其生长被阻滞。此外,为鉴定抗性分离株,鉴定了若干品种,所述品种在视觉检查时示出缺乏抗性。更详细地评价两个品系(pi341888和pi198735),其显示出夏孢子堆形成和孢子形成。将来自抗性品种的植株与缺乏抗性和允许孢子形成的品种杂交,并且将所得的f1植物自花授粉。杂交pi341888×pi220189(和交互)的f1植物用于构建第一作图群体。针对抗性和缺乏抗性来筛选所得的f2子代(总共500株植株),并显示15∶1比率,从而指示所述抗性由两个显性基因座调控。有意思的是,在f2群体中观察到多个表型;抗性表型(0型),两种类型的部分抗性(红棕色(rb)病变;2型和3型)和明显缺乏抗性(4型)f2子代。分离模式按照9∶3∶3∶1比率(9种抗性;6种不同的rb型抗性(部分抗性)和1种缺乏抗性)。这些结果表明存在于该群体中的两个抗性基因座以互补方式起作用,并且两种抗性均被需要以传递抗性。表6:来自亚洲的豌豆品种中对大豆锈菌ppufv02的抗性的源。病害症状记录为抗性(0),部分抗性(1-2)和缺乏抗性且具有夏孢子堆(3-4;参见表文字说明)。每种品种筛选两种植株,病害症状评分的变化指示品种内的变化。*病害分数。其中0=抗性;不存在宏观和微观症状。1=部分抗性;使用fitc-小麦胚芽凝集素(wga)染色,之后通过荧光显微镜可见的,由菌丝生长引起的红棕色坏死的小(≤250μm)斑块。2=部分抗性;使用fitc-wga染色,之后通过荧光显微镜可见的,由菌丝生长引起的≤1000μm红棕色坏死的感染斑块。3=缺乏抗性;使用亮视野显微镜可见的明显的感染结构和/或夏孢子堆、菌丝,无孢子形成。4=缺乏抗性;具有或不具有坏死的病变,存在完全成形的夏孢子堆和孢子形成。实施例10:用附加的大豆锈菌分离株测试ccrpp1转基因为了评估ccrpp1抵抗附加的当前美国田间分离株的有效性,用20,000sp/ml的ga15(乔治亚州)和ar15(阿肯色州)分离株独立地接种事件7.1的纯合小植株和空小植株。这些层锈菌属分离株得自分别在佐治亚州迪卡特市和阿拉斯加州芒德拜尤市的大豆田收获的受感染的叶。如前所述进行接种和温育,不同的是新鲜的孢子从易感品种收集,从而不需要热休克;评分在接种后15天进行。该实验在空植物和易感对照植物上呈现高度严重性,其具有多个棕褐色病变,然而纯合植物示出对两种分离株的免疫性,如表7中所概述。然而,样品尺寸太小不能进行统计分析。表7:测得的用两种田间分离株接种ccrpp7的性状。将接合性用作转基因拷贝数(空=0,半合子=1,纯合=2);r=抗性,pr=部分抗性,s=易感性;avglc/cm2=每单位面积(cm2)的平均病变计数。这些数据展示以纯合状态携带ccrpp1的转基因植物是抗性的,并且半合子植物示出对大豆锈菌的至少三种田间分离株的部分抗性。实施例11:利用ccrpp1构建体php74119转化大豆如实施例3所公开的,通过dna的基因枪递送实现ccrpp1转基因事件的分离。然后通过靶向整合方法,实现转基因和标记dna插入到大豆基因组中。该位点特异性整合(ssi)程序依赖于flp/frt重组体系,是植物转化领域的技术人员所熟知的,并且描述于li等人(2009)plantphysiol.151:1087中。实施例3的转基因事件通过将dna轰击到93b86-5.1转化品系中来产生。附加的ssi事件通过将dna递送到93b86-tb5品系中来获得,所述93b86-tb5品系与前一品系在整合位点的染色体位置方面不同。就该实验而言,经由将2791bp的ccrpp1编码区与包含大豆泛素启动子+内含子i的1959bp片段和880bp拟南芥属泛素终止子片段的连接来组装新的ccrpp1转化构建体(php74119)。大豆品系93b86-tb5的基因枪转化。转基因大豆品系通过使用利用bioradbiolisticpds1000/he仪的粒子轰击(美国专利号4,945,050)将dna(php74119和flp重组酶构建体)递送到胚芽发生悬浮愈伤组织培养物(93b86-tb5)中来获得。通过重组酶介导的盒式交换(rmce)实现gmubi-ccrpp1基因到大豆基因组中的位点特异性整合,如li等人,2009年所述。在对100ppb氯磺隆选择后鉴定转基因事件,并且体细胞胚再生以产生推进用于t1种子生产的t0小植株。rmce事件的鉴定和表征使用qpcr来进行,如li等人,2009年所述。测试93b86-tb5s中的php74119转基因事件。由2个转化实验(大豆5342和大豆5861),回收总共3个转基因事件,所述转基因事件通过qpcrf发现包含ccrpp1转基因的高质量单拷贝插入。使用实施例4中所述的方法,对三个事件全部均推进抗asr的t1测试。如实施例4中所述,将来自三个事件的t1种子种植、取样和接种;在接种后15天对植物进行评分。在来自两个事件(大豆5342.11.1和大豆5342.11.2)的植物中观察到并确认ccrpp1的效应。半合子植物示出具有减少量的rb病变的部分抗性,然而纯合植物是抗性的。在这种情况下,构建体对降低半合子植物的严重性的作用为约98%,这是比先前在5.1背景中检测到的更加显著的效果。验证实验的结果汇总于表8中。然而,第三事件(大豆5861.1.1)不示出转基因植物和空植物之间的病害严重性的差异。为了鉴定有效事件和无效事件之间的差异,通过qrt-pcr评价所有三个事件的转录物量。结果示出在两个有效事件中的较高表达,然而事件大豆5861.1.1显示出ccrpp1转录物的较低表达。表8:在tb5背景下.gmubi-ccrpp1的两个转化事件的测得的性状。将接合性用作转基因拷贝数(空=0,半合子=1,纯合=2);r=抗性,pr=部分抗性,s=易感性;avglc/cm2=每单位面积(cm2)的平均病变计数。这些结果证实ccrpp1基因抵抗大豆锈菌ga05的功效。在这些实验中,独立地生成第二过度表达ccrpp1构建体(php74119)并且将其引入附加的大豆品系(93b86-tb5)中,并且分离并测试高质量转化事件。asr感染测定提供对存在于纯合子中的具有抗性的ccrpp1基因的进一步验证,并且在半合子中观察到强的但是部分的抗性。本文称为seq.idno:1-8的序列以文本文件形式与本申请同时提交,并且全文并入本文中。除非上下文另外明确规定,如本文所用的单数形式“一个”、“一种”和“该”包括多个指代物。因此,例如,提及“一个细胞”包括多个此类细胞,并且提及“该蛋白”包括提及一种或多种蛋白以及本领域技术人员已知的其等同物,等等。除非另外明确指明,否则本文所用的所有技术和科学术语均具有本发明所属领域的普通技术人员通常所理解的相同含义。如说明书和权利要求中所用的,术语“包含”可包括“由...组成”和“基本上由...组成”的方面。“包含”还可表示“包括但不限于”。如本文所用,词语“或”是指具体列表中的任一个成员,并且还包括该列表中成员的任何组合。如本文所用,术语“大豆”是指大豆和利用大豆育种或培养的任何植物品种。范围在本文中可表示为从“约”一个具体值,和/或至“约”另一个具体值。当表达此类范围时,其它方面包括从一个具体值和/或至其它具体值。类似地,当值被表示为近似值时,应当理解,通过使用先行词“约”,该具体值形成另一方面。还应当理解,所述范围中每一个的端点既与另一端点显著相关又独立于另一端点。还应当理解,存在本文所公开的多个值并且除了该值本身之外,每个值在本文中也以“约”该具体值的形式公开。例如,如果公开了值“10”,则也公开了“约10”。还应当理解,还公开了两个特定单元之间的每个单元。例如,如果公开了10和15,则也公开了11、12、13和14。说明书中提到的所有公布和专利申请指示了本发明适合的本领域技术人员的水平。所有公布和专利申请以引用方式并入本文,如同每个单独的公布或专利申请被具体地和独立地指出以引用方式并入本文一样。虽然为了清楚理解的目的已通过例证和实例的方式详细地描述的前述发明,但一些变化和修改可在所附权利要求的范围内实施。序列表<110>twobladesfoundatione.i.dupontdenemoursandcompanyuniversidadefederaldevicosa<120>用于转移对亚洲大豆锈病抗性的多核苷酸和方法<130>36446.0235u1<150>62/159,718<151>2015-05-11<160>8<170>patentin版本3.5<210>1<211>2778<212>dna<213>木豆(cajanuscajan)<400>1atggcagacagtgtggtttcctttgttttagatcacttgtcccaactggtggaacatgaa60gcaaggttgctaagtggcgtggaagacaaggtgaagtccctcgagagggagcttcagatg120atcaacgtcatcctaagaaccacaaacagcaacaacgacattcagaaaacagtggtgagc180caaatcagagatgtggcccatgaagctgaggatgtcattgacacgtacgttgccaaagtg240gccctgcacaacaggagaaccatgctggggaggctactccatggagttgaccaaacaaag300ttgctccatgacgtatccgagaaaatagacgagatcataacaactctcaaccagatacgt360gaaaacaagatcaaatacagcgagttccaagaaagaaatcatcaatccatagcagaagag420gaggaggaggagaaggagagggagaggttacttcacaagctaagaagaaatgtagaggag480gaacatgtagtgggttttatccgtggatctcaagcaatcatcaagctactcaaggaaggt540ggctcacggcgtaatgtggtctccatcatcggcatgggggggttgggcaagaccaccctt600gcccgaaaagtttataatgatagcaaggtgaaacaaggctttagttgttgtgtttgggtg660tacgtgtcaaacgagtgtagagctaaggagcttttgcttagtcttcttaagcatttgagg720ccaaacctcgaaactgaacttcaagaagaaaacaacaaaggaaaaaaattcactgaagaa780caagacatttttaacttgagtgtggaggagctgaagaaactggtgcggcaatacttggag840aggaaaacaaggtatctggtggtcctcgatgacttgtggaaaacacaagattgggacgag900gtgcaagatgcttttcccgacaacaacagaggcaacagaatattgatcactagtcgtttg960aaagaggtggccttgcatactagtcttcatcctccatactaccttcaatttctcagccaa1020gaagaaagctgggagctctttcgtaggaaagtgtttagaggggaagaatgcccttttgaa1080ctagagcctctaggcaaacaaatagtggcaagttgtcgcggtttgccactctctattgtt1140gtattagcaggattgctagccaacaaggaaaagtcacacagggaatggtccaaagtggtg1200ggtcacgtcaactggtatcttactcaagacgagactcaagtgaaggatatagttctgaag1260ctcagttatgataacttgccaagaagattgaaaccatgctttctatattttgggatattc1320cctgaagactttgaaatccctgttaggccattactacaacaatgggttgcagaagggttt1380atacaagaaacaagaaatagagacccagatgatgtggcagaagactacttgtacgagctc1440attgatcgtagtttggtccaagtagcagcaataaagactagtggaggtgtgaaaacttgt1500cacatccatgatcttctccgagatctttgtgtatcgcagagcaaaggggacaagattttt1560gaagtctgctcagataatgacattcaaattctaacaaaacctcgcaggttgtccttccat1620tgtgacatgggccactacatttcttcaagcaacaaagaccattcatgtatccgttctttg1680ttcttctttggaatatattccaattttactgggaacgagtgggaatggcttttcaaaggc1740ttcaaattggttcgagtgttagagcttggaaaaaaccattgcgcaggaaagatcccatct1800aatttgggggactttatccacttaaggtatttgagaattgactcgaattttggtataatt1860attccaacttccatacttacccttcagaatttacaaacagtagatttaggtaattggttt1920agggaaatcccaatttgtttccctgctcaaatgtggaagctcaaacatttaaggcacctg1980tatgggcaaggacctgtgaagcttcaaggccactattcaggatcaaatgaggttatgtgg2040aatctccgaaccatcttccccattgatattgatacacaaacattgtctctgatgaagaaa2100ggaagcttccccaatcttgtgaaattggggttgtcaatcaattcggaccgccaaggtaag2160tggccaaagttgttgcagagcttacaagaattaagtcatttgaatatcttaaagatttgc2220ctccgaggggattttgatgcttcaataggcacagtgtcaagcatatggcggtttggttgt2280gagccacaggagctattacaaagcctagggttgttgactcatataactacgttgaaaatc2340accaatatctgcagccttatgataacggttcctccaaatgtcaccaagttaacattgcgt2400ggtattagtagcatcactagggaggggctgaatgcgttgagaaatcacaccaaactccaa2460attttgagtctatatggagactatggctctaacattaacctcaattgtgttgtaggcggc2520tttccacaactgcaagtattgcaattgaaaaagttcacctctgtaaattggaaattaggc2580aatggtgcaatgccacgtcttcacactctagtcatcatcaactgtcaaagtttagatgat2640cttccaaatgaattgtggtctctcactgccttcagaaaactgcatgtaaaacaaccctca2700caaccaatgcttcgtatgctacgggatttgaaaataaaggatagggttcaagtcatagtc2760gatgatcatgacaactag2778<210>2<211>925<212>prt<213>木豆<400>2metalaaspservalvalserphevalleuasphisleuserglnleu151015valgluhisglualaargleuleuserglyvalgluasplysvallys202530serleugluarggluleuglnmetileasnvalileleuargthrthr354045asnserasnasnaspileglnlysthrvalvalserglnileargasp505560valalahisglualagluaspvalileaspthrtyrvalalalysval65707580alaleuhisasnargargthrmetleuglyargleuleuhisglyval859095aspglnthrlysleuleuhisaspvalserglulysileaspgluile100105110ilethrthrleuasnglnilearggluasnlysilelystyrserglu115120125pheglngluargasnhisglnserilealagluglugluglugluglu130135140lysgluarggluargleuleuhislysleuargargasnvalgluglu145150155160gluhisvalvalglypheileargglyserglnalaileilelysleu165170175leulysgluglyglyserargargasnvalvalserileileglymet180185190glyglyleuglylysthrthrleualaarglysvaltyrasnaspser195200205lysvallysglnglyphesercyscysvaltrpvaltyrvalserasn210215220glucysargalalysgluleuleuleuserleuleulyshisleuarg225230235240proasnleugluthrgluleuglnglugluasnasnlysglylyslys245250255phethrglugluglnaspilepheasnleuservalglugluleulys260265270lysleuvalargglntyrleugluarglysthrargtyrleuvalval275280285leuaspaspleutrplysthrglnasptrpaspgluvalglnaspala290295300pheproaspasnasnargglyasnargileleuilethrserargleu305310315320lysgluvalalaleuhisthrserleuhisproprotyrtyrleugln325330335pheleuserglngluglusertrpgluleupheargarglysvalphe340345350argglygluglucysprophegluleugluproleuglylysglnile355360365valalasercysargglyleuproleuserilevalvalleualagly370375380leuleualaasnlysglulysserhisargglutrpserlysvalval385390395400glyhisvalasntrptyrleuthrglnaspgluthrglnvallysasp405410415ilevalleulysleusertyraspasnleuproargargleulyspro420425430cyspheleutyrpheglyilepheprogluaspphegluileproval435440445argproleuleuglnglntrpvalalagluglypheileglngluthr450455460argasnargaspproaspaspvalalagluasptyrleutyrgluleu465470475480ileaspargserleuvalglnvalalaalailelysthrserglygly485490495vallysthrcyshisilehisaspleuleuargaspleucysvalser500505510glnserlysglyasplysilephegluvalcysseraspasnaspile515520525glnileleuthrlysproargargleuserphehiscysaspmetgly530535540histyrileserserserasnlysasphissercysileargserleu545550555560phephepheglyiletyrserasnphethrglyasnglutrpglutrp565570575leuphelysglyphelysleuvalargvalleugluleuglylysasn580585590hiscysalaglylysileproserasnleuglyasppheilehisleu595600605argtyrleuargileaspserasnpheglyileileileprothrser610615620ileleuthrleuglnasnleuglnthrvalaspleuglyasntrpphe625630635640arggluileproilecyspheproalaglnmettrplysleulyshis645650655leuarghisleutyrglyglnglyprovallysleuglnglyhistyr660665670serglyserasngluvalmettrpasnleuargthrilepheproile675680685aspileaspthrglnthrleuserleumetlyslysglyserphepro690695700asnleuvallysleuglyleuserileasnseraspargglnglylys705710715720trpprolysleuleuglnserleuglngluleuserhisleuasnile725730735leulysilecysleuargglyasppheaspalaserileglythrval740745750serseriletrpargpheglycysgluproglngluleuleuglnser755760765leuglyleuleuthrhisilethrthrleulysilethrasnilecys770775780serleumetilethrvalproproasnvalthrlysleuthrleuarg785790795800glyileserserilethrarggluglyleuasnalaleuargasnhis805810815thrlysleuglnileleuserleutyrglyasptyrglyserasnile820825830asnleuasncysvalvalglyglypheproglnleuglnvalleugln835840845leulyslysphethrservalasntrplysleuglyasnglyalamet850855860proargleuhisthrleuvalileileasncysglnserleuaspasp865870875880leuproasngluleutrpserleuthralaphearglysleuhisval885890895lysglnproserglnprometleuargmetleuargaspleulysile900905910lysaspargvalglnvalilevalaspasphisaspasn915920925<210>3<211>2862<212>dna<213>大豆(glycinemax)<400>3atggcggacagtgtggtttcctttgtattagatcacttgtctcaactgatggcacgtgaa60gcaaagttgctgtgtggcgtggaagacaggatcaagtccctccaaaatgagcttgaaatg120atcaatgagttcctcaatacctcaaagagcaaaaaggggattgagaaaaaagtggtgagc180caaatcagagatgttgcccatctagctgaggatgtcatcgacacatacgttgccaaagtt240gccatacacgagaggagaaccatgatgggtaggctcctccatagcgttcaccaagcacgg300ttgctccatgacgtagctgagaaaatagaccagataaaaaacactgtcagtgagatacgc360aacaacaagatcaaatatgaagaattccaagaaagcaacaatcaatccacaacaaaagca420gaggaggaggagaaggaaagggcgcaattactacacaagataagaagaaatgtggaggaa480gaagatgtagtaggctttgtacgtgactccaacgtagtcatcaagcgactcctagaaggt540ggttcatctcgtaacgttgtctccatcattggcatgggtggattgggcaagaccaccctt600gcccgaaaggtctataacagcagagaggtgaaacaatactttagatgttgcgcgtgggtt660tatgtgtcaaacgagtgtagagtcaaggagatttttcttggccttcttaagcatttgatg720ccaagccttgaataccaatgtagaggcaacaagaaaggtaagaaacacacaagagaatta780aacatttctaaattaaacgaggaggagttgaagaaactggtgagagaatgcttggagagg840aaaaggtatctggtggtcctcgatgacctgtggaaaacacaagattgggacgagttgcac900gatgcttttcctgacgacaacaaaggcagcaggatattgattactagtcgtttgaaagag960gtggcgttgcatactagccatcatcctccttactatcttcagtttcttagtgaagatgaa1020agttgggagctcttctgcaagaaagtgtttaggggcgaagagtactcttctgatttggag1080cctttggggaaacagatagttcaaagttgtcgtggtttgccactctcaatcgtggtgtta1140gcagggttgctagccaacaaggaaaaatcatatagagaatggtctaaagtggtgggtcat1200gtcaactggtatcttactcaagatgagacccaagtgaaggatatagttctcaaactcagc1260tatgacaacctgccaaggagattgaaaccatgctttctgtttcttgggatattccccgaa1320gactttgaaatcccagttaggccattattgcaacgatgggttgccgaaggatttatacaa1380gaaacagggaatagagacccagatgatgttgctgaagactacttgtacgagctcattgat1440cgtagtttggtccaagtagcagctatgaagactagtggaggtgtgaagacttgtcacatc1500catgatcttcttcgagatctttgcatatcagagagcaaagaggacaaggttttccaagtt1560tgcacaggtaataacattctaatctccacaaaaccccgcagactgtccattcattgtaac1620atgggtgattacatttcttcaaataacaatgaccagtcatgtattcgttctttgttcatg1680tttggaccccattatttttttatcccaagcgagttgaaaagacttttcaaaggcttcaaa1740ttggttcgagtgttagagcttggaacagacagttgtggaggaaagattccatctaatttg1800ggggactttatccacttaaggtatttgagaattgactcgcaacatgttagaattattcca1860gattctatacttacccttcagaatttacaaaccgtagacctaggttgttggcgtttgact1920attccaatttctttccctgctcaaatatggaagctcaaacatttaaggcatttgtatgcg1980ccagggcctatcaagcttagaggccacaattcaaaaccaagtgaggttatgtggaatctt2040caaaccatgaacgccattgtgttggacgaacaaacatcatatttgataaataaaggaact2100ttccccaaccttaagaatttaagtctgcaaatatcttcgggtcgcaaggctaaatggcct2160aagttgttgcagagcctacaacaattaagtcatttgagtaagttaaggatttcctttgaa2220atcaatttttgtgaaggttcactgtccggaaactatatgaaaagcatggagtggcacatt2280ggttgtaagccgcaagaagtattacaatgcatagggcagttgagtcatgtaactacgctg2340aaaattgtcaatgccttggaccttctaacatgtagggtcacgtttcctccaaatgttata2400aagttaacattgacaggtattagttgtgtgactgatgagggaatggattctttgggaaat2460cacacgaaactccaaaaattgagactaactagaggaattttgtcagaatcctttgacctc2520aattgtgttgcaggaaggttcccacaactgcaggtgtttgagatgagtggtttgaaagtt2580agaaactggaaattaggcaacagtgcaatgttatgcctccaaagtctgatcatccacaaa2640tgtaaaatgttagatggcatcccaaatgaactgtggtctttgattgctttgagaaaagtg2700caagtaaagcaaccctcagaagcaatggctcgcatgctacaaaacttggaaatgaaggat2760ggggttgaacttatagttgaaccgaaggaacgtcatgattctactgtaatattatctatg2820gatgaaatttgggaggcatttaattcacgtggtatttgttag2862<210>4<211>953<212>prt<213>大豆<400>4metalaaspservalvalserphevalleuasphisleuserglnleu151015metalaargglualalysleuleucysglyvalgluaspargilelys202530serleuglnasngluleuglumetileasnglupheleuasnthrser354045lysserlyslysglyileglulyslysvalvalserglnileargasp505560valalahisleualagluaspvalileaspthrtyrvalalalysval65707580alailehisgluargargthrmetmetglyargleuleuhisserval859095hisglnalaargleuleuhisaspvalalaglulysileaspglnile100105110lysasnthrvalsergluileargasnasnlysilelystyrgluglu115120125pheglngluserasnasnglnserthrthrlysalagluglugluglu130135140lysgluargalaglnleuleuhislysileargargasnvalgluglu145150155160gluaspvalvalglyphevalargaspserasnvalvalilelysarg165170175leuleugluglyglyserserargasnvalvalserileileglymet180185190glyglyleuglylysthrthrleualaarglysvaltyrasnserarg195200205gluvallysglntyrpheargcyscysalatrpvaltyrvalserasn210215220glucysargvallysgluilepheleuglyleuleulyshisleumet225230235240proserleuglutyrglncysargglyasnlyslysglylyslyshis245250255thrarggluleuasnileserlysleuasngluglugluleulyslys260265270leuvalargglucysleugluarglysargtyrleuvalvalleuasp275280285aspleutrplysthrglnasptrpaspgluleuhisaspalaphepro290295300aspaspasnlysglyserargileleuilethrserargleulysglu305310315320valalaleuhisthrserhishisproprotyrtyrleuglnpheleu325330335sergluaspglusertrpgluleuphecyslyslysvalphearggly340345350gluglutyrserseraspleugluproleuglylysglnilevalgln355360365sercysargglyleuproleuserilevalvalleualaglyleuleu370375380alaasnlysglulyssertyrargglutrpserlysvalvalglyhis385390395400valasntrptyrleuthrglnaspgluthrglnvallysaspileval405410415leulysleusertyraspasnleuproargargleulysprocysphe420425430leupheleuglyilepheprogluaspphegluileprovalargpro435440445leuleuglnargtrpvalalagluglypheileglngluthrglyasn450455460argaspproaspaspvalalagluasptyrleutyrgluleuileasp465470475480argserleuvalglnvalalaalametlysthrserglyglyvallys485490495thrcyshisilehisaspleuleuargaspleucysilesergluser500505510lysgluasplysvalpheglnvalcysthrglyasnasnileleuile515520525serthrlysproargargleuserilehiscysasnmetglyasptyr530535540ileserserasnasnasnaspglnsercysileargserleuphemet545550555560pheglyprohistyrphepheileprosergluleulysargleuphe565570575lysglyphelysleuvalargvalleugluleuglythraspsercys580585590glyglylysileproserasnleuglyasppheilehisleuargtyr595600605leuargileaspserglnhisvalargileileproaspserileleu610615620thrleuglnasnleuglnthrvalaspleuglycystrpargleuthr625630635640ileproileserpheproalaglniletrplysleulyshisleuarg645650655hisleutyralaproglyproilelysleuargglyhisasnserlys660665670prosergluvalmettrpasnleuglnthrmetasnalailevalleu675680685aspgluglnthrsertyrleuileasnlysglythrpheproasnleu690695700lysasnleuserleuglnileserserglyarglysalalystrppro705710715720lysleuleuglnserleuglnglnleuserhisleuserlysleuarg725730735ileserphegluileasnphecysgluglyserleuserglyasntyr740745750metlyssermetglutrphisileglycyslysproglngluvalleu755760765glncysileglyglnleuserhisvalthrthrleulysilevalasn770775780alaleuaspleuleuthrcysargvalthrpheproproasnvalile785790795800lysleuthrleuthrglyilesercysvalthraspgluglymetasp805810815serleuglyasnhisthrlysleuglnlysleuargleuthrarggly820825830ileleusergluserpheaspleuasncysvalalaglyargphepro835840845glnleuglnvalpheglumetserglyleulysvalargasntrplys850855860leuglyasnseralametleucysleuglnserleuileilehislys865870875880cyslysmetleuaspglyileproasngluleutrpserleuileala885890895leuarglysvalglnvallysglnproserglualametalaargmet900905910leuglnasnleuglumetlysaspglyvalgluleuilevalglupro915920925lysgluarghisaspserthrvalileleusermetaspgluiletrp930935940glualapheasnserargglyilecys945950<210>5<211>2793<212>dna<213>大豆<400>5atggcggacagtgtggtttcctttgtattagatcacttgtctcaactgatggcacgtgaa60gcaaagttgctgtgtggcgtggaagacaggatcaagtccctccaaaatgagcttgaaatg120atcaatgagttcctcaatacctcaaagagcaaaaaggggattgagaaaaaagtggtgagc180caaatcagagatgttgcccatctagctgaggatgtcatcgacacatacgttgccaaagtt240gccatacacgagaggagaaccatgatgggtaggctcctccatagcgttcaccaagcaagg300ttgctccatgacgtagctgagaaaatagaccagataaaaaacactatcagtgagatacgc360aacaacaagatcaaatatgaagaattccaagaaagcaacagtcaatccacaacgaaagca420gaggaggaggagaaggagaaggaaagggcgcaattactacacaagctaagaagaaatgtg480gaggaagaagatgtagtaggctttgtacgtgactccaacgtagtcatcaggcgactccta540gaaggtggttcatctcgtaacgttgtctccatcattggcatgggtggattgggcaagacc600acccttgcccgaaaggtctataacagcagagaggtgaaacaatactttagatgttgcgcg660tgggtttatgtgtcggacgagtgtagagtcaaggagattttccttggccttcttaagcat720ttgatgccaaaccttgaataccaacgtagaggcaacacgaaaggtaagaaacgcacaaga780gaatttaacatttctaagttgagcgaggaggagttgaaaaaactggtgagagaatgcttg840gagaggaaaaggtatctggtggtcctcgatgacctgtggaaaacacaagattgggacgag900ttgcacgatgcttttcctgacgacaacaaaggcagcaggatattgattactagtcgtttg960aaagaggtggcgttgcatactagccatcatcctccttactatcttcagtttcttagtgaa1020gatgaaagttgggagctcttctgcaagaaagtgtttaggggcgaagagtactcttctgat1080ttggagcctttggggaaacagatagttcaaagttgtcgtggtttgccactctcaatcgtg1140gtgttagcagggttgctagccaacaaggaaaaatcatatagagaatggtctaaagtggtg1200ggtcatgtcaactggtatcttactcaagatgagacccaagtgaaggatatagttctcaaa1260ctcagctatgacaacctgccaaggagattgaaaccgtgctttctgtttcttgggatattc1320cccgaagactttgaaatcccagttaggccattattgcaacgatgggttgccgaaggattt1380atacaagaaacagggaatagagacccagatgatgttgttgaagactacttgtacgagctc1440attgatcgtagtttggtccaagtagcagcaatgaagacaagtggaggcgtgaagacttgt1500cacatccatgatcttcttcgagatctttgcatatcagagagcaaagaagacaaggttttc1560caagtttgcacaggtaataacattctaatctccacaaaaccccgcagactgtccattcat1620tgtaacatgggtgactacatttcttcaaacaacaatgaccagtcatgtatccgttctttg1680ttcatgtttgggccacgttatttttttattccaagcgagttgaaaagacttttcaaaggc1740ttcaaattggttcgagtgttagagcttggaacagacagttgtggaggaaagattccatct1800aatttgggggactttatccacttaaggtatttgagaattgactcgcaacatgttagaatt1860attccagcttctatacttacccttcagaatttacaaaccgtagacctaggttgctggcgt1920ttgactattccaatttctttccctgctcaaatatggaagctcaaacatttaaggcatttg1980tatgctccagggcctatcaagcttagaggccactattcaaaaccaagtgaggttatgtgg2040aatcttcaaaccatgaacgccattgtgttggacgaacaagcatcatatttgatatataaa2100ggaactttccccaaccttaagaatttaggtctgaaaatatcttcgggtcgcaaggctaaa2160tggcctaagttgttgcagagcctactacaattaagtcatttgagtaagttaaggatttcc2220tttgaaatgaagttgtttgaaggttcagtgtccggaaactatgtgaacagcatggagtgg2280cacattggttgtaagccacaagaagtattacaaagcatagggcagttgagtcatgtaact2340acgctgaatattcgcaatgccttggaccttctaacatgtagggtcacgtttcctccaaat2400gttataaagttaagattgacaggtattagttgtgtgactgatgagggaatggattctttg2460ggaaatcacaccaaactccaaaaattgagactaacaggaggaattttgtcagaatccttt2520gacctcaattgtgttgcaggaaggttcccacaactgcaggtgtttgagatgagtggtttg2580aaagttagaaactggaaattaggcaacagtgcaatgttatgcctccaaagtctgatcatc2640caccaatgtaaaatgttagatggcatcccaaatgaactgtggtctttgattgctttgaga2700aaagtgcaagtaaagcaaccctcagaagcaatggctcacatgctacaaaacttggaaatg2760aaggatggggttgaacttatagttgaaccgtag2793<210>6<211>930<212>prt<213>大豆<400>6metalaaspservalvalserphevalleuasphisleuserglnleu151015metalaargglualalysleuleucysglyvalgluaspargilelys202530serleuglnasngluleuglumetileasnglupheleuasnthrser354045lysserlyslysglyileglulyslysvalvalserglnileargasp505560valalahisleualagluaspvalileaspthrtyrvalalalysval65707580alailehisgluargargthrmetmetglyargleuleuhisserval859095hisglnalaargleuleuhisaspvalalaglulysileaspglnile100105110lysasnthrilesergluileargasnasnlysilelystyrgluglu115120125pheglngluserasnserglnserthrthrlysalagluglugluglu130135140lysglulysgluargalaglnleuleuhislysleuargargasnval145150155160gluglugluaspvalvalglyphevalargaspserasnvalvalile165170175argargleuleugluglyglyserserargasnvalvalserileile180185190glymetglyglyleuglylysthrthrleualaarglysvaltyrasn195200205serarggluvallysglntyrpheargcyscysalatrpvaltyrval210215220seraspglucysargvallysgluilepheleuglyleuleulyshis225230235240leumetproasnleuglutyrglnargargglyasnthrlysglylys245250255lysargthrargglupheasnileserlysleusergluglugluleu260265270lyslysleuvalargglucysleugluarglysargtyrleuvalval275280285leuaspaspleutrplysthrglnasptrpaspgluleuhisaspala290295300pheproaspaspasnlysglyserargileleuilethrserargleu305310315320lysgluvalalaleuhisthrserhishisproprotyrtyrleugln325330335pheleusergluaspglusertrpgluleuphecyslyslysvalphe340345350argglygluglutyrserseraspleugluproleuglylysglnile355360365valglnsercysargglyleuproleuserilevalvalleualagly370375380leuleualaasnlysglulyssertyrargglutrpserlysvalval385390395400glyhisvalasntrptyrleuthrglnaspgluthrglnvallysasp405410415ilevalleulysleusertyraspasnleuproargargleulyspro420425430cyspheleupheleuglyilepheprogluaspphegluileproval435440445argproleuleuglnargtrpvalalagluglypheileglngluthr450455460glyasnargaspproaspaspvalvalgluasptyrleutyrgluleu465470475480ileaspargserleuvalglnvalalaalametlysthrserglygly485490495vallysthrcyshisilehisaspleuleuargaspleucysileser500505510gluserlysgluasplysvalpheglnvalcysthrglyasnasnile515520525leuileserthrlysproargargleuserilehiscysasnmetgly530535540asptyrileserserasnasnasnaspglnsercysileargserleu545550555560phemetpheglyproargtyrphepheileprosergluleulysarg565570575leuphelysglyphelysleuvalargvalleugluleuglythrasp580585590sercysglyglylysileproserasnleuglyasppheilehisleu595600605argtyrleuargileaspserglnhisvalargileileproalaser610615620ileleuthrleuglnasnleuglnthrvalaspleuglycystrparg625630635640leuthrileproileserpheproalaglniletrplysleulyshis645650655leuarghisleutyralaproglyproilelysleuargglyhistyr660665670serlysprosergluvalmettrpasnleuglnthrmetasnalaile675680685valleuaspgluglnalasertyrleuiletyrlysglythrphepro690695700asnleulysasnleuglyleulysileserserglyarglysalalys705710715720trpprolysleuleuglnserleuleuglnleuserhisleuserlys725730735leuargileserpheglumetlysleuphegluglyservalsergly740745750asntyrvalasnsermetglutrphisileglycyslysproglnglu755760765valleuglnserileglyglnleuserhisvalthrthrleuasnile770775780argasnalaleuaspleuleuthrcysargvalthrpheproproasn785790795800valilelysleuargleuthrglyilesercysvalthraspglugly805810815metaspserleuglyasnhisthrlysleuglnlysleuargleuthr820825830glyglyileleusergluserpheaspleuasncysvalalaglyarg835840845pheproglnleuglnvalpheglumetserglyleulysvalargasn850855860trplysleuglyasnseralametleucysleuglnserleuileile865870875880hisglncyslysmetleuaspglyileproasngluleutrpserleu885890895ilealaleuarglysvalglnvallysglnproserglualametala900905910hismetleuglnasnleuglumetlysaspglyvalgluleuileval915920925glupro930<210>7<211>2847<212>dna<213>大豆<400>7atggcggacagtgtggtttcctttgtattagatcacttgtctcaactggtggcacgtgaa60gcaaagttgctgtgtggcgtggaagacaggatcaagtccctccaaaatgagcttgaaatg120atcaatgagttcctcaatacctcaaagagcaaaaaggggattgagaaaaaagtggtgagc180caaatcagagatgttgcccatctagctgaggatgtcatcgacacatacgttgccaaagtt240gccatacacgagaggaggaccatgatgggtaggctcctccatagcgttcaccaagcaagg300ttgctccatgatgtagctgagaaaatagaccagataaaaaacacaatcagtgagatacgc360aacaacaagatcaaatatgaagaattccaagaaagcaacagtcaatccacaacaaaagca420gaggaggaggaagaggagaaggaaagggcgcaattactacacaagctaagaagaaatgtg480gaggaagaagatgtagtaggctttgtacgtgactccaacgtagtcatcaagcgactccta540gacggtggttcatctcgtaacgttgtctccatcattggcatgggtggattgggcaagacc600acccttgcccgaaaggtctataacagcagagaggtgaaacaatacttcagaagttgcgcg660tgggtttatgtgtcaaacgagtgtagagtcaaagagattttccttggccttcttaagcat720ttgatgccaaaccttgaataccaacgtagaggcaacaagaaaggtaagaaacgcacaaga780gaatttaacatttctaagttgagcgaggaggagttgaagaaactggtgagagaatgcttg840gagaggaaaaggtatctggtggtcctcgatgacctgtggaaaacacaagattgggacgag900ttgcacgatgcctttcctgacgacaacaaaggcagcagaatattgattactagtcgtttg960aaagaggtggcgttgcatactagccatcatcctccttactatcttcagtttcttagtgaa1020gatgaaagttgggagctcttctgcaagaaagtgtttaggggcgaagagtactcttctgat1080ttggagcctttggggaaacagatagttcaaagttgtcgtggtttgccactctcaatcgtg1140gtgttagcagggttgctagccaacaaggaaaaatcatatagagaatggtctaaagtggtg1200ggtcatgtcaactggtatcttactcaagatgagacccaagtgaaggatatagttctcaaa1260ctcagctatgacaacctgccaaggagattgaaaccgtgctttctgtttcttgggatattc1320cccgaagactttgaaatcccagttaggccattattgcaacgatgggttgccgaaggattt1380atacaagaaacagggaatagagacccagatgatgttgctgaagactacttgtacgagctc1440attgatcgtagtttggtccaagtagcagcaatgaagacaagtggaggcgtgaagacttgt1500cacatccatgatcttcttcgagatctttgcatatcagagagcaaagaggacaaggttttc1560gaagtttgcacaggtaataacattctaatgtccacaaaaccccgcagattgtccattcat1620tgtaacatgggtgactacatttcttcaaataacaatgaccagtcatgtattcgttctttg1680ttcatgtttgggccacattattttttcatcccaagcgagttgaaaagacttttcaaaggc1740ttcaaattggttcgagtgttagagcttggaacagacagctgcggaggaaagattccatct1800aatttgggggactttatccacttaaggtatttgagaattgtctcgaaatatgttagaatt1860attcctgcttctatacttacccttcagaatttacaaaccgtagacctaggttgttggcgt1920tgggctactccaatttctttccctgtttcaatttctttcccggctcaaatatggaagctc1980aaacatttaaggcatttgtatgcgccagggcctatcaagcttagaggccactattcaaaa2040ccaagtgaggttatgtggaatcttcaaaccatgaatgccattgtgttggacgaacaaaca2100tcatatttgataaataaaggaactttccccaaccttaaggatttaggtctgcaaatatct2160tcgggtcgcaaggctaaatggcctaagttgttgcagagcctacaacaattaaatcatttg2220agtaagttaaggattttctttgaaatgaagtttcctgaaggttcagtgtccgaaaactat2280gtgaacagcatggagtggcacattggttgtaagccacaagaagtattacaatgcataggg2340cagttgagtcatgtaactacgctgaaaattgtcaatgccttggaccttctaacatgtagg2400gtcacgtttcctccaaatgttataaagttaacatttacaggtattagttatgtgactgat2460gagggaatggattctttgggaaatcacaccaaactccaaaaattgagactaaccggagga2520atttggtcagattcctttgacctcaattgtgttgcaggaaggttcccaaaactgcaggtg2580tttgagatgagtcgtttgaacgttagaaactggaaattaggcaatagtgcaatgttatgc2640ctccaaagtctgatcatccacaaatgtaaagtgttagatggcatcccaaatgaactgtgg2700tctttgattgccttgagaaaagtgcaagtaaagcaaccctcagaagcaatggctcacatg2760ctacaaaacttggaaatgaaggatggggttgaacttatagttgaaccggaggaacgtcat2820gattctactgtgattatctatggatga2847<210>8<211>948<212>prt<213>大豆<400>8metalaaspservalvalserphevalleuasphisleuserglnleu151015valalaargglualalysleuleucysglyvalgluaspargilelys202530serleuglnasngluleuglumetileasnglupheleuasnthrser354045lysserlyslysglyileglulyslysvalvalserglnileargasp505560valalahisleualagluaspvalileaspthrtyrvalalalysval65707580alailehisgluargargthrmetmetglyargleuleuhisserval859095hisglnalaargleuleuhisaspvalalaglulysileaspglnile100105110lysasnthrilesergluileargasnasnlysilelystyrgluglu115120125pheglngluserasnserglnserthrthrlysalagluglugluglu130135140gluglulysgluargalaglnleuleuhislysleuargargasnval145150155160gluglugluaspvalvalglyphevalargaspserasnvalvalile165170175lysargleuleuaspglyglyserserargasnvalvalserileile180185190glymetglyglyleuglylysthrthrleualaarglysvaltyrasn195200205serarggluvallysglntyrpheargsercysalatrpvaltyrval210215220serasnglucysargvallysgluilepheleuglyleuleulyshis225230235240leumetproasnleuglutyrglnargargglyasnlyslysglylys245250255lysargthrargglupheasnileserlysleusergluglugluleu260265270lyslysleuvalargglucysleugluarglysargtyrleuvalval275280285leuaspaspleutrplysthrglnasptrpaspgluleuhisaspala290295300pheproaspaspasnlysglyserargileleuilethrserargleu305310315320lysgluvalalaleuhisthrserhishisproprotyrtyrleugln325330335pheleusergluaspglusertrpgluleuphecyslyslysvalphe340345350argglygluglutyrserseraspleugluproleuglylysglnile355360365valglnsercysargglyleuproleuserilevalvalleualagly370375380leuleualaasnlysglulyssertyrargglutrpserlysvalval385390395400glyhisvalasntrptyrleuthrglnaspgluthrglnvallysasp405410415ilevalleulysleusertyraspasnleuproargargleulyspro420425430cyspheleupheleuglyilepheprogluaspphegluileproval435440445argproleuleuglnargtrpvalalagluglypheileglngluthr450455460glyasnargaspproaspaspvalalagluasptyrleutyrgluleu465470475480ileaspargserleuvalglnvalalaalametlysthrserglygly485490495vallysthrcyshisilehisaspleuleuargaspleucysileser500505510gluserlysgluasplysvalphegluvalcysthrglyasnasnile515520525leumetserthrlysproargargleuserilehiscysasnmetgly530535540asptyrileserserasnasnasnaspglnsercysileargserleu545550555560phemetpheglyprohistyrphepheileprosergluleulysarg565570575leuphelysglyphelysleuvalargvalleugluleuglythrasp580585590sercysglyglylysileproserasnleuglyasppheilehisleu595600605argtyrleuargilevalserlystyrvalargileileproalaser610615620ileleuthrleuglnasnleuglnthrvalaspleuglycystrparg625630635640trpalathrproileserpheprovalserileserpheproalagln645650655iletrplysleulyshisleuarghisleutyralaproglyproile660665670lysleuargglyhistyrserlysprosergluvalmettrpasnleu675680685glnthrmetasnalailevalleuaspgluglnthrsertyrleuile690695700asnlysglythrpheproasnleulysaspleuglyleuglnileser705710715720serglyarglysalalystrpprolysleuleuglnserleuglngln725730735leuasnhisleuserlysleuargilephepheglumetlysphepro740745750gluglyservalsergluasntyrvalasnsermetglutrphisile755760765glycyslysproglngluvalleuglncysileglyglnleuserhis770775780valthrthrleulysilevalasnalaleuaspleuleuthrcysarg785790795800valthrpheproproasnvalilelysleuthrphethrglyileser805810815tyrvalthraspgluglymetaspserleuglyasnhisthrlysleu820825830glnlysleuargleuthrglyglyiletrpseraspserpheaspleu835840845asncysvalalaglyargpheprolysleuglnvalpheglumetser850855860argleuasnvalargasntrplysleuglyasnseralametleucys865870875880leuglnserleuileilehislyscyslysvalleuaspglyilepro885890895asngluleutrpserleuilealaleuarglysvalglnvallysgln900905910proserglualametalahismetleuglnasnleuglumetlysasp915920925glyvalgluleuilevalgluproglugluarghisaspserthrval930935940ileiletyrgly945当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1