具有抗癌活性的肽以及均含有该肽作为活性成分的用于预防和治疗癌症的药物组合物和膳食补充组合物的制作方法
本发明涉及具有抗癌活性的肽,以及具有所述肽作为活性成分的癌症预防或癌症治疗的药物组合物和健康功能性食品组合物。
背景技术:
迄今为止已开发和使用了许多抗癌剂,包括天然产物、蛋白质能或肽能剂和合成小分子。然而,大多数这些抗癌剂对存活的正常细胞引起严重的副作用,并且可能对某些类型的癌不起作用。在一般情况下,抗癌剂的作用在患有同一类型的癌的患者中变化。在这样的情况下,已经在全世界进行了许多关于开发可以为上述问题提供解决方案,能够选择性地去除癌细胞而不影响存活的正常细胞,并且甚至可以消除任何类型的癌细胞之抗癌剂的新概念的研究。
转录因子cp2c(也称为cp2、tfcp2、lsf、lbp1或ubp1)在哺乳动物中广泛表达。当细胞从休止期(g0)进入dna复制期(s)时,cp2c的活性被精细地调节,并且其对于允许细胞有效地通过g1/s转变期至关重要。cp2c活性的调节主要通过翻译后修饰进行,并且其水平通常保持在低水平。然而,由于cp2c在肿瘤细胞中过表达,故其充当重要的癌基因在癌发生中起关键作用。
在这一点上,波士顿大学的一个研究小组报道了因子喹啉酮抑制剂1(factorquinolinoneinhibitor1,fqi1)作为抑制肝癌细胞系中cp2c的细胞活性的物质。fqi1及其衍生物通过化学文库筛选鉴定,并且在细胞和移植的小鼠模型中选择性地抑制癌细胞而不影响正常细胞方面是成功的(grant等,antiproliferativesmall-moleculeinhibitorsoftranscriptionfactorlsfrevealoncogeneaddictiontolsfinhepatocellularcarcinoma,proc.natl.acad.sci.2012;109(12):4503-4508)。
此外,本发明人已经报道了通过筛选肽展示文库示出的识别cp2c不同区域的四种新的肽基序(hxpr、phl、asr和pxhxh)(kang等,identificationandcharacterizationoffournovelpeptidemotifsthatrecognizedistinctregionsofthetranscriptionfactorcp2,febsjournal2005;272:1265-1277)。本发明人已经提出了cp2c识别靶蛋白的特异性结合基序并且与该蛋白质相互作用以调节多种细胞活性。基于可用于dna-蛋白质相互作用的高度特异且灵敏分析的dna免疫沉淀法,本发明人在其后续研究中已经通过体外测定筛选了抑制cp2c与dna结合的肽,结果,发现由12个氨基酸构成的肽5以浓度依赖性方式抑制cp2c-dna结合(kim等,adnaimmunoprecipitationassayusedinquantitativedetectionofinvitrodna-proteincomplexbinding,analyticalbiochemistry.2013;441:147-151)。
技术实现要素:
技术问题
因此,本发明是基于本发明人进行的研究结果进行的,并且旨在提供用于更有效地抑制cp2c(多种癌中已知的关键转录因子)活性的肽,含有所述肽作为活性成分的用于预防和治疗癌症的药物组合物,以及含有所述肽作为活性成分的用于预防和治疗癌症的健康功能性食品组合物。
技术方案
本发明提供了针对癌症具有预防和治疗活性的肽,其与转录因子cp2c结合并具有seqidno:1所示的氨基酸序列:
asn-tyr-pro-gln-arg-pro(1)。
根据本发明的一个实施方案,乙酰基和酰胺基可分别与具有seqidno:1所示氨基酸序列的肽的n端asn残基和c端pro残基连接(或键合)。
根据本发明的另一个实施方案,具有seqidno:1所示氨基酸序列的肽的c端pro残基可与具有seqidno:2所示氨基酸序列的肽连接:
cys-arg-gly-asp-lys-gly-pro-asp-cys(2)。
根据本发明的另一个实施方案,当具有seqidno:2所示氨基酸序列的肽与具有seqidno:1所示氨基酸序列的肽的c端pro残基连接时,乙酰基和酰胺基可分别与具有seqidno:1所示氨基酸序列的肽的n端asn残基和具有seqidno:2所示氨基酸序列的肽的c端cys残基键合。
根据本发明的另一个实施方案,具有seqidno:1所示氨基酸序列的肽的c端pro残基和n端asn残基可分别与具有seqidno:2所示氨基酸序列的肽和具有seqidno:3所示氨基酸序列的肽连接:
lys-cys-lys-gly-gly-ser-gly-gly-ser(3)
其中第一氨基酸lys和第三氨基酸lys表示6-氨基己酸,并且具有seqidno:3所示氨基酸序列的肽的n端lys残基可与异硫氰酸荧光素(fluoresceinisothiocyanate,fitc)连接。
根据本发明的另一个实施方案,当具有seqidno:2所示氨基酸序列的肽与具有seqidno:1所示氨基酸序列的肽的c端pro残基连接并且具有seqidno:3所示氨基酸序列的肽与具有seqidno:1所示氨基酸序列的肽的n端asn残基连接时,酰胺基可与具有seqidno:2所示氨基酸序列的肽的c端cys残基连接。
根据本发明的另一个实施方案,具有seqidno:1所示氨基酸序列的肽的n端asn残基可与具有seqidno:4所示氨基酸序列的肽键合:
lys-ile-lys-lys-val-lys-lys-lys-gly-arg-lys-gly-ser-lys-ile-lys-lys-val-lys-lys-lys-gly-arg-lys-gly-gly(4)。
根据本发明的另一个实施方案,当具有seqidno:4所示氨基酸序列的肽与具有seqidno:1所示氨基酸序列的肽的n端asn残基连接时,乙酰基和酰胺基可分别与具有seqidno:4所示氨基酸序列的肽的n端lys残基和具有seqidno:1所示氨基酸序列的肽的c端pro残基连接。
根据本发明的另一个实施方案,具有seqidno:2所示氨基酸序列的肽可与具有seqidno:1所示氨基酸序列的肽的c端pro残基连接,并且生物素标记的lys可与具有seqidno:2所示氨基酸序列的肽的c端cys残基的ε-nh2连接。
根据本发明的另一个实施方案,当具有seqidno:2所示氨基酸序列的肽与具有seqidno:1所示氨基酸序列的肽的c端pro残基连接,并且生物素标记的lys与具有seqidno:2所示氨基酸序列的肽的c端cys残基的ε-nh2连接时,乙酰基和酰胺基可分别与具有seqidno:1所示氨基酸序列的肽的n端asn残基和生物素标记的c端lys连接,所述生物素标记的c端lys与具有seqidno:2所示氨基酸序列的肽的c端cys残基的ε-nh2连接。
本发明还提供了用于预防和治疗癌症的含有至少一种调节cp2c活性的肽作为活性成分的药物组合物。
本发明还提供了用于预防和治疗癌症的含有至少一种调节cp2c活性的肽作为活性成分的健康功能性食品组合物。
本发明的效果
肽或药物组合物以非常高的效力穿透癌细胞的膜,能够与cp2c特异性结合,并且可以抑制cp2c与dna结合的能力。因此,本发明的肽或药物组合物具有抑制cp2c活性的能力。本发明的肽或药物组合物由于其能力而阻碍cp2c介导的癌细胞特异性转录活性,因此在特异性地治疗癌细胞中有效。此外,本发明的肽或药物组合物可以用于预防癌症。此外,本发明的肽可以用作用于癌症预防的健康食品添加剂。
附图说明
图1是由总共6个区域构成的cp2c全长氨基酸序列以及其中缺失cp2c序列n端和c端的多种cp2c突变体的氨基酸序列的示意图。
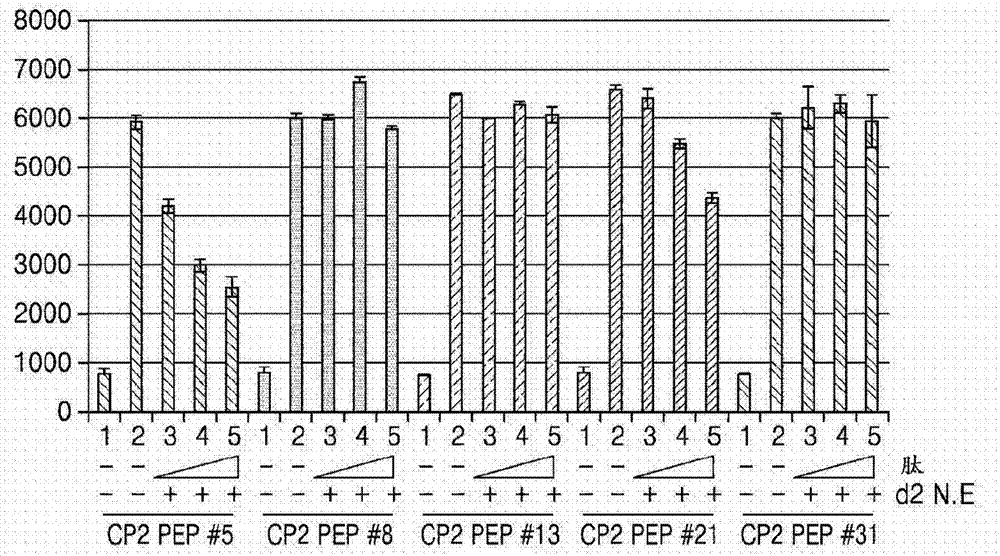
图2示出了通过dna免疫沉淀(dip)测定测量的dna-cp2c复合物对五种cp2c结合肽(肽5、肽8、肽13、肽21和肽31)的结合亲和力。
图3示出了通过dip测定的dna-cp2c复合物对肽5、肽8、肽5-1和肽5-2的结合亲和力。
图4a至4e是在将多种癌细胞系和正常人干细胞和分化细胞与fqi1、肽5c和肽5-2c培养之后获取的显微图像:具体地,图4a的图像示出了肝癌细胞系(hepg2、hep3b),人胚胎肾细胞系(293t),乳腺癌细胞系(mvcf7、mda-mb-231),血癌细胞系(k562、mel、hel、hl60)和成胶质细胞瘤细胞系(u251、u373mg、u87mg);图4b的图像示出了来自多种组织来源的表皮细胞系(mcf10a、beas2b),原代培养的小鼠t淋巴细胞(静息和活化的t淋巴细胞)和人间充质干细胞(hmsc);图4c的图像示出了在乳腺球培养期间的mcf7、293t和mba-mb-231细胞系;图4d的图像示出了在与成纤维细胞共培养期间的人胚胎干细胞(hesc);图4e的图像示出了人造血祖细胞(hpc)和在分化成红细胞期间来源于其的细胞(在分化诱导后第3、7和14天的细胞)。
图5a至5j示出了hepg2细胞系和hep3b细胞系(5a),mcf7细胞系和mda-mb-231细胞系(5b),u251细胞系,u87mg细胞系和u343细胞系(5c),k562细胞系、hel细胞系和hl60细胞系(5d),293t细胞系(5e),mcf10a细胞系和beas2b细胞系(5f),原代培养的小鼠t淋巴细胞(静息和活化t淋巴细胞)(5g),hmsc细胞系(5h),hesc细胞系(5i),造血祖细胞(hpc)和在诱导分化成红细胞期间来源于其的细胞(在第3、7和14天)(5j)在用浓度提高的fqi1、肽5c和肽5-2c处理一次后的相对生存力,而图5k示出了肽处理后96小时的孔板的图像。
图6a和6b是比较如通过western印迹确定的来自多种组织来源的6种结肠癌细胞系,6种肺癌细胞系,3种乳腺癌细胞系,3种成胶质细胞瘤细胞系和一种宫颈癌细胞系(6a)和血癌细胞系(mel、k562、hel、hl60),肝癌细胞系(hepg2、hep3b),人表皮样癌细胞系(a431),结肠癌细胞系(hct116),表皮细胞系(293t、beas2b、mcf10a),以及hmsc细胞系(6b)中的cp2c蛋白质表达水平的图像。
图7a和7b示出了用于在用肽5-2c处理细胞系之后计算图5所示细胞系的肽5-2c的ic50值的图以及计算的ic50值:具体地,图7a示出了用于在用肽5-2c处理每种细胞系一次后48小时计算ic50值的图以及计算的ic50值;图7b示出了在用肽5-2c处理成胶质细胞瘤细胞系u343、u373mg、u251和u87mg一次后48小时和96小时计算ic50值的图以及计算的ic50值。
图8示出了用于在用fqi1、肽5c和肽5-2c处理mcf10a细胞系和beas2b细胞系后48小时计算ic50值的图以及计算的ic50值。
图9a至9f示出了mda-mb-231细胞系和mcf7细胞系(9a),u343细胞系和u87mg细胞系(9b),hct116细胞系和ht29细胞系(9c),293t细胞系和hepg2细胞系(9d),u937细胞系、jurkat细胞系、hl60细胞系和hel细胞系(9e)以及mcf10a细胞系和beas2b细胞系(9f)在用浓度增加的肽5-2c和肽5-2d处理一次后的相对生存力,而图9g示出肽处理后96小时的孔板的图像。
图10示出了用于在用肽5-2c和肽5-2d处理细胞系后计算图9所示细胞系的肽5-2c和肽5-2d的ic50值的图以及计算的ic50值。
图11a至11g示出了移植有u343成胶质细胞瘤细胞系的小鼠模型中肽5-2c抑制肿瘤生长的能力和肽5-2c的生理特性。将u343成胶质细胞瘤细胞系(2.5×106个细胞/50μl)皮下注射到雄性balb/c裸鼠(n=6只/组)的背部,从肿瘤大小增加到约80mm3的时间起,每3天将肽5-2c(3mg/kg)直接注射到肿瘤部位,测量小鼠的体重和肿瘤的体积,在肽注射后第70天处死小鼠,并切除肿瘤、血液和主要器官。具体地,图11a示出了实验期间肿瘤的体积,图11b示出了小鼠的体重变化,图11c示出了在肽施用后第70天的小鼠的图像,图11d示出了切除的肿瘤的图像,图11e示出了从小鼠切除的肿瘤的重量及其平均值,图11f示出了对照组和肽处理组中小鼠的多种标准血液指标,图11g示出了主要器官(脾、肝、肺和肾)的h/e染色图像。
图12a至12d示出了移植有a431人表皮样癌细胞系的小鼠模型中肽5-2c抑制肿瘤生长的能力和肽5-2c的生理特性。将a431细胞系(5×106个细胞/100μl)皮下注射到雄性balb/c裸鼠(n=4只/组)的背部,从肿瘤大小增加到约50mm3的时间起,每隔一天将肽5-2c(1×ic50;1.7mg/kg,2×ic50;3.5mg/kg)通过尾静脉注射到小鼠中,总共5次,测量肿瘤体积,在肽注射后第13天处死小鼠,并切除肿瘤和主要器官。图12a示出了实验期间肿瘤的体积,图12b示出了小鼠和切除的肿瘤的图像,图12c示出了从小鼠切除的肿瘤的重量及其平均值,图12d示出了组的主要器官(脾、肝、肺、血管和肌肉)的h/e染色图像。
图13a至13d示出了移植有能够转移到肺组织的mda-mb-231细胞系(lm1细胞)的动物模型中肽5-2c抑制转移到肺组织和肿瘤生长的能力以及肽5-2c的生理特性。将mda-mb-231细胞系(lm1细胞;1×106个细胞/40μl)注射到12周龄雌性balb/c裸鼠(n=5只/组)的乳腺脂肪垫中,从肿瘤大小增加到约50mm3的时间起,每3天将肽5-2c(1×ic50;1.7mg/kg=52μg/小鼠,2×ic50;3.5mg/kg=104μg/小鼠)通过尾静脉注射到小鼠中,总共5次,测量肿瘤体积,在肽注射后第30天处死小鼠,并切除肿瘤和主要器官。图13a示出了具有肿瘤的小鼠和切除的肿瘤的图像,图13b示出了从小鼠切除的肿瘤的重量及其平均值,图13c示出了对照组和肽处理组中转移到肺的肿瘤灶数量的平均值和标准偏差,图13d示出了对照组和肽处理组中小鼠的血液指标。
图14a至14e示出了正常cp2c结合位点和cp2c突变体序列的荧光素酶报道基因的表达谱,以通过肽5c和肽5-2c分析cp2c的转录活性。
图15a至15c示出了在用0、1、2、3和10μm的多个浓度的肽5-2c(为了分析肽5-2c对细胞周期的影响)处理细胞系后48小时通过facs的多种细胞系(mel、k562、mcf-10a、mda-mb-231)的细胞周期分析结果:具体地,图15a示出了取决于facs期间的dna的量的细胞分布;图15b示出了不同细胞周期阶段中细胞的定量分布;图15c示出了在显示出凋亡的subg1期中细胞的扩大分布。
图16a至16d示出了当用不同浓度的肽5-2c处理mcf7细胞系不同的时间段时细胞周期相关标志物基因的表达谱:具体地,图16a示出了在用2μm肽5-2c处理72小时时如通过western印迹确定的多种细胞周期相关标志物基因蛋白质的时间依赖性表达谱,图16b示出了在用多个浓度(0、1、2和3μm)的肽5-2c处理48小时后如通过western印迹确定的细胞周期相关标志物基因蛋白质的表达谱。图16c和16d分别示出了图16a和16b中的标志物基因蛋白质表达的定量变化。
图17示出了在用2μm肽5-2c处理mda-mb-231细胞系和mcf7细胞系后凋亡相关标志物基因蛋白质的时间依赖性表达谱。
图18示出了显示出在用2μm肽5-2c处理mda-mb-231细胞系和mcf10a细胞系后24小时和48小时的细胞形态的电子显微图像,用生理盐水处理的细胞和用2μmfqi1处理的细胞分别用作阴性和阳性对照。
图19a至19d是确认随着时间的推移mda-mb-231细胞系中fitc-缀合的肽5-2c的细胞内迁移途径的图:具体地,图19a示出了如通过mtt测定确定的用浓度为0、0.5、1、2、3和10μm的生理盐水以及fitc-缀合的irgd、fitc-5-1c和fitc-5-2c肽处理mda-mb-231细胞系96小时后的细胞生长速率的变化;图19b示出了用于在处理后48小时计算处理组中肽的ic50值的图以及计算的ic50值;图19c示出了显示出在96小时时mtt测定的实例的孔板的图像;图19d示出了显示出在用肽处理后24小时fitc-缀合的肽(各2μm)的时间依赖性细胞内分布的图像。
图20a至20d比较了肽5-2cb(生物素标记的肽5-2c)对癌细胞存活的影响与肽5-2c对癌细胞存活的影响:具体地,图20a示出了在用浓度增加的肽5-2c和两个不同批次的肽5-2cb处理一次之后mda-mb-231细胞系的相对生存力;图20b示出了肽处理后96小时的孔板的图像;图20c示出了用于在用肽处理mda-mb-231细胞系后48小时计算ic50值的图以及计算的ic50值;图20d示出了用于在用肽处理mcf7细胞系后48小时计算ic50值的图以及计算的ic50值。
图21a至21d示出了用于鉴定与肽5-2c结合的cp2c蛋白质区域(对应的第306至396位氨基酸)中用于dna结合和肽5-2c结合的重要氨基酸的一系列实验的结果:具体地,图21a示出了与肽5-2c结合的区域的列表和通过用丙氨酸替换位点的氨基酸序列中的极性氨基酸而产生的19种gst-融合突变体;图21b示出了使用gst抗体珠之由大肠杆菌(e.coli)中过表达的突变体纯化的gst-融合蛋白与存在于放射性标记的α-球蛋白启动子中的cp2c结合位点的dna探针结合的dna-ip结果;图21c示出了纯化的gst-融合突变蛋白与肽5-2c结合的直接elisa测定结果;图21d示出了基于cp2c的第306-396位氨基酸区域的rosetta建模算法预测的用于dna与三级结构结合和肽5-2c与三级结构结合的重要氨基酸。
最佳实施方式
现在将更详细地描述本发明。
本发明提供了针对癌症具有治疗活性的肽,其与转录因子cp2c结合并具有seqidno:1所示的氨基酸序列:
asn-tyr-pro-gln-arg-pro(1)。
具有seqidno:1所示氨基酸序列的肽(在下文中也称为“肽5-2”)由6个氨基酸构成,并且与cp2c蛋白质相互作用以调节该蛋白质的活性,如从以下实施例部分可以具体看出。cp2c是一种在肿瘤细胞中特异性过表达的蛋白质。这种活性调节最终导致抗癌效果。
如上所述,本发明人已发现由12个氨基酸构成的肽5以浓度依赖性方式抑制cp2c-dna结合(kim等,adnaimmunoprecipitationassayusedinquantitativedetectionofinvitrodna-proteincomplexbinding,analyticalbiochemistry.2013;441:147-151)。然而,从以下实施例部分的结果可以看出,由12个氨基酸构成的肽5即使在正常对照细胞系中也抑制细胞生长和诱导凋亡,而由6个氨基酸(肽5的6个下游氨基酸)构成的本发明的肽5-2基本上不诱导正常对照细胞系的生长抑制和凋亡。另一方面,由肽5的6个上游氨基酸构成的肽5-1不能抑制cp2c-dna结合,并且用肽5-1处理不影响癌细胞系以及正常细胞对照组的生长抑制和凋亡诱导。因此,本发明的肽5-2由于其对癌细胞的特异性活性而不影响正常细胞提供了理想的癌症治疗的可能性。
本发明的肽5-2可通过修饰被赋予多种功能。例如,本发明的肽5-2可被修饰成以下肽。
首先,本发明的肽可通过修饰被赋予高稳定性。为此,乙酰基和酰胺基分别与具有seqidno:1所示氨基酸序列的肽的n端asn残基和c端pro残基连接。这种经修饰的肽也被称为“肽5-2a”。
其次,可通过修饰来提高具有seqidno:1所示氨基酸序列的肽的细胞穿透性效力。为此,用于与神经纤毛蛋白1受体结合的irgd肽序列(crgdkgpdc)与具有seqidno:1所示氨基酸序列的肽的c端pro残基连接。这种经修饰的肽也被称为“肽5-2b”。同样地,乙酰基和酰胺基团可分别与肽5-2b的n端asn残基和c端cys残基键合。这种经修饰的肽也被称为“肽5-2c”。
为了通过荧光/共焦显微术和生物成像容易地追踪肽5-2或5-2c的细胞内迁移途径和生物分布,可使含fitc荧光染料的化合物(fitc-(6-氨基己酸)-cys-(6-氨基己酸)-gly-gly-ser-gly-gly-ser-gly-gly)进一步与肽5-2或5-2c的n端asn残基连接。这种经修饰的肽也被称为“肽fitc-5-2”或“肽fitc-5-2c”。同样地,酰胺基团可与肽fitc-5-2c的c端cys键合。
此外,为了提高肽的血脑屏障(blood-brainbarrier,bbb)和细胞穿透效率,还可使具有这样功能的dnp2肽序列(kikkvkkkgrkgskikkvkkkgrkgg;lim等,natcommun(2015)6,8244)与肽5-2的n端asn残基连接。这种经修饰的肽也被称为“肽5-2d”。同样地,乙酰基和酰胺基可分别与肽5-2d的n端lys残基和c端pro残基连接。
为了便于分析肽5-2c与转录因子cp2c之间的结合,也可使生物素标记的lys与肽5-2c的c端cys残基的ε-nh2连接。这种经修饰的肽也被称为“肽5-2cb”。同样地,乙酰基和酰胺基可分别与肽5-2cb的n端asn残基和c端lys残基连接。
本发明还提供了用于预防和治疗癌症的含有具有seqidno:1所示氨基酸序列的肽作为活性成分的药物组合物,以及用于预防和治疗癌症的含有具有seqidno:1所示氨基酸序列的肽作为活性成分的健康功能性食品组合物。
如本文中使用的术语“治疗”意指通过施用根据本发明的肽或药物组合物,癌症症状已经好转或减轻的所有作用。
如本文中使用的术语“施用”意指通过任何合适的方法将物质(即本发明的肽衍生物或药物组合物)引入到对象中。本发明的肽衍生物或药物组合物可经由任何常见的途径施用,只要其能够到达期望的组织即可。具体地,考虑多种施用途径,包括腹膜内、静脉内、肌内、皮下、皮内、经口、经表面、鼻内、肺内和直肠内,但本发明不限于这些施用途径。由于本发明的经口施用的肽不被消化,故本发明的组合物优选用活性药物包被或者被保护免于在胃中降解。优选地,本发明的组合物可以以可注射制剂的形式施用。本发明的药物组合物可使用能够将活性物质递送到靶细胞的某种装置来施用。
如本文中使用的术语“含有活性成分”意指存在足以以可应用于任何医学治疗的合理收益/风险比治疗疾病的量。组合物的有效剂量水平可根据包括以下的因素决定:疾病的类型和严重程度、药物的活性、患者对药物的敏感性、施用时间、施用途径、排泄率、治疗持续时间、与组合物组合使用的药物,以及医学领域已知的其他因素。本发明的肽或药物组合物可单独地或与其他治疗剂组合施用,并且可依次地或者与常规治疗剂同时施用。本发明的肽或药物组合物可以以单一剂量或多剂量施用。考虑到所有上述因素,重要的是以可以表现出最大效果而不引起副作用的最小量施用本发明的肽或药物组合物。根据本发明的肽或药物组合物的量可以容易地由本领域技术人员确定。根据本发明的药物组合物的剂量和施用频率根据作为活性成分的药物种类以及多种相关因素,包括待治疗的疾病的类型,施用途径,患者的年龄、性别和体重和疾病的严重程度确定。
本发明的药物组合物还可包含一种或更多种可药用载体,只要其含有本发明的肽作为活性成分即可。合适的可药用载体的实例包括:用于经口施用的粘合剂、润滑剂、崩解剂、赋形剂、增溶剂、分散剂、稳定剂、助悬剂、着色剂和风味剂;用于可注射制剂的缓冲剂、防腐剂、疼痛缓解剂、增溶剂、等渗剂和稳定剂;以及用于表面施用的基质、赋形剂、润滑剂和防腐剂。本发明的药物组合物可用上述可药用载体进行配制。这些制剂的实例包括:用于经口施用的片剂、锭剂、胶囊剂、酏剂、混悬剂、糖浆剂和糯米纸囊剂以及用于可注射制剂的单位安瓿和多剂量形式。本发明的药物组合物还可配制成其他制剂,例如,溶液剂、混悬剂、片剂、丸剂、胶囊计和持续释放制剂。
适用于根据本发明的药物组合物制剂的载体、赋形剂和稀释剂的实例包括乳糖、右旋糖、蔗糖、山梨醇、甘露醇、木糖醇、赤藓糖醇、麦芽糖醇、淀粉、阿拉伯树胶橡胶、藻酸盐、明胶、磷酸钙、硅酸钙、纤维素、甲基纤维素、微晶纤维素、聚乙烯吡咯烷酮、水、羟基苯甲酸甲酯、羟基苯甲酸丙酯、滑石、硬脂酸镁和矿物油。本发明的药物组合物还可包含选自填料、抗凝剂、润滑剂、润湿剂、风味剂和防腐剂的至少一种添加剂。
本发明的肽用作健康功能性食品组合物的活性成分。本发明的食品组合物可与已知具有抗癌活性的一种或更多种活性成分组合使用。本发明的食品组合物还可包含一种或更多种营养学上可接受的食品补充剂。本发明的健康功能性食品组合物旨在包括用于所有类型的食品,例如功能性食品、营养补充剂、健康食品和食品添加剂的组合物。
食品组合物的类型可通过本领域已知的一般方法以多种形式制备。例如,健康食品可制备成多种制剂,例如片剂、丸剂、散剂、胶囊剂、口香糖、维生素混合物、汁和饮料。本发明的食品组合物可以通过造粒、包封或粉碎制成可食用制剂。本发明的食品组合物可包含通常添加用于食品生产的一种或更多种成分,例如蛋白质、碳水化合物、脂肪、营养物和调味剂。例如,本发明的食品组合物可制备成饮料。在这种情况下,本发明的食品组合物还可包含选自柠檬酸、高果糖玉米糖浆、糖、葡萄糖、乙酸、苹果酸、果汁、枣提取物和甘草提取物的至少一种添加剂。本发明的食品组合物还可包含选自本领域通常使用的食品添加剂(例如风味剂、甜味剂、着色剂、填料和稳定剂)的至少一种食品补充剂。本发明的食品组合物还可含有风味剂或天然碳水化合物,如一般饮料。这样的天然碳水化合物的具体实例包括单糖,例如葡萄糖和果糖;二糖,例如麦芽糖和蔗糖;多糖,例如糊精和环糊精;以及糖醇,例如木糖醇、山梨醇和赤藓糖醇。合适的风味剂的实例包括天然风味剂(索马甜和甜叶菊提取物(例如莱鲍迪苷a和甘草甜素))和合成风味剂(糖精和阿斯巴甜)。
此外,本发明的食品组合物还可含有选自以下的至少一种添加剂:营养补充剂、维生素、矿物质(电解质)、甜味剂(例如合成和天然的甜味剂)、着色剂、填料(乳酪和巧克力)、果胶酸及其盐、藻酸及其盐、有机酸、保护性胶体增稠剂、ph调节剂、稳定剂、防腐剂、甘油、醇和用于碳酸饮料中的碳酸化剂,其可单独地或组合使用。
具体实施方式
将参考以下实施例更详细地说明本发明。提供这些实施例以帮助理解本发明,并且不旨在限制本发明的范围。
实施例1:通过肽5抑制cp2c-dna结合
实施例1.1
使用由本发明人进行的先前研究中所公开的噬菌体展示技术鉴定的与鼠cp2c蛋白相互作用的5种肽基序(肽5、肽8、肽13、肽21和肽31)。
实施例1.2
为了分析实施例1.1中鉴定的五种肽是否与细胞中的cp2c结合以影响cp2c与dna结合的能力,如由本发明人进行的另一项先前研究所公开的进行dna免疫沉淀(dip)测定(kim等,adnaimmunoprecipitationassayusedinquantitativedetectionofinvitrodna-proteincomplexbinding,analyticalbiochemistry.2013;441:147-151)。
总之,通过用5mm六亚甲基双乙酰胺(hmba)处理来诱导鼠红细胞白血病(murineerythroidleukemia,mel)细胞系的分化,并且在分化之后第2天分离细胞核提取物。对于核提取物分离,首先收获所述细胞系的1×106个细胞并用pbs洗涤。在添加200μl的核提取缓冲液a(10mmhepes、10mmkcl、0.1mmedta、0.1mmegta、0.5mm苯甲磺酰氟、蛋白酶抑制剂混合物(roche))之后,使反应在4℃下进行15分钟。向反应混合物添加0.6%np-40。对所得的混浊溶液进行离心并弃去上清液。向离心之后剩余的沉淀物添加50μl的核提取缓冲液c(20mmhepes、400mmnacl、1mmedta、1mmegta、1mm苯甲磺酰氟、蛋白酶抑制剂混合物(roche))。通过在4℃下轻敲5分钟至10分钟使沉淀物松动,然后离心。仅将上清液用于后续的实验。
随后,使5μg的核提取物和[α-32p]dctp-标记的dna探针(对应于存在于小鼠α-球蛋白启动子中的共用cp2c结合位点以及在α-球蛋白基因的起始密码子中-156与-124位点之间的序列,对于所述序列的细节参见kim等的文章)与结合缓冲液(4%甘油、10mmtris-hcl、1mmdtt、1mmedta和0.1%np-40)反应15分钟,并且向其中添加逐渐增加量(0.2μg、0.5μg和1μg)的经鉴定的cp2c结合肽(肽5、肽8、肽13、肽21和肽31)。然后,在室温下继续再反应15分钟。向每个反应混合物添加20μl的50%蛋白g-琼脂糖珠悬液。所得的混合物在4℃下反应1小时(h)以除去非特异性结合。在添加2μg的抗-cp2c抗体(cosmogenetech)之后,反应在4℃下进行10小时。向与cp2c抗体结合的反应产物添加20μl的50%蛋白g-琼脂糖珠悬液。反应在4℃下进行2小时。将反应混合物用细胞裂解缓冲液(50mmtris-hcl、150mmnacl、1mmedta和1mm苯甲磺酰氟)洗涤三次。向cp2c结合的dna探针添加洗脱缓冲液(50mmtris-hcl、10mmedta和1%十二烷基硫酸钠)。使混合物在65℃下反应1小时。将cp2c蛋白从dna探针移除,并使用闪烁计数器测量dna探针的剩余量。
为了研究由总共6个区域构成的全长cp2c序列的哪个肽序列与cp2c区域相互作用,全长cp2c序列和其中缺失cp2c序列的n-端和c-端的多种cp2c突变体的序列示意性地在图1中示出。图2示出了通过经鉴定的五种cp2c-结合肽(肽5、肽8、肽13、肽21和肽31)的dna免疫沉淀(dip)测定测量的dna-cp2c复合物的结合亲和力。参照图1和图2,肽5以浓度依赖性方式特异性地抑制cp2c-dna结合。
实施例2:通过肽5-1和肽5-2抑制cp2c-dna结合
将由12个氨基酸构成的cp2c-结合肽5分为肽5-1和肽5-2,每个肽均由6个氨基酸构成。此后,通过与实施例1中所述相同的方法分析肽5-1和肽5-2抑制cp2c与dna结合能力的能力。
图3示出了通过对肽5、肽8、肽5-1和肽5-2的dip测定来测量dna-cp2c复合物的结合亲和力。参照图3,肽8和肽5-1可忽视地抑制cp2c结合dna的能力,而肽5-2特异性抑制cp2c与dna之间的结合,这与肽5相当。
实施例3:本发明合成肽的结构和表征
由于实施例1-2中所述的cp2c-结合肽非常不稳定,在培养期间处理细胞时它们倾向于在培养溶液中降解,并且在细胞处理时不容易穿透细胞膜,因此限制了其在体内实验中的使用。因此,需要修饰cp2c-结合肽以得到其高细胞穿透性。为了修饰每种肽以得到高稳定性,乙酰基和酰胺基分别与肽的n-端和c-端连接。作为得到高细胞穿透性的修饰,制备肽5c和肽5-2c,其中与神经纤毛蛋白1受体结合的irgd肽序列(crgdkgpdc)分别与肽5和肽5-2连接。经修饰的肽的肽序列列于表1中(肽在pepmicco.,ltd合成)。
表1
实施例4:显微观察肽5c和肽5-2c对癌细胞系的生长抑制和凋亡诱导
实施例4.1:显微观察肽5c和肽5-2c对正常细胞系与癌细胞系的生长抑制和凋亡诱导
肝癌细胞系(hepg2、hep3b),人胚胎肾细胞系(293t),乳腺癌细胞系(mcf-7、mda-mb-231),血液肿瘤细胞系(k562、hel、hl60)成胶质细胞瘤细胞系(u251、u373mg、u87mg),乳腺上皮细胞系(mcf10a),肺上皮细胞系(beas2b),人间充质干细胞(hmsc)和鼠原代t细胞(静息和活化的t细胞)以3000个细胞/孔的密度接种到96孔板中,补充50μl的培养溶液,并且在37℃和5%co2下培养1小时。将fqi1、肽5c和肽5-2c以2μm的浓度添加至细胞系接种的孔板,并在37℃和5%co2下培养48小时。培养完成之后,在倒置相差显微镜下观察细胞并且拍摄其形态变化。
图4a示出了癌细胞系的显微图像,并且图4b示出了正常细胞系、生物衍生的正常干细胞和分化细胞的显微图像。参照图4a和图4b,fqi1以浓度依赖性方式抑制正常细胞系以及癌细胞系的生长并诱导凋亡,而肽5c和肽5-2c特异性抑制癌细胞系的生长并诱导凋亡而基本上不影响正常细胞系。
实施例4.2:显微观察肽5-2c在乳腺球培养期间对癌细胞系的生长抑制和凋亡诱导
将癌细胞系(mcf-7、293t、mda-mb-231)以1×104个细胞/ml的密度接种到经聚hema涂覆的100phi培养皿中。生理盐水和肽5-2c以2μm的浓度添加至接种有所述细胞系的培养容器并且将细胞在37℃和5%co2下培养4天。在倒置相差显微镜下观察细胞每天一次并且拍摄其形态变化。图4c示出了乳腺球培养期间每天观察到的细胞系的显微图像。已知癌细胞系的乳腺球培养增加了癌干细胞的数量。参照图4c,通过用肽5-2c处理有效地抑制了乳腺球的形成并且降低了细胞生长,这表明肽5-2c将抑制癌干细胞的形成并诱导凋亡。
实施例4.3:显微观察肽5c和肽5-2c在胚胎上皮细胞上共培养期间对胚胎干细胞系的生长抑制和凋亡诱导
将每孔中的人胚胎干细胞系(h9)的两个集落接种到96孔板中,在其上铺板正常胚胎上皮细胞或用罗丹明荧光染色的胚胎上皮细胞,然后培养24小时。向接种细胞系的孔板以增加的浓度(0、0.5、1、2、3和10μm)添加生理盐水、fqi1、肽5c和肽5-2c,并将细胞在37℃和5%co2下培养3天。培养完成之后,在倒置相差显微镜下观察细胞并且拍摄其形态变化。
图4d示出了取决于处理组中处理浓度的72小时集落的显微图像(上)和代表性胚胎干细胞集落和荧光标记的共培养细胞的荧光显微图像(下)。图4d的最右边的红框图像(上)示出10μm处理组的图像中框的放大图像。参照图4d,对于fqi1和肽5c,胚胎干细胞和共培养的细胞系在≥3μm的浓度下死亡。相比之下,对于用肽5-2c处理的组,胚胎干细胞和共培养的细胞甚至在10μm的浓度下也没有死亡。这些结果表明,根据在实施例4.1中的正常细胞系中获得的结果,肽5-2c既不抑制全能干细胞的生长也不诱导凋亡。
实施例4.4:显微观察肽5c和肽5-2c对造血祖细胞和在分化成红细胞期间源自其的细胞的生长抑制和凋亡诱导
从人外周血中收集cd34+造血祖细胞(hematopoieticprogenitorcells,hpc)和分化成红细胞期间(分化之后3、7和14天)的细胞并且然后用增加浓度下的生理盐水、fqi1、肽5c和肽5-2处理72小时。在倒置相差显微镜下观察细胞,并拍摄其形态变化。将cd34+造血祖细胞在含有1%bsa的imdm培养基中培养。对于红细胞的分化诱导,前7天用epo、scf、il-3和氢化可的松培养细胞,并且然后在仅补充有epo、scf和il-3的培养基中培养。
图4e示出了处理组中造血祖细胞和分化诱导成红细胞期间(分化之后3天、7天和14天)的细胞的显微图像。参照图4e,当用包含浓度达10μm的肽5-2c的所有药物处理时,甚至没有观察到细胞的生长抑制和凋亡。
实施例5:肽5c和肽5-2c对癌细胞系的生长抑制和凋亡诱导:通过mtt测定定量测量细胞生存力
实施例5.1:分析当用肽5c和肽5-2c处理时肝癌细胞系的细胞生存力
将肝癌细胞系(hepg2、hep3b)以3000个细胞/孔的密度接种到96孔板中,并且然后向其中添加50μl的培养溶液。细胞在37℃和5%co2下培养1小时。fqi1、肽5c、肽5-2c以0、0.5、2、3和10μm的浓度添加到接种有肝癌细胞系的孔板。细胞在37℃和5%co2下培养24、48、72和96小时。培养完成之后,从孔板除去剩余的培养溶液,将培养物用mtt溶液(3-(4,5-二甲基噻唑-2-基)-2,5-二苯基四唑溴化物)稀释直到最终密度为500μg/ml,并且细胞在37℃和5%co2下培养3小时。除去剩余的mtt溶液,并且添加150μl的二甲基亚砜(dmso)。反应在室温下进行20分钟。在反应完成之后,使用微板读数仪测量吸光度。
图5a示出了当用增加浓度的肽处理一次后,hepg2细胞系和hep3b细胞系的相对生存力。参照图5a,肽5c和肽5-2c比fqi1更有效地抑制肝癌细胞系的生长并诱导凋亡。
实施例5.2:分析当用肽5c和肽5-2c处理时乳腺癌细胞系的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制乳腺癌细胞系(mcf7、mda-mb-231)的生长并诱导凋亡。
图5b示出了当用增加浓度的肽处理时,mcf7细胞系和mda-mb-231细胞系的相对生存力。参照图5b,肽5c和肽5-2c比fqi1更有效地抑制乳腺癌细胞系的生长并诱导凋亡。
实施例5.3:分析当用肽5c和5-2c处理时成胶质细胞瘤细胞系的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制成胶质细胞瘤细胞系(u251、u87mg、u343)的生长并诱导凋亡。
图5c示出了当用增加浓度的肽处理时,u251细胞系、u87mg细胞系和u343细胞系的相对生存力。参照图5c,肽5c和肽5-2c比fqi1更有效地抑制成胶质细胞瘤细胞系的生长并诱导凋亡。
实施例5.4:分析当用肽5c和肽5-2c处理时人血癌细胞系的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制人血癌细胞系(k562、hel、hl60)的生长并诱导凋亡。
图5d示出了当用增加浓度的肽处理时,k562细胞系、hel细胞系和hl60细胞系的相对生存力。参照图5d,肽5c和肽5-2c比fqi1稍微有效地抑制成胶质细胞瘤细胞系的生长并诱导凋亡。
实施例5.5:分析当用肽5c和肽5-2c处理时人胚胎肾癌细胞系的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制人胚胎肾癌细胞系(293t)的生长并诱导凋亡。
图5e示出了当用增加浓度的肽处理时293t细胞系的相对生存力。参照图5e,肽5c和肽5-2c类似于fqi1抑制成胶质细胞瘤细胞系的生长并诱导凋亡。
实施例5.6:分析当用肽5c和肽5-2c处理时人肺上皮细胞系和乳腺上皮细胞系的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制人肺上皮细胞系(beas2b)和乳腺上皮细胞系(mcf10a)的生长并诱导凋亡。
图5f示出了当用增加浓度的肽处理时,mcf10a细胞系和beas2b细胞系的相对生存力。
beas2b和mcf10a细胞系不是癌细胞但是是通常用作抗癌剂筛选实验中的正常对照组的正常上皮细胞系。参照图5f,fqi1显示浓度依赖性细胞毒性,而肽5c显示出比fqi1低的细胞毒性,肽5-2c甚至在10μm的浓度下也不产生显著的细胞毒性。
实施例5.7:分析当用肽5c和肽5-2c处理时原代培养的小鼠t淋巴细胞的细胞生存力
类似于实施例5.1的过程,确定fqi1、肽5c和肽5-2c是否有效地抑制原代培养的小鼠t淋巴细胞(静息和活化的t淋巴细胞)的生长并诱导凋亡。在本实验中,切除c57bl/6小鼠的脾,通过物理方法分离组织细胞,并通过0.45μm筛网以获得细胞悬液。使用macs将cd4+t细胞与悬浮细胞分离。将分离的t淋巴细胞(2.5×105个细胞/孔)在补充有10%血清的培养溶液中培养。通过在预先涂覆有抗-cd3和抗-cd28的板中培养来活化静息状态下所得的t淋巴细胞(2.5×105个细胞/孔)。
图5g示出了当用增加浓度的肽处理时原代培养的静息和活化的t淋巴细胞的相对生存力。参照图5g,即使在所有处理组(包括肽5-2c)中10μm浓度下,也没有观察到显著的细胞毒性。
实施例5.8:分析当用肽5c和肽5-2c处理时人间充质干细胞的细胞生存力
重复实施例5.1的过程以确定fqi1、肽5c和肽5-2c是否有效地抑制人间充质干细胞(hmsc)的生长并诱导凋亡。
图5h示出了当用增加浓度的肽处理时hmsc细胞系的相对生存力。与beas2b和mcf10a细胞系相似,hmsc细胞不是癌细胞,但是是正常细胞。参照图5h,fqi1显示出浓度依赖性细胞毒性而肽5-2c即使在高浓度下也没有引起显著的细胞毒性。
实施例5.9:分析当用肽5c和肽5-2c处理时人胚胎干细胞的细胞生存力
类似于实施例5.1的过程,确定fqi1、肽5c和肽5-2c是否有效地抑制通过与小鼠胚胎成纤维细胞(mef)共培养来保持多能性之人胚胎干细胞(hesc)的生长。在本实验中,将每个孔中的hesc(h9细胞)的两个集落共同培养在铺板有mef细胞的96-孔板中,mef细胞的生长已被丝裂霉素c(mmc)处理抑制。标记有罗丹明的mef细胞通常用于区分它们与hesc衍生的细胞。
图5i示出了当用增加浓度的肽处理时未标记的hesc(上)和罗丹明标记的hesc(下)的相对生存力。由于实施例5.6中上皮细胞显示出对所有处理的物质没有毒性,所以本实验中测量的细胞生存力可以被认为是hesc的生存力。参照图5i,fqi1显示出浓度依赖性细胞毒性,肽5c在高浓度下引起轻微的细胞毒性,而肽5-2c即使在高浓度下也没有引起显著的细胞毒性。
实施例5.10:分析当用肽5c和5-2c处理时人造血祖细胞和分化成红细胞期间由其衍生的细胞的细胞生存力
类似于实施例5.1的过程,确定fqi1、肽5c和肽5-2c是否有效地抑制cd34+造血祖细胞(hpc)和分化成红细胞期间(分化之后第3天、第7天和第14天)的细胞中癌细胞的生长并诱导凋亡。在本实验中,以与实施例4.4相同的方式制备和培养细胞。
图5j示出造血祖细胞和在分化诱导成红细胞期间(分化之后第3天、第7天和第14天)源自其的细胞的相对生存力。参照图5j,即使当用包含浓度达10μm的肽5-2c的所有药物处理时也没有观察到显著的细胞毒性。
图5k示出在实施例5a至实施例5j中进行的mtt测定数据中肽处理之后76小时(实施例5.10)和96小时的孔板的图像。
实施例6:分析多种癌细胞系中cp2c蛋白的表达水平
实施例6.1
通过western印迹比较并分析了总共19种细胞系(包括6种结肠癌细胞系,6种肺癌细胞系,3种乳腺癌细胞系,3种成胶质细胞瘤细胞系和1种宫颈癌细胞系)中的cp2c蛋白的表达水平。收集培养期间每种细胞系的1×106个细胞并用磷酸盐缓冲盐水(pbs)洗涤。使用细胞裂解缓冲液(50mmtris-hcl、ph7.4,150mmnacl、1mmedta、ph8,1%tritonx-100,1mm苯甲磺酰氟,蛋白酶抑制剂混合物(roche))获得细胞提取物。将预定量的蛋白质在sds-聚丙烯酰胺凝胶(7.5%至15%)上进行电泳,转移至pvdf膜,并在5%封闭溶液中反应1小时。将所述蛋白与作为一抗的抗-cp2cab(cosmogentech)在室温下反应1小时,并在室温下用作为二抗的辣根过氧化物酶缀合的抗兔ab(abcam)处理1小时。使用ecl系统(amersham-gehearlthcare)进行检测。
实验结果显示在图6a中。参照图6a,cp2c在多种癌细胞系中过表达。
实施例6.2
重复实施例6.1的过程以确认来自多种组织来源的血癌细胞系(mel、k562、hel、hl60),肝癌细胞系(hepg2、hep3b),表皮样癌细胞系(a431),结肠癌细胞系(hct116)和上皮细胞系(293t、beas2b、mcf10a)以及hmsc细胞系中cp2c蛋白的表达水平。
实验结果显示在图6b中。参照图6b,cp2c在多种癌细胞系中过表达,但是在正常细胞系(如hmsc)中没有观察到这种现象。
实施例7:测量肽5-2c处理之后多种癌细胞系中肽5-2c的ic50值
实施例7.1:测量肽5-2c处理之后多种癌细胞系和正常细胞系中肽5-2c的ic50值
基于在实施例5中对多种癌细胞系进行的mtt实验的结果,使用graphprismpad6程序计算肽5-2c处理48小时之后的ic50值(ic50是指将细胞的数目减少50%所需的每种物质的浓度)。
图7a示出了用于计算每种细胞系用肽5-2c处理48小时之后肽5-2c的ic50值的图以及计算的ic50值。参照图7a,肽5-2c基本上不影响正常对照细胞系的生长抑制和凋亡的诱导,但是ic50值平均为~1μm,与多种癌细胞系存在轻微的差异。
实施例7.2:测量肽5-2c处理不同时间段之后肽5-2c的ic50值
在用肽5-2c处理多种成胶质细胞瘤细胞系(u343、u373mg、u251和u87mg)一次之后,使用graphprismpad6程序随时间测量ic50值。
图7b示出了用于计算用肽5-2c处理每种细胞系一次之后48小时和96小时的ic50值的图以及计算的ic50值。参照图7b,48小时之后的ic50值类似于96小时之后的ic50值,表明肽5-2c对细胞生长的抑制和凋亡的诱导的作用维持96小时。
实施例7.3:测量在用fqi1、肽5c和肽5-2c处理之后mcf10a细胞系和beas2b细胞系中fqi1、肽5c和肽5-2c的ic50值
基于在实施例5中对肺上皮细胞系(beas2b)和乳腺上皮细胞系(mcf10a)进行的mtt实验的结果,使用sigma绘图程序计算fqi1、肽5c和肽5-2c处理之后的ic50值。
图8示出了用于计算用fqi1,肽5c和肽5-2c处理之后的ic50值的图以及计算的ic50值。参照图8,用fqi1处理正常对照细胞系抑制了细胞生长并诱导凋亡,类似于用肽5c的处理(fqi1的ic50值与mcf10a中肽5c的ic50值基本相同,但是肽5c的ic50值比beas2b中fqi1的ic50值高1.5倍),但是用肽5-2c处理基本上不影响正常对照细胞系的生长抑制和凋亡的诱导。
实施例8:比较并分析肽5-2c和肽5-2d对癌细胞系的生长抑制和凋亡的诱导:通过mtt测定定量测量细胞生存力
肽5-2c是肽5-2(seqidno:1)的变体,其末端pro残基与rgd肽序列(crgdkgpdc;seqidno:2)键合,所述rgd肽序列与神经纤毛蛋白1受体结合。这种修饰提高了肽的细胞穿透性。如上述实施例所证明的,肽5-2可以特异性抑制癌细胞系的生长并且可以诱导凋亡。有必要使用肽5-2和另一种细胞穿透肽的复合物,以确认肽5-2c对癌细胞的特异性生长抑制和凋亡的诱导的这些作用取决于肽5-2而不是rgd序列。为此,合成肽5-2d,其中已知dnp2肽序列(kikkvkkkgrkgskikkvkkkgrk;lim等,natcommun(2015)6,8244)具有穿透血脑屏障(bbb)和细胞的能力,将其与肽5-2的n-端asn残基连接。通过mtt测定定量测量当用肽5-2d处理时癌细胞系的细胞生存力并与肽5-2c的作用进行比较。
实施例8.1:分析当用肽5-2d处理时乳腺癌细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制乳腺癌细胞系(mcf7,mda-mb-231)的生长并诱导凋亡。
图9a示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时,mda-mb-231细胞系和mcf7细胞系的相对生存力。参照图9a,类似于肽5-2c,肽5-2d抑制乳腺癌细胞系的生长并有效地诱导凋亡。
实施例8.2:分析当用肽5-2d处理时成胶质细胞瘤细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制成胶质细胞瘤细胞系(u343,u87mg)的生长并诱导凋亡。
图9b示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时,u343细胞系和u87mg细胞系的相对生存力。参照图9b,类似于肽5-2c,肽5-2d抑制成胶质细胞瘤细胞系的生长并有效地诱导凋亡。
实施例8.3:分析当用肽5-2d处理时结肠癌细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制结肠癌细胞系(hct116、ht29)的生长并诱导凋亡。
图9c示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时hct116细胞系和ht29细胞系的相对生存力。参照图9c,类似于肽5-2c,肽5-2d抑制成胶质细胞瘤细胞系的生长并有效地诱导凋亡。
实施例8.4:分析当用肽5-2d处理时人胚胎肾癌细胞系和肝癌细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制人胚胎肾癌细胞系(293t)和肝癌细胞系(hepg2)的生长并诱导凋亡。
图9d示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时293t细胞系和hepg2细胞系的相对生存力。参照图9d,类似于肽5-2c,肽5-2d抑制人胚胎肾癌细胞系和肝癌细胞系的生长并有效地诱导凋亡。
实施例8.5:分析当用肽5-2d处理时人血癌细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制人血癌细胞系(u937、jurkat、hl60、hel)的生长并诱导凋亡。
图9e示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时,u937细胞系、jurkat细胞系、hl60细胞系和hel细胞系的相对生存力。参照图9e,类似于肽5-2c,肽5-2d抑制人胚胎肾癌细胞系和肝癌细胞系的生长并有效地诱导凋亡。
实施例8.6:分析当用肽5-2d处理时人乳腺上皮细胞系和肺上皮细胞系的细胞生存力
重复实施例5.1的过程以确定肽5-2d是否有效地抑制乳腺上皮细胞系(mcf10a)和肺上皮细胞系(beas2b)的生长并诱导凋亡。
图9f示出了当用增加浓度的作为阴性对照的生理盐水、作为阳性对照的肽5-2c、和肽5-2d处理时,mcf10a细胞系和beas2b细胞系的相对生存力。参照图9f,类似于肽5-2c,肽5-2d没有影响乳腺上皮细胞系和肺上皮细胞系的生长和凋亡。
图9g示出了在实施例8.1至实施例8.6中进行的mtt测定的数据中在肽处理之后96小时的孔板的图像。
实施例9:测量肽5-2d处理之后多种癌细胞系中肽5-2d的ic50值
基于在实施例8中对多种癌细胞系和正常细胞系进行的mtt实验的结果,使用graphprismpad6程序计算肽5-2c处理48小时之后的ic50值(ic50是指将细胞数目减少50%所需的每种物质的浓度)。
图10示出了用于计算用肽5-2c和肽5-2d处理每种细胞系一次之后48小时的ic50值的图以及计算的ic50值。参照图10,类似于肽5-2c,肽5-2d基本上没有影响正常对照细胞系的生长抑制和凋亡的诱导,但是ic50值平均为~3μm,与多种癌细胞系存在轻微的差异。
总之,当细胞穿透促进肽(例如rgd或dnp2)与肽5-2的c-端或n-端连接时,肽5-2可以特异性地抑制癌细胞的生长并诱导凋亡。
实施例10:分析肽5-2c处理对移植有多种癌细胞系的小鼠模型中肿瘤生长的抑制作用并预测一般生理毒性
实施例10.1:分析肽5-2c处理对移植有成胶质细胞瘤细胞系(u343)的小鼠模型中肿瘤生长的抑制作用并预测一般生理毒性
将u343成胶质细胞瘤细胞系(2.5×106个细胞/50μl)皮下注射到5周龄雄性balb/c裸鼠(n=6只/组)的背部,将肽5-2c(3mg/kg)从当肿瘤大小增加到约80mm3时每三天直接注射到肿瘤部位中,测量小鼠的体重和肿瘤的体积,在肽注射之后第70天处死小鼠,并切除肿瘤,血液和主要器官。使用游标卡尺测量肿瘤大小,并通过(长轴×短轴2)/2计算肿瘤体积。使用涂覆有edta的微毛细管从每只小鼠的眼动脉中收集外周血,并储存在涂覆有5.4mgedta的样品管中。通过解剖切除肿瘤和主要器官。使用coulterlh750血液学分析仪对收集的血液进行基本完全的血细胞计数(cbc)。肿瘤称重之后,将肿瘤和主要器官用4%甲醛染色。通过石蜡切片制备组织薄片并用苏木精/伊红染色。使用excel程序对获取的数据进行统计分析。
图11示出了在移植有u343成胶质细胞瘤细胞系的小鼠模型中肽5-2c抑制肿瘤生长的能力和肽5-2c的生理特性。具体地,图11a示出了实验期间的肿瘤体积,图11b示出了小鼠的体重变化,图11c示出了肽施用之后第70天小鼠的图像,图11d示出了切除的肿瘤的图像,图11e示出了从小鼠中切除的肿瘤的重量及其平均值,图11f示出了对照组和肽处理组中小鼠的多种标准血液指数,并且图11g示出了主要器官(脾、肝、肺和肾)的h/e染色图像。参照图11a至图11g,与仅施用生理盐水(对照)相比,当将肽5-2c直接施用于肿瘤部位时,小鼠的体重、小鼠的主要cbc指数以及主要器官的组织学/解剖特征不存在差异。然而,肿瘤体积和重量减少了~60%。总之,肽5-2c(3mg/kg)在移植有成胶质细胞瘤细胞系的肿瘤模型中有效地抑制了肿瘤的生长而不引起特别的生理毒性。
实施例10.2:分析肽5-2c处理对移植有表皮样癌细胞系(a431)的小鼠模型中肿瘤生长的抑制作用并预测一般生理毒性
将a431细胞系(5×106个细胞/100μl)皮下注射到7周龄雄性balb/c裸鼠(n=4只/组)的背部,肽5-2c(1×ic50;1.7mg/kg,2×ic50;3.5mg/kg)从当肿瘤大小增加到约50mm3时每隔一天通过尾静脉注射到小鼠中,共计5次,测量肿瘤体积,在肽注射之后第13天处死小鼠,并切除肿瘤和主要器官。肿瘤称重之后,将肿瘤和主要器官用4%甲醛染色。通过石蜡切片制备组织薄片并用苏木精/伊红染色。使用excel程序对获取的数据进行统计分析。
图12a至12d示出了在移植有a431人表皮样癌细胞系的小鼠模型中,肽5-2c抑制肿瘤生长的能力和肽5-2c的生理特性。图12a示出了实验期间肿瘤的体积,图12b示出了小鼠和切除的肿瘤的图像,图12c示出了从所述组中的小鼠切除的肿瘤的重量及其平均值,并且图12d示出了所述组的主要器官(脾、肝、肺、血管和肌肉)的h/e染色图像。参照图12a至图12d,当每隔一天通过静脉注射肽5-2c总共5次时,源自a431表皮样癌细胞系的肿瘤的体积和重量分别降低≥70%和≥60%,并且主要器官的组织学/解剖特征没有显著变化。总之,这些结果表明通过静脉内施用肽5-2c可以有效地抑制源自表皮样癌细胞系的肿瘤的生长。
实施例10.3:分析肽5-2c处理对移植有乳腺癌细胞系(mda-mb-231)的小鼠模型中肿瘤生长的抑制作用并预测一般生理毒性
将已确认转移至肺组织的mda-mb-231细胞系(lm1细胞;1×106个细胞/40μl)注射到12周龄雌性balb/c裸鼠(n=5只/组)的乳腺脂肪垫中,肽5-2c(1×ic50;1.7mg/kg=52μg/小鼠,2×ic50;3.5mg/kg=104μg/小鼠)从当肿瘤大小增加到约50mm3时每三天通过尾静脉注射到小鼠中,共计5次,测量肿瘤体积,在肽注射之后第30天处死小鼠,并切除肿瘤和主要器官。使用coulterlh750血液学分析仪对收集的血液进行基础cbc。肿瘤称重之后,将肿瘤和主要器官用4%甲醛染色。通过石蜡切片制备组织薄片,并用苏木精/伊红染色。使用excel程序对所获取的数据进行统计分析。
图13a至图13d示出了在移植有能够转移至肺组织的mda-mb-231细胞系(lm1细胞)的动物模型中肽5-2c抑制转移至肺组织和肿瘤生长的能力以及肽5-2c的生理特性。图13a示出了在注射乳腺癌细胞系之后第30天处死的小鼠和切除的肿瘤的图像,图13b示出了从对照组和肽处理组的小鼠切除的肿瘤的重量及其平均值,图13c示出了对照组和肽处理组中转移至肺的肿瘤灶的数量的平均值和标准偏差,并且图13d示出了对照组和肽处理组中的cbc指数。参照图13a至图13d,当将肽5-2c通过静脉注射到移植有已确认转移至肺组织的mda-mb-231乳腺癌细胞系的肿瘤小鼠模型中,与对照组中的那些相比,肿瘤重量减少了~30%并且转移至肺的肿瘤灶的数量减少了~50%。另一方面,肽5-2c注射组中的cbc指数与对照组中的那些相似,但是移植有乳腺癌细胞系的对照小鼠中的白细胞数目相当于正常小鼠中的两倍,并且在注射肽5-2c的小鼠中的白细胞数目接近正常值(图13d)。总之,在移植有乳腺癌细胞的小鼠模型中肽5-2c在抑制肿瘤的生长和癌症转移方面是有效的。此外,肽5-2c可以将由癌细胞移植引起的提高的wbc值恢复至正常水平而不引起任何血液学毒性。
实施例11:分析当用肽5c和肽5-2c处理时cp2c的转录活性
293t细胞以1×105个细胞/孔的密度接种到12孔板中。接种之后12小时,通过使用effectene方法用报道载体(野生型报道子将包含两次重复cp2c结合位点的gata-1基因的增强子序列连接到荧光素酶基因的上游,并且mut1/3、mut2/4和mut1-4报道子引起增强子序列中存在的cp2c结合位点的突变)与多种flag-cp2c、ha-cp2b和ha-pias1表达载体的组合来转染细胞。在培养1小时之后,将细胞用浓度为2μm的fqi1、肽5c和肽5-2c处理,然后培养48小时。收集细胞并在250μl的被动细胞裂解缓冲液(promega)中裂解以获得提取物。将20μl的细胞提取物和双荧光素酶测定试剂盒(promega)用于测量glomax(promega)中标记的荧光素酶基因的活性(通过western印迹确认每个表达载体的表达)。
结果示于图14a至图14e中。具体地,图14a示出了使用具有图14e所示的正常cp2c靶序列的报道载体所获得的结果。图14b至图14d分别示出了使用突变体2/4、1/3和1-4所获得的结果。图14e示出了正常序列和突变体的序列。参照图14a至图14e,肽5c和肽5-2c与fqi1相比有效地抑制了cp2c-介导的转录活性。在其中cp2c结合位点被突变的突变体中没有发现这种作用,并且具体观察到不仅在cp2c同源四聚体(jbiolchem,1994;genescells,1998)的位点起作用而且在cp2c/cp2b/pias1异源六聚体(cbpcomplex,nucleicacidsres.kang等,2010)的序列中起作用。
实施例12:分析肽5-2c处理对癌细胞特异性生长中断和凋亡诱导的作用机制
实施例12.1:分析当用肽5-2c处理时多种癌细胞系的细胞周期
将mel、k562、mcf-10a和mda-mb-231每种细胞系的5×105个细胞接种到100mm培养皿中。在接种之后24小时,将细胞用肽5-2c处理,以使得最终浓度为0、1、2、3和10μm。在肽5-2c处理之后48小时,通过胰蛋白酶处理收集细胞。然后,将1×106个细胞悬浮于0.5mlpbs中,并通过吹打分离成单个细胞。将细胞悬液转移到包含4.5ml70%乙醇的离心管中,并在4℃下静置2小时以固定细胞。通过离心获得细胞沉淀物,用5ml的pbs溶液洗涤,充分悬浮在1ml的pi染色溶液中,并在黑暗中染色30分钟。在bdfacsaria中测量细胞荧光,并使用由相同公司提供的软件分析细胞周期。
图15a至图15c示出在用0、1、2、3和10μm的多种浓度的肽5-2c处理细胞系之后48小时通过facs细胞周期分析多种细胞系(mel、k562、mcf-10a、mda-mb-231)的结果,从而分析肽5-2c对细胞周期的影响。具体地,图15a示出了在facs期间取决于dna的量的细胞分布,图15b示出了细胞周期阶段中的定量细胞分布,并且图15c示出了显示凋亡的subg1期中的细胞的放大分布。参照图15a至图15c,当用肽5-2c处理时,在其中生长中断和凋亡诱导不可能发生的mcf-10a细胞系中没有观察到细胞周期的显著变化。相比之下,在以高浓度(10μm)处理时诱导生长中断和凋亡的mel细胞系中,在facs分析中,仅在高浓度下,subg1期的细胞数目会显著增加。相比之下,在其中响应于肽5-2c敏感地诱导生长中断和凋亡的k562细胞系和mda-mb-231细胞系中,s期的细胞数目减少,g2/m期的细胞数目增加,并且subg1期的细胞数目增加取决于肽5-2c的浓度。从这些结果可以得出:通过肽5-2c的癌细胞特异性生长中断和凋亡诱导的现象是由抑制转化为细胞周期的g2/m期的细胞生长中断而产生的。
实施例12.2:分析当用不同浓度的肽5-2c处理不同时间段时mcf7细胞系中细胞周期相关的标志物基因的表达谱
将mcf7乳腺癌细胞系的1×105个细胞接种在6孔板中。接种之后24小时,将细胞用最终浓度为2μm的肽5-2c处理。在0、24、48和72小时之后,以与实施例6相同的方式收集细胞提取物。使用针对细胞周期相关的标志物基因的多种抗体进行western印迹。另一方面,从用最终浓度为1、2和3μm的肽5-2c处理48小时的组中获得细胞提取物并以相同的方式进行western印迹。
图16a至图16d示出了当用不同浓度的肽5-2c处理mcf7细胞系不同时间段时,细胞周期相关的标志物基因的表达谱。具体地,图16a示出了当用2μm肽5-2c处理72小时时多种细胞周期相关的标志物基因的时间依赖性表达谱,如通过western印迹测定。图16b示出了用多种浓度(0、1、2和3μm)的肽5-2c处理48小时之后多种细胞周期相关的标志物基因蛋白的表达谱,如通过western印迹测定。并且图16c和图16d分别示出了图16a和图16b中标志物基因蛋白表达的定量变化。参照图16a至图16d,随着肽5-2c处理时间和浓度的增加,细胞周期蛋白e、p21和p53的表达逐渐提高,并且cdk1和细胞周期蛋白b1的表达逐渐降低,表明肽5-2c在g2期中中断了细胞周期。
实施例12.3:在mda-mb-231细胞系和mcf7细胞系中凋亡相关的标志物基因的表达谱依赖于肽5-2c处理时间和浓度
以与实施例12.2相同的方式,将mda-mb-231和mcf7乳腺癌细胞系用生理盐水和2μm肽5-2c处理达72小时。在0、24、48和72小时之后,收集细胞提取物。使用针对细胞周期相关的标志物基因的多种抗体进行western印迹。
图17示出了用2μm肽5-2c处理mda-mb-231细胞系和mcf7细胞系之后凋亡相关的标志物基因蛋白的时间依赖性表达谱。参照图17,随着肽5-2c处理时间增加,抗凋亡相关的标志物基因mcl1和bcl2的表达逐渐降低,凋亡促进相关的标志物基因bim、bax、bak、切割的cas3和cas8、以及pro-cas11的表达提高,并且pro-cas3、pro-cas8、pro-cas9、pro-cas12的表达逐渐降低。这些结果可以得出这样的结论:随着肽5-2c处理时间的增加,抗凋亡相关的标志物基因的表达降低,并且凋亡促进相关的标志物基因的表达提高,从而导致凋亡的诱导。假设提高的p53通过线粒体途径诱导内在凋亡并且通过凋亡受体介导的pro-cas8活化诱导凋亡(ashkenazia,natrev,2008)。
实施例12.3:电子显微分析肽5-2c处理时mda-mb-231细胞系和mcf10a细胞系的细胞形态
将1.5×106个细胞的mda-mb-231细胞系和5×105个细胞的mcf10a细胞系接种到100mm培养皿中。接种1小时之后,用生理盐水、2μm肽5-2c和2μmfqi1处理细胞。在培养24和48小时之后,收集细胞。离心培养溶液并且通过胰蛋白酶处理收集表面粘附的细胞。将收集的细胞用生理盐水洗涤一次。将细胞沉淀物直接用2.5%水性戊二醛溶液处理两次(每次10分钟)。制备电子显微术的样品并根据一般方法成像。
图18示出了显示在用2μm肽5-2c处理mda-mb-231细胞系和mcf10a细胞系之后24小时和48小时细胞的形态的电子显微图像。用生理盐水处理的细胞和用2μmfqi1处理的细胞分别用作阴性对照和阳性对照。参照图18,对于发现生长中断和凋亡诱导对肽5-2c敏感的mda-mb-231细胞系,与fqi1处理组相比,在肽5-2c处理组中观察到显著不同的细胞内结构异常。对于通过肽5-2c处理未发现生长中断和凋亡诱导的mcf10a细胞系,在肽5-2c处理组中没有观察到细胞内结构异常,而细胞内结构异常在肽处理之后24小时开始出现并且在fqi1处理组中在肽处理之后48小时观察到相当大的核破裂。总之,通过肽5-2c的肿瘤细胞特异性生长中断和凋亡诱导基于与fqi1不同的机制。在正常的乳腺上皮细胞系mcf10a中,通过肽5-2c处理没有引起细胞内结构修饰,而fqi1引起了严重的细胞内结构修饰。这些结果与实施例4、实施例5、实施例7和实施例8中获得的结果非常一致。
实施例13:分析在mda-mb-231细胞系中fitc荧光标记的肽5-2c之细胞内迁移途径的跟踪
为了验证癌细胞特异性生长中断和凋亡诱导是由肽5-2c引起的,并且irgd简单地帮助肽的细胞穿透,合成三种fitc-荧光标记的肽(fitc-irgd、fitc-5-1c和fitc-5-2c)。以与实施例5.1相同的方式,将mda-mb-231细胞系用生理盐水和fitc缀合的肽处理96小时并且通过mtt测定计算细胞生存力和ic50值。作为参考,由肽5的n-端残基的前6个氨基酸构成的肽5-1不影响cp2c与dna结合的能力(参见图3)。每个fitc缀合的肽在实施例5.1所述的相同条件下处理30分钟。将孔用生理盐水洗涤三次,并用新的代替。随后获取延时相差显微图像和荧光显微图像,持续24小时(在肽处理之后1、2、4、8、12和24小时)。使用photoshop程序合并细胞图像和荧光图像。
图19a至图19d是确认在mda-mb-231细胞系中fitc-缀合的肽5-2c随时间经过的细胞内迁移途径的图。具体地,图19a示出了mda-mb-231细胞系用浓度为0、0.5、1、2、3和10μm的生理盐水和fitc-缀合的irgd、fitc-5-1c和fitc-5-2c肽处理96小时之后mtt测定的细胞生长速率的变化,图19b示出了用于计算在处理之后48小时处理组的ic50值的图以及计算的ic50值,图19c示出了在96小时显示示例性mtt测定的孔板的图像,并且图19d示出了显示在用肽处理之后24小时fitc-缀合的肽(各2μm)的时间依赖性细胞内分布的图像。参照图19a至图19d,仅fitc-缀合的肽5-2c抑制细胞生长并且其ic50值与非fitc-缀合的肽5-2c的ic50值相似。这证明了癌细胞特异性生长中断和凋亡诱导通过肽5-2c引起并且irgd简单地帮助肽的细胞穿透。细胞穿透在肽处理之后2小时开始出现,并且细胞穿透的肽5-2c累积在细胞核中,而其他肽(irgd和肽5-1c)保留在细胞质中,或者在8小时之后容易降解。
实施例14:鉴定与肽5-2c相互作用的cp2c蛋白区域以及该区域中的氨基酸
根据前述实施例获得的结果,可以认为由肽5-2引起的癌细胞特异性生长中断和凋亡诱导是因为肽5-2与cp2c结合以阻碍cp2c与表现出癌细胞特征的靶基因的调节位点的结合。为了支持这一假设,需要鉴定与肽5-2c相互作用的cp2c蛋白区域和直接与肽5-2c结合的区域中的氨基酸。因此,尝试获得突变体cp2c蛋白质,其中存在于与图1中所示的肽5结合的cp2c区域中的极性氨基酸被ala替换,并通过酶联免疫吸附测定(elisa)分析突变体cp2c蛋白与肽5-2c的结合。为了便于这种分析,合成了肽5-2cb,其中生物素标签与肽5-2c的c-端pro残基键合。为了确认肽5-2cb是否诱导癌细胞特异性生长中断和凋亡,如肽5-2c,通过与实施例5.1相同的方法在mda-mb-231细胞系和mcf7细胞系中的两个不同批次的肽5-2cb上进行mtt测定。还从mtt测定的结果计算ic50值。为了获得19个突变体cp2c蛋白质,其中存在于与肽5-2c结合的cp2c区域中的极性氨基酸被ala替换,合成了具有编码相应突变体氨基酸的序列的寡核苷酸并进行定点诱变。将编码突变体cp2c基因的碱基序列克隆到pgex4载体中,并使用与针对gst-标签的抗体结合的珠纯化突变体gst-融合cp2c蛋白。以与实施例1相同的方式确认突变体cp2c与cp2c结合位点(α-球蛋白启动子上的cp2c结合碱基序列)的结合。通过直接elisa确认每个突变体cp2c与肽5-2cb的结合,其用于通过噬菌体展示技术鉴定cp2c结合肽(kang等,febsj,2005;272:1265-1277)。使用能够在肽5-2cb中识别生物素标签的hrp-缀合的链霉亲和素珠来确认肽5-2cb与每个突变体cp2c蛋白之间的结合。基于rosetta建模算法预测了cp2c的306至396氨基酸区域的三级结构模型并且使用swissprot程序获得了靶dna结合和肽5-2c结合的重要氨基酸。
图20a至图20d比较了肽5-2cb(生物素标记的肽5-2c)对癌细胞存活的影响与肽5-2c的影响。具体地,图20a示出了用肽5-2c和两个不同批次的肽5-2cb以增加浓度处理一次之后mda-mb-231细胞系的相对生存力,图20b示出了在肽处理之后96小时的孔板的图像,图20c示出了用于计算用每种肽处理mda-mb-231细胞系之后48小时的ic50值的图以及计算的ic50值,并且图20d示出了用于计算用每种肽处理mcf7细胞系之后48小时的ic50值的以及计算的ic50值。参照图20a至图20d,类似于肽5-2c,肽5-2cb诱导癌细胞特异性细胞生长中断和凋亡,并具有与肽5-2c相似的ic50值。这些结果表明,肽5-2cb可以有效地用于图21a至图21d所示的实验。
图21a至图21d示出了用于鉴定肽5-2c结合的cp2c蛋白区域中的dna结合或肽5-2c结合之重要氨基酸的一系列实验的结果。具体地,图21a示出了肽5-2c和19个gst-融合突变体的结合区的列表,所述突变体由位点的氨基酸序列中的极性氨基酸被丙氨酸替换而产生,图21b示出了使用gst抗体珠与存在于放射性标记的α-球蛋白启动子中的cp2c结合位点的dna探针之从大肠杆菌中过表达的突变体纯化的gst-融合蛋白结合的dna-ip的结果,图21c示出了用于纯化的gst-融合蛋白与肽5-2c结合的直接elisa的结果,并且图21d示出了基于用于cp2c的306至396位氨基酸区域的rosetta建模算法预测的用于与三级结构的dna结合和肽5-2c结合的重要氨基酸。参照图21a至图21d,包含α-螺旋的cp2c蛋白的322至389位氨基酸区域识别cp2c的靶碱基序列,并参与与肽5-2的结合。特别地,e332、r339、r341和r387氨基酸对于结合dna是重要的,而e332、r339、r341和r347氨基酸对于结合肽5-2是重要的。总之,肽5-2竞争性地结合重要的e332、r339和r341以结合至cp2c的靶dna碱基序列,使得cp2c不可能结合至靶碱基序列。结果,肽5-2抑制参与转化至细胞周期g2/m期的cp2c靶基因的表达,从而诱导癌细胞特异性生长中断和凋亡。
工业适用性
本发明的肽5-2以非常高的效力穿透癌细胞的细胞膜,并且能够特异性结合至cp2c。因此,本发明的肽抑制了cp2c的活性,因此通过多种cp2c介导的癌细胞中的细胞生长抑制和凋亡诱导来有效地治疗癌症。
<110>industry-universitycooperationfoundationhanyanguniversity
<120>具有抗癌活性的肽,含有有效量的所述肽的用于预防和治疗癌症的药物组合物和健康功能性食物组合物
<130>jkpct-023
<160>4
<170>kopatentin2.0
<210>1
<211>6
<212>prt
<213>人工序列
<220>
<223>人工合成的肽
<400>1
asntyrproglnargpro
15
<210>2
<211>9
<212>prt
<213>人工序列
<220>
<223>人工合成的肽
<400>2
cysargglyasplysglyproaspcys
15
<210>3
<211>9
<212>prt
<213>人工序列
<220>
<223>人工合成的肽
<220>
<221>mod_res
<222>(1)..(3)
<223>no.1lys和no.3lys是6-氨基己酸
<400>3
lyscyslysglyglyserglyglyser
15
<210>4
<211>26
<212>prt
<213>人工序列
<220>
<223>人工合成的肽
<400>4
lysilelyslysvallyslyslysglyarglysglyserlysilelys
151015
lysvallyslyslysglyarglysglygly
2025
- 还没有人留言评论。精彩留言会获得点赞!