一种提高植物抵御RNA病毒的能力的方法与流程
本发明涉及植物培育技术领域,具体涉及一种提高植物抵御rna病毒的能力的方法。
背景技术:
植物病毒病会对许多植物造成危害,会极大的影响植物的生长,在农业生产中造成严重的损失。根据病毒的基因组构成,又分为rna病毒和dna病毒两种。其中植物rna病毒广泛存在于自然界中,具有非常广泛的宿主。已知的rna病毒中多数会通过各种途径传播,给经济性作物带来巨大的损失。
以黄瓜花叶病毒为例,它是一种由蚜虫传播的非常严重的植物病毒病害,病毒粒子呈等轴对称的二十面体,基因组为三分子线形正义rna。黄瓜花叶病毒是寄主范围最多、分布最广、最具经济重要性的植物病毒之一,可侵染1000多种双子叶和单子叶植物,呈世界性分布,尤其是在温带地区。
传统的在植物中抵御病毒的方式主要针对于抑制病毒的复制、组装或干扰其致病相关蛋白的表达。当病毒在侵染宿主细胞后,病毒会表达复制相关的蛋白,该蛋白会与病毒基因组结合并起始病毒的复制,同时病毒也会表达外壳蛋白,将复制后的病毒基因组进行组装形成新生成的病毒粒子。因此,通过转入全长或部分外源复制蛋白或外壳蛋白,使植物生成针对这些病毒蛋白的小干扰rna,从而抑制病毒的复制和组装。然而,高表达量、干扰宿主细胞生长、非普适性、抗性丢失等问题使得该技术具有巨大的劣势。也有报道利用rnai技术来干扰病毒致病蛋白的表达从而达到抗病毒的目的,但是该技术也因为其同源依赖性、以及病毒具有对抗rnai抑制子的问题而不具有普适性。
近年来随着基因编辑技术的发展,利用外源人工锌指蛋白系统(artificialzincfingerproteinazp)、转录激活因子样效应物核酸酶系统(transcriptionactivator-likeeffectornucleases,talen)和成簇的规律间隔短回文重复序列系统(clusteredregularlyinterspacedshortpalindromicrepeatsequences,crispr)开发了多种在植物上的抗病毒技术。尤其是利用细菌中获得的crispr系统,靶向性的切割外源病毒的基因组dna能够产生很高的抗病毒效率。但是,利用这些基因编辑技术的抗病毒体系,都是针对dna病毒的,目前依然没有发开出有效的基于基因编辑技术的抗rna病毒体系。
随着对不同细菌来源的crispr系统的深入研究和筛选,在一种叫新凶手弗朗西丝菌(francisellanovicida)的细菌中,发现其表达的cas9核酸内切酶(fncas9)能够靶向rna,不同于目前在基因编辑领域常用的来自酿脓链球菌(streptococcuspyogenes)的只能靶向dna的核酸内切酶(spcas9)。如果能将该针对rna的crispr系统表达于植物中,并靶向侵染植物的rna病毒,将能够显著提高植物的抗病毒能力。
技术实现要素:
本发明的目的在于克服现有技术存在的以上问题,提供一种提高植物抵御rna病毒的能力的方法,本发明仅基于病毒的基因组序列,而不需要了解病毒基因的具体功能,所以该方法可以广泛地应用于抵御序列已知的各种rna病毒。
为实现上述技术目的,达到上述技术效果,本发明通过以下技术方案实现:
一种提高对rna病毒抵御能力的植物的培育方法,包括以下步骤:
s1从待抵御的rna病毒的基因组序列中选取若干靶序列,针对若干所述靶序列,分别设计合成与所述靶序列反向互补的dna片段,所述dna片段为双链dna,所述靶序列的序列排列规则为5’-nx-c-3’;
其中,nx表示x个连续的核糖核苷酸,n表示a、g、c、u中的任一种,10≤x≤30,且x为整数;
s2将步骤s1中获得的若干个所述dna片段分别构建到用于表达crispr/cas9核酸酶的载体中,得到若干个重组载体;
s3将若干个所述重组载体分别导入受体植物,从导入所述重组载体的受体植物中获得对所述rna病毒抵御能力提高的植物。
优选地,所述重组载体能转录向导rna和表达fncas9蛋白,所述向导rna为由crrna和tracrrna通过碱基配对结合而成的具有回文结构的rna;所述crrna含有由所述dna片段转录所得的rna片段。
优选地,该方法用于提高植物抵御rna病毒能力,包括以下步骤:
s11获得目的dna片段:
s101从待抵御的rna病毒的基因组序列中选取若干靶序列,针对各靶序列,分别设计合成与所述靶序列反向互补的dna片段,所述dna片段为双链dna;
所述靶序列的序列排列规则为5’-nx-c-3’,其中,nx表示x个连续的核糖核苷酸,n表示a、g、c、u中的任一种,15≤x≤25,且x为整数;
s102将步骤s101中获得的若干个所述dna片段分别构建到用于表达crispr/cas9核酸酶的载体中,得到若干个重组载体一;
所述重组载体一能转录向导rna和表达fncas9蛋白,所述向导rna为由crrna和tracrrna通过碱基配对结合而成的具有回文结构的rna,所述crrna含有由所述dna片段转录所得的rna片段;
s103将若干个所述重组载体一分别导入受体植物一中,从导入所述重组载体一的受体植物一中获得对所述rna病毒抵御能力提高的植物,记为目的植物;
在所述目的植物中,被导入的所述重组载体一上携带的所述dna片段记为目的dna片段;
s12按照步骤s102和s103的方法将所述目的dna片段导入所述用于表达crispr/cas9核酸酶的载体中,获得重组载体二,将所述重组载体二导入受体植物二,提高受体植物二对所述rna病毒的抵御能力;
所述受体植物一与所述受体植物二为同一种受体植物,或者所述受体植物一与所述受体植物二为不同种受体植物;
从导入所述重组载体一的受体植物一中获得对所述rna病毒抵御能力提高的植物的方法包括以下步骤:
s201设置对照组植物,对照组植物为向所述受体植物一中导入空载体,所述空载体为用于表达crispr/cas9核酸酶的载体;
s201向若干导入所述重组载体一的受体植物一中分别导入表达所述rna病毒的表达载体,得到植物a,向导入所述空载体的受体植物一中导入所述表达载体,得到植物b;
s203检测所述植物a和所述植物b中所述rna病毒的含量,从所述植物a中选出rna病毒含量显著低于所述植物b中rna病毒含量的个体,记为植物a’,其中显著性水平p<0.05,所述植物a’为对所述rna病毒抵御能力提高的植物;
s204在s102中,在所述受体植物二中,由所述重组载体二转录出向导rna,并表达出所述fncas9蛋白;由所述向导rna和所述fncas9蛋白形成的crispr/cas9核酸酶能够抑制所述rna病毒在所述受体植物二体内的复制,提高所述受体植物二对所述rna病毒的抵御能力。
优选地,s203中,从所述植物a中选出rna病毒含量显著低于所述植物b中rna病毒含量的个体,记为植物a’,其中显著性水平p<0.01,所述植物a’为对所述rna病毒抵御能力提高的植物。
优选地,从所述植物a中选出无任何病态表型的个体,无任何病态表型的个体为对所述rna病毒抵御能力提高的植物。
优选地,在s1和s102中,所述rna片段能够与靶标片段互补结合,所述靶标片段为在rna病毒的基因组中对应于步骤s1中所述靶序列的“nx”所示序列的双链dna。
优选地,所述用于表达crispr/cas9核酸酶的载体为pcr01载体。
优选地,所述重组载体、重组载体一、重组载体二均为重组质粒,所述重组质粒为所述pcr01载体的两个酶切位点bsai之间插入dna片段后得到,所述dna片段为与所述靶序列反向互补的dna片段。
优选地,所述rna病毒为单链rna病毒。
优选地,所述单链rna病毒为黄瓜花叶病毒。
本发明的有益效果是:
本发明的方法是一种植物培育方法,该方法能够提高植物对rna病毒的抵御能力,模拟细菌的相应免疫系统(选取来自francisellanovicida细菌中的针对rna的crispr系统),在植物中表达与病毒靶位点特异识别的sgrna,引导fncas9蛋白特异性的清除机体内的病毒rna。本发明以烟草花叶病毒(cmv)为模式病毒,首次受限了利用crispr/cas9系统有效的抑制cmv在植物中的增殖,从而提高植物的抗病能力。本发明的方法设计仅基于病毒的基因组序列,而不需要了解病毒基因的具体功能,所以该方法可以广泛地应用于抵御序列已知的各种rna病毒。
上述说明仅是本发明技术方案的概述,为了能够更清楚了解本发明的技术手段,并可依照说明书的内容予以实施,以下以本发明的较佳实施例并配合附图详细说明如后。本发明的具体实施方式由以下实施例及其附图详细给出。
附图说明
为了更清楚地说明本发明实施例技术中的技术方案,下面将对实施例技术描述中所需要使用的附图作简单地介绍,显而易见地,下面描述中的附图仅仅是本发明的一些实施例,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他的附图。
图1是本发明中pcr01载体图;
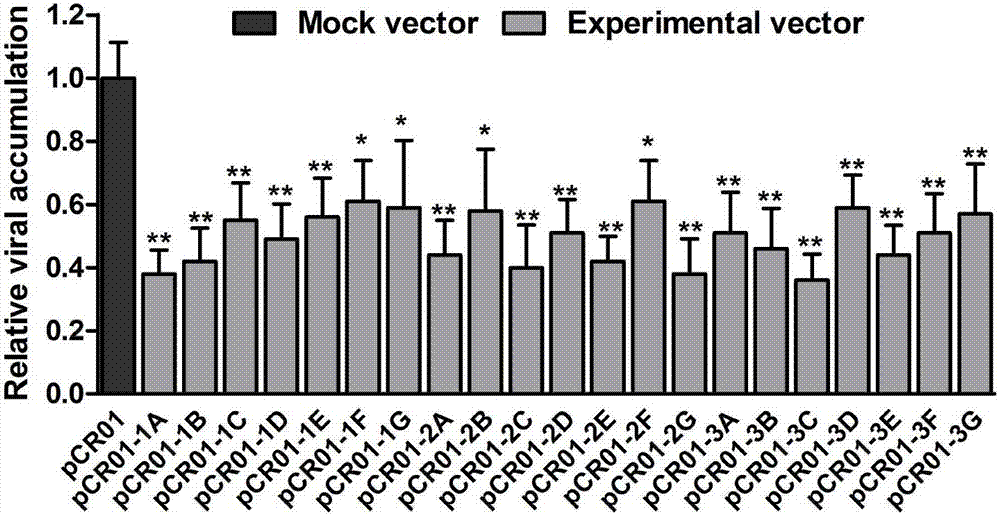
图2是本发明cmv病毒相对含量检测结果图;
图3是对照组烟草接种cmv后表型图;
图4是pcr01-1a实验组烟草接种cmv后表型图;
图5是pcr01-3c实验组烟草接种cmv后表型图。
具体实施方式
下面将结合本发明实施例中的附图,对本发明实施例中的技术方案进行清楚、完整地描述,显然,所描述的实施例仅仅是本发明一部分实施例,而不是全部的实施例。基于本发明中的实施例,本领域普通技术人员在没有作出创造性劳动前提下所获得的所有其他实施例,都属于本发明保护的范围。
下述实施例中所使用的材料、试剂等,如无特殊说明,均可以从商业途径得到。
pcr01载体,见图1,公众可从华南农业大学农学院植物病毒研究室获得。
本氏烟(nicotianabenthamiana),公众可从华南农业大学农学院植物病毒研究室获得。
黄瓜花叶病毒(cmv)毒源及侵染性克隆,公众可从华南农业大学农学院植物病毒研究室获得。
农杆菌菌株eha105:优宝生物公司产品,产品编号st1140。
实施例1
本实施例中公开了一种提高对rna病毒抵御能力的植物的培育方法,该方法包括以下步骤:
s1从待抵御的rna病毒的基因组序列中选取若干靶序列,针对若干所述靶序列,分别设计合成与所述靶序列反向互补的dna片段,所述dna片段为双链dna,所述靶序列的序列排列规则为5’-nx-c-3’;
其中,nx表示x个连续的核糖核苷酸,n表示a、g、c、u中的任一种,10≤x≤30,且x为整数;
s2将步骤s1中获得的若干个所述dna片段分别构建到用于表达crispr/cas9核酸酶的载体中,得到若干个重组载体;
s3将若干个所述重组载体分别导入受体植物,从导入所述重组载体的受体植物中获得对所述rna病毒抵御能力提高的植物。
所述重组载体能转录向导rna和表达fncas9蛋白,所述向导rna为由crrna和tracrrna通过碱基配对结合而成的具有回文结构的rna;所述crrna含有由所述dna片段转录所得的rna片段。
将上述方法用于提高植物抵御rna病毒能力,包括以下步骤:
s11获得目的dna片段:
s101从待抵御的rna病毒的基因组序列中选取若干靶序列,针对各靶序列,分别设计合成与所述靶序列反向互补的dna片段,所述dna片段为双链dna;
所述靶序列的序列排列规则为5’-nx-c-3’,其中,nx表示x个连续的核糖核苷酸,n表示a、g、c、u中的任一种,15≤x≤25,且x为整数;
s102将步骤s101中获得的若干个所述dna片段分别构建到用于表达crispr/cas9核酸酶的载体中,得到若干个重组载体一;
所述重组载体一能转录向导rna和表达fncas9蛋白,所述向导rna为由crrna和tracrrna通过碱基配对结合而成的具有回文结构的rna,所述crrna含有由所述dna片段转录所得的rna片段;
s103将若干个所述重组载体一分别导入受体植物一中,从导入所述重组载体一的受体植物一中获得对所述rna病毒抵御能力提高的植物,记为目的植物;
在所述目的植物中,被导入的所述重组载体一上携带的所述dna片段记为目的dna片段;
s12按照步骤s102和s103的方法将所述目的dna片段导入所述用于表达crispr/cas9核酸酶的载体中,获得重组载体二,将所述重组载体二导入受体植物二,提高受体植物二对所述rna病毒的抵御能力;
所述受体植物一与所述受体植物二为同一种受体植物,或者所述受体植物一与所述受体植物二为不同种受体植物;
从导入所述重组载体一的受体植物一中获得对所述rna病毒抵御能力提高的植物的方法包括以下步骤:
s201设置对照组植物,对照组植物为向所述受体植物一中导入空载体,所述空载体为用于表达crispr/cas9核酸酶的载体;
s201向若干导入所述重组载体一的受体植物一中分别导入表达所述rna病毒的表达载体,得到植物a,向导入所述空载体的受体植物一中导入所述表达载体,得到植物b;
s203检测所述植物a和所述植物b中所述rna病毒的含量,从所述植物a中选出rna病毒含量显著低于所述植物b中rna病毒含量的个体,记为植物a’,其中显著性水平p<0.05,所述植物a’为对所述rna病毒抵御能力提高的植物;
s204在s102中,在所述受体植物二中,由所述重组载体二转录出向导rna,并表达出所述fncas9蛋白;由所述向导rna和所述fncas9蛋白形成的crispr/cas9核酸酶能够抑制所述rna病毒在所述受体植物二体内的复制,提高所述受体植物二对所述rna病毒的抵御能力。
实施例2
实施例2以实施例1中的方法为基础,选用黄瓜花叶病毒(cmv)作为待抵御的rna病毒,选用拟南芥(arabidopsisthaliana)作为cmv攻击的植物体,说明建立提高植物抵御rna病毒的能力的方法。
cmv的基因组为单链rna,本实施例针对病毒rna基因组设计了多个特异性的sgrna,引导fncas9降低cmv在植物体内的积累。
一、双子叶crispr/cas9系统双元载体的构建
1、依据对黄瓜花叶病毒(cmv)的二代测序结果(序列xx-xx),在cmv基因组上选取x个靶标片段(表1)。另外构建一个非病毒序列的靶标片段作为对照。
表1靶标片段设计
其中,表1中下划线部分为pam序列。
2、根据表1设计的各靶标片段,合成带有粘性末端(小写部分)的单链引物(表2)。经过引物退火程序形成有粘性末端的双链dna(可编码与所述靶标片段互补结合的rna),正向插入到pcr01载体的两个酶切位点bsai之间,得到重组质粒。经测序表明在pcr01载体的两个酶切位点bsai之间正向插入表1中靶标片段的反向互补序列后得到的重组质粒为阳性,记为pcr01-sgrna。阳性重组质粒可转录出特异于相应靶标片段的向导rna(sgrna),并表达出fncas9蛋白。
表2对应于靶标片段的单链引物
二、烟草瞬时系统筛选活性sgrna
本实施例利用烟草建立了瞬时系统筛选对cmv具有抗性的活性sgrna。
1、烟草植株的准备
将本氏烟(nicotianabenthamiana)直接播种在营养土中,生长15天后移栽幼苗,每个直径8cm的培养杯移栽一棵小苗,生长15天后,长至8片叶片左右,选取2片大小相似位置相近的叶片待注射。温室培养条件:16h光照和8h黑暗,温度为24℃。
2、抗性载体以及病毒接种实验
液氮冻融法将pcr01-sgrna质粒转化进入农杆菌菌株eha105。接单菌落至2ml含kan和rif抗生素的yep中小摇过夜,而后接菌至10ml含kan和rif抗生素的yep中大摇过夜,5000rpm离心5min,用10mmmgcl2重悬并将之稀释至od600=0.8。加入千分之一体积的200mm乙酰丁香酮(as)并静置两小时,而后将之注射入烟草2片待注射叶片。
一天后,用同样方法将cmv的侵染性克隆注射入先前注入pcr01-sgrna的2片注射叶片。
实验设置pcr01质粒为对照组,至少选择3株烟草进行实验。
10天后,记录各组烟草植株的表型差异,并取样注射叶片。用液氮研磨至粉末,用trizol(invitrogen)提取rna。选取本氏烟(nicotianabenthamiana)中proteinphosphatase2a(pp2a)基因作为内参基因,选取一合适片段为123bp,其上下游引物为:
pp2a-f:5’-gaccctgatgttgatgttcgct-3’;
pp2a-r:5’-gagggatttgaagagagatttc-3’。
cmv病毒扩增片段大小为128bp,其上下游引物为:
cmv-f:5’-gtgaacgggttgtccatccag-3’;
cmv-r:5’-gaccgaagtccttccgaagaaac-3’。
qpcr所用试剂为vazymehiscriptiiqrtsupermixforqpcr。qpcr反应体系为:2×mix5μl;上下游引物各0.25μl;ddh2o3.5μl;rna模板1μl。
根据2-δδct法,计算各实验组烟草中cmv病毒相对于对照组的相对含量:
(1)将各组cmv含量归一化。病毒含量=2(ct(内参pp2a)-ct(cmv))
(2)实验组相对于对照组的病毒含量为病毒含量实验组/病毒含量对照组,并依据结果构建excel表格,并进行差异统计分析。
各组烟草植株中cmv病毒的相对含量结果如图2所示,由图2可以看出,与pcr01空载体对照组相比,各实验组中的cmv病毒的相对含量均较低。经过差异统计分析,各实验组除了1f、1g、2b、2f为显著差异(p<0.05)之外,其余各实验室都与空载体对照组具有极显著差异(p<0.01)。
pcr01空载体对照组中的烟草植株呈现明显的cmv病态,而差异最为明显的1a、3c实验组表型正常,表现出极强的抵御病毒的表型。并且发现各组中cmv病毒的相对含量与烟草植株的表型病态程度趋势一致(具有代表性的烟草植株表型结果见图3-5)。
该方法能够提高植物对rna病毒的抵御能力,模拟细菌的相应免疫系统(选取来自francisellanovicida细菌中的针对rna的crispr系统),在植物中表达与病毒靶位点特异识别的sgrna,引导fncas9蛋白特异性的清除机体内的病毒rna。以烟草花叶病毒(cmv)为模式病毒,首次受限了利用crispr/cas9系统有效的抑制cmv在植物中的增殖,从而提高植物的抗病能力。该方法设计仅基于病毒的基因组序列,而不需要了解病毒基因的具体功能,所以该方法可以广泛地应用于抵御序列已知的各种rna病毒。
对所公开的实施例的上述说明,使本领域专业技术人员能够实现或使用本发明。对这些实施例的多种修改对本领域的专业技术人员来说将是显而易见的,本文中所定义的一般原理可以在不脱离本发明的精神或范围的情况下,在其它实施例中实现。因此,本发明将不会被限制于本文所示的这些实施例,而是要符合与本文所公开的原理和新颖特点相一致的最宽的范围。
- 还没有人留言评论。精彩留言会获得点赞!