经过修饰的牛促生长素多肽和其用途的制作方法
本申请是国际申请日为2010年12月21日的国际申请pct/us2010/061669进入中国、申请号为201080058893.5的题为“经过修饰的牛促生长素多肽和其用途”的发明专利申请的分案申请。
本发明涉及任选用至少一个非天然编码的氨基酸修饰的牛促生长素(bst)多肽。
背景技术:
一些生物活性多肽只通过肠胃外投予极少剂量就能实现长期的活性,而一些生物活性多肽则需要达到足够血清浓度和/或在血清中的半衰期很短以致必须投予相当大剂量才能在较长时间(例如一周或更长时间)内提供所需的生物作用。促生长素(生长激素)是此类多肽的一个实例。
为了防止某些多肽不合需要地迅速释放到动物的血流中,已经利用可任选含有水合作用阻滞剂(抗水合剂)的液体媒剂或结合进一步降低多肽在体液中的溶解性的金属或金属化合物来肠胃外投予这些多肽。为避免对无法接受的大量此类媒剂的需求,且出于包括优良的长期释放性能在内的其它原因,宜采用含相当高浓度所述多肽的媒剂,例如,如颁予詹姆斯c.米奇尔(jamesc.mitchell)的美国专利第5,739,108号、让渡于艾利李莉(elililly)的美国专利第4,977,140号、让波于卢克有限公司(lucky,ltd.)的美国专利第5,520,927号和让渡于lg化学品有限公司(lgchemicalsltd.)的美国专利第5,744,163号中所示。然而,仍需要改进使这些多肽以生物活性形式释放到动物的血流中的效率(“生物利用率”)和/或在一些效用中其提供所需的动物生理反应的有效性(“功效”)。这些因素每一者都可实质上影响实现所需生物作用而必须投予的多肽量,并由此实质上影响每次投药的成本。多肽(例如促生长素)可在使用重组dna转化的原核生物体中产生,但仍然需要能提供改进的促生长素多肽(包括具有较长血清半衰期的促生长素多肽)的蛋白质调配物。用于投予生物活性组合物的各种方法和装置先前已经揭示于公开案中,且这些示范性公开案中的一些包括:克里斯特森(christensen)等人,wo97/03692,揭示一种生长激素与锌以及任选使用的赖氨酸或钙离子的调配物。以此方式配制的生长激素对脱酰胺作用显示出抗性。
董(dong)等人,wo00/13674,揭示一种定时释放药物的机构。此机构包含一种装载胶囊的具有半透性壁的容器,所述胶囊包含药物调配物、柱塞(piston)和渗透性组合物。这种给药机构以受控制的速率在长达24小时时间内通过通路释放药物调配物。艾克雷博(ekwuribe),美国专利第5,359,030号、第5,438,040号和第5,681,811号,揭示一种稳定的结合肽复合物,其包含以结合方式与包括亲脂性部分和亲水性部分的聚合物偶合的肽,适于肠胃外和非肠胃外投药。福格森(ferguson)等人,美国专利第4,977,140号,揭示一种持续释放调配物,其包含在载剂中的牛促生长素,所述载剂包含蜡(以重量计约1%到20%)和油(以重量计约80%到99%)。当注射于乳牛中时,此调配物使产奶量增加,持续28天。鲍曼de(baumande)等人在其论文中揭示在泌乳时使用外源bst,“外源牛促生长素对泌乳的影响(effectsofexogenousbovinesomatotropinonlactation)”(鲍曼等人;营养学年评(annurevnutr.)1993;13:437-61)。汉密尔顿(hamilton)等人,美国专利第4,816,568号,揭示动物生长激素与稳定剂的组合物。所述稳定剂可溶于水溶液中,并且一般极性极高。教示的这些稳定剂包括多元醇、氨基酸、在生理ph下具有带电侧基的氨基酸聚合物以及胆碱衍生物。此组合物的水性调配物可通过以下方式形成:(i)将稳定剂分散于水溶液中,和(ii)随后添加生长激素。固体调配物可通过以下方式形成:(i)混合稳定剂与生长激素,(ii)任选添加佐剂、粘合剂等到组合物中,和(iii)压缩组合物,形成片剂或丸粒。
金(kim)等人,美国专利第5,520,927号,揭示一种肠胃外投予的缓慢释放生物活性药物组合物,其包含促生长素、至少一种生育酚化合物和延迟释放剂。金等人,美国专利第5,744,163号,揭示一种供持续释放动物生长激素的调配物。此调配物包含用生物可降解聚合物和泊洛沙姆(poloxamer)构成的膜包覆含促生长素的丸粒。玛古德(magruder)等人,美国专利第5,034,229号,揭示一种用于向动物递送例如生长激素等有益试剂的装置。此装置还可递送多元醇作为一种粘度调节方式。马汀(martin),ep0216485,揭示一种制备生长激素与过渡金属的络合物的方法。还描述通过用过渡金属与生长激素的络合物处理动物来促进动物生长的方法。米奇尔(mitchell),美国专利第5,739,108号,揭示生物活性多肽的延长释放调配物,其包含以重量计约10%到以重量计约50%多肽于生物相容性油中的分散液。此多肽可与无毒金属或金属盐缔合。所述调配物还可包含抗水合剂,例如单硬脂酸铝。皮卡(pikal)等人,美国专利第5,612,315号,揭示用于肠胃外投予人生长激素的调配物,其包含人生长激素、甘氨酸和甘露糖醇。揭示的调配物被描述为提供针对蛋白质聚集的稳定作用。雷曼(raman)等人,美国专利第5,356,635号,揭示一种持续释放组合物,其包含生物活性剂,例如促生长素;生物可降解的非晶形碳水化合物玻璃基质,其内例如分散有促生长素;和疏水性物质。所述非晶形碳水化合物玻璃基质包含非晶形碳水化合物和再结晶延迟剂,并占组合物重量的约60%到组合物重量的90%。组合物在降到至少约18℃时为固体。
雷曼等人,wo93/13792,揭示一种植入式装置,其包含过渡金属-促生长素络合物与过渡金属-增溶物质的组合。过渡金属可为锌、锰或铜。金属-增溶物质可为氨基酸。可使用蔗糖来使促生长素稳定。此装置可包含硅树脂管或蜡。西里(seely)等人,wo93/19773,揭示包含以下各物的水溶液:(i)冻干的促生长素组合物,其包含促生长素和盐酸精氨酸;和(ii)包含edta的稀释剂、非离子型表面活性剂,和任选使用的缓冲液或非缓冲试剂,例如蔗糖或海藻糖。希瓦马克两南(sivaramakrishnan)等人,美国专利第5,219,572号,揭示一种用于控制释放例如促生长素等大分子蛋白质的装置。此装置包含完全包围含不均匀微型胶囊的内室的水溶性外壳。这些微型胶囊包含蜡壳,其包围核心基质。核心基质包含例如促生长素和任选使用的赋形剂、稳定剂、粘合剂等,例如硬脂酸镁或蔗糖。当外壳在动物体内的流体环境中溶解时,微型胶囊暴露于流体环境,并在暴露后各种时间时破裂。
斯特博(steber)等人,ep0523330a1,揭示一种压实、带刻痕、部分包覆的植入式组合物,其包含生物活性多肽(例如促生长素);脂肪、蜡或其混合物;和糖(例如单糖、二糖或三糖)。斯托斯(storrs)等人,美国专利第5,986,073号,揭示一种用于纯化和回收生物活性促生长素单体的方法。这项工作是基于发现促生长素单体与促生长素寡聚物尽管具有重叠的等电点,但仍可通过在极窄ph范围内进行选择性沉淀来分离。不合需要的杂质可借助此方法去除,并且回收的经过纯化的促生长素单体不经进一步纯化即适于肠胃外施用到目标动物。泰勒(tyle),美国专利第4,857,506号,揭示一种用于持续释放生长激素的水包油包水型多重乳液。生长激素分散于内部水相中;内部水相分散于不与水混溶的液相或油相中;且不与水混溶相分散于外部水相中。内部水相可包括以重量计高达40%的多元醇、二醇或糖。维斯瓦纳安(viswanathan)等人,美国专利第4,917,685号,揭示一种用于稳定动物生长激素的递送装置。此装置包含包围并界定储槽的器壁。至少一部分器壁为多孔的,以允许生长激素和稳定剂通过。生长激素与稳定剂的调配物实质上如上述汉密尔顿等人所揭示。
尽管以上概述的公开案中描述了众多尝试,但此项技术仍存在巨大改进空间。本发明通过提供改进的牛促生长素(bst)多肽满足此需求。本发明特别针对与bst多肽的活性和制备有关的问题,并且还针对具有改进的生物或药理学特性和/或改进的治疗半衰期的bst多肽的制备。
技术实现要素:
本发明提供包含一个或一个以上非天然编码氨基酸的bst多肽。
在一些实施例中,bst多肽包含一种或一种以上翻译后修饰。在一些实施例中,bst多肽与连接子、聚合物或生物活性分子连接。在一些实施例中,bst多肽与双官能聚合物、双官能连接子或至少另一种bst多肽连接。
在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。在一些实施例中,水溶性聚合物包含聚(乙二醇)部分。在一些实施例中,非天然编码氨基酸通过连接子与水溶性聚合物连接或与水溶性聚合物键接。在一些实施例中,聚(乙二醇)分子是一种双官能聚合物。在一些实施例中,双官能聚合物与第二多肽连接。在一些实施例中,第二多肽是bst多肽。
在一些实施例中,bst多肽包含至少两个连接到包含聚(乙二醇)部分的水溶性聚合物的氨基酸。在一些实施例中,至少一个氨基酸是非天然编码氨基酸。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9,10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191(即,具有190个氨基酸的牛生长激素氨基酸蛋白的羧基末端)。
在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:29、30、33、34、35、37、39、40、49、57、59、66、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、122、126、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、159、183、186和187(seqidno:1)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:29、30、33、34、35、37、39、40、49、57、59、66、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、122、126、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、159、183、186和187(seqidno:2)。在一些实施例中,一个或一个以上非天然编码氨基酸在具有190个氨基酸的牛生长激素蛋白的以下一个或一个以上位置进行取代:29、30、33、34、35、37、39、40、49、57、59、66、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、122、126、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、159、183、186和187。
在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:29、33、35、37、39、49、57、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、186和187(seqidno:1)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:29、33、35、37、39、49、57、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、186和187(seqidno:2)。在一些实施例中,一个或一个以上非天然编码氨基酸在具有190个氨基酸的牛生长激素蛋白的以下一个或一个以上位置进行取代:29、33、35、37、39、49、57、69、70、71、74、88、91、92、94、95、98、99、101、103、107、108、111、129、130、131、133、134、135、136、137、139、140、141、142、143、145、147、154、155、156、186和187。
在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:35、88、91、92、94、95、99、101、103、111、131、133、134、135、136、139、140、143、145和155(seqidno:1)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:35、88、91、92、94、95、99、101、103、111、131、133、134、135、136、139、140、143、145和155(seqidno:2)。在一些实施例中,一个或一个以上非天然编码氨基酸在具有190个氨基酸的牛生长激素蛋白的以下一个或一个以上位置进行取代:35、88、91、92、94、95、99、101、103、111、131、133、134、135、136、139、140、143、145和155。
在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:30、74、103(seqidno:1)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:35、92、143、145(seqidno:1)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:30、74、103(seqidno:2)。在一些实施例中,一个或一个以上非天然编码氨基酸在以下一个或一个以上位置进行取代:35、92、143、145(seqidno:2)。在一些实施例中,一个或一个以上非天然编码氨基酸在具有190个氨基酸的牛生长激素蛋白的以下一个或一个以上位置进行取代:30、74、103。在一些实施例中,一个或一个以上非天然编码氨基酸在具有190个氨基酸的牛生长激素蛋白的以下一个或一个以上位置进行取代:35、92、143、145。
在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst(例如bst)的亲和力相比较时,所述取代、添加或缺失将调节bst对bst(例如bgh多肽)受体的亲和力。在一些实施例中,bst(例如bst多肽)包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的稳定性相比较时,所述取代、添加或缺失将增加bst多肽的稳定性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的免疫原性相比较时,所述取代、添加或缺失将调节bst多肽的免疫原性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的血清半衰期或循环时间相比较时,所述取代、添加或缺失将调节bst多肽的血清半衰期或循环时间。在一些实施例中,本发明包含bgh多肽,其包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的亲和力相比较时,所述取代、添加或缺失将调节bgh对bgh受体的亲和力。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的稳定性相比较时,所述取代、添加或缺失将增加bgh多肽的稳定性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的免疫原性相比较时,所述取代、添加或缺失将调节bgh多肽的免疫原性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的血清半衰期或循环时间相比较时,所述取代、添加或缺失将调节bgh多肽的血清半衰期或循环时间。
共价连接亲水性聚合物聚(乙二醇)(缩写为peg)是一种增加许多生物活性分子(包括蛋白质、肽,尤其是疏水性分子)的水溶性、生物利用率;增加血清半衰期;增加治疗半衰期;调节免疫原性;调节生物活性;或延长循环时间的方法。peg已被广泛用于医药品或人工移植物以及生物相容性、无毒和无免疫原性极为重要的其它应用中。为了使peg的所需特性最大化,与生物活性分子连接的peg聚合物的总分子量和水合状态必须足够高,以便赋予通常与peg聚合物连接相关的有利特征,例如增加的水溶性和循环半衰期,同时不会不利地影响母体分子的生物活性。
peg衍生物常常是通过反应性化学官能团(例如赖氨酸、半胱氨酸和组氨酸残基、n末端和碳水化合物部分)与生物活性分子连接。蛋白质和其它分子通常具有少数反应性位点可供聚合物连接。通常,最适于经由聚合物连接而修饰的位点在受体结合中起到重要作用,并且是保留分子的生物活性所必需的。因此,不加选择地将聚合物链与生物活性分子上的这些反应性位点连接,通常会使经聚合物修饰的分子的生物活性显著降低或甚至完全丧失。r.克拉克(r.clark)等人,(1996),生物化学杂志(j.biol.chem.),271:21969-21977。为了形成赋予目标分子所需优势的具有足够聚合物分子量的结合物,现有技术方法通常涉及将多个聚合物臂与分子随机连接,由此增加了母体分子生物活性降低或甚至完全丧失的风险。
形成用于连接peg衍生物与蛋白质的基因座的反应性位点由蛋白质的结构指示。蛋白质(包括酶)是由各种α-氨基酸序列构成,这些序列的一般结构为h2n--chr--cooh。一个氨基酸的α氨基部分(h2n--)与相邻氨基酸的羧基部分(--cooh)接合形成酰胺键,其可表示为--(nh--chr--co)n--,其中下标“n”可等于数百或数千。由r表示的片段可含有提供蛋白质生物活性和连接peg衍生物的反应性位点。
举例来说,在氨基酸赖氨酸的情况下,在ε位以及α位中存在--nh2部分。ε--nh2在碱性ph条件下自由反应。用peg使蛋白质衍生化领域中的很多技术都是针对开发用于连接蛋白质中存在的赖氨酸残基的ε--nh2部分的peg衍生物。“用于先进聚乙二醇化技术的聚乙二醇和衍生物(polyethyleneglycolandderivativesforadvancedpegylation)”,奈克塔分子工程目录(nektarmolecularengineeringcatalog),2003,第1-17页。然而,这些peg衍生物都具有共同的局限性,即,无法将其选择性地安装到存在于蛋白质表面上的常见的多个赖氨酸残基上。在赖氨酸残基对于蛋白质活性极为重要(例如存在于酶活性位点中)的情况下,或在赖氨酸残基对于介导蛋白质与其它生物分子的相互作用起作用的情况下,如在受体结合位点的情况下,这可成为显著的局限性。
现有蛋白质聚乙二醇化方法的另一同等重要的限制因素在于,peg衍生物可与除所需残基外的残基发生不合需要的副反应。组氨酸含有结构以--n(h)--表示的反应性亚氨基部分,而许多与ε--nh2反应的化学反应性物质也可与--n(h)--反应。类似地,氨基酸半胱氨酸的侧链具有游离硫氢基,结构以-sh表示。在一些情况下,针对赖氨酸ε--nh2基团的peg衍生物也与半胱氨酸、组氨酸或其它残基反应。这样可产生经peg衍生化的生物活性分子的复杂不均匀混合物,并且有破坏目标生物活性分子活性的风险。需要开发出如下peg衍生物,其允许在蛋白质内的单一位点引入化学官能团,随后使一种或一种以上peg聚合物能够在蛋白质表面上明确界定且可预测的特定位点与生物活性分子选择性偶合。
除赖氨酸残基外,此项技术中还曾就开发靶向其它氨基酸侧链(包括半胱氨酸、组氨酸和n末端)的活性peg试剂进行了大量尝试。例如参看美国专利第6,610,281号,以引用的方式并入本文中;和“用于先进聚乙二醇化技术的聚乙二醇和衍生物”,奈克塔分子工程目录,2003,第1-17页。可以使用定点诱变和此项技术中已知的其它技术,将半胱氨酸残基位点选择性地引入蛋白质结构中,并且由此得到的游离硫氢基部分可与具有硫醇反应性官能团的peg衍生物反应。但这种方法比较麻烦,因为引入游离硫氢基会使所得蛋白质的表达、折叠和稳定性变得复杂。因此,本发明提供将化学官能团引入bst和bgh中的合意方式,其能使一种或一种以上peg聚合物与蛋白质选择性偶合,同时还与硫氢基以及蛋白质中常见的其它化学官能团相容(即,不会与之发生不合需要的副反应)。
从此项技术的抽样(sampling)中可以了解到,所开发的用于连接蛋白质侧链,尤其赖氨酸氨基酸侧链上的--nh2部分和半胱氨酸侧链上的-sh部分的许多此类衍生物都证实在合成和使用方面存在问题。一些衍生物与蛋白质形成不稳定键,这些键将经历水解,并因此分解、降解,或者以其它方式在水性环境(例如血流)中不稳定。一些衍生物形成较为稳定的键,但会在形成键之前经历水解,这意味着peg衍生物上的反应性基团可能会在连接蛋白质之前就失活。一些衍生物略带毒性,因此不太适于体内使用。一些衍生物反应过慢而无法实际使用。一些衍生物会因连接到负责蛋白质活性的位点而导致蛋白质活性丧失。一些衍生物对其将连接的位点不具特异性,这也会导致丧失所需活性以及缺乏结果的可再现性。为了克服用聚(乙二醇)部分修饰蛋白质所带来的问题,已经开发出更为稳定(例如,美国专利6,602,498,以引用的方式并入)或与分子和表面上的硫醇部分选择性反应(例如,美国专利6,610,281,以引用的方式并入)的peg衍生物。此项技术中无疑需要在生理环境中具化学惰性,而只有在需要的情况下才选择性反应形成稳定化学键的peg衍生物。
已经揭示了羟烷基淀粉,尤其是羟乙基淀粉(hydroxyethylstarch,hes)共价连接到多肽的结合物的使用,以便潜在地改变多肽的免疫原性和/或变应原性。羟乙基淀粉化是一项替代性技术,其揭示于让渡于费森尤斯卡比公司(freseniuskabiab)的一系列专利申请案中,包括美国专利公开案第20050063943号、第20060121073号、第20010100163号、第20050234230号、第20050238723号、第20060019877号、第20070134197号、第20070087961号,以及美国专利第7,285,661号,所有这些专利都以引用的方式并入本文中。hes是一种经过修饰的天然聚合物,在临床上用作血浆增量剂,而羟乙基淀粉化表示将原料药与hes衍生物偶合以改变药物特征(例如药物动力学或水溶性)的技术。这项技术还包括经由增加分子稳定性和降低肾清除率来延长蛋白质血浆循环,由此引起生物活性的增加。此外,也可能降低免疫原性或变应原性。通过改变不同的参数,例如hes的分子量,可以定制多种hes结合物。尽管如此,羟乙基淀粉仍与所有其它当前可用的聚合物共有共同的缺点:其多分散性。聚合物结合物是分子量分布在平均值周围的分子的混合物。此均质性的缺乏会引起较低水平的化学和生物化学特性表征,并且可阻止医药活性组分到达其作用位点(受体、酶等)。在这些情况下,活性药物需要以其原始的非结合形式递送,由此需要借助代谢反应使聚合物裂解以达到其医药功效。
本发明的蛋白质技术克服了与蛋白质其它位点特异性修饰有关的许多缺点。具体点说,已将新的组分添加到原核生物大肠杆菌(escherichiacoli,e.coli)(例如l.王(l.wang)等人,(2001),科学(science)292:498-500)和真核生物酿酒酵母菌(sacchromycescerevisia,s.cerevisiae)(例如j.秦(j.chin)等人,科学301:964-7(2003))的蛋白质生物合成机器中,使得所述机器能够在体内将非基因编码氨基酸并入蛋白质中。已使用这种方法响应琥珀密码子(tag)以高保真度将多种具有新颖化学、物理或生物特性的新氨基酸有效地并入大肠杆菌和酵母的蛋白质中,所述新氨基酸包括光亲和性标记和可光异构化氨基酸、光交联氨基酸(例如参看秦j.w.(chin,j.w.)等人(2002)美国国家科学院院刊(proc.natl.acad.sci.u.s.a.)99:11020-11024;和秦j.w.等人,(2002)美国化学协会杂志(j.am.chem.soc.)124:9026-9027)、酮基氨基酸、含有重原子的氨基酸和糖基化氨基酸。例如参看j.w.秦等人,(2002),美国化学协会杂志(journaloftheamericanchemicalsociety)124:9026-9027;j.w.秦和p.g.查鲁兹(p.g.schultz),(2002),化学与生物化学(chembiochem)3(11):1135-1137;j.w.秦等人,(2002),美国国家科学院院刊(pnasunitedstatesofamerica)99:11020-11024;以及l.王(l.wang)和p.g.查鲁兹,(2002),化学通讯(chem.comm.),1:1-11。所有参考文献都以全文引用的方式并入本文中。这些研究已证实,有可能选择性地以常规方式引入化学官能团,例如酮基、炔基和叠氮部分,这些官能团不存在于蛋白质中,对20种常见的基因编码氨基酸中所见的所有官能团具化学惰性,并且可用于有效且选择性地反应形成稳定共价键。
将非基因编码的氨基酸并入蛋白质中的能力允许引入可提供天然存在官能团(例如赖氨酸的ε-nh2、半胱氨酸的硫氢基-sh、组氨酸的亚氨基等)的有价值替代物的化学官能团。已知某些化学官能团对20种常见的基因编码氨基酸中所见的官能团呈惰性,但能干净且有效地反应形成稳定键。举例来说,此项技术中已知,叠氮基和乙炔基在催化量的铜存在下,可在水性条件中进行胡伊斯根(huisgen)[3+2]环加成反应。例如参见托尼诺(tornoe)等人,(2002)有机化学杂志(j.org.chem.)67:3057-3064;和罗斯托瑟夫(rostovtsev)等人,(2002)德国应用化学(angew.chem.int.ed.)41:2596-2599。举例来说,通过将叠氮部分引入蛋白质结构中,就能够并入对蛋白质中所见的胺、硫氢基、羧酸、羟基具化学惰性,但也可与乙炔部分平稳且有效地反应形成环加成产物的官能团。重要的是,在不存在乙炔部分的情况下,叠氮基仍保留化学惰性,而且在其它蛋白质侧链存在时在生理条件下不发生反应。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:seqidno:1中的3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:seqidno:2中的3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的以下一个或一个以上位置:3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:3、7、33、43、58、62、67、69、99、123、124、133、134、141、166,以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:3、7、33、43、58、62、67、69、99、123、124、133、134、141、166,以及其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的以下一个或一个以上位置:3、7、33、43、58、62、67、69、99、123、124、133、134、141,166和其任何组合。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:seqidno:1中的3、7、62、133、166,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:seqidno:2中的3、7、62、133、166,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的以下一个或一个以上位置:3、7、62、133、166,以及其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的62位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的62位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的62位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的133位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的133位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的133位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的92位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的92位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的92位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的35位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的35位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的35位。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的40位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的40位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的40位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的95位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的95位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的95位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的96位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的96位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的96位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的98位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的98位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的98位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的99位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的99位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的99位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的103位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的103位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的103位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的105位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的105位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的105位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的137位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的137位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的137位。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154,或其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154,或其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的以下一个或一个以上位置:tyr35、gln91、phe92、ser94、arg95、asn99、leu101、arg133、ala134、leu138、lys139、gln140、tyr142、lys144、leu149、arg150、ala154,或其任何组合。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的138位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的138位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的138位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的142位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的142位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的142位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的143位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的143位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的143位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的144位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的144位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的144位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的146位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的146位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的146位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的148位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的148位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的148位。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的153位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的153位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的153位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的154位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的154位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的154位。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的158位(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的158位(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh的158位。在一些实施例中,本发明的多肽包含一个或一个以上天然氨基酸取代、添加或缺失。在一些实施例中,将一个或一个以上非天然氨基酸并入在seqidno:1、2或者其它bst或bgh序列的n或c末端的前导序列或信号序列中。
在一些实施例中,将一个或一个以上所述位置的非天然存在氨基酸与水溶性聚合物连接,这些位置包括(但不限于)以下位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上所述位置的非天然存在氨基酸与水溶性聚合物连接,所述位置包括(但不限于)以下位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上所述位置的非天然存在氨基酸与水溶性聚合物连接,所述位置包括(但不限于)bgh蛋白的以下位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、61、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191(即,蛋白质的羧基末端)和其任何组合。
在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)以下位置:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154,或其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)以下位置:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154,或其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)bgh的以下位置:tyr35、gln91、phe92、ser94、arg95、asn99、leu101、arg133、ala134、leu138、lys139、gln140、tyr142、lys144、leu149、arg150、ala154,或其任何组合。
在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)以下位置:seqidno:1中的3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)以下位置:seqidno:2中的3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,将一个或一个以上所述位置的一个或一个以上非天然编码氨基酸与水溶性聚合物连接,这些位置包括(但不限于)bgh的以下位置:3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173,以及其任何组合。在一些实施例中,非天然存在氨基酸是在bgh、seqidno:1、2或者其它bst序列的n或c末端的信号序列或前导序列中,并与水溶性聚合物连接。
在一些实施例中,bst多肽包含取代、添加或缺失,其调节bst多肽对受体或结合搭配物(包括(但不限于)蛋白质、多肽、小分子或核酸)的亲和力。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的稳定性相比较时,所述取代、添加或缺失增加bst多肽的稳定性。稳定性和/或溶解性可以使用所属领域技术人员已知的多种不同分析来测量。所述分析包括(但不限于)se-hplc和rp-hplc。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的免疫原性相比较时,所述取代、添加或缺失将调节bst多肽的免疫原性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的血清半衰期或循环时间相比较时,所述取代、添加或缺失将调节bst多肽的血清半衰期或循环时间。
在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的水溶性相比较时,所述取代、添加或缺失将增加bst多肽的水溶性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的溶解性相比较时,所述取代、添加或缺失将增加宿主细胞中产生的bst多肽的溶解性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst在宿主细胞中的表达或在体外的合成相比较时,所述取代、添加或缺失将增加bst多肽的表达或增加其合成。包含此取代的bst多肽保留了激动剂活性,并且保持或改进在宿主细胞中的表达水平。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst的蛋白酶抗性相比较时,所述取代、添加或缺失将增加bst多肽的蛋白酶抗性。在一些实施例中,bst多肽包含取代、添加或缺失,当同受体与无取代、添加或缺失的相应bst多肽相互作用时的信号转导活性相比较时,所述取代、添加或缺失将调节受体的信号转导活性。在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst多肽与另一分子(例如受体)的结合相比较时,所述取代、添加或缺失将调节bst多肽的结合。在一些实施例中,bst多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bst多肽的造血作用相比较,所述取代、添加或缺失将调节造血作用。在一些实施例中,bst多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bst多肽的中性粒细胞增殖相比较,所述取代、添加或缺失将调节中性粒细胞的增殖。在一些实施例中,bst多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bst多肽的中性粒细胞成熟相比较,所述取代、添加或缺失将调节中性粒细胞的成熟。
在一些实施例中,bst多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bst与医药用防腐剂(例如间甲酚、苯酚、苯甲醇)的相容性相比较时,所述取代、添加或缺失将增加bst多肽的相容性。此增加的相容性将使保存的医药调配物制剂能够在储存期间保持蛋白质的生理化学特性和生物活性。
在一些实施例中,利用一个或一个以上非天然氨基酸产生一个或一个以上经工程改造的键。分子内键可以多种方式产生,包括(但不限于)在适合条件下,蛋白质中的两个氨基酸之间发生反应(一个或两个氨基酸可以为非天然氨基酸);在适合条件下,两个氨基酸与连接子、聚合物或其它分子发生反应,这两个氨基酸各自可为天然编码或非天然编码的,等等。
在一些实施例中,bgh多肽包含取代、添加或缺失,其调节bgh多肽对受体或结合搭配物(包括(但不限于)蛋白质、多肽、小分子或核酸)的亲和力。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的稳定性相比较时,所述取代、添加或缺失将增加bgh多肽的稳定性。稳定性和/或溶解性可以使用所属领域技术人员已知的多种不同分析来测量。所述分析包括(但不限于)se-hplc和rp-hplc。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的免疫原性相比较时,所述取代、添加或缺失将调节bgh多肽的免疫原性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的血清半衰期或循环时间相比较时,所述取代、添加或缺失将调节bgh多肽的血清半衰期或循环时间。
在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的水溶性相比较时,所述取代、添加或缺失将增加bgh多肽的水溶性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的溶解性相比较时,所述取代、添加或缺失将增加宿主细胞中产生的bgh多肽的溶解性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh在宿主细胞中的表达或在体外的合成相比较时,所述取代、添加或缺失将增加bgh多肽的表达或增加其合成。包含此取代的bgh多肽保留了激动剂活性,并且保持或改进在宿主细胞中的表达水平。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh的蛋白酶抗性相比较时,所述取代、添加或缺失将增加bgh多肽的蛋白酶抗性。在一些实施例中,bgh多肽包含取代、添加或缺失,当同受体与无取代、添加或缺失的相应bgh多肽相互作用时的信号转导活性相比较时,所述取代、添加或缺失将调节受体的信号转导活性。在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh多肽与另一分子(例如受体)的结合相比较时,所述取代、添加或缺失将调节bgh多肽的结合。在一些实施例中,bgh多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bgh多肽的造血作用相比较,所述取代、添加或缺失将调节造血作用。在一些实施例中,bgh多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bgh多肽的中性粒细胞增殖相比较,所述取代、添加或缺失将调节中性粒细胞的增殖。在一些实施例中,bgh多肽包含取代、添加或缺失,与无取代、添加或缺失的相应bgh多肽的中性粒细胞成熟相比较,所述取代、添加或缺失将调节中性粒细胞的成熟。
在一些实施例中,bgh多肽包含取代、添加或缺失,当与无取代、添加或缺失的相应bgh与医药用防腐剂(例如间甲酚、苯酚、苯甲醇)的相容性相比较时,所述取代、添加或缺失将增加bgh多肽的相容性。此增加的相容性将使保存的医药调配物制剂能够在储存期间保持蛋白质的生理化学特性和生物活性。
在一些实施例中,bst多肽中的一个或一个以上氨基酸可以用一个或一个以上天然存在或非天然存在氨基酸取代。在一些实施例中,bst多肽中的氨基酸可以用天然存在或非天然存在氨基酸取代,只要至少一个取代是用非天然编码氨基酸进行即可。在一些实施例中,bst多肽中的一个或一个以上氨基酸可以用一个或一个以上天然存在氨基酸取代,并且另外至少一个取代是用非天然编码氨基酸进行。
在一些实施例中,bgh多肽中的一个或一个以上氨基酸可以用一个或一个以上天然存在或非天然存在氨基酸取代。在一些实施例中,bgh多肽中的氨基酸可以用天然存在或非天然存在氨基酸取代,只要至少一个取代是用非天然编码氨基酸进行即可。在一些实施例中,bgh多肽中的一个或一个以上氨基酸可以用一个或一个以上天然存在氨基酸取代,并且另外至少一个取代是用非天然编码氨基酸进行。
在一些实施例中,非天然编码氨基酸包含羰基、乙酰基、氨氧基、肼基、酰肼基、氨基脲基、叠氮基或炔基。
在一些实施例中,非天然编码氨基酸包含羰基。在一些实施例中,非天然编码氨基酸具有以下结构:
其中n为0-10;r1为烷基、芳基、经取代烷基或经取代芳基;r2为h、烷基、芳基、经取代烷基和经取代芳基;且r3为h、氨基酸、多肽,或氨基末端修饰基团;且r4为h、氨基酸、多肽,或羧基末端修饰基团。
在一些实施例中,非天然编码氨基酸包含氨氧基。在一些实施例中,非天然编码氨基酸包含酰肼基。在一些实施例中,非天然编码氨基酸包含肼基。在一些实施例中,非天然编码氨基酸残基包含氨基脲基。
在一些实施例中,非天然编码氨基酸残基包含叠氮基。在一些实施例中,非天然编码氨基酸具有以下结构:
其中n为0-10;r1为烷基、芳基、经取代烷基、经取代芳基或不存在;x为o、n、s或不存在;m为0-10;r2为h、氨基酸、多肽或氨基末端修饰基团;且r3为h、氨基酸、多肽或羧基末端修饰基团。
在一些实施例中,非天然编码氨基酸包含炔基。在一些实施例中,非天然编码氨基酸具有以下结构:
其中n为0-10;r1为烷基、芳基、经取代烷基或经取代芳基;x为o、n、s或不存在;m为0-10;r2为h、氨基酸、多肽或氨基末端修饰基团;且r3为h、氨基酸、多肽或羧基末端修饰基团。
在一些实施例中,所述多肽为bst多肽激动剂、部分激动剂、拮抗剂、部分拮抗剂或反向激动剂。在一些实施例中,bst多肽激动剂、部分激动剂、拮抗剂、部分拮抗剂或反向激动剂包含与水溶性聚合物连接的非天然编码氨基酸。在一些实施例中,水溶性聚合物包含聚(乙二醇)部分。在一些实施例中,bst多肽激动剂、部分激动剂、拮抗剂、部分拮抗剂或反向激动剂包含非天然编码氨基酸,和一个或一个以上翻译后修饰、连接子、聚合物或生物活性分子。
本发明还提供分离的核酸,其包含在严格条件下与编码seqidno:1、2的多肽的核酸杂交的聚核苷酸。本发明也提供分离的核酸,其包含在严格条件下与编码seqidno:1、2所示多肽的聚核苷酸杂交的聚核苷酸,其中所述聚核苷酸包含至少一个选择密码子。本发明也提供分离的核酸,其包含编码seqidno.:1、2所示多肽的聚核苷酸。本发明也提供分离的核酸,其包含编码seqidno.:1、2所示具有一个或一个以上非天然编码氨基酸的多肽的聚核苷酸。所属领域技术人员易于了解,多种不同的聚核苷酸可编码本发明的任何多肽。
在一些实施例中,选择密码子选自由以下密码子组成的群组:琥珀密码子、赭石密码子、蛋白石密码子、独特密码子、稀有密码子、五碱基密码子和四碱基密码子。
本发明也提供制备与水溶性聚合物连接的bst多肽的方法。在一些实施例中,所述方法包含使包含非天然编码氨基酸的分离的bst多肽与水溶性聚合物接触,所述水溶性聚合物包含与所述非天然编码氨基酸反应的部分。在一些实施例中,并入bst多肽中的非天然编码氨基酸对水溶性聚合物具反应性,而水溶性聚合物另外对20种常见氨基酸中任一者均无反应性。在一些实施例中,并入bst多肽中的非天然编码氨基酸对连接子、聚合物或生物活性分子具反应性,而所述连接子、聚合物或生物活性分子另外对20种常见氨基酸中任一者均无反应性。
在一些实施例中,通过使包含含羰基氨基酸的bst多肽与包含氨氧基、肼基、酰肼基或氨基脲基的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bst多肽。在一些实施例中,氨氧基、肼基、酰肼基或氨基脲基是通过酰胺键与聚(乙二醇)分子连接。在一些实施例中,氨氧基、肼基、酰肼基或氨基脲基是通过氨基甲酸酯键与聚(乙二醇)分子连接。
在一些实施例中,通过使包含羰基的聚(乙二醇)分子与包含含氨氧基、肼基、酰肼基或氨基脲基的非天然编码氨基酸的多肽反应,来制备与水溶性聚合物连接的bst多肽。
在一些实施例中,通过使包含含炔氨基酸的bst多肽与包含叠氮部分的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bst多肽。在一些实施例中,叠氮基或炔基是通过酰胺键与聚(乙二醇)分子连接。
在一些实施例中,通过使包含含叠氮基氨基酸的bst多肽与包含炔部分的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bst多肽。在一些实施例中,叠氮基或炔基是通过酰胺键与聚(乙二醇)分子连接。
本发明也提供制备与水溶性聚合物连接的bgh多肽的方法。在一些实施例中,所述方法包含使包含非天然编码氨基酸的分离的bgh多肽与水溶性聚合物接触,所述水溶性聚合物包含与所述非天然编码氨基酸反应的部分。在一些实施例中,并入bgh多肽中的非天然编码氨基酸对水溶性聚合物具反应性,而水溶性聚合物另外对20种常见氨基酸中任一者均无反应性。在一些实施例中,并入bgh多肽中的非天然编码氨基酸对连接子、聚合物或生物活性分子具反应性,而所述连接子、聚合物或生物活性分子另外对20种常见氨基酸中任一者均无反应性。
在一些实施例中,通过使包含含羰基氨基酸的bgh多肽与包含氨氧基、肼基、酰肼基或氨基脲基的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bgh多肽。在一些实施例中,氨氧基、肼基、酰肼基或氨基脲基是通过酰胺键与聚(乙二醇)分子连接。在一些实施例中,氨氧基、肼基、酰肼基或氨基脲基是通过氨基甲酸酯键与聚(乙二醇)分子连接。
在一些实施例中,通过使包含羰基的聚(乙二醇)分子与包含含氨氧基、肼基、酰肼基或氨基脲基的非天然编码氨基酸的多肽反应,来制备与水溶性聚合物连接的bgh多肽。
在一些实施例中,通过使包含含炔氨基酸的bgh多肽与包含叠氮部分的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bgh多肽。在一些实施例中,叠氮基或炔基是通过酰胺键与聚(乙二醇)分子连接。
在一些实施例中,通过使包含含叠氮基氨基酸的bgh多肽与包含炔部分的聚(乙二醇)分子反应来制备与水溶性聚合物连接的bgh多肽。在一些实施例中,叠氮基或炔基是通过酰胺键与聚(乙二醇)分子连接。
在一些实施例中,聚(乙二醇)分子的分子量介于约0.1kda与约100kda之间。在一些实施例中,聚(乙二醇)分子的分子量介于0.1kda与50kda之间。
在一些实施例中,聚(乙二醇)分子为分支聚合物。在一些实施例中,聚(乙二醇)分支聚合物中各个分支的分子量介于1kda与100kda之间,或介于1kda与50kda之间。
在一些实施例中,与bst多肽连接的水溶性聚合物包含聚烷二醇部分。在一些实施例中,并入bst多肽中的非天然编码氨基酸残基包含羰基、氨氧基、酰肼基、肼基、氨基脲基、叠氮基或炔基。在一些实施例中,并入bst多肽中的非天然编码氨基酸残基包含羰基部分且水溶性聚合物包含氨氧基、酰肼、肼或氨基脲部分。在一些实施例中,并入bst多肽中的非天然编码氨基酸残基包含炔部分且水溶性聚合物包含叠氮部分。在一些实施例中,并入bst多肽中的非天然编码氨基酸残基包含叠氮部分且水溶性聚合物包含炔部分。
本发明也提供包含bst多肽和医药学上可接受的载剂的组合物,所述bst多肽包含非天然编码氨基酸。在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。
本发明还提供包含编码bst多肽的聚核苷酸的细胞,所述聚核苷酸包含选择密码子。在一些实施例中,所述细胞包含正交rna合成酶和/或正交trna,用以将非天然编码氨基酸取代入bst多肽中。
本发明也提供制备包含非天然编码氨基酸的bst多肽的方法。在一些实施例中,所述方法包含在允许表达bst多肽的条件下,培养包含一个或一个以上编码bst多肽的聚核苷酸、正交rna合成酶和/或正交trna的细胞;和从细胞和/或培养基中纯化bst多肽。
本发明也提供增加bst多肽的治疗半衰期、血清半衰期或循环时间的方法。本发明还提供调节bst多肽的免疫原性的方法。在一些实施例中,所述方法包含用非天然编码氨基酸取代天然存在bst多肽中的任一个或一个以上氨基酸,和/或将bst多肽与连接子、聚合物、水溶性聚合物或生物活性分子连接。
本发明也提供用有效量的本发明的bst分子治疗需要所述治疗的患者的方法。在一些实施例中,所述方法包含对患者投予治疗有效量的医药组合物,所述医药组合物包含bst多肽和医药学上可接受的载剂,所述bst多肽包含非天然编码氨基酸。在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。在一些实施例中,bst多肽经糖基化。在一些实施例中,bst多肽未糖基化。
在一些实施例中,与bgh多肽连接的水溶性聚合物包含聚烷二醇部分。在一些实施例中,并入bgh多肽中的非天然编码氨基酸残基包含羰基、氨氧基、酰肼基、肼基、氨基脲基、叠氮基或炔基。在一些实施例中,并入bgh多肽中的非天然编码氨基酸残基包含羰基部分,且水溶性聚合物包含氨氧基、酰肼、肼或氨基脲部分。在一些实施例中,并入bgh多肽中的非天然编码氨基酸残基包含炔部分且水溶性聚合物包含叠氮部分。在一些实施例中,并入bgh多肽中的非天然编码氨基酸残基包含叠氮部分且水溶性聚合物包含炔部分。
本发明也提供包含bgh多肽和医药学上可接受的载剂的组合物,所述bgh多肽包含非天然编码氨基酸。在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。
本发明还提供包含编码bgh多肽的聚核苷酸的细胞,所述聚核苷酸包含选择密码子。在一些实施例中,所述细胞包含正交rna合成酶和/或正交trna,用以将非天然编码氨基酸取代入bgh多肽中。
本发明也提供制备包含非天然编码氨基酸的bgh多肽的方法。在一些实施例中,所述方法包含在允许表达bgh多肽的条件下,培养包含一个或一个以上编码bgh多肽的聚核苷酸、正交rna合成酶和/或正交trna的细胞;和从细胞和/或培养基中纯化bgh多肽。
本发明也提供增加bgh多肽的治疗半衰期、血清半衰期或循环时间的方法。本发明还提供调节bgh多肽的免疫原性的方法。在一些实施例中,所述方法包含用非天然编码氨基酸取代天然存在bgh多肽中的任一个或一个以上氨基酸,和/或将bgh多肽与连接子、聚合物、水溶性聚合物或生物活性分子连接。
本发明也提供用有效量的本发明的bgh分子治疗需要所述治疗的患者的方法。在一些实施例中,所述方法包含对患者投予治疗有效量的医药组合物,所述医药组合物包含bgh多肽和医药学上可接受的载剂,所述bgh多肽包含非天然编码氨基酸。在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。在一些实施例中,bgh多肽经糖基化。在一些实施例中,bgh多肽未糖基化。
本发明也提供包含seqidno:1、2中所示序列或任何其它bst多肽序列的bst多肽,但至少一个氨基酸经非天然编码氨基酸取代。本发明还提供包含已知的bgh190个氨基酸的序列的bgh多肽,但至少一个氨基酸经非天然编码氨基酸取代。在一些实施例中,非天然编码氨基酸与水溶性聚合物连接。在一些实施例中,水溶性聚合物包含聚(乙二醇)部分。在一些实施例中,非天然编码氨基酸包含羰基、氨氧基、酰肼基、肼基、氨基脲基、叠氮基或炔基。
本发明也提供医药组合物,其包含医药学上可接受的载剂和bst多肽,所述bst多肽包含seqidno:1、2中所示序列,或任何其它bst多肽序列,其中至少一个氨基酸经非天然编码氨基酸取代。本发明也提供医药组合物,其包含医药学上可接受的载剂和bst多肽,所述bst多肽包含seqidno:1、2中所示的序列。在一些实施例中,非天然编码氨基酸包含糖部分。在一些实施例中,水溶性聚合物是经由糖部分与多肽连接。在一些实施例中,连接子、聚合物或生物活性分子经由糖部分与bst多肽连接。
本发明也提供一种bst多肽,其包含经由共价键与bst多肽的单一氨基酸连接的水溶性聚合物。在一些实施例中,水溶性聚合物包含聚(乙二醇)部分。在一些实施例中,与水溶性聚合物共价连接的氨基酸是多肽中存在的非天然编码氨基酸。
在本发明一些实施例中,包含经由共价键与bst多肽连接的hes的bst多肽是在单一氨基酸处连接。在一些实施例中,与hes共价连接的单一氨基酸是多肽中存在的非天然编码氨基酸。在本发明一些实施例中,bst多肽包含多个非天然编码氨基酸,这些氨基酸可与多个hes和/或peg分子连接。
本发明提供一种bst多肽,其包含至少一个连接子、聚合物或生物活性分子,其中所述连接子、聚合物或生物活性分子通过经核糖体并入多肽中的非天然编码氨基酸的官能团与多肽连接。在一些实施例中,多肽经单聚乙二醇化。本发明也提供一种bst多肽,其包含与一个或一个以上非天然编码氨基酸连接的连接子、聚合物或生物活性分子,其中所述非天然编码氨基酸是经核糖体并入多肽中预先选择的位点。
本发明的范围内包括与bst编码区接合的bst前导序列或信号序列,以及与bst编码区接合的异源信号序列。所选异源前导序列或信号序列应为例如由宿主细胞分泌系统分泌而经识别和加工且可能由宿主细胞的信号肽酶裂解的序列。用本发明的bst治疗病状或病症的方法意指用含或不含信号或前导肽的bst治疗。
本发明提供一种治疗和预防动物感染的方法。本发明也提供一种治疗和预防牛动物的乳腺炎和航运热的方法。本发明也提供一种治疗未积累细菌的菌株抗性的动物感染的方法。另外,本发明还提供经纯化和分离的多肽,其具有天然存在bst或bgh的部分或全部一级结构构象和一种或一种以上生物特性;以及编码所述bst或bgh的dna序列。
在本发明另一实施例中,将一个或一个以上其它集落刺激因子与促生长素和/或生长激素一起投予受感染的动物,所述其它集落刺激因子包括(但不限于)g-cst、gm-csf、m-csf和多集落刺激因子(il-3)。这些可一起或分开投予。在另一实施例中,bst治疗是以预防性方式使用。可使用bst,通过促生长素血清水平增加引起体重增益增加、产奶量增大或任何其它所需的生理反应。
在另一实施例中,包含一个或一个以上非天然存在氨基酸的bst多肽与另一分子(包括(但不限于)peg)的结合将因结合于所述非天然氨基酸所用的独特化学反应而提供实质上纯化的bst。在进行结合步骤之前或之后,可利用其它纯化技术将包含一个或一个以上非天然编码氨基酸的bst与另一分子(例如peg)结合,以提供实质上纯的bst。
在另一实施例中,包含一个或一个以上非天然存在氨基酸的bgh多肽与另一分子(包括(但不限于)peg)的结合将因结合于所述非天然氨基酸所用的独特化学反应而提供实质上纯化的bgh。在进行结合步骤之前或之后,可利用其它纯化技术将包含一个或一个以上非天然编码氨基酸的bgh与另一分子(例如peg)结合,以提供实质上纯的bgh。
附图说明
图1—显示考马斯蓝(coomassieblue)染色的sds-page,说明在以下各位置包含非天然编码氨基酸对乙酰基苯丙氨酸的hgh的表达:y35、f92、y111、g131、r134、k140、y143或k145。
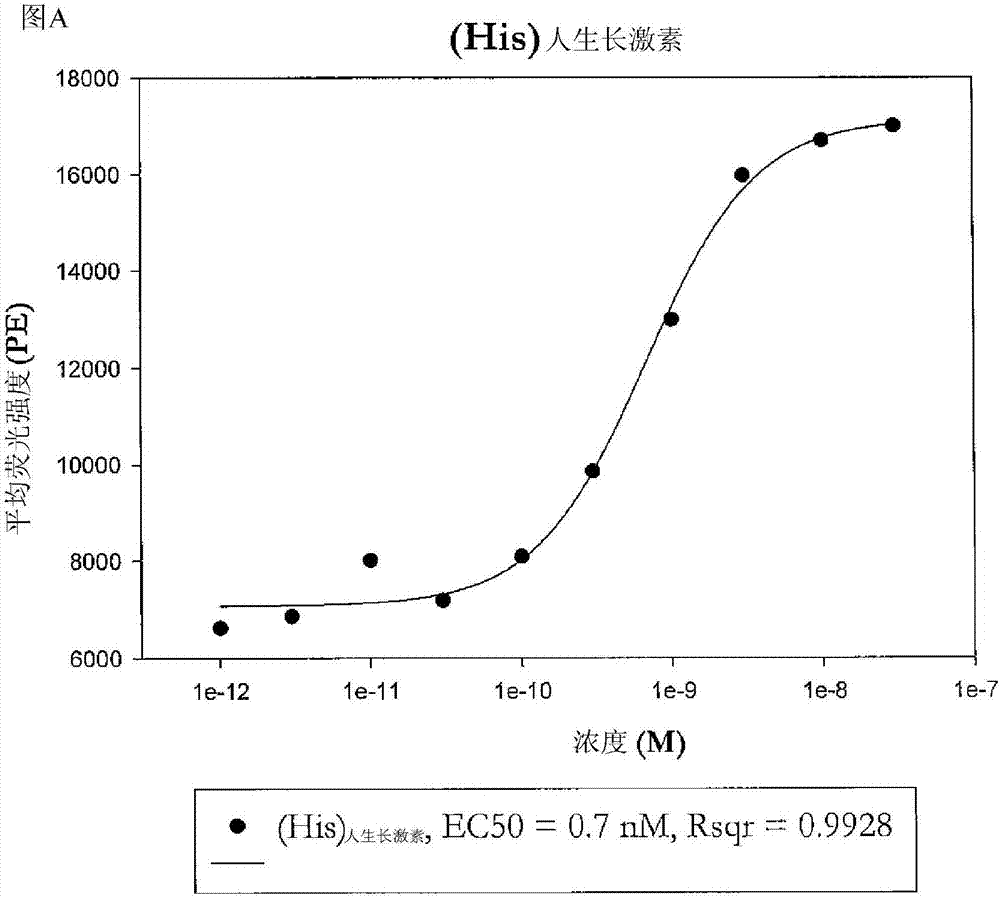
图2图a和图b—显示在im9细胞上包含非天然编码氨基酸的hgh(图b)和野生型hgh(图a)的生物活性的图。
图3—显示考马斯蓝染色的sds-page,说明通过peg(5、20和30kda)与非天然编码氨基酸的共价连接聚乙二醇化的包含所述非天然编码氨基酸的hgh的产生。
图4—显示的图说明在im9细胞上包含非天然编码氨基酸的hgh的各种聚乙二醇化形式的生物活性。
图5图a—本图描绘具有所示胰蛋白酶裂解位点和箭头说明的非天然氨基酸取代f92paf的hgh的一级结构(本图由贝克尔(becker)等人,生物技术与应用生物化学(biotechnolapplbiochem.)(1988)10(4):326-337修改得到)。图5图b—显示由聚乙二醇化的包含非天然编码氨基酸的hgh多肽产生的肽(标为a)、由包含非天然编码氨基酸的hgh多肽产生的肽(标为b)和由whorhgh产生的肽(标为c)的叠加胰蛋白酶图。图5图c—显示图b中峰9的放大图。
图6图a和图b显示纯化的peg-hgh多肽的考马斯蓝染色sds-page分析。
图7—显示在im9细胞上hgh二聚体分子的生物活性的图。
图8图a—显示im-9分析数据的图解,所述im-9分析数据是利用含g120r取代的hgh拮抗剂由pstat5磷酸化作用测量。图8图b—显示im-9分析数据的图解,所述im-9分析数据是利用在同一位置(g120)并入非天然氨基酸的hgh多肽由pstat5磷酸化作用测量。
图9—显示的图表明图8图b中所示hgh拮抗剂二聚体在im-9分析中也缺乏生物活性。
图10—显示的图比较聚乙二醇化的包含非天然编码氨基酸的hgh多肽与未聚乙二醇化的hgh多肽在大鼠中的血清半衰期。
图11-显示的图比较聚乙二醇化的包含非天然编码氨基酸的hgh多肽在大鼠中的血清半衰期。
图12-显示的图比较聚乙二醇化的包含非天然编码氨基酸的hgh多肽在大鼠中的血清半衰期。按2.1mg/kg对大鼠给药一次。
图13图a-显示在投予一剂聚乙二醇化(35位、92位)的包含非天然编码氨基酸的hgh多肽之后对大鼠体重增益的影响的图解。图13图b-显示在投予一剂聚乙二醇化(35位、92位)的包含非天然编码氨基酸的hgh多肽之后对循环血浆igf-1水平的影响的图解。图13图c-显示在投予一剂聚乙二醇化(92位、134位、145位、131位、143位)的包含非天然编码氨基酸的hgh多肽之后对大鼠体重增益的影响的图解。图13图d-显示在投予一剂聚乙二醇化(92位、134位、145位、131位、143位)的包含非天然编码氨基酸的hgh多肽之后对循环血浆igf-1水平的影响的图解。图13图e-显示的图比较聚乙二醇化(92位、134位、145位、131位、143位)的包含非天然编码氨基酸的hgh多肽在大鼠中的血清半衰期。
图14-显示线性30kda单甲氧基-聚(乙二醇)-2-氨氧基乙胺氨基甲酸酯盐酸盐的结构的图解。
图15-显示的图说明氨基甲酸酯连接的氧氨基衍生化peg的合成。
图16提供含peg试剂的说明性非限制性实例,所述含peg试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图17提供含peg试剂的合成的说明性非限制性实例,所述含peg试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图18提供含基于酰胺的羟基胺peg试剂的合成的说明性非限制性实例,所述试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图19提供基于氨基甲酸酯的含peg试剂的合成的说明性非限制性实例,所述试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图20提供基于氨基甲酸酯的含peg试剂的合成的说明性非限制性实例,所述试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图21提供简单的含peg试剂的合成的说明性非限制性实例,所述含peg试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图22提供含分支peg试剂的说明性非限制性实例以及使用一种此类试剂修饰基于羰基的非天然氨基酸多肽,所述含分支peg试剂可用于修饰非天然氨基酸多肽,形成肟连接的含peg非天然氨基酸多肽。
图23显示结合的非天然编码氨基酸的化学的图解,所述非天然编码氨基酸包括对乙酰基phe,或对乙酰基苯丙氨酸,或paf,或pacf;对氨基phe,或对氨基苯丙氨酸,或paf2,或panf;和对叠氮基phe,或对叠氮基苯丙氨酸,或paf3,或pazf。
图24显示在图(a)标准加工条件;以及图(b)利用所述方法内的溶解还原步骤时实例3的pst-f92物质的rp-hplc分析。
图25显示实例3的peg反应的sds-page分析,其中对照物在每一凝胶的最左边,且标为1到4的列是:1)预聚乙二醇化物质;2)按peg:蛋白质比率0.9:1进行peg反应;3)按peg:蛋白质比率1:1进行peg反应;和4)按peg:蛋白质比率1.5:1进行peg反应。
具体实施方式
定义
应了解,本发明不限于本文中描述的特定方法、方案、细胞系、构建体和试剂,因此其可变化。还应了解,本文中使用的术语只是出于描述特定实施例的目的,而不打算限制本发明的范围,本发明的范围应仅受所附权利要求书限制。
除非本文中另作清楚指示,否则如本文和随附权利要求中使用的单数形式“一个(种)”和“所述”包括多个(种)参考物。因此,例如,提到的一种“bst”、“牛st”、“牛促生长素”、“b促生长素”、“牛促生长素多肽”或“st多肽”以及各种带连字符和不带连字符形式是指一种或一种以上所述蛋白质,且包括所属领域技术人员已知的所述蛋白质的等效物等。
除非另作定义,否则本文中使用的所有科技术语都具有与本发明所属领域技术人员通常所了解相同的含义。尽管可使用与本文中所述相似或相同的任何方法、装置和材料来实施或测试本发明,但现仅描述优选方法、装置和材料。
本文中提到的所有出版物和专利都是以引用的方式并入本文中,旨在描述和揭示例如出版物中所述的可能与本发明结合使用的构建体和方法。本文中论述的出版物只提供在本申请案申请日期之前的揭示内容。本文在任何方面都不应解释为承认本发明者无权由于现有发明或任何其它原因使所述揭示案的日期提前。
术语“实质上纯化”是指bst多肽可实质上或基本上不含如在天然存在环境(即,天然细胞,或在重组产生bst多肽情况下是指宿主细胞)中所见的通常伴随所述蛋白质或与所述蛋白质相互作用的组分。可实质上不含细胞材料的bst多肽包括具有小于约30%、小于约25%、小于约20%、小于约15%、小于约10%、小于约5%、小于约4%、小于约3%、小于约2%或小于约1%(以干重计)污染蛋白质的蛋白质制剂。当由宿主细胞重组产生bst多肽或其变异体时,所述蛋白质可以按细胞干重计约30%、约25%、约20%、约15%、约10%、约5%、约4%、约3%、约2%或约1%或更低存在。当由宿主细胞重组产生bst多肽或其变异体时,所述蛋白质可以按细胞干重计约5g/l、约4g/l、约3g/l、约2g/l、约1g/l、约750mg/l、约500mg/l、约250mg/l、约100mg/l、约50mg/l、约10mg/l或约1mg/l或更低存在于培养基中。因此,如借助于例如sds/page分析、rp-hplc、sec和毛细管电泳等适当方法所测定,由本发明方法产生的“实质上纯化”的bst多肽可具有至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%的纯度水平,尤其至少约75%、80%、85%的纯度水平且更尤其至少约90%的纯度水平、至少约95%的纯度水平、至少约99%或更高的纯度水平。
“重组宿主细胞”或“宿主细胞”是指包括外源聚核苷酸的细胞,不管用于插入的方法如何,例如直接摄取、转导、f配对,或此项技术中已知用于产生重组宿主细胞的其它方法。外源聚核苷酸可保持非整合载体(例如质粒)的形式,或者可整合到宿主基因组中。
本文中使用的术语“培养基”包括任何培养基、溶液、固体、半固体或刚性支撑物,其可支撑或含有任何宿主细胞,包括细菌宿主细胞、酵母宿主细胞、昆虫宿主细胞、植物宿主细胞、真核生物宿主细胞、哺乳动物宿主细胞、cho细胞、原核生物宿主细胞、大肠杆菌或假单胞菌属(pseudomonas)宿主细胞,以及细胞内容物。因此,这一术语可涵盖生长有宿主细胞的培养基,例如分泌有bst多肽的培养基,包括增殖步骤之前或之后的培养基。这一术语还可涵盖含有宿主细胞溶解产物的缓冲液或试剂,例如bst多肽是在胞内产生并且宿主细胞溶解或破裂而释放bst多肽的情形。
本文中提到蛋白质再折叠时使用的“还原剂”定义为使硫氢基保持还原态并且还原分子内或分子间二硫键的任何化合物或材料。适当还原剂包括(但不限于)二硫苏糖醇(dtt)、2-巯基乙醇、二硫赤藓醇、半胱氨酸、半胱胺(2-氨基乙硫醇)和还原型谷胱甘肽。所属领域技术人员易于了解,多种还原剂适用于本发明的方法和组合物中。
本文中提到蛋白质再折叠时使用的“氧化剂”定义为能够从经氧化的化合物中去除电子的任何化合物或材料。适当氧化剂包括(但不限于)氧化型谷胱甘肽、胱氨酸、胱胺、氧化的二硫苏糖醇、氧化的赤藓醇和氧。所属领域技术人员易于了解,多种氧化剂适用于本发明的方法中。
本文中使用的“变性剂”定义为会使蛋白质可逆展开的任何化合物或材料。变性剂的强度将由特定变性剂的特性和浓度决定。适当变性剂可为离液剂、清洁剂、有机溶剂、水混溶性溶剂、磷脂或者两种或两种以上所述试剂的组合。适当离液剂包括(但不限于)脲、胍和硫代氰酸钠。有用的清洁剂可包括(但不限于)强清洁剂,例如十二烷基硫酸钠或聚氧乙烯醚(例如,吐温(tween)或曲通(triton)清洁剂)、月桂酰肌氨酸钠(sarkosyl);温和的非离子型清洁剂(例如,毛地黄皂苷);温和的阳离子型清洁剂,例如n→2,3-(二油酰基氧基)-丙基-n,n,n-三甲基铵;温和的离子型清洁剂(例如,胆酸钠或去氧胆酸钠);或两性离子型清洁剂,包括(但不限于)硫代甜菜碱(两性离子清洁剂(zwittergent))、3-(3-胆酰胺基丙基)二甲铵基-1-丙烷硫酸盐(chaps)和3-(3-胆酰胺基丙基)二甲铵基-2-羟基-1-丙烷磺酸盐(chapso)。水混溶性有机溶剂,例如乙腈、低碳数烷醇(尤其c2-c4烷醇,例如乙醇或异丙醇)或低碳数烷二醇(尤其c2-c4烷二醇,例如乙二醇),可用作变性剂。适用于本发明中的磷脂可为天然存在的磷脂,例如磷脂酰乙醇胺、磷脂酰胆碱、磷脂酰丝氨酸和磷脂酰肌醇,或合成磷脂衍生物或变异体,例如二己酰磷脂酰胆碱或二庚酰磷脂酰胆碱。
本文中使用的“再折叠”是针对二硫键,其描述将含有二硫键的多肽由不适当折叠或展开状态转化成天然或适当折叠的构象的任何过程、反应或方法。
本文中使用的“共折叠”尤其是指使用至少两个多肽的再折叠过程、反应或方法,这些多肽彼此相互作用,并且使展开或不适当折叠的多肽转化成天然的适当折叠的多肽。
本文中使用的“牛促生长素”、“牛st”或“bst”应包括具有bst的至少一种生物活性的那些多肽和蛋白质,以及bgh和bgh类似物、bst和bgh同工型、bst和bgh模拟物、bst和bgh片段、杂交bst和bgh蛋白、融合蛋白寡聚物和多聚体、同系物、糖基化形式变异体和突变蛋白,不管是否具有相同生物活性,而且也不管其合成或制造方法如何,所述方法包括(但不限于)重组(由cdna、基因组dna、合成dna或其它形式核酸制造)、合成、转基因和基因活化方法。bst的具体实例包括(但不限于)bst突变体、改变的糖基化bst、和peg结合的bst类似物。
术语“牛促生长素”或“bst”是指上文和本申请案通篇所述的牛bst或牛促生长素,以及保持天然存在bst的至少一种生物活性的多肽。bst多肽包括天然存在的牛促生长素的医药学上可接受的盐和前药,和所述盐的前药、多晶型物、水合物、溶剂化物、生物活性片段、生物活性变异体和立体异构体,以及天然存在的牛促生长素的激动剂、模拟物和拮抗剂变异体,和其多肽融合体。术语“bst多肽”内也涵盖在氨基末端、羧基末端或二者处包含其它氨基酸的融合体。示范性融合体包括(但不限于)例如由重组表达成熟形式的bst得到的甲硫氨酰基bst,其中甲硫氨酸与bst(例如seqidno:1或2的多肽)的n末端连接;用于纯化目的的融合体(包括(但不限于)聚组氨酸或亲和表位);与血清白蛋白结合肽的融合体;以及与例如血清白蛋白等血清蛋白的融合体。已知全长和成熟形式的天然存在bst核酸和氨基酸序列,以及变异体,例如单一氨基酸变异体和剪接变异体。有关成熟bst氨基酸序列以及甲硫氨酰基bst氨基酸序列,分别参见本文中的seqidno:1和seqidno:2。也已知编码hg-csf突变体的核酸分子以及突变hg-csf多肽。
已经描述bst中多个氨基酸位置的取代。取代包括(但不限于)调节医药稳定性、增加激动剂活性、增加蛋白酶抗性、将多肽转变成拮抗剂等的取代,并且都涵盖在术语“bst多肽”、“牛促生长素多肽”、“牛st”或“bst”中。
另一方面,本发明提供编码变异蛋白的重组核酸、含有变异核酸的表达载体、包含变异核酸和/或表达载体的宿主细胞,以及制造变异蛋白的方法。另一方面,本发明提供通过对动物投予治疗有效量的变异蛋白,通常连同医药载剂,来治疗感染。
在一些实施例中,本发明的bst多肽与seqidno:1、2或任何其它bst多肽序列实质上一致。众所周知编码包括突变体的bst多肽的核酸分子以及表达和纯化bst多肽的方法。
术语“bst多肽”还包括天然存在bst的医药学上可接受的盐和前药,和所述盐的前药、多晶型物、水合物、溶剂化物、生物活性片段、生物活性变异体和立体异构体,以及天然存在bst的激动剂、模拟物和拮抗剂变异体,和其多肽融合体。术语“bst多肽”内也涵盖在氨基末端、羧基末端或二者处包含其它氨基酸的融合体。示范性融合体包括(但不限于)例如由重组表达缺乏前导或信号肽或其部分的成熟形式的bst(通过重组表达,甲硫氨酸与bst的n末端连接)得到的甲硫氨酰基bst,其中甲硫氨酸与bst的n末端连接;用于纯化目的的融合体(包括(但不限于)聚组氨酸或亲和表位);与血清白蛋白结合肽的融合体;以及与例如血清白蛋白等血清蛋白的融合体。美国专利第5,750,373号(以引用的方式并入本文中)描述了一种用于选择新颖蛋白质(例如生长激素,和对个别受体分子的结合特性已改变的抗体片段变异体)的方法。所述方法包含将编码相关蛋白质的基因与丝状噬菌体m13的基因iii外壳蛋白的羧基末端域融合。嵌合分子包含bst和一个或一个以上其它分子。嵌合分子可含有bst和所述其它分子中一者或两者的特定区域或片段。任一所述片段都可由所述蛋白质借助于标准生物化学方法,或者通过表达编码所述片段的聚核苷酸制备得到。bst或其片段可制备成包含人血清白蛋白(hsa)、fc或其部分的融合蛋白的形式。所述融合构建体适于增强真核宿主细胞中bst或其片段的表达。示范性hsa部分包括n末端多肽(氨基酸1-369、1-419,和以氨基酸1开始的中等长度氨基酸),如美国专利第5,766,883号和公开案wo97/24445中所揭示,这些专利都以引用的方式并入本文中。其它嵌合多肽可包括hsa蛋白以及与hsa的c末端和n末端中每一者连接的bst或其片段。其它融合体可以由bst与a)免疫球蛋白的fc部分;b)免疫球蛋白fc部分的类似物;和c)免疫球蛋白fc部分的片段融合而产生。
多篇参考文献揭示了通过聚合物结合或糖基化来修饰多肽。术语“bst多肽”包括与例如peg等聚合物结合的多肽,且可包含半胱氨酸、赖氨酸或其它残基的一种或一种以上其它衍生作用。此外,bst多肽可包含连接子或聚合物,其中与所述连接子或聚合物结合的氨基酸可为根据本发明的非天然氨基酸,或可利用此项技术中已知的技术(例如与赖氨酸或半胱氨酸偶合)与天然编码氨基酸结合。
已经报导过多肽的聚合物修饰。ifnβ是作为属于生长激素超家族的多肽的一个实例而被提到。wo00/23114揭示了糖基化且聚乙二醇化的ifnβ。wo00/23472揭示了ifnβ融合蛋白。美国专利第4,904,584号揭示了耗尽聚乙二醇化赖氨酸的多肽,其中至少一个赖氨酸残基已缺失或经任何其它氨基酸残基置换。wo99/67291揭示了一种将蛋白质与peg结合的方法,其中所述蛋白质上至少一个氨基酸残基已缺失,并且所述蛋白质是在足以实现与所述蛋白质结合的条件下与peg接触。wo99/03887揭示了属于生长激素超家族的聚乙二醇化多肽变异体,其中半胱氨酸残基已经由位于所述多肽指定区域中的非必需氨基酸残基取代。wo00/26354揭示了一种制备变应原性较低的糖基化多肽变异体的方法,与相应的母体多肽相比,所述糖基化多肽变异体包含至少另一个糖基化位点。
术语“bst多肽”还包括糖基化bst,例如(但不限于)任一氨基酸位置糖基化的多肽、n连接或o连接糖基化形式的多肽。含有单一核苷酸改变的变异体也视作bst多肽的生物活性变异体。含有单一核苷酸改变的变异体也视作bst的生物活性变异体。此外,还包括剪接变异体。术语“bst多肽”也包括通过化学方式连接或表达为融合蛋白形式的任一种或一种以上bst或任何其它多肽、蛋白质、碳水化合物、聚合物、小分子、连接子、配体或任何类型的其它活性分子的bst杂二聚体、均二聚体、杂多聚体或均多聚体(参见美国专利第6,261,550号、第6,166,183号、第6,204,247号、第6,261,550号、第6,017,876号,以引用的方式并入本文中),以及含有例如特定缺失或其它修饰但仍保持生物活性的多肽类似物(美国专利第6,261,550号、第6,004,548号、第6,632,426号,以引用的方式并入本文中)。
除非另作说明(即,当规定比较是基于seqidno:2,或其它bst序列时),否则提到的本文所述的bst中的所有氨基酸位置都是基于seqidno:1中的位置。举例来说,seqidno:1的1位氨基酸为苏氨酸,而相应的苏氨酸位于seqidno:2中的2位。所属领域技术人员应理解,易于在任何其它bst分子(例如seqidno:2)中鉴别出对应于seqidno:1中各位置的氨基酸位置。所属领域技术人员应理解,易于在任何其它bst分子(例如bst融合体、变异体、片段等)中鉴别出对应于seqidno:1、2或任何其它bst序列中各位置的氨基酸位置。举例来说,可使用例如blast等序列比对程序来比对和鉴别一种蛋白质中与seqidno:1、2或其它bst序列中的位置对应的特定位置。本文中参照seqidno:1、2或其它bst序列描述的氨基酸取代、缺失或添加也打算指在本文中所述或此项技术中已知的bst融合体、变异体、片段等中相应位置的取代、缺失或添加,且这些都明确地涵盖在本发明内。
术语“bst多肽”或“bst”涵盖包含一个或一个以上氨基酸取代、添加或缺失的bst多肽。本发明的bst多肽可包含一个或一个以上天然氨基酸修饰与一个或一个以上非天然氨基酸修饰的组合。已经描述在天然存在bst多肽中多个氨基酸位置的示范性取代,包括(但不限于)调节医药稳定性、调节bst多肽的一种或一种以上生物活性的取代,所述调节生物活性为例如(但不限于)增加激动剂活性、增加多肽溶解性、降低蛋白酶易感性、将所述多肽转变成拮抗剂等,且这些取代都涵盖在术语“bst多肽”中。在一些实施例中,bst拮抗剂包含与水溶性聚合物连接的非天然编码氨基酸,其存在于bst分子的受体结合区内。
在一些实施例中,bst多肽另外包含调节bst多肽生物活性的添加、取代或缺失。在一些实施例中,bst多肽另外包含调节bst多肽的中性粒细胞增殖、功能和/或分化的添加、取代或缺失。举例来说,所述添加、取代或缺失可调节bst的一种或一种以上特性或活性。举例来说,所述添加、取代或缺失可调节对受体的亲和力、调节循环半衰期、调节治疗半衰期、调节多肽稳定性、调节蛋白酶的裂解作用、调节剂量、调节释放或生物利用率、促进纯化,或者改进或改变特定投药途径。类似地,bst多肽可包含蛋白酶裂解序列、反应性基团、抗体结合域(包括(但不限于)flag或聚组氨酸)或其它基于亲和力的序列(包括(但不限于)flag、聚组氨酸、gst等)或改进对于多肽的检测(包括(但不限于)gfp)、纯化或其它特质的连接分子(包括(但不限于)生物素)。
术语“bst多肽”还涵盖均二聚体、杂二聚体、均多聚体和杂多聚体,其与相同或不同非天然编码氨基酸侧链、天然编码氨基酸侧链连接,包括(但不限于)经由非天然编码氨基酸侧链直接连接或经由连接子间接连接。示范性连接子包括(但不限于)小有机化合物;各种长度的水溶性聚合物,例如聚(乙二醇)或葡聚糖;或各种长度的多肽。
“非天然编码氨基酸”是指不属于20种常见氨基酸中一者或者不为吡咯赖氨酸或硒代半胱氨酸的氨基酸。可与术语“非天然编码氨基酸”同义使用的其它术语为“非天然氨基酸”、“非天然存在氨基酸”以及其各种带连字符和不带连字符的型式。术语“非天然编码氨基酸”也包括(但不限于)通过修饰(例如,翻译后修饰)天然编码氨基酸(包括(但不限于)20种常见氨基酸,或吡咯赖氨酸和硒代半胱氨酸)而产生,但本身未通过翻译复合物天然地并入到生长多肽链中的氨基酸。所述非天然存在氨基酸的实例包括(但不限于)n-乙酰基氨基葡糖基-l-丝氨酸、n-乙酰基氨基葡糖基-l-苏氨酸和o-磷酸酪氨酸。
“氨基末端修饰基团”是指可与多肽的氨基末端连接的任何分子。类似地,“羧基末端修饰基团”是指可与多肽的羧基末端连接的任何分子。末端修饰基团包括(但不限于)各种水溶性聚合物、肽或蛋白质(例如血清白蛋白),或者增加肽的血清半衰期的其它部分。
此项技术中和本文中使用的术语“官能团”、“活性部分”、“活化基团”、“离去基团”、“反应位点”、“化学反应性基团”和“化学反应性部分”是指分子中截然不同的、可定义的部分或单元。这些术语在某种程度上与在化学技术中的含义同义,且在本文中用于指示分子中执行某种功能或活性并与其它分子反应的部分。
本文中使用的术语“键”或“连接子”是指通常由化学反应形成的基团或键,且通常为共价键。水解稳定性键意思是指,这些键在水中实质上稳定,并且在有用的ph值下(包括(但不限于)在生理学条件下)可在一段较长时间内(可能甚至无限期)不与水反应。水解不稳定或可降解性键意思是指,这些键可在水或水溶液(包括例如血液)中降解。酶不稳定或可降解性键意思是指,这些键可经一种或一种以上酶降解。此项技术中应了解,peg和相关聚合物可在聚合物主链中或在聚合物主链与聚合物分子的一个或一个以上末端官能团之间的连接基团中包括可降解的键。举例来说,由peg羧酸或活性peg羧酸与生物活性剂上的醇基反应而形成的酯键一般会在生理学条件下水解,释放出所述试剂。其它水解可降解键包括(但不限于)碳酸酯键;由胺与醛反应产生的亚胺键;通过醇与磷酸酯基反应形成的磷酸酯键;作为酰肼与醛的反应产物的腙键;作为醛与醇的反应产物的缩醛键;作为甲酸盐与醇的反应产物的原酸酯键;由(包括(但不限于))聚合物(例如peg)末端的胺基与肽的羧基形成的肽键;和由(包括(但不限于))聚合物末端的亚磷酰胺(phosphoramidite)基与寡核苷酸的5'羟基形成的寡核苷酸键。
当用于本文中时,术语“生物活性分子”、“生物活性部分”或“生物活性剂”意思是可影响与生物体相关的生物系统、路径、分子或相互作用的任何物理或生物化学特性的任何物质,所述生物体包括(但不限于)病毒、细菌、噬菌体、转座子、朊病毒、昆虫、真菌、植物、动物和人类。具体说来,本文中使用的生物活性分子包括(但不限于)打算用于诊断、治愈、减轻、治疗或预防人类或其它动物的疾病,或以其它方式增强人类或动物的身体或精神良好状态的任何物质。生物活性分子的实例包括(但不限于)肽、蛋白质、酶、小分子药物、疫苗、免疫原、硬药(harddrug)、软药、碳水化合物、无机原子或分子、染料、脂质、核苷、放射性核素、寡核苷酸、类毒素、毒素、原核生物和真核生物细胞、病毒、多糖、核酸和其得自或来源于病毒、细菌、昆虫、动物或任何其它细胞或细胞类型的部分,脂质体、微粒和胶束。bst多肽可添加到胶束调配物中。适用于本发明的生物活性剂的种类包括(但不限于)药物、前药、放射性核素、显影剂、聚合物、抗生素、杀真菌剂、抗病毒剂、消炎剂、抗肿瘤剂、心血管药、抗焦虑剂、激素、生长因子、类固醇药、微生物来源的毒素等。
“双官能聚合物”是指包含两个个别官能团的聚合物,这两个官能团能够与其它部分(包括(但不限于)氨基酸侧基)特异性反应,形成共价或非共价键。双官能连接子具有一个可与特定生物活性组分上的基团反应的官能团,以及可与第二生物组分上的基团反应的另一基团,由此可用于形成一种结合物,这种结合物包括第一生物活性组分、双官能连接子和第二生物活性组分。已知许多用于连接各种化合物与肽的程序和连接分子。例如参见欧洲专利申请案第188,256号;美国专利第4,671,958号、第4,659,839号、第4,414,148号、第4,699,784号、第4,680,338号和第4,569,789号,这些专利都以引用的方式并入本文中。“多官能聚合物”是指包含两个或两个以上个别官能团的聚合物,这些官能团能够与其它部分(包括(但不限于)氨基酸侧基)特异性反应,形成共价或非共价键。双官能聚合物或多官能聚合物可为任何所需长度或分子量,并且可经选择以使一个或一个以上与bst连接的分子与其受体或bst之间具有特定所需间隔或构象。
当用从左向右书写的常规化学式说明取代基时,这些取代基同样涵盖由从右向左书写结构所得到的化学上一致的取代基,例如,结构-ch2o-与结构-och2-相同。
术语“取代基”包括(但不限于)“非干扰性取代基”。“非干扰性取代基”是得到稳定化合物的基团。适合非干扰性取代基或基团包括(但不限于)卤基、c1-c10烷基、c2-c10烯基、c2-c10炔基、c1-c10烷氧基、c2-c12芳烷基、c1-c12烷芳基、c3-c12环烷基、c3-c12环烯基、苯基、经取代苯基、甲苯酰基、二甲苯基、联苯基、c2-c12烷氧基烷基、c2-c12烷氧基芳基、c7-c12芳氧基烷基、c7-c12氧基芳基、c1-c6烷基亚磺酰基、c1-c10烷基磺酰基、--(ch2)m--o--(c1-c10烷基)(其中m为1到8)、芳基、经取代芳基、经取代烷氧基、氟烷基、杂环基、经取代杂环基、硝基烷基、-no2、--cn、--nrc(o)--(c1-c10烷基)、--c(o)--(c1-c10烷基)、c2-c10烷基硫烷基、--c(o)o-(-c1-c10烷基)、--oh、--so2、=s、--cooh、--nr2、羰基、--c(o)--(c1-c10烷基)-cf3、--c(o)—cf3、--c(o)nr2、--(c1-c10芳基)-s--(c6-c10芳基)、--c(o)--(c1-c10芳基)、--(ch2)m--o--(--ch2)m--o--(c1-c10烷基)(其中各m为1到8)、--c(o)nr2、--c(s)nr2、--so2nr2、--nrc(o)nr2、--nrc(s)nr2,其盐等。本文中使用的各r为h、烷基或经取代烷基、芳基或经取代芳基、芳烷基或烷芳基。
术语“卤素”包括氟、氯、碘和溴。
除非另作规定,否则单独或作为另一取代基的部分的术语“烷基”意思是指具有指定碳原子数(即,c1-c10意思是指1到10个碳)的直链或支链或者环状烃基或其组合,其可为完全饱和、单或多不饱和的,并且可包括二价和多价基团。饱和烃基的实例包括(但不限于)例如以下基团:甲基、乙基、正丙基、异丙基、正丁基、叔丁基、异丁基、仲丁基、环己基、(环己基)甲基、环丙基甲基;例如正戊基、正己基、正庚基、正辛基的同系物和异构物等。不饱和烷基是具有一个或一个以上双键或三键的烷基。不饱和烷基的实例包括(但不限于)乙烯基、2-丙烯基、巴豆基(crotyl)、2-异戊烯基、2-(丁二烯基)、2,4-戊二烯基、3-(1,4-戊二烯基)、乙炔基、1-和3-丙炔基、3-丁炔基以及高级同系物和异构物。除非另作说明,否则术语“烷基”还打算包括下文更为详细地定义的烷基衍生物,例如“杂烷基”。局限于烃基的烷基称为“同系烷基(homoalkyl)”。
单独或作为另一取代基的部分的术语“亚烷基”意思是指衍生自烷烃的二价基团,例如(但不限于)结构-ch2ch2-和-ch2ch2ch2ch2-,且另外包括下文描述为“亚杂烷基”的基团。通常,烷基(或亚烷基)将具有1到24个碳原子,且具有10个或更少碳原子的基团为本文所述方法和组合物的特定实施例。“低碳数烷基”或“低碳数亚烷基”是一般具有8个或更少碳原子的短链烷基或亚烷基。
术语“烷氧基”、“烷基氨基”和“烷硫基”(或硫烷氧基)是以其常规含义使用,并且指分别经由氧原子、氨基或硫原子与分子剩余部分连接的烷基。
除非另作规定,否则单独或与另一术语组合的术语“杂烷基”意思是指稳定的直链或支链或者环状烃基,或其组合,其是由规定数量的碳原子和至少一个选自由o、n、si和s组成的群组的杂原子组成,并且其中氮和硫原子可任选经氧化,且氮杂原子可任选经季铵化。杂原子o、n以及s和si可位于杂烷基的任一内部位置或在烷基与分子其余部分连接的位置。实例包括(但不限于)-ch2-ch2-o-ch3、-ch2-ch2-nh-ch3、-ch2-ch2-n(ch3)-ch3、-ch2-s-ch2-ch3、-ch2-ch2,-s(o)-ch3、-ch2-ch2-s(o)2-ch3、-ch=ch-o-ch3、-si(ch3)3、-ch2-ch=n-och3和-ch=ch-n(ch3)-ch3。最多两个杂原子可为连续的,例如-ch2-nh-och3和-ch2-o-si(ch3)3。类似地,单独或作为另一取代基的部分的术语“亚杂烷基”意思是指衍生自杂烷基的二价基团,例如(但不限于)-ch2-ch2-s-ch2-ch2-和-ch2-s-ch2-ch2-nh-ch2-。对于亚杂烷基,相同或不同杂原子也可占据任一个或两个链末端(包括(但不限于)亚烷基氧基、亚烷基二氧基、亚烷基氨基、亚烷基二氨基、氨氧基亚烷基等)。另外,对于亚烷基和亚杂烷基连接基团,连接基团化学式的书写方向并不表示连接基团的定向。举例来说,式-c(o)2r'-表示-c(o)2r'-和-r'c(o)2-。
除非另作规定,否则单独或与其它术语组合时术语“环烷基”和“杂环烷基”分别表示“烷基”和“杂烷基”的环状形式。因此,环烷基或杂环烷基包括饱和、部分不饱和和完全不饱和的环键。此外,对于杂环烷基,杂原子可占据杂环与分子其余部分连接的位置。环烷基的实例包括(但不限于)环戊基、环己基、1-环己烯基、3-环己烯基、环庚基等。杂环烷基的实例包括(但不限于)1-(1,2,5,6-四氢吡啶基)、1-哌啶基、2-哌啶基、3-哌啶基、4-吗啉基、3-吗啉基、四氢呋喃-2-基、四氢呋喃-3-基、四氢噻吩-2-基、四氢噻吩-3-基、1-哌嗪基、2-哌嗪基等。此外,这一术语还涵盖双环和三环结构。类似地,单独或作为另一取代基的部分的术语“亚杂环烷基”意思是指衍生自杂环烷基的二价基团,并且单独或作为另一取代基的部分的术语“亚环烷基”意思是指衍生自环烷基的二价基团。
本文中使用的术语“水溶性聚合物”是指可溶于水性溶剂的任何聚合物。水溶性聚合物与bst多肽连接可产生改变,包括(但不限于)相对于未经修饰的形式,血清半衰期增加或经调节,或者治疗半衰期增加或经调节;免疫原性经调节;物理缔合特征(例如聚集和多聚体形成)经调节;受体结合改变;与一种或一种以上结合搭配物的结合改变;和受体二聚化或多聚化改变。所述水溶性聚合物可具有或可不具有其自身的生物活性,并且可用作连接子,将bst与其它物质(包括(但不限于)一个或一个以上bst多肽,或者一个或一个以上生物活性分子)连接。适合的聚合物包括(但不限于)聚乙二醇、聚乙二醇丙醛、其单c1-c10烷氧基或芳氧基衍生物(描述于美国专利第5,252,714号中,其以引用的方式并入本文中)、单甲氧基-聚乙二醇、聚乙烯吡咯烷酮、聚乙烯醇、聚氨基酸、二乙烯基醚马来酸酐、n-(2-羟基丙基)-甲基丙烯酰胺、葡聚糖、葡聚糖衍生物(包括硫酸葡聚糖)、聚丙二醇、聚氧化丙烯/氧化乙烯共聚物、聚氧乙基化多元醇、肝素、肝素片段、多糖、寡糖、聚糖、纤维素和纤维素衍生物(包括(但不限于)甲基纤维素和羧甲基纤维素)、淀粉和淀粉衍生物、多肽、聚烷二醇和其衍生物、聚烷二醇共聚物和其衍生物、聚乙烯基乙基醚和α-β-聚[(2-羟乙基)-dl-天冬酰胺等,或其混合物。所述水溶性聚合物的实例包括(但不限于)聚乙二醇和血清白蛋白。wo03/074087和wo03/074088描述了蛋白质或小分子与羟烷基淀粉(has)的结合。羟烷基淀粉的实例包括(但不限于)羟乙基淀粉。羟烷基淀粉与另一分子的结合物例如可包含has的末端醛基与另一分子的反应性基团之间的共价连接。
本文中使用的术语“聚烷二醇”或“聚(烷二醇)”是指聚乙二醇(聚(乙二醇))、聚丙二醇、聚丁二醇和其衍生物。术语“聚烷二醇”涵盖线性与分支聚合物,且平均分子量介于0.1kda与100kda之间。其它示范性实施例列于例如商业供应商目录中,例如肖尔沃特公司(shearwater)的目录“生物医学用聚乙二醇和衍生物(polyethyleneglycolandderivativesforbiomedicalapplications)”(2001)。
除非另作规定,否则术语“芳基”意思是指可为单环或者稠合在一起或共价连接的多个环(包括(但不限于)1到3个环)的多不饱和、芳香族烃取代基。术语“杂芳基”是指含有1到4个选自n、o和s的杂原子的芳基(或环),其中氮和硫原子任选经氧化,且氮原子任选经季铵化。杂芳基可经由杂原子与分子剩余部分连接。芳基和杂芳基的非限制性实例包括苯基、1-萘基、2-萘基、4-联苯基、1-吡咯基、2-吡咯基、3-吡咯基、3-吡唑基、2-咪唑基、4-咪唑基、吡嗪基、2-噁唑基、4-噁唑基、2-苯基-4-噁唑基、5-噁唑基、3-异噁唑基、4-异噁唑基、5-异噁唑基、2-噻唑基、4-噻唑基、5-噻唑基、2-呋喃基、3-呋喃基、2-噻吩基、3-噻吩基、2-吡啶基、3-吡啶基、4-吡啶基、2-嘧啶基、4-嘧啶基、5-苯并噻唑基、嘌呤基、2-苯并咪唑基、5-吲哚基、1-异喹啉基、5-异喹啉基、2-喹喔啉基、5-喹喔啉基、3-喹啉基和6-喹啉基。上述每一芳基和杂芳基环系统的取代基都选自下文所述的可接受取代基的群组。
为简洁起见,术语“芳基”当与其它术语(包括(但不限于)芳氧基、芳硫氧基、芳烷基)组合使用时,包括上文所定义的芳基和杂芳基环。因此,术语“芳烷基”拟包括芳基与烷基连接的基团(包括(但不限于)苯甲基、苯乙基、吡啶基甲基等),所述烷基包括碳原子(包括(但不限于)亚甲基)已经由例如氧原子置换的烷基(包括(但不限于)苯氧基甲基、2-吡啶氧基甲基、3-(1-萘氧基)丙基等)。
上述每一术语(包括(但不限于)“烷基”、“杂烷基”、“芳基”和“杂芳基”)都打算包括所述基团的经取代和未取代的形式。下文将提供各类基团的示范性取代基。
烷基和杂烷基(包括常常称为亚烷基、烯基、亚杂烷基、杂烯基、炔基、环烷基、杂环烷基、环烯基和杂环烯基的基团)的取代基可为选自包括(但不限于)以下各基团的多个基团中的一个或一个以上基团:-or'、=o、=nr'、=n-or'、-nr'r"、-sr'、-卤素、-sir'r"r"'、-oc(o)r'、-c(o)r'、-co2r'、-conr'r"、-oc(o)nr'r"、-nr"c(o)r'、-nr'-c(o)nr"r"'、-nr"c(o)2r'、-nr-c(nr'r"r"')=nr""、-nr-c(nr'r")=nr'"、-s(o)r'、-s(o)2r'、-s(o)2nr'r"、-nrso2r'、-cn和-no2,其数量在0到(2m'+1)的范围内,其中m'为所述基团中碳原子的总数。r'、r"、r'"和r""各自独立地指氢、经取代或未取代的杂烷基、经取代或未取代的芳基(包括(但不限于)1到3个卤素取代的芳基)、经取代或未取代的烷基、烷氧基或硫烷氧基,或芳基烷基。当本发明化合物包括一个以上r基团时,例如,各r基团是如各r'、r"、r'"和r""基团当存在超过一个时那样独立地选择。当r'与r"连接到同一氮原子时,其可与所述氮原子组合形成5元、6元或7元环。举例来说,-nr'r"拟包括(但不限于)1-吡咯烷基和4-吗啉基。所属领域技术人员从上文有关取代基的论述将了解到,术语“烷基”打算包括碳原子与除氢基外的基团结合的基团,例如卤烷基(包括(但不限于)-cf3和-ch2cf3)和酰基(包括(但不限于)-c(o)ch3、-c(o)cf3、-c(o)ch2och3等)。
与针对烷基所述的取代基类似,芳基和杂芳基的取代基可变化且选自(但不限于):卤素、-or'、=o、=nr'、=n-or'、-nr'r"、-sr'、-卤素、-sir'r"r"'、-oc(o)r'、-c(o)r'、-co2r'、-conr'r"、-oc(o)nr'r"、-nr"c(o)r'、-nr'-c(o)nr"r"'、-nr"c(o)2r'、-nr-c(nr'r"r"')=nr""、-nr-c(nr'r")=nr'"、-s(o)r'、-s(o)2r'、-s(o)2nr'r"、-nrso2r'、-cn和-no2、-r'、-n3、-ch(ph)2、氟(c1-c4)烷氧基和氟(c1-c4)烷基,其数量在0到芳环系统上开放价态总数的范围内;且其中r'、r"、r'"和r""独立地选自氢、烷基、杂烷基、芳基和杂芳基。当本发明化合物包括一个以上r基团时,例如,r基团是如各r'、r"、r'"和r""基团在存在超过一个时那样各自独立地选择。
本文中使用的术语“经调节的血清半衰期”意思是指经修饰bst的循环半衰期相对于其未修饰形式的正或负改变。血清半衰期是通过在投予bst后于各个时间点时取得血液样本,并测定各样本中所述分子的浓度来测量。血清浓度与时间的相关性允许计算血清半衰期。血清半衰期理想地增加至少约两倍,但例如在其增加可获得令人满意的给药方案或避免有毒作用的情况下,较少的增加将有益。在一些实施例中,所述增加为至少约三倍、至少约五倍或至少约十倍。
本文中使用的术语“经调节的治疗半衰期”意思是指治疗有效量bst的半衰期相对于其未修饰形式的正或负改变。治疗半衰期是通过测量投药后各个时间点时的药物动力学和/或药效特性来测量。治疗半衰期增加可理想地得到特别有益的给药方案、特别有益的总剂量,或避免不合需要的作用。在一些实施例中,治疗半衰期增加是由效力增加、经修饰分子与其目标的结合增加或减少、酶(例如蛋白酶)对所述分子的分解的增加或减少,或未修饰分子的另一参数或作用机制的增加或减少,或者所述分子的受体介导的清除增加或减少引起。
当用于核酸或蛋白质时,术语“分离的”表示,所述核酸或蛋白质不含至少某些与其天然状态缔合的细胞组分,或者已将所述核酸或蛋白质浓缩到大于其在体内或体外制造时的浓度的程度。其可为均质态。分离的物质可为干燥或半干燥状态,或呈溶液形式,包括(但不限于)水溶液。其可为医药组合物的一种组分,所述医药组合物还包含医药学上可接受的载剂和/或赋形剂。纯度和均质性通常是使用分析化学技术,例如聚丙烯酰胺凝胶电泳或高效液相色谱法测定。蛋白质,作为一种在制剂中存在的主要物质,是实质上纯化的。具体说来,分离的基因与侧接所述基因且编码除相关基因外的蛋白质的开放式阅读框分离。术语“纯化的”表示核酸或蛋白质在电泳凝胶中实质上产生一条谱带。具体说来,这一术语意思是指,所述核酸或蛋白质为至少85%纯、至少90%纯、至少95%纯、至少99%纯或更高纯度。
术语“核酸”是指单链或双链形式的脱氧核糖核苷酸、脱氧核糖核苷、核糖核苷或核糖核苷酸,以及其聚合物。除非特别限制,否则这一术语涵盖含有天然核苷酸的已知类似物的核酸,这些核酸的结合特性与参考核酸类似,并且代谢方式也与天然存在的核苷酸类似。除非另作明确限制,否则这一术语也指寡核苷酸类似物,包括pna(肽基核酸(peptidonucleicacid))、反义技术中使用的dna类似物(硫代磷酸酯、氨基磷酸酯等)。除非另作指示,否则特定核酸序列也隐含地涵盖其保守修饰变异体(包括(但不限于)简并密码子取代)和互补序列,以及明确指出的序列。具体来说,可通过产生一个或一个以上所选(或所有)密码子的第三位经混合碱基和/或脱氧肌苷残基取代的序列来实现简并密码子取代(贝赞(batzer)等人,核酸研究(nucleicacidres.)19:5081(1991);大冢(ohtsuka)等人,生物化学杂志(j.biol.chem.)260:2605-2608(1985);罗斯里尼(rossolini)等人,分子与细胞探针(mol.cell.probes)8:91-98(1994))。
术语“多肽”、“肽”和“蛋白质”在本文中可互换使用,指的是氨基酸残基的聚合物。也就是说,针对多肽的描述同样适用于肽的描述和蛋白质的描述,反之亦然。这些术语适用于天然存在氨基酸的聚合物,以及一个或一个以上氨基酸残基为非天然编码氨基酸的氨基酸聚合物。本文中使用的这些术语涵盖任何长度的氨基酸链,包括全长蛋白质,其中氨基酸残基是借助于共价肽键连接。
术语“氨基酸”是指天然存在和非天然存在的氨基酸,以及作用方式与天然存在氨基酸类似的氨基酸类似物和氨基酸模拟物。天然编码氨基酸为20种常见氨基酸(丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和缬氨酸)以及吡咯赖氨酸和硒代半胱氨酸。氨基酸类似物是指基础化学结构与天然存在氨基酸相同的化合物,即,α碳与氢、羧基、氨基和r基团结合,例如高丝氨酸、正亮氨酸、甲硫氨酸亚砜、甲硫氨酸甲基锍。这些类似物具有经修饰的r基团(例如,正亮氨酸)或经修饰的肽主链,但仍保持与天然存在氨基酸相同的基础化学结构。提到的氨基酸包括例如天然存在的成蛋白质性l-氨基酸;d-氨基酸,经化学修饰的氨基酸,例如氨基酸变异体和衍生物;天然存在的非成蛋白质性氨基酸,例如β-丙氨酸、鸟氨酸等;以及具有此项技术中已知为氨基酸的特征的特性的化学合成的化合物。非天然存在氨基酸的实例包括(但不限于)α-甲基氨基酸(例如α-甲基丙氨酸)、d-氨基酸、组氨酸样氨基酸(例如2-氨基-组氨酸、β-羟基-组氨酸、高组氨酸(homohistidine)、α-氟甲基-组氨酸和α-甲基-组氨酸)、侧链中具有其它亚甲基的氨基酸(“高”氨基酸),以及侧链中的羧酸官能团经磺酸基置换的氨基酸(例如磺基丙氨酸)。宜以多种不同方式将非天然氨基酸(包括合成非天然氨基酸、经取代氨基酸,或者一个或一个以上d-氨基酸)并入本发明蛋白质中。含d-氨基酸的肽等在体外或体内展现出比含l-氨基酸的对应物高的稳定性。因此,当需要或必需较高胞内稳定性时,构建并入了d-氨基酸的肽等将特别有用。更具体地说,d-肽等对内源肽酶和蛋白酶具有抗性,由此在需要这些特性时,改进在体内分子的生物利用率并延长寿命。另外,d-肽等无法有效加工以使ii类主要组织相容性复合物限制性呈递到辅助t细胞,因此,其不太可能诱导整个生物体的体液免疫反应。
氨基酸可在本文中以其通常已知的三字母符号或iupac-iub生物化学命名委员会(biochemicalnomenclaturecommission)所推荐的单字母符号提及。同样,核苷酸也可以用其通常认可的单字母代码提及。
“保守修饰变异体”适用于氨基酸和核酸序列。对于特定核酸序列,“保守修饰变异体”是指编码一致或基本上一致的氨基酸序列的核酸,或者当核酸不编码氨基酸序列时,也指基本上一致的序列。由于遗传密码具有简并性,故大量功能一致的核酸编码任何指定蛋白质。举例来说,密码子gca、gcc、gcg和gcu都编码氨基酸丙氨酸。因此,在密码子指定丙氨酸的每一位置,在不改变编码多肽的情况下,所述密码子可变成任一所述的相应密码子。此类核酸变异为“沉默变异(silentvariation)”,是保守修饰变异的一种。本文中编码多肽的每一核酸序列也描述这一核酸的每一种可能的沉默变异。所属领域技术人员将认识到,核酸中的每一密码子(通常为甲硫氨酸的唯一密码子的aug,和通常为色氨酸的唯一密码子的tgg除外)可经修饰而得到功能一致的分子。因此,编码多肽的核酸的每一沉默变异是每一所述序列中所固有的。
对于氨基酸序列,所属领域技术人员将认识到,改变、添加或缺失编码序列中的单一氨基酸或小百分比氨基酸的核酸、肽、多肽或蛋白质序列的个别取代、缺失或添加为“保守修饰变异”,其中所述改变引起氨基酸的缺失、氨基酸的添加或者化学上类似的氨基酸对某一氨基酸的取代。所属领域技术人员众所周知提供功能类似的氨基酸的保守取代表。所述保守修饰变异体除为多形态变异体外且不排除多形态变异体,还为本发明的种间同系物和等位基因。
所属领域技术人员已知可提供功能相似的氨基酸的保守取代表。以下八个组各自含有彼此互为保守取代的氨基酸:
1)丙氨酸(a)、甘氨酸(g);
2)天冬氨酸(d)、谷氨酸(e);
3)天冬酰胺(n)、谷氨酰胺(q);
4)精氨酸(r)、赖氨酸(k);
5)异亮氨酸(i)、亮氨酸(l)、甲硫氨酸(m)、缬氨酸(v);
6)苯丙氨酸(f)、酪氨酸(y)、色氨酸(w);
7)丝氨酸(s)、苏氨酸(t);和
8)半胱氨酸(c)、甲硫氨酸(m),
(例如参见科雷敦(creighton),蛋白质结构和分子特性(proteins:structuresandmolecularproperties)(wh弗雷曼出版社(whfreeman&co.);第2版(1993年12月))。
在两个或两个以上核酸或多肽序列的情况下,术语“一致”或“一致性”百分比是指两个或两个以上序列或子序列相同。当使用以下序列比较算法(或所属领域技术人员可用的其它算法)中的一者或通过手工比对和目测,在比较窗或指定测量的区域上比较和比对最大对应性时,如果各序列具有一定百分比的相同氨基酸残基或核苷酸(即,在指定区域内约60%一致、约65%、约70%、约75%、约80%、约85%、约90%或约95%一致),那么所述序列“实质上一致”。此定义也指测试序列的互补性。一致性可存在于至少约50个氨基酸或核苷酸长的区域,或75到100个氨基酸或核苷酸长的区域,或者(未指定时)跨整个聚核苷酸或多肽序列。获得编码本发明多肽(包括来自除人类外的物种的同系物)的聚核苷酸的方法可包含以下步骤:在严格杂交条件下,用具有本发明聚核苷酸序列或其片段的经标记探针筛选文库,以及分离含有所述聚核苷酸序列的全长cdna和基因组克隆。所属领域技术人员众所周知所述杂交技术。
对于序列比较,通常一个序列充当参考序列,用以与测试序列相比较。当使用序列比较算法时,将测试序列和参考序列输入计算机内,必要时指定子序列坐标,并指定序列算法程序参数。可使用默认程序参数,或者可指定替代参数。随后,序列比较算法将基于这些程序参数计算测试序列相对于参考序列的序列一致性百分比。
本文中使用的“比较窗”包括提及具有选自由20到600,通常约50到约200,更通常约100到约150组成的群组的邻近位置数量中的任一者的区段,其中在最佳比对序列与邻近位置数量相同的参考序列后可将这两个序列相比较。所属领域技术人员众所周知供比较的序列比对方法。可通过(包括(但不限于))史密斯(smith)和沃特曼(waterman)(1970)应用数学进展(adv.appl.math.)2:482c的局部同源性算法;尼德尔曼(needleman)和伍思奇(wunsch)(1970)分子生物学杂志(j.mol.biol.)48:443的同源性比对算法;皮尔森(pearson)和利普曼(lipman)(1988)美国国家科学院院刊,85:2444的相似性搜索方法;计算机实施这些算法(wisconsingenetics软件包中的gap、bestfit、fasta和tfasta,遗传学计算机组(geneticscomputergroup),575位理学博士,威斯康星州麦迪逊(madison,wi));或者人工比对和目测(例如参见奥苏贝尔(ausubel)等人,现代分子生物学技术(currentprotocolsinmolecularbiology)(1995增补版)),来进行供比较的序列的最佳比对。
适用于测定序列一致性和序列相似性百分比的算法的一个实例为blast和blast2.0算法,其分别描述于奥特查尔(altschul)等人(1997)核酸研究,25:3389-3402;和奥特查尔等人(1990)分子生物学杂志(j.mol.biol.)215:403-410中。执行blast分析的软件可通过万维网ncbi.nlm.nih.gov可见的美国生物技术信息中心(nationalcenterforbiotechnologyinformation)公开可用。blast算法参数w、t和x决定比对的灵敏性和速度。blastn程序(用于核苷酸序列)使用如下默认参数:字长(w)为11,期望值(e)为10,m=5,n=-4,和比较两条链。对于氨基酸序列,blastp程序使用如下默认参数:字长为3且期望值(e)为10,和blosum62计分矩阵(参见赫尼霍夫(henikoff)和赫尼霍夫(1992)美国国家科学院院刊,89:10915),比对值(b)为50,期望值(e)为10,m=5,n=-4,以及比较两条链。在执行blast算法时通常关闭“低复杂度”过滤器。
blast算法还执行两个序列之间相似性的统计分析(例如参见卡尔琳(karlin)和奥特查尔(altschul)(1993)美国国家科学院院刊,90:5873-5787)。blast算法所提供的一种相似性测量法为最小和概率(smallestsumprobability,p(n)),其可指示两个核苷酸或氨基酸序列之间偶然出现的匹配的概率。举例来说,如果在测试核酸与参考核酸的比较中,最小和概率小于约.2,或小于约0.01,或小于约0.001,那么认为所述核酸与参考序列类似。
短语“与……选择性(或特异性)杂交”是指当特定核苷酸序列存在于复杂混合物(包括(但不限于)全细胞或者文库dna或rna)中时,分子只与所述序列在严格杂交条件下结合、形成双链体或杂交。
短语“严格杂交条件”是指dna、rna、pna或其它核酸模拟物或其组合的序列在此项技术中已知的低离子强度和高温条件下杂交。通常,在严格条件下,探针将与其在复杂核酸混合物(包括(但不限于)全细胞或者dna或rna文库)中的目标子序列杂交,而不与这一复杂混合物中的其它序列杂交。严格条件与序列相关,并且会随不同环境而不同。较长的序列在较高温度下特异性杂交。有关核酸杂交的详尽指导可见于迪杰森(tijssen),生物化学和分子生物学实验技术--核酸探针杂交(laboratorytechniquesinbiochemistryandmolecularbiology--hybridizationwithnucleicprobes),“杂交原理和核酸分析策略综述(overviewofprinciplesofhybridizationandthestrategyofnucleicacidassays)”(1993)中。通常,在指定离子强度ph值下,选择的严格条件比特定序列的热熔点(tm)低约5-10℃。tm为平衡时有50%与目标互补的探针与目标序列杂交的温度(在指定离子强度、ph值和核酸浓度下)(当目标序列过量存在时,在tm下,平衡时有50%探针被占据)。严格条件可为在ph7.0到8.3下盐浓度小于约1.0m钠离子,通常为约0.01到1.0m钠离子浓度(或其它盐)且温度对于短探针(包括(但不限于)10到50个核苷酸)为至少约30℃且对于长探针(包括(但不限于)大于50个核苷酸)为至少约60℃的条件。严格条件也可通过添加例如甲酰胺等去稳定剂来实现。对于选择性或特异性杂交,正信号可为背景杂交的至少两倍,任选为背景杂交的10倍。示范性严格杂交条件如下:50%甲酰胺、5×ssc和1%sds,在42℃下培育;或5×ssc、1%sds,在65℃下培育,且在65℃下于0.2×ssc和0.1%sds中洗涤。所述洗涤可执行5、15、30、60、120分钟或更长时间。
本文中使用的术语“真核生物”是指属于真核系统发生域的生物体,例如动物(包括(但不限于)哺乳动物、昆虫、爬行动物、鸟类等)、纤毛虫、植物(包括(但不限于)单子叶植物、双子叶植物、藻类等)、真菌、酵母、鞭毛虫、小孢子虫、原生生物等。
本文中使用的术语“非真核生物”是指非真核生物体。举例来说,非真核生物体可属于真菌系统发生域,包括(但不限于)大肠杆菌(escherichiacoli)、极端嗜热细菌(thermusthermophilics)、嗜热脂肪芽孢杆菌(bacillusstearothermophilus)、荧光假单胞杆菌(pseudomonasfluoresceins)、铜绿假单胞菌(pseudomonasaeruginosa)、恶臭假单胞菌(pseudomonasputida)等;或古菌系统发生域,包括(但不限于)詹氏甲烷球菌(methanococcusjannaschii)、嗜热自养甲烷杆菌(methanobacteriumthermoautotrophicum)、嗜盐菌(halobacterium)(例如沃氏嗜盐富饶菌(haloferaxvolcanii)和嗜盐菌属nrc-1(halobacteriumspeciesnrc-1))、超嗜热古菌(archaeoglobusfulgidus)、强烈嗜热球菌(pyrococcusfuriosus)、堀越火球菌(pyrococcushorikoshii)、嗜热泉生古细菌(aeuropyrumpernix)等。
本文中使用的术语“个体”是指作为治疗、观察或实验的对象的动物,在一些实施例中为哺乳动物,而在其它实施例中为人类。动物可为伴侣动物(例如狗、猫等)、家畜(例如奶牛、绵羊、猪、马等)或实验动物(例如大鼠、小鼠、豚鼠等)。
本文中使用的术语“有效量”是指所投予的将在一定程度上缓解所治疗的疾病、病状或病症的一种或一种以上症状的经修饰非天然氨基酸多肽的量。可投予含有本文中所述的经修饰非天然氨基酸多肽的组合物,以实现预防、增强和/或治疗性治疗。
术语“增强”意思是指增加或延长所需作用的效力或持续时间。因此,就增强治疗剂的作用来说,术语“增强”是指增加或延长其它治疗剂对系统的作用效力或持续时间的能力。本文中使用的“增强有效量”是指足以增强所需系统中另一治疗剂的作用的量。当用于动物时,适于此用途的有效量将取决于疾病、病症或病状的严重程度和病程、先前疗法、动物的健康状态和对药物的反应,以及治疗兽医的判断。
本文中使用的术语“经修饰”是指对指定多肽进行的任何改变,例如多肽长度、氨基酸序列、化学结构的改变;多肽的共翻译修饰或翻译后修饰。“(经修饰)”形式的术语意思是指,所论述的多肽任选经修饰,也就是说,所论述的多肽可经修饰或不经修饰。
术语“翻译后修饰”是指天然或非天然氨基酸在并入多肽链后发生的对所述氨基酸的任何修饰。举个例子,这一术语涵盖共翻译体内修饰、共翻译体外修饰(例如,在无细胞翻译系统中)、翻译后体内修饰和翻译后体外修饰。
在预防性应用中,将含有bst多肽的组合物投予易患特定疾病、病症或病状或者另外有患特定疾病、病症或病状风险的动物。其量定义为“预防有效量”。在此使用中,确切量也取决于动物的健康状态、体重等。认为通过常规实验(例如,剂量递增临床实验)确定所述预防有效量完全在所属领域的技能范围内。
术语“经保护”是指存在防止化学反应性官能团在某些反应条件下反应的“保护基”或部分。保护基将视所保护的化学反应性基团的类型而变化。举例来说,如果化学反应性基团为胺或酰肼,那么保护基可选自叔丁氧羰基(t-boc)和9-芴基甲氧基羰基(fmoc)的群组。如果化学反应性基团为硫醇,那么保护基可为邻吡啶基二硫化物。如果化学反应性基团为羧酸(例如丁酸或丙酸)或羟基,那么保护基可为苯甲基或烷基(例如甲基、乙基或叔丁基)。也可将此项技术中已知的其它保护基用于本文中所述的方法和组合物中,包括光不稳定性基团,例如nvoc和menvoc。此项技术中已知的其它保护基也可用于本文中所述的方法和组合物中。
仅举例来说,封端/保护基可选自:
其它保护基描述于格里尼(greene)和伍兹(wuts),有机合成中的保护基(protectivegroupsinorganicsynthesis),第3版,约翰威立父子出版公司(johnwiley&sons),纽约州纽约(newyork,ny),1999中,所述文献的全文以引用的方式并入本文中。
在治疗性应用中,将含有经修饰非天然氨基酸多肽的组合物以足以治愈疾病、病症或病状或者至少部分抑制疾病、病症或病状的症状的量投予已罹患所述疾病、病症或病状的动物。所述量定义为“治疗有效量”,且将取决于疾病、病症或病状的严重程度和病程、先前疗法、动物的健康状况和对药物的反应,以及治疗兽医的判断。认为通过常规实验(例如,剂量递增临床实验)确定所述治疗有效量完全在所属领域的技能范围内。
术语“治疗”用于指预防和/或治疗性治疗。
本文所提供的非天然编码氨基酸多肽可包括同位素标记的化合物,在这些化合物中,一个或一个以上原子经原子质量或质量数与自然界常见的原子质量或质量数不同的原子置换。可并入本发明的化合物中的同位素的实例包括氢、碳、氮、氧、氟和氯的同位素,分别例如2h、3h、13c、14c、15n、18o、17o、35s、18f、36cl。本文中所述的某些经同位素标记的化合物,例如并有如3h和14c等放射性同位素的化合物,可用于药物和/或基质组织分布分析中。此外,用同位素(例如氘,即2h)取代可能因代谢稳定性较高(例如,体内半衰期增加或剂量需求降低)而提供某些治疗优势。
所有异构体,包括(但不限于)非对映异构体、对映异构体和其混合物,都可视为本文中所述组合物的一部分。在额外或其它实施例中,在投予有需要的生物体非天然编码氨基酸多肽后,这些多肽代谢产生代谢物,随后使用这些代谢物产生所需作用,包括所需治疗作用。其它或额外实施例为非天然编码氨基酸多肽的活性代谢物。
在一些情形中,非天然编码氨基酸多肽可以互变异构体形式存在。此外,本文中所述的非天然编码氨基酸多肽可以未溶剂化形式以及与医药学上可接受的溶剂(例如水、乙醇等)形成的溶剂化形式存在。本文中也揭示所述溶剂化形式。所属领域技术人员将认识到,本文中的一些化合物可以数种互变异构体形式存在。所有这些互变异构形式都可视为本文中所述组合物的一部分。
除非另作指示,否则在此项技术的技术领域内的质谱、nmr、hplc、蛋白质化学、生物化学、重组dna技术和药理学的常规方法都可使用。
具体实施方式
i.引言
本发明中提供包含至少一个非天然氨基酸的b-gcsf分子。在本发明某些实施例中,具有至少一个非天然氨基酸的b-gcsf多肽包括至少一个翻译后修饰。在一个实施例中,所述至少一个翻译后修饰包含利用所属领域技术人员已知适于特定反应性基团的化学方法使包含第二反应性基团的分子与包含第一反应性基团的至少一个非天然氨基酸连接,所述分子包括(但不限于)羟烷基淀粉(has)、羟乙基淀粉(hes)、标记、染料、聚合物、水溶性聚合物、聚乙二醇衍生物、光交联剂、放射性核素、细胞毒性化合物、药物、亲和标记、光亲和标记、反应性化合物、树脂、第二蛋白质或多肽或者多肽类似物、抗体或抗体片段、金属螯合剂、辅因子、脂肪酸、碳水化合物、聚核苷酸、dna、rna、反义聚核苷酸、糖、水溶性树突状聚合物、环糊精、抑制性核糖核酸、生物材料、纳米粒子、自旋标记、荧光团、含金属部分、放射性部分、新颖官能团、与其它分子共价或非共价相互作用的基团、光笼蔽部分(photocagedmoiety)、光化辐射可激发部分、可光异构化部分、生物素、生物素衍生物、生物素类似物、并有重原子的部分、可化学裂解基团、可光裂解基团、延长的侧链、碳连接糖、氧化还原活性剂、氨基硫代酸、有毒部分、同位素标记的部分、生物物理探针、磷光基团、化学发光基团、电子致密基团、磁性基团、插入基团、发色团、能量转移剂、生物活性剂、可检测标记、小分子、量子点、纳米递质、放射性核苷酸、放射性递质、中子捕获剂,或上述物质的任何组合,或任何其它所需化合物或物质。举例来说,第一反应性基团为炔基部分(包括(但不限于)在非天然氨基酸对炔丙氧基苯丙氨酸中,其中所述炔丙基有时也称为乙炔部分),且第二反应性基团为叠氮基部分,并利用[3+2]环加成化学方法。在另一实例中,第一反应性基团为叠氮基部分(包括(但不限于)在非天然氨基酸对叠氮基-l-苯丙氨酸中),且第二反应性基团为炔基部分。在本发明经修饰b-gcsf多肽的某些实施例中,使用至少一个包含至少一个翻译后修饰的非天然氨基酸(包括(但不限于)含有酮基官能团的非天然氨基酸),其中所述至少一个翻译后修饰包含糖部分。在某些实施例中,翻译后修饰是在真核细胞或非真核细胞中于体内进行。连接子、聚合物、水溶性聚合物或其它分子可将所述分子与多肽连接。所述分子可与多肽直接连接。
在某些实施例中,蛋白质包括至少一个由一个宿主细胞在体内进行的翻译后修饰,其中所述翻译后修饰通常不是由另一宿主细胞类型进行。在某些实施例中,蛋白质包括至少一个由真核细胞在体内进行的翻译后修饰,其中所述翻译后修饰通常不是由非真核细胞进行。翻译后修饰的实例包括(但不限于)糖基化、乙酰化、酰基化、脂质修饰、棕榈酰化、棕榈酸盐加成、磷酸化、糖脂键修饰等。
在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸用于多肽的糖基化、乙酰化、酰基化、脂质修饰、棕榈酰化、棕榈酸盐加成、磷酸化或糖脂键修饰。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸用于多肽的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上天然编码氨基酸用于多肽的糖基化、乙酰化、酰基化、脂质修饰、棕榈酰化、棕榈酸盐加成、磷酸化或糖脂键修饰。在一些实施例中,b-gcsf多肽包含一个或一个以上天然编码氨基酸用于多肽的糖基化。
在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上缺失,用以增进多肽的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽中不同氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上缺失,用以增进多肽中不同氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽中非天然编码氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽中天然编码氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上天然编码氨基酸的添加和/或取代,用以增进多肽中不同氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽中天然编码氨基酸的糖基化。在一些实施例中,b-gcsf多肽包含一个或一个以上非天然编码氨基酸的添加和/或取代,用以增进多肽中非天然编码氨基酸的糖基化。
在一个实施例中,翻译后修饰包含通过glcnac-天冬酰胺键将寡糖与天冬酰胺连接(包括(但不限于)其中所述寡糖包含(glcnac-man)2-man-glcnac-glcnac等)。在另一实施例中,翻译后修饰包含通过galnac-丝氨酸、galnac-苏氨酸、glcnac-丝氨酸或glcnac-苏氨酸键将寡糖(包括(但不限于)gal-galnac、gal-glcnac等)与丝氨酸或苏氨酸连接。在某些实施例中,本发明蛋白质或多肽可包含分泌或定位序列、表位标签、flag标签、聚组氨酸标签、gst融合体等。分泌信号序列的实例包括(但不限于)原核分泌信号序列、真核分泌信号序列、针对细菌表达而5'-优化的真核分泌信号序列、新颖分泌信号序列、果胶酸裂解酶分泌信号序列、ompa分泌信号序列和噬菌体分泌信号序列。分泌信号序列的实例包括(但不限于)stii(原核)、fdgiii和m13(噬菌体)、bgl2(酵母)和来源于转座子的信号序列bla。任何所述序列都可进行修饰以使多肽具有所需的结果,包括(但不限于)用一个信号序列取代不同的信号序列;用一个前导序列取代不同的前导序列等。
相关蛋白质或多肽可含有至少1个、至少2个、至少3个、至少4个、至少5个、至少6个、至少7个、至少8个、至少9个或者10个或更多个非天然氨基酸。这些非天然氨基酸可相同或不同,例如,蛋白质中的1、2、3、4、5、6、7、8、9、10个或更多不同位点可包含1、2、3、4、5、6、7、8、9、10个或更多不同的非天然氨基酸。在某些实施例中,天然存在的蛋白质型式中存在的至少一个(但少于全部)特定氨基酸经非天然氨基酸取代。
本发明提供基于包含至少一个非天然编码氨基酸的b-gcsf的方法和组合物。将至少一个非天然编码氨基酸引入b-gcsf中,可允许应用结合化学,其涉及(包括(但不限于))与一个或一个以上非天然编码氨基酸发生特定化学反应,而不与通常存在的20种氨基酸反应。在一些实施例中,包含非天然编码氨基酸的b-gcsf经由非天然编码氨基酸的侧链与水溶性聚合物(例如聚乙二醇(peg))连接。本发明提供一种利用peg衍生物选择性修饰蛋白质的高效方法,其涉及对选择密码子反应而在蛋白质中选择性并入非基因编码氨基酸,包括(但不限于)含有未见于20种天然并入的氨基酸中的官能团或取代基(包括(但不限于)酮基、叠氮或乙炔部分)的氨基酸,随后利用适当的反应性peg衍生物修饰所述氨基酸。并入后,随后可利用所属领域技术人员已知适于非天然编码氨基酸中存在的特定官能团或取代基的化学方法来修饰氨基酸侧链。多种已知的化学方法都适用于本发明中,用以将水溶性聚合物并入蛋白质中。所述方法包括(但不限于)分别利用(但不限于)乙炔或叠氮化物衍生物的胡伊斯根[3+2]环加成反应(例如参见帕德瓦a.(padwa,a.)有机合成大全(comprehensiveorganicsynthesis),第4卷.(1991)托斯特b.m.(trost,b.m.)编,派格蒙出版社(pergamon),牛津(oxford),第1069-1109页;和胡伊斯根r.(huisgen,r.)1,3-偶极环加成化学(1,3-dipolarcycloadditionchemistry),(1984),帕德瓦a.编,威立出版公司(wiley),纽约,第1-176页)。
由于胡伊斯根[3+2]环加成方法涉及环加成而非亲核取代反应,故可以极高的选择性修饰蛋白质。所述反应可在室温下于水性条件中,通过将催化量的cu(i)盐添加到反应混合物中以优良的区位选择性(1,4>1,5)进行。例如参见托尼诺等人,(2002)有机化学杂志67:3057-3064;和罗斯托瑟夫等人,(2002)德国应用化学41:2596-2599;和wo03/101972。可通过[3+2]环加成反应加到本发明蛋白质中的分子包括具有适当官能团或取代基(包括(但不限于)叠氮基或乙炔衍生物)的实际上任何分子。这些分子可分别加到具有乙炔基(包括(但不限于)对炔丙氧基苯丙氨酸)或叠氮基(包括(但不限于)对叠氮基-苯丙氨酸)的非天然氨基酸中。
由胡伊斯根[3+2]环加成反应产生的5元环在还原环境中通常是不可逆的,且可在水性环境中对水解可稳定较长时间。因此,可在要求的水性条件下,用本发明的活性peg衍生物改变多种物质的物理和化学特征。甚至更重要的是,由于叠氮基和乙炔部分彼此具特异性(并且例如不会与20种常见的基因编码氨基酸中的任一者反应),故可在一个或一个以上特定位点中以极高的选择性修饰蛋白质。
本发明还提供peg衍生物的水溶性和水解稳定性衍生物以及具有一个或一个以上乙炔或叠氮部分的相关亲水聚合物。含有乙炔部分的peg聚合物衍生物以极高选择性与作为对选择密码子的反应而选择性引入蛋白质中的叠氮部分偶合。类似地,含有叠氮部分的peg聚合物衍生物以极高选择性与作为对选择密码子的反应而选择性引入蛋白质中的乙炔部分偶合。
更具体说来,叠氮部分包含(但不限于)烷基叠氮化物、芳基叠氮化物和这些叠氮化物的衍生物。烷基和芳基叠氮化物的衍生物可包括其它取代基,只要保持乙炔特异性反应性即可。乙炔部分包含烷基和芳基乙炔,以及各自的衍生物。烷基和芳基乙炔的衍生物可包括其它取代基,只要保持叠氮基特异性反应性即可。
本发明提供具有多种官能团、取代基或部分的物质与其它物质的结合物,所述其它物质包括(但不限于)羟烷基淀粉(has)、羟乙基淀粉(hes)、标记、染料、聚合物、水溶性聚合物、聚乙二醇衍生物、光交联剂、放射性核素、细胞毒性化合物、药物、亲和标记、光亲和标记、反应性化合物、树脂、第二蛋白质或多肽或多肽类似物、抗体或抗体片段、金属螯合剂、辅因子、脂肪酸、碳水化合物、聚核苷酸、dna、rna、反义聚核苷酸、糖、水溶性树突状聚合物、环糊精、抑制性核糖核酸、生物材料、纳米粒子、自旋标记、荧光团、含金属部分、放射性部分、新颖官能团、与其它分子共价或非共价相互作用的基团、光笼蔽部分、光化辐射可激发部分、可光异构化部分、生物素、生物素衍生物、生物素类似物、并有重原子的部分、可化学裂解基团、可光裂解基团、延长的侧链、碳连接糖、氧化还原活性剂、氨基硫代酸、有毒部分、同位素标记的部分、生物物理探针、磷光基团、化学发光基团、电子致密基团、磁性基团、插入基团、发色团、能量转移剂、生物活性剂、可检测标记、小分子、量子点、纳米递质、放射性核苷酸、放射性递质、中子捕获剂;或上述物质的任何组合,或任何其它所需化合物或物质。本发明还包括具有叠氮或乙炔部分的物质与具有相应乙炔或叠氮部分的peg聚合物衍生物的结合物。举例来说,可在蛋白质中含有带乙炔官能团的非基因编码氨基酸的位置处,将含有叠氮部分的peg聚合物与生物活性分子偶合。使peg与生物活性分子偶合的键包括(但不限于)胡伊斯根[3+2]环加成产物。
此项技术中充分确定,可使用peg修饰生物材料的表面(例如参见美国专利6,610,281;米赫瓦r.(mehvar,r.),药学与药理学杂志(j.pharmpharmsci.),3(1):125-136(2000),以引用的方式并入本文中)。本发明还包括生物材料,所述生物材料包含具有一个或一个以上反应性叠氮基或乙炔位点的表面,和通过胡伊斯根[3+2]环加成键与所述表面偶合的一种或一种以上本发明的含叠氮基或含乙炔聚合物。生物材料和其它物质也可通过除叠氮键或乙炔键外的键(例如通过包含羧酸、胺、醇或硫醇部分的键)与叠氮化物或乙炔活化的聚合物衍生物偶合,由此留下叠氮或乙炔部分用于后续反应。
本发明包括一种合成本发明的含叠氮基和含乙炔聚合物的方法。在含叠氮基peg衍生物的情况下,叠氮基可与聚合物的碳原子直接键接。或者,可将在一端具有叠氮部分的连接剂与常规活化聚合物连接,从而使所得聚合物在其一端具有叠氮部分,以此来制备含叠氮基的peg衍生物。在含乙炔peg衍生物的情况下,乙炔可与聚合物的碳原子直接键接。或者,可将一端具有乙炔部分的连接剂与常规活化聚合物连接,从而使所得聚合物在其一端具有乙炔部分,以此制备含乙炔的peg衍生物。
更具体地说,在含叠氮基peg衍生物的情况下,具有至少一个活性羟基部分的水溶性聚合物发生反应,产生带有更强反应性部分(例如甲磺酸酯基、三氟乙磺酸酯基、甲苯磺酸酯基或卤素离去基团)的经取代聚合物。所属领域技术人员已知含有磺酸卤化物、卤素原子和其它离去基团的peg衍生物的制备和使用。随后,所得经取代聚合物可发生反应,以用聚合物末端的叠氮部分取代所述反应性更强的部分。或者,具有至少一个活性亲核部分或亲电子部分的水溶性聚合物可与一端具有叠氮基的连接剂发生反应,由此在peg聚合物与所述连接剂之间形成共价键,且所述叠氮部分位于聚合物的一端。所属领域技术人员已知亲核部分和亲电子部分,包括胺、硫醇、酰肼、肼、醇、羧酸酯、醛、酮、硫酯等。
更具体点说,在含乙炔peg衍生物的情况下,具有至少一个活性羟基部分的水溶性聚合物可发生反应,以置换含有乙炔部分的前体中的卤素或其它活化的离去基团。或者,具有至少一个活性亲核部分或亲电子部分的水溶性聚合物可与一端具有乙炔的连接剂发生反应,由此在peg聚合物与所述连接剂之间形成共价键,且所述乙炔部分位于聚合物的一端。卤素部分、活化的离去基团、亲核和亲电子部分在有机合成中的使用以及peg衍生物的制备和使用已经得到了此项技术中的从业人员的充分确认。
本发明也提供一种用于选择性修饰蛋白质以将其它物质添加到经修饰蛋白质中的方法,所述其它物质包括(但不限于)水溶性聚合物,例如peg和含有叠氮或乙炔部分的peg衍生物。在生物相容性、稳定性、溶解性和免疫原性缺乏极为重要的情况下,可以使用含叠氮基和含乙炔的peg衍生物改变表面和分子的特性,同时提供一种选择性高于此项技术中先前已知者的将peg衍生物与蛋白质连接的方式。
ii.牛促生长素
本发明的bst多肽可用来改善或预防动物感染。所属领域技术人员已知牛和人g-csf的生物活性以及表征牛和人g-csf的生物活性的分析。所属领域技术人员已知涉及评估中性粒细胞数量和中性粒细胞功能的分析。
iii.用于本发明的一般重组核酸方法
在本发明的多个实施例中,将使用重组方法分离、克隆和通常改变编码相关bst多肽的核酸。这些实施例用于(包括(但不限于))蛋白质表达或产生来源于bst多肽的变异体、衍生物、表达盒或其它序列的过程中。在一些实施例中,编码本发明多肽的序列可操作性地连接到异源启动子。宿主细胞中hg-csf的分离和g-csf的产生描述于例如美国专利第4,810,643号、第4,999,291号、第5,580,755号和第6,716,606号中,各专利以引用的方式并入本文中。
可根据母体多肽的氨基酸序列,包括(但不限于)具有seqidno:1、2中所示的氨基酸序列,合成编码包含非天然编码氨基酸的bst多肽的核苷酸序列,随后改变所述核苷酸序列,从而引入(即,并入或取代)或去除(即,缺失或取代)相关氨基酸残基。宜根据常规方法,利用定点诱变来修饰核苷酸序列。或者,可利用化学合成来制备核苷酸序列,所述化学合成包括(但不限于)使用寡核苷酸合成仪(其中寡核苷酸是根据所需多肽的氨基酸序列来设计)且优选选择产生重组多肽的宿主细胞偏好的那些密码子。举例来说,可通过pcr、接合或接合链式反应来合成和组装数个编码所需多肽多个部分的小寡核苷酸。例如参见布兰妮(barany)等人,美国国家科学院院刊,88:189-193(1991);美国专利6,521,427,都以引用方式并入本文中。
本发明利用重组遗传学领域中的常规技术。揭示本发明的一般使用方法的基础教材包括萨姆布鲁克(sambrook)等人,分子克隆实验手册(molecularcloning,alaboratorymanual)(第3版,2001);克林格(kriegler),基因转移和表达实验手册(genetransferandexpression:alaboratorymanual)(1990);和现代分子生物学技术(currentprotocolsinmolecularbiology)(奥斯贝尔等人编,1994)。
描述分子生物技术的一般教材包括伯格(berger)和吉米尔(kimmel),分子克隆技术指南,酶学方法(guidetomolecularcloningtechniques.methodsinenzymology)第152卷,学术出版公司(academicpress,inc.),加利福尼亚州圣地亚哥(sandiego,ca)(伯格);萨姆布鲁克(sambrook)等人,分子克隆实验手册(第2版)第1-3卷,冷泉港实验室(coldspringharborlaboratory),纽约州冷泉港(coldspringharbor,newyork),1989(“萨姆布鲁克”);和现代分子生物学技术,f.m.奥斯贝尔(f.m.ausubel)等人编,现代技术(currentprotocols),格里尼出版公司(greenepublishingassociates,inc.)与约翰威立父子出版公司的合资公司,(1999增刊)(“奥斯贝尔”)。这些教材描述了诱变、载体的使用、启动子和许多其它相关的主题,所述相关主题涉及(包括(但不限于))包括用于产生包括非天然氨基酸的蛋白质的选择密码子、正交trna、正交合成酶和其对的基因或聚核苷酸的产生。
多类诱变方法可用于本发明中以实现多种目的,包括(但不限于)产生新颖合成酶或trna、使trna分子突变、使编码合成酶的聚核苷酸突变、产生trna文库、产生合成酶文库、产生选择密码子、插入编码相关蛋白质或多肽中的非天然氨基酸的选择密码子。所述诱变方法包括(但不限于)定点诱变、随机点诱变、同源重组、dna改组或其它递归诱变方法、嵌合构建、使用含尿嘧啶的模板的诱变、寡核苷酸定向诱变、硫代磷酸酯修饰dna诱变、使用带缺口双链体dna的诱变等,pcr介导的诱变,或其任何组合。其它适合的方法包括点错配修复、使用修复缺陷型宿主株的诱变、限制性选择和限制性纯化、缺失诱变、通过总基因合成诱变、双链断裂修复等。本发明中还包括(包括(但不限于))涉及嵌合构建体的诱变。在一个实施例中,可利用有关天然存在分子或者改变或突变的天然存在分子的已知信息,包括(但不限于)序列、序列比较、物理特性、二级、三级或四级结构、晶体结构等,来指导诱变。
本文中所见的文章和实例描述这些程序。其它信息可见于文中所引用的以下出版物和参考文献:林(ling)等人,dna诱变方法综述(approachestodnamutagenesis:anoverview),分析生物化学(analbiochem.)254(2):157-178(1997);戴勒(dale)等人,使用硫代磷酸酯法的寡核苷酸定向随机诱变法(oligonucleotide-directedrandommutagenesisusingthephosphorothioatemethod),分子生物学方法(methodsmol.biol.)57:369-374(1996);史密斯(smith),体外诱变(invitromutagenesis),遗传学评述(ann.rev.genet.)19:423-462(1985);博思汀(botstein)和肖特勒(shortle),体外诱变策略和应用(strategiesandapplicationsofinvitromutagenesis),科学229:1193-1201(1985);卡特(carter),定点诱变(site-directedmutagenesis),生物化学杂志(biochem.j.)237:1-7(1986);库克尔(kunkel),寡核苷酸定向诱变效率(theefficiencyofoligonucleotidedirectedmutagenesis),核酸与分子生物学(nucleicacids&molecularbiology)(艾克斯廷f.(eckstein,f.)和李磊d.m.j.(lilley,d.m.j.)编,柏林施普林格出版社(springerverlag,berlin))(1987);库克尔,在无表型选择情况下快速而高效地定点诱变(rapidandefficientsite-specificmutagenesiswithoutphenotypicselection),美国国家科学院院刊82:488-492(1985);库克尔等人,在无表型选择情况下快速而高效地定点诱变,酶学方法(methodsinenzymol.)154,367-382(1987);贝斯(bass)等人,具有新dna结合特异性的突变trp阻遏子(mutanttrprepressorswithnewdna-bindingspecificities);科学242:240-245(1988);佐勒(zoller)和史密斯(smith),使用m13源性载体的寡核苷酸定向诱变:一种在任何dna片段中产生点突变的通用有效程序(oligonucleotide-directedmutagenesisusingm13-derivedvectors:anefficientandgeneralprocedurefortheproductionofpointmutationsinanydnafragment),核酸研究,10:6487-6500(1982);佐勒和史密斯,克隆到m13载体中的dna片段的寡核苷酸定向诱变(oligonucleotide-directedmutagenesisofdnafragmentsclonedintom13vectors),酶学方法(methodsinenzymol.)100:468-500(1983);佐勒和史密斯,寡核苷酸定向诱变:一种使用两个寡核苷酸引物和单链dna模板的简单方法(oligonucleotide-directedmutagenesis:asimplemethodusingtwooligonucleotideprimersandasingle-strandeddnatemplate),酶学方法154:329-350(1987);泰勒(taylor)等人,在限制性酶反应中使用硫代磷酸酯修饰的dna来制备带切口dna(theuseofphosphorothioate-modifieddnainrestrictionenzymereactionstopreparenickeddna),核酸研究13:8749-8764(1985);泰勒等人,使用硫代磷酸酯修饰的dna以高频率快速产生寡核苷酸定向突变(therapidgenerationofoligonucleotide-directedmutationsathighfrequencyusingphosphorothioate-modifieddna),核酸研究13:8765-8785(1985);纳卡玛(nakamaye)和艾克斯廷(eckstein),硫代磷酸酯基团的限制性内切核酸酶ncii裂解抑制以及其在寡核苷酸定向诱变中的应用(inhibitionofrestrictionendonucleasenciicleavagebyphosphorothioategroupsanditsapplicationtooligonucleotide-directedmutagenesis),核酸研究14:9679-9698(1986);塞尔斯(sayers)等人,用于基于硫代磷酸酯的寡核苷酸定向诱变中的5'-3'外切核酸酶(5'-3'exonucleasesinphosphorothioate-basedoligonucleotide-directedmutagenesis),核酸研究16:791-802(1988);塞尔斯等人,含硫代磷酸酯的dna通过在溴化乙锭存在下与限制性内切核酸酶反应来进行链特异性裂解(strandspecificcleavageofphosphorothioate-containingdnabyreactionwithrestrictionendonucleasesinthepresenceofethidiumbromide),(1988)核酸研究16:803-814;克拉莫(kramer)等人,用于构建寡核苷酸定向突变的缺口双链dna方法(thegappedduplexdnaapproachtooligonucleotide-directedmutationconstruction),核酸研究12:9441-9456(1984);克拉莫和菲特兹(fritz),经由带缺口双链dna的寡核苷酸定向突变构建(oligonucleotide-directedconstructionofmutationsviagappedduplexdna),酶学方法154:350-367(1987);克拉莫等人,在寡核苷酸定向突变构建的带缺口双链dna方法中酶促体外反应的改进(improvedenzymaticinvitroreactionsinthegappedduplexdnaapproachtooligonucleotide-directedconstructionofmutations),核酸研究16:7207(1988);菲特兹等人,寡核苷酸定向突变构建:无体外酶促反应的缺口双链dna程序(oligonucleotide-directedconstructionofmutations:agappedduplexdnaprocedurewithoutenzymaticreactionsinvitro),核酸研究16:6987-6999(1988);克拉莫等人,利用大肠杆菌的甲基定向dna错配修复系统以不同效率校正不同的碱基/碱基错配(differentbase/basemismatchesarecorrectedwithdifferentefficienciesbythemethyl-directeddnamismatch-repairsystemofe.coli),细胞(cell)38:879-887(1984);卡特(carter)等人,使用m13载体改进寡核苷酸定点诱变(improvedoligonucleotidesite-directedmutagenesisusingm13vectors),核酸研究13:4431-4443(1985);卡特,使用m13载体改进寡核苷酸定向诱变(improvedoligonucleotide-directedmutagenesisusingm13vectors),酶学方法154:382-403(1987);艾格达乍德赫(eghtedarzadeh)和汉尼科夫(henikoff),使用寡核苷酸产生大缺失(useofoligonucleotidestogeneratelargedeletions),核酸研究14:5115(1986);威尔斯(wells)等人,形成氢键对于稳定枯草杆菌蛋白酶的过渡态的重要性(importanceofhydrogen-bondformationinstabilizingthetransitionstateofsubtilisin),英国皇家化学协会期刊(phil.trans.r.soc.lond.)a317:415-423(1986);纳姆拜耳(nambiar)等人,编码核糖核酸酶s蛋白的基因的总合成和克隆(totalsynthesisandcloningofagenecodingfortheribonucleasesprotein),科学223:1299-1301(1984);萨克玛(sakmar)和克拉那(khorana),牛视杆外节鸟嘌呤核苷酸结合蛋白(转导蛋白)α亚单元的基因的总合成和表达(totalsynthesisandexpressionofageneforthealpha-subunitofbovinerodoutersegmentguaninenucleotide-bindingprotein(transducin)),核酸研究14:6361-6372(1988);威尔斯等人,盒式诱变:一种在指定位点产生多个突变的有效方法(cassettemutagenesis:anefficientmethodforgenerationofmultiplemutationsatdefinedsites),基因(gene)34:315-323(1985);格拉托姆
通常根据布克吉(beaucage)和卡鲁斯(caruthers),四面体通讯(tetrahedronletts.)22(20):1859-1862,(1981)中所述的固相亚磷酰胺三酯法,例如使用尼德尔曼-范德瓦特(needham-vandevanter)等人,核酸研究,12:6159-6168(1984)所述的自动合成仪,以化学方式合成寡核苷酸,例如用于本发明的诱变过程中,例如使合成酶文库突变,或改变trna。
本发明还涉及经由正交trna/rs对在体内并入非天然氨基酸的真核宿主细胞、非真核宿主细胞和生物体。利用本发明的聚核苷酸或包括本发明聚核苷酸的构建体,包括(但不限于)本发明的载体(其可为例如克隆载体或表达载体),对宿主细胞进行基因工程改造(包括(但不限于)转化、转导或转染)。举例来说,将正交trna、正交trna合成酶和待衍生化的蛋白质的编码区可操作性地连接到在所需宿主细胞中起作用的基因表达控制元件。载体可为例如质粒、科斯质粒(cosmid)、噬菌体、细菌、病毒、裸聚核苷酸或结合的聚核苷酸的形式。可通过标准方法将载体引入细胞和/或微生物中,所述方法包括电穿孔(弗洛姆(fromm)等人,美国国家科学院院刊82,5824(1985))、通过病毒载体感染、在小珠粒或粒子基质内或在表面上用具有核酸的小粒子高速弹道渗透(科莱恩(klein)等人,自然327,70-73(1987))等。适于在体外将核酸转移到细胞中的技术包括使用脂质体、显微注射、细胞融合、deae-葡聚糖、磷酸钙沉淀法等。体内基因转移技术包括(但不限于)用病毒(通常为逆转录病毒)载体转染和病毒外壳蛋白-脂质体介导的转染[迪泽(dzau)等人,生物技术趋势(trendsinbiotechnology)11:205-210(1993)]。在一些情况下,可能需要利用靶向目标细胞的试剂,例如对细胞表面膜蛋白或目标细胞具特异性的抗体、目标细胞上的受体的配体等,来提供核酸来源。当使用脂质体时,可使用以胞吞作用缔合的结合于细胞表面膜蛋白的蛋白质以供靶向和/或便利摄取,例如对特定细胞类型具嗜性的衣壳蛋白或其片段、在循环中经历内化的蛋白质的抗体以及靶向胞内定位并增加胞内半衰期的蛋白质。
可在经改变而适用于例如筛选步骤、活化启动子或选择转化株等活动的常规营养培养基中培养经工程改造的宿主细胞。这些细胞可任选培养入转基因生物体中。关于(包括(但不限于))细胞分离和培养(例如用于随后的核酸分离)的其它有用参考文献包括弗莱希利(freshney)(1994)动物细胞培养基础技术手册(cultureofanimalcells,amanualofbasictechnique),第3版,纽约威立-利斯出版公司,及其中引用的参考文献;佩尼(payne)等人(1992)在液体系统中的植物细胞和组织培养(plantcellandtissuecultureinliquidsystems),约翰威立父子公司,纽约州纽约;盖姆伯格(gamborg)和菲利普斯(phillips)(编)(1995)植物细胞、组织和器官培养(plantcell,tissueandorganculture);基本方法施普林格实验手册(fundamentalmethodsspringerlabmanual),施普林格-维拉格(springer-verlag)(柏林,海德堡纽约(berlinheidelbergnewyork),以及奥特拉斯(atlas)和帕克斯(parks)(编)微生物培养基手册(thehandbookofmicrobiologicalmedia)(1993)crc出版社,佛罗里达州伯克莱屯(bocaraton,fl)。
可使用将目标核酸引入细胞中的数种众所周知的方法,其中任一种都可用于本发明中。这些方法包括:受体细胞与含有dna的细菌原生质体融合、电穿孔、基因枪法(projectilebombardment)和病毒载体(下文中进一步论述)感染等。细菌细胞可用于扩增含有本发明的dna构建体的质粒的数目。使细菌生长到对数生长期,并且可通过此项技术中已知的多种方法分离细菌中的质粒(例如参见萨姆布鲁克)。另外,也可购买试剂盒从细菌中纯化质粒(例如参见easypreptm、flexipreptm,都来自法玛西亚生物技术公司(pharmaciabiotech);stratacleantm,来自赛特杰公司(stratagene);和qiapreptm,来自凯杰公司(qiagen))。随后进一步操作分离和纯化的质粒以产生其它质粒,将这些质粒用于转染细胞,或并入相关载体中以感染生物体。典型载体含有转录和翻译终止子、转录和翻译起始序列以及可用于调控特定目标核酸表达的启动子。载体任选包含通用表达盒,其含有至少一个独立终止子序列、允许表达盒在真核细胞或原核细胞或两者中复制的序列(包括(但不限于)穿梭载体),以及用于原核系统与真核系统的选择标记物。载体适于在原核细胞、真核细胞或两者中复制和整合。参见盖里姆(gillam)和史密斯(smith),基因,8:81(1979);罗伯茨(roberts)等人,自然,328:731(1987);斯奇尼德尔e.(schneider,e.)等人,蛋白质表达和纯化(proteinexpr.purif.)6(1):10-14(1995);奥斯贝尔,萨姆布鲁克,伯格(berger)(都同上文)。举例来说,atcc(例如由atcc出版的细菌和噬菌体的atcc目录(theatcccatalogueofbacteriaandbacteriophage)(1992)吉赫纳(gherna)等人(编))提供了可用于克隆的细菌和噬菌体的目录。用于测序、克隆和分子生物学的其它方面的其它基本程序以及基础理论讨论也见于沃特森(watson)等人(1992)重组dna(recombinantdna)第2版,科学美国人(scientificamericanbooks),纽约(ny)。另外,基本上任何核酸(和几乎任何带标记的核酸,无论标准或非标准)都可从多种商业来源中的任一者定制或标准定购,这些商业来源例如米德兰标准试剂公司(midlandcertifiedreagentcompany)(德克萨斯州米德兰(midland,tx),可在万维网mcrc.com上获得)、美国基因公司(thegreatamericangenecompany)(加利福尼亚州雷蒙钠(ramona,ca),可在万维网genco.com上获得)、艾普斯基因公司(expressgeninc.)(伊利诺伊州芝加哥(chicago,il),可在万维网expressgen.com上获得)、欧普隆技术公司(operontechnologiesinc.)(加利福尼亚州阿拉米达(alameda,ca))和许多其它来源。
选择密码子
本发明的选择密码子可扩充蛋白质生物合成机器的遗传密码子框架。举例来说,选择密码子包括(但不限于)独特的三碱基密码子;无义密码子,例如终止密码子,包括(但不限于)琥珀密码子(uag)、赭石密码子或蛋白石密码子(uga);非天然密码子;四个或四个以上碱基的密码子;稀有密码子等。所属领域技术人员易于了解,可引入所需基因或聚核苷酸中的选择密码子的数目范围很广,包括(但不限于)在编码至少一部分bst多肽的单一聚核苷酸中存在一个或一个以上、两个或两个以上、三个或三个以上、4个、5个、6个、7个、8个、9个、10个或更多个选择密码子。
在一个实施例中,所述方法涉及使用作为终止密码子的选择密码子,用于在体内并入一个或一个以上非天然氨基酸。举例来说,产生识别终止密码子(包括(但不限于)uag)的o-trna,并通过带有所需非天然氨基酸的o-rs使其氨酰基化。天然存在的宿主氨酰基trna合成酶不识别这一o-trna。可以使用常规定点诱变在相关多肽中的相关位点引入终止密码子,包括(但不限于)tag。例如参见赛尔斯j.r.(sayers,j.r.)等人,(1988),用于基于硫代磷酸酯的寡核苷酸定向诱变中的5'-3'外切核酸酶(5'-3'exonucleaseinphosphorothioate-basedoligonucleotide-directedmutagenesis.)核酸研究,16:791-802。当在体内组合o-rs、o-trna和编码相关多肽的核酸时,对uag密码子反应而并入非天然氨基酸,从而得到在特定位置含有非天然氨基酸的多肽。
可在不显著干扰真核宿主细胞的情况下在体内并入非天然氨基酸。举例来说,由于对于uag密码子的抑制效率取决于o-trna(包括(但不限于)琥珀抑制trna)与真核释放因子(包括(但不限于)erf)(其结合于终止密码子且启动核糖体释放生长肽)之间的竞争,故可通过(包括(但不限于))增加o-trna和/或抑制trna的表达水平来调节抑制效率。
也可用稀有密码子编码非天然氨基酸。举例来说,当降低体外蛋白质合成反应中的精氨酸浓度时,已证实,稀有精氨酸密码子agg可有效地利用经丙氨酸酰基化的合成trna插入ala。例如参见马(ma)等人,生物化学(biochemistry.)32:7939(1993)。在此情况下,合成trna将与作为大肠杆菌中的次要物质存在的天然存在trnaarg竞争。一些生物体不使用所有三联体密码子。已经利用藤黄微球菌(micrococcusluteus)中的未指定密码子aga在体外转录/翻译提取物中插入氨基酸。例如参见科瓦尔(kowal)和奥利弗(oliver),核酸研究25:4685(1997)。可产生本发明的组分以在体内使用这些稀有密码子。
选择密码子还包含延长密码子,包括(但不限于)四个或四个以上碱基的密码子,例如四个、五个、六个或六个以上碱基的密码子。四碱基密码子的实例包括(但不限于)agga、cuag、uaga、cccu等。五碱基密码子的实例包括(但不限于)aggac、ccccu、cccuc、cuaga、cuacu、uaggc等。本发明的特征包括使用基于移码抑制的延长密码子。四个或四个以上碱基的密码子可将(包括(但不限于))一个或多个非天然氨基酸插入同一蛋白质中。举例来说,在突变o-trna(包括(但不限于)具有反密码子环(例如,具有至少8-10个核苷酸的反密码子环)的特定移码抑制trna)存在下,四个或四个以上碱基的密码子被读成单一氨基酸。在其它实施例中,反密码子环可解码(包括(但不限于))至少一个四碱基密码子、至少一个五碱基密码子或至少一个六碱基密码子或更多碱基的密码子。由于存在256个可能的四碱基密码子,故可在同一细胞中使用四个或四个以上碱基的密码子编码多个非天然氨基酸。参见安德森(anderson)等人,(2002)探究密码子和反密码子尺寸的极限(exploringthelimitsofcodonandanticodonsize),化学与生物学(chemistryandbiology.)9:237-244;麦格赖瑞(magliery),(2001)扩充遗传密码:大肠杆菌中四碱基密码子的有效抑制剂的选择和利用文库方法鉴别“移码”四碱基密码子(expandingthegeneticcode:selectionofefficientsuppressorsoffour-basecodonsandidentificationof"shifty"four-basecodonswithalibraryapproachinescherichiacoli.)分子生物学杂志(j.mol.biol.)307:755-769。
举例来说,已使用四碱基密码子,使用体外生物合成方法将非天然氨基酸并入蛋白质中。例如参见马等人,(1993)生物化学,32:7939;和阳司(hohsaka)等人,(1999)美国化学协会杂志,121:34。cggg和aggu用于在体外利用两种以化学方式酰基化的移码抑制trna将2-萘基丙氨酸和赖氨酸的nbd衍生物同时并入抗生物素蛋白链菌素中。例如参见阳司等人,(1999)美国化学协会杂志,121:12194。在体内研究中,莫尔(moore)等人检查了具有ncua反密码子的trnaleu衍生物抑制uagn密码子(n可为u、a、g或c)的能力,并且发现四联体uaga可以由具有ucua反密码子的trnaleu以13%到26%的效率解码,其中在0或-1框内极少解码。参见莫尔等人,(2000)分子生物学杂志,298:195。在一个实施例中,可将基于稀有密码子或无义密码子的延长密码子用于本发明中,这样可减少错义通读和在其它不想要的位点的移码抑制。
对于给定系统,选择密码子也可包括天然三碱基密码子中的一种,其中内源系统不使用(或很少使用)天然碱基密码子。举例来说,这包括缺乏识别天然三碱基密码子的trna的系统和/或三碱基密码子为稀有密码子的系统。
选择密码子任选包括非天然碱基对。这些非天然碱基对进一步扩充现有的遗传代码。一个额外的碱基对使三联体密码子的数量从64增加到125。第三碱基对的特性包括稳定且具选择性的碱基配对、利用聚合酶以高保真度有效酶促并入dna中,以及在合成新的非天然碱基对之后有效的持续引物延伸。适用于各方法和组合物的非天然碱基对的描述包括例如海奥(hirao)等人,(2002)用于将氨基酸类似物并入蛋白中的非天然碱基对(anunnaturalbasepairforincorporatingaminoacidanaloguesintoprotein),自然生物技术(naturebiotechnology),20:177-182。还参见吴y.(wu,y.)等人(2002)美国化学协会杂志124:14626-14630。其它相关出版物列于下文中。
对于体内使用,非天然核苷可透过膜,并且经磷酸化形成相应的三磷酸酯。此外,增加的遗传信息也很稳定,不会被细胞酶破坏。本纳(benner)和其它人先前的尝试利用了不同于标准沃特森-克里克(watson-crick)对的氢键模式,其中最值得关注的实例为iso-c:iso-g对。例如参见斯维泽(switzer)等人,(1989)美国化学协会杂志,111:8322;和皮克瑞利(piccirilli)等人,(1990)自然,343:33;库尔(kool),(2000)现代化学生物学观点(curr.opin.chem.biol.),4:602。一般说来,这些碱基在某种程度上与天然碱基错配,并且无法酶促复制。库尔和同事证实,碱基之间的疏水堆积相互作用可替代氢键来驱动碱基对的形成。参见库尔,(2000)现代化学生物学观点,4:602;和库克安(guckian)以及库尔,(1998)德国应用化学杂志(英文版)(angew.chem.int.ed.engl.),36,2825。在致力于开发满足所有上述需求的非天然碱基对的过程中,查鲁兹(schultz)、罗姆斯伯格(romesberg)和同事已经系统地合成和研究了一系列非天然疏水性碱基。发现pics:pics自身对比天然碱基对稳定,并且能够利用大肠杆菌dna聚合酶i的克列诺(klenow)片段(kf)有效地并入dna中。例如参见麦克米恩(mcminn)等人,(1999)美国化学协会杂志,121:11585-6;以及小川(ogawa)等人,(2000)美国化学协会杂志,122:3274。3mn:3mn自身对可通过kf以足以获得生物功能的效率和选择性合成。例如参见小川等人,(2000)美国化学协会杂志,122:8803。然而,这两种碱基都充当进一步复制的链终止子。近来已开发出可用于复制pics自身对的突变dna聚合酶。此外,还可复制7ai自身对。例如参见泰厄(tae)等人,(2001)美国化学协会杂志,123:7439。也已开发出新颖的金属碱基对dipic:py,其在结合cu(ii)后形成稳定对。参见麦格斯(meggers)等人,(2000)美国化学协会杂志,122:10714。由于延长密码子和非天然密码子固有地与天然密码子正交,故本发明的方法可利用这一特性产生其正交trna。
翻译旁路系统(translationalbypassingsystem)也可用于在所需多肽中并入非天然氨基酸。在翻译旁路系统中,将大序列并入基因中,但并不翻译成蛋白质。所述序列含有的结构可充当诱导核糖体越过所述序列并继续在插入序列下游翻译的提示(cue)。
在某些实施例中,本发明方法和/或组合物中的相关蛋白质或多肽(或其部分)是由核酸编码。通常,核酸包含至少一个选择密码子、至少两个选择密码子、至少三个选择密码子、至少四个选择密码子、至少五个选择密码子、至少六个选择密码子、至少七个选择密码子、至少八个选择密码子、至少九个选择密码子、十个或更多个选择密码子。
可使用所属领域技术人员众所周知且本文中描述的方法来诱变编码相关蛋白质或多肽的基因,以使其包括例如一个或一个以上用于并入非天然氨基酸的选择密码子。举例来说,诱变相关蛋白质的核酸,以使其包括一个或一个以上选择密码子,从而并入一个或一个以上非天然氨基酸。本发明包括例如包括至少一个非天然氨基酸的任何蛋白质的任何此种变异体(包括(但不限于)突变体)形式。类似地,本发明还包括相应核酸,即,具有一个或一个以上编码一个或一个以上非天然氨基酸的选择密码子的任何核酸。
可容易地使编码相关蛋白质(例如bst多肽)的核酸分子突变,以在多肽的任何所需位置引入半胱氨酸。半胱氨酸广泛用于将反应性分子、水溶性聚合物、蛋白质或多种其它分子引入相关蛋白质上。所属领域技术人员众所周知适于将半胱氨酸并入多肽的所需位置中的方法,例如美国专利第6,608,183号(以引用的方式并入本文中)中所述的那些方法,以及标准诱变技术。
iv.非天然编码氨基酸
多种非天然编码氨基酸适用于本发明中。许多非天然编码氨基酸都可引入bst多肽中。一般说来,引入的非天然编码氨基酸对20种常见基因编码的氨基酸(即,丙氨酸、精氨酸、天冬酰胺、天冬氨酸、半胱氨酸、谷氨酰胺、谷氨酸、甘氨酸、组氨酸、异亮氨酸、亮氨酸、赖氨酸、甲硫氨酸、苯丙氨酸、脯氨酸、丝氨酸、苏氨酸、色氨酸、酪氨酸和缬氨酸)实质上呈化学惰性。在一些实施例中,非天然编码氨基酸包括与未见于20种常见氨基酸中的官能团(包括(但不限于)叠氮基、酮基、醛基和氨氧基)有效选择性反应形成稳定结合物的侧链官能团。举例来说,可以使包括含叠氮基官能团的非天然编码氨基酸的bst多肽与聚合物(包括(但不限于)聚(乙二醇))或者含炔部分的第二多肽反应,形成稳定结合物,这一结合物是由叠氮与炔官能团选择性反应而形成胡伊斯根[3+2]环加成产物得到。
α-氨基酸的一般结构如下所示(式i):
i
非天然编码氨基酸通常为具有上文所列式的任何结构,其中r基团是除20种天然氨基酸中所用外的任何取代基,并且都可适用于本发明中。由于本发明非天然编码氨基酸通常仅侧链结构不同于天然氨基酸,故非天然编码氨基酸与其它氨基酸(包括(但不限于)天然或非天然编码氨基酸)形成酰胺键的方式与在天然存在多肽中形成酰胺键的方式相同。然而,非天然编码氨基酸的侧链基团使其与天然氨基酸得以区别。举例来说,r任选包含烷基-、芳基-、酰基-、酮基-、叠氮基-、羟基-、肼、氰基-、卤基-、酰肼、烯基、炔基、醚、硫醇、硒基-、磺酰基-、硼酸酯基(borate)、硼酸酯基(boronate)、磷酸基、膦酸基、膦、杂环、烯酮、亚胺、醛、酯、硫代酸、羟胺、氨基等,或其任何组合。适用于本发明中的其它相关的非天然存在氨基酸包括(但不限于)包含可光活化交联剂的氨基酸、自旋标记的氨基酸、荧光氨基酸、金属结合氨基酸、含金属氨基酸、放射性氨基酸、具有新颖官能团的氨基酸、与其它分子共价或非共价相互作用的氨基酸、光笼蔽和/或可光异构化氨基酸、包含生物素或生物素类似物的氨基酸、糖基化氨基酸(例如糖取代的丝氨酸)、其它碳水化合物修饰的氨基酸、含酮基氨基酸、包含聚乙二醇或聚醚的氨基酸、重原子取代的氨基酸、可化学裂解和/或可光裂解氨基酸、侧链比天然氨基酸长的氨基酸(包括(但不限于)聚醚或长链烃,包括(但不限于)大于约5个或大于约10个碳)、含碳连接糖的氨基酸、氧化还原活性氨基酸、含有氨基硫代酸的氨基酸和包含一个或一个以上有毒部分的氨基酸。
适用于本发明中并且适用于与水溶性聚合物反应的示范性非天然编码氨基酸包括(但不限于)具有羰基、氨氧基、肼、酰肼、氨基脲、叠氮和炔反应性基团的氨基酸。在一些实施例中,非天然编码氨基酸包含糖部分。这些氨基酸的实例包括n-乙酰基-l-氨基葡糖基-l-丝氨酸、n-乙酰基-l-氨基半乳糖基-l-丝氨酸、n-乙酰基-l-氨基葡糖基-l-苏氨酸、n-乙酰基-l-氨基葡糖基-l-天冬酰胺和o-氨基甘露糖基-l-丝氨酸。所述氨基酸的实例还包括氨基酸与糖之间的天然存在的n键或o键经自然界中不常见的共价键(包括(但不限于)烯烃、肟、硫醚、酰胺等)置换的实例。所述氨基酸的实例还包括天然存在的蛋白质中不常见的糖,例如2-脱氧-葡萄糖、2-脱氧半乳糖等。
本文中提供的许多非天然编码氨基酸可购自例如西格玛-阿尔德里奇公司(sigma-aldrich)(美国密苏里州圣路易斯(st.louis,mo,usa))、诺瓦生物科技公司(novabiochem)(emd生物科技公司(emdbiosciences)的分公司,位于德国达姆施塔特(darmstadt,germany))或派普科技公司(peptech)(美国马萨诸塞州伯灵顿(burlington,ma,usa))。非市售的非天然编码氨基酸可任选根据本文中提供或使用所属领域技术人员已知的标准方法合成。有关有机合成技术,例如参见弗雷斯敦(fessendon)和弗雷斯敦的有机化学(organicchemistry),(1982,第2版,维拉德格兰特出版社(willardgrantpress),马萨诸塞州波士顿(bostonmass.));马奇(march)的高等有机化学(advancedorganicchemistry)(第3版,1985,威立父子出版公司,纽约);以及卡雷(carey)和萨德伯格(sundberg)的高等有机化学(第3版,第a和b部分,1990,派拉蒙出版社(plenumpress),纽约)。还参见美国专利第7,045,337号和第7,083,970号,都以引用的方式并入本文中。除含有新颖侧链的非天然氨基酸外,适用于本发明中的非天然氨基酸还任选包含经修饰的主链结构,包括(但不限于)如式ii和iii的结构所示:
ii
iii
其中z通常包含oh、nh2、sh、nh-r'或s-r';x和y可相同或不同,通常包含s或o;且r和r'任选相同或不同,通常选自与上文关于具有式i的非天然氨基酸所述的r基团的组分相同的清单以及氢。举例来说,如式ii和iii所示,本发明的非天然氨基酸任选在氨基或羧基中包含取代。此类非天然氨基酸包括(但不限于)α-羟基酸、α-硫代酸、α-氨基硫代羧酸酯,包括(但不限于)具有与常见20种天然氨基酸对应的侧链或非天然侧链。此外,α-碳的取代任选包括(但不限于)l、d或α-α双取代氨基酸,例如d-谷氨酸、d-丙氨酸、d-甲基-o-酪氨酸、氨基丁酸等。其它结构替代物包括环状氨基酸,例如脯胺酸类似物,以及3元、4元、6元、7元、8元和9元环脯胺酸类似物;β和γ氨基酸,例如经取代β-丙氨酸和γ-氨基丁酸。
许多非天然氨基酸都是基于天然氨基酸,例如酪胺酸、谷氨酰胺、苯丙氨酸等,并且都适用于本发明中。酪氨酸类似物包括(但不限于)对位取代的酪氨酸、邻位取代的酪氨酸和间位取代的酪氨酸,其中这些经取代酪氨酸包含(包括(但不限于))酮基(包括(但不限于)乙酰基)、苯甲酰基、氨基、肼、羟胺、硫醇基、羧基、异丙基、甲基、c6-c20直链或支链烃、饱和或不饱和烃、o-甲基、聚醚基、硝基、炔基等。此外,也涵盖多取代的芳基环。适用于本发明中的谷氨酰胺类似物包括(但不限于)α-羟基衍生物、γ取代的衍生物、环状衍生物和酰胺取代的谷氨酰胺衍生物。适用于本发明中的苯丙氨酸类似物的实例包括(但不限于)对位取代的苯丙氨酸、邻位取代的苯丙氨酸和间位取代的苯丙氨酸,其中所述取代基包含(包括(但不限于))羟基、甲氧基、甲基、烯丙基、醛、叠氮基、碘、溴、酮基(包括(但不限于)乙酰基)、苯甲酰基、炔基等。适用于本发明中的非天然氨基酸的具体实例包括(但不限于)对乙酰基-l-苯丙氨酸、o-甲基-l-酪氨酸、l-3-(2-萘基)丙氨酸、3-甲基-苯丙氨酸、o-4-烯丙基-l-酪氨酸、4-丙基-l-酪氨酸、三-o-乙酰基-glcnacβ-丝氨酸、l-多巴(l-dopa)、氟代苯丙氨酸、异丙基-l-苯丙氨酸、对叠氮基-l-苯丙氨酸、对酰基-l-苯丙氨酸、对苯甲酰基-l-苯丙氨酸、l-磷酸丝氨酸、膦酰基丝氨酸、膦酰基酪氨酸、对碘-苯丙氨酸、对溴苯丙氨酸、对氨基-l-苯丙氨酸、异丙基-l-苯丙氨酸和对炔丙氧基-苯丙氨酸等。适用于本发明中的多种非天然氨基酸结构的实例提供于例如名为“体内并入非天然氨基酸(invivoincorporationofunnaturalaminoacids)”的wo2002/085923中。关于其它甲硫氨酸类似物,还参见柯克(kiick)等人,(2002)利用施陶丁格尔接合将叠氮基并入重组蛋白中以进行化学选择性修饰(incorporationofazidesintorecombinantproteinsforchemoselectivemodificationbythestaudingerligtation),pnas99:19-24,其以引用的方式并入本文中。国际申请案第pct/us06/47822号,名为“含有非天然氨基酸和多肽的组合物、涉及非天然氨基酸和多肽的方法以及非天然氨基酸和多肽的用途(compositionscontaining,methodsinvolving,andusesofnon-naturalaminoacidsandpolypeptides)”(以引用的方式并入本文中),描述芳香族胺部分(包括(但不限于)对氨基-苯丙氨酸)的还原烷基化,以及还原胺化。
在一个实施例中,提供bst多肽的组合物,所述bst多肽包括非天然氨基酸(例如对-(炔丙氧基)-苯丙氨酸)。还提供包含对-(炔丙氧基)-苯丙氨酸且包括(但不限于)蛋白质和/或细胞的各种组合物。一方面,包括对-(炔丙氧基)-苯丙氨酸非天然氨基酸的组合物进一步包括正交trna。非天然氨基酸可与正交trna键接(包括(但不限于)共价),包括(但不限于)经由氨酰基键与正交trna共价键接、与正交trna的末端核糖的3'oh或2'oh共价键接等。
可经由非天然氨基酸并入蛋白质中的化学部分将为蛋白质提供多种益处和操作。举例来说,酮基官能团的独特反应性允许在体外和体内利用多种含肼或含羟胺试剂中的任一种选择性修饰蛋白质。重原子非天然氨基酸例如可用于定相x射线结构数据。使用非天然氨基酸位点特异性地引入重原子还为选择重原子的位置提供选择性和灵活性。光反应性非天然氨基酸(包括(但不限于)具有二苯甲酮和叠氮芳基(包括(但不限于)叠氮苯基)侧链的氨基酸)例如允许在体内和体外有效地光交联蛋白质。光反应性非天然氨基酸的实例包括(但不限于)对叠氮基-苯丙氨酸和对苯甲酰基-苯丙氨酸。随后可通过激发光反应性基团(提供时间控制)使具有光反应性非天然氨基酸的蛋白质随意交联。在一个实例中,非天然氨基酸的甲基可经作为局部结构和动力学探针(包括(但不限于)使用核磁共振和振动光谱)的同位素标记(包括(但不限于))甲基取代。举例来说,炔基或叠氮基官能团允许利用分子通过[3+2]环加成反应来选择性修饰蛋白质。
并入多肽氨基末端的非天然氨基酸可由以下各物构成:r基团,其为除20种天然氨基酸中所使用者外的任何取代基;和第二反应性基团,其不同于通常存在于α-氨基酸中的nh2基团(参见式i)。可并入羧基末端的类似非天然氨基酸中的第二反应性基团不同于通常存在于α-氨基酸中的cooh基团(参见式i)。
本发明的非天然氨基酸可经选择或设计,从而提供在20种天然氨基酸中不可得到的其它特征。举例来说,可任选对非天然氨基酸进行设计或选择,从而改变例如并有这些非天然氨基酸的蛋白质的生物特性。举例来说,任选通过在蛋白质中包括非天然氨基酸来改变以下特性:毒性、生物分布、溶解性、稳定性(例如热稳定性、水解稳定性、氧化稳定性、对酶降解的抗性等)、纯化和加工的便利性、结构特性、光谱特性、化学和/或光化学特性、催化活性、氧化还原电位、半衰期、与其它分子反应(例如共价或非共价)的能力等。
非天然氨基酸的结构和合成:羰基、类羰基、经遮蔽羰基、经保护羰基和羟胺基
在一些实施例中,本发明提供的bst经由肟键与水溶性聚合物(例如peg)连接。
许多类型的非天然编码氨基酸适于形成肟键。这些氨基酸包括(但不限于)含有羰基、二羰基或羟胺基的非天然编码氨基酸。所述氨基酸描述于以下专利文献中:美国专利公开案第2006/0194256号、第2006/0217532号和第2006/0217289号,以及wo2006/069246,名为“含有非天然氨基酸和多肽的组合物、涉及非天然氨基酸和多肽的方法以及非天然氨基酸和多肽的用途(compositionscontaining,methodsinvolving,andusesofnon-naturalaminoacidsandpolypeptides)”,这些专利文献的全文都以引用的方式并入本文中。非天然编码氨基酸也描述于美国专利第7,083,970号和美国专利第7,045,337号中,这些专利的全文都以引用的方式并入本文中。
本发明一些实施例利用一个或一个以上位置经对乙酰基苯丙氨酸氨基酸取代的bst多肽。对乙酰基-(+/-)-苯丙氨酸和间乙酰基-(+/-)-苯丙氨酸的合成描述于张z.(zhang,z.)等人,生物化学(biochemistry)42:6735-6746(2003)中,以引用的方式并入本文中。所属领域技术人员可以用类似方式制备其它含羰基或含二羰基的氨基酸。此外,本文中包括的非天然氨基酸的非限制性示范性合成法提供于美国专利第7,083,970号(以全文引用的方式并入本文中)图4、24-34和36-39中。
具有亲电反应性基团的氨基酸允许经由亲核加成反应等连接分子的各种反应。所述亲电反应性基团包括羰基(包括酮基和二羰基)、类羰基基团(反应性类似于羰基(包括酮基和二羰基)且结构类似于羰基)、经遮蔽的羰基(易于转化成羰基(包括酮基和二羰基))或经保护羰基(脱除保护基后,反应性类似于羰基(包括酮基和二羰基))。所述氨基酸包括具有式(iv)结构的氨基酸:
其中:
a为任选存在的,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
b为任选存在的,并且当存在时,其为选自由以下组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
j为
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
各r"独立地为h、烷基、经取代烷基或保护基,或者当存在一个以上r"基团时,两个r"任选形成杂环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
r3和r4各独立地为h、卤素、低碳数烷基或经取代低碳数烷基,或者r3和r4或两个r3基团任选形成环烷基或杂环烷基;
或-a-b-j-r基团一起形成包含至少一个羰基的双环或三环环烷基或杂环烷基,所述羰基包括二羰基、经保护羰基(包括经保护二羰基)或经遮蔽羰基(包括经遮蔽二羰基);
或-j-r基团一起形成包含至少一个羰基的单环或双环环烷基或杂环烷基,所述羰基包括二羰基、经保护羰基(包括经保护二羰基)或经遮蔽羰基(包括经遮蔽二羰基);
条件是,当a为亚苯基且各r3为h时,b存在;且当a为-(ch2)4-且各r3为h时,b不为-nhc(o)(ch2ch2)-;且当a和b都不存在,且各r3为h时,r不为甲基。
此外,还包括具有式(v)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
条件是,当a为亚苯基时,b存在;且当a为-(ch2)4-时,b不为-nhc(o)(ch2ch2)-;且当a和b不存在时,r不为甲基。
此外,还包括具有式(vi)结构的氨基酸:
其中:
b为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基。
此外,还包括以下氨基酸:
此外,还包括以下具有式(vii)结构的氨基酸:
其中
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基;且n为0到8;
条件是,当a为-(ch2)4-时,b不为-nhc(o)(ch2ch2)-。
此外,还包括以下氨基酸:
此外,还包括以下具有式(viii)结构的氨基酸:
其中a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸。
此外,还包括以下具有式(ix)结构的氨基酸:
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
其中各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基。
此外,还包括以下氨基酸:
此外,还包括以下具有式(x)结构的氨基酸:
其中b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基;且n为0到8。
此外,还包括以下氨基酸:
本文中所述的非天然氨基酸除包括单羰基结构外,也可包括例如二羰基、类二羰基、经遮蔽二羰基和经保护二羰基等基团。
举例来说,包括以下具有式(xi)结构的氨基酸:
其中a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸。
此外,还包括以下具有式(xii)结构的氨基酸:
b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
其中各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基。
此外,还包括以下氨基酸:
此外,还包括以下具有式(xiii)结构的氨基酸:
其中b任选存在,并且当存在时,其为选自由以下各基团组成的群组的连接子:低碳数亚烷基、经取代低碳数亚烷基、低碳数亚烯基、经取代低碳数亚烯基、低碳数亚杂烷基、经取代低碳数亚杂烷基、-o-、-o-(亚烷基或经取代亚烷基)-、-s-、-s-(亚烷基或经取代亚烷基)-、-s(o)k-(其中k为1、2或3)、-s(o)k(亚烷基或经取代亚烷基)-、-c(o)-、-c(o)-(亚烷基或经取代亚烷基)-、-c(s)-、-c(s)-(亚烷基或经取代亚烷基)-、-n(r')-、-nr'-(亚烷基或经取代亚烷基)-、-c(o)n(r')-、-con(r')-(亚烷基或经取代亚烷基)-、-csn(r')-、-csn(r')-(亚烷基或经取代亚烷基)-、-n(r')co-(亚烷基或经取代亚烷基)-、-n(r')c(o)o-、-s(o)kn(r')-、-n(r')c(o)n(r')-、-n(r')c(s)n(r')-、-n(r')s(o)kn(r')-、-n(r')-n=、-c(r')=n-、-c(r')=n-n(r')-、-c(r')=n-n=、-c(r')2-n=n-和-c(r')2-n(r')-n(r')-,其中各r'独立地为h、烷基或经取代烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基;且n为0到8。
此外,还包括以下氨基酸:
此外,还包括以下具有式(xiv)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
x1为c、s或s(o);且l为亚烷基、经取代亚烷基、n(r')(亚烷基)或n(r')(经取代亚烷基),其中r'为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xiv-a)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
l为亚烷基、经取代亚烷基、n(r')(亚烷基)或n(r')(经取代亚烷基),其中r'为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xiv-b)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
l为亚烷基、经取代亚烷基、n(r′)(亚烷基)或n(r′)(经取代亚烷基),其中r′为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xv)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
x1为c、s或s(o);且n为0、1、3、4或5;且各cr8r9基团上的r8和r9各独立地选自由以下各基团组成的群组:h、烷氧基、烷基胺、卤素、烷基、芳基,或者任何r8和r9可一起形成=o或环烷基,或任何相邻的r8基团可一起形成环烷基。
此外,还包括以下具有式(xv-a)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,具为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
n为0、1、3、4或5;且各cr8r9基团上的r8和r9各独立地选自由以下各基团组成的群组:h、烷氧基、烷基胺、卤素、烷基、芳基,或者任何r8和r9可一起形成=o或环烷基,或任何相邻的r8基团可一起形成环烷基。
此外,还包括以下具有式(xv-b)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
n为0、1、3、4或5;且各cr8r9基团上的r8和r9各独立地选自由以下各基团组成的群组:h、烷氧基、烷基胺、卤素、烷基、芳基,或者任何r8和r9可一起形成=o或环烷基,或任何相邻的r8基团可一起形成环烷基。
此外,还包括以下具有式(xvi)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
x1为c、s或s(o);且l为亚烷基、经取代亚烷基、n(r′)(亚烷基)或n(r′)(经取代亚烷基),其中r′为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xvi-a)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
l为亚烷基、经取代亚烷基、n(r')(亚烷基)或n(r')(经取代亚烷基),其中r'为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xvi-b)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
r为h、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
l为亚烷基、经取代亚烷基、n(r')(亚烷基)或n(r')(经取代亚烷基),其中r'为h、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括具有式(xvii)结构的氨基酸:
其中:
a任选存在,并且当存在时,其为低碳数亚烷基、经取代低碳数亚烷基、低碳数亚环烷基、经取代低碳数亚环烷基、低碳数亚烯基、经取代低碳数亚烯基、亚炔基、低碳数亚杂烷基、经取代亚杂烷基、低碳数亚杂环烷基、经取代低碳数亚杂环烷基、亚芳基、经取代亚芳基、亚杂芳基、经取代亚杂芳基、亚烷芳基、经取代亚烷芳基、亚芳烷基或经取代亚芳烷基;
m为-c(r3)-、
r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基;
t3为一键、c(r)(r)、o或s,且r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸。
此外,还包括具有式(xviii)结构的氨基酸:
其中:
m为-c(r3)-、
r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基;
t3为一键、c(r)(r)、o或s,且r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基;
r1任选存在,且当存在时,其为h、氨基保护基、树脂、氨基酸、多肽或聚核苷酸;且
r2任选存在,且当存在时,其为oh、酯保护基、树脂、氨基酸、多肽或聚核苷酸;
各ra独立地选自由以下各基团组成的群组:h、卤素、烷基、经取代烷基、-n(r')2、-c(o)kr'(其中k为1、2或3)、-c(o)n(r')2、-or'和-s(o)kr',其中各r'独立地为h、烷基或经取代烷基。
此外,还包括具有式(xix)结构的氨基酸:
其中:
r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基;且
t3为o或s。
此外,还包括具有式(xx)结构的氨基酸:
其中:
r为h、卤素、烷基、经取代烷基、环烷基或经取代环烷基。
此外,还包括以下具有式(xxi)结构的氨基酸:
在一些实施例中,对包含非天然氨基酸的多肽进行化学修饰以产生反应性羰基或二羰基官能团。举例来说,可由具有相邻氨基和羟基的官能团产生适用于结合反应的醛官能团。当生物活性分子为多肽时,例如,可使用n末端丝氨酸或苏氨酸(通常存在或者可经由化学或酶消化而暴露),在温和氧化裂解条件下使用高碘酸盐产生醛官能团。例如参见盖特纳(gaertner)等人,生物结合物化学(bioconjug.chem.)3:262-268(1992);乔治格k.(geoghegan,k.)和斯托赫j.(stroh,j.),生物结合物化学,3:138-146(1992);盖特纳等人,生物化学杂志(j.biol.chem.)269:7224-7230(1994)。然而,此项技术中已知的方法局限于在肽或蛋白质n末端的氨基酸。
在本发明中,可将带有相邻羟基和氨基的非天然氨基酸以“经遮蔽”醛官能团的形式并入多肽中。举例来说,5-羟基赖氨酸在邻近ε胺处带有羟基。用于产生醛的反应条件通常涉及在温和条件下添加摩尔数过量的偏高碘酸钠,以避免在多肽内的其它位点发生氧化。氧化反应的ph值通常为约7.0。典型反应涉及将约1.5摩尔过量的偏高碘酸钠添加到多肽缓冲溶液中,随后在暗处培育约10分钟。例如参见美国专利第6,423,685号。
在温和条件下,羰基或二羰基官能团可与含羟胺试剂在水溶液中选择性反应,形成在生理条件下稳定的相应肟键。例如参见杰克斯w.p.(jencks,w.p.),美国化学协会杂志,81,475-481(1959);邵j.(shao,j.)和泰姆j.p.(tam,j.p.),美国化学协会杂志,117:3893-3899(1995)。此外,羰基或二羰基的独特反应性允许在其它氨基酸侧链存在下进行选择性修饰。例如参见柯尼西v.w.(cornish,v.w.)等人,美国化学协会杂志,118:8150-8151(1996);乔治格k.f.(geoghegan,k.f.)和斯托赫j.g.(stroh,j.g.),生物结合物化学,3:138-146(1992);马赫尔l.k.(mahal,l.k.)等人,科学,276:1125-1128(1997)。
非天然氨基酸的结构和合成:含羟胺氨基酸
美国临时专利申请案第60/638,418号是以全文引用的方式并入本文中。因此,美国临时专利申请案第60/638,418号中第v节(名为“非天然氨基酸(non-naturalaminoacids)”)第b部分(名为“非天然氨基酸的结构和合成:含羟胺氨基酸(structureandsynthesisofnon-naturalaminoacids:hydroxylamine-containingaminoacids)”)中提供的揭示内容完全适用于制备、纯化、表征和使用本文中所述的非天然氨基酸、非天然氨基酸多肽和经修饰非天然氨基酸多肽的方法、组合物(包括式i-xxxv)、技术和策略,其适用程度就如同所述揭示内容完全提供于本文中。美国专利公开案第2006/0194256号、第2006/0217532号和第2006/0217289号,以及wo2006/069246,名为“含有非天然氨基酸和多肽的组合物、涉及非天然氨基酸和多肽的方法以及非天然氨基酸和多肽的用途(compositionscontaining,methodsinvolving,andusesofnon-naturalaminoacidsandpolypeptides)”,都以全文引用的方式并入本文中。
非天然氨基酸的化学合成
适用于本发明中的许多非天然氨基酸例如购自西格玛公司(sigma)(美国)或阿尔德里奇公司(aldrich)(美国威斯康辛州密尔沃基(milwaukee,wi,usa))。非市售的非天然氨基酸任选可根据本文中所提供或根据各种公开案中所提供或者使用所属领域技术人员已知的标准方法合成。有关有机合成技术,例如参见弗雷斯敦和弗雷斯敦的有机化学,(1982,第2版,维拉德格兰特出版社,马萨诸塞州波士顿);马奇的高等有机化学(第3版,1985,威立父子出版公司,纽约);以及卡雷和萨德伯格的高等有机化学(第3版,第a和b部分,1990,派拉蒙出版社,纽约)。描述非天然氨基酸合成的其它出版物例如包括:wo2002/085923,名为“体内并入非天然氨基酸(invivoincorporationofunnaturalaminoacids)”;马托索卡斯(matsoukas)等人,(1995)药物化学杂志(j.med.chem.),38-4669;金f.e.(king,f.e.)和凯德d.a.a.(kidd,d.a.a.)(1949)由邻苯二甲酰化中间物合成谷氨酰胺以及谷氨酸γ-二肽的新方法。(anewsynthesisofglutamineandofγ-dipeptidesofglutamicacidfromphthylatedintermediates.)化学协会杂志(j.chem.soc.)3315-3319;弗雷德曼o.m.(friedman,o.m.)和查特瑞提r.(chatterrji,r.)(1959)作为抗肿瘤剂的模型底物的谷氨酰胺衍生物的合成。(synthesisofderivativesofglutamineasmodelsubstratesforanti-tumoragents.)美国化学协会杂志81,3750-3752;克莱格j.c.(craig,j.c.)等人(1988)7-氯-4[[4-(二乙氨基)-1-甲基丁基]氨基]喹啉(氯喹)的绝对构型。(absoluteconfigurationoftheenantiomersof7-chloro-4[[4-(diethylamino)-1-methylbutyl]amino]quinoline(chloroquine).)有机化学杂志53,1167-1170;阿佐雷m.(azoulay,m.),维尔蒙特m.(vilmont,m.)和弗莱皮尔f.(frappier,f.)(1991)作为潜在抗疟药的谷氨酰胺类似物(glutamineanaloguesaspotentialantimalarials),欧洲药物化学杂志(eur.j.med.chem.)26,201-5;库斯基宁a.m.p.(koskinen,a.m.p.)和雷珀特h.(rapoport,h.)(1989)合成4-经取代脯氨酸作为构象受限的氨基酸类似物。(synthesisof4-substitutedprolinesasconformationallyconstrainedaminoacidanalogues.)有机化学杂志54,1859-1866;克雷斯廷b.d.(christie,b.d.)和雷珀特h.,(1985)由l-天冬酰胺合成光学纯六氢吡啶的方法。(synthesisofopticallypurepipecolatesfroml-asparagine.)经由氨基酸脱羰基和亚胺离子环化全合成(+)阿扑长春胺的应用。(applicationtothetotalsynthesisof(+)-apovincaminethroughaminoaciddecarbonylationandiminiumioncyclization.)有机化学杂志50:1239-1246;巴顿(barton)等人,(1987)使用自由基化学合成新颖的α-氨基酸和衍生物:合成l-和d-α-氨基-己二酸、l-α-氨基庚二酸和适合的不饱和衍生物。(synthesisofnovelalpha-amino-acidsandderivativesusingradicalchemistry:synthesisofl-andd-alhpa-amino-adipicacids,l-alpha-aminopimelicacidandappropriateunsaturatedderivatives.)四面体通讯(tetrahedron)43:4297-4308;和萨贝辛赫(subasinghe)等人,(1992)使君子氨酸类似物:合成β-杂环2-氨基丙酸衍生物和其在新颖的使君子氨酸敏感性位点的活性。(quisqualicacidanalogues:synthesisofbeta-heterocyclic2-aminopropanoicacidderivativesandtheiractivityatanovelquisqualate-sensitizedsite.)药物化学杂志35:4602-7。还参见美国专利公开案第us2004/0198637号,名为“蛋白质阵列(proteinarrays)”,以引用的方式并入本文中。
a.羰基反应性基团
具有羰基反应性基团的氨基酸能够进行多种反应,经由亲核加成或醛醇缩合反应等连接分子(包括(但不限于)peg或其它水溶性分子)。
示范性含羰基氨基酸可如下所示:
其中n为0到10;r1为烷基、芳基、经取代烷基或经取代芳基;r2为h、烷基、芳基、经取代烷基和经取代芳基;且r3为h、氨基酸、多肽或氨基末端修饰基团;且r4为h、氨基酸、多肽或羧基末端修饰基团。在一些实施例中,n为1,r1为苯基且r2为简单烷基(即,甲基、乙基或丙基),并且酮部分位于烷基侧链的对位。在一些实施例中,n为1,r1为苯基且r2为简单烷基(即,甲基、乙基或丙基),并且酮部分位于烷基侧链的间位。
对乙酰基-(+/-)-苯丙氨酸和间乙酰基-(+/-)-苯丙氨酸的合成描述于张z.等人,生物化学42:6735-6746(2003)中,其以引用的方式并入本文中。所属领域技术人员可以用类似方式制备其它含羰基氨基酸。
在一些实施例中,对包含非天然编码氨基酸的多肽进行化学修饰以产生反应性羰基官能团。举例来说,可由具有相邻氨基和羟基的官能团产生适用于结合反应的醛官能团。当生物活性分子为多肽时,例如,可使用n末端丝氨酸或苏氨酸(通常存在或者可经由化学或酶消化而暴露),在温和氧化裂解条件下使用高碘酸盐产生醛官能团。例如参见盖特纳等人,生物结合物化学,3:262-268(1992);乔治格k.和斯托赫j.,生物结合物化学,3:138-146(1992);盖特纳等人,生物化学杂志269:7224-7230(1994)。然而,此项技术中已知的方法局限于在肽或蛋白质n末端的氨基酸。
在本发明中,可将带有相邻羟基和氨基的非天然编码氨基酸以“经遮蔽”醛官能团的形式并入多肽中。举例来说,5-羟基赖氨酸在邻近ε胺处带有羟基。用于产生醛的反应条件通常涉及在温和条件下添加摩尔数过量的偏高碘酸钠,以避免在多肽内的其它位点发生氧化。氧化反应的ph值通常为约7.0。典型反应涉及将约1.5摩尔过量的偏高碘酸钠添加到多肽缓冲溶液中,随后在暗处培育约10分钟。例如参见美国专利第6,423,685号,其以引用的方式并入本文中。
在温和条件下,羰基官能团可与含肼、含酰肼、含羟胺或含氨基脲的试剂在水溶液中选择性反应,从而分别形成在生理条件下稳定的相应腙、肟或缩氨基脲键。例如参见杰克斯w.p.,美国化学协会杂志,81,475-481(1959);邵j.和泰姆j.p.,美国化学协会杂志,117:3893-3899(1995)。此外,羰基的独特反应性允许在其它氨基酸侧链存在下进行选择性修饰。例如参见柯尼西v.w.等人,美国化学协会杂志,118:8150-8151(1996);乔治格k.f.和斯托赫j.g.,生物结合物化学,3:138-146(1992);马赫尔l.k.等人,科学,276:1125-1128(1997)。
b.肼、酰肼或氨基脲反应性基团
含有亲核基团(例如肼、酰肼或氨基脲)的非天然编码氨基酸允许与各种亲电基团反应形成结合物(包括(但不限于)与peg或其它水溶性聚合物形成结合物)。
示范性含肼、含酰肼或含氨基脲的氨基酸可如下所示:
其中n为0-10;r1为烷基、芳基、经取代烷基、经取代芳基,或不存在;x为o、n、s,或不存在;r2为h、氨基酸、多肽或氨基末端修饰基团;且r3为h、氨基酸、多肽或羧基末端修饰基团。
在一些实施例中,n为4,r1不存在且x为n。在一些实施例中,n为2,r1不存在且x不存在。在一些实施例中,n为1,r1为苯基,x为o,且氧原子位于芳环上脂肪族基团的对位。
含酰肼、含肼和含氨基脲的氨基酸可从商业来源获得。举例来说,l-谷氨酸-γ-酰肼可购自西格玛化学品公司(sigmachemical)(密苏里州圣路易斯(st.louis,mo))。其它非市售氨基酸可由所属领域技术人员制备。例如参见美国专利第6,281,211号,其以引用的方式并入本文中。
含有带酰肼、肼或氨基脲官能团的非天然编码氨基酸的多肽可与多个含有醛或其它具有类似化学反应性的官能团的分子有效地选择性反应。例如参见邵j.和泰姆j.,美国化学协会杂志,117:3893-3899(1995)。与20种常见氨基酸上存在的亲核基团(包括(但不限于)丝氨酸或苏氨酸的羟基,或赖氨酸和n末端的氨基)相比较,酰肼、肼和氨基脲官能团的独特反应性使其对醛、酮和其它亲电基团的反应性明显更高。
c.含氨氧基的氨基酸
含有氨氧基(也称为羟胺)的非天然编码氨基酸能够与各种亲电子基团反应,形成结合物(包括(但不限于)与peg或其它水溶性聚合物形成结合物)。与肼、酰肼和氨基脲类似,增强氨氧基的亲核性使其能够与多个含有醛或其它具有类似化学反应性的官能团的分子有效地选择性反应。例如参见邵j.和泰姆j.,美国化学协会杂志,117:3893-3899(1995);h.杭(h.hang)和c.贝托兹(c.bertozzi),化学研究述评(acc.chem.res.)34:727-736(2001)。与肼基反应得到相应腙,而肟一般是由氨氧基与含羰基基团(例如酮)反应得到。
示范性含氨氧基的氨基酸可如下所示:
其中n为0到10;r1为烷基、芳基、经取代烷基、经取代芳基,或不存在;x为o、n、s,或不存在;m为0到10;y为=c(o),或不存在;r2为h、氨基酸、多肽或氨基末端修饰基团;且r3为h、氨基酸、多肽或羧基末端修饰基团。在一些实施例中,n为1,r1为苯基,x为o,m为1且y存在。在一些实施例中,n为2,r1和x不存在,m为0且y不存在。
含氨氧基的氨基酸可由容易得到的氨基酸前体(高丝氨酸、丝氨酸和苏氨酸)制备。例如参见m.卡拉索科(m.carrasco)和r.布朗(r.brown),有机化学杂志,68:8853-8858(2003)。已经从天然来源中分离出某些含氨氧基的氨基酸,例如l-2-氨基-4-(氨氧基)丁酸(罗森纳g.(rosenthal,g.),生命科学(lifesci.)60:1635-1641(1997))。所属领域技术人员可制备出其它含氨氧基的氨基酸。
d.叠氮基和炔反应性基团
叠氮基和炔官能团的独特反应性使其极其适用于选择性修饰多肽和其它生物分子。有机叠氮化物,尤其是脂肪族叠氮化物,以及炔一般对常用化学反应条件稳定。具体说来,叠氮基和炔官能团对天然存在多肽中所见的20种常见氨基酸的侧链(即,r基)呈惰性。但当紧密接近时,会呈现叠氮基和炔基的“弹簧加压(spring-loaded)”性,并且其经由胡伊斯根[3+2]环加成反应有效选择性反应,产生相应的三唑。例如参见秦j.(chinj.)等人,科学,301:964-7(2003);王q.(wang,q.)等人,美国化学协会杂志,125,3192-3193(2003);秦j.w.等人,美国化学协会杂志,124:9026-9027(2002)。
由于胡伊斯根环加成反应涉及选择性环加成反应(例如参见帕德瓦a.(padwa,a.),有机合成大全(comprehensiveorganicsynthesis),第4卷,(托斯特b.m.(trost,b.m.)编,1991),第1069-1109页;胡伊斯根r.(huisgen,r.),1,3-偶极环加成化学(1,3-dipolarcycloadditionchemistry),(帕德瓦a.编,1984),第1-176页)而非亲核取代反应,故并入带有含叠氮和含炔侧链的非天然编码氨基酸允许在非天然编码氨基酸位置对所得多肽进行选择性修饰。可在室温下,于水性条件下,通过在用于将cu(ii)还原成cu(i)的还原剂存在下,添加催化量的cu(ii)(包括(但不限于)催化量的cuso4形式),原位进行涉及含叠氮或含炔的bst多肽的环加成反应。例如参见王q.(wang,q.)等人,美国化学协会杂志,125,3192-3193(2003);托尼诺c.w.(tornoe,c.w.)等人,有机化学杂志,67:3057-3064(2002);罗斯托瑟夫(rostovtsev)等人,德国应用化学,41:2596-2599(2002)。示范性还原剂包括(包括(但不限于))抗坏血酸盐、金属铜、奎宁(quinine)、氢醌、维生素k、谷胱甘肽、半胱氨酸、fe2+、co2+和外加电位。
在需要叠氮化物与炔之间发生胡伊斯根[3+2]环加成反应的一些情况下,bst多肽包含包括炔部分的非天然编码氨基酸,且待与所述氨基酸连接的水溶性聚合物包含叠氮部分。或者,也可执行逆反应(即,氨基酸上的叠氮部分与水溶性聚合物上存在的炔部分反应)。
叠氮官能团也可与含有芳基酯且经芳基膦部分适当官能化的水溶性聚合物选择性反应,从而产生酰胺键。芳基膦基在原位还原叠氮基,随后所得胺与邻近的酯键有效反应,产生相应酰胺。例如参见e.萨龙(e.saxon)和c.波特兹(c.bertozzi),科学,287,2007-2010(2000)。含叠氮基的氨基酸可以是烷基叠氮化物(包括(但不限于)2-氨基-6-叠氮基-1-己酸)或芳基叠氮化物(对叠氮基苯丙氨酸)。
含有芳基酯和膦部分的示范性水溶性聚合物可如下所示:
其中x可为o、n、s,或不存在,ph为苯基,w为水溶性聚合物,且r可为h、烷基、芳基、经取代烷基和经取代芳基。示范性r基包括(但不限于)-ch2、-c(ch3)3、-or'、-nr'r"、-sr'、-卤素、-c(o)r'、-conr'r"、-s(o)2r'、-s(o)2nr'r"、-cn和-no2。r'、r"、r'"和r""各独立地指氢、经取代或未经取代的杂烷基、经取代或未经取代的芳基(包括(但不限于)经1到3个卤素取代的芳基)、经取代或未经取代的烷基、烷氧基或硫烷氧基,或芳基烷基。当本发明化合物包括一个以上r基团时,例如,各r基团是如各r'、r"、r'"和r""基团当存在超过一个时那样独立地选择。当r'与r"连接到同一氮原子时,其可与所述氮原子组合形成5元、6元或7元环。举例来说,-nr'r"意欲包括(但不限于)1-吡咯烷基和4-吗啉基。根据上文有关取代基的论述,所属领域技术人员将了解到,术语“烷基”意欲包括包含与除氢基外的基团结合的碳原子的基团,例如卤烷基(包括(但不限于)-cf3和-ch2cf3)和酰基(包括(但不限于)-c(o)ch3、-c(o)cf3、-c(o)ch2och3等)。
叠氮官能团还可与含有硫酯且经芳基膦部分适当官能化的水溶性聚合物选择性反应,从而产生酰胺键。芳基膦基在原位还原叠氮基,随后所得胺与硫酯键有效反应,产生相应酰胺。含有硫酯和膦部分的示范性水溶性聚合物可如下所示:
其中n为1-10;x可为o、n、s,或不存在,ph为苯基,且w为水溶性聚合物。
示范性含炔氨基酸可如下所示:
其中n为0到10;r1为烷基、芳基、经取代烷基或经取代芳基,或不存在;x为o、n、s,或不存在;m为0到10;r2为h、氨基酸、多肽或氨基末端修饰基团;且r3为h、氨基酸、多肽或羧基末端修饰基团。在一些实施例中,n为1,r1为苯基,x不存在,m为0,且乙炔部分位于烷基侧链的对位。在一些实施例中,n为1,r1为苯基,x为o,m为1,且炔丙氧基位于烷基侧链的对位(即,o-炔丙基-酪氨酸)。在一些实施例中,n为1,r1和x不存在,且m为0(即,炔丙基甘氨酸)。
含炔氨基酸在市面上有售。举例来说,炔丙基甘氨酸可购自派普科技公司(马萨诸塞州伯灵顿)。或者,可根据标准方法制备含炔氨基酸。举例来说,可例如根据戴特斯a.(deiters,a.)等人,美国化学协会杂志,125:11782-11783(2003)中所述,合成对炔丙氧基苯丙氨酸,并且可根据凯瑟b.(kayser,b.)等人,四面体通讯53(7):2475-2484(1997)中所述,合成4-炔基-l-苯丙氨酸。所属领域技术人员可制备出其它含炔氨基酸。
示范性含叠氮基氨基酸可如下所示:
其中n为0到10;r1为烷基、芳基、经取代烷基、经取代芳基,或不存在;x为o、n、s,或不存在;m为0到10;r2为h、氨基酸、多肽,或氨基末端修饰基团;且r3为h、氨基酸、多肽,或羧基末端修饰基团。在一些实施例中,n为1,r1为苯基,x不存在,m为0,且叠氮部分位于烷基侧链的对位。在一些实施例中,n为0到4,且r1和x不存在,且m=0。在一些实施例中,n为1,r1为苯基,x为o,m为2,且β-叠氮基乙氧基部分位于烷基侧链的对位。
含叠氮基氨基酸可从商业来源得到。举例来说,4-叠氮基苯丙氨酸可从凯姆英派国际公司(chem-impexinternational,inc.)(伊利诺伊州伍德戴尔(wooddale,il))获得。对于非市售的含叠氮基氨基酸,可使用所属领域技术人员已知的标准方法,包括(但不限于)经由置换适当离去基团(包括(但不限于)卤基、甲磺酸酯基、甲苯磺酸酯基)或经由使适当保护的内酯开环,相对容易地制备出叠氮基。例如参见马奇的高等有机化学(第3版,1985,威立父子出版公司,纽约)。
e.氨基硫醇反应性基团
β取代的氨基硫醇官能团的独特反应性使其极其适用于经由形成噻唑烷选择性修饰含有醛基的多肽和其它生物分子。例如参见j.邵(j.shao)和j.泰姆(j.tam),美国化学协会杂志,1995,117(14)3893-3899。在一些实施例中,可将β取代的氨基硫醇氨基酸并入bst多肽中,随后与包含醛官能团的水溶性聚合物反应。在一些实施例中,可经由形成噻唑烷将水溶性聚合物、药物结合物或其它有效负荷与包含β取代的氨基硫醇氨基酸的bst多肽偶合。
f.其它反应性基团
以下专利公开案(都以全文引用的方式并入本文中)中描述可并入本发明bst多肽中的其它反应性基团和非天然编码氨基酸(包括(但不限于)对氨基-苯丙氨酸):美国专利公开案第2006/0194256号、美国专利公开案第2006/0217532号、美国专利公开案第2006/0217289号、美国临时专利第60/755,338号、美国临时专利第60/755,711号、美国临时专利第60/755,018号、国际专利申请案第pct/us06/49397号、wo2006/069246、美国临时专利第60/743,041号、美国临时专利第60/743,040号、国际专利申请案第pct/us06/47822号、美国临时专利第60/882,819号、美国临时专利第60/882,500号,和美国临时专利第60/870,594号。这些申请案中也论述了peg或其它聚合物上可能存在的供结合的反应性基团,包括(但不限于)羟胺(氨氧基)。
非天然氨基酸的细胞摄取
细胞对非天然氨基酸的摄取是在设计和选择(包括(但不限于))并入蛋白质中的非天然氨基酸时常常考虑的一个问题。举例来说,α-氨基酸的高电荷密度表明,这些化合物不可能渗透细胞。天然氨基酸是经由一系列基于蛋白质的转运系统吸收到真核细胞中。可进行快速筛选来评估哪些非天然氨基酸(如果存在)被细胞吸收。例如参见,例如美国专利公开案第us2004/0198637号,名为“蛋白质阵列”(其以引用的方式并入本文中);以及林d.r.(liu,d.r.)和查鲁兹p.g.(schultz,p.g.)(1999)具有扩充的遗传密码的生物体的进化发展(progresstowardtheevolutionofanorganismwithanexpandedgeneticcode.)美国国家科学院院刊(pnasunitedstates)96:4780-4785中的毒性分析。尽管易于利用各种分析法来分析摄取,但设计适用于细胞摄取路径的非天然氨基酸的替代方法是提供在体内产生氨基酸的生物合成路径。
非天然氨基酸的生物合成
细胞中已存在许多生物合成路径来产生氨基酸和其它化合物。特定非天然氨基酸的生物合成方法可能不存在于自然界(包括(但不限于)细胞)中,而本发明将提供此类方法。举例来说,在宿主细胞中,通过添加新颖酶,或改变现存的宿主细胞路径,来任选产生非天然氨基酸的生物合成路径。其它新颖酶任选为天然存在酶或人工开发酶。举例来说,对氨基苯丙氨酸的生物合成(如名为“体内并入非天然氨基酸”的wo2002/085923中的实例所提供)依赖于添加来自其它生物体的已知酶的组合。可通过用包含这些酶的基因的质粒转化细胞来将所述基因引入真核细胞中。当在细胞中表达时,这些基因提供合成所需化合物的酶促路径。任选添加的酶类型的实例提供于下文的实例中。其它酶序列可见于例如基因库(genbank)中。也任选以相同方式将人工开发酶添加到细胞中。以此方式,操纵细胞的细胞机器和资源产生非天然氨基酸。
多种方法可用于产生用于生物合成路径中或用于发展现存路径的新颖酶。举例来说,任选使用包括(但不限于)麦克西根公司(maxygen,inc.)开发的递归重组法(recursiverecombination)(可见于万维网maxygen.com)来开发新颖酶和路径。例如参见斯蒂姆(stemmer)(1994),在体外利用dna改组快速进化蛋白质(rapidevolutionofaproteininvitrobydnashuffling),自然370(4):389-391;和斯蒂姆,(1994),通过随机断裂和再组装进行dna改组:体外重组以供分子进化(dnashufflingbyrandomfragmentationandreassembly:invitrorecombinationformolecularevolution),美国国家科学院院刊91:10747-10751。类似地,任选使用杰能科公司(genencor)(可见于万维网genencor.com)开发的designpathtm进行代谢路径的工程改造,包括(但不限于)工程改造某条路径以在细胞中产生o-甲基-l-酪氨酸。这项技术使用新基因(包括(但不限于)利用功能性基因组学以及分子进化和设计所鉴别的基因)的组合在宿主生物体中重建现存路径。迪文萨公司(diversacorporation)(可见于万维网diversa.com)也提供迅速筛选基因文库和基因路径(包括(但不限于)建立新路径)的技术。
通常,利用本发明的经工程改造的生物合成路径所产生的非天然氨基酸的浓度足以进行有效蛋白质生物合成(包括(但不限于)天然细胞量),但未达到影响其它氨基酸的浓度或耗尽细胞资源的程度。在体内以此方式产生的典型浓度为约10mm到约0.05mm。在用包含用于产生特定路径所需的酶的基因的质粒转化细胞,并产生非天然氨基酸后,任选使用体内选择,以针对核糖体蛋白质合成和细胞生长进一步优化产生非天然氨基酸。
具有非天然氨基酸的多肽
并入非天然氨基酸可实现多个目的,包括(但不限于)定制蛋白质结构和/或功能的改变;改变尺寸、酸度、亲核性、氢键结、疏水性、蛋白酶目标位点的可接近性;靶向部分(包括(但不限于)用于蛋白质阵列);添加生物活性分子;连接聚合物;连接放射性核素;调节血清半衰期;调节组织渗透(例如肿瘤);调节主动转运;调节组织、细胞或器官特异性或分布;调节免疫原性;调节蛋白酶抗性等。包括非天然氨基酸的蛋白质可具有较强的或甚至全新的催化或生物物理特性。举例来说,任选通过在蛋白质中包括非天然氨基酸来改变以下特性:毒性、生物分布、结构特性、光谱特性、化学和/或光化学特性、催化能力、半衰期(包括(但不限于)血清半衰期)、与其它分子反应(包括(但不限于)共价或非共价)的能力等。包括包含至少一个非天然氨基酸的蛋白质的组合物适用于(包括(但不限于))新颖治疗剂、诊断剂、催化酶、工业酶、结合蛋白质(包括(但不限于)抗体)以及(包括(但不限于))蛋白质结构和功能研究。例如参见道格悌(dougherty),(2000)作为蛋白质结构和功能探针的非天然氨基酸(unnaturalaminoacidsasprobesofproteinstructureandfunction),现代化学生物学观点(currentopinioninchemicalbiology.)4:645-652。
在本发明一方面中,组合物包括至少一种具有至少一个(包括(但不限于)至少两个、至少三个、至少四个、至少五个、至少六个、至少七个、至少八个、至少九个或至少十个或更多个非天然氨基酸的蛋白质。这些非天然氨基酸可相同或不同,包括(但不限于)在蛋白质中的1、2、3、4、5、6、7、8、9、10个或更多个不同位点可包含1、2、3、4、5、6、7、8、9、10个或更多个不同的非天然氨基酸。另一方面,组合物包括蛋白质中存在的至少一个(但少于全部)特定氨基酸经非天然氨基酸取代的蛋白质。对于具有一个以上非天然氨基酸的指定蛋白质,非天然氨基酸可相同或不同(包括(但不限于),所述蛋白质可包括两种或两种以上不同类型的非天然氨基酸,或者可包括两个相同的非天然氨基酸)。对于具有两个以上非天然氨基酸的指定蛋白质,非天然氨基酸可相同、不同,或为多个相同种类的非天然氨基酸与至少一个不同非天然氨基酸的组合。
具有至少一个非天然氨基酸的相关蛋白质或多肽为本发明的一个特征。本发明还包括使用本发明组合物和方法产生的具有至少一个非天然氨基酸的多肽或蛋白质。赋形剂(包括(但不限于)医药学上可接受的赋形剂)也可与蛋白质一起存在。
通过在真核细胞中产生具有至少一个非天然氨基酸的相关蛋白质或多肽,蛋白质或多肽通常会包括真核翻译后修饰。在某些实施例中,蛋白质包括至少一个非天然氨基酸和至少一个由真核细胞在体内进行的翻译后修饰,其中所述翻译后修饰不是由原核细胞进行的。举例来说,翻译后修饰包括(包括(但不限于))乙酰化、酰基化、脂质修饰、棕榈酰化、棕榈酸盐加成、磷酸化、糖脂键修饰、糖基化等。一方面,翻译后修饰包括寡糖(包括(但不限于)(glcnac-man)2-man-glcnac-glcnac))经由glcnac-天冬酰胺键与天冬酰胺连接。参见表1,其列出了真核蛋白质的n连接寡糖的一些实例(也可存在其它未显示的残基)。另一方面,翻译后修饰包括寡糖(包括(但不限于)gal-galnac、gal-glcnac等)经由galnac-丝氨酸或galnac-苏氨酸键或者glcnac-丝氨酸或glcnac-苏氨酸键与丝氨酸或苏氨酸连接。
表1:经由glcnac键连接的寡糖实例
另一方面,翻译后修饰包括前体(包括(但不限于)降钙素前体、降钙素基因相关性肽前体、前甲状旁腺激素原、前胰岛素原、前胰岛素、前阿黑皮素原(prepro-opiomelanocortin)、阿黑皮素原等)的蛋白水解处理;组装成多亚单元蛋白质或大分子组装物;翻译到细胞中另一位点(包括(但不限于)细胞器,例如内质网、高尔基体(golgiapparatus)、核、溶酶体、过氧化物酶体、线粒体、叶绿体、液泡等,或通过分泌路径)。在某些实施例中,蛋白质包含分泌或定位序列、表位标签、flag标签、聚组氨酸标签、gst融合体等。
非天然氨基酸的一个优势在于,其提供可用于添加其它分子的其它化学部分。这些修饰可在真核或非真核细胞体内进行或在体外进行。因此,在某些实施例中,翻译后修饰是通过非天然氨基酸进行。举例来说,翻译后修饰可通过亲核-亲电反应进行。当前用于选择性修饰蛋白质的大部分反应涉及亲核与亲电反应搭配物之间的共价键形成,包括(但不限于)α-卤代酮与组氨酸或半胱氨酸侧链的反应。在这些情况中,选择性是由蛋白质中亲核残基的数量和可接近性决定。在本发明蛋白质中,可在体外和体内使用其它更具选择性的反应,例如非天然酮基氨基酸与酰肼或氨氧基化合物的反应。例如参见柯尼西(cornish)等人(1996),美国化学协会杂志118:8150-8151;马赫尔(mahal)等人(1997),科学276:1125-1128;,王(wang)等人(2001),科学292:498-500;秦(chin)等人(2002),美国化学协会杂志124:9026-9027;秦等人(2002),美国国家科学院院刊.99:11020-11024;王等人(2003),美国国家科学院院刊,100:56-61;张(zhang)等人(2003),生物化学(biochemistry),42:6735-6746;和秦等人(2003),科学,301:964-7,所有这些文献都以引用的方式并入本文中。这使得能够用包括荧光团、交联剂、糖衍生物和细胞毒性分子在内的多种试剂选择性标记几乎任何蛋白质。还参见美国专利第6,927,042号,名为“糖蛋白的合成(glycoproteinsynthesis)”,以引用的方式并入本文中。翻译后修饰(包括(但不限于)经由叠氮基氨基酸修饰)也可通过施陶丁格尔接合(staudingerligation)(包括(但不限于)用三芳基膦试剂)进行。例如参见柯克等人,(2002)利用施陶丁格尔接合将叠氮基并入重组蛋白中以进行化学选择性修饰(incorporationofazidesintorecombinantproteinsforchemoselectivemodificationbythestaudingerligtation),pnas99:19-24。
本发明提供另一种选择性修饰蛋白质的高效方法,其涉及对选择密码子反应来将(包括(但不限于))含叠氮基或含炔基部分的非天然氨基酸以遗传方式并入蛋白质中。随后,可通过与(包括(但不限于))炔基或叠氮衍生物分别发生(包括(但不限于))胡伊斯根[3+2]环加成反应(例如参见帕德瓦a.,有机合成大全,第4卷.(1991)托斯特b.m.编,波格蒙出版社(pergamon),牛津,第1069-1109页;和胡伊斯根r.1,3-偶极环加成化学,(1984),帕德瓦a.编,威立出版公司,纽约,第1-176页)来修饰这些氨基酸侧链。由于此方法涉及环加成而非亲核取代反应,故可以极高的选择性修饰蛋白质。所述反应可在室温下,于水性条件中,通过将催化量的cu(i)盐添加到反应混合物中以优良的区位选择性(1,4>1,5)进行。例如参见托尼诺(tornoe)等人,(2002)有机化学杂志67:3057-3064;和罗斯托瑟夫(rostovtsev)等人,(2002)德国应用化学41:2596-2599。使用的另一方法是具有四半胱氨酸(tetracysteine)基元的双砷化合物的配体交换,例如参见格里夫林(griffin)等人,(1998)科学281:269-272。
可经由[3+2]环加成反应加到本发明蛋白质中的分子包括实际上任何具有叠氮基或炔基衍生物的分子。分子包括(但不限于)染料、荧光团、交联剂、糖衍生物、聚合物(包括(但不限于)聚乙二醇衍生物)、光交联剂、细胞毒性化合物、亲和标记、生物素衍生物、树脂、珠粒、第二蛋白质或多肽(或更多)、聚核苷酸(包括(但不限于)dna、rna等)、金属螯合剂、辅因子、脂肪酸、碳水化合物等。这些分子可分别添加到具有炔基(包括(但不限于)对炔丙氧基苯丙氨酸)或叠氮基(包括(但不限于)对叠氮基-苯丙氨酸)的非天然氨基酸中。
v.体内产生包含非天然编码氨基酸的bst多肽
可以在体内使用经修饰trna和trna合成酶,添加或取代天然存在系统中不编码的氨基酸,来产生本发明bst多肽。
产生可使用天然存在系统中不编码的氨基酸的trna和trna合成酶的方法描述于例如美国专利第7,045,337号和第7,083,970号中,其以引用的方式并入本文中。这些方法涉及产生独立于翻译系统内源性合成酶和trna起作用(因此有时称为“正交”)的翻译机器。通常,翻译系统包含正交trna(o-trna)和正交氨酰基trna合成酶(o-rs)。通常,o-rs优先使翻译系统中具有至少一个非天然存在氨基酸的o-trna氨酰基化,并且o-trna识别至少一个不被所述系统中的其它trna所识别的选择密码子。因此,翻译系统对所编码的选择密码子反应而将非天然编码氨基酸插入所述系统中产生的蛋白质中,由此将氨基酸“取代”入编码多肽的某一位置中。
此项技术中已经描述多种用于将特定合成氨基酸插入多肽中的正交trna和氨酰基trna合成酶,并且其一般都适用于本发明中。举例来说,酮基特异性o-trna/氨酰基trna合成酶描述于王l(wang,l)等人,美国国家科学院院刊,100:56-61(2003)和张z.(zhang,z.)等人,生物化学(biochem.)42(22):6735-6746(2003)中。示范性o-rs或其部分是由聚核苷酸序列编码,并且包括美国专利第7,045,337号和第7,083,970号(各自以引用的方式并入本文)中所揭示的氨基酸序列。与o-rs一起使用的相应o-trna分子也描述于美国专利第7,045,337号和第7,083,970号中,其以引用的方式并入本文中。o-trna/氨酰基trna合成酶对的其它实例描述于wo2005/007870、wo2005/007624和wo2005/019415中。
叠氮基特异性o-trna/氨酰基trna合成酶系统的实例描述于秦j.w.等人,美国化学协会杂志,124:9026-9027(2002)中。对叠氮基-l-phe的示范性o-rs序列包括(但不限于)美国专利第7,083,970号(以引用的方式并入本文中)中所揭示的核苷酸序列seqidno:14-16和29-32以及氨基酸序列seqidno:46-48和61-64。适用于本发明中的示范性o-trna序列包括(但不限于)美国专利第7,083,970号(以引用的方式并入本文中)中所揭示的核苷酸序列seqidno:1-3。对特定非天然编码氨基酸具特异性的o-trna/氨酰基trna合成酶对的其它实例描述于美国专利第7,045,337号中,其以引用的方式并入本文中。在酿酒酵母中并入含酮基氨基酸与含叠氮基氨基酸的o-rs和o-trna描述于秦j.w.等人,科学,301:964-967(2003)中。
已经报导了数种其它正交对。已针对在大肠杆菌中并入非天然氨基酸的可能性描述了来源于酿酒酵母菌trna的谷氨酰胺酰基(例如参见刘d.r.(liu,d.r.)和查鲁兹p.g.(schultz,p.g.)(1999)美国国家科学院院刊96:4780-4785)、天冬氨酰基(例如参见帕斯纳克m.(pastrnak,m.)等人,(2000)瑞士化学学报(helv.chim.acta)83:2277-2286)和酪氨酰基(例如参见大野s.(ohno,s.)等人,(1998)生物化学杂志(日本东京)(j.biochem.(tokyo,jpn.))124:1065-1068;和卡瓦尔a.k.(kowal,a.k.)等人,(2001)美国国家科学院院刊98:2268-2273)系统和合成酶。也已描述将来源于大肠杆菌谷氨酰胺酰基(例如参见卡瓦尔a.k.(kowal,a.k.)等人,(2001)美国国家科学院院刊98:2268-2273)和酪氨酰基(例如参见爱德华兹h.(edwards,h.)和斯奇莫p.(schimmel,p.)(1990)分子细胞生物学(mol.cell.biol.)10:1633-1641)合成酶的系统用于酿酒酵母中。已使用大肠杆菌酪氨酰基系统在体内将3-碘-l-酪氨酸并入哺乳动物细胞中。参见坂本k.(sakamoto,k.)等人,(2002)核酸研究30:4692-4699。
使用o-trna/氨酰基trna合成酶涉及选择编码非天然编码氨基酸的特定密码子。尽管可使用任何密码子,但一般理想地选择在细胞中很少或从未用于表达o-trna/氨酰基trna合成酶的密码子。举例来说,示范性密码子包括无义密码子,例如终止密码子(琥珀、赭石和蛋白石密码子);四个或四个以上碱基的密码子;和稀有或从未使用的其它天然三碱基密码子。
可使用此项技术中已知的诱变方法(包括(但不限于)位点特异性诱变、盒式诱变、限制性选择性诱变等),将特定的选择密码子引入bst聚核苷酸编码序列的适当位置中。
用于产生可用于并入非天然编码氨基酸的蛋白质合成机器组分(例如o-rs、o-trna和正交o-trna/o-rs对)的方法描述于王l.(wang,l.)等人,科学,292:498-500(2001);秦j.w.等人,美国化学协会杂志,124:9026-9027(2002);张z(zhang,z)等人,生物化学(biochemistry)42:6735-6746(2003)中。用于体内并入非天然编码氨基酸的方法和组合物描述于美国专利第7,045,337号中,其以引用的方式并入本文中。用于选择在生物体体内翻译系统中使用的正交trna-trna合成酶对的方法也描述于美国专利第7,045,337号和第7,083,970号中,所述专利以引用的方式并入本文中。pct公开案第w004/035743号,名为“酮基氨基酸位点特异性并入蛋白质中(sitespecificincorporationofketoaminoacidsintoproteins)”(以全文引用的方式并入本文中),描述用于并入酮基氨基酸的正交rs和trna对。pct公开案第wo04/094593号,名为“扩充真核遗传密码(expandingtheeukaryoticgeneticcode)”(其以全文引用的方式并入本文中),描述用于将非天然编码氨基酸并入真核宿主细胞中的正交rs和trna对。
产生至少一种重组正交氨酰基trna合成酶(o-rs)的方法包含:(a)由第一生物体产生来源于至少一种氨酰基trna合成酶(rs)的(任选突变)rs文库,所述第一生物体包括(但不限于)原核生物体(例如詹氏甲烷球菌、嗜热自养甲烷杆菌、嗜盐菌、大肠杆菌、超嗜热古菌、强烈嗜热球菌、堀越火球菌(p.horikoshii)、嗜热泉生古细菌、极端嗜热细菌等)或真核生物体;(b)在rs(任选突变rs)文库中选择(和/或筛选)在非天然编码氨基酸和天然氨基酸存在下使正交trna(o-trna)氨酰基化的成员,由此提供活性(任选突变)rs的池;和/或(c)在所述池中选择(任选通过阴性选择)在不存在非天然编码氨基酸的情况下优先使o-trna氨酰基化的活性rs(包括(但不限于)突变rs),由此提供所述至少一种重组o-rs;其中所述至少一种重组o-rs优先使具有非天然编码氨基酸的o-trna氨酰基化。
在一个实施例中,rs为无活性rs。可通过使活性rs突变来产生无活性rs。举例来说,可通过使至少约1个、至少约2个、至少约3个、至少约4个、至少约5个、至少约6个或至少约10个或更多个氨基酸突变成不同氨基酸(包括(但不限于)丙氨酸)来产生无活性rs。
可使用此项技术中已知的各种技术产生突变rs文库,所述技术包括(但不限于)基于蛋白质三维rs结构的合理设计,或在随机或合理设计技术中rs核苷酸的诱变。举例来说,可利用位点特异性突变、随机突变、产生多样性的重组突变、嵌合构建体、合理设计和本文中所述或此项技术中已知的其它方法产生突变rs。
在一个实施例中,在rs(任选突变rs)文库中选择(和/或筛选)(包括(但不限于))在非天然编码氨基酸和天然氨基酸存在下使正交trna(o-trna)氨酰基化的活性成员包括:将阳性选择或筛选标记物(包括(但不限于)抗生素抗性基因等)和(任选突变)rs文库引入多个细胞中,其中所述阳性选择和/或筛选标记物包含至少一个选择密码子(包括(但不限于)琥珀密码子、赭石密码子或蛋白石密码子);使所述多个细胞在选择剂存在下生长;通过抑制阳性选择或筛选标记物中的所述至少一个选择密码子来鉴别在选择剂和/或筛选剂存在下存活(或显示特异性反应)的细胞,从而提供含有活性(任选突变)rs的池的经阳性选择的细胞的子集。任选可改变选择剂和/或筛选剂的浓度。
一方面,阳性选择标记物为氯霉素(chloramphenicol)乙酰基转移酶(cat)基因,且在cat基因中,选择密码子为琥珀终止密码子。阳性选择标记物任选为β-内酰胺酶基因,且在β-内酰胺酶基因中,选择密码子为琥珀终止密码子。另一方面,阳性筛选标记物包含荧光或发光筛选标记物,或基于亲和性的筛选标记物(包括(但不限于)细胞表面标记物)。
在一个实施例中,在池中阴性选择或筛选在不存在非天然编码氨基酸的情况下优先使o-trna氨酰基化的活性rs(任选突变体)包括:将阴性选择或筛选标记物与来自阳性选择或筛选的活性(任选突变)rs的池引入第二生物体的多个细胞中,其中阴性选择或筛选标记物包含至少一个选择密码子(包括(但不限于)抗生素抗性基因,包括(但不限于)氯霉素乙酰基转移酶(cat)基因);以及鉴别在补充有非天然编码氨基酸和筛选剂或选择剂的第一培养基中存活或显示特异性筛选反应,但在未补充非天然编码氨基酸和选择剂或筛选剂的第二培养基中不能存活或不显示特异性反应的细胞,从而提供具有至少一种重组o-rs的存活细胞或经筛选细胞。举例来说,在测定适当o-rs重组体时,cat鉴别方案任选充当阳性选择和/或阴性筛选。举例来说,在存在或不存在一个或一个以上非天然编码氨基酸的情况下,任选于含有cat(其包含至少一个选择密码子)的生长板上复制克隆池。因此,认为仅在含有非天然编码氨基酸的板上生长的集落含有重组o-rs。一方面,改变选择(和/或筛选)剂的浓度。在一些方面中,第一生物体与第二生物体不同。因此,第一和/或第二生物体任选包含:原核生物、真核生物、哺乳动物、大肠杆菌、真菌、酵母、古细菌、真细菌、植物、昆虫、原生生物等。在其它实施例中,筛选标记物包含荧光或发光筛选标记物,或基于亲和性的筛选标记物。
在另一个实施例中,在池中筛选或选择(包括(但不限于)阴性选择)活性(任选突变)rs包括:自阳性选择步骤(b)分离出活性突变rs的池;将阴性选择或筛选标记物和活性(任选突变)rs的池引入第二生物体的多个细胞中,其中阴性选择或筛选标记物包含至少一个选择密码子(包括(但不限于)包含至少一个选择密码子的毒性标记基因,包括(但不限于)核糖核酸酶barnase基因);以及鉴别在未补充非天然编码氨基酸的第一培养基中存活或显示特异性筛选反应,但在补充有非天然编码氨基酸的第二培养基中不能存活或不显示特异性筛选反应的细胞,从而提供具有至少一种重组o-rs的存活细胞或经筛选细胞,其中所述至少一种重组o-rs对非天然编码氨基酸具特异性。一方面,所述至少一个选择密码子包含约两个或两个以上选择密码子。这些实施例任选可包括所述至少一个选择密码子包含两个或两个以上选择密码子,且第一生物体与第二生物体不同(包括(但不限于),各生物体任选为(包括(但不限于))原核生物、真核生物、哺乳动物、大肠杆菌、真菌、酵母、古细菌、真细菌、植物、昆虫、原生生物等)的情形。另外,一些方面包括阴性选择标记物包含核糖核酸酶barnase基因(其包含至少一个选择密码子)的情形。其它方面包括筛选标记物任选包含荧光或发光筛选标记物或基于亲和性的筛选标记物的情形。在本文中的实施例中,筛选和/或选择任选包括筛选和/或选择严格度的变化。
在一个实施例中,用于产生至少一种重组正交氨酰基-trna合成酶(o-rs)的方法可进一步包含:(d)分离所述至少一种重组o-rs;(e)产生来源于所述至少一种重组o-rs的第二组o-rs(任选突变);以及(f)重复步骤(b)和(c),直到获得能够优先使o-trna氨酰基化的突变o-rs。任选将步骤(d)到(f)重复(包括(但不限于))至少约两次。一方面,可通过诱变(包括(但不限于)随机诱变、位点特异性诱变、重组或其组合)产生来源于至少一种重组o-rs的第二组突变o-rs。
上述方法中选择/筛选步骤(包括(但不限于)阳性选择/筛选步骤(b)、阴性选择/筛选步骤(c)或阳性与阴性选择/筛选步骤(b)与(c))的严格度任选包括改变选择/筛选严格度。在另一个实施例中,阳性选择/筛选步骤(b)、阴性选择/筛选步骤(c)或阳性与阴性选择/筛选步骤(b)与(c)包含使用报告基因,其中报告基因是利用荧光活化细胞分选法(facs)检测,或其中报告基因是通过发光检测。报告基因任选展示于细胞表面上、噬菌体展示上等,且其是根据涉及非天然编码氨基酸或其类似物的亲和性或催化活性选择。在一个实施例中,突变合成酶是展示于细胞表面上、噬菌体展示上等。
用于产生重组正交trna(o-trna)的方法包括:(a)由第一生物体产生来源于至少一种trna(包括(但不限于)抑制trna)的突变trna文库;(b)在所述文库中选择(包括(但不限于)阴性选择)或筛选在不存在第一生物体的rs的情况下由第二生物体的氨酰基trna合成酶(rs)氨酰基化的(任选突变)trna,从而提供trna(任选突变体)的池;以及(c)在所述trna(任选突变体)池中选择或筛选经引入的正交rs(o-rs)氨酰基化的成员,从而提供至少一种重组o-trna;其中所述至少一种重组o-trna识别选择密码子且并不由第二生物体的rs有效识别,并由o-rs优先氨酰基化。在一些实施例中,所述至少一种trna为抑制trna,和/或包含具有天然和/或非天然碱基的独特三碱基密码子,或为无义密码子、稀有密码子、非天然密码子、包含至少4个碱基的密码子、琥珀密码子、赭石密码子或蛋白石终止密码子。在一个实施例中,重组o-trna的正交性得到改进。应了解,在一些实施例中,无需修饰即可任选将o-trna从第二生物体引入第一生物体中。在各种实施例中,第一生物体与第二生物体相同或不同,并且任选选自(包括(但不限于))原核生物(包括(但不限于)詹氏甲烷球菌、嗜热自养甲烷杆菌、大肠杆菌、嗜盐菌等)、真核生物、哺乳动物、真菌、酵母、古细菌、真细菌、植物、昆虫、原生生物等。另外,重组trna任选经非天然编码氨基酸氨酰基化,其中所述非天然编码氨基酸是在体内天然生物合成的,或通过基因操作生物合成。任选将非天然编码氨基酸添加到至少第一或第二生物体的生长培养基中。
一方面,在文库中选择(包括(但不限于)阴性选择)或筛选经氨酰基trna合成酶氨酰基化的(任选突变)trna(步骤(b))包括:将毒性标记基因和(任选突变)trna文库引入第二生物体的多个细胞中,其中毒性标记基因包含至少一个选择密码子(或导致产生毒性剂或抑制剂(staticagent)的基因,或生物体必需的基因,其中所述标记基因包含至少一个选择密码子);和选择存活细胞,其中存活细胞含有包含至少一个正交trna或非功能性trna的(任选突变)trna的池。举例来说,可使用比较比率细胞密度分析(comparisonratiocelldensityassay)来选择存活细胞。
另一方面,毒性标记基因可包括两个或两个以上选择密码子。在所述方法的另一个实施例中,毒性标记基因为核糖核酸酶barnase基因,其中核糖核酸酶barnase基因包含至少一个琥珀密码子。核糖核酸酶barnase基因可任选包括两个或两个以上琥珀密码子。
在一个实施例中,在(任选突变)trna的池中选择或筛选经引入的正交rs(o-rs)氨酰基化的成员可包括:将阳性选择或筛选标记基因连同o-rs和(任选突变)trna的池引入第二生物体的多个细胞中,其中阳性标记基因包含药物抗性基因(包括(但不限于)β-内酰胺酶基因,其包含至少一个选择密码子,例如至少一个琥珀终止密码子)或生物体必需的基因,或使毒性剂解毒的基因;和鉴别在选择剂或筛选剂(包括(但不限于)抗生素)存在下生长的存活或经筛选细胞,从而提供具有至少一种重组trna的细胞的池,其中所述至少一种重组trna是由o-rs氨酰基化,并对所述至少一个选择密码子反应而将氨基酸插入由阳性标记基因编码的翻译产物中。在另一个实施例中,选择剂和/或筛选剂的浓度可变化。
本发明提供用于产生特异性o-trna/o-rs对的方法。所述方法包括:(a)由第一生物体产生来源于至少一种trna的突变trna的文库;(b)在所述文库中阴性选择或筛选在不存在第一生物体的rs的情况下由第二生物体的氨酰基trna合成酶(rs)氨酰基化的(任选突变)trna,从而提供(任选突变)trna的池;(c)在(任选突变)trna的池中选择或筛选经引入的正交rs(o-rs)氨酰基化的成员,从而提供至少一种重组o-trna。所述至少一种重组o-trna识别选择密码子,且并不由第二生物体的rs有效识别,并由o-rs优先氨酰基化。所述方法还包括(d)由第三生物体产生来源于至少一种氨酰基trna合成酶(rs)的(任选突变)rs的文库;(e)在突变rs文库中选择或筛选在非天然编码氨基酸和天然氨基酸存在下优先使所述至少一种重组o-trna氨酰基化的成员,从而提供活性(任选突变)rs的池;和(f)在所述池中阴性选择或筛选在不存在非天然编码氨基酸的情况下优先使所述至少一种重组o-trna氨酰基化的活性(任选突变)rs,从而提供至少一种特异性o-trna/o-rs对,其中所述至少一种特异性o-trna/o-rs对包含至少一种对非天然编码氨基酸具特异性的重组o-rs和至少一种重组o-trna。本发明包括由所述方法产生的特异性o-trna/o-rs对。举例来说,特异性o-trna/o-rs对可包括(包括(但不限于))mutrnatyr-muttyrrs对,例如mutrnatyr-ss12tyrrs对、mutrnaleu-mutleurs对、mutrnathr-mutthrrs对、mutrnaglu-mutglurs对等。另外,所述方法包括第一生物体与第三生物体相同(包括(但不限于)詹氏甲烷球菌)的情形。
本发明中还包括用于选择用于第二生物体的体内翻译系统中的正交trna-trna合成酶对的方法。所述方法包括:将标记基因、trna和从第一生物体分离或获得的氨酰基trna合成酶(rs)引入第二生物体的第一组细胞中;将标记基因和trna引入第二生物体的复制细胞组中;和选择在复制细胞组中不能存活且在第一组中存活的细胞,或筛选显示特异性筛选反应但在复制细胞组中不能提供这种反应的细胞,其中第一组与复制细胞组都是在选择剂或筛选剂存在下生长,其中存活或经筛选细胞包含用于第二生物体的体内翻译系统中的正交trna-trna合成酶对。在一个实施例中,比较和选择或筛选包括体内互补分析。选择剂或筛选剂的浓度可变化。
本发明的生物体包含多种生物体和多种组合。举例来说,本发明方法中的第一生物体与第二生物体可相同或不同。在一个实施例中,生物体任选为原核生物体,包括(但不限于)詹氏甲烷球菌、嗜热自养甲烷杆菌、嗜盐菌、大肠杆菌、超嗜热古菌、强烈嗜热球菌、堀越火球菌、嗜热泉生古细菌、极端嗜热菌等。或者,生物体任选包含真核生物体,包括(但不限于)植物(包括(但不限于)复杂植物,例如单子叶植物或双子叶植物)、藻类、原生生物、真菌(包括(但不限于)酵母等)、动物(包括(但不限于)哺乳动物、昆虫、节肢动物等)等。在另一个实施例中,第二生物体任选为原核生物体,包括(但不限于)詹氏甲烷球菌、嗜热自养甲烷杆菌、嗜盐菌、大肠杆菌、超嗜热古菌、嗜盐菌、强烈嗜热球菌、堀越火球菌、嗜热泉生古细菌、极端嗜热菌等。或者,第二生物体可为真核生物体,包括(但不限于)酵母、动物细胞、植物细胞、真菌、哺乳动物细胞等。在各个实施例中,第一生物体与第二生物体不同。
vi.bst多肽中非天然存在氨基酸的定位
本发明涵盖将一个或一个以上非天然存在氨基酸并入bst多肽中。可将一个或一个以上非天然存在氨基酸并入不破坏多肽活性的特定位置。这可以通过进行“保守”取代实现,包括(但不限于)用疏水性氨基酸取代疏水性氨基酸;用大体积氨基酸取代大体积氨基酸;用亲水性氨基酸取代亲水性氨基酸,和/或将非天然存在氨基酸插入非活性所需的位置中。
可使用多种生物化学和结构方法来选择bst多肽内供非天然编码氨基酸取代的合意位点。所属领域技术人员易于了解,多肽链中的任何位置都适于选择用于并入非天然编码氨基酸,并且选择可建立在合理设计的基础上或者通过随机选择以实现任何或并非特别需要的目的。选择的所需位点可用于产生具有任何所需特性或活性的bst分子,包括(但不限于)激动剂、超级激动剂、反向激动剂、拮抗剂、受体结合调节剂、受体活性调节剂;用于形成二聚体或多聚体;活性或特性相比天然分子无改变;或者操纵多肽的任何物理或化学特性,例如溶解性、聚集或稳定性。举例来说,可使用此项技术中已知的点突变分析、丙氨酸扫描、饱和诱变和生物活性筛选或同系物扫描法,来鉴别bst多肽的生物活性所必需的多肽位置。其它可用于鉴别供修饰bst多肽的残基的方法包括(但不限于)序列谱(sequenceprofiling)、转子文库选择、残基成对势(residuepairpotential)和使用自动蛋白质设计技术(proteindesign
也可检查含有缺失的bst多肽突变体的结构和活性,以确定可能耐受非天然编码氨基酸取代的蛋白质区域。以类似方式,可使用蛋白酶消化和单克隆抗体来鉴别bst负责结合受体的区域。莱顿等人(2001)jbc276(39)36779-36787描述利用hg-csf和其受体进行的抗体研究。一旦去除了可能无法耐受非天然编码氨基酸取代的残基,就可以检查所建议的取代在各剩余位置处的影响。可以由其它csf家族成员和csf受体的三维晶体结构产生模型。蛋白质数据库(proteindatabank,pdb;可见于万维网rcsb.org)是一个含有蛋白质和核酸大分子的三维结构数据的中央数据库。如果无法得到三维结构数据,那么可建立研究多肽二级结构和三级结构的模型。hg-csf的x射线结晶学和nmr结构可见于蛋白质数据库(其中pdbid为:1cd9、1pgr、1rhg、1gnc)以及美国专利第5,581,476号和第5,790,421号中,各专利以引用的方式并入本文中。因此,所属领域技术人员可容易地鉴别出可经非天然编码氨基酸取代的氨基酸位置。
在一些实施例中,本发明的bst多肽包含一个或一个以上位于不破坏多肽结构的蛋白质区域中的非天然存在氨基酸。
并入非天然编码氨基酸的示范性残基可为以下残基:排除在潜在受体结合区外;可完全或部分溶剂暴露;与邻近残基具有最低限度或无氢键结相互作用;可在最低限度上暴露于邻近反应性残基;可在一个或一个以上暴露面上;可为一个或一个以上与第二bst或者其它分子或其片段毗邻的位点;如由结合或未结合于受体或者与另一生物活性分子偶合或未偶合的bst的三维二级、三级或四级结构所预测,可在具有高度柔性或刚性结构的区域中;或者可通过视需要改变完整结构的柔性或刚性来调节bst本身或者包含一个或一个以上bst的二聚体或多聚体的构象。
所属领域技术人员应认识到,这种针对bst的分析能够确定与埋在蛋白质三级结构内的氨基酸残基相比较,何种氨基酸残基为表面暴露的。因此,本发明的一个实施例是用非天然编码氨基酸取代作为表面暴露残基的氨基酸。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入st中的以下一个或一个以上位置:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16,17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:2)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bgh中的以下一个或一个以上位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191(即,蛋白质的羧基末端)和其任何组合。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的一个或一个以上位置(seqidno:1),且所述一个或一个以上非天然编码氨基酸不包括组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸,例如β-丙氨酸、鸟氨酸等。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的一个或一个以上位置(seqidno:2),且所述一个或一个以上非天然编码氨基酸不包括组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸,例如β-丙氨酸、鸟氨酸等。在一些实施例中,一个或一个以上所述位置处的一个或一个以上非天然编码氨基酸是除组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸(例如β-丙氨酸、鸟氨酸等)外的氨基酸,并在以下一个或一个以上位置出现:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154或其任何组合(seqidno:1)。在一些实施例中,一个或一个以上所述位置处的一个或一个以上非天然编码氨基酸是除组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸(例如β-丙氨酸、鸟氨酸等)外的氨基酸,并在以下一个或一个以上位置出现:3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173和其任何组合(seqidno:1)。在一些实施例中,一个或一个以上所述位置处的一个或一个以上非天然编码氨基酸是除组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸(例如β-丙氨酸、鸟氨酸等)外的氨基酸,并在以下一个或一个以上位置出现:35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154或其任何组合(seqidno:2)。在一些实施例中,一个或一个以上所述位置处的一个或一个以上非天然编码氨基酸是除组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸(例如β-丙氨酸、鸟氨酸等)外的氨基酸,并在以下一个或一个以上位置出现:3、7、11、33、43、58、62、67、69、98、99、123、124、125、133、134、136、141、159、166、169、170、173和其任何组合(seqidno:2)。在一些实施例中,bgh多肽中一个或一个以上所述位置处的一个或一个以上非天然编码氨基酸是除组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸(例如β-丙氨酸、鸟氨酸等)外的氨基酸,并在以下一个或一个以上位置出现:tyr35、gln91、phe92、ser94、arg95、asn99、leu101、arg133、ala134、leu138、lys139、gln140、tyr142、lys144、leu149、arg150、ala154或其任何组合。
在一些实施例中,一个或一个以上非天然编码氨基酸经核糖体并入bst的一个或一个以上位置(seqidno:1),且所述一个或一个以上非天然编码氨基酸不包括组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸,例如β-丙氨酸、鸟氨酸等。在一些实施例中,一个或一个以上非天然编码氨基酸经核糖体并入bst的一个或一个以上位置(seqidno:2),且所述一个或一个以上非天然编码氨基酸不包括组氨酸、精氨酸、赖氨酸、异亮氨酸、苯丙氨酸、亮氨酸、色氨酸、丙氨酸、半胱氨酸、天冬酰胺、缬氨酸、甘氨酸、丝氨酸、谷氨酰胺、酪氨酸、天冬氨酸、谷氨酸、苏氨酸,或天然存在的非成蛋白质性氨基酸,例如β-丙氨酸、鸟氨酸等。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的一个或一个以上位置(seqidno:1),其中所述一个或一个以上非天然编码氨基酸具有一个或一个以上不被内源rs识别的官能团。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的一个或一个以上位置(seqidno:2),其中所述一个或一个以上非天然编码氨基酸具有一个或一个以上不被内源rs识别的官能团。
对bst或bst家族成员的晶体结构以及其与bst和/或bgh受体相互作用的检查可指示哪些氨基酸残基的侧链完全或部分接近溶剂。这些位置的非天然编码氨基酸的侧链可远离蛋白质表面向外伸入溶剂中。
多种非天然编码氨基酸可取代bst多肽的指定位置或并入bst多肽的指定位置中。一般说来,根据对bst多肽或其它g-csf家族成员与其受体的三维晶体结构的检查,来选择供并入的特定非天然编码氨基酸,优选保守取代(即,用含芳基的非天然编码氨基酸,例如对乙酰基苯丙氨酸或o-炔丙基酪氨酸,取代phe、tyr或trp),以及希望引入bst多肽中的特定结合化学(例如,当希望实现与带有炔部分的水溶性聚合物的胡伊斯根[3+2]环加成,或与带有芳基酯的水溶性聚合物形成酰胺键,接着并入膦部分时,可引入4-叠氮基苯丙氨酸)。
在一实施例中,所述方法进一步包括将非天然氨基酸并入蛋白质中,其中所述非天然氨基酸包含第一反应性基团;和使所述蛋白质与包含第二反应性基团的分子(包括(但不限于)羟烷基淀粉(has)、羟乙基淀粉(hes)、标记、染料、聚合物、水溶性聚合物、聚乙二醇衍生物、光交联剂、放射性核素、细胞毒性化合物、药物、亲和标记、光亲和标记、反应性化合物、树脂、第二蛋白质或多肽或多肽类似物、抗体或抗体片段、金属螯合剂、辅因子、脂肪酸、碳水化合物、聚核苷酸、dna、rna、反义聚核苷酸、糖、水溶性树突状聚合物、环糊精、抑制性核糖核酸、生物材料、纳米粒子、自旋标记、荧光团、含金属部分、放射性部分、新颖官能团、与其它分子共价或非共价相互作用的基团、光笼蔽部分、光化辐射可激发部分、可光异构化部分、生物素、生物素衍生物、生物素类似物、并有重原子的部分、可化学裂解基团、可光裂解基团、延长的侧链、碳连接糖、氧化还原活性剂、氨基硫代酸、有毒部分、同位素标记的部分、生物物理探针、磷光基团、化学发光基团、电子致密基团、磁性基团、插入基团、发色团、能量转移剂、生物活性剂、可检测标记、小分子、量子点、纳米递质、放射性核苷酸、放射性递质、中子捕获剂,或上述各物的任何组合,或任何其它合乎需要的化合物或物质)接触。第一反应性基团与第二反应性基团发生[3+2]环加成反应,以将所述分子与非天然氨基酸连接。在一个实施例中,第一反应性基团为炔基或叠氮基部分,且第二反应性基团为叠氮基或炔基部分。举例来说,第一反应性基团为炔基部分(包括(但不限于)在非天然氨基酸对炔丙氧基苯丙氨酸中),且第二反应性基团为叠氮基部分。在另一实例中,第一反应性基团为叠氮基部分(包括(但不限于)在非天然氨基酸对叠氮基-l-苯丙氨酸中),且第二反应性基团为炔基部分。
在一些情况下,将非天然编码氨基酸取代与bst多肽内的其它添加、取代或缺失组合,以影响bst多肽的其它生物特质。在一些情况下,所述其它添加、取代或缺失可增加bst多肽的稳定性(包括(但不限于)对蛋白水解降解的抗性),或增加bst多肽对其受体的亲和力。在一些情况下,所述其它添加、取代或缺失可增加bst多肽的医药稳定性。在一些情况下,所述其它添加、取代或缺失可增加bst多肽的生物活性。在一些情况下,所述其它添加、取代或缺失可增加bst多肽的溶解性(包括(但不限于)当在大肠杆菌或其它宿主细胞中表达时)。在一些实施例中,在大肠杆菌或其它重组宿主细胞中表达后,添加、取代或缺失可增加bst多肽的溶解性。在一些实施例中,除选择在大肠杆菌或其它重组宿主细胞中表达后,并入的非天然氨基酸使多肽的溶解性增加的位点外,还选择供天然编码或非天然氨基酸取代的其它位点。在一些实施例中,bst多肽包含另一添加、取代或缺失,其调节对受体、结合蛋白或缔合配体的亲和力;在结合于受体后,调节信号转导;调节循环半衰期;调节释放或生物利用率;便利纯化;或者改进或改变特定投药途径。在一些实施例中,bst多肽包含增加bst变异体对其受体的亲和力的添加、缺失或取代。类似地,bst多肽可包含化学或酶裂解序列、蛋白酶裂解序列、反应性基团、抗体结合域(包括(但不限于)flag或聚组氨酸)或其它基于亲和性的序列(包括(但不限于)flag、聚组氨酸、gst等)或连接分子(包括(但不限于)生物素),其可改进检测(包括(但不限于)gfp)、纯化、通过组织或细胞膜的转运、前药释放或活化、bst尺寸减小或多肽的其它特质。
在一些实施例中,非天然编码氨基酸的取代产生bst拮抗剂。在一些实施例中,将非天然编码氨基酸取代或添加到受体结合所涉及的区域中。在一些实施例中,bst拮抗剂包含至少一个使bst充当拮抗剂的取代。在一些实施例中,bst拮抗剂包含与水溶性聚合物连接的非天然编码氨基酸,其存在于bst分子的受体结合区内。
在一些实施例中,非天然编码氨基酸的取代产生bst拮抗剂。在一些实施例中,bst拮抗剂包含与水溶性聚合物连接的非天然编码氨基酸,其存在于bst分子的受体结合区内。
在一些情况下,用一个或一个以上非天然编码氨基酸取代1、2、3、4、5、6、7、8、9、10或更多个氨基酸。在一些情况下,bst多肽进一步包括1、2、3、4、5、6、7、8、9、10或更多个天然存在氨基酸经一个或一个以上非天然编码氨基酸取代。举例来说,在一些实施例中,bst中的一个或一个以上残基经一个或一个以上非天然编码氨基酸取代。在一些情况下,一个或一个以上非天然编码残基与一种或一种以上分子量较低的线性或分支peg连接,由此得到相比与单一较高分子量的peg连接的物质有所增强的结合亲和力和与之相当的血清半衰期。
在一些实施例中,bst的以下残基中最多两个经一个或一个以上非天然编码氨基酸取代。
vii.非真核生物和真核生物中的表达
为了克隆的bst聚核苷酸的高水平表达,通常将编码本发明bst多肽的聚核苷酸亚克隆到一种表达载体中,这种表达载体含有可引导转录的强启动子、转录/翻译终止子,以及(如果就编码蛋白质的核酸来说)用于起始翻译的核糖体结合位点。所属领域技术人员已知适当的细菌启动子,且这些启动子例如描述于萨姆布鲁克等人和奥斯贝尔(ausubel)等人中。
用于表达本发明bst多肽的细菌表达系统可从(包括(但不限于))大肠杆菌、芽孢杆菌属(bacillussp.)、荧光假单胞菌、铜绿假单胞菌、恶臭假单胞菌和沙门氏菌(salmonella)中获得(帕尔瓦(palva)等人,基因(gene)22:229-235(1983);莫斯贝奇(mosbach)等人,自然(nature)302:543-545(1983))。这些表达系统的试剂盒在市面上有售。所属领域技术人员众所周知哺乳动物细胞、酵母和昆虫细胞的真核表达系统,且在市面上也有销售。在使用正交trna和氨酰基trna合成酶(上文所述)表达本发明bst多肽的情况中,用于表达的宿主细胞是根据其使用正交组分的能力来选择。示范性宿主细胞包括革兰氏阳性细菌(包括(但不限于)短小芽孢杆菌(b.brevis)、枯草杆菌(b.subtilis)或链霉菌(streptomyces))和革兰氏阴性细菌(大肠杆菌、荧光假单胞菌、铜绿假单胞菌、恶臭假单胞菌),以及酵母和其它真核细胞。如本文中所述,可使用包含o-trna/o-rs对的细胞。
本发明的真核宿主细胞或非真核宿主细胞能够合成较大有效量的包含非天然氨基酸的蛋白质。一方面,组合物任选包括(包括(但不限于))至少10微克、至少50微克、至少75微克、至少100微克、至少200微克、至少250微克、至少500微克、至少1毫克、至少10毫克、至少100毫克、至少1克或更多包含非天然氨基酸的蛋白质,或者可利用体内蛋白质制备方法达到的量(关于重组蛋白制备和纯化的细节如本文中所提供)。另一方面,组合物中存在的蛋白质在(包括(但不限于))细胞溶解产物、缓冲液、医药缓冲液或其它液体悬浮液(包括(但不限于)体积(包括(但不限于))在约1nl到约100l间的任何体积或更高体积)中的浓度任选为(包括(但不限于))每升至少10微克蛋白质、每升至少50微克蛋白质、每升至少75微克蛋白质、每升至少100微克蛋白质、每升至少200微克蛋白质、每升至少250微克蛋白质、每升至少500微克蛋白质、每升至少1毫克蛋白质,或每升至少10毫克蛋白质或更高的浓度。本发明的一个特征为,在真核细胞中产生大量(包括(但不限于)大于用其它方法(包括(但不限于)体外翻译)通常可能获得的量)包括至少一个非天然氨基酸的蛋白质。
本发明的真核宿主细胞或非真核宿主细胞能够生物合成较大有效量的包含非天然氨基酸的蛋白质。举例来说,产生的包含非天然氨基酸的蛋白质在细胞提取物、细胞溶菌液、培养基、缓冲液等中的浓度为(包括(但不限于))至少10微克/升、至少50微克/升、至少75微克/升、至少100微克/升、至少200微克/升、至少250微克/升或至少500微克/升、至少1毫克/升、至少2毫克/升、至少3毫克/升、至少4毫克/升、至少5毫克/升、至少6毫克/升、至少7毫克/升、至少8毫克/升、至少9毫克/升、至少10毫克/升、至少20、30、40、50、60、70、80、90、100、200、300、400、500、600、700、800、900毫克/升、1克/升、5克/升、10克/升或更多的蛋白质。
多种适于表达bst的载体在市面上有售。适用于真核宿主的表达载体包括(但不限于)包含来自sv40的表达控制序列的载体、牛乳头瘤病毒、腺病毒和细胞肥大病毒。这些载体包括pcdna3.1(+)\hyg(英杰公司(invitroge),美国加利福尼亚州卡尔斯巴德(carlsbad,calif.,usa))和pci-neo(斯塔杰公司(stratagene),美国加利福尼亚州拉霍亚(lajolla,calif.,usa))。细菌质粒,例如来自大肠杆菌的质粒(包括pbr322、pet3a和pet12a);宿主范围较广的质粒,例如rp4;噬菌体dna,例如λ噬菌体的各种衍生物,例如nm989,及其它dna噬菌体,例如m13和丝状单链dna噬菌体,都可以使用。2μ质粒和其衍生物、pot1载体(美国专利第4,931,373号,以引用的方式并入本文中)、pjso37载体(描述于欧凯尔(okkels),纽约科学院年报(ann.newyorkaced.sci.)782,202207,1996)和ppicza、b或c(英杰公司(invitrogen)),都可用于酵母宿主细胞中。昆虫细胞载体包括(但不限于)pvl941、pbg311(凯特(cate)等人,“缪勒式抑制物质的牛和人类基因的分离以及人类基因在动物细胞中的表达(isolationofthebovineandhumangenesformullerianinhibitingsubstanceandexpressionofthehumangeneinanimalcells)”,细胞(cell),45,第685-698页(1986))、pbluebac4.5和pmelbac(英杰公司,加利福尼亚州卡尔斯巴德(carlsbad,ca))。
编码bst多肽的核苷酸序列也可包括或可不包括编码信号肽的序列。当多肽是由表达其的细胞分泌时,存在信号肽。所述信号肽可以为任何序列。信号肽可以是原核或真核的。科勒玛m(coloma,m)(1992)(免疫学方法杂志(j.imm.methods)152:89104)描述用于哺乳动物细胞中的信号肽(鼠类igκ轻链信号肽)。其它信号肽包括(但不限于)来自酿酒酵母菌的α-因子信号肽(美国专利第4,870,008号,以引用的方式并入本文中)、小鼠唾液淀粉酶信号肽(o.哈根布奇尔(o.hagenbuchle)等人,自然(nature)289,1981,第643-646页)、经修饰的羧肽酶信号肽(l.a.威尔斯(l.a.vails)等人,细胞(cell)48,1987,第887-897页)、酵母bar1信号肽(wo87/02670,以引用的方式并入本文中)和酵母天冬氨酸蛋白酶3(yap3)信号肽(参见m.伊格尔-米坦尼(m.egel-mitani)等人,酵母(yeast)6,1990,第127-137页)。
所属领域技术人员已知适当的哺乳动物宿主细胞的实例。所述宿主细胞可以是中国仓鼠卵巢(cho)细胞(例如cho-k1;atccccl-61)、绿猴细胞(greenmonkeycell,cos)(例如cos1(atcccrl-1650)、cos7(atcccrl-1651))、小鼠细胞(例如ns/o)、幼仓鼠肾(babyhamsterkidney,bhk)细胞系(例如atcccrl-1632或atccccl-10)和人细胞(例如hek293(atcccrl-1573)),以及组织培养中的植物细胞。这些细胞系和其它细胞系都可从公共保藏处获得,例如美国典型菌种保藏中心(americantypeculturecollection)(位于马里兰州罗克维尔(rockville,md))。为了改进bst多肽的糖基化,可以对哺乳动物宿主细胞进行修饰,以表达唾液酸转移酶,例如1,6-唾液酸转移酶,例如,如美国专利第5,047,335号(以引用的方式并入本文中)中所述。
用于将外源dna引入哺乳动物宿主细胞中的方法包括(但不限于)磷酸钙介导的转染、电穿孔、deae-葡聚糖介导的转染、脂质体介导的转染、病毒载体,以及生命科技有限公司(lifetechnologiesltd)(位于英国佩斯里(paisley,uk)所述的使用lipofectamin2000进行的转染法和罗氏诊断试剂公司(rochediagnosticscorporation)(位于美国印第安纳波利斯(indianapolis,usa))所述使用fugene6进行的转染法。此项技术中众所周知这些方法,并描述于奥斯贝尔(ausbel)等人(编),1996,现代分子生物学技术(currentprotocolsinmolecularbiology),约翰威立父子出版公司,美国纽约(newyork,usa)中。哺乳动物细胞的培养可以根据已经确立的方法执行,例如(动物细胞生物技术、方法和方案(animalcellbiotechnology),尼戈尔杰金斯(nigeljenkins)编辑,1999,人民出版公司(humanpressinc.),美国新泽西州托托瓦(totowa,n.j.,usa);以及哈里森玛斯(harrisonmass)和雷阿if(raeif),细胞培养的一般技术(generaltechniquesofcellculture),剑桥大学出版社(cambridgeuniversitypress)1997)中所揭示的方法。
i.表达系统、培养和分离
bst多肽可在许多适当的表达系统(包括例如酵母、昆虫细胞、哺乳动物细胞和细菌)中表达。下文将提供示范性表达系统的描述。
酵母本文中使用的术语“酵母”包括能够表达编码bst多肽的基因的各种酵母中的任一种。这些酵母包括(但不限于)产子囊酵母(ascosporogenousyeast)(内孢霉目(endomycetales))、担孢子酵母(basidiosporogenousyeast)和属于半知菌(fungiimperfecti)(芽孢纲(blastomycetes))类的酵母。产子囊酵母分为两个科:蚀精霉科(spermophthoraceae)和酵母菌科(saccharomycetaceae)。酵母菌科是由四个亚科构成,即裂殖酵母亚科(schizosaccharomycoideae)(例如,裂殖酵母属(schizosaccharomyces))、拿逊酵母亚科(nadsonioideae)、油脂酵母亚科(lipomycoideae)和酵母亚科(saccharomycoideae)(例如,毕赤酵母属(pichia)、克鲁维酵母属(kluyveromyces)和酵母属(saccharomyces))。担孢子酵母包括白冬孢酵母属(leucosporidium)、红冬孢酵母属(rhodosporidium)、锁掷酵母属(sporidiobolus)、线黑粉酵母属(filobasidium)和香灰拟锁担菌属(filobasidiella)。属于半知菌(芽孢纲)类的酵母分为两个科:掷孢酵母科(sporobolomycelaceae)(例如,掷孢酵母属(sporoholomyces)和布勒掷孢酵母属(bullera))和隐球酵母科(cryptococcaceae)(例如,假丝酵母属(candida))。
本发明使用的特别值得关注的为毕赤酵母属、克鲁维酵母属、酵母菌属(saccharomyces)、裂殖酵母属(schizosaccharomyces)、汉逊酵母属(hansenula)、球拟酵母属(torulopsis)和假丝酵母属内的种类,包括(但不限于)巴斯德毕赤酵母(p.pastoris)、谷勒氏酵母(p.guillerimondii)、酿酒酵母、卡尔斯伯酵母(s.carlsbergensis)、糖化酵母(s.diastaticus)、道格拉斯酵母(s.douglasii)、克鲁维酵母(s.kluyveri)、诺本酵母(s.norbensis)、卵形酵母(s.oviformis)、乳酸克鲁维酵母(k.lactis)、脆壁克鲁维酵母(k.fragilis)、白色念珠菌(c.albicans)、麦芽糖假丝酵母(c.maltosa)和汉森酵母(h.polymorpha)。
选择适于表达bst多肽的酵母是在所属领域技术人员的技术范围内。在选择用于表达的酵母宿主时,适合的宿主可包括显示具有例如良好分泌能力、低蛋白水解活性、良好可溶性蛋白质产生能力和总体稳固性的酵母宿主。酵母通常可从多种来源获得,包括(但不限于)加利福尼亚大学生物物理和医学物理系酵母基因保藏中心(加利福尼亚州伯克利)(yeastgeneticstockcenter,departmentofbiophysicsandmedicalphysics,universityofcalifornia(berkeley,ca))和美国典型菌种保臧中心(“atcc”)(北弗吉尼亚州马纳萨斯(manassas,va))。
术语“酵母宿主”或“酵母宿主细胞”包括可用作或已用作重组载体或其它转移dna受体的酵母。这一术语包括已接收重组载体或其它转移dna的原始酵母宿主细胞的后代。应了解,由于偶发突变或故意突变,单一母体细胞后代的形态或与原始母体互补的基因组dna或总dna可能未必完全相同。在此定义所指的后代中包括与欲以相关特性(例如存在编码bst多肽的核苷酸序列)表征的母体足够类似的母体细胞后代。
已经开发出包括染色体外复制子或整合载体在内的表达和转化载体用于转化入许多酵母宿主中。举例来说,已经开发出用于以下各生物体的表达载体:酿酒酵母(希寇斯基(sikorski)等人,遗传学(genetics)(1989)122:19;埃托(ito)等人,应用细菌学杂志(j.bacteriol.)(1983)153:163;海因伦(hinnen)等人,美国国家科学院院刊(proc.natl.acad.sci.usa)(1978)75:1929);白色念珠菌(科特兹(kurtz)等人,分子细胞生物学(mol.cell.biol.)(1986)6:142);麦芽糖假丝酵母(库尼则(kunze)等人,基础微生物学杂志(j.basicmicrobiol.)(1985)25:141);汉森酵母(格林森(gleeson)等人,普通与应用微生物学杂志(j.gen.microbiol.)(1986)132:3459;罗格坎普(roggenkamp)等人,分子遗传学和基因组(mol,geneticsandgenomics)(1986)202:302);脆壁克鲁维酵母(达斯(das)等人,应用细菌学杂志(1984)158:1165);乳酸克鲁维酵母(德拉文科特(delouvencourt)等人,应用细菌学杂志(1983)154:737;范登伯格(vandenberg)等人,生物技术(纽约)(biotechnology(ny))(1990)8:135);谷勒氏酵母(库尼则等人,基础微生物学杂志(1985)25:141);巴斯德毕赤酵母(美国专利第5,324,639号;第4,929,555号和第4,837,148号;克雷格(cregg)等人,分子细胞生物学(1985)5:3376);粟酒裂殖酵母(schizosaccharomycespombe)(比奇(beach)等人,自然(nature)(1982)300:706);和解脂耶氏酵母(y.lipolytica);构巢曲霉(a.nidulans)(伯伦斯(ballance)等人,生物化学与生物物理研究通讯(biochem.biophys.res.commun)(1983)112:284-89;迪尔伯恩(tilburn)等人,基因(gene)(1983)26:205-221;和耶尔敦(yelton)等人,美国国家科学院院刊(1984)81:1470-74);黑曲霉(a.niger)(凯里(kelly)和海因斯(hynes),欧洲分子生物学会杂志(emboj.)(1982)4:475-479);里氏木霉(t.reesia)(ep0244234);和丝状真菌,例如脉孢菌(neurospora)、青霉菌(penicillium)、弯颈霉(tolypocladium)(wo91/00357),各文献以引用的方式并入本文中。
所属领域技术人员众所周知用于酵母载体的控制序列,且其包括(但不限于)以下基因的启动子区:例如醇脱氢酶(adh)(ep0284044);烯醇化酶;葡萄糖激酶;葡萄糖-6-磷酸异构酶;甘油醛-3-磷酸-脱氢酶(gap或gapdh);己糖激酶;磷酸果糖激酶;3-磷酸甘油酸变位酶;和丙酮酸激酶(pyk)(ep0329203)。编码酸性磷酸酶的酵母pho5基因也可提供有用的启动子序列(宫之原(miyanohara)等人,美国国家科学院院刊,(1983)80:1)。适用于酵母宿主的其它启动子序列可包括3-磷酸甘油酸激酶(海泽曼(hitzeman)等人,生物化学杂志(j.biol.chem)(1980)255:12073);和其它糖解酶(例如丙酮酸脱羧酶、磷酸丙糖异构酶和磷酸葡萄糖异构酶(霍兰德(holland)等人,生物化学(biochemistry)(1978)17:4900;海斯(hess)等人,酶调控进展杂志(j.adv.enzymereg.)(1969)7:149))的启动子。具有由生长条件控制的转录的其它优点的诱导性酵母启动子可包括醇脱氢酶2、异细胞色素c、酸性磷酸酶、金属硫蛋白、甘油醛-3-磷酸脱氢酶、氮代谢相关性降解酶以及负责麦芽糖和半乳糖利用的酶的启动子区。适用于酵母表达中的载体和启动子进一步描述于ep0073657中。
酵母增强子也可与酵母启动子一起使用。另外,合成启动子也可充当酵母启动子。举例来说,酵母启动子的上游活化序列(upstreamactivatingsequence,uas)可与另一酵母启动子的转录活化区连接,产生合成杂合启动子。所述杂合启动子的实例包括与gap转录活化区连接的adh调控序列。参见美国专利第4,880,734号和第4,876,197号,其以引用的方式并入本文中。杂合启动子的其它实例包括由adh2、gal4、gal10或pho5基因的调控序列与糖解酶基因(例如gap或pyk)的转录活化区组成的启动子。参见ep0164556。此外,酵母启动子可包括能够结合酵母rna聚合酶并起始转录的非酵母来源的天然存在启动子。
可构成酵母表达载体的一部分的其它控制元件包括例如来自gapdh或烯醇酶基因的终止子(霍兰德(holland)等人,生物化学杂志(j.biol.chem.)(1981)256:1385)。另外,来自2μ质粒来源的复制起点也适用于酵母。适用于酵母中的选择基因为酵母质粒中存在的trp1基因。参见迪斯查普(tschumper)等人,基因(gene)(1980)10:157;金戈斯曼(kingsman)等人,基因(1979)7:141。trp1基因可为不能够在色氨酸中生长的酵母突变菌株提供选择标记物。类似地,leu2缺陷型酵母菌株(atcc20,622或38,626)是由带有leu2基因的已知质粒补充。
所属领域技术人员众所周知将外源dna引入酵母宿主中的方法,且其通常包括(但不限于)转化原生质球状体或转化经碱金属阳离子处理的完整酵母宿主细胞。举例来说,可根据萧(hsiao)等人,美国国家科学院院刊,(1979)76:3829和范索里格(vansolingen)等人,应用细菌学杂志(j.bact.)(1977)130:946中所述的方法来转化酵母。然而,也可如萨姆布鲁克(sambrook)等人,分子克隆实验手册(molecularcloning:alab.manual)(2001)中通常所述,使用其它方法将dna引入细胞中,例如核注射法、电穿孔法或原生质体融合法。随后,可使用所属领域技术人员已知的标准技术来培养酵母宿主细胞。
所属领域技术人员已知在酵母宿主细胞中表达异源蛋白质的其它方法。一般参见美国专利公开案第20020055169号;美国专利第6,361,969号、第6,312,923号、第6,183,985号、第6,083,723号、第6,017,731号、第5,674,706号、第5,629,203号、第5,602,034号和第5,089,398号;美国复审专利第re37,343号和第re35,749号;pct公开专利申请案wo99/07862、wo98/37208和wo98/26080;欧洲专利申请案ep0946736、ep0732403、ep0480480、wo90/10277、ep0340986、ep0329203、ep0324274和ep0164556中。还参见格里森(gellissen)等人,列文虎克(antonievanleeuwenhoek)(1992)62(1-2):79-93;罗曼诺斯(romanos)等人,酵母(yeast)(1992)8(6):423-488;格利德尔(goeddel),酶学方法(methodsinenzymology)(1990)185:3-7,各自以引用的方式并入本文中。
在使用所属领域技术人员已知的标准进料分批发酵方法的扩增阶段期间,酵母宿主菌株可在发酵罐中生长。发酵方法可用于解释特定酵母宿主的碳利用路径或表达控制模式的差异。举例来说,酵母菌酵母宿主的发酵可能需要单一葡萄糖进料、复合氮源(例如,酪蛋白水解产物)和多种维生素补充。相比之下,甲基营养型酵母巴斯德毕赤酵母可能需要甘油、甲醇和微量矿物进料,而只需要简单铵(氮)盐以进行最佳生长和表达。例如参见美国专利第5,324,639号;埃利奥特(elliott)等人,蛋白质化学杂志(j.proteinchem.)(1990)9:95;和菲斯查克(fieschko)等人,生物技术与生物工程(biotech.bioeng.)(1987)29:1113,以引用的方式并入本文中。
然而,这些发酵方法可能具有某些与所用酵母宿主菌株无关的常见特征。举例来说,在扩增期间,可将限制生长的养分(通常为碳)添加到发酵罐中以允许最大生长。另外,发酵方法使用的发酵培养基一般设计成含有足量碳、氮、基本盐、磷和其它微量养分(维生素、微量矿物和盐等)。适用于毕赤酵母的发酵培养基的实例描述于美国专利第5,324,639号和第5,231,178号中,其以引用的方式并入本文中。
感染杆状病毒的昆虫细胞术语“昆虫宿主”或“昆虫宿主细胞”是指可用作或已用作重组载体或其它转移dna的受体的昆虫。这一术语包括经过转染的原始昆虫宿主细胞的后代。应了解,由于偶发突变或故意突变,单一母体细胞的后代的形态或与原始母体互补的基因组dna或总dna可能未必完全相同。在此定义所指的后代中包括与欲以相关特性(例如存在编码bst多肽的核苷酸序列)表征的母体足够类似的母体细胞的后代。
所属领域技术人员已知适用于表达bst多肽的昆虫细胞的选择。此项技术中充分描述数种昆虫种类,并且可在市面上购得,包括埃及伊蚊(aedesaegypti)、桑蚕(bombyxmori)、黑腹果蝇(drosophilamelanogaster)、草地贪夜蛾(spodopterafrugiperda)和粉纹夜蛾(trichoplusiani)。在选择用于表达的昆虫宿主时,合适的宿主可包括显示(尤其)具有良好分泌能力、低蛋白水解活性和总体稳固性的宿主。昆虫一般可从多种来源获得,包括(但不限于)加利福尼亚大学生物物理和医学物理系昆虫基因保藏中心(加利福尼亚州伯克利)(insectgeneticstockcenter,departmentofbiophysicsandmedicalphysics,universityofcalifornia(berkeley,ca));和美国典型菌种保藏中心(“atcc”)(北弗吉尼亚州马纳萨斯)。
一般说来,感染杆状病毒的昆虫表达系统的组分包括:转移载体,通常为细菌质粒,其含有杆状病毒基因组的片段和供插入欲表达的异源基因的便利限制性位点;序列与转移载体中的杆状病毒特异性片段同源的野生型杆状病毒(此使得异源基因能同源重组到杆状病毒基因组中);以及适当昆虫宿主细胞,和生长培养基。此项技术中已知用于构建载体、转染细胞、挑选斑块、使细胞在培养物中生长等的材料、方法和技术,并且可以使用描述这些技术的手册。
在将异源基因插入转移载体中后,将载体和野生型病毒基因组转染到昆虫宿主细胞中,在宿主细胞中,载体和病毒基因组重组。表达经包装的重组病毒,并且鉴别和纯化重组体斑块。用于杆状病毒/昆虫细胞表达系统的材料和方法可以试剂盒形式购自例如英杰公司(invitrogencorp.)(加利福尼亚州卡尔斯巴德)。所属领域技术人员一般已知这些技术,且其充分描述于萨姆斯(summers)和史密斯(smith),德克萨斯农业实验站会刊(texasagriculturalexperimentstationbulletin)第1555期(1987)中,此文献以引用的方式并入本文中。还参见理查德森(richardson),39分子生物学方法:杆状病毒表达技术(methodsinmolecularbiology:baculovirusexpressionprotocols)(1995);奥斯贝尔(ausubel)等人,现代分子生物学技术(currentprotocolsinmolecularbiology)16.9-16.11(1994);金(king)和普斯伊(possee),杆状病毒系统实验指南(thebaculovirussystem:alaboratoryguide)(1992);和奥雷利(o'reilly)等人,杆状病毒表达载体实验手册(baculovirusexpressionvectors:alaboratorymanual)(1992)。
事实上,所属领域技术人员已知使用杆状病毒/昆虫细胞表达系统来产生各种异源蛋白质。例如参见,美国专利第6,368,825号;第6,342,216号;第6,338,846号;第6,261,805号;第6,245,528号;第6,225,060号;第6,183,987号;第6,168,932号;第6,126,944号;第6,096,304号;第6,013,433号;第5,965,393号;第5,939,285号;第5,891,676号;第5,871,986号;第5,861,279号;第5,858,368号;第5,843,733号;第5,762,939号;第5,753,220号;第5,605,827号;第5,583,023号;第5,571,709号;第5,516,657号;第5,290,686号;wo02/06305;wo01/90390;wo01/27301;wo01/05956;wo00/55345;wo00/20032;wo99/51721;wo99/45130;wo99/31257;wo99/10515;wo99/09193;wo97/26332;wo96/29400;wo96/25496;wo96/06161;wo95/20672;wo93/03173;wo92/16619;wo92/02628;wo92/01801;wo90/14428;wo90/10078;wo90/02566;wo90/02186;wo90/01556;wo89/01038;wo89/01037;wo88/07082,都以引用的方式并入本文中。
此项技术中已知适用于杆状病毒/昆虫细胞表达系统中的载体,且其包括例如来源于杆状病毒苜蓿银纹夜蛾(autographacalifornica)核型多角体病毒(acnpv)的昆虫表达和转移载体,这是一种不依赖于辅助细胞的病毒表达载体。来源于此系统的病毒表达载体通常使用强病毒多角体蛋白基因启动子来驱动异源基因的表达。一般参见奥雷利(o'reilly)等人,杆状病毒表达载体实验手册(baculovirusexpressionvectors:alaboratorymanual)(1992)。
在将外来基因插入杆状病毒基因组中之前,通常将包含启动子、前导序列(必要时)、相关编码序列和转录终止序列的上述组分组装到中间错位构建体(intermediatetransplacementconstruct)(转移载体)中。中间错位构建体通常保持在能够稳定保持在宿主(例如细菌)中的复制子(例如染色体外元件,例如质粒)中。复制子将具有复制系统,由此使其能保持在适于克隆和扩增的宿主中。更具体来说,质粒可含有多角体蛋白聚腺苷酸化信号(米勒(miller),微生物学年鉴(ann.rev.microbiol.)(1988)42:177)和用于在大肠杆菌中选择和繁殖的原核抗氨比西林(ampicillin)(amp)基因和复制起点。
一种将外来基因引入acnpv中的常用转移载体为pac373。也已经对所属领域技术人员已知的许多其它载体(包括例如pvl985)进行设计,这种设计将多角体蛋白的起始密码子由atg变为att,并将bamhi克隆位点引入att下游32个碱基对处。参见鲁克诺(luckow)和萨姆斯(summers),病毒学(virology)170:31(1989)。其它市售载体包括例如pbluebac4.5/v5-his、pbluebachis2、pmelbac、pbluebac4.5(英杰公司(invitrogencorp.),加利福尼亚州卡尔斯巴德)。
在插入异源基因之后,将转移载体和野生型杆状病毒基因组共转染到昆虫细胞宿主中。此项技术中已知将异源dna引入杆状病毒中的所需位点中的方法。参见萨姆斯(summers)和史密斯(smith),德克萨斯农业实验站会刊(texasagriculturalexperimentstationbulletin)第1555期(1987);史密斯(smith)等人,分子细胞生物学(mol.cell.biol.)(1983)3:2156;鲁克诺(luckow)和萨姆斯(summers),病毒学(virology)(1989)170:31中。举例来说,可通过同源双交叉重组来插入例如多角体蛋白基因等基因中;也可插入所需杆状病毒基因中经工程改造的限制酶位点中。参见米勒(miller)等人,生物学论文集(bioessays)(1989)11(4):91。
可利用电穿孔法来实现转染。参见托特(trotter)和伍德(wood),39分子生物学方法(methodsinmolecularbiology)(1995);曼恩(mann)和金(king),普通病毒学杂志(j.gen.virol.)(1989)70:3501。或者,可以使用脂质体,利用重组表达载体和杆状病毒转染昆虫细胞。例如参见,利伯曼(liebman)等人,生物技术(biotechniques)(1999)26(1):36;格拉芙斯(graves)等人,生物化学(biochemistry)(1998)37:6050;野村(nomura)等人,生物化学杂志(j.biol.chem.)(1998)273(22):13570;斯奇米特(schmidt)等人,蛋白质表达和纯化(proteinexpressionandpurification)(1998)12:323;西弗特(siffert)等人,自然·遗传(naturegenetics)(1998)18:45;迪尔金斯(tilkins)等人,细胞生物学实验手册(cellbiology:alaboratoryhandbook)145-154(1998);蔡(cai)等人,蛋白质表达和纯化(proteinexpressionandpurification)(1997)10:263;多夫林(dolphin)等人,自然·遗传(1997)17:491;科斯特(kost)等人,基因(gene)(1997)190:139;杰克博森(jakobsson)等人,生物化学杂志(1996)271:22203;洛韦思(rowles)等人,生物化学杂志(1996)271(37):22376;雷沃瑞(reverey)等人,生物化学杂志(1996)271(39):23607-10;斯坦利(stanley)等人,生物化学杂志(1995)270:4121;西斯科(sisk)等人,病毒学杂志(j.virol.)(1994)68(2):766;以及彭(peng)等人,生物技术(biotechniques)(1993)14(2):274。市售脂质体包括例如
杆状病毒表达载体通常含有杆状病毒启动子。杆状病毒启动子是能够结合杆状病毒rna聚合酶并起始编码序列(例如结构基因)下游(3′)转录成mrna的任何dna序列。启动子将具有转录起始区,其通常最接近编码序列5′端。此转录起始区通常包括rna聚合酶结合位点和转录起始位点。杆状病毒启动子也可具有称作增强子的第二区域,当存在时,其通常在结构基因的末梢。此外,表达可为受到调控或为组成性的。
在感染周期末期大量转录的结构基因将提供特别有用的启动子序列。实例包括来源于编码病毒多面体蛋白的基因(弗雷森(friesen)等人,杆状病毒基因表达的调控(theregulationofbaculovirusgeneepression),杆状病毒分子生物学(themolecularbiologyofbaculoviruses)(1986);ep0127839和0155476)和编码p10蛋白的基因(沃拉克(vlak)等人,普通病毒学杂志(j.gen.virol.)(1988)69:765)的序列。
新近形成的杆状病毒表达载体被包装到感染性重组杆状病毒中,随后可利用所属领域技术人员已知的技术纯化生长的斑块。参见米勒(miller)等人,生物学论文集(bioessays)(1989)11(4):91;萨姆斯(summers)和史密斯(smith),德克萨斯农业实验站会刊(texasagriculturalexperimentstationbulletin)第1555期(1987)。
已经开发出了用于感染到数种昆虫细胞中的重组杆状病毒表达载体。举例来说,已开发(尤其)用于埃及伊蚊(atcc第ccl-125号)、桑蚕(atcc第crl-8910号)、黑腹果蝇(atcc第1963号)、草地贪夜蛾和粉纹夜蛾的重组杆状病毒。参见怀特(wright),自然(nature)(1986)321:718;卡博耐尔(carbonell)等人,病毒学杂志(j.virol.)(1985)56:153;史密斯(smith)等人,分子细胞生物学(mol.cell.biol.)(1983)3:2156。一般参见弗雷瑟(fraser)等人,体外细胞和发育生物学(invitrocell.dev.biol.)(1989)25:225。更具体来说,用于杆状病毒表达载体系统的细胞系通常包括(但不限于)sf9(草地贪夜蛾)(atcc第crl-1711号)、sf21(草地贪夜蛾)(英杰公司(invitrogencorp.),目录号11497-013(加利福尼亚州卡尔斯巴德(carlsbad,ca)))、tri-368(粉纹夜蛾)和high-fivetmbti-tn-5b1-4(粉纹夜蛾)。
可在市面上购得用于在杆状病毒/细胞表达系统中直接和融合表达异源多肽的细胞和培养基,且所属领域技术人员一般已知细胞培养技术。
大肠杆菌、假单胞菌和其它原核生物所属领域技术人员已知细菌表达技术。多种载体可用于细菌宿主中。这些载体可以是单拷贝或者低或高多拷贝载体。载体可用于克隆和/或表达。鉴于存在丰富的有关载体的文献、许多载体在市面上有售以及甚至存在描述载体和其限制性图谱及特征的手册,故无需在本文中深入讨论。众所周知,载体通常涉及允许选择的标记物,这些标记物可提供细胞毒性剂抗性、原营养或免疫性。常常会存在多个标记物,其将提供不同特征。
细菌启动子为能够结合细菌rna聚合酶并起始编码序列(例如结构基因)下游(3')转录成mrna的任何dna序列。启动子将具有转录起始区,其通常最接近编码序列的5'端。此转录起始区通常包括rna聚合酶结合位点和转录起始位点。细菌启动子也可具有称作操纵子的第二区域,其可与开始rna合成的相邻rna聚合酶结合位点重叠。操纵子允许负调控(诱导性)转录,这是因为基因抑制蛋白可结合操纵子,并由此抑制特定基因的转录。在不存在负调控元件(例如操纵子)的情况下,可发生组成性表达。另外,利用基因活化蛋白结合序列可实现正调控,当存在这一序列时,其通常最接近rna聚合酶结合序列的(5')。基因活化蛋白的实例为代谢物活化蛋白(cataboliteactivatorprotein,cap),其有助于起始大肠杆菌中lac操纵子的转录[雷波德(raibaud)等人,遗传学年鉴(annu.rev.genet.)(1984)18:173]。因此,表达的调控可为正调控或负调控,由此增强或减弱转录。
编码代谢路径酶的序列将提供特别有用的启动子序列。实例包括来源于糖代谢酶(例如半乳糖、乳糖(lac)[常(chang)等人,自然(nature)(1977)198:1056]和麦芽糖)的启动子序列。其它实例包括来源于生物合成酶(例如色氨酸(trp))的启动子序列[古德尔(goeddel)等人,核酸研究,(1980)8:4057;耶尔弗顿(yelverton)等人,核酸研究,(1981)9:731;美国专利第4,738,921号;欧洲公开案第036776号和第121775号,其以引用的方式并入本文中]。β-半乳糖苷酶(bla)启动子系统[威瑟曼(weissmann)(1981)“干扰素和其它错误的克隆(thecloningofinterferonandothermistakes.)”,干扰素(interferon)3(i.格雷瑟(i.gresser)编)]、噬菌体λpl[施马塔克(shimatake)等人,自然(nature)(1981)292:128]和t5[美国专利第4,689,406号,以引用的方式并入本文中]启动子系统也可提供有用的启动子序列。本发明的优选方法利用强启动子(例如t7启动子)来诱导高水平的bst多肽。所属领域技术人员已知这些载体的实例,且其包括来自诺杰公司(novagen)的pet29系列,以及wo99/05297(以引用的方式并入本文中)中所述的ppop载体。这些表达系统可在宿主中产生高水平bst多肽,同时不会损害宿主细胞生存力或生长参数。pet19(诺杰公司(novagen))为此项技术中已知的另一载体。
另外,自然界中不存在的合成启动子也可充当细菌启动子。举例来说,可将一个细菌或噬菌体启动子的转录活化序列与另一细菌或噬菌体启动子的操纵子序列连接,从而产生合成的杂合启动子[美国专利第4,551,433号,以引用的方式并入本文中]。举例来说,tac启动子是由trp启动子与lac操纵子序列构成的杂合trp-lac启动子,其受lac阻遏物调控[阿曼恩(amann)等人,基因(gene)(1983)25:167;德波尔(deboer)等人,美国国家科学院院刊,(1983)80:21]。此外,细菌启动子可包括非细菌来源的天然存在的启动子,其能够与细菌rna聚合酶结合并起始转录。也可将非细菌来源的天然存在的启动子与相容性rna聚合酶偶合以便在原核生物中高水平表达一些基因。噬菌体t7rna聚合酶/启动子系统为偶合启动子系统的一个实例[斯蒂尔(studier)等人,分子生物学杂志(j.mol.biol.)(1986)189:113;塔波(tabor)等人,美国国家科学院院刊,(1985)82:1074]。另外,杂合启动子也可由噬菌体启动子和大肠杆菌操纵子区组成(欧洲公开案第267851号)。
除功能性启动子序列之外,有效核糖体结合位点也可用于在原核生物中表达外来基因。在大肠杆菌中,核糖体结合位点称作沙恩-达尔加诺(shine-dalgarno,sd)序列,且包括起始密码子(atg)和位于起始密码子上游3-11个核苷酸处的3-9个核苷酸长的序列[沙恩(shine)等人,自然(nature)(1975)254:34]。据悉,sd序列通过sd序列与大肠杆菌16srrna的3'端之间的碱基配对来促进mrna与核糖体的结合[斯特兹(steitz)等人,“信使rna中的基因信号和核苷酸序列(geneticsignalsandnucleotidesequencesinmessengerrna)”,生物调控和发育:基因表达(biologicalregulationanddevelopment:geneexpression)(r.f.古德伯格(r.f.goldberger)编,1979)]。表达具有弱核糖体结合位点的真核基因和原核基因[萨姆布鲁克(sambrook)等人,“在大肠杆菌中表达克隆的基因(expressionofclonedgenesinescherichiacoli)”,分子克隆实验手册(molecularcloning:alaboratorymanual),1989]。
术语“细菌宿主”或“细菌宿主细胞”是指可用作或已用作重组载体或其它转移dna的受体的细菌。这一术语包括经过转染的原始细菌宿主细胞的后代。应了解,由于偶发突变或故意突变,单一母体细胞的后代的形态或与原始母体互补的基因组dna或总dna可能未必完全相同。在此定义所指的后代中包括与欲以相关特性(例如存在编码bst多肽的核苷酸序列)表征的母体足够类似的母体细胞的后代。
所属领域技术人员已知适于表达bst多肽的宿主细菌的选择。在选择供表达的细菌宿主时,合适宿主可包括显示(尤其)具有良好包涵体形成能力、低蛋白水解活性和总体稳固性的宿主。细菌宿主通常可从多种来源获得,包括(但不限于)加利福尼亚大学生物物理和医学物理系细菌基因保藏中心(加利福尼亚州伯克利)(bacterialgeneticstockcenter,departmentofbiophysicsandmedicalphysics,universityofcalifornia(berkeley,ca))和美国典型菌种保藏中心(“atcc”)(北弗吉尼亚州马纳萨斯)。工业/医药发酵一般使用来源于k菌株(例如w3110)的细菌或来源于b菌株(例如bl21)的细菌。这些菌株因其生长参数众所周知且稳定而特别有用。另外,这些菌株为非病原性的,而出于安全性和环境原因,其在商业上相当重要。适当大肠杆菌宿主的其它实例包括(但不限于)菌株bl21、dh10b或其衍生物。在本发明方法的另一个实施例中,大肠杆菌宿主为缺乏蛋白酶的菌株,包括(但不限于)omp-和lon-。宿主细胞株可为假单胞菌属,包括(但不限于)荧光假单胞菌、铜绿假单胞菌和恶臭假单胞菌。已知荧光假单胞菌生物型1(命名为菌株mb101)适用于重组制备,并且可用于治疗性蛋白质的制备工艺中。假单胞菌表达系统的实例包括从陶氏化学公司(dowchemicalcompany)得到的呈宿主菌株的系统(密歇根州米德兰(midland,mi),可见于万维网dow.com)。
一旦形成了重组宿主细胞株(即,已将表达构建体引入宿主细胞中,并分离出具有合适表达构建体的宿主细胞),就在适于产生bst多肽的条件下培养这一重组宿主细胞株。所属领域技术人员将了解,培养重组宿主细胞株的方法将取决于所用表达构建体的性质和宿主细胞的身份。通常使用所属领域技术人员已知的方法来培养重组宿主菌株。重组宿主细胞通常是在含有可吸收的碳源、氮源和无机盐源且任选含有维生素、氨基酸、生长因子和所属领域技术人员已知的其它蛋白质培养补充物的液体培养基中培养。用于培养宿主细胞的液体培养基可任选含有抗生素或抗真菌剂以防止不需要的微生物生长和/或化合物(包括(但不限于)用于选择含有表达载体的宿主细胞的抗生素)。
重组宿主细胞可以分批或连续模式培养,其中细胞采集(在bst多肽积聚于胞内的情况下)或培养物上清液的采集为分批或连续模式。对于在原核宿主细胞中制备,优选分批培养和细胞采集。
本发明的bst多肽通常是在重组系统中表达之后纯化。可以利用此项技术中已知的多种方法,从宿主细胞或培养基中纯化出bst多肽。美国专利第5,849,883号和wo89/10932(以全文引用的方式并入本文中)描述宿主细胞中b-gcsf和其类似物的克隆以及分离和纯化方法。在细菌宿主细胞中产生的bst多肽可具有弱溶解性或不可溶(呈包涵体形式)。在本发明一个实施例中,利用本文中揭示的方法以及此项技术中已知的方法,可容易地在bst多肽中进行氨基酸取代,选择的这些取代应能增加重组产生的蛋白质的溶解性。在不溶性蛋白质的情况下,可通过离心收集宿主细胞溶解产物中的蛋白质,随后可进一步进行细胞的均质化。在具有弱溶解性蛋白质的情况下,可添加包括(但不限于)聚乙烯亚胺(polyethyleneimine,pei)的化合物以诱导部分溶解的蛋白质沉淀。随后宜通过离心作用收集沉淀蛋白质。可使用所属领域技术人员已知的多种方法,使重组宿主细胞破裂或均质化以自细胞内部释放包涵体。可使用众所周知的技术来使宿主细胞破裂或均质化,这些技术包括(但不限于)酶促细胞破裂、超声波降解法、杜恩斯均质化(douncehomogenization)或高压释放破裂。在本发明方法的一个实施例中,使用高压释放技术来使大肠杆菌宿主细胞破裂,从而释放出bst多肽的包涵体。在处理bst多肽包涵体时,宜使均质化时间的重复最少,以便使包涵体产率达到最大,同时不会因例如溶解、机械剪切或蛋白质水解等因素而造成损失。
随后,可使用此项技术中已知的多种适合的增溶剂中的任一种来溶解不溶性或沉淀的bst多肽。可利用尿素或盐酸胍来溶解bst多肽。应使溶解bst多肽的体积最小,由此可使用易于操作的批量来进行大批量制备。在重组宿主可以生长达到数千升体积批量的大规模商业装置中,这个因素相当重要。另外,当在大规模商业装置中制造bst多肽时,特别是对于人类医药用途来说,如果可能的话,那么应避免可损害机器和容器的苛性化学品(harshchemicals)或蛋白质产物本身。本发明方法中已经显示,较温和的变性剂尿素可代替较苛刻的变性剂盐酸胍用于溶解bst多肽包涵体。尿素的使用在有效溶解bst多肽包涵体的同时,显著降低了对在bst多肽的制造和纯化过程中所利用的不锈钢设备造成损害的风险。
在可溶性bst蛋白质的情况下,bst可被分泌到周质空间或培养基中。另外,可溶性bst可存在于宿主细胞的细胞质中。可能需要在执行纯化步骤之前浓缩可溶性bst。可使用所属领域技术人员已知的标准技术浓缩来自例如细胞溶解产物或培养基等的可溶性bst。另外,可使用所属领域技术人员已知的标准技术使宿主细胞破裂,并从宿主细胞的细胞质或周质空间释放出可溶性bst。
当制备呈融合蛋白形式的bst多肽时,可去除融合序列。可通过酶促裂解或化学裂解来去除融合序列。可使用所属领域技术人员已知的方法来酶促去除融合序列。如所属领域技术人员所了解的,对用于去除融合序列的酶的选择将由融合体的身份决定,而反应条件将由酶的选择指定。可使用所属领域技术人员已知的试剂,包括(但不限于)溴化氰、tev蛋白酶和其它试剂,来实现化学裂解。可利用所属领域技术人员已知的方法从裂解的融合序列中纯化出裂解的bst多肽。如所属领域技术人员所了解的,这些方法将由融合序列和bst多肽的身份和特性决定。纯化方法可包括(但不限于)尺寸排阻色谱法、疏水相互作用色谱法、离子交换色谱法或透析法,或者其任何组合。
也可纯化bst多肽以从蛋白质溶液中去除dna。可利用例如沉淀或离子交换色谱法等此项技术中已知的任何适当方法来去除dna,但也可通过用核酸沉淀剂(例如(但不限于)硫酸鱼精蛋白)沉淀来去除dna。可使用包括(但不限于)离心或过滤的众所周知的标准方法使bst多肽与沉淀的dna分离。在欲使用bst多肽治疗动物或人类的情况中,宿主核酸分子的去除是一个重要因素,且本发明的方法将宿主细胞dna减少到医药学上可接受的水平。
小规模或大规模发酵方法也可用于蛋白质表达中,这些方法包括(但不限于)发酵罐、振荡烧瓶、流化床生物反应器、中空纤维生物反应器、滚瓶培养系统和搅拌槽生物反应器系统。这些方法中的每一种都可通过分批、馈料分批或连续模式过程执行。
一般使用此项技术中的标准方法来回收本发明bst多肽。举例来说,可对培养基或细胞溶解产物进行离心或过滤,以去除细胞碎片。可将上清液浓缩或稀释到所需体积或者透滤到适当缓冲液中以调节制剂以供进一步纯化。本发明bst多肽的进一步纯化包括从完整形式中分离出脱酰胺化和剪短形式的bst多肽变异体。
以下任一种示范性程序可用于纯化本发明的bst多肽:亲和色谱;阴离子或阳离子交换色谱(使用(包括(但不限于)deaesepharose);二氧化硅色谱;高效液相色谱(highperformanceliquidchromatography,hplc);反相hplc;凝胶过滤(使用(包括(但不限于))sephadexg-75);疏水相互作用色谱;尺寸排阻色谱;金属螯合物色谱;超滤/透滤;乙醇沉淀;硫酸铵沉淀;色谱聚焦;置换色谱;电泳程序(包括(但不限于)制备型等电聚焦);差示溶解度(包括(但不限于)硫酸铵沉淀);sds-page;或萃取。
根据所属领域技术人员已知且使用的标准程序,可将本发明蛋白质(包括(但不限于),包含非天然氨基酸的蛋白质、包含非天然氨基酸的肽、针对包含非天然氨基酸的蛋白质的抗体、包含非天然氨基酸的蛋白质的结合搭配物等)部分或实质上纯化到均质。因此,可利用所属领域技术人员已知的多种方法中的任一种来回收和纯化本发明的多肽,这些方法包括(但不限于)硫酸铵或乙醇沉淀、酸或碱萃取、柱色谱、亲和柱色谱、阴离子或阳离子交换色谱、磷酸纤维素色谱、疏水相互作用色谱、羟基磷灰石色谱、凝集素色谱、凝胶电泳等。必要时,在制备正确折叠的成熟蛋白质时,可使用蛋白质再折叠步骤。可将高效液相色谱(hplc)、亲和色谱或其它适合的方法用于需要高纯度的最终纯化步骤中。在一个实施例中,将制备的针对非天然氨基酸(或包含非天然氨基酸的蛋白质或肽)的抗体用作纯化试剂(包括(但不限于))以基于亲和力来纯化包含一个或一个以上非天然氨基酸的蛋白质或肽。必要时,在部分纯化或达到均质后,任选将多肽用于多种应用,包括(但不限于)作为分析组分、治疗剂、预防剂、诊断剂、研究试剂和/或作为产生抗体的免疫原。可使用常规方案,通过对动物,优选非人类动物投予多肽或带有表位的片段,来获得针对本发明多肽产生的抗体。所属领域技术人员可使用多种已知技术产生抗体。另外,也可使用转基因小鼠或其它生物体(包括其它哺乳动物)来表达人类化抗体。上述抗体可用来分离或鉴别表达多肽的克隆,或者用于纯化多肽。针对本发明多肽的抗体也可用于治疗疾病。
本发明的多肽和聚核苷酸也可用作疫苗。因此,另一方面,本发明涉及一种诱导哺乳动物的免疫反应的方法,其包含用适于产生抗体和/或t细胞(包括例如产生细胞因子的t细胞或细胞毒性t细胞)免疫反应的本发明多肽接种所述哺乳动物,以保护所述动物免受疾病影响,不管这种疾病是否已经在个体体内产生。诱导哺乳动物免疫反应的方法也可包含经由载体递送本发明多肽,引导在体内表达聚核苷酸和编码多肽,以便诱导所述免疫反应,从而产生抗体,保护所述动物免受本发明疾病的影响。一种投予载体的方式是将载体以粒子上的涂层或其它形式投予以加快使其进入所需细胞中。所述核酸载体可包含dna、rna、经修饰的核酸或dna/rna杂合体。对于用作疫苗,多肽或核酸载体通常是以疫苗调配物(组合物)的形式提供。所述调配物可进一步包含适合的载剂。由于多肽可能会在胃中降解,使得可以肠胃外(例如皮下、肌肉内、静脉内或皮内注射)投予所述多肽。适于肠胃外投予的调配物包括水性和非水性无菌注射液,这些注射液可含有抗氧化剂、缓冲剂、抑菌剂,和使调配物与接受者血液等渗的溶质;以及水性和非水性无菌悬浮液,其可包括悬浮剂或增稠剂。疫苗调配物还可包括所属领域技术人员已知的佐剂系统,用于增强调配物的免疫原性。剂量将取决于疫苗的比活性,并且易于借助常规实验确定。
除了本文中所述的其它参考文献之外,所属领域技术人员已知多种纯化/蛋白质折叠方法,包括(但不限于)以下文献中描述的方法:r.斯科普斯(r.scopes),蛋白质纯化(proteinpurification),施普林格-维拉格出版公司(springer-verlag),纽约(n.y.)(1982);道彻尔(deutscher),酶学方法(methodsinenzymology)第182卷:蛋白质纯化指南(guidetoproteinpurification.)学术出版公司(academicpress,inc.)纽约(1990);沙德拉(sandana),(1997)蛋白质的生物分离(bioseparationofproteins).学术出版公司;博拉格(bollag)等人(1996)蛋白质方法(proteinmethods),第2版威立-利斯出版公司(wiley-liss),纽约;沃克尔(walker),(1996)蛋白质技术手册(theproteinprotocolshandbook)哈曼纳出版社(humanapress),新泽西(nj);哈瑞丝(harris)和安吉拉(angal),(1990)蛋白质纯化应用:实用方法(proteinpurificationapplications:apracticalapproach)牛津irl出版社(irlpressatoxford),英格兰牛津(oxford,england);哈瑞丝和安吉拉,蛋白质纯化应用:实用方法牛津irl出版社,英格兰牛津;斯科普(scopes),(1993)蛋白质纯化:原理和实践(proteinpurification:principlesandpractice)第3版施普林格-维拉格出版公司,纽约;杰森(janson)和雷顿(ryden),(1998)蛋白质纯化:原理、高分辨率方法和应用(proteinpurification:principles.highresolutionmethodsandapplications),第2版威立vch出版公司(wiley-vch),纽约;和沃克尔(1998),有关cd-rom的蛋白质技术(proteinprotocolsoncd-rom)哈曼纳出版社,新泽西;以及其中所引用的参考文献。
在真核宿主细胞或非真核宿主细胞中产生具有非天然氨基酸的相关蛋白质或多肽的一个优点在于,蛋白质或多肽通常折叠成其天然构象。然而,在本发明的某些实施例中,所属领域技术人员将认识到,在合成、表达和/或纯化之后,蛋白质或肽可具有与相关多肽的所需构象不同的构象。在本发明一个方面中,表达的蛋白质或多肽任选变性,随后复性。此举可利用此项技术中已知的方法来实现,所述方法包括(但不限于)将伴侣蛋白(chaperonin)添加到相关蛋白质或多肽中;将蛋白质溶解于例如盐酸胍等离液剂中;利用蛋白质二硫化物异构酶等。
一般来说,有时候需要使表达的多肽变性和还原,随后使所述多肽再折叠成优选的构象。举例来说,可将胍、尿素、dtt、dte和/或伴侣蛋白添加到相关翻译产物中。所属领域技术人员已知还原、变性和复性蛋白质的方法(参见上述参考文献,以及德宾斯基(debinski)等人,(1993)生物化学杂志(j.biol.chem.),268:14065-14070;克雷特曼(kreitman)和帕斯坦(pastan)(1993)生物结合物化学,4:581-585;和布奇纳(buchner)等人,(1992)分析生物化学,205:263-270)。举例来说,德宾斯基(debinski)描述在胍-dte中使包涵体蛋白质变性和还原。这些蛋白质可在含有(包括(但不限于))氧化谷胱甘肽和l-精氨酸的氧化还原缓冲液中再折叠。再折叠试剂可流动或以其它方式移动,以便与一种或一种以上多肽或其它表达产物接触,或者反之亦然。
在原核产生bst多肽的情况下,由此产生的bst多肽可能错误折叠,并因此缺乏生物活性或具有较低生物活性。可通过“再折叠”来恢复蛋白质的生物活性。一般使用例如一种或一种以上离液剂(例如尿素和/或胍)和能够还原二硫键的还原剂(例如硫苏糖醇(dtt)或2-巯基乙醇(2-me))来溶解(其中bst多肽也不可溶)、展开和还原bst多肽链,以使错误折叠的bst多肽再折叠。随后在中等浓度的离液剂下,添加氧化剂(例如氧、胱氨酸或胱胺),以便再形成二硫键。可使用此项技术中已知的标准方法再折叠bst多肽,例如美国专利第4,511,502号、第4,511,503号和第4,512,922号中所述的方法,这些专利都以引用的方式并入本文中。bst多肽也可与其它蛋白质共折叠,以形成异二聚体或异多聚体。
再折叠之后,可进一步纯化bst。可使用所属领域技术人员已知的多种技术来纯化bst,包括(但不限于)疏水相互作用色谱、尺寸排阻色谱、离子交换色谱、反相高效液相色谱、亲和色谱等,或其任何组合。其它纯化还可包括干燥或沉淀纯化的蛋白质的步骤。
纯化后,可将bst交换到不同缓冲液中,和/或利用此项技术中已知的多种方法中的任一种进行浓缩,所述方法包括(但不限于)透滤和透析。提供的呈单一纯化蛋白质形式的bst可经历聚集和沉淀。
纯化的bst可为至少90%纯(如通过反相高效液相色谱rp-hplc或十二烷基硫酸钠-聚丙烯酰胺凝胶电泳sds-page所测量)或至少95%纯,或至少98%纯,或至少99%或更高纯度。不管bst纯度的确切数值如何,bst的纯度应足够高以用作医药产品或用于进一步处理,例如,与例如peg等水溶性聚合物结合。
某些bst分子可在不存在其它活性成分或蛋白质(除赋形剂、载剂和稳定剂、血清白蛋白等外)的情况下用作治疗剂,或其可与另一蛋白质或聚合物复合。
根据本发明,动物可为肉可供消费的任何经济动物,且优选可为奶牛。动物优选为乳牛或肉用奶牛。在一个实施例中,奶牛是小母牛或种牛。在替代性实施例中,奶牛选自黑白花牛、杂色牛、昂古斯牛(angus)、海福特牛(hereford)、夏洛来牛(charlois)等品种。
可使用本发明的bst类似物,通过在断奶时到屠宰前之间任何时间投予来增加肉用牛的生长速率。在本发明一个实施例中,对肉用牛投予bst最少30天。在本发明另一实施例中,根据屠宰的所需时间,投予bst最多450天或更短时间。在另一实施例中,对用于提供小牛肉的动物投予bst类似物直到屠宰年龄,以实现所需生长速率增加。在另一实施例中,本发明的bst多肽用于增加牛、尤其乳牛的泌乳;产后投予bst类似物介于30天与90天之间的时间,并持续最多300天。在另一实施例中,投予本发明bst类似物以增加例如山羊和绵羊等其它经济产奶动物的泌乳。
一般纯化方法:可以任何适当次序对包含bst多肽的细胞溶解产物、提取物、培养基、包涵体、宿主细胞的周质空间、宿主细胞的细胞质或其它材料或者由任何分离步骤产生的任何bst多肽混合物执行多种分离步骤中的任一种,所述分离步骤包括(但不限于)亲和色谱、离子交换色谱、疏水相互作用色谱、凝胶过滤色谱、高效液相色谱(“hplc”)、反相hplc(“rp-hplc”)、膨胀床吸附,或者其任何组合和/或重复。
用于执行本文中所述的技术的设备和其它必要材料在市面上有售。泵、馏分收集器、监测器、记录器和整个系统都可从例如应用生物系统公司(appliedbiosystems)(加利福尼亚州福斯特城(fostercity,ca))、拜耳雷德实验公司(bio-radlaboratories,inc.)(加利福尼亚州埃库莱斯(hercules,ca))和安玛西亚生物科技公司(amershambiosciences,inc.)(新泽西州皮斯卡塔韦(piscataway,nj))获得。包括(但不限于)交换基质材料、培养基和缓冲液等色谱材料也可从这些公司获得。
可使用专用设备(例如泵)更快速地实现平衡,以及本文中所述的柱色谱过程中的其它步骤,例如洗涤和洗脱。市售的泵包括(但不限于)
馏分收集器的实例包括redifrac馏分收集器、frac-100和frac-200馏分收集器以及
可使用任何市售监测器来监测色谱过程。这些监测器可用于收集如uv、ph值和传导率等信息。检测器的实例包括监测器uv-1、
在本发明一个实施例中,例如,可通过首先在尿素中使所得的纯化bst多肽变性,接着在合适ph值下于含有还原剂(例如dtt)的tris缓冲液中稀释来还原bst多肽,并使其变性。在另一个实施例中,在尿素中使浓度介于约2m到约9m之间的范围内的bst多肽变性,接着在ph值范围为约5.0到约8.0的tris缓冲液中稀释。随后可培育此实施例的再折叠混合物。在一个实施例中,在室温下,将再折叠混合物培育4小时到24小时。随后可进一步分离或纯化经还原且变性的bst多肽混合物。
如本文中所述,在执行任何后续分离步骤之前,可调整第一bst多肽混合物的ph值。另外,可使用此项技术中已知的技术来浓缩第一bst多肽混合物或其任何后续混合物。此外,可使用所属领域技术人员已知的技术将包含第一bst多肽混合物或其任何后续混合物的洗脱缓冲液换成适于下一分离步骤的缓冲液。
离子交换色谱在一个实施例中且作为任选的额外步骤,可对第一bst多肽混合物执行离子交换色谱。一般参见离子交换色谱法的原理和方法(ionexchangechromatography:principlesandmethods)(目录号18-1114-21,安玛西亚生物科技公司(amershambiosciences,inc.)(新泽西州皮斯卡塔韦(piscataway,nj))。市售离子交换柱包括
阳离子交换基质可为任何适当的阳离子交换剂,包括强和弱阳离子交换剂。强阳离子交换剂可在较宽的ph值范围内保持离子化,并因此能够在较宽的ph值范围内结合bst。然而,弱阳离子交换剂可随ph值变化而失去离子化。举例来说,当ph值降到约ph4或ph5以下时,弱阳离子交换剂可失去电荷。适当的强阳离子交换剂包括(但不限于)带电荷的官能团,例如磺丙基(sulfopropyl,sp)、甲磺酸根(methylsulfonate,s)或磺乙基(sulfoethyl,se)。阳离子交换基质可为优选在约ph2.5到约ph6.0的ph值范围内结合bst的强阳离子交换剂。或者,强阳离子交换剂可在约ph2.5到约ph5.5的ph值范围内结合bst。阳离子交换基质可为在约ph3.0下结合bst的强阳离子交换剂。或者,阳离子交换基质可为优选在约ph6.0到约ph8.0的ph值范围内结合bst的强阳离子交换剂。阳离子交换基质可为优选在约ph8.0到约ph12.5的ph值范围内结合bst的强阳离子交换剂。或者,强阳离子交换剂可在约ph8.0到约ph12.0的ph值范围内结合bst。
在装载bst之前,可例如使用数个柱体积的稀弱酸(例如4个柱体积的20mm乙酸,ph3)平衡阳离子交换基质。平衡后,可添加bst,并且在洗脱实质上纯化的bst前,也使用例如弱乙酸或磷酸溶液等弱酸溶液洗涤柱1次到数次。举例来说,可使用约2-4个柱体积的20mm乙酸(ph3)洗涤柱。也可使用其它洗涤液,例如使用2-4个柱体积的0.05m乙酸钠(ph5.5),或者0.05m乙酸钠与0.1m氯化钠的混合物(ph5.5)。或者,使用此项技术中已知的方法,使用数个柱体积的稀弱碱来平衡阳离子交换基质。
另外,可通过使阳离子交换基质与ph值或离子强度足够低的缓冲液接触,从所述基质中置换出bst,从而洗脱实质上纯化的bst。洗脱缓冲液的ph值可在约ph2.5到约ph6.0的范围内。更具体来说,洗脱缓冲液的ph值可在约ph2.5到约ph5.5、约ph2.5到约ph5.0的范围内。洗脱缓冲液的ph值可为约3.0。此外,洗脱缓冲液的量可变化极大,且一般在约2到约10个柱体积的范围内。
在将bst多肽吸附到阳离子交换基质上后,可通过使基质与ph值或离子强度足够高的缓冲液接触,从所述基质中置换出bst多肽,从而洗脱实质上纯化的bst多肽。适用于高ph值洗脱实质上纯化的bst多肽的缓冲液包括(但不限于)浓度在至少约5mm到至少约100mm范围内的柠檬酸盐、磷酸盐、甲酸盐、乙酸盐、hepes和mes缓冲液。
反相色谱可根据所属领域技术人员已知的适合方案执行rp-hplc以纯化蛋白质。例如参见皮尔森(pearson)等人,分析生物化学(analbiochem.)(1982)124:217-230(1982);瑞文斯(rivier)等人,色谱学杂志(j.chrom.)(1983)268:112-119;国谷(kunitani)等人,色谱学杂志(1986)359:391-402。可对bst多肽执行rp-hplc以分离出实质上纯化的bst多肽。在这点上,可使用具有多种长度(包括(但不限于)至少约c3到至少约c30、至少约c3到至少约c20或至少约c3到至少约c18)的烷基官能团的二氧化硅衍生化树脂。或者,可使用聚合性树脂。举例来说,可使用tosohaasamberchromecg1000sd树脂,其为苯乙烯聚合物树脂。也可使用具有多种烷基链长度的氰基或聚合性树脂。此外,可用例如乙醇的溶剂洗涤rp-hplc柱。sourcerp柱为rp-hplc柱的另一实例。
含有离子配对剂和有机改性剂(例如甲醇、异丙醇、四氢呋喃、乙腈或乙醇)的合适洗脱缓冲液都可用于从rp-hplc柱洗脱bst多肽。最常用的离子配对剂包括(但不限于)乙酸、甲酸、高氯酸、磷酸、三氟乙酸、七氟丁酸、三乙胺、四甲基铵、四丁基铵和三乙基乙酸铵。可使用一种或一种以上梯度或等度条件执行洗脱,其中优选减少分离时间且降低峰宽度的梯度条件。另一种方法涉及使用具有不同溶剂浓度范围的两种梯度。适用于本文中的洗脱缓冲液的实例可包括(但不限于)乙酸铵和乙腈溶液。
疏水相互作用色谱纯化技术可对bst多肽执行疏水相互作用色谱(hic)。一般参见疏水相互作用色谱手册:原理和方法(hydrophobicinteractionchromatographyhandbook:principlesandmethods)(目录号18-1020-90,安玛西亚生物科技公司(amershambiosciences,inc.)(新泽西州皮斯卡塔韦(piscataway,nj)),以引用的方式并入本文中。适合的hic基质可包括(但不限于)经烷基或芳基取代的基质,例如经丁基、己基、辛基或苯基取代的基质,所述基质包括琼脂糖、交联琼脂糖、琼脂糖凝胶、纤维素、二氧化硅、葡聚糖、聚苯乙烯、聚(甲基丙烯酸酯)基质,以及混合模式树脂,包括(但不限于)聚乙二胺树脂或者经丁基或苯基取代的聚(甲基丙烯酸酯)基质。疏水相互作用柱色谱的市售来源包括(但不限于)
简单地说,在装载之前,可使用所属领域技术人员已知的标准缓冲液(例如乙酸/氯化钠溶液,或含有硫酸铵的hepes)来平衡hic柱。可将硫酸铵用作缓冲液来装载hic柱。在装载bst多肽后,随后可使用标准缓冲液和条件来洗涤柱以去除不想要的材料,但将bst多肽留在hic柱上。可用约3到约10个柱体积的标准缓冲液(例如含有edta和浓度低于平衡缓冲液的硫酸铵的hepes缓冲液,或乙酸/氯化钠缓冲液)来洗脱bst多肽。使用例如磷酸钾梯度的逐渐降低的线性盐梯度也可用来洗脱bst分子。随后可例如通过过滤(例如透滤或超滤)来浓缩洗出液。可利用透滤来去除用于洗脱bst多肽的盐。
其它纯化技术可对第一bst多肽混合物或其任何后续混合物执行使用例如凝胶过滤(凝胶过滤的原理和方法(gelfiltration:principlesandmethods)(目录号18-1022-18,安玛西亚生物科技公司,新泽西州皮斯卡塔韦),以引用的方式并入本文中)、羟基磷灰石色谱(适合基质包括(但不限于)ha-ultrogel、highresolution(卡尔生物化学公司(calbiochem))、cht陶瓷羟基磷灰石(拜耳雷德公司(biorad))、bio-gelhtp羟基磷灰石(拜耳雷德公司))、hplc、膨胀床吸附、超滤、透滤、冻干等的另一分离步骤,以去除任何过量盐,并用适合的缓冲液代替所述缓冲液以用于下个分离步骤或甚至最终药物产品的调配。
在本文中所述的各个步骤中,可使用所属领域技术人员已知的技术监测bst多肽(包括实质上纯化的bst多肽)的产率。这些技术也可用于在最后的分离步骤后评估实质上纯化的bst多肽的产率。举例来说,可使用具有多种烷基链长度的数个反相高压液相色谱柱(例如氰基rp-hplc、c18rp-hplc,以及阳离子交换hplc和凝胶过滤hplc)中的任一种来监测bst多肽的产率。
在本发明的特定实施例中,在各纯化步骤后,bst的产率可为各纯化步骤原料中bst的至少约30%、至少约35%、至少约40%、至少约45%、至少约50%、至少约55%、至少约60%、至少约65%、至少约70%、至少约75%、至少约80%、至少约85%、至少约90%、至少约91%、至少约92%、至少约93%、至少约94%、至少约95%、至少约96%、至少约97%、至少约98%、至少约99%、至少约99.9%或至少约99.99%。
可使用例如sds-page等标准技术,或通过使用蛋白免疫印迹法(westernblot)和elisa分析测量bst多肽,来测定纯度。举例来说,可产生针对由阴性对照酵母发酵和阳离子交换回收而分离的蛋白质的多克隆抗体。这些抗体也可用于探查污染性宿主细胞蛋白质的存在。
rp-hplc材料vydacc4(维达科公司(vydac))由硅胶粒子组成,其表面带有c4烷基链。bst多肽与蛋白质杂质的分离是基于疏水相互作用强度的差别。洗脱是用稀三氟乙酸中的乙腈梯度执行的。使用不锈钢柱(填充2.8到3.2升vydacc4硅胶)执行制备型hplc。通过添加三氟乙酸来酸化羟基磷灰石ultrogel洗出液,并将其装载到vydacc4柱上。使用稀三氟乙酸中的乙腈梯度来进行洗涤和洗脱。收集洗脱部分,并立即用磷酸盐缓冲液中和。汇集在ipc限度内的bst多肽洗脱部分。
deaesepharose(法玛西亚生物技术公司(pharmacia))材料由二乙氨基乙基(deae)基团共价结合于琼脂糖凝胶珠粒的表面而组成。bst多肽与deae基团的结合是由离子相互作用所介导。乙腈和三氟乙酸无滞留地通过柱。在洗掉这些物质之后,通过用低ph值的乙酸盐缓冲液洗涤柱来去除微量杂质。随后,用中性磷酸盐缓冲液洗涤柱,并用离子强度较高的缓冲液洗脱bst多肽。用deaesepharosefastflow填充柱。调整柱体积,以确保bst多肽的装载量在每毫升凝胶3-10mgbst多肽的范围内。用水和平衡缓冲液(磷酸钠/钾)洗涤柱。装载汇集的hplc洗出液中的洗脱部分,并用平衡缓冲液洗涤柱。随后用洗涤缓冲液(乙酸钠缓冲液)洗涤柱,接着用平衡缓冲液洗涤。随后,用洗脱缓冲液(氯化钠、磷酸钠/钾)从柱洗脱bst多肽,并根据主要洗脱曲线收集于单一洗脱部分中。将deaesepharose柱的洗出液调整到指定传导率。使所得原料药无菌过滤到铁氟隆(teflon)瓶中,并在-70℃下储存。
使用的其它方法包括(但不限于)去除内毒素的步骤。内毒素是位于革兰氏阴性宿主细胞(例如大肠杆菌)的外侧膜上的脂多糖(lps)。所属领域技术人员已知用于降低内毒素含量的方法,且这些方法包括(但不限于)使用二氧化硅支撑物、玻璃粉或羟基磷灰石的纯化技术;反相色谱、亲和色谱、尺寸排阻色谱、阴离子交换色谱;疏水相互作用色谱;这些方法的组合等。可能需要修改或其它方法以从相关多肽中去除污染物,例如共迁移蛋白质。用于测量内毒素含量的方法已为所属领域技术人员所知,且这些方法包括(但不限于)鲎变形细胞溶解物(limulusamebocytelysate,lal)分析。endosafetm-pts分析是一种单管比色系统,其利用预先装载有lal试剂、发色底物和对照标准品内毒素的药包以及手持分光光度计。其它方法包括(但不限于)动力学lal法,其为比浊度分析法,并且使用96孔形式。
多种方法和程序可用来评估包含一个或一个以上非天然编码氨基酸的bst蛋白的产率和纯度,包括(但不限于)布莱德福德分析(bradfordassay)、sds-page、银染色sds-page、考马斯染色sds-page(coomassiestainedsds-page)、质谱(包括(但不限于)maldi-tof),以及所属领域技术人员已知的其它表征蛋白质的方法。
其它方法包括(但不限于):sds-page联合蛋白质染色方法、免疫印迹、基质辅助激光解吸附/离子化质谱(matrixassistedlaserdesorption/ionization-massspectrometry,maldi-ms)、液相色谱/质谱、等电聚焦、分析型阴离子交换、色谱聚焦和圆二色谱。
viii.替代系统中的表达
已使用数种策略以在非重组宿主细胞、诱变的宿主细胞或无细胞系统中将非天然氨基酸引入蛋白质中。这些系统也适用于制备本发明的bst多肽。具有反应性侧链的氨基酸(例如lys、cys和tyr)的衍生化会使得赖氨酸转化为n2-乙酰基-赖氨酸。化学合成也是一种并入非天然氨基酸的直截了当的方法。随着近来肽片段的酶促接合和天然化学接合的发展,有可能制造出更大的蛋白质。例如参见p.e.达沃森(p.e.dawson)和s.b.h.肯特(s.b.h.kent),生物化学年评(annu.rev.biochem),69:923(2000)。化学肽接合和天然化学接合方法描述于美国专利第6,184,344号、美国专利公开案第2004/0138412号、美国专利公开案第2003/0208046号、wo02/098902和wo03/04223中,这些专利都以引用的方式并入本文中。已使用将经所需非天然氨基酸以化学方式酰基化的抑制trna加到能够支持蛋白质生物合成的体外提取物中的一般体外生物合成方法,将超过100个非天然氨基酸位点特异性地并入实际上任何尺寸的多种蛋白质中。例如参见v.w.柯尼西(v.w.cornish),d.孟德尔(d.mendel)和p.g.查鲁兹(p.g.schultz),德国应用化学(英文版)1995,34:621(1995);c.j.诺伦(c.j.noren),s.j.安托尼-卡黑尔(s.j.anthony-cahill),m.c.格雷夫斯(m.c.griffith),p.g.查鲁兹(p.g.schultz),用于将非天然氨基酸位点特异性并入蛋白质中的一般方法(ageneralmethodforsite-specificincorporationofunnaturalaminoacidsintoproteins),科学244:182-188(1989);和j.d.贝恩(j.d.bain),c.g.格雷布(c.g.glabe),t.a.迪克斯(t.a.dix),a.r.查柏林(a.r.chamberlin),e.s.迪阿拉(e.s.diala),用于将非天然氨基酸并入多肽中的生物合成方法(biosyntheticsite-specificincorporationofanon-naturalaminoacidintoapolypeptide),美国化学协会杂志111:8013-8014(1989)。已经将多种官能团引入蛋白质中以供蛋白质稳定性、蛋白质折叠、酶机理和信号转导的研究。
已开发出一种称作选择性压力并入(selectivepressureincorporation)的体内方法,来研究野生型合成酶的杂乱性。例如参见n.鲍里斯(n.budisa),c.明克斯(c.minks),s.阿尔弗雷德(s.alefelder),w.温格尔(w.wenger),f.m.董(f.m.dong),l.莫罗德(l.moroder)和r.胡波(r.huber),美国实验生物学联合会会刊(fasebj.),13:41(1999)。使向细胞供应特定天然氨基酸的相关代谢路径关闭的营养缺陷型菌株生长于含有有限浓度天然氨基酸的基本培养基中,而目标基因的转录受到阻遏。在稳定生长期开始时,天然氨基酸被耗尽,并且被非天然氨基酸类似物置换。诱导重组蛋白表达将使含有非天然类似物的蛋白质积聚。举例来说,曾使用这种策略将邻氟苯丙氨酸、间氟苯丙氨酸和对氟苯丙氨酸并入蛋白质中,且蛋白质在uv光谱中展现两个易于鉴别的特征性肩峰,例如参见c.明克斯(c.minks),r.胡波(r.huber),l.莫罗德(l.moroder)和n.鲍里斯(n.budisa),分析生物化学(anal.biochem.),284:29(2000);曾使用三氟甲硫氨酸代替噬菌体t4溶菌酶中的甲硫氨酸,并利用19fnmr研究其与壳寡糖配体的相互作用,例如参见h.杜威尔(h.duewel),e.道博(e.daub),v.罗宾逊(v.robinson)和j.f.霍尼克(j.f.honek),生物化学(biochemistry),36:3404(1997);而且已并入三氟亮氨酸代替亮氨酸,从而使亮氨酸拉链蛋白的热稳定性和化学稳定性增加。例如参见y.杨(y.tang),g.格瑞兰达(g.ghirlanda),w.a.派特塔(w.a.petka),t.中岛(t.nakajima),w.f.德格雷多(w.f.degrado)和d.a.斯瑞尔(d.a.tirrell),德国应用化学(英文版).40:1494(2001)。此外,还将硒代甲硫氨酸和碲代甲硫氨酸并入各种重组蛋白中以促进x射线结晶学中的相溶解。例如参见w.a.汉迪克森(w.a.hendrickson),j.r.霍顿(j.r.horton)和d.m.雷玛斯特(d.m.lemaster),欧洲分子生物学组织期刊(emboj.),9:1665(1990);j.o.波利斯(j.o.boles),k.莱文斯基(k.lewinski),m.坤克(m.kunkle),j.d.奥多姆(j.d.odom),b.顿雷普(b.dunlap),l.勒博达(l.lebioda)和m.哈塔达(m.hatada),自然结构生物学(nat.struct.biol.).1:283(1994);n.布迪萨(n.budisa),b.斯特普(b.steipe),p.德曼基(p.demange),c.艾克斯柯恩(c.eckerskorn),j.科勒曼(j.kellermann)和r.胡波(r.huber),欧洲生物化学杂志(eur.j.biochem.).230:788(1995);和n.布迪萨(n.budisa),w.卡恩博克(w.karnbrock),s.斯坦恩布奇(s.steinbacher),a.汉姆(a.humm),l.普雷德(l.prade),t.纽费恩德(t.neuefeind),l.莫罗德(l.moroder)和r.胡波(r.huber),分子生物学杂志(j.mol.biol.).270:616(1997)。也已有效地并入具有烯或炔官能团的甲硫氨酸类似物,由此允许借助化学方式对蛋白质进行其它修饰。例如参见j.c.m.范海斯特(j.c.vanhest)和d.a.特瑞尔(d.a.tirrell),欧洲生物学化学会联盟通讯(febslett.),428:68(1998);j.c.范海斯特,k.l.奇科(k.l.kiick)和d.a.特瑞尔,美国化学协会杂志.122:1282(2000);以及k.l.奇科和d.a.特瑞尔,四面体通讯(tetrahedron),56:9487(2000);美国专利第6,586,207号;美国专利公开案第2002/0042097号,都以引用的方式并入本文中。
这种方法的成功取决于氨酰基trna合成酶对非天然氨基酸类似物的识别,而所述合成酶一般需要高选择性来确保蛋白质翻译的保真度。扩展这种方法的范围的一种方式是放宽氨酰基trna合成酶的底物特异性,这已在少数情况下实现。举例来说,在大肠杆菌苯丙氨酰基trna合成酶(phers)中用gly置换ala294可增加底物结合口袋的尺寸,并引起对氯苯丙氨酸(p-cl-phe)对trnaphe的酰基化。参见m.爱博(m.ibba),p.卡斯特(p.kast)和h.汉尼克(h.hennecke),生物化学(biochemistry),33:7107(1994)。带有这种突变phers的大肠杆菌菌株能够并入对氯苯丙氨酸或对溴苯丙氨酸来替代苯丙氨酸。例如参见m.爱博(m.ibba)和h.汉尼克(h.hennecke),欧洲生物学化学会联盟通讯(febslett.).364:272(1995);以及n.夏尔玛(n.sharma),r.福特(r.furter),p.卡斯特(p.kast)和d.a.特瑞尔(d.a.tirrell),欧洲生物学化学会联盟通讯,467:37(2000)。类似地,显示接近大肠杆菌酪氨酰基trna合成酶的氨基酸结合位点的点突变phe130ser并入比酪氨酸有效地并入重氮酪氨酸。参见f.滨野高久(f.hamano-takaku),t.岩间(t.iwama),s.齐藤矢野(s.saito-yano),k.高久(k.takaku),y.门田(y.monden),m.北田(m.kitabatake),d.索尔(d.soll)和s.西村(s.nishimura),生物化学杂志(j.biol.chem.),275:40324(2000)。
在体内将非天然氨基酸并入蛋白质中的另一策略为修饰具有校对机制的合成酶。这些合成酶不能区分结构与同源天然氨基酸类似的氨基酸,并因此将其活化。此错误在单独位点上得到校正,使得来自trna的错配氨基酸去酰基化以保持蛋白质翻译的保真度。如果合成酶失去校对活性,那么错误活化的结构类似物可避开编辑功能且被并入。近来已利用缬氨酰基trna合成酶(valrs)来证实这种方法。参见v.道宁(v.doring),h.d.磨兹(h.d.mootz),l.a.南格(l.a.nangle),t.l.汉迪克森(t.l.hendrickson),范德克雷兹拉格德(v.decrecy-lagard),p.斯奇迈尔(p.schimmel)和p.玛丽尔(p.marliere),科学,292:501(2001)。valrs可使带有cys、thr或氨基丁酸(abu)的trnaval错误氨酰基化;随后经由编辑域水解这些非同源氨基酸。在随机诱变大肠杆菌染色体之后,选择在valrs的编辑位点中具有突变的突变大肠杆菌菌株。这种编辑缺陷型valrs错误地使trnaval装有cys。由于abu与cys在空间上类似(cys的-sh基团经abu中的-ch3置换),以致当这种突变大肠杆菌菌株在abu存在下生长时,突变valrs也将abu并入蛋白质中。质谱分析显示,在天然蛋白质的各缬氨酸位置处约24%的缬氨酸经abu置换。
固相合成和半合成方法也已允许合成多种含有新颖氨基酸的蛋白质。举例来说,参见以下出版物和其中所引用的参考文献:克里克f.j.c.(crick,f.j.c.),巴雷特l.(barrett,l.),布雷纳s.(brenner,s.),沃特托比r.(watts-tobin,r.)蛋白质遗传代码的一般性质。(generalnatureofthegeneticcodeforproteins.)自然(nature),192:1227-1232(1961);霍夫曼k.(hofmann,k.),伯恩h.(bonn,h.)有关多肽的研究。xxxvi.吡唑-咪唑置换对s-肽片段的s-蛋白质活化效力的影响。(studiesonpolypeptides.xxxvi.theeffectofpyrazole-imidazolereplacementsonthes-proteinactivatingpotencyofans-peptidefragment),美国化学杂志(j.amchem),88(24):5914-5919(1966);卡瑟e.t.(kaiser,e.t.)生物活性肽和包括酶在内的蛋白质的合成方法(syntheticapproachestobiologicallyactivepeptidesandproteinsincludingenyzmes),化学研究述评(acechemres),22:47-54(1989);中塚t.(nakatsuka,t.),佐佐木t.(sasaki,t.),卡瑟e.t.(kaiser,e.t.)由半合成酶硫代枯草杆菌蛋白酶催化的肽片段偶合(peptidesegmentcouplingcatalyzedbythesemisyntheticenzymethiosubtilisin),美国化学协会杂志,109:3808-3810(1987);斯奇诺泽m.(schnolzer,m.),肯特sbh.(kent,sbh.)通过接合未受保护的合成肽构建蛋白质:主链经工程改造的hiv蛋白酶(constructingproteinsbydovetailingunprotectedsyntheticpeptides:backbone-engineeredhivprotease),科学,256(5054),221-225(1992);查肯i.m.(chaiken,i.m.)半合成肽和蛋白质(semisyntheticpeptidesandproteins),生物化学学报(crccritrevbiochem),11(3):255-301(1981);奥夫德r.e.(offord,r.e.)如何借助化学方式进行蛋白质工程改造?(proteinengineeringbychemicalmeans?)蛋白质工程(proteineng.),1(3):151-157(1987);和杰克森d.y.(jackson,d.y.),伯尼尔j.(burnier,j.),权c(quan,c),斯坦利m.(stanley,m.),汤姆j.(tom,j.),威尔斯j.a.(wells,j.a.)用于总体合成带有非天然催化残基对核糖核酸酶a的经过设计的肽接合酶(adesignedpeptideligasefortotalsynthesisofribonucleaseawithunnaturalcatalyticresidues),科学,266,(5183):243(1994)。
已使用化学修饰在体外将包括辅因子、自旋标记和寡核苷酸在内的多种非天然侧链引入蛋白质中。例如参见,克雷d.r.(corey,d.r.),查鲁兹p.g.(schultz,p.g.)产生杂交序列特异性单链脱氧核糖核酸酶(generationofahybridsequence-specificsingle-strandeddeoxyribonuclease),科学,238(4832):1401-1403(1987);卡瑟e.t.(kaiser,e.t.),劳伦斯d.s.(lawrenced.s.),洛克塔s.e.)(rokita,s.e.)酶特异性的化学修饰(thechemicalmodificationofenzymaticspecificity),生物化学年评(annurevbiochem),54:565-595(1985);卡瑟e.t.,劳伦斯d.s.,酶活性位点的化学突变(chemicalmutationofenyzmeactivesites),科学,226(4674):505-511(1984);尼特k.e.(neet,k.e.),南西a(nancia),柯西兰德d.e.(koshland,d.e.)硫醇-枯草杆菌蛋白酶的特性(propertiesofthiol-subtilisin),生物化学杂志(jbiol.chem.)243(24):6392-6401(1968);博拉格l.(polgar,l.)(编),m.l.本德尔(m.l.bender.)含有合成形成的活性位点的新酶。硫醇-枯草杆菌蛋白酶。(anewenzymecontainingasyntheticallyformedactivesite.thiol-subtilisin.)美国化学协会杂志,88:3153-3154(1966);和波拉克s.j.(pollack,s.j.),中山g.(nakayama,g.),查鲁兹p.g.(schultz,p.g.)将亲核基团和光谱探针引入抗体组合位点中(introductionofnucleophilesandspectroscopicprobesintoantibodycombiningsites),科学,242(4881):1038-1040(1988)。
另外,也已经使用以化学方式修饰的氨酰基trna的生物合成方法来将数种生物物理探针并入在体外合成的蛋白质中。参见以下出版物和其中引用的参考文献:布伦纳j.(brunner,j.)新的光标记和交联方法(newphotolabelingandcrosslinkingmethods),生物化学年评(annu.revbiochem),62:483-514(1993);和克雷格u.c.(krieg,u.c.),沃特p.(walter,p.),霍恩森a.e.(hohnson,a.e.)信号识别粒子的54kda多肽新生催乳激素前体的信号序列的光交联(photocrosslinkingofthesignalsequenceofnascentpreprolactinofthe54-kilodaltonpolypeptideofthesignalrecognitionparticle),美国国家科学院院刊,83(22):8604-8608(1986)。
先前已证实,在体外通过将以化学方式氨酰基化的抑制trna添加到经含有所需琥珀无义突变的基因编程的蛋白质合成反应中,可将非天然氨基酸位点特异性地并入蛋白质中。使用这些方法,可使用对于特定氨基酸来说为营养缺陷型的菌株,用密切结构同系物取代20种常见氨基酸中的多种氨基酸,例如,用氟苯丙氨酸取代苯丙氨酸。例如参见诺伦c.j.(noren,c.j.),安托尼-卡黑尔(anthony-cahill),格雷夫斯m.c.(griffith,m.c.),查鲁兹p.g.(schultz,p.g.)用于奖非天然氨基酸位点特异性并入蛋白质中的一般方法(ageneralmethodforsite-specificincorporationofunnaturalaminoacidsintoproteins),科学,244:182-188(1989);m.w.诺瓦克(m.w.nowak)等人,科学,268:439-42(1995);贝恩j.d.(bain,j.d.),格雷布c.g.(glabe,c.g.),迪克斯t.a.(dix,t.a.),查柏林a.r.(chamberlin,a.r.),迪阿拉e.s.(diala,e.s.)将非天然氨基酸位点特异性并入多肽中的生物合成方法(biosyntheticsite-specificincorporationofanon-naturalaminoacidintoapolypeptide),美国化学协会杂志,111:8013-8014(1989);n.鲍里斯(n.budisa)等人,美国实验生物学联合会会刊(fasebj.)13:41-51(1999);埃尔曼j.a.(ellman,j.a.),孟德尔d.(mendel,d.),安托尼-卡黑尔s.(anthony-cahill,s.),诺伦c.j.,查鲁兹p.g.,用于将非天然氨基酸位点特异性引入蛋白质中的生物合成方法(biosyntheticmethodforintroducingunnaturalaminoacidssite-specificallyintoproteins),酶学方法(methodsinenz.),第202卷,301-336(1992);以及孟德尔d.,柯尼西v.w.(cornish,v.w.)和查鲁兹p.g.,利用扩充的遗传代码进行定点诱变(site-directedmutagenesiswithanexpandedgeneticcode),生物物理与生物大分子结构评述(annurevbiophys.biomolstruct.)24,435-62(1995)。
举例来说,制备识别终止密码子uag的抑制trna,并用非天然氨基酸以化学方式使其氨酰基化。使用常规定点诱变在蛋白质基因中的相关位点处引入终止密码子tag。例如参见赛尔j.r.(sayers,j.r.),斯奇米特w.(schmidt,w.),艾克斯坦f.(eckstein,f.)5'-3'核酸外切酶在基于硫代磷酸酯的寡核苷酸引导的诱变中的应用(5'-3'exonucleaseinphosphorothioate-basedoligonucleotide-directedmutagenesis),核酸研究,16(3):791-802(1988)。当将酰基化抑制trna和突变基因组合于体外转录/翻译系统中时,对uag密码子反应而并入非天然氨基酸,得到在指定位置含有所述氨基酸的蛋白质。使用[3h]-phe进行的实验和使用α-羟基酸进行的实验证实,仅在uag密码子指定的位置处并入所需氨基酸,而且未在蛋白质中的任何其它位点并入此氨基酸。例如参见,诺伦(noren)等人,同上文;小林(kobayashi)等人,(2003)自然结构生物学(naturestructuralbiology)10(6):425-432;和埃尔曼j.a.(ellman,j.a.),孟德尔d.(mendel,d.),查鲁兹p.g.(schultz,p.g.)将新颖主链结构位点特异性并入蛋白质中(site-specificincorporationofnovelbackbonestructuresintoproteins),科学,255(5041):197-200(1992)。
可借助于任何方法或技术(包括(但不限于)化学或酶促氨酰基化)利用所需氨基酸将trna氨酰基化。
氨酰基化可利用氨酰基trna合成酶或其它酶分子(包括(但不限于)核糖酶)实现。术语“核糖酶”可与“催化性rna”互换。克奇(cech)和同事(克奇,1987,科学.236:1532-1539;麦克科勒(mccorkle)等人,1987,生物化学概念(conceptsbiochem.)64:221-226)证实存在可充当催化剂的天然存在的rna(核糖酶)。然而,尽管仅显示这些天然rna催化剂对核糖核酸底物作用以供裂解和剪接,但近来核糖酶人工开发的发展已将催化谱系扩展到各种化学反应。多项研究已鉴别出可催化自身(2')3'末端氨酰基rna键的rna分子(爱兰克卡尔(illangakekare)等人,1995,科学,267:643-647),以及可将氨基酸从一个rna分子转移到另一个的rna分子(罗斯(lohse)等人,1996,自然(nature)381:442-444)。
美国专利申请公开案2003/0228593(以引用的方式并入本文中)描述构建核糖体的方法和其于利用天然编码和非天然编码氨基酸使trna氨酰基化中的用途。可使trna氨酰基化的酶分子(包括(但不限于)核糖酶)的底物固定形式能够有效亲和纯化氨酰基化的产物。适合底物的实例包括琼脂糖、琼脂糖凝胶和磁珠。用于氨酰基化的底物固定形式核糖酶的制备和用途描述于化学与生物学(chemistryandbiology)2003,10:1077-1084和美国专利申请公开案2003/0228593中,其以引用的方式并入本文中。
化学氨酰基化方法包括(但不限于)以下文献中介绍的避免在氨酰基化过程中使用合成酶的方法:海奇特(hecht)和同事(海奇特s.m.(hecht,s.m.)化学研究述评(ace.chem.res.)1992,25,545;海克勒t.g.(heckler,t.g.);罗塞尔j.r.(roesser,j.r.);徐c(xu,c);常p.(chang,p.);海奇特s.m.,生物化学(biochemistry)1988,27,7254;海奇特s.m.;阿尔福德b.l.(alford,b.l.);黑田y.(kuroda,y.);北野s.(kitano,s.),生物化学杂志(j.biol.chem.)1978,253,4517);和查鲁兹(schultz),查柏林(chamberlin),道赫提(dougherty)等人(柯尼西v.w.(cornish,v.w.);孟德尔d.(mendel,d.);查鲁兹p.g.(schultz,p.g.)德国应用化学(英文版)1995,34,621;罗宾森s.a.(robertson,s.a.);埃尔曼j.a.(ellman,j.a.);查鲁兹p.g.,美国化学协会杂志1991,113,2722;诺伦c.j.(noren,c.j.);安托尼-卡黑尔s.j.(anthony-cahill,s.j.);格雷夫斯m.c.(griffith,m.c.);查鲁兹p.g.,科学1989,244,182;贝恩j.d.(bain,j.d.);格雷布c.g.(glabe,c.g.);迪克斯t.a.(dix,t.a.);查柏林a.r.(chamberlin,a.r.),美国化学协会杂志1989,111,8013;贝恩j.d.等人,自然(nature)1992,356,537;盖里梵j.p.(gallivan,j.p.);莱斯特h.a.(lester,h.a.);道赫提d.a(dougherty,d.a);化学与生物学(chem.biol.)1997,4,740;特卡提(turcatti)等人,生物化学杂志(j.biol.chem.)1996,271,19991;诺瓦克m.w.(nowak,m.w.)等人科学,1995,268,439;萨克斯m.e.(saks,m.e.)等人生物化学杂志1996,271,23169;村上t.(hohsaka,t.)等人美国化学协会杂志1999,121,34)。所述方法或其它化学氨酰基化方法都可用于使trna分子氨酰基化。
产生催化性rna的方法可涉及产生单独的随机核糖酶序列的池;对所述池执行定向演化;针对所需氨酰基化活性筛选所述池;和选择展现所需氨酰基化活性的核糖酶序列。
核糖酶可包含促进酰基化活性的基序和/或区域,例如ggu基序和富集u的区域。举例来说,已报导富集u的区域可促进氨基酸底物的识别,而ggu基序可与trna的3'末端形成碱基对。ggu基序与富集u区域的组合将促进同时识别氨基酸与trna,并因此促进trna3'末端的氨酰基化。
可通过使用与trnaasncccg结合的部分随机r24mini进行体外选择,随后系统的工程改造活性克隆中所见的一致序列,来产生核糖酶。利用此方法获得的示范性核糖酶称为“fx3核糖酶”,并且描述于美国公开申请案第2003/0228593号(其内容以引用的方式并入本文中)中,所述核糖酶将充当合成载有同源非天然氨基酸的各种氨酰基化trna的通用催化剂。
在基质上固定可用于促成氨酰基化trna的有效亲和纯化。适合底物的实例包括(但不限于)琼脂糖、琼脂糖凝胶和磁珠。可利用rna的化学结构将核糖酶固定于树脂上,例如,rna核糖上的3'-顺-二醇可经高碘酸盐氧化以得到相应的二醛,以便利将rna固定于树脂上。可使用各种类型的树脂,包括廉价的酰肼树脂,其中还原胺化使得树脂与核糖酶之间相互作用而形成不可逆键。可借助于此种柱上氨酰基化技术来显著促进氨酰基trna的合成。卡罗利斯(kourouklis)等人,方法(methods)2005;36:239-4中描述一种基于柱的氨酰基化系统。
氨酰基化trna的分离可借助多种方式实现。一种适合的方法是利用缓冲液(例如含有10mmedta的乙酸钠溶液;含有50mmn-(2-羟乙基)哌嗪-n'-(3-丙烷磺酸)、12.5mmkcl(ph7.0)、10mmedta的缓冲液;或者简单的edta缓冲的水(ph7.0))从柱中洗脱出氨酰基化trna。
可将氨酰基化trna加到翻译反应中,以便将使trna氨酰基化的氨基酸并入翻译反应所产生的多肽中的所选位置中。可使用本发明的氨酰基化trna的翻译系统的实例包括(但不限于)细胞溶解产物。细胞溶解产物提供由输入mrna体外翻译多肽必需的反应组分。所述反应组分的实例包括(但不限于)核糖体蛋白、rrna、氨基酸、trna、gtp、atp、翻译起始因子和延长因子以及其它与翻译有关的因子。此外,翻译系统也可为批量翻译或分隔翻译(compartmentalizedtranslation)。批量翻译系统组合单一隔室中的反应组分,而分隔翻译系统使翻译反应组分与可抑制翻译效率的反应产物分离。这些翻译系统在市面上都有售。
此外,可使用偶合式转录/翻译系统。偶合式转录/翻译系统允许将输入dna转录成相应的mrna,而mrna又经由反应组分翻译。市售偶合式转录/翻译的实例为rapidtranslationsystem(rts,罗氏公司(rocheinc.))。这一系统包括含有大肠杆菌溶解产物的混合物,用于提供例如核糖体和翻译因子等翻译组分。此外,还包括rna聚合酶以将输入dna转录成mrna模板,以便用于翻译。rts可经由插入反应隔室(包括供应/废料隔室和转录/翻译隔室)之间的膜来分隔反应组分。
可利用包括(但不限于)转移酶、聚合酶、催化性抗体、多功能蛋白等其它试剂使trna氨酰基化。
史蒂芬(stephan)于2005年10月10日的科学家(scientist)第30-33页中描述其它将非天然编码氨基酸并入蛋白质中的方法。鲁(lu)等人于2001年10月的分子细胞(molcell.);8(4):759-69中描述一种以化学方式将蛋白质与含有非天然氨基酸的合成肽接合的方法(称为蛋白质接合法)。
也已使用微注射技术将非天然氨基酸并入蛋白质中。例如参见,m.w.诺瓦克(m.w.nowak),p.c.卡尔雷(p.c.kearney),j.r.萨普森(j.r.sampson),m.e.萨克斯(m.e.saks),c.g.拉伯卡(c.g.labarca),s.k.希文曼(s.k.silverman),w.g.钟(w.g.zhong),j.托森(j.thorson),j.n.阿贝森(j.n.abelson),n.戴维森(n.davidson),p.g.查鲁兹(p.g.schultz),d.a.道赫提(d.a.dougherty)和h.a.莱斯特(h.a.lester),科学,268:439(1995);以及d.a.道赫提,化学生物学现代观点(curr.opin.chem.biol.),4:645(2000)。将爪蟾卵母细胞(xenopusoocyte)与体外产生的以下两种rna物质共注射:在相关氨基酸位置具有uag终止密码子的编码目标蛋白的mrna,和经所需非天然氨基酸氨酰基化的琥珀抑制trna。随后,卵母细胞的翻译机器在uag指定的位置插入非天然氨基酸。这种方法允许进行一般不适用于体外表达系统的整合膜蛋白的体内结构-功能研究。实例包括将荧光氨基酸并入速激肽神经激肽-2受体中,通过荧光共振能量转移来测量距离,例如参见g.特卡提(g.turcatti),k.尼曼斯(k.nemeth),m.d.艾德格顿(m.d.edgerton),u.米瑟斯(u.meseth),f.塔拉波特(f.talabot),m.佩斯奇(m.peitsch),j.诺里斯(j.knowles),h.沃基尔(h.vogel)和a.查利特(a.chollet),生物化学杂志(j.biol.chem.),271:19991(1996);并入生物素化氨基酸以鉴别离子通道中表面暴露的残基,例如参见j.p.盖里梵(j.p.gallivan),h.a.莱斯特(h.a.lester)和d.a.道赫提(d.a.dougherty),化学与生物学(chem.biol.),4:739(1997);使用经笼蔽的酪氨酸类似物实时监测离子通道的构象变化,例如参见j.c米勒(j.c.miller),s.k.希文曼(s.k.silverman),p.m.英格兰德(p.m.england),d.a.道赫提和h.a.莱斯特,神经元(neuron),20:619(1998);以及使用α羟基氨基酸改变离子通道主链以探查其门控机制。例如参见p.m.英格兰德(p.m.england),y.张(y.zhang),d.a.道赫提(d.a.dougherty)和h.a.莱斯特(h.a.lester),细胞(cell),96:89(1999);以及t.鲁(t.lu),a.y.亭格(a.y.ting),j.玛恩兰德(j.mainland),l.y.简(l.y.jan),p.g.查鲁兹(p.g.schultz)和j.杨(j.yang),自然—神经科学(nat.neurosci.),4:239(2001)。
在体内直接将非天然氨基酸并入蛋白质中的能力具有多种益处,包括(但不限于)突变蛋白的高产率、技术简易性、在细胞中或在活生物体中研究突变蛋白的可能性以及这些突变蛋白用于治疗性治疗和诊断应用的用途。将具有各种尺寸、酸度、亲核性、疏水性和其它特性的非天然氨基酸包括在蛋白质中的能力可极大地扩展合理且系统地操纵蛋白质结构的能力,从而探查蛋白质功能,并产生具有新颖特性的新蛋白质或生物体。
在位点特异性地并入p-f-phe的一次尝试中,将酵母琥珀抑制trnaphecua/苯丙氨酰基trna合成酶对用于p-f-phe抗性、phe营养缺陷型大肠杆菌菌株中。例如参见r.佛特(r.furter),蛋白质科学(proteinsci.),7:419(1998)。
也可能使用无细胞(体外)翻译系统来表达本发明的bst聚核苷酸。翻译系统可为细胞或无细胞翻译系统,并且可为原核或真核翻译系统。细胞翻译系统包括(但不限于)全细胞制剂,例如可将所需核酸序列转录成mrna并且翻译mrna的渗透细胞或细胞培养物。无细胞翻译系统在市面上有售,并且众所周知多种不同类型和系统。无细胞系统的实例包括(但不限于)原核细胞溶解产物,例如大肠杆菌溶解产物;和真核细胞溶解产物,例如麦胚提取物、昆虫细胞溶解产物、兔网织红细胞溶解产物、兔卵母细胞溶解产物和人细胞溶解产物。当糖基化、磷酸化或以其它方式修饰所得蛋白质时,由于许多修饰只可能在真核系统中发生,故优选真核提取物或溶解产物。这些提取物和溶解产物中有一些在市面上有售(普洛麦格公司(promega),位于威斯康星州麦迪逊(madison,wis.);赛特杰公司(stratagene),位于加利福尼亚州拉荷亚(lajolla,calif.);安玛西亚生物技术公司(amersham),位于伊利诺伊州阿灵顿海茨(arlingtonheights,ill.);gibco/brl公司,位于纽约州格兰德岛(grandisland,n.y.))。也可使用膜提取物(例如含有微粒体膜的犬胰腺提取物),其适用于翻译分泌蛋白质。在这些可包括mrna作为模板(体外翻译)或dna作为模板(组合式体外转录与翻译)的系统中,体外合成是由核糖体所引导。曾进行过相当多的尝试来开发无细胞蛋白质表达系统。例如参见,肯姆d.m.(kim,d.m.)和j.r.斯沃兹(j.r.swartz),生物技术与生物工程(biotechnologyandbioengineering),74:309-316(2001);肯姆d.m.和j.r.斯沃兹,生物技术通讯(biotechnologyletters),22,1537-1542,(2000);肯姆d.m.和j.r.斯沃兹,生物技术进展(biotechnologyprogress),16,385-390,(2000);肯姆d.m.和j.r.斯沃兹,生物技术与生物工程,66,180-188,(1999);以及帕特耐克r.(patnaik,r.)和j.r.斯沃兹,,生物技术(biotechniques)24,862-868,(1998);美国专利第6,337,191号;美国专利公开案第2002/0081660号;wo00/55353;wo90/05785,都是以引用的方式并入本文中。另一种可用于表达包含非天然编码氨基酸的bst多肽的方法包括mrna-肽融合技术。例如参见r.罗伯特(r.roberts)和j.斯佐塔克(j.szostak),美国国家科学院院刊(usa)94:12297-12302(1997);a.弗兰克尔(a.frankel)等人,化学与生物学(chemistry&biology)10:1043-1050(2003)。在本方法中,在核糖体上将连接嘌呤霉素(puromycin)的mrna模板翻译成肽。如果已对一个或一个以上trna分子进行修饰,那么也可将非天然氨基酸并入所述肽中。在读取了最后一个mrna密码子后,嘌呤霉素将捕获肽的c末端。如果发现所得mrna-肽结合物在体外分析中具有引人关注的特性,那么可容易地由mrna序列揭示其身份。以此方式,可筛选出包含一个或一个以上非天然编码氨基酸的bst多肽的文库,以鉴别具有所需特性的多肽。最近,已报导利用经纯化组分进行的体外核糖体翻译,其允许合成经非天然编码氨基酸取代的肽。例如参见a.福斯特(a.forster)等人,美国国家科学院院刊,100:6353(2003)。
还可使用重构翻译系统。已成功地使用纯化的翻译因子混合物以及溶解产物或补充有纯化翻译因子(例如起始因子-1(if-1)、if-2、if-3(α或β)、延长因子t(ef-tu)或终止因子)的溶解产物的组合将mrna翻译成蛋白质。无细胞系统也可为偶合式转录/翻译系统,其中如现代分子生物学技术(currentprotocolsinmolecularbiology)(f.m.奥斯贝尔(f.m.ausubel)等人编辑,威立在线出版公司(wileyinterscience),1993)(以引用的方式清楚地并入本文中)中所述,将dna引入所述系统中,转录成mrna并翻译mrna。在真核转录系统中转录的rna可为异核rna(hnrna)或者5'端戴帽(7-甲基鸟苷)且3'端加多聚腺苷酸尾的成熟mrna(其宜用于某些翻译系统中)形式。举例来说,在网织红细胞溶解产物系统中,以高效率翻译戴帽的mrna。
ix.与bst多肽偶合的大分子聚合物
可使用本文中所述的组合物、方法、技术和策略对本文中所述的非天然氨基酸多肽进行各种修饰。这些修饰包括将其它官能团并入多肽的非天然氨基酸组分上,所述官能团包括(但不限于)羟烷基淀粉(has)、羟乙基淀粉(hes)、标记、染料、聚合物、水溶性聚合物、聚乙二醇衍生物、光交联剂、放射性核素、细胞毒性化合物、药物、亲和标记、光亲和标记、反应性化合物、树脂、第二蛋白质或者多肽或多肽类似物、抗体或抗体片段、金属螯合剂、辅因子、脂肪酸、碳水化合物、聚核苷酸、dna、rna、反义聚核苷酸、糖、水溶性树突状聚合物、环糊精、抑制性核糖核酸、生物材料、纳米粒子、自旋标记、荧光团、含金属部分、放射性部分、新颖官能团、与其它分子共价或非共价相互作用的基团、光笼蔽部分、光化辐射可激发部分、可光异构化部分、生物素、生物素衍生物、生物素类似物、并有重原子的部分、可化学裂解基团、可光裂解基团、延长的侧链、碳连接糖、氧化还原活性剂、氨基硫代酸、有毒部分、同位素标记的部分、生物物理探针、磷光基团、化学发光基团、电子致密基团、磁性基团、插入基团、发色团、能量转移剂、生物活性剂、可检测标记、小分子、量子点、纳米递质、放射性核苷酸、放射性递质、中子捕获剂,或上述物质的任何组合,或任何其它所需化合物或物质。作为本文中所述组合物、方法、技术和策略的说明性非限制性实例,以下描述将集中在将大分子聚合物添加到非天然氨基酸多肽中,同时应了解,相关组合物、方法、技术和策略也适用于(必要时利用适当修饰,且所属领域技术人员可利用本文的揭示内容进行)添加其它官能团,包括(但不限于)上文所述的官能团。
可将多种大分子聚合物和其它分子连接到本发明的bst多肽,以调节bst多肽的生物特性,和/或向bst分子提供新颖生物特性。这些大分子聚合物可经由天然编码氨基酸、非天然编码氨基酸,或者天然或非天然氨基酸的任何官能取代基,或添加到天然或非天然氨基酸的任何取代基或官能团与bst多肽连接。聚合物的分子量可在广泛范围内,包括(但不限于)介于约100da与约100,000da之间或更高。聚合物的分子量可介于约100da与约100,000da之间,包括(但不限于)100,000da、95,000da、90,000da、85,000da、80,000da、75,000da、70,000da、65,000da、60,000da、55,000da、50,000da、45,000da、40,000da、35,000da、30,000da、25,000da、20,000da、15,000da、10,000da、9,000da、8,000da、7,000da、6,000da、5,000da、4,000da、3,000da、2,000da、1,000da、900da、800da、700da、600da、500da、400da、300da、200da和100da。在一些实施例中,聚合物的分子量介于约100da与约50,000da之间。在一些实施例中,聚合物的分子量介于约100da与约40,000da之间。在一些实施例中,聚合物的分子量介于约1,000da与约40,000da之间。在一些实施例中,聚合物的分子量介于约5,000da与约40,000da之间。在一些实施例中,聚合物的分子量介于约10,000da与约40,000da之间。
本发明提供实质上均质的聚合物:蛋白质结合物制剂。本文中使用的“实质上均质”意指据观察,聚合物:蛋白质结合物分子大于总蛋白质的一半。聚合物:蛋白质结合物具有生物活性,并且本文中提供的本发明的“实质上均质”的聚乙二醇化bst多肽制剂是足够均质以展现均质制剂的优势(例如,在临床应用中易于预测批间药物动力学)的多肽制剂。
还可选择制备聚合物:蛋白质结合物分子的混合物,且本文中提供的优势在于,可选择欲包括在混合物中的单一聚合物:蛋白质结合物的比例。因此,必要时,可制备各种蛋白质与各种数量的连接的聚合物部分(即,二聚体、三聚体、四聚体等)的混合物,并将所述结合物与使用本发明方法制备的单聚物:蛋白质结合物组合,且得到具有预定的单聚物:蛋白质结合物比例的混合物。
所选聚合物可为水溶性聚合物,以致与其连接的蛋白质不会在水性环境(例如生理学环境)中沉淀。聚合物可为分支或未分支聚合物。对治疗性使用最终产品制剂来说,聚合物应为医药学上可接受的。
聚合物的实例包括(但不限于)聚烷基醚和其烷氧基封端的类似物(例如,聚氧乙二醇、聚氧乙烯/丙二醇和其经甲氧基或乙氧基封端的类似物,尤其聚氧乙二醇,后者也称为聚乙二醇或peg);聚乙烯吡咯烷酮;聚乙烯烷基醚;聚噁唑啉、聚烷基噁唑啉和聚羟基烷基噁唑啉;聚丙烯酰胺、聚烷基丙烯酰胺和聚羟烷基丙烯酰胺(例如,聚羟丙基甲基丙烯酰胺和其衍生物);聚丙烯酸羟烷基酯;聚唾液酸和其类似物;亲水性肽序列;多糖和其衍生物,包括葡聚糖和葡聚糖衍生物,例如羧甲基葡聚糖、硫酸葡聚糖、氨基葡聚糖、纤维素和其衍生物,例如羧甲基纤维素、羟基烷基纤维素;壳多糖和其衍生物,例如壳聚糖、琥珀酰基壳聚糖、羧甲基壳多糖、羧甲基壳聚糖;透明质酸和其衍生物;淀粉;海藻酸酯;硫酸软骨素;白蛋白;支链淀粉(pullulan)和羧甲基支链淀粉;聚氨基酸和其衍生物,例如聚谷氨酸、聚赖氨酸、聚天冬氨酸、聚天冬酰胺;马来酸酐共聚物,例如苯乙烯马来酸酐共聚物、二乙烯基乙基醚马来酸酐共聚物、聚乙烯醇;其共聚物;其三聚体;其混合物;羟烷基淀粉(has),包括(但不限于)羟乙基淀粉(hes);和前述物质的衍生物。
聚乙二醇分子与蛋白质分子的比例可变化,二者在反应混合物中的浓度也将变化。一般来说,可通过所选聚乙二醇的分子量和可用反应性基团的可用数量来确定最佳比率(就反应效率来说,因为存在极少量过量的未反应的蛋白质或聚合物)。对于分子量,通常聚合物的分子量越高,则可与蛋白质连接的聚合物分子的数量就越少。类似地,当优化这些参数时,应考虑聚合物的分支情况。一般来说,分子量越高(或分支越多),则聚合物:蛋白质的比率就越高。
当涵盖peg:bst多肽结合物时,本文中使用的术语“治疗有效量”是指对动物提供所需益处的量。所述量将随个体差异而不同,并且视多种因素而定,包括患者的总体身体状况和欲治疗的病状的潜在病因。疗法中使用的bst多肽的量将提供可接受的改变速率,并将所需反应保持在有益的水平。所属领域技术人员使用公开可用的材料和程序可容易地确定本发明组合物的治疗有效量。
水溶性聚合物可为任何结构形式,包括(但不限于)线性、叉状或分支形式。通常,水溶性聚合物为聚(烷二醇),例如聚(乙二醇)(peg),但也可使用其它水溶性聚合物。举例来说,使用peg描述本发明的某些实施例。
peg是一种众所周知的水溶性聚合物,其为市售产品,或者可根据所属领域的一般技术人员所知的方法,通过使乙二醇开环聚合来制备(萨德勒(sandler)和卡罗(karo),聚合物合成(polymersynthesis),学术出版社(academicpress),纽约,第3卷,第138-161页)。术语“peg”广泛用于涵盖任何聚乙二醇分子(不考虑peg的尺寸或其末端修饰),并且可由下式表示成与bst多肽连接:
xo-(ch2ch2o)n-ch2ch2-y
其中n为2到10,000,且x为h或末端修饰,包括(但不限于)c1-4烷基、保护基或末端修饰基。
在一些情况下,本发明中使用的peg的一个末端以羟基或甲氧基封端,即,x为h或ch3(“甲氧基peg”)。或者,peg可以用反应性基团封端,由此形成双官能聚合物。典型的反应性基团可包括常用于与20种常见氨基酸中所见的官能团反应的反应性基团(包括(但不限于)马来酰亚胺基、活性碳酸酯(包括(但不限于)对硝基苯酯)、活性酯(包括(但不限于)n-羟基琥珀酰亚胺、对硝基苯酯)和醛)以及对20种常见氨基酸呈惰性但与非天然编码氨基酸中存在的互补官能团特异性反应的官能团(包括(但不限于)叠氮基、炔基)。应注意,上式中以y表示的peg的另一末端将直接或者经由天然存在或非天然编码氨基酸间接连接到bst多肽。举例来说,y可为与多肽胺基(包括(但不限于)赖氨酸的ε胺或n末端)形成的酰胺、氨基甲酸酯或脲键。或者,y可为与硫醇基(包括(但不限于)半胱氨酸的硫醇基)形成的马来酰亚胺键。或者,y可为与经由20种常见氨基酸通常不可得的残基形成的键。举例来说,peg上的叠氮基可与bst多肽上的炔基反应形成胡伊斯根[3+2]环加成产物。或者,peg上的炔基可与非天然编码氨基酸中存在的叠氮基反应形成类似产物。在一些实施例中,强亲核试剂(包括(但不限于)肼、酰肼、羟胺、氨基脲)可与非天然编码氨基酸中存在的醛基或酮基反应形成腙、肟或缩氨基脲,适当时,在一些情况下可利用适当的还原剂处理,再将其还原。或者,可经由非天然编码氨基酸将强亲核基团并入bst多肽中,并使用其优先与水溶性聚合物中存在的酮基或醛基反应。
peg的任何分子质量可视实际需要使用,包括(但不限于)约100da到100,000da,或必要时更高(包括(但不限于)有时为0.1-50kda或10-40kda)。peg的分子量可在广泛范围内,包括(但不限于)介于约100da与约100,000da之间或为更高。peg可介于约100da与约100,000da之间,包括(但不限于)100,000da、95,000da、90,000da、85,000da、80,000da、75,000da、70,000da、65,000da、60,000da、55,000da、50,000da、45,000da、40,000da、35,000da、30,000da、25,000da、20,000da、15,000da、10,000da、9,000da、8,000da、7,000da、6,000da、5,000da、4,000da、3,000da、2,000da、1,000da、900da、800da、700da、600da、500da、400da、300da、200da和100da。在一些实施例中,peg介于约100da与约50,000da之间。在一些实施例中,peg介于约100da与约40,000da之间。在一些实施例中,peg介于约1,000da与约40,000da之间。在一些实施例中,peg介于约5,000da与约40,000da之间。在一些实施例中,peg介于约10,000da与约40,000da之间。还可使用支链peg,包括(但不限于)各链的mw在1-100kda(包括(但不限于)1-50kda或5-20kda)范围内的peg分子。支链peg各链的分子量可(包括(但不限于))介于约1,000da与约100,000da之间或为更高。支链peg各链的分子量可介于约1,000da与约100,000da之间,包括(但不限于)100,000da、95,000da、90,000da、85,000da、80,000da、75,000da、70,000da、65,000da、60,000da、55,000da、50,000da、45,000da、40,000da、35,000da、30,000da、25,000da、20,000da、15,000da、10,000da、9,000da、8,000da、7,000da、6,000da、5,000da、4,000da、3,000da、2,000da和1,000da。在一些实施例中,支链peg各链的分子量介于约1,000da与约50,000da之间。在一些实施例中,支链peg各链的分子量介于约1,000da与约40,000da之间。在一些实施例中,支链peg各链的分子量介于约5,000da与约40,000da之间。在一些实施例中,支链peg各链的分子量介于约5,000da与约20,000da之间。多种peg分子描述于(包括(但不限于))肖尔沃特聚合物公司(shearwaterpolymers,inc.)目录、奈克塔治疗品公司(nektartherapeutics)目录中,二者都以引用方式并入本文中。
一般来说,peg分子的至少一个末端可与非天然编码氨基酸反应。举例来说,可使用带有与氨基酸侧链反应的炔和叠氮部分的peg衍生物将peg与本文中所述的非天然编码氨基酸连接。如果非天然编码氨基酸包含叠氮基,那么peg通常会含有炔部分以形成[3+2]环加成产物,或者会含有含膦基的活性peg物质(即,酯、碳酸酯)以形成酰胺键。或者,如果非天然编码氨基酸包含炔,那么peg通常会含有叠氮部分以形成[3+2]胡伊斯根环加成产物。如果非天然编码氨基酸包含羰基,那么peg通常会包含有效的亲核基团(包括(但不限于)酰肼、肼、羟胺或氨基脲官能团),以便分别形成相应腙、肟和缩氨基脲键。在其它替代性实例中,可逆转上述反应性基团的定向,即,非天然编码氨基酸中的叠氮部分可与含有炔的peg衍生物反应。
在一些实施例中,具有peg衍生物的bst多肽变异体含有可与非天然编码氨基酸侧链上存在的化学官能团反应的化学官能团。
在一些实施例中,本发明提供含叠氮和乙炔的聚合物衍生物,其包含平均分子量为约800da到约100,000da的水溶性聚合物主链。水溶性聚合物的聚合物主链可为聚(乙二醇)。然而,应了解,多种水溶性聚合物,包括(但不限于)聚(乙二醇)和其它相关聚合物(包括聚(葡聚糖)和聚(丙二醇)),也适用于实践本发明,且术语peg和聚(乙二醇)的使用意图涵盖且包括所有这些分子。术语peg包括(但不限于)任何形式的聚(乙二醇),包括双官能peg、多臂peg、衍生化peg、叉状peg、分支peg、侧接peg(即,具有一个或一个以上侧接到聚合物主链的官能团的peg或相关聚合物),或具有可降解键的peg。
peg通常澄清,无色,无臭,可溶于水,对热稳定,对许多化学剂呈惰性,不水解或变质,并且一般无毒。聚(乙二醇)被认为是生物可相容的,据悉,peg能够与活组织或生物体共存而不引起危害。更具体地说,peg实质上为非免疫原性的,据悉,peg不会倾向于在体内产生免疫反应。当与体内具有某种所需功能的分子(例如生物活性剂)连接时,peg倾向于遮蔽这种试剂,并且可减少或消除任何免疫反应,从而使生物体能够耐受所述试剂的存在。peg结合物不会倾向于产生实质免疫反应,或者引起凝固或其它不合需要的作用。具有式--ch2ch2o--(ch2ch2o)n--ch2ch2--(其中n为约3到约4000,通常为约20到约2000)的peg适用于本发明中。在本发明一些实施例中,分子量为约800da到约100,000da的peg尤其适用作聚合物主链。peg的分子量可在广泛范围内,包括(但不限于)介于约100da与约100,000da之间或为更高。peg的分子量可介于约100da与约100,000da之间,包括(但不限于)100,000da、95,000da、90,000da、85,000da、80,000da、75,000da、70,000da、65,000da、60,000da、55,000da、50,000da、45,000da、40,000da、35,000da、30,000da、25,000da、20,000da、15,000da、10,000da、9,000da、8,000da、7,000da、6,000da、5,000da、4,000da、3,000da、2,000da、1,000da、900da、800da、700da、600da、500da、400da、300da、200da和100da。在一些实施例中,peg的分子量介于约100da与约50,000da之间。在一些实施例中,peg的分子量介于约100da与约40,000da之间。在一些实施例中,peg的分子量介于约1,000da与约40,000da之间。在一些实施例中,peg的分子量介于约5,000da与约40,000da之间。在一些实施例中,peg的分子量介于约10,000da与约40,000da之间。
聚合物主链可为线性或分支的。此项技术中一般已知分支聚合物主链。通常,分支聚合物具有中央分支核心部分和与中央分支核心连接的多个线性聚合物链。常用的peg为分支形式,这种形式可通过将环氧乙烷与各种多元醇(例如甘油、甘油寡聚物、季戊四醇和山梨糖醇)加成来制备。中央分支部分也可从数种氨基酸(例如赖氨酸)衍生得到。分支聚(乙二醇)可以通式r(-peg-oh)m表示,其中r衍生自核心部分,例如甘油、甘油寡聚物或季戊四醇,且m表示臂的数量。多臂peg分子(例如美国专利第5,932,462号、第5,643,575号、第5,229,490号、第4,289,872号;美国专利申请案2003/0143596、wo96/21469和wo93/21259中所述的分子,各专利以全文引用的方式并入本文中)也可用作聚合物主链。
分支peg也可为以peg(--ychz2)n表示的叉状peg的形式,其中y为连接子,且z为经由指定长度的原子链与ch连接的活性末端基团。
另一分支形式侧接peg具有沿peg主链而不是在peg链末端的反应性基团,例如羧基。
除这些形式的peg外,聚合物还可制备成在主链中具有弱键或可降解键。举例来说,peg可制备成在聚合物主链中具有易于水解的酯键。如下文所示,此水解导致聚合物裂解成分子量较低的片段:
-peg-co2-peg-+h2o→peg-co2h+ho-peg-
所属领域技术人员应了解,术语聚(乙二醇)或peg表示或包括此项技术中已知的所有形式,包括(但不限于)本文中揭示的形式。
许多其它聚合物也适用于本发明中。在一些实施例中,具有2到约300个末端的水溶性聚合物主链尤其适用于本发明中。适合聚合物的实例包括(但不限于)其它聚(烷二醇),例如聚(丙二醇)(“ppg”)、其共聚物(包括(但不限于)乙二醇与丙二醇的共聚物)、其四聚体、其混合物等。尽管聚合物主链各链的分子量可变化,但通常在约800da到约100,000da、通常约6,000da到约80,000da的范围内。聚合物主链各链的分子量可介于约100da与约100,000da之间,包括(但不限于)100,000da、95,000da、90,000da、85,000da、80,000da、75,000da、70,000da、65,000da、60,000da、55,000da、50,000da、45,000da、40,000da、35,000da、30,000da、25,000da、20,000da、15,000da、10,000da、9,000da、8,000da、7,000da、6,000da、5,000da、4,000da、3,000da、2,000da、1,000da、900da、800da、700da、600da、500da、400da、300da、200da和100da。在一些实施例中,聚合物主链各链的分子量介于约100da与约50,000da之间。在一些实施例中,聚合物主链各链的分子量介于约100da与约40,000da之间。在一些实施例中,聚合物主链各链的分子量介于约1,000da与约40,000da之间。在一些实施例中,聚合物主链各链的分子量介于约5,000da与约40,000da之间。在一些实施例中,聚合物主链各链的分子量介于约10,000da与约40,000da之间。
所属领域技术人员将认识到,实质上水溶性主链的上述清单一点也不详尽且仅为说明性的,并且预期具有上述品质的所有聚合物材料都适用于本发明中。
在本发明一些实施例中,聚合物衍生物为“多官能的”,意思是聚合物主链的至少两个末端且可能多达约300个末端经官能团官能化或活化。多官能聚合物衍生物包括(但不限于)具有两个末端的线性聚合物,各末端与相同或不同官能团键接。
在一个实施例中,聚合物衍生物具有以下结构:
x—a—poly—b—n=n=n
其中:
n=n=n为叠氮部分;
b为连接部分,其可能存在或不存在;
poly为水溶性非抗原性聚合物;
a为连接部分,其可能存在或不存在,并且可与b相同或不同;且
x为第二官能团。
连接部分a和b的实例包括(但不限于)含有至多18个且可含有介于1到10个之间的碳原子的多官能烷基。烷基链中可包括杂原子,例如氮、氧或硫。烷基链还可在杂原子处分支。连接部分a和b的其它实例包括(但不限于)含有至多10个且可含有5到6个碳原子的多官能芳基。芳基可经一个或一个以上碳原子、氮、氧或硫原子取代。适合连接基团的其它实例包括美国专利第5,932,462号、第5,643,575号以及美国专利申请公开案2003/0143596中所述的连接基团,所述专利各以引用的方式并入本文中。所属领域技术人员将认识到,连接部分的上述清单一点也不详尽且仅为说明性的,并且预期具有上述品质的所有连接部分都适用于本发明中。
适用作x的官能团的实例包括(但不限于)羟基、经保护羟基、烷氧基、活性酯(例如n-羟基琥珀酰亚胺酯和1-苯并三唑酯)、活性碳酸酯(例如碳酸n-羟基琥珀酰亚胺酯和碳酸1-苯并三唑酯)、缩醛、醛、水合醛、烯基、丙烯酸酯、甲基丙烯酸酯、丙烯酰胺、活性砜、胺、氨氧基、经保护胺、酰肼、经保护酰肼、经保护硫醇、羧酸、经保护羧酸、异氰酸酯、异硫代氰酸酯、马来酰亚胺、乙烯基砜、二硫代吡啶、乙烯基吡啶、碘代乙酰胺、环氧基、乙二醛、二酮、甲磺酸酯、甲苯磺酸酯、三氟甲磺酸酯、烯烃、酮和叠氮基。如所属领域技术人员所了解,所选x部分应能与叠氮基相容,因此不会与叠氮基发生反应。含叠氮聚合物衍生物可为同双官能性(homobifunctional)的,意指第二官能团(即,x)也为叠氮部分;或异双官能性的,意指第二官能团为不同官能团。
术语“经保护”是指存在防止化学反应性官能团在某些反应条件下反应的保护基或部分。保护基将视所保护的化学反应性基团的类型而变化。举例来说,如果化学反应性基团为胺或酰肼,那么保护基可选自叔丁氧羰基(t-boc)和9-芴基甲氧基羰基(fmoc)的群组。如果化学反应性基团为硫醇,那么保护基可为邻吡啶基二硫化物。如果化学反应性基团为羧酸(例如丁酸或丙酸)或羟基,那么保护基可为苯甲基或烷基(例如甲基、乙基或叔丁基)。此项技术中已知的其它保护基也可用于本发明中。
文献中末端官能团的具体实例包括(但不限于)碳酸n-琥珀酰亚胺基酯(例如参见,美国专利第5,281,698号、第5,468,478号)、胺(例如参见,布克曼(buckmann)等人,高分子化学(makromol.chem.),182:1379(1981);泽利浦斯基(zalipsky)等人,欧洲聚合物杂志(eur.polym.j.)19:1177(1983))、酰肼(例如参见,安德瑞兹(andresz)等人,高分子化学,179:301(1978))、丙酸琥珀酰亚胺酯和丁酸琥珀酰亚胺酯(例如参见,奥尔森(olson)等人,聚(乙二醇)的化学与生物应用(poly(ethyleneglycol)chemistry&biologicalapplications),第170-181页,哈里斯(harris)和泽利浦斯基编,acs出版社,华盛顿哥伦比亚特区(washington,d.c.),1997;还参见美国专利第5,672,662号)、琥珀酸琥珀酰亚胺酯(例如参见,阿布托斯奇(abuchowski)等人,癌症生物化学与生物物理学(cancerbiochem.biophys.)7:175(1984),以及卓普吉(joppich)等人,高分子化学,180:1381(1979))、琥珀酰亚胺酯(例如参见,美国专利第4,670,417号)、苯并三唑碳酸酯(例如参见,美国专利第5,650,234号)、缩水甘油醚(例如参见,皮萨(pitha)等人,欧洲生物化学杂志(eur.jbiochem.),94:11(1979);艾琳(elling)等人,应用生物化学与生物技术(biotech.appl.biochem.)13:354(1991))、氧基羰基咪唑(例如参见,比奥查普(beauchamp)等人,分析生物化学(anal.biochem.),131:25(1983);托德力(tondelli)等人,控释杂志(j.controlledrelease)1:251(1985))、碳酸对硝基苯酯(例如参见,沃尼斯(veronese)等人,应用生物化学与生物技术,11:141(1985);以及萨托(sartore)等人,应用生物化学与生物技术,27:45(1991))、醛(例如参见,哈里斯(harris)等人,聚合物科学杂志第a辑:聚合物化学版(j.polym.sci.chem.ed.)22:341(1984);美国专利第5,824,784号、美国专利第5,252,714号)、马来酰亚胺(例如参见,古德森(goodson)等人,生物技术(纽约)(biotechnology)(ny)8:343(1990);罗曼尼(romani)等人,肽和蛋白质化学(chemistryofpeptidesandproteins)2:29(1984)),以及克甘(kogan),合成通讯(syntheticcomm.)22:2417(1992))、原吡啶基二硫化物(例如参见,沃赫伦(woghiren)等人,生物结合物化学(bioconj.chem.)4:314(1993))、丙烯醇(例如参见,萨赫雷(sawhney)等人,高分子(macromolecules),26:581(1993))、乙烯基亚砜(例如参见,美国专利第5,900,461号)。上述所有参考文献和专利都以引用的方式并入本文中。
在本发明某些实施例中,本发明的聚合物衍生物包含具有以下结构的聚合物主链:
x—ch2ch2o--(ch2ch2o)n--ch2ch2-n=n=n
其中:
x为如上文所述的官能团;且
n为约20到约4000。
在另一实施例中,本发明的聚合物衍生物包含具有以下结构的聚合物主链:
x—ch2ch2o--(ch2ch2o)n--ch2ch2-o-(ch2)m-w-n=n=n
其中:
w为包含1到10个碳原子的脂肪族或芳香族连接部分;
n为约20到约4000;且
x为如上文所述的官能团;m介于1与10之间。
可通过此项技术中已知和/或本文中揭示的多种方法制备本发明的含叠氮基peg衍生物。在下文所示的一种方法中,平均分子量为约800da到约100,000da的水溶性聚合物主链(所述聚合物主链的第一末端与第一官能团键接且第二末端与适当离去基团键接)与叠氮阴离子(其可与多种适合的反离子中的任一种配对,包括钠、钾、叔丁基铵等)反应。离去基团经历亲核置换且经叠氮部分置换,从而提供所需含叠氮基的peg聚合物。
x-peg-l+n3-→x-peg-n3
如所示,适用于本发明中的聚合物主链具有式x-peg-l,其中peg为聚(乙二醇),且x为不与叠氮基反应的官能团,且l为适当的离去基团。适合的官能团的实例包括(但不限于)羟基、经保护羟基、缩醛、烯基、胺、氨氧基、经保护胺、经保护酰肼、经保护硫醇、羧酸、经保护羧酸、马来酰亚胺、二硫代吡啶和乙烯基吡啶,以及酮。适合离去基团的实例包括(但不限于)氯基、溴基、碘基、甲磺酸酯基、三氟甲磺酸酯基和甲苯磺酸酯基。
在另一种制备本发明的含叠氮基聚合物衍生物的方法中,使带有叠氮官能团的连接剂与平均分子量为约800da到约100,000da的水溶性聚合物主链接触,其中所述连接剂带有会与peg聚合物上的化学官能团选择性反应形成含叠氮基聚合物衍生物产物的化学官能团,其中所述叠氮基经连接基团与聚合物主链分开。
示范性反应方案显示如下:
x-peg-m+n-连接基团-n=n=n→pg-x-peg-连接基团-n=n=n
其中:
peg为聚(乙二醇),且x为封端基团,例如烷氧基或如上文所述的官能团;且
m为不与叠氮官能团反应但与n官能团有效选择性反应的官能团。
适合官能团的实例包括(但不限于):如果n为胺,那么m为羧酸、碳酸酯或活性酯;如果n为酰肼或氨氧基部分,那么m为酮;如果n为亲核基团,那么m为离去基团。
可利用已知方法纯化粗产物,所述方法包括(但不限于)沉淀产物,随后视需要进行色谱分离。
更具体的实例显示于下文中peg二胺的情况下,其中一个胺经例如叔丁基-boc等保护基保护,且所得单保护peg二胺与带有叠氮官能团的连接部分反应:
bochn-peg-nh2+ho2c-(ch2)3-n=n=n
在此情况下,可使用多种活化剂,例如亚硫酰氯或碳化二亚胺试剂和n-羟基琥珀酰亚胺或n-羟基苯并三唑,将胺基与羧酸基偶合,从而在单胺peg衍生物与带有叠氮基的连接部分之间产生酰胺键。在成功形成酰胺键后,可直接使用所得含n-叔丁基-boc保护的叠氮基的衍生物修饰生物活性分子,或者可进一步精细设计所述衍生物以安装其它有用官能团。举例来说,可通过用强酸处理来水解n-t-boc基团,以产生ω-氨基-peg-叠氮化物。所得胺可用作合成处理剂(handle)以安装其它有用官能团,例如马来酰亚胺基、活性二硫化物、活性酯等,以便产生有价值的异双官能试剂。
当需要将不同分子与聚合物各末端连接时,异双官能衍生物特别有用。举例来说,ω-n-氨基-n-叠氮基peg允许将具有活性亲电基团的分子(例如醛、酮、活性酯、活性碳酸酯等)与peg的一个末端连接,并将具有乙炔基的分子与peg的另一末端连接。
在本发明另一实施例中,聚合物衍生物具有以下结构:
x—a—poly—b—c≡c-r
其中:
r可为h或烷基、烯烃、烷氧基,或者芳基或经取代芳基;
b为连接部分,其可能存在或不存在;
poly为水溶性非抗原性聚合物;
a为连接部分,其可能存在或不存在,并且可与b相同或不同;且
x为第二官能团。
连接部分a和b的实例包括(但不限于)含有至多18个且可含有介于1到10个之间的碳原子的多官能烷基。烷基链中可包括杂原子,例如氮、氧或硫。烷基链还可在杂原子处分支。连接部分a和b的其它实例包括(但不限于)含有至多10个且可含有5到6个碳原子的多官能芳基。芳基可经一个或一个以上碳原子、氮、氧或硫原子取代。适合的连接基团的其它实例包括美国专利第5,932,462号和第5,643,575号以及美国专利申请公开案2003/0143596中所述的连接基团,所述专利各以引用的方式并入本文中。所属领域技术人员将认识到,连接部分的上述清单一点也不详尽且仅打算为说明性的,并且预期具有上述品质的多种连接部分都适用于本发明中。
适用作x的官能团的实例包括:羟基、经保护羟基、烷氧基、活性酯(例如n-羟基琥珀酰亚胺酯和1-苯并三唑酯)、活性碳酸酯(例如碳酸n-羟基琥珀酰亚胺酯和碳酸1-苯并三唑酯)、缩醛、醛、水合醛、烯基、丙烯酸酯、甲基丙烯酸酯、丙烯酰胺、活性砜、胺、氨氧基、经保护胺、酰肼、经保护酰肼、经保护硫醇、羧酸、经保护羧酸、异氰酸酯、异硫代氰酸酯、马来酰亚胺、乙烯基砜、二硫代吡啶、乙烯基吡啶、碘代乙酰胺、环氧化物、乙二醛、二酮、甲磺酸酯、甲苯磺酸酯和三氟甲磺酸酯、烯烃、酮和乙炔。如应了解的,所选x部分应能与乙炔基相容,以致不会与乙炔基发生反应。含乙炔的聚合物衍生物可为同双官能性的,意指第二官能团(即,x)也为乙炔部分;或异双官能性的,意指第二官能团为不同官能团。
在本发明另一实施例中,聚合物衍生物包含具有以下结构的聚合物主链:
x—ch2ch2o--(ch2ch2o)n--ch2ch2-o-(ch2)m-c≡ch
其中:
x为如上文所述的官能团;
n为约20到约4000;且
m介于1与10之间。
各异双官能peg聚合物的具体实例显示于下文中。
可使用所属领域技术人员已知和/或本文中揭示的方法制备本发明的含乙炔peg衍生物。在一种方法中,平均分子量为约800da到约100,000da的水溶性聚合物主链(所述聚合物主链的第一末端与第一官能团键接且第二末端与适当亲核基团键接)与带有乙炔官能团和适于与peg上的亲核基团反应的离去基团的化合物反应。当带有亲核部分的peg聚合物与带有离去基团的分子组合时,所述离去基团经历亲核置换且经亲核部分置换,从而得到所需的含乙炔聚合物。
x-peg-nu+l-a-c→x-peg-nu-a-c≡cr'
如所示,用于所述反应中的优选聚合物主链具有式x-peg-nu,其中peg为聚(乙二醇),nu为亲核部分,且x为不与nu、l或乙炔官能团反应的官能团。
nu的实例包括(但不限于)主要经由sn2型机制反应的胺、烷氧基、芳氧基、巯基、亚氨基、羧酸酯基、酰肼基、氨氧基。nu基团的其它实例包括主要经由亲核加成反应来反应的官能团。l基团的实例包括氯基、溴基、碘基、甲磺酸酯基、三氟甲磺酸酯基和甲苯磺酸酯基,和预期会经历亲核置换的其它基团,以及酮、醛、硫酯、烯烃、α-β不饱和羰基、碳酸酯基,和预期会经历亲核试剂加成的其它亲电基团。
在本发明另一实施例中,a为具有1到10个碳原子的脂肪族连接基团或具有6到14个碳原子的经取代芳基环。x为不与叠氮基反应的官能团,且l为适当的离去基团。
在另一种制备本发明的含乙炔聚合物衍生物的方法中,使平均分子量为约800da到约100,000da且在一末端带有经保护官能团或封端剂以及在另一末端带有适当离去基团的peg聚合物与乙炔阴离子接触。
示范性反应方案显示如下:
x-peg-l+-c≡cr'→x-peg-c≡cr'
其中:
peg为聚(乙二醇),且x为封端基团,例如烷氧基或如上文所述的官能团;且
r'为h、烷基、烷氧基、芳基或芳氧基或者经取代烷基、烷氧基、芳基或芳氧基。
在上述实例中,离去基团l应具有足够的反应性以在与足够浓度的乙炔阴离子接触时发生sn2型置换。所属领域技术人员已知实现乙炔阴离子对离去基团的sn2置换所需的反应条件。
通常可借助于此项技术中已知的方法纯化粗产物,所述方法包括(但不限于)沉淀产物,随后视需要进行色谱分离。
可将水溶性聚合物与本发明的bst多肽连接。水溶性聚合物可经由并入bst多肽中的非天然编码氨基酸,或非天然编码或天然编码氨基酸的任何官能团或取代基,或添加到非天然编码或天然编码氨基酸中的任何官能团或取代基连接。或者,水溶性聚合物经由天然存在氨基酸(包括(但不限于)半胱氨酸、赖氨酸,或n末端残基的胺基)与并有非天然编码氨基酸的bst多肽连接。在一些情况下,本发明的bst多肽包含1、2、3、4、5、6、7、8、9、10个非天然氨基酸,其中一个或一个以上非天然编码氨基酸与水溶性聚合物(包括(但不限于)peg和/或寡糖)连接。在一些情况下,本发明的bst多肽另外包含1、2、3、4、5、6、7、8、9、10或10个以上与水溶性聚合物连接的天然编码氨基酸。在一些情况下,本发明的bst多肽包含一个或一个以上与水溶性聚合物连接的非天然编码氨基酸和一个或一个以上与水溶性聚合物连接的天然存在氨基酸。在一些实施例中,相对于未结合形式,本发明中使用的水溶性聚合物将增加bst多肽的血清半衰期。
可调节与本发明的bst多肽连接的水溶性聚合物的数量(即,聚乙二醇化或糖基化程度),从而提供改变(包括(但不限于)增加或降低)的药理学、药物动力学或药效学特征,例如体内半衰期。在一些实施例中,bst的半衰期增加超过未经修饰多肽的至少约10%、20%、30%、40%、50%、60%、70%、80%、90%、2倍、5倍、6倍、7倍、8倍、9倍、10倍、11倍、12倍、13倍、14倍、15倍、16倍、17倍、18倍、19倍、20倍、25倍、30倍、35倍、40倍、50倍或至少约100倍。
含有强亲核基团(即,酰肼、肼、羟胺或氨基脲)的peg衍生物
在本发明一个实施例中,利用含有直接与peg主链连接的末端肼、羟胺、酰肼或氨基脲部分的peg衍生物修饰包含含羰基非天然编码氨基酸的bst多肽。
在一些实施例中,以羟胺为末端的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-onh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000(即,平均分子量介于5到40kda之间)。
在一些实施例中,含肼或含酰肼的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-x-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000,并且x任选为存在或不存在的羰基(c=o)。
在一些实施例中,含氨基脲的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-nh-c(o)-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000。
在本发明另一实施例中,利用含有借助酰胺键与peg主链连接的末端羟胺、酰肼、肼或氨基脲部分的peg衍生物修饰包含含羰基氨基酸的bst多肽。
在一些实施例中,以羟胺为末端的peg衍生物具有以下结构:
ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)(ch2)m-o-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000(即,平均分子量介于5到40kda之间)。
在一些实施例中,含肼或含酰肼的peg衍生物具有以下结构:
ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)(ch2)m-x-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,n为100到1,000,并且x任选为存在或不存在的羰基(c=o)。
在一些实施例中,含氨基脲的peg衍生物具有以下结构:
ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)(ch2)m-nh-c(o)-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000。
在本发明另一实施例中,利用含有末端肼、羟胺、酰肼或氨基脲部分的分支peg衍生物修饰包含含羰基氨基酸的bst多肽,其中所述分支peg各链的mw在10到40kda的范围内且可为5到20kda。
在本发明另一实施例中,利用具有分支结构的peg衍生物修饰包含非天然编码氨基酸的bst多肽。举例来说,在一些实施例中,以肼或以酰肼为末端的peg衍生物将具有以下结构:
[ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)]2ch(ch2)m-x-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),m为2到10且n为100到1,000,并且x任选为存在或不存在的羰基(c=o)。
在一些实施例中,含有氨基脲基的peg衍生物将具有以下结构:
[ro-(ch2ch2o)n-o-(ch2)2-c(o)-nh-ch2-ch2]2ch-x-(ch2)m-nh-c(o)-nh-nh2
其中r为简单烷基(甲基、乙基、丙基等),x任选为nh、o、s、c(o),或不存在,m为2到10且n为100到1,000。
在一些实施例中,含有羟胺基的peg衍生物将具有以下结构:
[ro-(ch2ch2o)n-o-(ch2)2-c(o)-nh-ch2-ch2]2ch-x-(ch2)m-o-nh2
其中r为简单烷基(甲基、乙基、丙基等),x任选为nh、o、s、c(o),或不存在,m为2到10且n为100到1,000。
水溶性聚合物与bst多肽连接的程度和位点可调节bst多肽与受体的结合。在一些实施例中,键的布置应使得通过平衡结合分析所测量(例如斯派瑟(spencer)等人,生物化学杂志(j.biol.chem.),263:7862-7867(1988)中所述的分析),bst多肽与受体结合的kd为约400nm或更低、150nm或更低,且在一些情况下kd为100nm或更低。
活化聚合物以及结合肽的方法和化学研究描述于文献中且为此项技术中所知。活化聚合物的常用方法包括(但不限于)用溴化氰、高碘酸盐、戊二醛、双环氧化物、表氯醇、二乙烯基砜、碳化二亚胺、磺酰基卤化物、三氯三嗪等活化官能团(参见r.f.泰勒(r.f.taylor),(1991),蛋白质固定、基本原则和应用(proteinimmobilisation.fundamentalandapplications),马克戴克出版公司(marceldekker),纽约(n.y.);s.s.王(s.s.wong),(1992),蛋白质结合和交联化学研究(chemistryofproteinconjugationandcrosslinking),crc出版社,博卡拉顿(bocaraton);g.t.赫曼森(g.t.hermanson)等人,(1993),固定亲和配体技术(immobilizedaffinityligandtechniques),学术出版社(academicpress),纽约(n.y.);度恩(dunn,r.l.)等人编,聚合物药物与药物递送系统(polymericdrugsanddrugdeliverysystems),美国化学学会论文集(acssymposiumseries),第469卷,美国化学学会(americanchemicalsociety),华盛顿哥伦比亚特区(washington,d.c.),1991)。
可利用有关peg官能化和结合的数篇评论和专题文章。例如参见,哈里斯(harris),高分子化学与物理学(macromol.chem.phys.)c25:325-373(1985);司考顿(scouten),酶学方法(methodsinenzymology)135:30-65(1987);王(wong)等人,酶学与微生物技术(enzymemicrob.technol.)14:866-874(1992);戴尔盖多(delgado)等人,治疗性药物载体系统的评述(criticalreviewsintherapeuticdrugcarriersystems)9:249-304(1992);泽利浦斯基(zalipsky),生物结合物化学,6:150-165(1995)。
聚合物活化以及活性聚合物与酶之间结合的方法也可见于wo94/17039、美国专利第5,324,844号、wo94/18247、wo94/04193、美国专利第5,219,564号、美国专利第5,122,614号、wo90/13540、美国专利第5,281,698号和wo93/15189中,所述酶包括(但不限于)凝血因子viii(wo94/15625)、血红蛋白(wo94/09027)、载氧分子(美国专利第4,412,989号)、核糖酶和超氧化物歧化酶(沃伦瑟(veronese)等人,应用生物化学与生物技术(app.biochem.biotech.)11:141-52(1985))。引用的所有参考文献和专利都以引用的方式并入本文中。
含有非天然编码氨基酸(例如对叠氮基-l-苯丙氨酸)的bst多肽的聚乙二醇化(即,添加任何水溶性聚合物)是利用任何便利的方法进行。举例来说,利用经炔封端的mpeg衍生物使bst多肽聚乙二醇化。简单地说,室温下,在搅拌下将过量的固体mpeg(5000)-o-ch2-c≡ch添加到含对叠氮基-l-phe的bst多肽水溶液中。通常,用pka接近欲进行反应的ph值(通常为约ph4-10)的缓冲液对水溶液进行缓冲。适于ph7.5下进行聚乙二醇化的缓冲液的实例例如包括(但不限于)hepes、磷酸盐、硼酸盐、tris-hcl、epps和tes。持续监测ph值,并视需要加以调节。反应通常持续约1到48小时。
随后,使反应产物经历疏水相互作用色谱以使聚乙二醇化的bst多肽变异体与游离mpeg(5000)-o-ch2-c≡ch和聚乙二醇化bst多肽的任何高分子量复合物分离,所述高分子量复合物可在活化未封端peg的分子两端以使bst多肽变异体分子交联时形成。疏水相互作用色谱期间的条件应使游离mpeg(5000)-o-ch2-c≡ch流过柱,而任何交联的聚乙二醇化bst多肽变异体复合物在含有与一个或一个以上peg基团结合的一个bst多肽变异体分子的所需形式后洗脱。适当条件视交联复合物相对于所需结合物的相对尺寸而变化,并且易于由所属领域技术人员确定。利用超滤法浓缩含有所需结合物的洗脱液,并利用透滤法脱盐。
必要时,可借助于一种或一种以上所属领域技术人员已知的程序进一步纯化由疏水性色谱获得的聚乙二醇化bst多肽,所述程序包括(但不限于)亲和色谱、阴离子或阳离子交换色谱(使用(包括(但不限于)deaesepharose)、二氧化硅色谱、反相hplc、凝胶过滤(使用(包括(但不限于)sephadexg-75)、疏水相互作用色谱、尺寸排阻色谱、金属螯合色谱、超滤/透滤、乙醇沉淀、硫酸铵沉淀、色谱聚焦、置换色谱、电泳程序(包括(但不限于)制备型等电聚焦)、差示溶解度(包括(但不限于)硫酸铵沉淀)或萃取。可通过gpc经由与球蛋白标准品比较来估算表观分子量(普伦塔az(prenetaaz),蛋白质纯化法:一种实用方法(proteinpurificationmethods,apracticalapproach)(哈里斯(harris)与安吉拉(angal)编)irl出版社,1989,293-306)。可通过蛋白水解降解(包括(但不限于)胰蛋白酶裂解)、随后质谱分析来评估bst-peg结合物的纯度。派普斯基rb(pepinskyrb).等人,药理学与实验治疗学杂志(j.pharmcol.&exp.ther.)297(3):1059-66(2001)。
可在不受限制的情况下进一步衍生化或取代与本发明bst多肽的氨基酸连接的水溶性聚合物。
含叠氮基peg衍生物
在本发明另一实施例中,用含有会与非天然编码氨基酸侧链上存在的炔部分反应的叠氮部分的peg衍生物修饰bst多肽。一般来说,peg衍生物的平均分子量在1到100kda范围内,且在一些实施例中在10到40kda范围内。
在一些实施例中,以叠氮基为末端的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-n3
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000(即,平均分子量介于5到40kda之间)。
在另一实施例中,以叠氮基为末端的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-nh-c(o)-(ch2)p-n3
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,p为2到10且n为100到1,000(即,平均分子量介于5到40kda之间)。
在本发明另一实施例中,利用含有末端叠氮部分的分支peg衍生物修饰包含含炔氨基酸的bst多肽,其中所述分支peg各链的mw在10到40kda的范围内且可为5到20kda。举例来说,在一些实施例中,以叠氮基为末端peg衍生物将具有以下结构:
[ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)]2ch(ch2)m-x-(ch2)pn3
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,p为2到10且n为100到1,000,并且在各情况下x任选为可存在或不存在的o、n、s或羰基(c=o)。
含炔peg衍生物
在本发明另一实施例中,用含有会与非天然编码氨基酸侧链上存在的叠氮部分反应的炔部分的peg衍生物修饰bst多肽。
在一些实施例中,以炔为末端的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-c≡ch
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,且n为100到1,000(即,平均分子量介于5到40kda之间)。
在本发明另一实施例中,利用含有借助酰胺键与peg主链连接的末端叠氮或末端炔部分的peg衍生物修饰包含含炔非天然编码氨基酸的bst多肽。
在一些实施例中,以炔为末端的peg衍生物将具有以下结构:
ro-(ch2ch2o)n-o-(ch2)m-nh-c(o)-(ch2)p-c≡ch
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,p为2到10且n为100到1,000。
在本发明另一实施例中,利用含有末端炔部分的分支peg衍生物修饰包含含叠氮基氨基酸的bst多肽,其中所述分支peg各链的mw在10到40kda的范围内且可为5到20kda。举例来说,在一些实施例中,以炔为末端peg衍生物将具有以下结构:
[ro-(ch2ch2o)n-o-(ch2)2-nh-c(o)]2ch(ch2)m-x-(ch2)pc≡ch
其中r为简单烷基(甲基、乙基、丙基等),m为2到10,p为2到10且n为100到1,000,并且x任选为o、n、s或羰基(c=o),或不存在。
含膦peg衍生物
在本发明另一实施例中,用含有活性官能团(包括(但不限于)酯、碳酸酯)且另外包含会与非天然编码氨基酸侧链上存在的叠氮部分反应的芳基膦基的peg衍生物修饰bst多肽。一般来说,peg衍生物的平均分子量在1到100kda范围内,且在一些实施例中在10到40kda范围内。
在一些实施例中,peg衍生物将具有以下结构:
其中n为1到10;x可为o、n、s,或不存在,ph为苯基,且w为水溶性聚合物。
在一些实施例中,peg衍生物将具有以下结构:
其中x可为o、n、s,或不存在,ph为苯基,w为水溶性聚合物,且r可为h、烷基、芳基、经取代烷基和经取代芳基。示范性r基包括(但不限于)-ch2、-c(ch3)3、-or'、-nr'r"、-sr'、-卤素、-c(o)r'、-conr'r"、-s(o)2r'、-s(o)2nr'r"、-cn和-no2。r'、r"、r'"和r""各独立地指氢、经取代或未经取代的杂烷基、经取代或未经取代的芳基(包括(但不限于)经1到3个卤素取代的芳基)、经取代或未经取代的烷基、烷氧基或硫烷氧基,或芳基烷基。当本发明化合物包括一个以上r基团时,例如,各r基团是如各r'、r"、r'"和r""基团当存在超过一个时那样独立地选择。当r'与r"连接到同一氮原子时,其可与所述氮原子组合形成5元、6元或7元环。举例来说,-nr'r"意欲包括(但不限于)1-吡咯烷基和4-吗啉基。所属领域技术人员从上文有关取代基的论述将了解到,术语“烷基”意欲包括碳原子与除氢基外的基团结合的基团,例如卤烷基(包括(但不限于)-cf3和-ch2cf3)和酰基(包括(但不限于)-c(o)ch3、-c(o)cf3、-c(o)ch2och3等)。
其它peg衍生物和通用聚乙二醇化技术
可与bst多肽连接的其它示范性peg分子以及聚乙二醇化方法包括(但不限于)例如以下专利中所述者:美国专利公开案第2004/0001838号、第2002/0052009号、第2003/0162949号、第2004/0013637号、第2003/0228274号、第2003/0220447号、第2003/0158333号、第2003/0143596号、第2003/0114647号、第2003/0105275号、第2003/0105224号、第2003/0023023号、第2002/0156047号、第2002/0099133号、第2002/0086939号、第2002/0082345号、第2002/0072573号、第2002/0052430号、第2002/0040076号、第2002/0037949号、第2002/0002250号、第2001/0056171号、第2001/0044526号、第2001/0021763号;美国专利第6,646,110号、第5,824,778号、第5,476,653号、第5,219,564号、第5,629,384号、第5,736,625号、第4,902,502号、第5,281,698号、第5,122,614号、第5,473,034号、第5,516,673号、第5,382,657号、第6,552,167号、第6,610,281号、第6,515,100号、第6,461,603号、第6,436,386号、第6,214,966号、第5,990,237号、第5,900,461号、第5,739,208号、第5,672,662号、第5,446,090号、第5,808,096号、第5,612,460号、第5,324,844号、第5,252,714号、第6,420,339号、第6,201,072号、第6,451,346号、第6,306,821号、第5,559,213号、第5,747,646号、第5,834,594号、第5,849,860号、第5,980,948号、第6,004,573号、第6,129,912号;wo97/32607、ep229,108、ep402,378、wo92/16555、wo94/04193、wo94/14758、wo94/17039、wo94/18247、wo94/28024、wo95/00162、wo95/11924、wo95/13090、wo95/33490、wo96/00080、wo97/18832、wo98/41562、wo98/48837、wo99/32134、wo99/32139、wo99/32140、wo96/40791、wo98/32466、wo95/06058、ep439508、wo97/03106、wo96/21469、wo95/13312、ep921131、wo98/05363、ep809996、wo96/41813、wo96/07670、ep605963、ep510356、ep400472、ep183503和ep154316,所述专利都以引用的方式并入本文中。本文中所述的任何peg分子都可以任何形式使用,包括(但不限于)单链、支链、多臂链、单官能、双官能、多官能或其任何组合。
其它聚合物和peg衍生物,包括(但不限于)羟胺(氨氧基)peg衍生物,描述于以下专利申请案中(全文以引用的方式并入本文中):美国专利公开案第2006/0194256号、美国专利公开案第2006/0217532号、美国专利公开案第2006/0217289号、美国临时专利第60/755,338号、美国临时专利第60/755,711号、美国临时专利第60/755,018号、国际专利申请案第pct/us06/49397号、wo2006/069246、美国临时专利第60/743,041号、美国临时专利第60/743,040号、国际专利申请案第pct/us06/47822号、美国临时专利第60/882,819号、美国临时专利第60/882,500号,和美国临时专利第60/870,594号。
异源fc融合蛋白
上述bst化合物可直接或经由肽连接子与免疫球蛋白的fc部分融合。免疫球蛋白是含有利用二硫键保持在一起的多肽链的分子,通常具有两条轻链和两条重链。在各链中,视分子的抗体特异性而定,一个域(v)具有可变氨基酸序列。其它域(c)具有同类分子共有的相当恒定的序列。
本文中使用的免疫球蛋白fc部分的含义与该术语在免疫学领域中的含义相同。具体点说,此术语是指通过去除抗体中的两个抗原结合区(fab片段)得到的抗体片段。一种去除fab片段的方式是用木瓜蛋白酶消化免疫球蛋白。因此,fc部分是由来自两条重链的恒定区的尺寸大致相等的片段形成,这两个片段经由非共价相互作用和二硫键缔合。fc部分可包括铰链区,并延伸通过ch2和ch3域,到达抗体c末端。人类和小鼠免疫球蛋白的示范性铰链区可见于抗体工程实践指南(antibodyengineering,apracticalguide),博瑞贝克c.a.k(borrebaeck,c.a.k)编,w.h.弗雷曼出版社(w.h.freemanandco.),1992,其教示以引用的方式并入本文中。fc部分可进一步包括一个或一个以上糖基化位点。此项技术中众所周知含有铰链区、ch2和ch3域以及一个n-糖基化位点的多种代表性fc蛋白质的氨基酸序列。
存在5种类型的具有不同效应功能和药物动力学特性的人免疫球蛋白fc区:igg、iga、igm、igd和ige。igg是血清中含量最丰富的免疫球蛋白。igg在血清任何免疫球蛋白中半衰期也最长(23天)。与其它免疫球蛋白不同,igg在结合于fc受体后会有效再循环。igg具有4个亚类:g1、g2、g3和g4,每一类具有不同的效应功能。g1、g2和g3可结合c1q并固定补体,而g4不能。尽管与g1相比,g3能够更有效地结合c1q,但在介导补体引导的细胞溶解方面,g1更有效。g2固定补体的效率极低。igg中的c1q结合位点位于ch2域的羧基末端区。
所有igg亚类都能够结合于fc受体(cd16、cd32、cd64),其中g1和g3的效率高于g2和g4。igg的fc受体结合区是由位于ch2域的铰链区和羧基末端区中的残基形成。
iga可以单体形式以及借助于j链保持在一起的二聚体形式存在。iga是血清中含量第二丰富的免疫球蛋白,但其半衰期只有6天。iga具有3种效应功能。其结合于巨噬细胞和嗜曙红细胞上的iga特异性受体,从而分别驱动胞噬作用和细胞脱粒。其也可经由未知的替代性路径固定补体。
igm是表达为五聚体或六聚体,二者都经由j链保持在一起。igm的血清半衰期为5天。其经由位于其ch3域中的结合位点较弱地结合于c1q。igd的血清半衰期为3天。何种效应功能是由此种免疫球蛋白引起尚不明了。ige是一种单体免疫球蛋白,且血清半衰期为2.5天。ige结合于两个fc受体,以驱动细胞脱粒,并引起促发炎因子的释放。
视所需体内效应而定,本发明的异源融合蛋白可含有上述任一同种型,或者可含有突变的fc区,其中补体和/或fc受体结合功能已经改变。因此,本发明的异源融合蛋白可含有与bst化合物融合的免疫球蛋白的完整fc部分、免疫球蛋白fc部分的片段,或其类似物。
本发明的融合蛋白可由单链蛋白组成或呈多链多肽的形式。可以产生两个或两个以上fc融合蛋白,以致其经由fc区之间天然形成的二硫键相互作用。这些多聚体可关于bst化合物为同源的,或者其可含有不同bst化合物融合于融合蛋白fc部分的n末端。
不管融合蛋白的最终结构如何,fc区或类fc区可用于延长融合于n末端的bst化合物的体内血浆半衰期。另外,融合蛋白化合物的bst组分应保持bst的至少一种生物活性。使用本文中所述或此项技术中已知的方法可以证实治疗或循环半衰期的增加,其中将融合蛋白的半衰期与单独bst化合物的半衰期相比较。生物活性可以借助于此项技术中已知的体外和体内方法测定。
由于由蛋白质水解产生的igg的fc区的体内半衰期与完整igg分子相同,且fab片段迅速降解,以致相信用于延长半衰期的相关序列存在于ch2和/或ch3域中。此外,文献中已经显示,不结合高亲和力fc受体或c1q的igg变异体的代谢速率与亲代野生型抗体的清除速率难以区别,表明代谢位点不同于fc受体或c1q结合所涉及的位点。[瓦泽纳克(wawrzynczak)等人,(1992)分子免疫学(molecularimmunology)29:221]。使用鼠类igg1fc区进行的定点诱变研究表明,igg1fc区中控制代谢速率的位点位于ch2-ch3域界面处。可以修饰fc区的代谢位点,以优化融合蛋白的半衰期。用于本发明融合蛋白的fc区可来源于igg1或igg4fc区,并且可含有ch2和ch3区,包括铰链区。
异源白蛋白融合蛋白
本文中描述的bst可直接或经由肽连接子、水溶性聚合物或前药连接子与白蛋白或其类似物、片段或衍生物融合。一般说来,作为本发明融合蛋白的一部分的白蛋白可来源于从包括人类在内的任何物种克隆得到的白蛋白。人血清白蛋白(humanserumalbumin,hsa)由具有585个氨基酸的单一未糖基化多肽链组成,所述多肽链的化学式分子量为66,500。已知人hsa的氨基酸序列[参见米洛恩(meloun)等人,(1975)欧洲生物学化学会联盟通讯(febsletters)58:136;比伦斯(behrens)等人,(1975)联盟论文集(fed.proc.)34:591;劳恩(lawn)等人(1981)核酸研究,9:6102-6114;米赫提(minghetti)等人(1986)生物化学杂志(j.biol.chem.)261:6747,各自以引用的方式并入本文中]。已经描述白蛋白的多种多晶型变异体以及类似物和片段。[参见维特坎普(weitkamp)等人(1973)人类遗传学记事(ann,hum.genet.)37:219]。举例来说,在ep322,094中,描述各种较短形式的hsa。揭示这些hsa片段中的一部分,包括hsa(1-373)、hsa(1-388)、hsa(1-389)、hsa(1-369)和hsa(1-419),以及在1-369与1-419之间的片段。ep399,666揭示的白蛋白片段包括hsa(1-177)和hsa(1-200)以及在hsa(1-177)与hsa(1-200)之间的片段。
应了解,本发明的异源融合蛋白包括与任何白蛋白(包括片段、类似物和衍生物)偶合的bst化合物,其中所述融合蛋白具有生物活性,并且血浆半衰期比单独bst化合物长。因此,融合蛋白中的白蛋白部分的血浆半衰期不必与天然人白蛋白相当。片段、类似物和衍生物为已知的,或者可以产生具有较长半衰期或半衰期在天然人白蛋白与相关bst化合物之间的片段、类似物和衍生物。
本发明的异源融合蛋白涵盖在融合蛋白的bst化合物和/或fc或白蛋白部分中具有保守氨基酸取代的蛋白质。“保守取代”是利用另一具有相同净电荷以及大致相同的尺寸和形状的氨基酸代替某一氨基酸。当侧链中碳和杂原子总数的差异不超过约4个时,具有脂肪族或经取代脂肪族氨基酸侧链的氨基酸具有大致相同的尺寸。当侧链中分支的数量差异不超过1个时,其具有大致相同的形状。侧链中具有苯基或经取代苯基的氨基酸被认为具有几乎相同的尺寸和形状。除本文中另外明确提供者外,优选利用天然存在的氨基酸进行保守取代。
野生型白蛋白和免疫球蛋白可以从多种来源获得。举例来说,可以由以可检测的水平表达相关mrna的组织或细胞制得的cdna文库得到这些蛋白质。可以用使用公开的dna或相关特定蛋白质的蛋白质序列设计的探针筛选文库。举例来说,免疫球蛋白轻链或重链恒定区描述于以下文献中:亚当斯(adams)等人(1980)生物化学(biochemistry)19:2711-2719;高尔夫特(goughet)等人(1980)生物化学,19:2702-2710;多尔比(dolby)等人(1980)美国国家科学院院刊,77:6027-6031;莱斯(rice)等人(1982)美国国家科学院院刊,79:7862-7862;弗克纳(falkner)等人(1982)自然(nature)298:286-288;和莫里森(morrison)等人(1984)免疫学年评(ann.rev.immunol.)2:239-256。揭示白蛋白和dna序列的一些参考文献包括米洛恩(meloun)等人,(1975)欧洲生物学化学会联盟通讯(febsletters)58:136;比伦斯(behrens)等人,(1975)联盟论文集(fed.proc.)34:591;劳恩(lawn)等人(1981)核酸研究,9:6102-6114;和米赫提(minghetti)等人(1986)生物化学杂志(j.biol.chem.)261:6747。
本发明异源融合蛋白的表征
多种方法可用于表征本发明的融合蛋白。这些方法中有一些包括(但不限于):sds-page联合蛋白质染色方法或使用抗igg或抗has抗体的免疫印迹法。其它方法包括例如基质辅助激光解吸附/离子化质谱(matrixassistedlaserdesorption/ionization-massspectrometry,maldi-ms)、液相色谱/质谱、等电聚焦、分析型阴离子交换、色谱聚焦和圆二色谱。
增强对血清白蛋白的亲和力
也可以将各种分子与本发明的bst多肽融合以调节bst多肽的血清半衰期。在一些实施例中,将分子与本发明的bst多肽连接或融合以增强对动物体内内源血清白蛋白的亲和力。
举例来说,在一些情况下,制备bst多肽与白蛋白结合序列的重组融合体。示范性白蛋白结合序列包括(但不限于)来自链球菌蛋白g的白蛋白结合域(例如参见马克雷德(makrides)等人,药理学与实验治疗学杂志(j.pharmacol.exp.ther)277:534-542(1996),以及索兰德尔(sjolander)等人,免疫学方法杂志(j,immunol.methods)201:115-123(1997)),或白蛋白结合肽,例如丹尼斯(dennis)等人,生物化学杂志(j.biol.chem.)277:35035-35043(2002)中所述的肽。
在其它实施例中,利用脂肪酸使本发明的bst多肽酰基化。在一些情况下,脂肪酸促进与血清白蛋白的结合。例如参见卡兹霍尔(kurtzhals)等人,生物化学杂志312:725-731(1995)。
在其它实施例中,将本发明的bst多肽与血清白蛋白(包括(但不限于)人血清白蛋白)直接融合。所属领域技术人员将认识到,也可将多种其它分子与本发明中的bst连接,以调节与血清白蛋白或其它血清组分的结合。
x.bst多肽的糖基化
本发明包括并有一个或一个以上带有糖残基的非天然编码氨基酸的bst多肽。糖残基可为天然(包括(但不限于)n-乙酰基氨基葡糖)或非天然(包括(但不限于)3-氟半乳糖)的。可通过n或o连接糖苷键(包括(但不限于)n-乙酰基半乳糖-l-丝氨酸)或非天然键(包括(但不限于)肟或者相应的c或s连接糖苷)将糖与非天然编码氨基酸连接。
可将糖(包括(但不限于)糖基)部分在体内或体外添加到bst多肽中。在本发明一些实施例中,用经氨氧基衍生化的糖修饰包含含羰基非天然编码氨基酸的bst多肽,以产生经由肟键连接的相应糖基化多肽。一旦与非天然编码氨基酸连接,即可利用糖基转移酶和其它酶处理来进一步精心制作糖,从而产生与bst结合的寡糖。例如参见h.刘(h.liu)等人,美国化学协会杂志,125:1702-1703(2003)。
在本发明一些实施例中,利用制备成氨氧基衍生物且具有指定结构的聚糖直接修饰包含含羰基非天然编码氨基酸的bst多肽。所属领域技术人员将认识到,可使用其它官能团,包括叠氮基、炔、酰肼、肼和氨基脲,将糖与非天然编码氨基酸连接。
在本发明一些实施例中,随后可通过包括(但不限于)分别与(包括(但不限于))炔基或叠氮基衍生物发生胡伊斯根[3+2]环加成反应,来修饰包含含叠氮基或含炔基非天然编码氨基酸的bst多肽。这种方法允许以极高选择性修饰蛋白质。
xi.bst二聚体和多聚体
本发明也提供bst和bst类似物组合,例如均二聚体、杂二聚体、均多聚体或杂多聚体(即,三聚体、四聚体等),其中含有一个或一个以上非天然编码氨基酸的bst与另一bst或其bst变异体,或者不为bst或其bst变异体的任何其它多肽结合,这种结合可以是与多肽主链直接结合或经由连接子结合。由于与单体相比,bst二聚体或多聚体结合物的分子量有所增加,故其可展现新颖或所需的特性,包括(但不限于)相对于单体bst不同的药理学、药物动力学、药效学;经调节的治疗半衰期;或经调节的血浆半衰期。在一些实施例中,本发明的bst二聚体将调节g-csf受体的信号转导。在其它实施例中,本发明的bst二聚体或多聚体将充当受体拮抗剂、激动剂或调节剂。
在一些实施例中,含有bst的二聚体或多聚体中存在的一个或一个以上bst分子包含非天然编码氨基酸与水溶性聚合物连接。
在一些实施例中,bst多肽是直接连接,包括(但不限于)经由asn-lys酰胺键或cys-cys二硫键连接。在一些实施例中,bst多肽和/或连接的非bst分子将包含不同的非天然编码氨基酸来促进二聚化,包括(但不限于)第一bst多肽的一个非天然编码氨基酸中的炔与第二分子的第二非天然编码氨基酸中的叠氮基将经由胡伊斯根[3+2]环加成结合。或者,可将包含含酮非天然编码氨基酸的bst和/或连接的非bst分子与包含含羟胺非天然编码氨基酸的第二多肽结合,且所述多肽经由形成相应肟来反应。
或者,两个bst多肽和/或连接的非bst分子经由连接子连接。可使用任何异双官能连接子或同双官能连接子连接两个分子,和/或连接的非bst分子,其可具有相同或不同的一级序列。在一些情况下,用于将bst和/或连接的非bst分子系在一起的连接子可为双官能peg试剂。连接子可具有多种分子量或分子长度。可以使用分子量较大或较小的连接子在bst与连接的实体之间或在bst与其受体之间,或者在连接的受体与其结合搭配物之间(如果存在的话)提供所需的空间关系或构象。也可以使用具有较大或较小分子长度的连接子在bst与连接的实体之间,或在连接的实体与其结合搭配物之间(如果存在的话)提供所需的空间或柔性。
在一些实施例中,本发明提供具有哑铃结构的水溶性双官能连接子,其包括:a)在聚合物主链至少第一端上的含叠氮、炔、肼、酰肼、羟胺或羰基的部分;和b)在聚合物主链第二端上的至少第二官能团。第二官能团与第一官能团可相同或不同。在一些实施例中,第二官能团不与第一官能团反应。在一些实施例中,本发明提供包含分支分子结构的至少一个臂的水溶性化合物。举例来说,分支分子结构可为树突状。
在一些实施例中,本发明提供包含一个或一个以上bst多肽的多聚体,其是由bst多肽与水溶性活性聚合物反应而形成,所述聚合物具有以下结构:
r-(ch2ch2o)n-o-(ch2)m-x
其中n为约5到约3,000,m为2到10,x可为含叠氮、炔、肼、酰肼、氨氧基、羟胺、乙酰基或羰基的部分,且r为可与x相同或不同的封端基、官能团或离去基团。r例如可为选自由以下各基团组成的群组的官能团:羟基、经保护羟基、烷氧基、n-羟基琥珀酰亚胺基酯、1-苯并三唑酯、n-羟基琥珀酰亚胺基碳酸酯、1-苯并三唑基碳酸酯、缩醛、醛、水合醛、烯基、丙烯酸酯、甲基丙烯酸酯、丙烯酰胺、活性砜、胺、氨氧基、经保护胺、酰肼、经保护酰肼、经保护硫醇、羧酸、经保护羧酸、异氰酸酯、异硫代氰酸酯、顺丁烯二酰亚胺、乙烯基砜、二硫代吡啶、乙烯基吡啶、碘代乙酰胺、环氧基、乙二醛、二酮、甲磺酸酯、甲苯磺酸酯和三氟甲磺酸酯、烯烃和酮。
xii.bst多肽活性和bst对受体的亲和力的测量
可使用标准或已知的体外或体内分析测定bst多肽的活性。可以借助于此项技术中已知的适合方法分析bst多肽的生物活性。所述分析包括(但不限于)赫达瑞(hedari)等人,兽医免疫和免疫病理(veterinaryimmunologyandimmunopathology)(2001)81:45-57中所述的分析和评估hg-csf生物活性的分析。
可以分析bst多肽上调中性粒细胞中cd11a、cd11b、cd11c和/或cd18的能力。可以利用赫达瑞(hedari)等人(同上文)所述的facs法测量这种活性。所属领域技术人员已知的其它分析也可测量中性粒细胞的活化,包括(但不限于)测量l-选择素(l-selectin)的分析。也可执行其它分析来评估本发明的bst多肽对细胞增殖和/或分化的影响。
可以分析bst多肽结合于受体的能力。可使用所属领域技术人员已知的技术和方法制备g-csf受体。hg-csf受体可如美国专利第5,574,136号(以引用的方式并入本文中)中所述制备。举例来说,可以使用对g-csf起反应而作用或结合g-csf的细胞或细胞系(包括(但不限于)含有活性g-csf受体的细胞,例如重组g-csf受体产生细胞)来监测bst受体结合。对于包含非天然氨基酸的未聚乙二醇化或聚乙二醇化bst多肽来说,可使用biacoretm生物传感器(法玛西亚公司(pharmacia))测量bst对其受体或对另一g-csf受体的亲和力。适合的结合分析包括(但不限于)biacore分析(皮尔斯(pearce)等人,生物技术(biochemistry)38:81-89(1999))和alphascreentm分析(珀金埃尔默公司(perkinelmer))。alphascreentm是一种基于珠粒的非放射性发光近距离分析(non-radioactiveluminescentproximityassay),其中供体珠粒在680nm激光激发下,释放出单线态氧。单线态氧扩散并与受者珠粒表面上的二甲基噻吩衍生物反应,从而在约600nm下发出荧光。只有在供者与受者珠粒各自分别与配体和受体连接时发生分子相互作用,从而使二者紧密靠近的情况下才会发射荧光。可以使用受体结合变异体竞争消除此配体-受体相互作用,而非结合变异体不会竞争。
可使用标准或已知的体外或体内分析测定bst多肽的活性。举例来说,可使用在hg-csf存在下增殖或结合hg-csf的细胞或细胞系(包括(但不限于)含有活性g-csf受体的细胞,例如小鼠骨髓细胞、wehi-3b(d+)、aml-193(atcc)或重组g-csf受体产生细胞)监测bst受体结合。例如参见金(king)等人,实验血液学(exp.hematol.)20:223(1992);美国专利第6,385,505号,都以引用的方式并入本文中。用于测试hg-csf活性的体内动物模型以及人类临床试验包括例如美国专利第6,166,183号、第6,565,841号、第6,162,426号、第5,718,893号中所述者,各专利以引用的方式并入本文中。所述模型可用于评价bst活性。
不管使用哪些方法来产生本发明的bst类似物,这些类似物都需经历生物活性分析。可以进行氚化胸苷分析来确定细胞分裂的程度。但也可使用其它生物分析来确定所需活性。生物分析,例如针对诱导小鼠wehi-3b(d+)白血病细胞系终末分化的能力的分析,也指示g-csf活性。参见尼科拉(nicola)等人,血液学(blood)54:614-27(1979)。可以使用其它体外分析来确定生物活性。参见尼科拉,生物化学年评(ann.rev.biochem.)58:45-77(1989)。一般说来,针对生物活性的测试应提供针对所需结果进行的分析,例如生物活性增加或降低(与未改变的g-csf相比较)、不同的生物活性(与未改变的g-csf相比较)、受体或结合搭配物亲和力分析、bst本身或其受体的构象或结构改变(与经修饰的bst相比较),或血清半衰期分析。
先前曾报导,来自新近诊断的白血病的wehi-3bd+细胞和人白血病细胞将结合125i标记的鼠类g-csf,并且通过添加未标记的g-csf或人csf-β可与这一结合竞争。测试天然g-csf和bst与125i-g-csf竞争结合于人类和鼠类白血病细胞的能力。用碘处理高度纯化的天然g-csf(纯度>95%;1μg)[特赫多尔(tejedor)等人,分析生物化学(anal.biochem.),127,143(1982)],并通过凝胶过滤和离子交换色谱法与反应物分离。天然125i-g-csf的比活性为约100μci/μg蛋白质。
上述有关分析方法的参考文献的汇编并不详尽,并且所属领域技术人员将认识到适用于测试所需最终结果的其它分析。所属领域技术人员已知对这些分析的改变。
xiii.效力、功能性体内半衰期和药物动力学参数的测量
本发明的一个重要方面是,通过构建结合或不结合水溶性聚合物部分的b-gcsf多肽,来获得较长的生物半衰期。投药后bst多肽血清浓度的迅速降低对于评估用结合和未结合的bst多肽和其变异体进行治疗的生物反应极为重要。本发明的结合和未结合的bst多肽和其变异体在经由例如皮下或静脉内投药后,具有较长的血清半衰期,使得可能通过例如elisa方法或初级筛选分析进行测量。可以使用商业来源的elisa或ria试剂盒。用于测量hg-csf或其变异体体内半衰期的分析的另一实例描述于美国专利第5,824,778号中,该案以引用的方式并入本文中。体内生物半衰期的测量是如本文中所述进行的。
包含非天然编码氨基酸的hg-csf多肽的效力和功能性体内半衰期可根据以下专利中所述的方案测定:美国专利第6,646,110号、第6,555,660号、第6,166,183号、第5,985,265号、第5,824,778号、第5,773,581号,各案以引用的方式并入本文中。这些方案也可用于bst。
可利用正常的斯普拉格-道来(sprague-dawley)雄性大鼠评估包含非天然编码氨基酸的bst多肽的药物动力学参数(每一治疗组n=5只动物)。使动物接受静脉内25微克/大鼠或皮下50微克/大鼠的单次剂量,并根据预定时程取得约5到7个血样,对于包含非天然编码氨基酸且未结合水溶性聚合物的bst多肽,所述时程一般包含约6小时,而对于包含非天然编码氨基酸且与水溶性聚合物结合的bst多肽,所述时程一般包含约4天。可将不含非天然编码氨基酸的bst的药物动力学数据直接与由包含非天然编码氨基酸的bst多肽获得的数据相比较。
bst多肽的药物动力学研究可以在小鼠、大鼠或灵长类动物(例如猕猴)中执行。通常,经皮下或静脉内投予单次注射液,并随时间监测血清bst含量。
所属领域技术人员已知用于评估动物健康状况、产奶量、生长和其它参数的方法。所属领域技术人员已知可用于评估本发明bst多肽的其它模型和这些模型。
用于测量hg-csf或其变异体的体内生物活性的分析的其它实例描述于美国专利第5,681,720号、第5,795,968号、第5,824,778号、第5,985,265号以及鲍文(bowen)等人,实验血液学,27:425-432(1999)中,各文献以引用的方式并入本文中。
xiv.投药和医药组合物
本发明的多肽或蛋白质(包括(但不限于)包含一个或一个以上非天然氨基酸的bst、合成酶、蛋白质等)任选与(包括(但不限于))适当医药载剂组合用于治疗用途。所述组合物例如包含治疗有效量的化合物和医药学上可接受的载剂或赋形剂。所述载剂或赋形剂包括(但不限于)生理盐水、缓冲生理盐水、右旋糖、水、甘油、乙醇和/或其组合。调配物被制备成与投药模式相适应。一般来说,所属领域技术人员已知投予蛋白质的方法,并且这些方法适用于投予本发明的多肽。组合物可呈水溶性形式,例如以医药学上可接受的盐形式存在,其中意欲包括酸和碱加成盐。bst调配和投药可通过所属领域技术人员已知的方法实现。论述了包含硫酸根离子,例如硫酸铵、硫酸钠、硫酸镁和其混合物;以及缓冲剂,例如乙酸盐、柠檬酸盐、磷酸盐、hepes、bes、taps、epps、tes和其混合物的盐。
根据所属领域技术人员已知的方法,任选在一个或一个以上适当的体外和/或体内动物疾病模型中测试包含一种或一种以上本发明多肽的治疗组合物,以便确定功效、组织代谢并估计剂量。具体说来,最初可通过本文中非天然氨基酸相对于天然氨基酸同系物的活性、稳定性或其它适当测量(包括(但不限于),经修饰以包括一个或一个以上非天然氨基酸的bst多肽与天然氨基酸bst多肽相比较和经修饰以包括一个或一个以上非天然氨基酸的bst多肽与目前可用的bst处理相比较)(即,在相关分析中)来测定剂量。
投药是通过常用于引入分子最终使其与血液或组织细胞接触的任何途径进行。本发明的非天然氨基酸多肽是以任何适当的方式任选与一种或一种以上医药学上可接受的载剂一起投予。向患者投予本发明上下文中所述的这些多肽的适当方法都可使用,而且,尽管可使用一种以上途径投予特定组合物,但特定途径通常可提供比另一途径快捷和有效的作用或反应。
医药学上可接受的载剂部分地是由所投予的特定组合物以及用于投予组合物的特定方法确定。因此,存在多种适当的本发明医药组合物的调配物。
可通过适于蛋白质或肽的任何常规途径投予本发明的bst多肽,所述途径包括(但不限于)肠胃外方式,例如注射,包括(但不限于)皮下注射或静脉内注射,或任何其它注射或输注形式。多肽组合物可以多种途径投予,包括(但不限于)经口、静脉内、腹膜内、肌肉内、透皮、皮下、局部、舌下、血管内、乳腺内或直肠方式。还可经由脂质体投予包含经修饰或未经修饰的非天然氨基酸多肽的组合物。所属领域技术人员一般已知所述投药途径和适当的调配物。bst多肽可单独使用或与其它适合的组分(例如医药载剂)组合使用。bst多肽可与其它试剂或治疗剂组合使用。
也可将单独或与其它适当组分组合的包含非天然氨基酸的bst多肽制成气雾剂调配物(即,其可经“雾化”)以经由吸入投予。可将气雾剂调配物放入加压可接受的推进剂(例如二氯二氟甲烷、丙烷、氮气等)中。
适于肠胃外投予(例如,经由关节内(关节中)、静脉内、肌肉内、皮内、腹膜内和皮下途径投予)的调配物包括水性与非水性等渗无菌注射液,其可含有抗氧化剂、缓冲剂、抑菌剂和使调配物与预定接受者的血液等渗的溶质;以及水性与非水性无菌悬浮液,其可包括悬浮剂、增溶剂、增稠剂、稳定剂和防腐剂。bst调配物可提供于单位剂量或多剂量密封容器(例如安瓿和小瓶)中。
肠胃外投药和静脉内投药是优选的投药方法。具体说来,已经用于天然氨基酸同系物治疗剂的投药途径(包括(但不限于),通常用于epo、gh、g-csf、gm-csf、ifn、白细胞介素、抗体、fgf和/或任何其它以医药学方式递送的蛋白质的途径)连同当前使用的调配物,将提供优选的投药途径和本发明多肽的调配物。
在本发明上下文中,投予动物的剂量足以在动物体内随时间引起有益的治疗反应或视应用而定引起其它适当的活性。所述剂量是由特定载体或调配物的功效和所用非天然氨基酸多肽的活性、稳定性或血清半衰期,和动物的病状,以及欲治疗动物的体重或表面积确定。剂量大小也可通过在特定动物体内伴随投予特定载体、调配物等的任何不利副作用的存在、性质和程度来确定。
在确定治疗或预防疾病时欲投予的载体或调配物的有效量时,兽医将评估循环血浆含量、调配物毒性、疾病进程和/或(相关时)抗非天然氨基酸多肽抗体的产生。
投药剂量通常在等于当前使用的治疗蛋白质的剂量的范围内,所述范围可针对相关组合物的活性或血清半衰期的改变而调整。本发明的载体或医药调配物可通过任何已知的常规疗法,包括抗体投药、疫苗投药、细胞毒性剂、天然氨基酸多肽、核酸、核苷酸类似物、生物反应修饰剂的投药等来补充治疗条件。
对于投药,本发明的调配物是以由相关调配物的ld-50或ed-50和/或在(包括(但不限于))适用于动物的质量和总体健康状况的各种浓度下,非天然氨基酸多肽的任何副作用的观察结果所确定的施用率投予。投药可经由单次剂量或分次剂量实现。
如果输注调配物的动物出现发烧、寒战或肌肉疼痛,那么其可接受适当剂量的阿斯匹林(aspirin)、布洛芬(ibuprofen)、乙酰胺苯酚或其它适于动物的疼痛/发烧控制药物。经历例如发烧、肌肉疼痛和寒战等输注反应的动物,在将进行输注之前30分钟,用阿斯匹林、乙酰胺苯酚或(包括(但不限于))苯海拉明(diphenhydramine),或另一适于动物的药物预先用药。派替啶(meperidine)可用于不能对退热剂和抗组织胺快速作出反应的较为严重的寒战和肌肉疼痛。视反应的严重性而减慢或中断细胞输注。
本发明的bst多肽可直接投予动物个体。投药是通过通常用于将bst多肽引入个体的任何途径进行。根据本发明实施例的bst多肽组合物包括适于以下投药的组合物:经口、直肠、局部、吸入(包括(但不限于)通过气雾剂)、颊内(包括(但不限于)舌下)、阴道、肠胃外(包括(但不限于)皮下、肌内、皮內、关节内、胸膜内、腹膜內、脑内、动脉内或静脉内)、局部(意即,皮肤和粘膜表面,包括气道表面)、肺、眼内、鼻内和透皮投药,但在任何给定情况下,最适合的途径将取决于所治疗的病状的性质和严重性。投药可为局部的或全身投药。化合物调配物可提供于单位剂量或多剂量密封容器(例如安瓿和小瓶)中。本发明的bst多肽可与医药学上可接受的载剂制备成呈单位剂量可注射形式(包括(但不限于)溶液、悬浮液或乳液)的混合物。本发明的bst多肽也可通过连续输注(包括(但不限于),使用例如渗透泵等微型泵)、单次团注或缓慢释放储槽式调配物投予。
适于投药的调配物包括可含有抗氧化剂、缓冲剂、抑菌剂和使调配物等渗的溶质的水性与非水性溶液、等渗无菌溶液,以及可包括悬浮剂、增溶剂、增稠剂、稳定剂和防腐剂的水性与非水性无菌悬浮液。溶液和悬浮液可由前文所述种类的无菌粉末、颗粒和片剂制备而来。
冷冻干燥是用于将水从相关蛋白质制剂中去除的提供蛋白质的常用技术。冷冻干燥或冻干法是一种首先将欲干燥的材料冷冻,随后通过在真空环境中升华去除冰或冷冻溶剂的方法。预先冻干的调配物中可包括赋形剂,以在冷冻干燥过程中增强稳定性和/或改进储存时冻干产物的稳定性。皮卡尔m.(pikal,m.),生物医药(biopharm.)3(9)26-30(1990),和荒川(arakawa)等人,医药研究(pharm.res.)8(3):285-291(1991)。
所属领域技术人员也已知医药品的喷雾干燥。例如参见波罗德海德j.(broadhead,j.)等人,“医药品的喷雾干燥(thespraydryingofpharmaceuticals)”,药物开发与工业制药(drugdev.ind.pharm),18(11和12),1169-1206(1992)中。除小分子医药品之外,也已将各种生物材料喷雾干燥,并且所述生物材料包括:酶、血清、血浆、微生物和酵母。喷雾干燥是一项有用的技术,这是因为其可在单步骤方法中将液体医药制剂转化成精细、无尘的或成团的粉末。基本技术包含以下四个步骤:a)将进料溶液雾化成喷雾;b)喷雾-空气接触;c)干燥喷雾;和d)使干燥产物与干燥空气分离。美国专利第6,235,710号和第6,001,800号(以引用的方式并入本文中)中描述通过喷雾干燥制备重组促红细胞生成素。
本发明的医药组合物和调配物可包含医药学上可接受的载剂、赋形剂或稳定剂。医药学上可接受的载剂部分地是由所投予的特定组合物以及用于投予组合物的特定方法确定。因此,存在本发明医药组合物(包括任选使用的医药学上可接受的载剂、赋形剂或稳定剂)的多种适当调配物(例如参见雷氏药学大全(remington'spharmaceuticalsciences,)第17版,1985)。
适合的载剂包括(但不限于)含有琥珀酸盐、磷酸盐、硼酸盐、hepes、柠檬酸盐、组氨酸、咪唑、乙酸盐、碳酸氢盐和其它有机酸的缓冲液;抗氧化剂,包括(但不限于)抗坏血酸;低分子量多肽,包括(但不限于)小于约10个残基的多肽;蛋白质,包括(但不限于)血清白蛋白、明胶或免疫球蛋白;亲水性聚合物,包括(但不限于)聚乙烯吡咯烷酮;氨基酸,包括(但不限于)甘氨酸、谷氨酰胺、天冬酰胺、精氨酸、组氨酸或组氨酸衍生物、甲硫氨酸、谷氨酸或赖氨酸;单糖、二糖和其它碳水化合物,包括(但不限于)海藻糖、蔗糖、葡萄糖、甘露糖或糊精;螯合剂,包括(但不限于)edta和依地酸二钠(edentatedisodium);二价金属离子,包括(但不限于)锌、钴或铜;糖醇,包括(但不限于)甘露糖醇或山梨糖醇;成盐反离子,包括(但不限于)钠和氯化钠;填充剂,例如微晶纤维素、乳糖、玉米和其它淀粉;粘合剂;甜味剂和其它调味剂;着色剂;和/或非离子型表面活性剂,包括(但不限于)tweentm(包括(但不限于)tween80(聚山梨酸酯80)和tween20(聚山梨酸酯20))、pluronicstm和其它普流尼克酸(pluronicacid)(包括(但不限于)普流尼克酸f68(泊洛沙姆188(poloxamer188)))或peg。适合的表面活性剂例如包括(但不限于)基于聚(氧化乙烯)-聚(氧化丙烯)-聚(氧化乙烯),即(peo-ppo-peo)的聚醚;或基于聚(氧化丙烯)-聚(氧化乙烯)-聚(氧化丙烯),即(ppo-peo-ppo)的聚醚;或其组合。peo-ppo-peo和ppo-peo-ppo在市面上以商品名pluronicstm、r-pluronicstm、tetronicstm和r-tetronicstm(巴斯夫怀恩多特公司(basfwyandottecorp.),密歇根州怀恩多特(wyandotte,mich.))销售,并且进一步描述于美国专利第4,820,352号中,所述专利以全文引用的方式并入本文中。其它乙烯/聚丙烯嵌段聚合物可为适合的表面活性剂。表面活性剂或表面活性剂组合可用于使聚乙二醇化bst对一种或一种以上应力(包括(但不限于)由搅动产生的应力)稳定。上述一些表面活性剂可称为“增积剂”。一些也可称为“张力改变剂(tonicitymodifier)”。抗菌防腐剂也适用于产品稳定性和抗菌有效性;适合的防腐剂包括(但不限于)苯甲醇、苯扎氯铵(benzalkoniumchloride)、间甲酚、对羟基苯甲酸甲酯/对羟基苯甲酸丙酯、甲酚和苯酚,或其组合。美国专利第7,144,574号(以引用的方式并入本文中)描述适用于本发明的医药组合物和调配物以及其它递送制剂中的其它材料。
本发明的bst多肽,包括与例如peg等水溶性聚合物连接的bst多肽,也可通过持续释放系统或作为持续释放系统的一部分投予。持续释放组合物包括(包括(但不限于))呈包括(但不限于)膜或微胶囊的成形物件形式的半渗透聚合物基质。持续释放基质包括生物相容性材料,例如聚(甲基丙烯酸2-羟乙酯)(朗格(langer)等人,生物医用材料研究(j.biomed.mater.res.),15:267-277(1981);朗格,化学技术(chem.tech.),12:98-105(1982))、乙烯乙酸乙烯酯(朗格等人,同上文)或聚d-(-)-3-羟基丁酸(ep133,988)、聚丙交酯(聚乳酸)(美国专利第3,773,919号;ep58,481)、聚乙交酯(乙醇酸聚合物)、聚丙交酯共聚乙交酯(乳酸与乙醇酸的共聚物)、聚酐、l-谷氨酸与γ-乙基-l-谷氨酸的共聚物(赛德曼(sidman)等人,生物聚合物(biopolymers),22,547-556(1983))、聚(原酸)酯、多肽、透明质酸、胶原蛋白、硫酸软骨素、羧酸、脂肪酸、磷脂、多糖、核酸、聚氨基酸、氨基酸(例如苯丙氨酸、酪氨酸、异亮氨酸)、聚核苷酸、聚乙烯丙烯、聚乙烯吡咯烷酮和硅酮。持续释放组合物还包括脂质体包埋的化合物。可利用本身已知的方法制备含有化合物的脂质体:de3,218,121;艾普斯坦(eppstein)等人,美国国家科学院院刊,82:3688-3692(1985);黄(hwang)等人,美国国家科学院院刊,77:4030-4034(1980);ep52,322;ep36,676;美国专利第4,619,794号;ep143,949;美国专利第5,021,234号;日本专利申请案83-118008;美国专利第4,485,045号和第4,544,545号;以及ep102,324。引用的所有参考文献和专利都以引用的方式并入本文中。
可利用以下文献中所述的方法制备经脂质体包埋的bst多肽:例如de3,218,121;艾普斯坦(eppstein)等人,美国国家科学院院刊,82:3688-3692(1985);黄(hwang)等人,美国国家科学院院刊,77:4030-4034(1980);ep52,322;ep36,676;美国专利第4,619,794号;ep143,949;美国专利第5,021,234号;日本专利申请案83-118008;美国专利第4,485,045号和第4,544,545号;以及ep102,324。脂质体的组成和尺寸已为人们所熟知或者能够凭所属领域技术人员的经验容易地确定。脂质体的一些实例描述于例如帕克jw(parkjw)等人,美国国家科学院院刊,92:1327-1331(1995);拉斯科d(lasicd)和帕普卓普洛斯d(papahadjopoulosd)(编):脂质体的医学应用(medicalapplicationsofliposomes)(1998);杜蒙德dc(drummonddc)等人,用于癌症疗法的脂质体药物递送系统(liposomaldrugdeliverysystemsforcancertherapy),泰彻尔b(teicherb)(编):癌症药物新发现和开发(cancerdrugdiscoveryanddevelopment)(2002);帕克jw等人,临床癌症研究(clin.cancerres.)8:1172-1181(2002);尼尔森ub(nielsenub)等人,生物化学与生物物理学文献(biochim.biophys.acta)1591(1-3):109-118(2002);玛莫特c(mamotc)等人,癌症研究(cancerres).63:3154-3161(2003)中。引用的所有参考文献和专利都以引用的方式并入本文中。已经描述了多种hg-csf调配物,且都为所属领域技术人员所知。
在本发明的上下文中,投予动物的剂量应足以随时间在个体体内引起有益的反应。一般来说,每剂肠胃外投予的本发明bst多肽的总医药有效量在每天每公斤动物体重约0.01微克到约100微克,或者每天每公斤动物体重约0.05毫克到约1毫克的范围内,但所述有效量应服从治疗判断。给药的频率也应服从治疗判断,并且可比批准用于动物的市售bst多肽产品的频率更高或更低。一般可通过上述任何投药途径投予本发明的聚乙二醇化bst多肽。
xv.本发明bst多肽的治疗用途
本发明的bst多肽适用于治疗多种病症。投予bst产物引起产奶量增加、体重增益增加等。因此,投予本发明的bst多肽适用于预防有感染风险的动物发生感染。在本发明一个实施例中,在介于产犊前两周与一天间对动物投予本发明的聚乙二醇化bst多肽。在本发明一个实施例中,在介于产犊前两周与一天间对动物投予本发明的聚乙二醇化bst多肽,且再在产犊当天或在产犊后至多一周投药。在本发明一个实施例中,在介于产犊前两周与一天间对动物投予本发明的bst多肽。在本发明一个实施例中,在介于产犊前两周与一天间对动物投予本发明的bst多肽,且再在产犊当天或在产犊后至多一周投药。在本发明一个实施例中,在介于产犊前一周与一天间对动物投予本发明的聚乙二醇化bst多肽。在本发明一个实施例中,在介于产犊前一周与一天间对动物投予本发明的聚乙二醇化bst多肽,且再在产犊当天或在产犊后至多一周投药。在本发明一个实施例中,在介于产犊前一周与一天间对动物投予本发明的bst多肽。在本发明一个实施例中,在介于产犊前一周与一天间对动物投予本发明的bst多肽,且再在产犊当天或在产犊后至多一周投药。
在本发明一个实施例中,在介于航运前两周与当天间对动物投予本发明的bst多肽。在本发明一个实施例中,在介于航运前一周与一天间对动物投予本发明的bst多肽。在本发明一个实施例中,在介于航运前一周与一天间对动物投予本发明的bst多肽,且再在航运当天或在航运后至多一周投药。
在本发明一个实施例中,在产犊前7天对动物投予本发明的聚乙二醇化bst多肽。在本发明一个实施例中,在产犊前7天对动物投予本发明的聚乙二醇化bst多肽,且再在产犊当天或在产犊后至多一周投药。在本发明一个实施例中,在产犊前7天对动物投予本发明的聚乙二醇化bst多肽,且再在产犊当天投药。在本发明一个实施例中,在产犊前7天对动物投予本发明的bst多肽。在本发明一个实施例中,在产犊前一周对动物投予本发明的bst多肽,且再在产犊当天或在产犊后至多一周投药。在本发明一个实施例中,在产犊前一周对动物投予本发明的bst多肽,且再在产犊当天投药。在本发明一个实施例中,在产犊前或在产犊当天对奶牛投予本发明的bst多肽,以预防小牛患病。在本发明一个实施例中,在产犊前或在产犊当天对奶牛投予本发明的聚乙二醇化bst多肽,以预防小牛患病。在本发明一个实施例中,在产犊前对奶牛投予本发明的bst多肽,以预防小牛患病。在本发明一个实施例中,在产犊前对奶牛投予本发明的聚乙二醇化bst多肽,以预防小牛患病。在一个实施例中,本发明bst多肽的投药剂量为0.01、0.02、0.03、0.04、0.05、0.06、0.07、0.08、0.09、0.10、0.11、0.12、0.13、0.14、0.15、0.16、0.17、0.18、0.19、0.20、0.21、0.22、0.23、0.24、0.25、0.26、0.27、0.28、0.29、0.30、0.31、0.32、0.33、0.34、0.35、0.36、0.37、0.38、0.39、0.40、0.41、0.42、0.43、0.44、0.45、0.46、0.47、0.48、0.49或0.50μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.01、0.02、0.03、0.04、0.05、0.06、0.07、0.08、0.09、0.10、0.11、0.12、0.13、0.14、0.15、0.16、0.17、0.18、0.19、0.20、0.21、0.22、0.23、0.24、0.25、0.26、0.27、0.28、0.29、0.30、0.31、0.32、0.33、0.34、0.35、0.36、0.37、0.38、0.39、0.40、0.41、0.42、0.43、0.44、0.45、0.46、0.47、0.48、0.49或0.50μg/kg。在一个实施例中,本发明bst多肽经聚乙二醇化,且投药剂量为0.01、0.02、0.03、0.04、0.05、0.06、0.07、0.08、0.09、0.10、0.11、0.12、0.13、0.14、0.15、0.16、0.17、0.18、0.19、0.20、0.21、0.22、0.23、0.24、0.25、0.26、0.27、0.28、0.29、0.30、0.31、0.32、0.33、0.34、0.35、0.36、0.37、0.38、0.39、0.40、0.41、0.42、0.43、0.44、0.45、0.46、0.47、0.48、0.49或0.50μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽经聚乙二醇化,且投药剂量为0.01、0.02、0.03、0.04、0.05、0.06、0.07、0.08、0.09、0.10、0.11、0.12、0.13、0.14、0.15、0.16、0.17、0.18、0.19、0.20、0.21、0.22、0.23、0.24、0.25、0.26、0.27、0.28、0.29、0.30、0.31、0.32、0.33、0.34、0.35、0.36、0.37、0.38、0.39、0.40、0.41、0.42、0.43、0.44、0.45、0.46、0.47、0.48、0.49或0.50μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.01μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.01μg/kg。
在一个实施例中,本发明bst多肽的投药剂量为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8或1.0μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8或1.0μg/kg。在一个实施例中,本发明bst多肽经聚乙二醇化,且投药剂量为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8或1.0μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽经聚乙二醇化,且投药剂量为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8或1.0μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.1μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.1μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.2μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.2μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.3μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.3μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.4μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.4μg/kg。在一个实施例中,本发明bst多肽的投药剂量为0.5μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为0.5μg/kg。
在一个实施例中,本发明bst多肽的投药剂量为1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60μg/kg。在一个实施例中,本发明bst多肽的投药剂量为10μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为10μg/kg。在一个实施例中,本发明bst多肽的投药剂量为20μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为20μg/kg。在一个实施例中,本发明bst多肽的投药剂量为30μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为30μg/kg。在一个实施例中,本发明bst多肽的投药剂量为40μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为40μg/kg。在一个实施例中,本发明bst多肽的投药剂量为50μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量为50μg/kg。在一个实施例中,本发明bst多肽的投药剂量高于0.5μg/kg。在一个实施例中,本发明的聚乙二醇化bst多肽的投药剂量高于0.5μg/kg。
含有bst的医药组合物的调配浓度可有效以各种方式投予罹患以产生较少或缺陷型白细胞为特征的病症的动物,所述特征是单独出现或作为病状或疾病的一部分出现。bst的平均量可不同,具体地说,所述平均量应基于有资质的兽医的推荐和处方。bst的确切量应优先考虑例如以下因素:所治疗病状的确切类型、所治疗动物的状况以及组合物中的其它成分。本发明也投予治疗有效量的另一活性剂。所属领域技术人员可根据利用bst的疗法容易地确定欲给予的量。由此,可以使用本发明的bst刺激产奶和生长等。
本发明的医药组合物可以用常规方式制备。
实例
提供的以下实例将说明而非限制主张的发明。
实例1
在bst中并入非天然编码氨基酸的位点选择
本实例描述用于选择在bst中并入非天然编码氨基酸的位点的众多可能标准组中的一部分。
已知牛促生长素的晶体结构,并且所选供取代的可能残基包括(但不限于)保守取代位点以及使用cx程序具有最大溶剂可接近性的残基(品塔(pintar)等人(2002)生物信息学(bioinformatics),18(7):980-4)。鉴别出的适于用对乙酰基苯丙氨酸取代的保守取代位点包括(但不限于)含有带电或不带电的疏水性核心的酪氨酸、苯丙氨酸和精氨酸残基。没有选择与结构相关的残基进行取代,包括(但不限于)甘氨酸、脯氨酸以及螺旋封端所涉及的残基。也不能选择已知受体结合区中的残基进行取代。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst中的以下一个或一个以上位置:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16,17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:2)。
在一些实施例中,将一个或一个以上非天然编码氨基酸并入bst的以下一个或一个以上位置:seqidno:1或seqidno:2的35、91、92、94、95、99、101、133、134、138、139、140、142、144、149、150、154,和其任何组合。
在一些实施例中,将一个或一个以上所述位置的非天然存在氨基酸与水溶性聚合物连接,所述位置包括(但不限于)以下位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1)。在一些实施例中,将一个或一个以上所述位置的非天然存在氨基酸与水溶性聚合物连接,所述位置包括(但不限于)以下位置:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:2)。
实例2
在大肠杆菌中产生的含有非天然编码氨基酸的bst多肽的克隆和表达
本实例详述在大肠杆菌中包括非天然编码氨基酸的bst多肽的克隆和表达,以及评估经修饰bst多肽生物活性的方法。
所属领域技术人员已知克隆bst的方法。bst的多肽和聚核苷酸序列和宿主细胞中bst的克隆以及bst的纯化详述于美国专利第5,849,883号(全文以引用的方式并入本文中)和海达瑞(heidari)等人,兽医免疫学与免疫病理学(veterinaryimmunologyandimmunopathology)(2001)81:45-57中。
使用引入的包含正交trna(o-trna)和正交氨酰基trna合成酶(o-rs)的翻译系统表达含有非天然编码氨基酸的bst。o-rs优先利用非天然编码氨基酸使o-trna氨酰基化。接着,翻译系统对编码的选择密码子反应将非天然编码氨基酸插入bst中。适合的o-rs和o-trna序列描述于以下专利中:wo2006/068802,名为“氨酰基trna合成酶的组合物以及其用途(compositionsofaminoacyl-trnasynthetaseandusesthereof)”(在本申请案中,e9-seqidno:22和e9的d286r突变体-seqidno:24);和wo2007/021297,名为“trna的组合物以及其用途(compositionsoftrnaandusesthereof)”(在本申请案中,f13;seqidno:23),所述专利都以全文引用的方式并入本文中。
表2:o-rs和o-trna序列。
用含有经修饰bst聚核苷酸序列和正交氨酰基trna合成酶/trna对(对所需非天然编码氨基酸具特异性)的质粒转化大肠杆菌,从而将非天然编码氨基酸位点特异性地并入bst多肽中。作为实例显示的相关基因是用选择密码子(琥珀密码子)代替seqidno:1或2中一个或一个以上密码子的bst。所得bst多肽中以下一个位置处的天然编码氨基酸经非天然编码氨基酸对乙酰基苯丙氨酸(paf;pacf)取代:seqidno:1或2的1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191。
产生所选位置和突变bst多肽,验证序列,将质粒转化到w3110b2细胞中,并使集落在氨必西林板上生长。使用这些集落以1:1000的氨必西林培养物稀释度接种到5mllb中,使其在37℃下生长到o.d.600=0.8。随后将paf(对乙酰基苯丙氨酸)添加到15种不同培养物中,达到4mm最终浓度。约30分钟后,用l-阿拉伯糖诱导培养物,达到0.2%最终浓度,并在37℃再培育培养物5小时。此时,从每一培养物中取出500μl样品,并以13,000rpm短暂离心4分钟。丢弃上清液,并将离心块再悬浮于含有1μldnase的150μlb-per中,且在室温下培育过夜。次日清晨,添加4×lds样品缓冲液(英杰公司,加利福尼亚州卡尔斯巴德),将样品加热到95℃,保持5分钟,并添加10×样品还原剂(英杰公司,加利福尼亚州卡尔斯巴德)。随后,借助于sds-page在含4到12%梯度凝胶(英杰公司,加利福尼亚州卡尔斯巴德)的mes缓冲液中解析样品,并使用simplybluesafestain(英杰公司,加利福尼亚州卡尔斯巴德)观察。图1显示由hgh培养物产生的样品在4到12%梯度凝胶上分析以及考马斯染色后的结果。
包涵体的预备溶解
将细胞糊状物混合于4℃的包涵体(inclusionbody,ib)缓冲液i(50mmtrisph8.0;100mmnacl;1mmedta;1%tritonx-100;4℃)中达到最终10%固体,来使细胞糊状物再悬浮。使再悬浮的物质通过微量流化床共计2次,来溶解细胞。在4℃下,以10,000g离心样品15分钟,并倾析上清液。将包涵体(ib)小球再悬浮于另一体积ib缓冲液i(50mmtrisph8.0;100mmnacl;1mmedta;1%tritonx-100;4℃)中来进行洗涤,并使再悬浮的物质通过微量流化床共计2次。随后在4℃下,以10,000g离心样品15分钟,并倾析上清液。将ib小球再悬浮于1体积缓冲液ii(50mmtrisph8.0;100mmnacl;1mmedta;4℃)中。再悬浮后,在4℃下,以10,000g离心样品15分钟,并倾析上清液。接着将ib小球再悬浮于1/2体积缓冲液ii(50mmtrisph8.0;100mmnacl;1mmedta;4℃)中。接着将ib等分到适宜的容器中。在4℃下,以10,000g离心样品15分钟,并倾析上清液。接着将包涵体溶解或在-80℃储存待用。
包涵体溶解
在溶解缓冲液(20mmtris,ph8.0;8m胍;10mmβ-me)中溶解包涵体,达到介于10到15mg/ml之间的最终浓度。室温下,在恒定混合下,培育溶解的ib1小时或直到其完全溶解。如果蛋白质浓度较高,那么通过再用溶解缓冲液稀释来调整蛋白质浓度。
再折叠
通过在0.5m精氨酸(ph8.0;4℃)中稀释样品达到0.5mg/ml的最终蛋白质浓度,来执行再折叠。在4℃下,使样品再折叠48到72小时。
纯化
在小心混合下,将固体(nh4)2so4添加到样品中,达到20%最终浓度。在4℃下小心混合样品30分钟。以12,000g离心15分钟来使沉淀的蛋白质(含有bst)聚结成球。去除上清液,并将小球再悬浮于1/2再折叠体积的20mmnaac(ph4.5)中。所有小球都不再溶解于溶液中。只有bst溶解于溶液中。以12,000g离心15分钟来使未溶解的物质聚结成球。倾析样品,并保留上清液。使bst物质滤过0.45μm滤膜。随后,将物质装载到在缓冲液a(20mmnaac,ph4.5)中平衡的cmff柱(ge医疗集团(gehealthcare))上。在装载到柱上之前,物质<10m/s。利用约10个柱体积达到100%缓冲液b(20mmnaac,ph4.5;500mmnacl)的线性梯度从柱中洗脱出bst。
聚乙二醇化和纯化
利用50%冰乙酸将cm池的ph值调到ph4.0。随后,将所述池浓缩到约4.0mg/ml蛋白质。将12:1或8:1摩尔过量的羟胺peg:bst添加到所述池中。在28℃下培育混合物48到72小时。随后,用水(<8m/s)稀释混合物8到10倍,接着装载到在缓冲液a(20mmnaac,ph4.5)中平衡的sphp柱(ge医疗集团(gehealthcare))上。利用约40个柱体积达到100%缓冲液b(20mmnaac,ph4.5;500mmnacl)的线性梯度洗脱出聚乙二醇化bst。
汇集聚乙二醇化bst洗脱部分,并针对bst调配物缓冲液(4.26mmnaac,ph4.0;0.565mmnacl;0.0033%tween20;5%山梨糖醇)进行透析。将peg物质浓缩到浓度介于6到8mg/ml蛋白质之间,并使用0.22μmpes滤膜进行无菌过滤。将蛋白质储存于4℃下,或者速冻并在-80℃储存以延长储存期。图6显示聚乙二醇化之前和之后bst的sds-page分析。
bst的肽图分析(胰蛋白酶/胞内蛋白酶glu-c)
执行肽图分析以确定对乙酰基苯丙氨酸(paf)并入bst多肽中。将尚未聚乙二醇化的纯化的bst以及野生型bst稀释到最终6m盐酸胍、50mmtris(ph7.8)中,并在37℃下用10mmdtt还原1小时。室温下,于暗处用20mmiaa使样品烷基化40分钟,并通过添加dtt达到20mm的最终浓度来中止反应。将物质透析到100mm碳酸氢铵(ph7.7)中,并在37℃下,用1:50(蛋白质:酶)胰蛋白酶处理4小时。在此反应后,在25℃下添加1:20的glu-c,保持过夜。通过添加tfa达到0.1%的最终浓度,来中止消化。将样品施加到串联thermofinniganlcqdeca离子阱质谱计的gracevydacc8反相柱上。以98%流动相a(含0.05%tfa的水)开始等度梯度洗脱,持续8分钟,随后匀变到60%流动相b(含0.05%tfa的乙腈),持续90分钟,同时在214nm和250nm下检测。所用流速为0.2ml/min且柱温为40℃。毛细管电压设置为15v,且完整扫描范围为100到2000m/z。ms/ms的碰撞电压(collisionvoltage)为标准化电压的42%。
bst的肽图分析(胞内蛋白酶glu-c)
将尚未聚乙二醇化的纯化的bst稀释到最终6m盐酸胍、50mmtris(ph7.8)中,并在37℃下用10mmdtt还原1小时。室温下,于暗处用20mmiaa使样品烷基化40分钟,并通过添加dtt达到20mm的最终浓度来中止反应。将物质透析到100mm碳酸氢铵(ph7.7)中,并在25℃下,用1:20(蛋白质:酶)glu-c处理过夜。通过添加tfa达到0.1%的最终浓度,来中止消化。将样品施加到串联thermofinniganlcqdeca离子阱质谱计的gracevydacc8反相柱上。以98%流动相a(含0.05%tfa的水)开始等度梯度洗脱,持续8分钟,随后匀变到60%流动相b(含0.05%tfa的乙腈),持续90分钟,同时在214nm和250nm下检测。所用流速为0.2ml/min且柱温为40℃。毛细管电压设置为15v,且完整扫描范围为100到2000m/z。ms/ms的碰撞电压为标准化电压的42%。
bst多肽的rp-hplc和sec-hplc分析
纯化后,使用rp-hplc和sec-hplc分析样品的纯度并确定其身份。利用调配物缓冲液(4.26mm乙酸钠(ph4.0)、0.565mm氯化钠、0.0033%tween-20和5%山梨糖醇)将纯化的聚乙二醇化bst稀释到1mg/ml,并将10μl注射到j.t.baker宽孔辛基(c8)反相柱(4.6×100mm;5μm)上。以50%流动相a(含0.1%tfa的水)开始梯度洗脱,并经26分钟匀变到70%流动相b(含0.1%tfa的乙腈)。使柱再生,测量流速,并应用60℃柱温,同时在214nm下检测。可使用安捷伦化学工作站(agilentchemstation)软件执行分析。
m-nfs60增殖分析
为了评价bst分子的效力,可执行增殖分析。每两天分裂细胞,并以0.02×106个细胞/毫升接种。
分析前一天,细胞分裂为0.1×106个细胞/毫升。16到24小时后,细胞以10,000个细胞/孔接种到黑色平底96孔板中的分析培养基中,并一式两份添加bst化合物的连续稀释液。每孔的总体积为100μl,且分析培养基(assaymedium)为rpmi1640+10%fbs+p/s。也一式两份将标准品(例如
在数据分析中,对每种化合物的一式两份的连续稀释液求取平均值,并且在sigmaplot中计算ec50值。列出所有化合物的原始ec50值,并计算倍数差异(聚乙二醇化牛gcsf化合物与wtbst比较)。执行实验多次,以确定批内cv<20%且批间cv<30%。
实例3
可使用e9rs在琥珀密码子处使bsttrna装有paf,并且还可使用e9rs在琥珀密码子处使bsttrna装有paf3(有关paf3,例如参见图式)。并入paf3后,paf3可在还原条件下转变成paf2。在本实例中,再折叠前paf3转变成paf2,且此转变允许基于还原烷基化的聚乙二醇化。这一转变是在猪促生长素存在下进行,并且评价在包括包涵体洗涤、预先聚乙二醇化和溶解的3个步骤中paf3还原成paf2的情况。在包涵体洗涤步骤中,向ib洗涤缓冲液中添加不同浓度的dtt。在最终洗涤缓冲液中使用最高20mm的不同浓度的dtt。还原水平为约90%。在预先聚乙二醇化步骤中,培育是在4℃下进行,并使用0.1mm到0.5mmdtt浓度。在0.2mmdtt存在下o/n培育后,观察到95%和95%以上的高还原水平。在溶解步骤中,dtt浓度增加到10mm,并且再培育2小时(3小时总培育时间),且实现95%和95%以上高还原水平和高产率。由此得到的结果可见于图24中。在paf3还原成paf2后,进行聚乙二醇化。利用10%hoac将蛋白质降到4.0ph,缓冲液更换成20mmnaoac,并浓缩到约3.0mg/ml。添加nacnbh3达到5mm最终浓度,并按以下比率(peg:蛋白质)添加peg-醛:0.9:1、1:1、1.5:1。在室温下混合/培育物质,并在1小时、2小时、3小时、4小时和24小时时借助sds-page进行分析,且由此得到的结果可见于图25中。
实例4
在本实例中,用f92paf-30k聚乙二醇化bst处理3组乳牛。经过处理的奶牛是产犊并处于哺乳期的奶牛。第1组接受0mg/kg;第2组接受1mg/kg;且第3组接受5mg/kg,并在第1天以及接下来时间给予奶牛一次以测定对产奶量的影响,且预期经处理组的产奶量高于负对照组。
实例5
在本实例中,用f92paf-30k聚乙二醇化bst处理3组乳牛。奶牛的处理是在产犊前进行。第1组接受0mg/kg;第2组接受1mg/kg;且第3组接受5mg/kg,并在第1天以及接下来时间给予奶牛一次以测定对产奶量的影响,且预期经处理组的产奶量高于负对照组。
实例6
本实例详述大肠杆菌中包括非天然编码氨基酸的人生长激素(hgh)多肽的克隆和表达。本实例还描述一种评估经修饰hgh多肽的生物活性的方法。
克隆hgh和其片段的方法详细描述于美国专利第4,601,980号、第4,604,359号、第4,634,677号、第4,658,021号、第4,898,830号、第5,424,199号和第5,795,745号中,所述专利以引用的方式并入本文中。编码全长hgh或缺乏n末端信号序列的成熟形式hgh的cdna分别以seqidno:21和seqidno:22显示。
使用引入的包含正交trna(o-trna)和正交氨酰基trna合成酶(o-rs)的翻译系统表达含有非天然编码氨基酸的hgh。o-rs优先利用非天然编码氨基酸将o-trna氨酰基化。接着,翻译系统对编码的选择密码子反应而将非天然编码氨基酸插入hgh中。
用含有经修饰hgh基因和正交氨酰基trna合成酶/trna对(对所需非天然编码氨基酸具特异性)的质粒转化大肠杆菌,从而将非天然编码氨基酸位点特异性地并入hgh多肽中。37℃下在含有介于0.01到100mm之间的特定非天然编码氨基酸的培养基中生长的经转化大肠杆菌以高保真度和效率表达经修饰hgh。大肠杆菌宿主细胞以包涵体或聚集体的形式产生his标记的含有非天然编码氨基酸的hgh。溶解聚集体,并在变性条件下于6m盐酸胍中亲和纯化。通过在4℃下于50mmtris-hcl(ph8.0)、40μmcuso4和2%(w/v)月桂酰肌氨酸钠(sarkosyl)中透析过夜,来执行再折叠。随后针对20mmtris-hcl(ph8.0)、100mmnacl、2mmcacl2透析所述材料,接着去除his标签。参见波塞尔(boissel)等人,生物化学杂志,(1993)268:15983-93。所属领域技术人员已知hgh的纯化方法并且通过sds-page、蛋白质印迹分析或电喷雾离子阱质谱等加以确定。
图1是纯化的hgh多肽的sds-page。使用probond镍螯合树脂(英杰公司,加利福尼亚州卡尔斯巴德),经由制造商提供的标准his标记的蛋白质纯化程序,随后利用阴离子交换柱,接着装载到凝胶上,来纯化his标记的突变hgh蛋白。泳道1显示分子量标记物,且泳道2表示未并入非天然氨基酸的n-hishgh。泳道3到10含有n-hishgh突变体,其分别在以下各位置包含非天然氨基酸对乙酰基苯丙氨酸:y35、f92、y111、g131、r134、k140、y143和k145。
为进一步评估经修饰hgh多肽的生物活性,使用测量hgh与其受体相互作用的下游标记物的分析。hgh与其内源产生的受体相互作用引起人类im-9淋巴细胞细胞系中转录家族成员stat5的信号转导子和活化子的酪氨酸磷酸化。stat5的两种形式stat5a和stat5b是从im-9cdna文库鉴别。例如参见卡兹霍尔等人,生物化学杂志,(1996)10(5):508-518。im-9细胞上的人生长激素受体对人生长激素具选择性,因为大鼠生长激素和人催乳激素都无法引起可检测的stat5磷酸化。重要的是,大鼠ghr(l43r)胞外域和带有g120r的hgh竞争有效对抗hgh刺激的pstat5磷酸化。
用本发明hgh多肽刺激im-9细胞。人im-9淋巴细胞购自atcc(弗吉尼亚州马纳萨斯)并在补充有丙酮酸钠、青霉素(penicillin)、链霉素(streptomycin)(英杰公司,圣地亚哥卡尔斯巴德(carlsbad,sandiego))和10%热灭活胎牛血清(海康公司(hyclone),犹他州罗根市(logan,ut))的rpmi1640中生长。使im-9细胞在分析培养基(无酚红rpmi、10mmhepes、1%热灭活木炭/葡聚糖处理的fbs、丙酮酸钠、青霉素和链霉素)中饥饿过夜,随后在37℃下用12点剂量范围的hgh多肽刺激10分钟。用1%甲醛固定刺激的细胞,随后在冰上用90%冰冷的甲醇渗透1小时。通过在室温下用一次磷酸化stat5抗体(信号传导技术公司(cellsignalingtechnology),马萨诸塞州贝弗利(beverly,ma))胞内染色30分钟,随后用pe结合的二次抗体进行胞内染色,以此检测stat5磷酸化的水平。样品获取是在facs阵列上进行,并在flowjo软件(树星公司(treestarinc.),俄勒冈州阿什兰德(ashland,or))上分析获取的数据。根据利用sigmaplot绘制的平均荧光强度(mfi)针对蛋白质浓度的剂量反应曲线得到ec50值。
下表3概述利用突变hgh多肽产生的im-9数据。如所述,利用人im-9细胞测试在不同位置具有非天然氨基酸的各种hgh多肽。具体说来,图7图a显示his标记的hgh多肽的im-9数据,且图7图b显示his标记的包含非天然氨基酸对乙酰基-苯丙氨酸取代y143的hgh的im-9数据。使用相同分析来评估包含非天然氨基酸的聚乙二醇化hgh多肽的生物活性。
实例7
引入含羰基氨基酸和与含氨氧基peg的后续反应
本实例描述一种产生bst多肽的方法,所述bst多肽并有含酮非天然编码氨基酸,这一氨基酸随后将与mw为约5,000的含氨氧基peg反应。用具有以下结构的非天然编码氨基酸分别取代以下各残基:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,蛋白质的羧基末端),以及其任何组合(seqidno:1或seqidno:2):
修饰后,就使包含含羰基氨基酸的bst多肽变异体与以下形式的含氨氧基peg衍生物反应:
r-peg(n)-o-(ch2)n-o-nh2
其中r为甲基,n为3且n为约5,000mw。使以10mg/ml溶解于25mmmes(西格玛化学品公司,密苏里州圣路易斯)(ph6.0)、25mmhepes(西格玛化学品公司,密苏里州圣路易斯)(ph7.0)中或者溶解于10mm乙酸钠(西格玛化学品公司,密苏里州圣路易斯)(ph4.5)中的含对乙酰基苯丙氨酸的经纯化bst与10到100倍过量的含氨氧基peg反应,随后在室温下搅拌10到16小时(杰克斯(jencks,w.j.),美国化学协会杂志,1959,81,第475页)。随后将peg-bst稀释于适当缓冲液中以便立即进行纯化和分析。
实例8
由羟胺基经由酰胺键与peg连接组成的与peg的结合物
使用实例3中所述的程序将具有以下结构的peg试剂与含酮非天然编码氨基酸偶合:
r-peg(n)-o-(ch2)2-nh-c(o)(ch2)n-o-nh2
其中r=甲基,n=4且n为约20,000mw。反应条件、纯化条件和分析条件如实例3中所述。
实例9
将两种不同的非天然编码氨基酸引入bst多肽中
本实例描述一种产生在以下残基中两个位置并有包含酮官能团的非天然编码氨基酸的bst多肽的方法:1位前(即,n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,所述蛋白质的羧基末端),以及其任何组合(seqidno:1或seqidno:2)。如实例1和2中所述制备bst多肽,但在核酸内两个不同位点引入选择密码子。
实例10
bst多肽与含酰肼peg结合以及随后原位还原
根据实例2和3中所述的程序制备并有含羰基氨基酸的bst多肽。修饰后,就将具有以下结构的含酰肼peg与bst多肽结合:
r-peg(n)-o-(ch2)2-nh-c(o)(ch2)n-x-nh-nh2
其中r=甲基,n=2且n=10,000mw,且x为羰基(c=o)。使以0.1到10mg/ml溶解于25mmmes(西格玛化学品公司,密苏里州圣路易斯)(ph6.0)、25mmhepes(西格玛化学品公司,密苏里州圣路易斯)(ph7.0)中或者溶解于10mm乙酸钠(西格玛化学品公司,密苏里州圣路易斯)(ph4.5)中的含对乙酰基苯丙氨酸的经纯化b-gcsf与1到100倍过量的含酰肼peg反应,且通过添加溶解于h2o中的1mnacnbh3(西格玛化学品公司,密苏里州圣路易斯)储备液达到10到50mm最终浓度,来原位还原相应腙。所述反应在4℃到室温下于暗处进行18到24小时。通过添加约ph7.6的1mtris(西格玛化学品公司,密苏里州圣路易斯)达到50mm的最终tris浓度或者稀释于适当缓冲液中来停止反应以供立即纯化。
实例11
将含炔氨基酸引入bst多肽中并用mpeg-叠氮化物衍生化
用以下非天然编码氨基酸各自取代以下残基:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,在蛋白质的羧基末端),以及其任何组合(seqidno:1、seqidno:2):
用于将对炔丙基-酪氨酸位点特异性并入bst中的序列可为上文实例2中所述的seqidno:1或2、seqidno:3(muttrna,詹氏甲烷球菌
将以0.1到10mg/ml溶解于pb缓冲液(100mm磷酸钠、0.15mnacl,ph=8)中的含炔丙基-酪氨酸的经纯化bst和10到1000倍过量的含叠氮基peg添加到反应混合物中。随后将催化量的cuso4和cu线添加到反应混合物中。培育混合物(包括(但不限于)在室温或37℃下约4小时或者在4℃下过夜)后,添加h2o并使混合物滤过透析膜。借助于包括(但不限于)类似于实例3中所述的程序分析添加的样本。
在本实例中,peg将具有以下结构:
r-peg(n)-o-(ch2)2-nh-c(o)(ch2)n-n3
其中r为甲基,n为4且n为10,000mw。
实例12
用炔丙基酪氨酸取代bst多肽中的较大的疏水性氨基酸
用以下实例7中所述的非天然编码氨基酸取代bst中以下一个区域内存在的phe、trp或tyr残基:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175、176、177、178、179、180、181、182、183、184、185、186、187、188、189、190、191、192(即,在蛋白质的羧基末端),以及其任何组合(seqidno:1,或seqidno:2中的相应氨基酸,或者另一bst多肽中的相应氨基酸):
修饰后,将peg与包含含炔氨基酸的bst多肽变异体连接。peg将具有以下结构:
me-peg(n)-o-(ch2)2-n3
且偶合程序将遵循实例7中的程序。这将产生包含非天然编码氨基酸的bst多肽变异体,所述非天然编码氨基酸与天然存在的较大疏水性氨基酸中的一者大致等同排列且在所述多肽内的不同位点经peg衍生物修饰。
实例13
产生由一个或一个以上peg连接子分隔的bst多肽均二聚体、杂二聚体、均多聚体或杂多聚体
使实例7中制得的含炔bst多肽变异体与以下形式的双官能peg衍生物反应:
n3-(ch2)n-c(o)-nh-(ch2)2-o-peg(n)-o-(ch2)2-nh-c(o)-(ch2)n-n3
其中n为4,且peg的平均mw为约5,000;由此产生两个bst分子经由peg物理分隔的相应bst多肽均二聚体。可以用类似方式使bst多肽与一种或一种以上其它多肽偶合,以形成杂二聚体、均多聚体或杂多聚体。偶合、纯化和分析如实例7和3中般执行。
实例14
糖部分与bst多肽偶合
如实例3中所述,用以下非天然编码氨基酸取代一个以下残基:1位前(即,在n末端)、1、2、3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139、140、141、142、143、144、145、146、147、148、149、150、151、152、153、154、155、156、157、158、159、160、161、162、163、164、165、166、167、168、169、170、171、172、173、174、175(即,在蛋白质的羧基末端),以及其任何组合(seqidno:1,或seqidno:2的相应氨基酸,或者另一bst多肽中的相应氨基酸)。
修饰后,使包含含羰基氨基酸的bst多肽变异体与n-乙酰基氨基葡糖(glcnac)的β连接的氨氧基类似物反应。将bst多肽变异体(10mg/ml)和氨氧基糖(21mm)混入100mm乙酸钠缓冲水溶液(ph5.5)中,并在37℃下培育7到26小时。通过在环境温度下,将结合糖的bst多肽(5mg/ml)与udp-半乳糖(16mm)和β-1,4-半乳糖基转移酶(0.4个单位/毫升)在150mmhepes缓冲液(ph7.4)中培育48小时,在酶作用下,将第二糖与第一糖偶合(斯查布奇尔(schanbacher)等人,生物化学杂志,1970,245,5057-5061)。
实例15
产生聚乙二醇化bst多肽拮抗剂
如实例3中所述,用以下非天然编码氨基酸取代残基,包括(但不限于)bst受体结合所涉及的残基。
修饰后,使包含含羰基氨基酸的bst多肽变异体与以下形式的含氨氧基peg衍生物反应:
r-peg(n)-o-(ch2)n-o-nh2
其中r为甲基,n为4且n为20,000mw;由此产生包含非天然编码氨基酸且在多肽的单一位点经peg衍生物修饰的b-gcsf多肽拮抗剂。偶合、纯化和分析都是如实例3中般执行。
实例16
产生直接连接bst分子的bst多肽均二聚体、杂二聚体、均多聚体或杂多聚体
可以将包含含炔氨基酸的bst多肽变异体与包含含叠氮基氨基酸的另一bst多肽变异体直接偶合。可以用类似方式使bst多肽变异体与一种或一种以上其它多肽偶合,形成杂二聚体、均多聚体或杂多聚体。偶合、纯化和分析如实例3、6和7中般执行。
实例17
使聚烷二醇(p-oh)与烷基卤化物(a)反应,形成醚(b)。在这些化合物中,n为0到9的整数,且r′可为直链或支链、饱和或不饱和的c1到c20烷基或杂烷基。r′也可以为c3到c7饱和或不饱和环烷基或环杂烷基、经取代或未经取代的芳基或杂芳基,或经取代或未经取代的烷芳基(烷基为c1到c20饱和或不饱和烷基)或杂烷芳基。通常,peg-oh是分子量为800到40,000道尔顿(da)的聚乙二醇(peg)或单甲氧基聚乙二醇(mpeg)。
实例18
mpeg-oh+br-ch2-c≡ch→mpeg-o-ch2-c≡ch
用含nah(12mg,0.5mmol)的thf(35ml)处理分子量为20,000da的mpeg-oh(mpeg-oh20kda;2.0g,0.1mmol;新日生物技术公司(sunbio))。随后将溶解于二甲苯中成为80重量%溶液的炔丙基溴溶液(0.56ml,5mmol,50当量,阿尔德里奇公司)和催化量的ki添加到溶液中,并将所得混合物加热到回流,保持2小时。接着添加水(1ml),并在真空下去除溶剂。向残余物中添加ch2cl2(25ml),并分离有机层,用无水na2so4干燥,并将体积减少到约2ml。将此ch2cl2溶液逐滴添加到乙醚(150ml)中。收集得到的沉淀,用数份冷乙醚洗涤,并干燥,得到炔丙基-o-peg。
实例19
mpeg-oh+br-(ch2)3-c≡ch→mpeg-o-(ch2)3-c≡ch
用含nah(12mg,0.5mmol)的thf(35ml)处理分子量为20,000da的mpeg-oh(mpeg-oh20kda;2.0g,0.1mmol;新日生物技术公司)。随后,将50当量5-溴-1-戊炔(0.53ml,5mmol,阿尔德里奇公司)和催化量的ki添加到混合物中。将所得混合物加热到回流,保持16小时。接着添加水(1ml),并在真空下去除溶剂。向残余物中添加ch2cl2(25ml),并分离有机层,用无水na2so4干燥,并将体积减少到约2ml。将此ch2cl2溶液逐滴添加到乙醚(150ml)中。收集得到的沉淀,用数份冷乙醚洗涤,并干燥,得到相应的炔。在类似反应中可以使用5-氯-1-戊炔。
实例20
产生mpeg-q-ch2-c6h4o-ch2-c□ch
(1)m-hoch2c6h4oh+naoh+br-ch2-c≡ch→m-hoch2c6h4o-ch2-c≡ch
(2)m-hoch2c6h4o-ch2-och+mscl+n(et)3→m-msoch2c6h4o-ch2-c≡ch
(3)m-msoch2c6h4o-ch2-c≡ch+libr→m-br-ch2c6h4o-ch2-c≡ch
(4)mpeg-oh+m-br-ch2c6h4o-ch2-c≡ch→mpeg-o-ch2-c6h4o-ch2-c≡ch
向3-羟基苯甲醇(2.4g,20mmol)于thf(50ml)和水(2.5ml)中的溶液中首先添加粉末状氢氧化钠(1.5g,37.5mmol),随后添加溶解于二甲苯中成为80重量%溶液的炔丙基溴溶液(3.36ml,30mmol)。在回流下加热反应混合物6小时。向混合物中添加10%柠檬酸(2.5ml),并在真空下去除溶剂。用乙酸乙酯(3×15ml)萃取残余物,并用饱和nacl溶液(10ml)洗涤合并的有机层,用mgso4干燥并浓缩,得到3-炔丙氧基苯甲醇。
在0℃下,将甲烷磺酰氯(2.5g,15.7mmol)和三乙胺(2.8ml,20mmol)添加到化合物3(2.0g,11.0mmol)于ch2c12中的溶液中,并将反应物放入冰箱中,保持16小时。常规处理后,得到浅黄色油状的甲磺酸酯。将此油状物(2.4g,9.2mmol)溶解于thf(20ml)中,并添加libr(2.0g,23.0mmol)。将反应混合物加热到回流,保持1小时,随后将其冷却到室温。向混合物中添加水(2.5ml),并在真空下去除溶剂。用乙酸乙酯(3×15ml)萃取残余物,并用饱和nacl溶液(10ml)洗涤合并的有机层,用无水na2so4干燥并浓缩,得到所需溴化物。
将mpeg-oh20kda(1.0g,0.05mmol,新日生物技术公司)溶解于thf(20ml)中,并在冰浴中冷却溶液。在用力搅拌下,经数分钟添加nah(6mg,0.25mmol),随后添加由上述获得的溴化物(2.55g,11.4mmol)和催化量的ki。移开冷却浴,并将所得混合物加热到回流,保持12小时。将水(1.0ml)添加到混合物中,并在真空下去除溶剂。向残余物中添加ch2cl2(25ml),并分离有机层,用无水na2so4干燥,并将体积减少到约2ml。逐滴添加到乙醚溶液(150ml)中,得到白色沉淀,收集沉淀,得到peg衍生物。
实例21
mpeg-nh2+x-c(o)-(ch2)n-c≡cr'→mpeg-nh-c(o)-(ch2)n-c≡cr'
也可通过使含末端官能团的聚(乙二醇)聚合物与上文所示的含有炔官能团的反应分子偶合,获得含末端炔的聚(乙二醇)聚合物。n介于1与10之间。r'可为h或c1到c4小烷基。
实例22
制备mpeg-nh-c(o)-(ch2)2-c≡ch
(1)ho2c-(ch2)2-c≡ch+nhs+dcc→nhso-c(o)-(ch2)2-c≡ch
(2)mpeg-nh2+nhso-c(o)-(ch2)2-c≡ch→mpeg-nh-c(o)-(ch2)2-c≡ch
将4-戊炔酸(2.943g,3.0mmol)溶解于ch2cl2(25ml)中。添加n-羟基琥珀酰亚胺(3.80g,3.3mmol)和dcc(4.66g,3.0mmol),并在室温下搅拌溶液过夜。不经进一步纯化即将所得粗nhs酯7用于下一反应中。
将分子量为5,000da的mpeg-nh2(mpeg-nh2,1g,新日生物技术公司)溶解于thf(50ml)中,并将混合物冷却到4℃。在用力搅拌下,分数份添加nhs酯7(400mg,0.4mmol)。混合物搅拌3小时,同时升温到室温。接着添加水(2ml),并在真空下去除溶剂。向残余物中添加ch2cl2(50ml),并分离有机层,用无水na2so4干燥,并将体积减少到约2ml。将此ch2cl2溶液逐滴添加到乙醚(150ml)中。收集所得沉淀,并在真空中干燥。
实例23
制备聚(乙二醇)的甲烷磺酸酯或甲磺酸酯
本实例提供聚(乙二醇)甲烷磺酰基酯(也可称为聚(乙二醇)甲烷磺酸酯或甲磺酸酯)的制备。可以用类似程序制备相应的甲苯磺酸酯和卤化物。
mpeg-oh+ch3so2cl+n(et)3→mpeg-o-so2ch3→mpeg-n3
在氮气下,将mpeg-oh(mw=3,400,25g,10mmol)在150ml甲苯中共沸蒸馏2小时,并将溶液冷却到室温。将40ml无水ch2cl2和2.1ml无水三乙胺(15mmol)添加到溶液中。在冰浴中冷却溶液,并逐滴添加1.2ml蒸馏过的甲烷磺酰氯(15mmol)。室温下,在氮气下搅拌溶液过夜,并通过添加2ml无水乙醇中止反应。在真空下蒸发混合物,去除溶剂(主要是除甲苯外的溶剂),过滤,再次在真空下浓缩,随后沉淀到100ml乙醚中。用数份冷乙醚洗涤滤液,并在真空中干燥,得到甲磺酸酯。
将甲磺酸酯(20g,8mmol)溶解于75mlthf中,并将溶液冷却到4℃。向冷却的溶液中添加叠氮化钠(1.56g,24mmol)。在氮气下,将反应物加热到回流,保持2小时。随后蒸发溶剂,并用ch2cl2(50ml)稀释残余物。用nacl溶液洗涤有机部分,并用无水mgso4干燥。将体积减少到20ml,并通过添加到150ml冷无水乙醚中沉淀产物。
实例24
产生mpeg-o-ch2-c6h4-n3
(1)n3-c6h4-co2h→n3-c6h4ch2oh
(2)n3-c6h4ch2oh→br-ch2-c6h4-n3
(3)mpeg-oh+br-ch2-c6h4-n3→mpeg-o-ch2-c6h4-n3
可使用美国专利5,998,595(以引用的方式并入本文中)中所述的方法制备4-叠氮基苯甲醇。在0℃下,将甲烷磺酰氯(2.5g,15.7mmol)和三乙胺(2.8ml,20mmol)添加到4-叠氮基苯甲醇(1.75g,11.0mmol)的ch2c12溶液中,并将反应物放入冰箱中,保持16小时。常规处理后,得到浅黄色油状的甲磺酸酯。将此油状物(9.2mmol)溶解于thf(20ml)中,并添加libr(2.0g,23.0mmol)。将反应混合物加热到回流,保持1小时,随后将其冷却到室温。向混合物中添加水(2.5ml),并在真空下去除溶剂。用乙酸乙酯(3×15ml)萃取残余物,并用饱和nacl溶液(10ml)洗涤合并的有机层,用无水na2so4干燥并浓缩,得到所需溴化物。
用含nah(12mg,0.5mmol)的thf(35ml)处理mpeg-oh20kda(2.0g,0.1mmol,新日生物技术公司),并将溴化物(3.32g,15mmol)与催化量的ki一起添加到混合物中。将所得混合物加热到回流,保持12小时。将水(1.0ml)添加到混合物中,并在真空下去除溶剂。向残余物中添加ch2cl2(25ml),并分离有机层,用无水na2so4干燥,并将体积减少到约2ml。逐滴添加到乙醚溶液(150ml)中,得到沉淀,收集沉淀,得到mpeg-o-ch2-c6h4-n3。
实例25
nh2-peg-o-ch2ch2co2h+n3-ch2ch2co2nhs→n3-ch2ch2-c(o)nh-peg-o-ch2ch2co2h
将nh2-peg-o-ch2ch2co2h(mw3,400da,2.0g)溶解于饱和nahco3水溶液(10ml)中,并将溶液冷却到0℃。在用力搅拌下,添加丙酸3-叠氮基-1-n-羟基琥珀酰亚胺酯(5当量)。3小时后,添加20mlh2o,并在室温下,再搅拌混合物45分钟。用0.5nh2so4将ph值调到3,并添加nacl达到约15重量%浓度。用ch2cl2(100ml×3)萃取反应混合物,用na2so4干燥并浓缩。用冷乙醚沉淀后,通过过滤收集产物,并在真空下干燥,得到ω-羧基-叠氮peg衍生物。
实例26
mpeg-oms+hc≡cli→mpeg-o-ch2-ch2-c≡c-h
用力搅拌下,向如此项技术中所知般制备并冷却到-78℃的乙炔锂(4当量)于thf中的溶液中逐滴添加mpeg-oms溶解于thf中得到的溶液。3小时后,使反应物升温到室温,并通过添加1ml丁醇中止反应。随后添加20mlh2o,并在室温下,再搅拌混合物45分钟。用0.5nh2so4将ph值调到3,并添加nacl达到约15重量%浓度。用ch2cl2(100ml×3)萃取反应混合物,用na2so4干燥并浓缩。用冷乙醚沉淀后,通过过滤收集产物,并在真空下干燥,得到1-(丁-3-炔基氧基)-甲氧基聚乙二醇(mpeg)。
实例27
并入含叠氮基氨基酸和含乙炔的氨基酸
可以使用以下文献中所述的方法,将含叠氮基氨基酸和含乙炔的氨基酸位点选择性地并入蛋白质中:l.王等人(2001),科学292:498-500;j.w.秦(j.w.chin)等人,科学301:964-7(2003);j.w.秦等人(2002),美国化学协会杂志124:9026-9027;j.w.秦和p.g.查鲁兹(p.g.schultz),(2002),化学与生物化学(chembiochem)3(11):1135-1137;j.w.秦等人(2002),美国国家科学院院刊,99:11020-11024;以及l.王和p.g.查鲁兹,(2002),化学通讯,1:1-11。在并入氨基酸后,在37℃下,于2mmpeg衍生物、1mmcuso4和约1mg铜线存在下,利用含0.01mm蛋白质的磷酸盐缓冲液(phosphatebuffer,pb;ph8)进行环加成反应4小时。
实例28
合成对乙酰基-d,l-苯丙氨酸(paf)和m-peg-羟胺衍生物
使用先前于张z.,史密斯b.a.c(smith,b.a.c),王l.,布洛克a.(brock,a.),曹c(cho,c)和查鲁兹p.g.,生物化学,(2003)42,6735-6746中描述的程序,合成外消旋paf。
为了合成m-peg-羟胺衍生物,完成以下程序。在室温(rt)下,搅拌(n-t-boc-氨氧基)乙酸(0.382g,2.0mmol)和1,3-二异丙基碳化二亚胺(0.16ml,1.0mmol)于二氯甲烷(dcm,70ml)中的溶液1小时,向其中添加甲氧基-聚乙二醇胺(m-peg-nh2,7.5g,0.25mmol,mt.30k,来自拜耳维特公司(biovectra))和二异丙基乙胺(0.1ml,0.5mmol)。在室温下搅拌反应物48小时,随后浓缩到约100ml。将混合物逐滴添加到冷乙醚(800ml)中。经t-boc保护的产物沉淀析出,并通过过滤收集,用乙醚洗涤3次,每次100ml。通过将其再溶解于dcm(100ml)中,并沉淀于乙醚(800ml)中2次,来进行进一步纯化。真空干燥产物,得到7.2g(96%),借助于nmr和茚三酮测试确定。
在0℃下,在50%tfa/dcm(40ml)中保持1小时,随后在室温下保持1.5小时,来脱除上文获得的经保护产物(7.0g)的保护基boc。真空中去除大部分tfa后,通过向残余物中添加4nhcl的二噁烷溶液(1ml),将羟胺衍生物的tfa盐转化成hcl盐。将沉淀溶解于dcm(50ml)中,并再沉淀于乙醚(800ml)中。通过过滤收集终产物(6.8g,97%),用乙醚洗涤3次,每次100ml,真空干燥,在氮气下储存。使用相同程序合成其它peg(5k,20k)羟胺衍生物。
实例29
聚乙二醇化bst的体外和体内活性
对小鼠或大鼠投予peg-bst、未修饰的bst和缓冲溶液。结果显示,在每只小鼠使用相同剂量时,如由中性粒细胞数量显著增加和白细胞计数最大值移位所指示,与未修饰的bst相比较,本发明的聚乙二醇化bst具有优良的活性和较长的半衰期。
药物动力学分析
经静脉内或皮下途径投予小鼠本发明的bst多肽。在给药前以及给药后各时间点抽取动物血液。收集每个样品的血浆,并通过放射免疫分析进行分析。可计算消除半衰期,并将本发明的包含非天然编码氨基酸的bst多肽与野生型bst或各种形式的bst多肽相比较。类似地,可以将本发明的bst多肽投予猕猴。在给药前以及给药后各时间点抽取动物的血液。收集每个样品的血浆,并通过放射免疫分析进行分析。
可以将本发明多肽投予动物疾病模型。可执行的动物研究涉及经溶血性巴斯德氏菌激发的牛、用细菌激发乳腺/乳腺炎激发(肺炎克氏杆菌)的牛。可以执行其它研究来评估牛呼吸系统疾病的控制、发生率和持续时间,或大肠杆菌型乳腺炎的预防。所属领域技术人员已知评价动物健康、产奶量、中性粒细胞计数和其它参数的方法。可用于评价本发明bst多肽的其它模型包括(但不限于)感染或暴露于感染的动物模型,例如绿脓杆菌肺炎的仓鼠模型、白色念珠菌肾盂肾炎的大鼠模型、涉及新生驹的模型以及涉及生长的猪的模型。这些模型中有一些描述于美国专利第5,849,883号和wo89/10932中。所属领域技术人员已知例如此类的模型。
3h-胸苷分析。使用标准方法执行3h-胸苷分析。骨髓是从处死的雌性balbc小鼠或其它动物得到。短暂悬浮骨髓细胞,离心,并将其再悬浮于生长培养基中。将160μl含有约10,000个细胞的等分试样放入96孔微量滴定板的每一孔中。将经纯化g-csf类似物的样品(如上文所述制备)添加到每一孔中,并培育68小时。将氚化胸苷添加到孔中,并且再培育5小时。5小时培育时间后,采集细胞,过滤并充分漂洗。将滤膜添加到含有闪烁液的小瓶中。对β发射进行计数(lkbbetaplate闪烁计数器)。一式三份,分析标准品和类似物,并通过适当稀释对实质上落在标准曲线上方或下方的样品再次进行分析。结果报导为三次重复类似物数据的平均值相对于未改变的bst标准结果。
根据3h-胸苷并入量的增加,分析对人骨髓细胞增殖的诱导。利用聚蔗糖-泛影葡胺(ficoll-hypaque)(1.077g/ml,法玛西亚公司)对来自健康供体的人骨髓进行密度分割(densitycut),并将低密度细胞悬浮于含有10%胎牛血清和谷氨酰胺青霉素/链霉素(glutaminepen-strep)的伊斯科文培养基(iscove'smedium;gibco公司)中。随后,在96孔平底板中,在37℃下,于含有5%co2的空气中,将2×104个人骨髓细胞与对照培养基或者来源于大肠杆菌的重组bst物质培育2天。一式两份进行样品分析,且浓度在10,000倍范围内变化。随后,用0.5μci/孔3h-胸苷(新英格兰核公司(newenglandnuclear),马萨诸塞州波士顿(boston,mass.))使培养物脉冲4小时。如维努塔(venuta)等人,血液(blood),61,781(1983)中所述测量3h-胸苷的摄取。
wehi-3bd+分化诱导。如米特卡夫(metcalf),国际癌症杂志(int.j.cancer),25,225(1980)中所述,在半固体琼脂培养基中分析本发明的bst多肽诱导鼠类骨髓单核白血病细胞系wehi-3bd+分化的能力。在37℃下,于含有5%co2的空气中,将重组bst产物和培养基对照品与约60个wehi-3bd+细胞/孔培育7天。在24孔平底板中培育样品,且浓度在2000倍范围内变化。将集落归类为未分化的、部分分化的或完全分化的,并通过显微镜对细胞集落数量进行计数。
测量结合和未结合的bst和其变异体的体内半衰期。使用雄性斯普拉格-道来大鼠(约7周龄)。投药当天,测量每只动物的重量。可使用此项技术中已知的方法测定剂量,例如,在3只大鼠的尾静脉中分别按每公斤体重100μg静脉内注射未结合和结合的bst样品。注射后1分钟、30分钟、1小时、2小时、4小时、6小时和24小时,在co2麻醉下从每只大鼠抽取500μl血液。在室温下储存血样1.5小时,随后通过离心(4℃,18000×g,5分钟)分离出血清。在-80℃下储存血清样品,直到分析当天。在冰上解冻样品后,利用bst体外活性分析来定量血清样品中活性bst的量。
测量结合和未结合的bst和其变异体在健康大鼠中的体内生物活性。通过测量bst在spf斯普拉格道来大鼠中的体内生物作用,来评价结合和未结合的bst和其变异体的生物功效。到达当天,将大鼠随机分组,每组6只。使动物休息7天,其中排除处于不良状况或极端体重的个体。在休息期开始时,大鼠的重量范围为250-270g。
投药当天,使大鼠禁食16小时,随后皮下注射每公斤体重100μgbst或其变异体。将各bst样品注射到一组6只随机大鼠中。在给药前以及给药后6、12、24、36、48、72、96、120和144小时,从大鼠的尾静脉抽取经300μgedta稳定的血液的血样。分析血样的以下血液学参数:血红蛋白、红细胞计数、血细胞比容、平均细胞体积、平均细胞血红蛋白浓度、平均细胞血红蛋白、白细胞计数、分化的白细胞计数(中性粒细胞、淋巴细胞、嗜曙红细胞、嗜碱细胞、单核细胞)。根据这些测量结果,评价结合和未结合的bst以及其变异体的生物功效。
测量结合和未结合的bst和其变异体在患有化疗诱发的中性白细胞减少症的大鼠中的体内生物活性。本分析利用spf斯普拉格道来大鼠。到达当天,将大鼠随机分组,每组6只。使动物休息7天,其中排除处于不良状况或极端体重的个体。在休息期开始时,大鼠的重量范围为250-270g。
投予bst样品前24小时,对大鼠腹膜内注射每公斤体重50mg环磷酰胺(cyclophosphamide,cpa)以诱发中性粒细胞减少症,来模拟由抗癌化疗引起的中性粒细胞减少症。第0天,皮下注射每公斤体重100μgbst或其变异体。将各bst样品注射到一组6只随机大鼠中。在给药前以及给药后6、12、24、36、48、72、96、120、144和168小时,从大鼠的尾静脉抽取经300μledta稳定的血液的血样。分析血样的以下血液参数:血红蛋白、红细胞计数、血细胞比容、平均细胞体积、平均细胞血红蛋白浓度、平均细胞血红蛋白、白细胞计数、分化的白细胞计数(中性粒细胞、淋巴细胞、嗜曙红细胞、嗜碱细胞、单核细胞)。根据这些测量结果,评价结合和未结合的bst以及其变异体的生物功效。
应了解,本文中所述的实例和实施例仅出于说明目的,并且将对所属领域技术人员提出根据所述实例和实施例的各种修改或改变,且所述修改和改变都将包括在本申请案的精神和范围以及所附权利要求书的范围内。本申请案中引用的所有出版物、专利、专利申请案和/或其它文献都以全文引用的方式并入本文中用于所有目的,引用的程度就如同将各个别出版物、专利、专利申请案和/或其它文献个别地以引用的方式并入本文中用于所有目的一般。
表4:引用的序列。
序列表
<110>haysputnam,anna-mariaa.
<120>经过修饰的牛促生长素多肽和其用途
<130>ambx-0166.00pct
<150>61/288,764
<151>2009-12-21
<160>19
<170>patentinversion3.4
<210>1
<211>191
<212>prt
<213>牛
<400>1
alapheproalametserleuserglyleuphealaasnalavalleu
151015
argalaglnhisleuhisglnleualaalaaspthrphelysgluphe
202530
gluargthrtyrileprogluglyglnargtyrserileglnasnthr
354045
glnvalalaphecysphesergluthrileproalaprothrglylys
505560
asnglualaglnglnlysseraspleugluleuleuargileserleu
65707580
leuleuileglnsertrpleuglyproleuglnpheleuserargval
859095
phethrasnserleuvalpheglythrseraspargvaltyrglulys
100105110
leulysaspleuglugluglyileleualaleumetarggluleuglu
115120125
aspglythrproargalaglyglnileleulysglnthrtyrasplys
130135140
pheaspthrasnmetargseraspaspalaleuleulysasntyrgly
145150155160
leuleusercysphearglysaspleuhislysthrgluthrtyrleu
165170175
argvalmetlyscysargargpheglyglualasercysalaphe
180185190
<210>2
<211>191
<212>prt
<213>牛
<400>2
alapheproalametserleuserglyleuphealaasnalavalleu
151015
argalaglnhisleuhisglnleualaalaaspthrphelysgluphe
202530
gluargthrtyrileprogluglyglnargtyrserileglnasnthr
354045
glnvalalaphecysphesergluthrileproalaprothrglylys
505560
asnglualaglnglnlysseraspleugluleuleuargileserleu
65707580
leuleuileglnsertrpleuglyproleuglnpheleuserargval
859095
phethrasnserleuvalpheglythrseraspargvaltyrglulys
100105110
leulysaspleuglugluglyileleualaleumetarggluvalglu
115120125
aspglythrproargalaglyglnileleulysglnthrtyrasplys
130135140
pheaspthrasnmetargseraspaspalaleuleulysasntyrgly
145150155160
leuleusercysphearglysaspleuhislysthrgluthrtyrleu
165170175
argvalmetlyscysargargpheglyglualasercysalaphe
180185190
<210>3
<211>77
<212>dna
<213>詹氏甲烷球菌
<400>3
ccggcggtagttcagcagggcagaacggcggactctaaatccgcatggcgctggttcaaa60
tccggcccgccggacca77
<210>4
<211>88
<212>dna
<213>人工序列
<220>
<223>优化的琥珀抑制trna
<400>4
cccagggtagccaagctcggccaacggcgacggactctaaatccgttctcgtaggagttc60
gagggttcgaatcccttccctgggacca88
<210>5
<211>89
<212>dna
<213>人工序列
<220>
<223>优化的agga移码抑制trna
<400>5
gcgagggtagccaagctcggccaacggcgacggacttcctaatccgttctcgtaggagtt60
cgagggttcgaatccctcccctcgcacca89
<210>6
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-l-苯丙氨酸的氨酰基trna合成酶
<400>6
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralagly
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserthrpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnthrtyrtyr
145150155160
tyrleuglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>7
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对苯甲酰基-l-苯丙氨酸的氨酰基trna合成酶
<400>7
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralagly
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserserpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnthrserhis
145150155160
tyrleuglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>8
<211>305
<212>prt
<213>人工序列
<220>
<223>用于并入炔丙基-苯丙氨酸的氨酰基trna合成酶
<400>8
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaala
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserpropheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnalailetyr
145150155160
leualavalaspvalalavalglyglymetgluglnarglysilehis
165170175
metleualaarggluleuleuprolyslysvalvalcysilehisasn
180185190
provalleuthrglyleuaspglygluglylysmetserserserlys
195200205
glyasnpheilealavalaspaspserproglugluileargalalys
210215220
ilelyslysalatyrcysproalaglyvalvalgluglyasnproile
225230235240
metgluilealalystyrpheleuglutyrproleuthrilelysarg
245250255
proglulyspheglyglyaspleuthrvalasnsertyrglugluleu
260265270
gluserleuphelysasnlysgluleuhisprometaspleulysasn
275280285
alavalalaglugluleuilelysileleugluproilearglysarg
290295300
leu
305
<210>9
<211>305
<212>prt
<213>人工序列
<220>
<223>用于并入炔丙基-苯丙氨酸的氨酰基trna合成酶
<400>9
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaala
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserpropheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnileprotyr
145150155160
leuprovalaspvalalavalglyglymetgluglnarglysilehis
165170175
metleualaarggluleuleuprolyslysvalvalcysilehisasn
180185190
provalleuthrglyleuaspglygluglylysmetserserserlys
195200205
glyasnpheilealavalaspaspserproglugluileargalalys
210215220
ilelyslysalatyrcysproalaglyvalvalgluglyasnproile
225230235240
metgluilealalystyrpheleuglutyrproleuthrilelysarg
245250255
proglulyspheglyglyaspleuthrvalasnsertyrglugluleu
260265270
gluserleuphelysasnlysgluleuhisprometaspleulysasn
275280285
alavalalaglugluleuilelysileleugluproilearglysarg
290295300
leu
305
<210>10
<211>305
<212>prt
<213>人工序列
<220>
<223>用于并入炔丙基-苯丙氨酸的氨酰基trna合成酶
<400>10
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaala
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserlyspheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnalailetyr
145150155160
leualavalaspvalalavalglyglymetgluglnarglysilehis
165170175
metleualaarggluleuleuprolyslysvalvalcysilehisasn
180185190
provalleuthrglyleuaspglygluglylysmetserserserlys
195200205
glyasnpheilealavalaspaspserproglugluileargalalys
210215220
ilelyslysalatyrcysproalaglyvalvalgluglyasnproile
225230235240
metgluilealalystyrpheleuglutyrproleuthrilelysarg
245250255
proglulyspheglyglyaspleuthrvalasnsertyrglugluleu
260265270
gluserleuphelysasnlysgluleuhisprometaspleulysasn
275280285
alavalalaglugluleuilelysileleugluproilearglysarg
290295300
leu
305
<210>11
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>11
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralathr
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserasnpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnproleuhis
145150155160
tyrglnglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>12
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>12
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralathr
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserserpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnproleuhis
145150155160
tyrglnglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>13
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>13
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaleu
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserthrpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnprovalhis
145150155160
tyrglnglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>14
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>14
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralathr
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserserpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnproserhis
145150155160
tyrglnglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>15
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对乙酰基-苯丙氨酸的氨酰基trna合成酶
<400>15
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaleu
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserglupheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnglycyshis
145150155160
tyrargglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>16
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对乙酰基-苯丙氨酸的氨酰基trna合成酶
<400>16
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaleu
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserglupheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnglythrhis
145150155160
tyrargglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>17
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对乙酰基-苯丙氨酸的氨酰基trna合成酶
<400>17
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaala
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserglupheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnglyglyhis
145150155160
tyrleuglyvalaspvalilevalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>18
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>18
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralaala
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserargpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnvalilehis
145150155160
tyraspglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
<210>19
<211>306
<212>prt
<213>人工序列
<220>
<223>用于并入对叠氮基-苯丙氨酸的氨酰基trna合成酶
<400>19
metaspglupheglumetilelysargasnthrsergluileileser
151015
gluglugluleuarggluvalleulyslysaspglulysseralagly
202530
ileglyphegluproserglylysilehisleuglyhistyrleugln
354045
ilelyslysmetileaspleuglnasnalaglypheaspileileile
505560
leuleualaaspleuhisalatyrleuasnglnlysglygluleuasp
65707580
gluilearglysileglyasptyrasnlyslysvalpheglualamet
859095
glyleulysalalystyrvaltyrglyserthrpheglnleuasplys
100105110
asptyrthrleuasnvaltyrargleualaleulysthrthrleulys
115120125
argalaargargsermetgluleuilealaarggluaspgluasnpro
130135140
lysvalalagluvaliletyrproilemetglnvalasnthrtyrtyr
145150155160
tyrleuglyvalaspvalalavalglyglymetgluglnarglysile
165170175
hismetleualaarggluleuleuprolyslysvalvalcysilehis
180185190
asnprovalleuthrglyleuaspglygluglylysmetserserser
195200205
lysglyasnpheilealavalaspaspserproglugluileargala
210215220
lysilelyslysalatyrcysproalaglyvalvalgluglyasnpro
225230235240
ilemetgluilealalystyrpheleuglutyrproleuthrilelys
245250255
argproglulyspheglyglyaspleuthrvalasnsertyrgluglu
260265270
leugluserleuphelysasnlysgluleuhisprometaspleulys
275280285
asnalavalalaglugluleuilelysileleugluproilearglys
290295300
argleu
305
- 还没有人留言评论。精彩留言会获得点赞!