非生物胁迫耐性的植物和方法与流程
本发明涉及植物育种和遗传学领域,特别是,涉及用于提高植物耐诸如干旱和冷胁迫等非生物胁迫的重组dna构建体,以及用于调控植物开花期和/或抽穗期的重组dna构建体。
背景技术:
生物和非生物原因可以对植物产生胁迫,例如造成生物胁迫的原因包括病原菌感染、昆虫取食、一种植物对另一种植物的寄生,例如槲寄生;非生物胁迫包括诸如有效水分的过量或者不足、极限温度和诸如除草剂的合成化学药品。
非生物胁迫是造成世界范围内农作物减产的主要原因,造成主要农作物平均减产50%以上(boyer,j.s.(1982)science218:443-448;bray,e.a.等(2000)inbiochemistryandmolecularbiologyofplants,editedbybuchannan,b.b.等,amer.soc.plantbiol.,pp.1158-1249)。植物固着于地面,必须调整以适应周围的环境条件,这就导致了植物发育中基因调控、形态形成和新陈代谢的巨大可塑性。植物的适应和防御策略包括激活编码重要蛋白的基因,这些蛋白可以使植物适应或防御不同胁迫条件。
干旱(有效水分不足)是一种主要的非生物胁迫,限制了世界范围内农作物的生产。在植物生长发育阶段,将植物暴露于水分限制环境下,将会激活植物各种不同生理和发育变化。遗传学研究表明植物的耐旱性是一个由多基因调控的数量性状,分子标记辅助育种能够提高农作物的耐旱性。转基因途径提高农作物的耐旱性已经取得了很大进展(vinocurb.andaltmana.(2005)curr.opin.biotechnol.16:123-132;lawlordw.(2013)j.exp.bot.64:83-108)。
发明概述
本发明包括以下具体实施例:
在一个实施例中,本发明包括一种分离的多核苷酸,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:2、5、8、11、14、17、20或23的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:3、6、9、12、15、18、21或24的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或(d)核苷酸序列(a)、(b)或(c)的全长互补序列,其中过量表达所述多核苷酸能够提高植物的耐旱性。所述分离的多核苷酸包括seqidno:2、3、5、6、8、9、11、12、14、15、17、18、20、21、23或24所示的核苷酸序列;所述多肽包括seqidno:4、7、10、13、16、19、22或25所示的氨基酸序列。进一步的,过量表达所述多核苷酸提高植物在正常条件下的籽粒产量。改变所述多核苷酸在植物中的表达可以调控植物的开花时间,其中,增加所述多核苷酸的表达,促进植物早开花,降低所述多核苷酸的表达延长植物开花时间。过量表达所述多核苷酸提高植物的氮胁迫耐性或者提高nue。
在另一个实施例中,本发明包括一种重组dna构建体,所述重组dna构建体包括一种分离的多核苷酸和与其可操作的连接至少一个异源调控序列,其中所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5、6、8、9、11、12、14、15、17、18、20、21、23或24的序列一致性至少为85%;(b)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或者(c)核苷酸序列(a)或(b)的全长互补序列。
在另一个实施例中,本发明包括一种修饰的植物或种子,所述植物或种子包含编码icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1、dn-dtp7多肽的至少一种多核苷酸,与对照植物或种子相比,所述多核苷酸的表达是提高的,其中,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5、6、8、9、11、12、14、15、17、18、20、21、23或24的序列一致性至少为85%;(b)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或(c)核苷酸序列(a)或(b)的全长互补序列。
所述修饰的植物,在其基因组中包含一种重组dna构建体,所述重组dna构建体包含一种多核苷酸和与其可操作连接的至少一个异源调控元件,其中,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:2、3、5、6、8、9、11、12、14、15、17、18、20、21、23或24的序列一致性至少为85%;(b)一种多核苷酸序列,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或(c)核苷酸序列(a)或(b)的全长互补序列,其中,与对照植物相比,所述植物显示出增强的耐旱性,所述增强的耐旱性可以表现为在干旱条件下存活率增加、卷叶程度降低、种子结实率增加或籽粒产量增加。
所述植物包含修饰的调控元件用于增加内源多核苷酸的表达,其中,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:2、5、8、11、14、17、20或23的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:3、6、9、12、15、18、21或24的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或(d)核苷酸序列(a)、(b)或(c)的全长互补序列,其中,与对照植物相比,所述植物显示出增强的耐旱性。
与对照植物相比,所述植物在正常条件下显示出增加的籽粒产量。
与相同条件下种植的对照植物相比,所述植物显示早开花性状,进一步的,与对照植物相比,所述植物显示大约6-11天早开花的时间。进一步的,所述植物是水稻并种植在大约北纬30度以上的纬度。
与对照植物相比,所述植物显示提高的氮素耐性或者nue。
一种修饰植物包含一种多核苷酸,与对照植物相比,所述多核苷酸的表达是降低的,其中,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:14的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:15的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:16的序列一致性至少为90%;或(d)核苷酸序列(a)、(b)或(c)的全长互补序列,其中,与种植在相同条件下的对照植物相比,所述植物显示较晚的开花时间。
在另一实施例中,公开了提高植物耐旱性的方法,所述方法包括,与对照植物相比,增加编码icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1、dn-dtp7多肽的至少一个多核苷酸的表达,其中,所述多核苷酸包括为:(a)一种多核苷酸,其核苷酸序列与seqidno:2、5、8、11、14、17、20或23的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:3、6、9、12、15、18、21或24的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;其中,与对照植物相比,获得的植物显示出增强的耐旱性,所述提高的耐旱性能可以表现为在干旱条件下存活率增加、卷叶程度降低、种子结实率增加或籽粒产量增加。
进一步的,所述多核苷酸的表达通过以下步骤增加:(a)通过重组dna构建体增加编码icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1、dn-dtp7多肽的多核苷酸在植物中的表达,其中所述重组dna构建体包含编码icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1、dn-dtp7多肽的多核苷酸和与其可操作连接的至少一个异源调控元件,所述多核苷酸编码多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%;或(b)增加内源多核苷酸的表达,所述内源多核苷酸编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的序列一致性至少为90%。
在另一实施例中,公开了评估植物耐旱性的方法,所述方法包括的步骤为:(a)将重组dna构建体转入植物可再生细胞,所述重组dna构建体包括一种多核苷酸和与其可操作连接的至少一个异源调控序列,其中所述多核苷酸编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少80%的序列一致性;(b)步骤(a)后由植物可再生细胞再生出转基因植物,其中所述转基因植物在其基因组中含有重组dna构建体;(c)获得转基因植物的子代植物,其中所述子代植物在其基因组中含有重组dna构建体;和(d)与不含有重组dna构建体的对照植物相比,评估所述子代植物的耐旱性能。
在另一实施例中,公开了一个方法,所述方法中,与对照植物相比,能够提高水稻籽粒产量,其中所述植物在正常和/或胁迫条件下显示增强的籽粒产量,所述方法包括的步骤为增加水稻中内源icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7基因或异源icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7基因的表达。
在另一实施例中,提供了调控水稻开花时间的方法,所述方法包括改变编码mbd2多肽的多核苷酸在水稻植株中的表达,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:14的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:15的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:16的序列一致性至少为90%。所述多核苷酸的表达通过以下步骤改变:(a)通过重组dna构建体增加编码mbd2多肽的多核苷酸在植物中的表达,其中所述重组dna构建体包含编码mbd2多肽的多核苷酸和与其可操作连接的至少一个异源调控元件,所述多核苷酸编码多肽的氨基酸序列与seqidno:16的序列一致性至少为90%;(b)增加或降低内源多核苷酸的表达,所述内源多核苷酸编码的多肽的氨基酸序列与seqidno:16的序列一致性至少为90%;(c)通过重组dna构建体降低编码mbd2多肽的多核苷酸在植物中的表达,其中所述重组dna构建体包含一个沉默元件,能够下调内源多核苷酸的表达,所述多核苷酸编码多肽的氨基酸与seqidno:16的序列一致性至少为90%。进一步的,与没有所述增加表达的对照植物相比,水稻植株中所述多核苷酸表达的增加促进开花;所述水稻植株比对照植株早开花6-11天;与没有所述降低表达的对照植物相比,水稻植株中所述多核苷酸表达的降低延长开花时间。
在另一实施例中,提供了提高植物低氮耐性或nue的方法,所述方法包括增加编码mtn3l或ankl1多肽的至少一种多核苷酸在植物中的表达,所述植物不含有所述增加的表达,所述多核苷酸包括(a)一种多核苷酸,其核苷酸序列与seqidno:5或11的序列一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:6或12的序列一致性至少为85%;(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:7或13的序列一致性至少为90%。所述多核苷酸的表达通过以下步骤增加:(a)通过重组dna构建体增加编码mtn3l或ankl1多肽的多核苷酸在植物中的表达,其中所述重组dna构建体包含编码mtn3l或ankl1多肽的多核苷酸和与其可操作连接的至少一个异源调控元件,所述多核苷酸编码多肽的氨基酸序列与seqidno:7或13的序列一致性至少为90%;或(b)增加内源多核苷酸的表达,所述内源多核苷酸编码的多肽的氨基酸序列与seqidno:7或13的序列一致性至少为90%。
在另一个实施例中,公开了一种方法,所述方法中,当与对照植物中野生型icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7多肽的活性相比,增加植物中内源icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7多肽的表达或活性,与所述对照植物相比,所述植物显示出至少一种表型,所述表现选自增加的耐旱性、增加的籽粒产量、增加的非生物胁迫耐性或增加的生物量,所述方法包括的步骤为在内源基因icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7基因的基因组区域(包括启动子和基因区域)(i)导入一个dna片段或删除一个dna片段或(ii)导入一个或多个核苷酸的改变,其中所述dna片段或核苷酸的改变能够有效增加内源icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7多肽的表达或活性;所述改变可以通过锌指核酸酶、转录激活子样效应核酸酶(talen)、成簇的规则间隔的短回文重复序列技术(crispr)、引导cas核酸内切酶或归位核酸酶切酶介导实现。
在另一实施例中,植物包含本发明的dna构建体,其中,当与对照植物相比,所述植物中icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7基因的表达是增加的;所述植物显示出至少一种表型,所述表型选自籽粒产量的增加、非生物胁迫耐性的增加、生物量的增加;所述植物显示增加的非生物胁迫耐性,所述非生物胁迫是干旱胁迫。
在另一个实施例中,本发明包括任一公开的植物,所述植物选自水稻、玉米、大豆、向日葵、高粱、油菜、小麦、苜蓿、棉花、大麦、粟、甘蔗或柳枝稷。
在另一个实施例中,公开了一种在水稻植株群体中鉴定一个或多个等位基因的方法,所述等位基因与籽粒产量的增加相关,所述方法包括的步骤为(a)检测水稻植株群体中一个或多个多态性,所述多态性位于(i)多肽编码的基因组区域或(ii)调控所述多肽表达的调控区域,所述多肽包含的氨基酸序列选自seqidno:4、7、10、13、16、19、22或25,或与seqidno:4、7、10、13、16、19、22或25序列一致性为90%以上的氨基酸序列;所述多肽编码基因组区域或所述调控多肽表达的调控区域的一个或多个多态性与籽粒产量相关;和(b)鉴定一个或多个等位基因,所述等位基因位于增加产量相关的一个或多个多态性中,所述增加产量相关的一个或多个等位基因可用于分子标记辅助选择增加籽粒产量的水稻植株。所述一个或多个多态性位于多核苷酸的编码区,所述调控区域是启动子。
在另一个实施例中,本发明涉及重组dna构建体,所述重组dna构建体包含本发明任一分离的多核苷酸和与其可操作连接至少一个调控序列;以及含有所述重组dna构建体的细胞、植物或种子。所述细胞可以是真核细胞,如酵母、昆虫或植物细胞;也可以是原核细胞,诸如细菌细胞。
附图说明和序列表
根据以下发明详述和附图及序列表,可以更全面的理解本发明,以下发明详述和附图及序列表形成本发明的一部分。
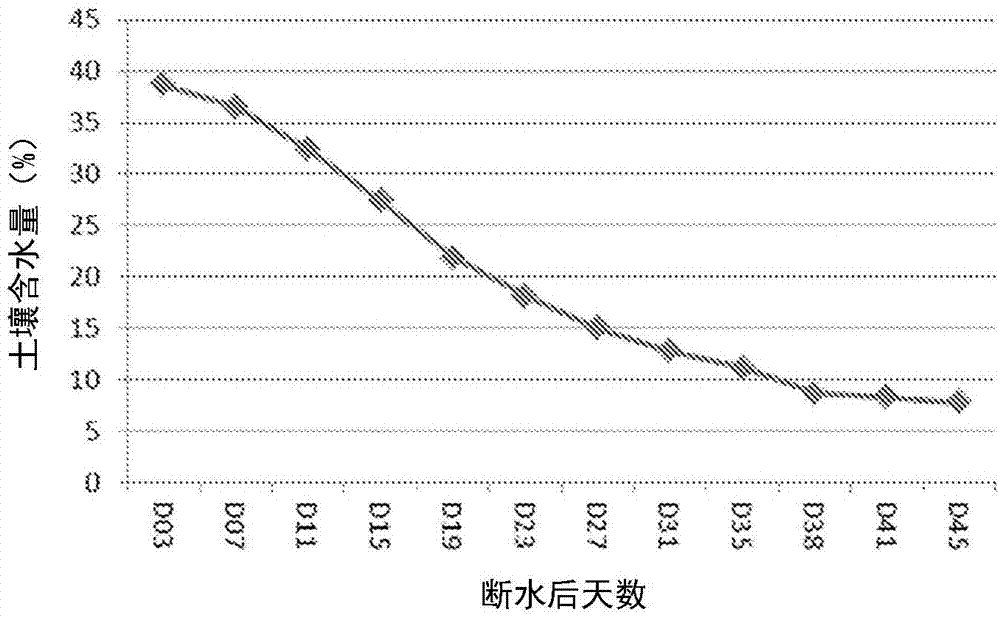
图1为第一次田间干旱试验中海南田间在osicdh1转基因水稻不同发育时期的土壤含水量。osicdh1转基因水稻在断水后25天开始抽穗。
图2为第二次田间干旱验证中海南田间在osicdh1转基因水稻不同发育时期的土壤含水量。osicdh1转基因水稻的主茎穗在断水后处于幼穗分化ⅳ-ⅵ期。
图3为干旱试验中海南田间在osmtn3l转基因水稻不同发育时期土壤含水量。osmtn3l转基因水稻在断水后36天开始抽穗。
图4为干旱验证中宁夏田间在osmtn3l转基因水稻不同发育时期的土壤含水量。osmtn3l转基因水稻在断水后47天开始抽穗
图5为干旱试验中海南田间在osdn-dtp6和osankl1转基因水稻不同发育时期土壤含水量。osdn-dtp6和osankl1转基因水稻在断水后38天开始抽穗。
图6为第二次干旱试验中田间在osdn-dtp6和osankl1转基因水稻不同发育时期的土壤含水量。osdn-dtp6和osankl1转基因水稻在断水后25天开始抽穗。
图7为干旱试验中海南田间在osmbd2转基因水稻不同发育阶段的土壤含水量。osmbd2转基因水稻在断水后37天开始抽穗。
图8为第二次干旱试验中田间在osmbd2转基因水稻在不同发育阶段的土壤含水量。osmbd2转基因水稻在断水后33天开始抽穗。
图9为干旱试验中海南田间试验在ostp1和osacoat1转基因水稻不同发育阶段土壤含水量。ostp1和osacoat1转基因水稻在断水后37天开始抽穗。
图10为干旱试验中宁夏田间在ostp1和osacoat1转基因水稻不同发育阶段土壤含水量。ostp1和osacoat1转基因水稻在断水后44天开始抽穗。
图11为干旱试验中海南田间在osdn-dtp7转基因水稻不同发育阶段土壤含水量。osdn-dtp7转基因水稻在断水后35天开始抽穗。
图12为第二次干旱试验中田间在osdn-dtp7转基因水稻不同发育阶段土壤含水量。osdn-dtp7转基因水稻在断水后26天开始抽穗。
图13为实时pcr分析测定的不同转基因水稻株系叶片中osmbd2基因相对表达水平,dp0158中基本表达水平设定为1.00,转基因事件的表达量为与dp0158水稻相比的倍数。
表1.序列表中核苷酸和氨基酸序列的编号
序列描述以及关联的序列表遵循如37c.f.r.§1.821-1.825中所列出的管理专利申请中的核苷酸和/或氨基酸序列公开的规则。序列表包含核苷酸序列字符的单字母码以及氨基酸的三字母码,如遵照iupac-iubmb标准所定义的,该标准在nucleicacidsres.13:3021-3030(1985)以及在biochemicalj.219(no.2):345-373(1984)中有所描述,这两篇文献以引用的方式并入本发明。用于核苷酸和氨基酸序列数据的符号和格式遵循37c.f.r.§1.822中列出的规则。
详细描述
本发明中所列出的每篇参考文献的公开内容的全文均以引用的方式并入本发明。
如本发明所用的并在所附权利要求书中的单数形式“一个”和“所述”包括复数涵义,除非上下文中清楚地另有指明。因此,例如,“一株植物”的涵义包括多株该类植物。“一个细胞”的涵义包括一个或多个细胞及其本领域的技术人员已知的等同物,等等。
如本发明所述:
“osicdh1”是nadp依赖的异柠檬酸脱氢酶(nadp-dependentisocitratedehydrogenase),涉及水稻基因位点loc_os01g14580.1编码的能够赋予植物耐旱性表型的水稻多肽。“icdh1多肽”此处涉及osicdh1多肽和源于其他植物的同源物。
osicdh1多肽(seqidno:4)是水稻基因位点loc_os01g14580.1的编码序列(cds)(seqidno:3)或核酸序列(seqidno:2)编码的氨基酸序列。该多肽在tigr(theinternetatplantbiologymsu.edu/index.shtml)中注释为“脱氢酶,推测,表达”,在ncbi(ontheworldwebatncbi.nlm.nih.gov)注释为“nadp依赖的异柠檬酸脱氢酶”,但是没有在先的功能介绍。
“osmtn3l”是mtn3类似蛋白,涉及水稻基因位点loc_os01g40960.1编码的能够赋予植物耐旱性表型的水稻多肽。“mtn3l多肽”此处涉及osmtn3l多肽和源于其他植物的同源物。
osmtn3l多肽(seqidno:7)是水稻基因位点loc_os01g40960.1的编码序列(cds)(seqidno:6)或核酸序列(seqidno:5)编码的氨基酸序列。该多肽在tigr中注释为“根瘤素mtn3家族蛋白,推测,表达”,在ncbi注释为“mtn3-类似”,但是没有在先的功能介绍。
“osdn-dtp6”是耐旱性蛋白6(droughttoleranceprotein6),涉及水稻基因位点loc_os09g15190.1编码的能够赋予植物耐旱性表型水稻多肽。“dn-dtp6多肽”此处涉及osdn-dtp6多肽和源于其他植物的同源物。
osdn-dtp6多肽(seqidno:10)是水稻基因位点loc_os09g15190.1的编码序列(cds)(seqidno:9)或核酸序列(seqidno:8)编码的氨基酸序列。该多肽在tigr中注释为“表达蛋白”。
“osankl1”是锚蛋白类似蛋白(ankyrinlikeprotein),涉及水稻基因位点loc_os09g15200.1编码的能够赋予植物耐旱性表型水稻多肽。“ankl1多肽”此处涉及osankl1多肽和源于其他植物的同源物。
osankl1多肽(seqidno:13)是水稻基因位点loc_os09g15200.1的编码序列(cds)(seqidno:12)或核酸序列(seqidno:11)编码的氨基酸序列。该多肽在tigr中注释为“锚蛋白重复家族蛋白,推测,表达”,在ncbi注释为“锚蛋白类似蛋白”,但是没有在先的功能介绍。
“osmbd2”是甲基结合结构域蛋白(methyl-bindingdomainprotein,mbd2),涉及水稻基因位点loc_os06g48870.1编码的能够赋予植物耐旱性和调控水稻开花性状的多肽。“mbd2多肽”此处涉及osmbd2多肽和源于其他植物的同源物。
osmbd2多肽(seqidno:16)是水稻基因位点loc_os06g48870.1的编码序列(cds)(seqidno:15)或核酸序列(seqidno:14)编码的氨基酸序列。tigr中,该多肽注释为“甲基结合结构域蛋白mbd,推测,表达”。
“ostp1”是海藻糖磷酸酶1(trehalosephosphatase1),涉及水稻基因位点loc_os05g03810.1编码的能够赋予植物耐旱性表型水稻多肽。“tp1多肽”此处涉及ostp1多肽和源于其他植物的同源物。
ostp1多肽(seqidno:19)是水稻基因位点loc_os05g03810.1的编码序列(cds)(seqidno:18)或核酸序列(seqidno:17)编码的氨基酸序列。该多肽在tigr中注释为“海藻糖磷酸酶,推测,表达”。
“osacoat1”是乙酰鸟苷酸转氨酶(acetylornithineaminotransferase),涉及水稻基因位点loc_os05g03830.1编码的能够赋予植物耐旱性表型水稻多肽。“acoat1多肽”此处涉及osacoat1多肽和源于其他植物的同源物。
osacoat1多肽(seqidno:22)是水稻基因位点loc_os05g03830.1的编码序列(cds)(seqidno:21)或核酸序列(seqidno:20)编码的氨基酸序列。该多肽在tigr中注释为“转氨酶,推测,表达”,在ncbi注释为“乙酰鸟苷酸转氨酶”。
“osdn-dtp7”是耐旱性蛋白7(adroughttoleranceprotein7),涉及水稻基因位点loc_os03g15720.1编码的能够赋予植物耐旱性表型水稻多肽。“dn-dtp7多肽”此处涉及osdn-dtp7多肽和源于其他植物的同源物。
osdn-dtp7多肽(seqidno:25)是水稻基因位点loc_os03g15720.1的编码序列(cds)(seqidno:24)或核酸序列(seqidno:23)编码的氨基酸序列。该多肽在tigr中注释为“表达的蛋白”。
本发明中的单子叶植物包括禾本科的植物;双子叶植物包括十字花科、豆科和茄科的植物。
“开花”指开花的过程,即在适宜的温度和湿度下,颖壳开裂,花药散射,或花的形成过程。此处开花用于表示从幼穗分化、成熟到幼穗抽出的过程。
“花发育”是指花或花序从花分生组织的开始到发育成熟花的发育。
“生殖发育”是指花或花序从花分生组织开始经过授粉到果实成熟的发育。
此处“早开花时间”的植物指开花早于对照的植物,因此这个词指的是显示开花起始较早的植物。植物的开花时间指播种到第一个花序出现的天数,例如,植物的“开花时间”可以使用wo2007/093444中描述的方法确定。
“抽穗”此处指禾谷类作物发育完全的幼穗从剑叶鞘内伸出的时期或状态。
“抽穗期”和“抽穗时间”本发明可互换使用,指从种子播种到50%幼穗从水稻植株剑叶鞘抽出的天数。抽穗期是重要的农业性状,由基本的营养基因和光周期敏感基因调控,在水稻品种的适应性和地理分布起关键的作用。适当的抽穗期是获得理想产量的前提。
通常情况下,水稻穗抽出后即开花,因此本发明抽穗期也用于表示开花期。
“植株高度”本发明指从地面到单株植株最高穗或叶顶部的高度。
“全长互补序列”是指给定核苷酸序列的互补序列,互补序列和核苷酸序列含有相同的核苷酸数,并且100%的互补。
“性状”指植物或特定植物材料或细胞的生理的、形态的、生化的或物理的特征。在一些实施例中,这些特征可以是肉眼可见的,比如种子、植株的大小等;可用生物化学技术测定的指标,如种子或叶片中蛋白、淀粉或油份的含量等;可观察的代谢或生理过程,如测定对水分胁迫、特定盐、糖或氮浓度的耐性;可检测的基因表达水平;或可观察渗透胁迫的耐性或产量等农艺性状。
“农艺性状”是可测量的指标参数,包括但不限于:叶片绿色、籽粒产量、生长速率、总生物量或积累速率、成熟时的鲜重、成熟时的干重、果实产量、种子产量、植物总氮含量、果实氮含量、种子氮含量、植物营养组织氮含量、植物总游离氨基酸含量、果实游离氨基酸含量、种子游离氨基酸含量、植物营养组织游离氨基酸含量、植物总蛋白含量、果实蛋白含量、种子蛋白含量、植物营养组织蛋白质含量、耐旱性、氮的吸收、根的倒伏、收获指数、茎的倒伏、株高、穗高、穗长、耐盐性、分蘖数、圆锥花序的大小、早期苗的活力和低温胁迫下的出苗状况。
可以测量增加的生物量,例如与对照植物相比,增加的植物高度、植物总叶面积、植物鲜重、植物干重或植物种子产量等。
增加植株生物量或大小的能力具有多种重要商业应用,作物栽培品种可用于产生较高植物营养组织部分,用于食品、饲料、纤维和/或生物燃料。
人们对叶片大小的增加特别有兴趣。增加的叶片的生物量可用于增加植物源药或工业产品的生产,增加的分蘖数可以增加产量,增加植物叶片面积可提高植物总的光合作用,增加的光合合成能力可以增加特定植物组织的产量,并使植物在低光强或高光强下生长,所述特定植物组织包括叶片、根、果实或种子。
其它组织如根的生物量的改变有利于提高植物在严酷条件下生长的能力,所述严酷条件包括干旱、营养贫瘠,因为大量的根系可以更好地吸收水分和营养。
对于观赏植物,人们所期望获得较大的品种,许多植物包括果树,用于木材生产的树,作为视图或风帘的乔木或灌木,可以通过增加其大小以获得较大的产量或增加屏障。
“转基因的”指其基因组因异源核酸(如重组dna构建体)的存在而发生改变的任何细胞、细胞系、愈伤组织、组织、植物部分或植物,包括那些最初的转基因事件以及从最初的转基因事件通过有性杂交或无性生殖而产生的那些。如本发明所用的术语“转基因”不涵盖通过常规植物育种方法或通过诸如随机异花受精、非重组病毒感染、非重组细菌转化、非重组转座或自发突变之类的自然发生事件导致的基因组(染色体基因组或染色体外基因组)改变。
“对照”、“对照植物”或“对照植物细胞”为测定受测试植物或植物细胞的表型变化提供参考,由于转化,受测植物或植物细胞的基因组改变影响到目的基因,测试的植物或植物细胞可以是转基因植物或转基因植物细胞的子代。
对照植物或对照植物细胞包括,例如:(a)用于基因改变产生受测植物或细胞的野生型植物或细胞;(b)相同基因组作为起始材料但是转入空载体(如带有标记基因的并对目标的性状没有影响的载体)的植物或植物细胞;(c)转基因植物或植物细胞性状分离,获得的非转基因的子代植物或植物细胞;(d)没有暴露于可以诱导基因表达的条件或刺激下的,与转基因植物或植物细胞基因组相同的植物或植物细胞;(e)特定的目标基因不表达情况下的,转基因植物或者植物细胞本身。对照可以包括多种代表上述一种或多种类型的个体,例如,(c)中性状分离后非转基因的材料的混合通常认为是bulknull。
本发明中,zh11-tc和dp0158指对照植物。zh11-tc表示通过组织培养中花11获得的水稻植株,dp0158表示转化空载体dp0158获得水稻植株。
“基因组”在用于植物细胞时不仅涵盖存在于细胞核中的染色体dna,而且还包括存在于细胞的亚细胞组分(如线粒体、质粒)中的细胞器dna。
“植物”包括整个植株、植物器官、植物组织、种子和植物细胞以及同一植株的子代。植物细胞包括但不限于得自下列物质的细胞:种子、悬浮培养物、胚、分生区域、愈伤组织、叶、根、芽、配子体、孢子体、花粉和小孢子。
“子代”包括植物的任何后续世代。
“修饰的植物”包括在其基因组内包含异源多核苷酸或修饰的基因或启动子的植物。例如异源多核苷酸能够稳定地整合进基因组中,并遗传连续的世代。异源多核苷酸可单独地或作为重组dna构建体的部分整合进基因组中。t0植物直接源于转化和再生过程,t0植物的子代为t1代(第一个子代),t2代(第二个子代)等。
针对序列而言的“异源”意指来自外来物种的序列,或者如果来自相同物种,则指通过蓄意的人为干预而从其天然形式发生了组成和/或基因座的显著改变的序列。
“多核苷酸”、“核酸序列”、“核苷酸序列”或“核酸片段”可互换使用并且是任选含有合成的、非天然的或改变的核苷酸碱基的单链或双链rna或dna聚合物。核苷酸(通常以它们的5′-单磷酸形式存在)通过如下它们的单个字母名称来指代:“a”为腺苷酸或脱氧腺苷酸(分别对应rna或dna),“c”表示胞苷酸或脱氧胞苷酸,“g”表示鸟苷酸或脱氧鸟苷酸,“u”表示尿苷酸,“t”表示脱氧胸苷酸,“r”表示嘌呤(a或g),“y”表示嘧啶(c或t),“k”表示g或t,“h”表示a或c或t,“i”表示肌苷,并且“n”表示任何核苷酸。
“多肽”、“肽”、“氨基酸序列”和“蛋白质”在本发明中可互换使用,指氨基酸残基的聚合物。该术语适用于其中一个或多个氨基酸残基是相应的天然存在的氨基酸的人工化学类似物的氨基酸聚合物,以及适用于天然存在的氨基酸聚合物。术语“多肽”、“肽”、“氨基酸序列”和“蛋白质”还可包括修饰形式,包括但不限于糖基化、脂质连接、硫酸盐化、谷氨酸残基的γ羧化、羟化和adp-核糖基化。
“信使rna(mrna)”指无内含子并且可通过细胞翻译成蛋白质的rna。
“cdna”指与mrna模板互补并且利用逆转录酶从mrna模板合成的dna。cdna可为单链的或者可用dna聚合成酶i的klenow片段合成双链形式。
“成熟”蛋白质指经翻译后加工的多肽;即已经去除了存在于初级翻译产物中的任何前肽或原肽的多肽。
“前体”蛋白质指mrna的翻译初级产物;即带有前肽和原肽的蛋白质。前肽和原肽可为并且不限于细胞内定位信号。
“分离的”指物质,例如核酸和/或蛋白质,该物质基本上不含在天然存在的环境中通常伴随该物质或与其反应的组分,或者说是该物质被从所述组分移出。分离的多核苷酸可从它们天然存在的宿主细胞中纯化。技术人员已知的常规核酸纯化方法可用于获得分离的多核苷酸。该术语也涵盖重组多核苷酸和化学合成的多核苷酸。
“重组体”指(例如)通过化学合成或者通过用基因工程技术操纵分离的核酸片段来实现的两个原本分离的序列片段的人工组合。“重组体”也包括指已经通过引入异源核酸而进行了修饰的细胞或载体,或源于经这样修饰的细胞的细胞,但不涵盖由天然发生的事件(如自发突变、自然转化/转导/转座)对细胞或载体的改变,例如没有蓄意人为干扰而发生的那些。
“重组dna构建体”指在自然界中通常不会一起存在的核酸片段的组合。因此,重组dna构建体可包含源于不同来源的调控序列和编码序列,或源于相同来源但以不同于通常天然存在的方式排列的调控序列和编码序列。
术语“入门克隆”和“入门载体”本发明可互换使用。
“调控序列”和“调控元件”可互换使用,指位于编码序列的上游(5'非编码序列)、中间或下游(3'非编码序列),并且影响相关编码序列的转录、rna加工或稳定性或者翻译的核苷酸序列。调控序列可包括但不限于启动子、翻译前导序列、内含子和多腺苷酸化识别序列。
“启动子”指能够控制另一核酸片段转录的核酸片段。
“植物中有功能启动子”是能够控制植物细胞中基因转录的启动子,无论其是否来源于植物细胞。
“组织特异性启动子”和“组织优选启动子”可互换使用,并且指主要但非必须专一地在一种组织或器官中表达,而且也可在一种特定细胞或细胞型中表达的启动子。
“发育调控启动子”指其活性由发育事件决定的启动子。
术语“可操作地连接”指核酸片段连接成单一片段,使得其中一个核酸片段的功能受到另一个核酸片段的调控。例如,在启动子能够调节核酸片段的转录时,该启动子与该核酸片段进行了可操作地连接。
“表达”指功能产物的产生。例如,核酸片段的表达可指核酸片段的转录(如转录生成mrna或功能rna)和/或rna翻译成前体或成熟蛋白质。
“表型”意指细胞或生物体的可检测的特征。
有关将核酸片段(例如重组dna构建体)插入细胞内的“导入”是指“转染”或“转化”或“转导”,并且包括指将核酸片段整合进真核或原核细胞中,在该细胞中核酸片段可整合进细胞的基因组(如染色体、质粒、质体或线粒体dna)内,转变成自主的复制子或瞬时表达(如转染的mrna)。
“转化细胞”是将核酸片段(如重组dna构建体)导入其中的任何细胞。
本发明所用的“转化”指稳定转化和瞬时转化两者。
“稳定转化”指将核酸片段导入宿主生物体的基因组中,导致基因稳定遗传。一旦稳定转化,核酸片段稳定地整合进宿主生物体和任何连续世代的基因组中。
“瞬时转化”指将核酸片段导入宿主生物体的核中或包含dna的细胞器中,引起基因表达而没有基因稳定遗传。
“等位基因”是占据染色体上给定位点的基因的几种供选择形式的其中一种。当二倍体植物中一对同源染色体上给定基因座上存在的等位基因相同时,该植物在该基因座处是纯合的。如果二倍体植物中一对同源染色体上给定基因座上存在的等位基因不同,则该植物在该基因座处是杂合的。如果转基因存在于二倍体植物中一对同源染色体中的其中之一上,则该植物在该基因座处是半合子的。
“叶绿体转运肽”是与蛋白协同翻译并将蛋白导向叶绿体或翻译蛋白的细胞中存在的其它质体类型的氨基酸序列。“叶绿体转运序列”指编码叶绿体转运肽的核苷酸序列。“信号肽”是一种与蛋白协同翻译并将蛋白导向分泌器官的氨基酸序列(chrispeels,m.(1991)ann.rev.plantphys.plantmol.biol.42:21-53)。如果将所述蛋白导向液泡,可另外加上液泡靶向信号,或者如果将所述蛋白导向内质网,可加上内质网驻留信号。如果将蛋白导向细胞核,将移除任何存在的信号肽并用以核定位信号替代(raikhel(1992)plantphys.100:1627-1632)。“线粒体信号肽”是引导前体蛋白进入线粒体的氨基酸序列(zhang和glaser(2002)trendsplantsci7:14-21)。
dna核酸酶和其他的突变酶结构域可以与dna结合结构域结合在靶dna上产生双链断裂(dsb),dna结合结构域包括,例如一个特异dna结合结构域,或一个位点特异dna结合结构域。位点特异dna结合结构域包括但不限于tal(转录激活样效应因子)或锌指结合结构域。
dna结合结构域与dna核酸酶融合的例子包括但不限于talen和多talen。转录激活样效应因子核酸酶(talens)是tal效应因子dna结合结构域融合到dna酶的结构域产生的人工限制性酶。tal蛋白是细菌产生的,包括高度保守的33~34氨基酸dna结合结构域序列(pct公开号wo2014127287,美国专利公开号us20140087426)。
起初的talen嵌合体是由野生型的foki核酸内切酶结构域制备的,而talen也包括源于foki核酸内切酶结构域变异提高了切割特异性和切割活性突变嵌合体。在一些例子中,多个talen可以靶向多个基因区域表达。
锌指是另一种类型的dna结合结构域,可以将突变引入靶dna。
各种蛋白工程技术可以用于改变锌指dna结合特异性,这样设计的锌指的串联重复序列可用于靶向期望的基因组dna序列。融合第二个蛋白结构域例如转录抑制子到锌指,可以与附近的yep基因启动子结合,从而降低icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1或dn-dtp7基因表达水平。
在一个实施例中,一个调控元件驱动内源基因表达或编码序列本身,例如,可能会通过crispr/cas9体系的基因组编辑技术被编辑或插入植物中。
crispr位点(成簇的规律间隔短回文重复,也被称作spidrs,间隔穿插的顺向重复)构成近来描述的dna位点家族,crispr位点由短且高度保守的dna重复(典型的24~40bp,重复1~140次,也称为crispr重复)组成,crispr位点为部分回文。重复序列(通常具有品种特异性)被固定长度(代表性的20~58bp,根据crispr位点而不同)的可变序列间隔开(公布与2007年3月1日的wo2007/025097)。
cas基因指通常耦合的,关联或邻近crispr位点或在crispr位点侧翼的附近的基因。术语“cas基因”、“crispr关联(cas)基因”此处可以互换使用。(haft等(2005),计算生物学,ploscomputbiol1(6):e60,doi:10.1371/journal.pcbi.0010060)。如本发明所描述,除了四个已知基因家族外,41个crispr关联(cas)基因家族进行了描述。这表明crispr系统属于不同的类别,具有不同的重复模式,基因组别和物种范围。一个给定的crispr位点不同物种间的cas基因数目不同。
cas核酸内切酶指cas基因编码的cas蛋白,其中,所述cas蛋白可以向靶dna序列导入双链断裂。cas核酸切酶由引导多聚核苷酸引导识别和选择性的向细胞基因组特定靶位点导入双链断裂(u.s.2015/0082478)。引导多聚核苷酸/cas核酸内切酶体系包括cas核酸内切酶和引导多聚核苷酸组成的复合体,所述引导多聚核苷酸能够向dna靶序列导入双链断裂。如果一个正确的前间区序列邻近基序(pam)位于的靶序列的3’端,cas核酸内切酶有引导rna引导解开基因组靶位点邻近dna双链,切割靶序列dna双链的识别位点。cas核酸内切酶可以通过本领域已知的方法直接导入细胞,例如,转染和/或局部适用,但不限于瞬时导入方法。
如本发明中使用,术语“引导rna”指crrna(crisprrna)和tracrrna两个rna分子合成的融合,所述crrna包括一个可变的靶结构域。在一个实施例中,引导rna包括一个12~30bp核苷酸序列的可变的靶结构域和可以与cas核酸内切酶相互作用的rna片段。
如本发明所述,术语“引导多聚核苷酸”指可以与cas核酸内切酶构成复合体的多聚核苷酸序列,并且可以使cas核酸内切酶识别并随意切割dna靶位点(u.s.2015/0082478)。引导多聚核苷酸可以是单链分子也可以是双链分子,引导多核苷酸序列可以是rna序列,dna序列或它们的组合(rna-dna组合序列)。可选的,引导多聚核苷酸包括至少一个核苷酸,磷酸二酯键或修饰键,例如,但不限于锁定核苷酸(lna)、5-甲基脱氧胞苷、2,6-二氨基嘌呤、2-氟腺苷,2-氟脲苷、2-甲基rna、硫代磷酸酯键,一种胆固醇分子链、聚乙二醇分子链、以间隔18(六乙二醇链)分子链、或5至3共价键产生的环。引导多聚核苷酸仅包括也被称作“引导rna”的脱氧核糖核酸。
术语“可变靶结构域”或“vt结构域”此处可互换使用,包括与双链dna靶位点的一条单链(核苷酸序列)互补的核苷酸序列。第一个核苷酸序列结构域(vt结构域)与靶序列的互补百分数至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、63%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%。可变靶结构域的长度至少为12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29或者30bp的核苷酸。在一些实施例中,可变靶结构域由邻近延伸的12~30bp核苷酸组成。可变靶结构域是由dna序列、rna序列、修饰的dna序列、修饰的rna序列或它们的任意组合组成。
术语“cas核酸内切酶识别结构域”或引导多聚核苷酸的“cer结构域”此处可互换使用,包括与cas核酸内切酶多肽相互作用的核苷酸序列(诸如引导多聚核苷酸的第二核苷酸序列)。cer结构域由dna序列、rna序列、修饰的dna序列、修饰的rna序列(例如此处描述的修饰)或它们的任意组合构成。
核苷酸序列连接单引导多聚核苷酸的crnucleotide和tracrnucleotide可以组成rna序列、dna序列或rna-dna组合序列。在一个实施例中,核苷酸序列连接单引导多聚核苷酸的crnucleotide和tracrnucleotide组成长度至少为3、4、5、6、7、8、9、10、11、12、13、14、15、16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99或100bp。在另一个实施例中,核苷酸序列连接单多聚核苷酸的crnucleotide和tracrnucleotide组成四核苷酸环序列,诸如,但不限于gaaa四核苷酸环序列。
目前已知有多种可以用来研究各种多聚核苷酸和多肽序列之间关系的方法。本发明中,“参考序列”是一个界定序列,用作序列比对的基础。参考序列是一个特定序列的子集或全部,例如全长cdna/基因序列的片段,抑或完整的cdna/基因序列。此处用到的“比对窗口”,涉及到一个多聚核苷酸或多肽序列的共有或特定片段,其中两个序列的最佳比对时,与参考序列(没有添加或者删除)相比,比较窗口中序列可能会包含添加或删除(换言之既间隙)。通常比对窗口中至少包含20个连续的碱基或氨基酸,还可以选择30、40、50、100甚至更长。本领域的技术人员为了避免因引入序列间隙而导致的与参考序列的高相似性,引入空位罚分,并减去相应的匹配数目。
使用缺省参数就可以通过上述程序开展序列联配工作。higgins等(gene73:237-244,1988),higgins等(cabios5:151-153,1989),corpet等(nucleicacidsres.16:10881-10890,1988),huang等(cabios8:155-165,1992)和pearson等(meth.mol.biol.24:307-331,1994)对clustal进行了详细的描述;align程序则以myers和miller提出的supra算法(1988)为基础。进行氨基酸序列比对时,当使用pam120残基重量表,且空位罚分为12和空位罚分为4时,可以与align程序相结合对氨基酸序列进行比较。blast程序(altschul等,j.mol.biol.215:403,1990)以karlin和altschul的supra算法(1990)为基础,可以使用blastn对核苷酸序列进行检索,score=100,wordlength=12,以获得本发明所涉及蛋白质对应编码核苷酸序列的同源序列;可以使用blastx,score=50,wordlength=3,以获得本发明所涉及蛋白质或多肽的同源氨基酸序列。gappedblast(blast2.0)(altschul等,(1997)nucleicacidsres.25:3389)可以用于gap比对。作为一种选择,psi-blast(blast2.0)可以开展交互式搜索,从而检测到亲缘关系较远的两个分子(altschul等(1997)supra)。当使用blast时,gappedblast、psi-blast和相应程序的缺省参数(例如用于核甘酸序列的blastn,用于蛋白质的blastx)也可以一并使用(美国政府国立卫生研究院国立医学图书馆国家生物技术信息中心)。序列联配也可以用于人工检索。
通过使用gap(ver10)和以下参数可以得到配对序列一致性/相似性值:使用gapweight50、lengthweight3以及nwsgapdna.cmp得分矩阵来计算核酸序列的一致性(%)和相似性(%);使用gapweight8、lengthweight2以及blosum62得分矩阵可以计算氨基酸序列的一致性(%)和相似性(%);或者任何等同程序。任何序列比较程序都可以被看作“等同程序”,因为任意两条序列进行比较,都会产生一个联配结果,包含一致的核苷酸或氨基酸碱基配对。当与gapver10所产生序列联配结果进行比较时,还会产生序列一致性百分比。
gap利用needleman和wunsch算法((1970)j.mol.biol.48:443-453),以发现两个全长序列联配的最大匹配数和最小间隙数。gap考虑所有可能的联配和gap位点,并创造联配位点数目最大且gap数目结果,它允许匹配碱基单元中存在间隙创造罚分和间隙延展罚分,gap必须利用每个插入间隙的间隙创造罚分匹配数,另外,如果选择间隙延展罚分大于0,gap必须利用每个插入间隙的长度间隙次数,间隙延展罚分。gcgwisconsin遗传学软件包ver10中,缺省间隙创造罚分值和间隙延展罚分值分别为8和2,核苷酸序列中,缺省间隙创造罚分是50,缺省间隙延展罚分为3,间隙创造和间隙延展罚分可以是0-200中的整数,因此间隙创造和间隙延展罚分可以是0、1、2、3、4、5、6、7、8、9、10、15、20、25、30、35、40、45、50、55、60、65或更大。
除非特别说明,本发明中的多序列比对采用clustalv比对方法(higgins和sharp.(1989)cabios.5:151-153)和缺省参数(间隙罚分=10,间隙长度罚分=10),两个比对的缺省参数和计算氨基酸序列一致性百分数使用clustalv方法:ktuple=1,间隙罚分=3,窗口=5和对角线拯救=5;用于核酸序列的参数ktuple=2,间隙罚分=5,窗口=4和对角线拯救=4。序列比对后应用clustalv程序,通过观察“序列距离”表,可以得到“一致性百分比”和“散度”值,除非特别说明,一致性百分比和散度值以相同的方式获得。
本发明中,当在特定比对窗口中最大一致性比对时,两个多聚核苷酸或多肽序列的“序列一致性”或“一致性”指参考两序列中残基相同,当序列一致性百分数用于提及的蛋白质,被认为不同残基位置与保守氨基酸替代不同,氨基酸残基被具有相似化学性质的其它氨基酸残基(如电荷或疏水性)替代,而不改变分子的功能特性,当序列在保守区替代不同,序列一致性百分数可以上调以更正替代的保守性,保守替代不同的序列被认为具有“序列相似性”或“相似性”。本领域的技术人员熟悉进行调整的方法,包括计算作为部分的而不是全长错配的保守替代,因而增加序列一致性百分数,因此,相同氨基酸给与1分,不保守的替代给与0分,保守替代给与0-1分,计算保守替代的得分,如在pc/gene执行(intelligenetics,mountainview,加州)。
本发明中“序列一致性百分数”的计算包括确定两个序列中相同核苷酸碱基或氨基酸残基的位置数目,获得匹配位置数,然后将匹配的位置数除以比对窗口中位置总数再乘以100。
本发明中使用的标准重组dna和分子克隆技术为本领域技术人员熟知,并且在如下文献中有更全面的描述:sambrook,j.,fritsch,e.f.和maniatis,t.,molecularcloning:alaboratorymanual;coldspringharborlaboratorypress:coldspringharbor,1989(下文称为“sambrook”)。
现在转向实施方案:
实施方案包括分离的多核苷酸和多肽、赋予耐旱性的重组dna构建体、包含重组dna构建体的诸如植物或种子的组合物、以及利用这些重组dna构建体的方法。
分离的多核苷酸和多肽:
本发明包括如下分离的多核苷酸和多肽:
一种分离的多核苷酸,包括(i)一种核酸序列,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)核酸序列(i)的全长互补序列,其中核酸序列(i)与全长互补序列具有相同数目的核苷酸,并且100%的互补。前述任一分离的多核苷酸可用于构建本发明的任一重组dna构建体,过量表达所述编码的多肽提高植物的耐旱性和/或百草枯耐性。过量表达所述编码的多肽增加正常条件下的籽粒产量。
一种分离的多肽,所述多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性。所述多肽优选为耐旱性多肽,过量表达所述多肽增加植物的耐旱性和/或百草枯耐性。过量表达所述编码的多肽增加正常条件下的籽粒产量。
一种分离的多核苷酸,包括(i)一种核酸序列,其核苷酸序列与seqidno:3、6、9、12、15、18、21或24相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(ii)一种核酸序列,其核苷酸序列与seqidno:2、5、8、11、14、17、20或23相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(iii)核酸序列(i)或(ii)的全长互补序列。前述任一分离的多核苷酸可用于构建本发明任一重组dna构建体。所述分离的多核苷酸优选为编码耐旱性的多肽,过量表达所述多肽提高植物的耐旱性和/或百草枯耐性。过量表达所述编码的多肽增加正常条件下的籽粒产量。
一种分离的多核苷酸包括(i)一种核酸序列,所述核酸序列编码的多肽的氨基酸序列与seqidno:16的相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或者(ii)核酸序列(i)的全长互补序列,其中核酸序列(i)与全长互补序列核苷酸数目相同且100%互补。过量表达编码的多肽促进植物营养生长向生殖生长的转变。
一种分离的多肽,其氨基酸序列与seqidno:16相比具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性。所述多肽为mbd2,可以调控植物开花时间。
一种分离的多核苷酸,包括(i)一种多核苷酸,其核苷酸序列与seqidno:14或15相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)核酸序列(i)的全长互补序列。所述分离的多核苷酸序列优选为编码能够调控开花时间的蛋白。过量表达所述多核苷酸能够促进植物从营养生长向生殖生长的转变;减少所述多核苷酸的表达水平延长植物从营养生长向生殖生长的转变的时间。
一种分离的多核苷酸,其包括(i)一种核酸序列,其编码的多肽与seqidno:7或13相比具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(ii)核酸序列(i)的全长互补序列,其中核酸序列(i)与全长互补序列核苷酸数目相同且100%互补。前述任一分离的多核苷酸可用于构建本发明的任一重组dna构建体,过量表达所述编码的多肽提高植物的低氮耐性和nue。
一种分离的多肽,其氨基酸序列与seqidno:7或13相比具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性。过量表达所述多肽提高植物的低氮耐性和nue。
一种分离的多核苷酸,包括(i)一种核酸序列,其核苷酸序列与seqidno:6或12相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(ii)一种核酸序列,其核苷酸序列与seqidno:5或11相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(iii)核酸序列(i)或(ii)的全长互补序列。前述任一分离的多核苷酸可用于构建本发明任一重组dna构建体。过量表达所述多肽提高植物的低氮耐性和nue。
重组dna构建体和抑制dna构建体:
一方面,本发明包括重组dna构建体和抑制dna构建体。
在一个实施方案中,一种重组dna构建体包含一种多核苷酸和与其可操作连接的至少一个调控序列(如植物中有功能的启动子),其中所述多核苷酸包括(i)一种核酸序列,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)核酸序列(i)的全长互补序列。
在另一个实施方案中,一种重组dna构建体包含一种多核苷酸和与其可操作连接的至少一个调控序列(如植物中有功能的启动子),其中,所述多核苷酸包括(i)一种核酸序列与seqidno:3、6、9、12、15、18、21或24相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(ii)一种核酸序列与seqidno:2、5、8、11、14、17、20或23相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(iii)核酸序列(i)或(ii)的全长互补序列。
在一个实施方案中,一种重组dna构建体包含一个多核苷酸和与其可操作连接的至少一个调控序列(如植物中用功能的启动子),其中所述多核苷酸编码一个耐旱性的多肽。所述多肽优选为具有耐旱性、低氮耐性和/或百草枯耐性,并可来自,例如水稻(oryzasativa)、拟南芥(arabidopsisthaliana)、玉米(zeamays)、大豆(glycinemax)、烟豆(glycinetabacina)、野大豆(glycinesoja)和短绒野大豆(glycinetomentella)。
在另一实施方案中,一种重组dna构建体包含一种多核苷酸和与其可操作连接的至少一个调控序列(如植物中有功能的启动子),其中所述多核苷酸编码mbd2蛋白,所述多肽可调控开花时间,并可选自,例如水稻(oryzasativa)、野生稻(oryzaaustraliensis)、短舌野生稻(oryzabarthii)、非洲型稻(oryzaglaberrima)、阔叶稻(oryzalatifolia)、长雄野生稻(oryzalongistaminata)、南方野生稻(oryzameridionalis)、药用野生稻(oryzaofficinalis)、斑点野生稻(oryzapunctata)、普通野生稻(oryzarufipogon)(红稻)、印度野生稻(oryzanivara)、拟南芥(arabidopsisthaliana)、玉米(zeamays)、大豆(glycinemax)、烟豆(glycinetabacina)、野大豆(glycinesoja)和短绒野大豆(glycinetomentella)。
在另一方面,本发明包括抑制dna构建体。
一种抑制dna构建体包含至少一个调控序列(如植物中有功能的启动子)和与其可操作连接的(a)全部或部分(i)一种核酸序列,其编码的多肽的氨基酸序列与seqidno:16相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(ii)核酸序列(i)的全长互补序列;或(b)源于目标基因的正义链或反义链的全部或部分的区域,所述区域的核酸序列与该目标基因的正义链或反义链的全部或部分相比具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%的序列一致性,其中所述目标基因编码花期调控多肽mbd2;或者(c)全部或部分(i)与seqidno:15的核苷酸序列一致性至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%的核苷酸序列,或者(ii)核酸序列(c)(i)的全长互补序列。抑制dna构建体包括反义构建体、病毒抑制构建、发夹抑制构建体、茎-环抑制构建体、双链rna产生构建体、rnai构建体或小rna构建体(例如sirna构建体或mirna构建体)。
应当理解(正如本领域技术人员将会理解的),本发明不仅仅涵盖这些具体的示例性序列。导致给定位点处产生化学上等价的氨基酸但不影响所编码多肽的功能特性的核酸片段中的改变是本领域众所周知的。例如,丙氨酸(一种疏水性氨基酸)的密码子可被编码另一个疏水性较弱的残基(例如甘氨酸)或疏水性较强的残基(例如缬氨酸、亮氨酸或异亮氨酸)的密码子取代。类似地,导致一个带负电荷的残基替换为另一个带负电荷的残基(例如,天冬氨酸替代谷氨酸)或者一个带正电荷的残基替换为另一个带正电荷的残基(例如,赖氨酸替换精氨酸)的改变也可预期产生功能上等价的产物。导致多肽分子的n-末端和c-末端部分改变的核苷酸变化也将预计不会改变多肽的活性。所提出的修饰中的每一种均完全在本领域常规技术内,如测定所编码的产物的生物活性的保留。
“抑制dna构建体”是在转化或稳定整合进植物基因组时,导致该植物中的靶基因“沉默”的重组dna构建体。对该植物来说,该靶基因可为内源性的基因或是转入的基因。如本文针对靶基因所使用的,“沉默”通常指在由靶基因表达的mrna或蛋白质/酶的水平上的抑制,和/或在酶活性或蛋白质功能性的水平上的抑制。本文中可交换使用的术语“抑制”、“抑制性”以及“沉默”包括降低、减少、减退、减小、抑制、消除或防止。“沉默”或“基因沉默”不特指机理并且包括但不限于反义、病毒-抑制、发夹抑制、茎-环抑制、基于rnai的方法以及基于小rnai的方法。
抑制dna构建体可包含源自所关注的靶基因的区域并且可包含所关注的靶基因的有义链(或反义链)的核酸序列的全部或部分。取决于所要利用的方法,所述区域可与所关注基因的有义链(或反义链)的全部或部分100%相同或者具有少于100%序列的一致性(如,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%的一致性)。
抑制dna构建体是本领域所熟知的,一旦选定所关注的靶基因就很容易构建,并且包括但不限于反义构建体、病毒-抑制构建体、发夹抑制构建体、茎-环抑制构建体、产生双链rna的构建体,以及更通常的是,rnai(rna干扰)构建体和小rna构建体,例如sirna(短干扰rna)构建体和mirna(微rna)构建体。
“反义抑制”指产生能够抑制靶基因或基因产物表达的反义rna转录物。“反义rna”指与靶初级转录物或mrna的全部或部分互补,并阻断分离的靶核酸片段表达的rna转录物(美国专利号5,107,065)。反义rna可与特定基因转录物的任何部分,即5′非编码序列、3′非编码序列、内含子或编码序列互补。
rna干扰(rnai)是指由短干扰性rna(sirna)介导的动物中序列特异性转录后基因沉默的过程(fire等人,(1998)nature391:806)。在植物中的对应过程通常称为转录后基因沉默(ptgs)或rna沉默,并且在真菌中也称为阻抑作用(quelling)。据信ptgs转录后基因沉默过程是用于防止外来基因表达的进化保守性细胞防御机制,并且通常由不同植物区系和门所共有(fire等人,(1999)trendsgenet.15:358)。
小rna在控制基因表达中起重要作用。很多发育过程(包括开花)的调节是由小rna控制的。现在有可能通过使用在植物中产生小rna的转基因构建体来以基因工程手段改变植物基因的表达。
小rna似乎是通过与互补rna或dna靶序列碱基配对来行使功能的。当与rna结合时,小rna或者引发靶序列的rna裂解或者引发翻译抑制。当与dna靶序列结合时,据信小rna可介导靶序列的dna甲基化。无论具体机制是什么,这些事件的后果是基因表达受到抑制。
微rna(mirna)是长度为约19至约24个核苷酸(nt)的已经在动物和植物中鉴定出的非编码rna(lagos-quintana等人,(2001)science294:853-858;lagos-quintana等人,(2002)curr.biol.12:735-739;lau等人,(2001)science294:858-862;lee和ambros,science294:(2001)862-864;llave等人,(2002)plantcell14:1605-1619;mourelatos等人,(2002)genes.dev.16:720-728;park等人,(2002)curr.biol.12:1484-1495;reinhart等人,(2002)genes.dev.16:1616-1626)。它们是由大小为大约70至200nt较长的前体转录物加工生成的,并且这些前体转录物能够形成稳定的发夹结构。
微rna(mirna)看起来通过与位于由这些基因产生的转录物中的互补序列结合来调节靶基因。看起来有可能mirna可进入至少两条靶基因调控途径:(1)翻译抑制;和(2)rna裂解。进入rna裂解途径的微rna类似于在动物中的rna干涉作用(rnai)和植物中的转录后基因沉默(ptgs)期间生成的21-25nt的短干涉rna(sirna),并且可能掺入了rna诱导沉默复合物(risc),该复合物与rnai相似或相同。
调控序列:
本发明的重组dna构建体(包括抑制dna构建体)包含至少一个调控元件。
调控元件可以是启动子或增强子。
多个启动子可用于本发明的重组dna构建体,启动子根据期望的结果选择,可包括宿主生物体中表达的组成型、组织特异型、诱导型、或其他启动子。
一般将引起基因在多数细胞类型中在多数情况下表达的启动子称为“组成型启动子”。
当组成型启动子驱动候选基因表达时能够评估候选基因的效应,但候选基因在35s或ubi启动子控制下的高水平、组成型表达可具有多重效应。使用组织特异和/或胁迫特异启动子可消除不需要的效应但保留提高植物耐旱性的能力。在拟南芥属中已经观察到了该效应(kasuga等人(1999)naturebiotechnol.17:287-91)。
适用于植物宿主细胞的组成型启动子包括(例如)rsyn7启动子的核心启动子和在wo99/43838和美国专利6,072,050中公开的其他组成型启动子;camv35s核心启动子(odell等人,(1985)nature313:810-812);稻肌动蛋白(mcelroy等人,(1990)plantcell2:163-171);泛素启动子(christensen)等人,(1989)plantmol.biol.12:619-632和christensen等人,(1992)plantmol.biol.18:675-689);pemu(last等人,(1991)theor.appl.genet.81:581-588);mas(velten等人,(1984)emboj.3:2723-2730);als启动子(美国专利5,659,026)等。其他组成型启动子包括例如在美国专利5,608,149、5,608,144、5,604,121、5,569,597、5,466,785、5,399,680、5,268,463、5,608,142和6,177,611中公开的那些启动子。
在选择启动子用于本发明方法时,期望使用组织特异性启动子或发育调节启动子。
组织特异性启动子或发育调节启动子是这样的dna序列,其调节dna序列选择性地在对雄穗发育、结籽或两者重要的植物细胞/组织中表达,并限制这种dna序列只在植物的雄穗发育或种子成熟期间表达。任何引起所需时空表达的可鉴定启动子均可用于本发明的方法中。
本领域的技术人员知道多种叶片优选的启动子(yamamoto等(1997)plantj.12(2):255-265;kwon等(1994)plantphysiol.105:357-367;yamamoto等(1994)plantcellphysiol.35(5):773-778;gotor等(1993)plantj.3:509-518;orozco等(1993)plantmol.biol.23(6):1129-1138;和matsuoka等(1993)proc.natl.acad.sci.usa90(20):9586-9590)。
可用于本发明的种子或胚特异性启动子包括大豆kunitz胰蛋白酶抑制剂(kti3,jofuku和goldberg,(1989)plantcell1:1079-1093),convicilin、豌豆球蛋白和豆球蛋白(豌豆子叶)(rerie,w.g.等,(1991)mol.gen.genet.259:149-157;newbigin,e.j.等,(1990)planta180:461-470;higgins,t.j.v.等,(1988)plant.mol.biol.11:683-695),玉米蛋白(玉米胚乳)(schemthaner,j.p.等,(1988)emboj.7:1249-1255),菜豆蛋白(菜豆子叶)(segupta-gopalan,c.等,(1985)proc.natl.acad.sci.u.s.a.82:3320-3324),植物血球凝集素(菜豆子叶)(voelker,t.等,(1987)emboj.6:3571-3577),b-伴球蛋白和大豆球蛋白(大豆子叶)(chen,z-l等,(1988)emboj.7:297-302),谷蛋白(稻胚乳),大麦醇溶蛋白(大麦胚乳)(marris,c.等,(1988)plantmol.biol.10:359-366),麦谷蛋白和麦醇溶蛋白(小麦胚乳)(colot,v.等,(1987)emboj.6:3559-3564)。可操作地连接至嵌合基因构建体异源编码区的种子特异性基因的启动子在转基因植物中保持它们的时空表达模式。这样的实施例包括在拟南芥和甘蓝型油菜(brassicanapus)种子中表达脑啡肽的拟南芥属2s种子储藏蛋白基因启动子(vanderkerckhove等,(1989)bio/technology7:l929-932),表达荧光素酶的菜豆凝集素和β-菜豆蛋白启动子(riggs等,(1989)plantsci.63:47-57),以及表达氯霉素乙酰转移酶的小麦谷蛋白启动子(colot等,(1987)emboj6:3559-3564)。
诱导启动子响应内源性或外源性刺激的存在,例如,通过化合物(化学诱导剂),或响应环境、激素、化学信号和/或发育信号而选择性表达可操纵连接的dna序列。诱导启动子或受调控的启动子包括(例如)受光、热、胁迫、水涝或干旱、植物激素、创伤或诸如乙醇、茉莉酮酸酯、水杨酸或安全剂之类的化学品调控的启动子。
用于本发明的启动子包括以下启动子:1)胁迫诱导型rd29a启动子(kasuga等,(1999)naturebiotechnol.17:287-91);2)胁迫诱导启动子rab17(vilardell等,(1991)plantmol.bio.17:985-993;kampbusk等(1997)plantj11(6):1285-1295);3)大麦启动子b22e;b22e是发育中玉米籽粒中的柄所特异表达的启动子(“primarystructureofanovelbarleygenedifferentiallyexpressedinimmaturealeuronelayers”(在未成熟糊粉层中差异表达的新大麦基因的一级结构)klemsdal,s.s.等,(1991)mol.gen.genet.228(1/2):9-16);和4)玉米启动子zag2(“identificationandmolecularcharacterizationofzag1,themaizehomologofthearabidopsisfloralhomeoticgeneagamous”,schmidt,r.j.等,(1993)plantcell5(7):729-737;“structuralcharacterization,chromosomallocalizationandphylogeneticevaluationoftwopairsofagamous-likemads-boxgenesfrommaize”,theissen等,(1995)gene156(2):155-166;ncbigenbank登录号x80206))。zag2转录物可在授粉前5天至授粉后(dap)7至8天被检测到,并且引导ciml在发育中的雌花序的心皮中表达,ciml是发育玉米籽粒的籽仁特异的启动子。ciml转录物在授粉前4至5天至授粉后6至8天被检测到。其他可用的启动子包括可源自其表达与发育中的雌小花母系相关的基因的任何启动子。
对于在发育种子组织中表达的多聚核苷酸,特定的启动子包括种子优选的启动子,尤其是早期籽粒/胚胎启动子和晚期籽粒/胚乳启动子,授粉后籽粒的发育大致可以分为三个基本阶段,籽粒生长的停滞期起始于授粉后0天到10-12天,在此期间,籽粒不再明显的生长,但是决定籽粒活力的重要事件将在此期间发生(如细胞建成数目)。线性籽粒灌浆期起始于授粉后的10-12天并延续到授粉后的40天左右,在此籽粒发育期间,籽粒达到最终的质量,并产生多种储藏物质如淀粉、蛋白质和油等;成熟期起始于授粉后大约40天到收获,在这一过程中,籽粒开始休眠,变干并为萌芽前的种子休眠做准备。本发明中的“早期籽粒/胚乳启动子”指主要在种子发育的停滞期(也就是授粉第0天到授粉后第12天期间)驱动基因表达的启动子;“后期籽粒/胚乳启动子”主要驱动基因在授粉后12天到成熟过程中的种子中基因表达;表达窗口可能会有一些重叠,将根据使用的aba偶联的序列和期望的表型选择启动子。
早期籽粒/胚胎启动子包括,cim1,其在授粉后第5天活跃在特定组织中(wo00/11177);其它的早期籽粒/胚胎启动子包括种子偏爱启动子end1,其在授粉后7-10天表达,和end2,其在授粉后9-14天在整个籽粒中表达,在授粉后10天在胚乳和果皮中表达(wo00/12733),此处以全文引用方式并入本文。本发明中的特定方法使用的其它早期籽粒/胚乳启动子包括种子偏爱启动子ltp2(美国专利号5,525,716);玉米zm40启动子(美国专利号6,403,862);玉米nuc1c(美国专利号6,407,315);玉米ckx1-2启动子(美国专利号6,921,815和美国专利申请公开号2006/0037103);玉米lec1启动子(美国专利号7,122,658);玉米esr启动子(美国专利号7,276,596);玉米zap启动子(美国专利申请公开号20040025206和20070136891);玉米启动子eep1(美国专利申请公开号20070169226);和玉米启动子adf4(美国专利申请号60/963,878,2007年8月7日申请)。
本发明中调控核酸序列在植物中表达的其它启动子是茎特异启动子,包括苜蓿s2a启动子(genbank登录号ef030816;abrahams等(1995)plantmol.biol.27:513-528)和s2b启动子(genbank登录号ef030817)和类似的启动子。
启动子可以整个源于天然基因,或者由源于不同的天然存在的启动子的不同元件组成,或者甚至包括合成的dna片段。
用于本发明特定实施例的启动子包括:rip2、mlip15、zmcor1、rab17、camv35s、rd29a、b22e、zag2、sam合成酶、泛素、camv19s、nos、adh、蔗糖合成酶、r-等位基因、维管组织偏爱启动子s2a(genbank登录号ef030816)和s2b(genbank登录号ef030817)及来自玉米的组成型启动子gos2。其他启动子还包括根偏爱启动子,例如玉米nas2启动子、玉米cyclo启动子(us2006/0156439,公开于2006年7月13日)、玉米rootmet2启动子(wo05063998,公开于2005年7月14日)、crlbio启动子(wo06055487,公开于2006年5月26日)、crwaq81(wo05035770,公开于2005年4月21日)和玉米zrp2.47启动子(ncbi登录号:u38790;gino.1063664)。
本发明的重组dna构建体也可包括其他调控序列,包括但不限于翻译前导序列、内含子和多腺苷酸化识别序列。在特定实施方案中,重组dna构建体还包括增强子或沉默子。
内含子序列可加至5’非翻译区、蛋白编码区或3’非翻译区以增加积聚在胞浆中的成熟信息的量。已经显示,在植物和动物两者的表达构建体的转录单位中包含可剪接内含子可使基因表达在mrna和蛋白质水平上均增强高达1000倍。参见buchman和berg,(1988)mol.cellbiol.8:4395-4405;callis等,(1987)genesdev.1:1183-1200。
任何植物都可以选择用来鉴定将用于本发明重组dna构建体的调控序列和基因。适用于分离基因和调控序列的靶植物的实例应该包括但不限于苜蓿、苹果、杏、拟南芥、洋蓟、芝麻菜、芦笋、鳄梨、香蕉、大麦、豆类、甜菜、黑莓、蓝莓、西兰花、抱子甘蓝、卷心菜、加拿大油菜、香瓜、胡萝卜、木薯、蓖麻子、菜花、芹菜、樱桃、菊苣、芫荽、柑桔、克莱门氏小柑橘、三叶草、椰子、咖啡、玉米、棉、蔓越莓、黄瓜、花旗松、茄子、菊苣、茅菜、桉树、茴香、无花果、大蒜、葫芦、葡萄、柚子树、白兰瓜、豆薯、猕猴桃、生菜、韭葱、柠檬、酸橙、火炬松、亚麻子、芒果、甜瓜、蘑菇、油桃、坚果、燕麦、油棕、油菜、秋葵、橄榄树、洋葱、橙、观赏植物、棕榈、木瓜树、欧芹、欧洲防风草、豌豆、桃树、花生、梨树、胡椒、柿树、松树、菠萝、车前草、李树、石榴树、白杨、马铃薯、南瓜、温柏、辐射松、红菊苣、萝卜、油菜、树莓、稻、黑麦、高粱、南方松、大豆、菠菜、南瓜、草莓、甜菜、甘蔗、向日葵、甘薯、枫香树、柑橘、茶、烟草、蕃茄、黑小麦、草皮草、芜菁、葡萄树、西瓜、小麦、薯蓣和西葫芦。
组合物:
本发明的组合物是其基因组中包含本发明的任何重组dna构建体或抑制dna构建体(例如上面所讨论的任何一种构建体)的植物。组合物还包括所述植物的任何子代,以及获取自所述植物或其子代的任何种子,其中所述子代或种子在其基因组中包含重组dna构建体或抑制dna构建体。所述子代包括植物通过自花授粉或异型杂交而获得的连续世代。子代也包括杂交种和自交系。
在杂交种子繁殖的农作物中,成熟的转基因植物可自花授粉而产生纯合的自交系植物。该自交系植物产生的种子含有新导入的重组dna构建体或抑制dna构建体。这些种子可生长成植物,所述植物表现出改变的农学特性(如水分限制条件下增加农业特性),或者可用于育种程序以产生杂交种子,这些杂交种子可生长成植物,所述植物将会表现出如改变的农学特性。种子可以是玉米种子或水稻种子。
所述种子可为玉米种子或水稻种子。植物可为单子叶植物或双子叶植物,例如水稻、玉米或大豆植物,如玉米杂种植物或玉米自交系植物。植物还可为向日葵、高梁、油菜、小麦、苜蓿、棉花、大麦、粟、甘蔗或柳枝稷。
重组dna构建体可稳定地整合进植物的基因组中。
实施方案包括但不限于以下实施方案:
1.在其基因组中包含重组dna构建体的转基因植物,如水稻、玉米或大豆,所述重组dna构建体包含一种多核苷酸和与其可操作连接至少一个调控序列,其中所述多核苷酸编码一个多肽,所述多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;与对照植物相比,所述植物表现出增加的耐旱性和/或百草枯耐性,且所述植物进一步表现出至少一个改变的农艺性状。
2.在其基因组中包含重组dna构建体的转基因植物,如水稻、玉米或大豆,所述重组dna构建体包含一个多核苷酸和与其可操作连接至少一个调控序列,其中所述多核苷酸编码一个多肽,与对照植物相比,所述植物表现出增加的耐旱性和/或百草枯耐性,且所述植物进一步表现出至少一个改变的农艺性状。
3.在其基因组中包含重组dna构建体的转基因植物,例如水稻、玉米或大豆,所述重组dna构建体包含一个多核苷酸和与其可操作连接的至少一个调控序列,其中所述多核苷酸编码一个多肽,与对照植物相比,所述植物表现出至少一个农艺性状的变化。
4.在其基因组中包含重组dna构建体的转基因植物,如水稻、玉米或大豆,所述重组dna构建体包含一种多核苷酸和与其可操作连接的至少一调控序列,其中所述多核苷酸编码一个多肽,所述多肽的氨基酸序列与seqidno:16相比,具有50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;与对照植物相比,所述植物表现出较早的开花时间,且所述植物进一步表现为至少一个农艺性状的变化。
5.在其基因组中包含抑制dna构建体的转基因植物,如水稻、玉米或大豆,所述抑制dna构建体包含至少一个调控序列和与其可操作连接的源于靶基因正义链或反义链全部或部分片段,所述片段的核苷酸序列与所述靶基因的正义链或反义链的全部或部分序列一致性至少为50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%;所述靶基因编码开花时间调控的多肽;与对照植物相比,所述植物表现出延迟的开花时间。
6.在其基因组中包含抑制dna构建体的转基因植物,如水稻、玉米或大豆,所述抑制dna构建体包含至少一个调控序列和与其可操作连接的全部或部分(a)一种核酸序列,其编码一个多肽,所述多肽的氨基酸序列与seqidno:16相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;或(b)核酸序列(a)的全长互补序列,其中与对照植物相比,所述植物表现出延迟的开花时间。
7.实施方案1-6所述的植物的任何子代植物,实施方案1-6所述的植物的任何种子,实施方案1-6所述的植物的任何子代植物的种子,和源于实施方案1-6的植物和其子代植物的细胞。
前述实施方案1-7或其它实施方案中,所述耐旱性多肽可来自水稻(oryzasativa)、野生稻(oryzaaustraliensis)、短舌野生稻(oryzabarthii)、非洲型稻(oryzaglaberrima)、阔叶稻(oryzalatifolia)、长雄野生稻(oryzalongistaminata)、南方野生稻(oryzameridionalis)、药用野生稻(oryzaofficinalis)、斑点野生稻(oryzapunctata)、普通野生稻(oryzarufipogon)(红稻)、印度野生稻(oryzanivara)、拟南芥(arabidopsisthaliana)、玉米(zeamays)、大豆(glycinemax)、烟豆(glycinetabacina)、野大豆(glycinesoja)和短绒野大豆(glycinetomentella)。
前述实施方案1-7或其它实施方案中,重组dna构建体或抑制dna构建体包含至少一个植物中有功能的启动子作为调控序列。
前述实施方案1-7或其它实施方案中,至少一种农艺性状的变化可以使增加也可以是减少。
前述实施方案1-7或其他实施方案中,至少一种农艺性状选自叶片绿色、籽粒产量、生长速率、生物量、成熟时的鲜重、成熟时的干重、果实产量、种子产量、植物总氮含量、果实氮含量、种子氮含量、植物营养组织氮含量、植物总游离氨基酸含量、果实游离氨基酸含量、种子游离氨基酸含量、植物营养组织游离氨基酸含量、植物总蛋白含量、果实蛋白含量、种子蛋白含量、植物营养组织蛋白含量、耐旱性、氮吸收能力、根倒伏、收获指数、茎倒伏、株高、穗高、穗长、耐盐性、分蘖数、圆锥花序大小、幼苗活力和低温胁迫下的出苗情况。例如,至少一个农艺性状的改变可以是籽粒产量、叶绿素含量或生物量的增加。
前述1-7实施方案或其它实施方案中,与对照植物相比,所述植物在水分限制条件下表现出至少一个农艺性状的变化。
前述实施方案1-7其它实施方案中,与对照植物相比,所述植物在氧化胁迫即百草枯胁迫下表现出至少一个农艺性状的变化。
“干旱”指植物可用水分的减少,特别是较长时间的缺水或者在植物重要的生长阶段发生,会造成植物损伤,或阻止植物的生长(限制植物的生长,降低籽粒的产量)。
“耐旱性”指干旱胁迫下能够存活并且没有实质性的生理或物理改变的能力,和/或一段时间干旱后复水能够恢复的能力。
多肽的“耐旱能力”表示与参照或对照植物相比,过表达该多肽可以提高转基因植物的耐旱性能。
植物“增强的耐旱性”是与参照或对照植物相比测定的,反映了植物在干旱胁迫下存活的能力,并且与参照或对照相比,具有较小的生理或物理损伤,或者干旱胁迫后复水时,恢复较快的能力。
“环境条件”指植物生长的环境,诸如可用的水分、可用的营养物质或昆虫或病害的存在。
“百草枯”(1,1-二甲基-4,4-联吡啶二氯化物)是一类叶片喷施的非选择性的吡啶除草剂,能够引起光氧化胁迫,并进一步造成植物的损害或者阻止植物的正常生长。
“百草枯耐性”是植物的一种性状,反映了百草枯溶液处理后,与参照或对照植物相比,植物的存活或生长良好的能力。
植物“增加的百草枯耐性”是相对于参照或对照植物测定的,反映了植物在百草枯溶液处理后,与参照或对照植物相比,能够存活并具有较小的生理或物理伤害的能力。通常,针对较低浓度的百草枯溶液的耐性用作诸如干旱胁迫的非生物胁迫耐性的一种指标。
“氧化胁迫”反映了活性氧分子的生成和生物系统清除活性氧中间体或修复损伤的能力之间的不平衡。扰乱细胞正常的氧化还原状态会导致产生过氧化氢和自由基的毒性效应,过氧化氢和自由基能够损害包括蛋白、脂质和dna在内的细胞组成部件。
下面的例子描述一些代表性的方法或技术用于模拟干旱条件和/或评估耐旱性;模拟氧化条件。
本领域的技术人员也可以测试模拟或者自然发生的干旱条件下的植物保持足够产量(至少75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或者100%的产量)的能力评估耐旱性,例如,干旱条件下产生与非干旱条件下相比实质相当的产量;与对照或参照植物相比,干旱条件下产量减少较少。
本发明中涉及的参数包括与对照细胞或对照植物相比的恢复度、存活率、百草枯抗性率、基因表达水平、水分利用效率、编码蛋白的水平或活性和其他参数。“对照”、“对照植物”或“对照植物细胞”为测定受测试植物或植物细胞的表型变化提供参考,由于转化,受测植物或植物细胞的基因组改变影响到目的基因,测试的植物或植物细胞可以是转基因植物或转基因植物细胞的子代,并含有上述的基因的改变。本领域技术人员很容易找到适当的对照或参照植物,当采用本发明描述的组合物或方法评估或测定转基因植物一个农艺性状或表型时。例如,但不限于下述举例:
1.转化植物的子代,所述转化植物对于重组dna构建体或抑制dna构建体来说是半合子的,所述子代分离成包含或不包含该dna构建体或抑制dna构建体的植株:包含该重组dna构建体或抑制dna构建体的子代将通常相对于不包含该重组dna构建体或抑制dna构建体的子代来进行测量(即,不包含该重组dna构建体或抑制dna构建体的子代是对照或参照植物)。
2.重组dna构建体或抑制dna构建体基因渗入至近交系中,例如在水稻和玉米中,或基因渗入进变种中,例如在大豆中:基因渗入品系将通常相对于亲本近交系或变种品系进行测量(即,亲本近交系或品种品系是对照或参照植物)。
3.双杂交系,其中第一杂交系由两个亲本近交系产生,而第二杂交系由相同的两个亲本近交系产生,不同的是其中一个亲本近交系含有重组dna构建体或抑制dna构建体:第二杂交系通常将相对于第一杂交系进行测量(即第一杂交系为对照植物或参照植物)。
4.包含重组dna构建体或抑制dna构建体的的植物:该植株可以相对于这样的对照植物进行评价或测量,该对照植物不包含重组dna构建体或抑制dna构建体,但具有与该植株相当的遗传背景(例如,与包含重组dna构建体或抑制dna构建体的植株相比较,该核遗传物质具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列同一性)。存在许多可用于分析、比较和表征植物遗传背景的实验室的技术;其中这些技术是同工酶电泳、限制性片段长度多态性(rflps)、随机扩增多态性dna(rapds)、任何引物聚合成酶链式反应(ap-pcr)、dna扩增指纹(daf)、序列特异扩增区域(scars)、扩增片段长度多态性
方法:
方法包括但不限于:提高植物耐旱性的方法,评估植物耐旱性的方法,提高植物百草枯耐受性的方法,改变植物农艺性状的方法,确定植物农艺性状变化的方法,调控植物开花时间的方法,观察和/或评估植物农艺性状的方法,和生产种子的方法。植物可以是单子叶植物,也可以是双子叶植物,例如水稻、玉米或大豆,植物也可以是向日葵、油菜、小麦、苜蓿、棉花、大麦、粟、甘蔗或高粱,种子是玉米或大豆种子,例如玉米杂交种子或玉米自交种子。
方法包括但不限于:
转化细胞方法包括将本发明公开的任意一个或多个分离的多核苷酸转化细胞,其中,在特定的实施方案中,所述细胞是真核细胞,如酵母、昆虫或植物细胞;或原核细胞如细菌细胞。
产生转基因植物的方法包括转化将本发明公开的任意分离的核苷酸或重组dna构建体或抑制dna构建体转化植物细胞和由转化的植物细胞再生转基因植物,其中本方法获得转基因植物和转基因种子可用于本发明的其他方法。
从细胞或细胞培养物中分离本发明的多肽的方法,其中,所述细胞包含重组dna构建体,所述重组dna构建体包含本发明的一个多核苷酸和与其可操作连接的至少一个调控序列,所述宿主细胞在适于重组dna构建体表达的条件下生长。
改变本发明多肽在宿主细胞中表达水平的方法包括:(a)将本发明重组dna构建体转化宿主细胞;和(b)使转化宿主细胞在适于重组dna构建体表达的条件下生长,其中重组dna构建体的表达导致转化宿主细胞中本发明多肽含量的变化。
提高植物耐旱性的方法,包括增加植物中编码icdh1、mtn3l、dn-dtp6、ankl1、mbd2、tp1、acoat1、dn-dtp7多肽的至少一个多核苷酸的表达,其中,所述多核苷酸包括:(a)一种多核苷酸,其核苷酸序列与seqidno:2、5、8、11、14、17、20或23的一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:3、6、9、12、15、18、21或24的一致性至少为85%;和(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25的一致性至少为90%。
提高植物耐旱性和/或百草枯耐受性的方法,包括(a)将重组dna构建体导入到可再生的植物细胞中,所述重组dna构建体包含一种多核苷酸和与其可操作地连接至少一种调控序列(例如在植物中有功能的启动子),其中所述多核苷酸编码多肽的氨基酸序列在与seqidno:4、7、10、13、16、19、22或25进行比较时,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(b)在步骤(a)之后,由所述可再生的植物细胞再生转基因植物,其中所述转基因植物在其基因组中包含所述重组dna构建体并且在与对照植物进行比较时表现出提高的耐旱性和/或百草枯耐性;和(c)获得来源于所述转基因植物的子代植物,其中所述子代植物在其基因组中包含重组dna构建体,与对照植物相比,表现出提高的耐旱性和/或百草枯耐性。
评估植物耐旱性和/或百草枯耐性的方法,包括(a)获得转基因植物,所述转基因植物在其基因组中包含重组dna构建体,所述重组dna构建体包含一个多核苷酸和与其可操作连接的至少一个调控序列(如,植物中有功能的启动子)其中,所述多核苷酸编码一个多肽,所述多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(b)获得源于所述转基因植物的子代植物,其中所述子代植物在其基因组中包含重组dna构建体;和(c)与对照植物相比,评估子代植物的耐旱性和/或百草枯耐性。
确定植物一种农艺性状变化的方法,包括(a)获得转基因植物,所述转基因植物在其基因组中包含重组dna构建体,所述重组dna构建体包含一个多核苷酸和与其可操作连接至少一个调控序列(如,植物中有功能的启动子),其中所述多核苷酸编码一个多肽,所述多肽的氨基酸序列与seqidno:4、7、10、13、16、19、22或25相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;(b)获得源于所述转基因植物的子代植物,其中所述子代植物的基因组中包含重组dna构建体;和(c)水分限制条件下,确定与对照植物相比,所述子代植物是否表现出至少一个农艺性状的变化。
调控植物开花时间的方法,包括(a)将重组dna构建体导入到可再生的植物细胞中,所述重组dna构建体包含一个核苷酸和与其可操作地连接至少一种调控序列(例如在植物中有功能的启动子),其中所述多核苷酸编码一个多肽,所述多肽的氨基酸序列与seqidno:16相比,具有至少50%、51%、52%、53%、54%、55%、56%、57%、58%、59%、60%、61%、62%、63%、64%、65%、66%、67%、68%、69%、70%、71%、72%、73%、74%、75%、76%、77%、78%、79%、80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%的序列一致性;和(b)在步骤(a)之后,由所述可再生的植物细胞再生转基因植物,其中所述转基因植物在其基因组中包含所述重组dna构建体并且在与不包含所述重组dna构建体的对照植物进行比较时表现出改变的开花时间。所述方法还可包括(c)获得来源于所述转基因植物的子代植物,其中所述子代植物在其基因组中包含重组dna构建体并且在与不包含所述重组dna构建体的对照植物进行比较时表现改变的开花时间。
提高植物低氮耐性或nue的方法,包括增加编码mtn3l或ankl1多肽的至少一个多核苷酸在植物中表达,所述多核苷酸包括:(a)一种多核苷酸,其核苷酸序列与seqidno:5或11的一致性至少为85%;(b)一种多核苷酸,其核苷酸序列与seqidno:6或12的一致性至少为85%;和(c)一种多核苷酸,其编码的多肽的氨基酸序列与seqidno:7或13的一致性至少为90%,其中,所述多核苷酸的表达通过以下步骤增加:(a)通过重组dna构建体增加编码mtn3l或ankl1多肽的多核苷酸在植物中的表达,所述重组dna构建体包含编码mtn3l或ankl1多肽的多核苷酸和与其可操作连接的至少一个异源调控元件,所述多核苷酸编码多肽的氨基酸序列与seqidno:7或13的序列一致性至少为90%;或(b)增加内源多核苷酸的表达,所述内源多核苷酸编码的多肽的氨基酸序列与seqidno:7或13的序列一致性至少为90%。
生产种子的方法包括前述的任一方法,进一步还包括从所述子代植物获得种子,其中所述种子在其基因组中包含重组dna构建体或抑制dna构建体。
在前述任一的方法或本文其他实施方案披露的任意方法中,所述引入步骤中所述再生植物细胞可包括愈伤细胞、胚胎愈伤细胞、配子细胞、分生细胞、或不成熟的胚胎细胞。可再生植物细胞可源于自交玉米植株。
在前述任一方法或本文披露的其他实施方案中的任一方法中,所述再生步骤可包括:(i)在含有胚促激素培养基上培养所述转化的植物细胞直至长出愈伤组织;(ii)将步骤(i)所述的转化植物细胞转移至含有组织促激素的第一培养基上;和(iii)将步骤(ii)所述转化植物细胞接种到第二培养基上,使其茎伸长、根发育或两者均发育。
在前述任一方法或本发明的其他实施方案的方法中,确定转基因植物农艺性状的变化的步骤,如果可行,可包括在可变的环境条件下,确定与不含有重组dna构建体的对照植物相比,转基因植物是否表现出至少一个农艺性状的变化。
在前述任一方法或者本发明其他实施方案的任一方法中,如果可能,确定子代植物农艺性状变化的步骤包括可变的环境条件下,确定与不含有重组dna构建体的对照植物相比,所述子代植物是否表现出至少一个农艺性状的变化。
在前述任一方法或者本发明其他实施方案的任一方法中,与对照植物相比,植物在水分限制条件下表现出至少一个农艺性状的变化。
在前述任一方法或者本发明其他实施方案的任一方法中,存在将重组dna构建体导入可再生植物的供选方案,所述重组dna构建体包含一个多核苷酸和与其可操作连接的至少一个调控序列,例如可以将一个调控序列(如一个或多个增强子、任选的转座子元件的一部分)导入可再生植物细胞,接着筛选带有调控序列和与其可操作连接的编码本发明多肽的内源基因的转基因事件。
可以通过任何适当的技术将本发明的重组dna构建体导入植物中,所述技术包括但不限于直接dna吸收、化学药品处理、电穿孔、显微注射、细胞融合、感染、载体介导的dna转移、基因枪轰击或农杆菌的转化。植物转化和再生的技术在国际专利公开号wo2009/006276中描述,其全部内容并入作为参考。
另外,也有修饰或改变宿主内源基因组dna的方法,包括改变宿主天然dna序列或包括调控元件、编码或非编码序列等前体转基因序列。这些方法也可以用于将核酸序列靶向到基因组中改造目标识别序列。例如本文中转基因修饰的细胞或植物使用传统的基因工程核酸酶如产生修饰植物基因组的归位核酸内切酶产生(例如wo2009/114321;gao等(2010)plantjournal1:176-187)。其他的位点定向工程是通过使用锌指结构域识别偶联的限制性内切酶的限制性特点修饰内源基因(例如urnov等(2010)natrevgenet.11(9):636-46;shukla等(2009)nature459(7245):437-41)。转录激活样(tal)效应因子-dna修饰酶(transcriptionactivator-likeeffector-dnamodifyingenzyme,tale或talen)可用于基因工程修饰植物基因组,参见例如us20110145940,cermak等(2011)nucleicacidsres.39(12)和boch等(2009),science326(5959):1509-12。植物基因组定点修饰也可以使用细菌ii型crispr(成簇的规律的间隔的短回文的重复序列,clusteredregularlyinterspacedshortpalindromicrepeats)/cas(crispr关联蛋白,crispr-associated)系统,参考例如belhaj等(2013),plantmethods9:39.crispr/cas系统可以允许可定制的小的非编码rna引导的基因组dna靶向切割。
本领域的技术人员熟悉将编码期望多肽的外源基因转化植物和培育再生植物的方法。再生植物可以自花授粉产生纯合子转基因植物,或者将再生植物的花粉与种子长出的农艺重要的植物杂交,或者农艺重要的植物的花粉与再生的转基因植物杂交。本领域的技术人员熟知培育本文披露的含有期望多肽的转基因植物的方法。
转基因植物中性状的重叠
转基因植物可包含本发明公开的一个或多个耐旱性多核苷酸与一个或多个另外的多核苷酸的堆叠,从而导致多个多肽序列的产生或抑制。包含多核苷酸序列堆叠的转基因植物可通过传统育种方法或通过遗传工程方法中的一者或两者来获得。这些方法包括但不限于培育各自包含靶多核苷酸的单独品系,用后续基因转化包含本发明公开的基因的转基因植物,以及将基因共转化进单个植物细胞中。如本发明所用,术语“堆叠”包括具有存在于同一植物中的多种性状(例如,两种性状均掺入核基因组中,一种性状掺入核基因组中且一种性状掺入质体的基因组中,或者两种性状均掺入质体的基因组中)。在一个非限制性例子中,“堆叠性状”包含序列彼此物理相邻的分子堆叠。如本发明所用的性状是指衍自特定序列或序列群组的表型。可使用包含多个基因的单个转化载体或单独地携带于多个载体上的基因进行基因的共转化。如果序列通过遗传转化植株来堆叠,则靶多核苷酸序列可在任何时间以任何顺序进行组合。可以用共转化方案将性状与靶多核苷酸同时引入,所述多核苷酸由转化盒的任何组合提供。例如,如果将要引入两条序列,则可将该两条序列包含在单独的转化盒中(反式)或包含在同一转化盒中(顺式),可通过相同启动子或不同启动子驱动所述序列表达。在某些情况中,可能期望引入会抑制靶多核苷酸的表达的转化盒。这可以与其他抑制盒或过表达盒的任何组合进行组合以在植物中生成所需的性状组合。还认识到可使用位点特异性重组系统在所需的基因组位置堆叠多核苷酸序列。参见,例如,wo1999/25821、wo1999/25854、wo1999/25840、wo1999/25855和wo1999/25853,上述专利均以引用的方式并入本文。
实施例
本文的具体实施进一步在下列实例中示明。在这些实例中,除非特别说明,采用摄氏/公制。在这些实例中,仅用举例说明具体的实施过程。通过上述讨论和具体事例,本领域的专业人员可以查明本发明的基本特征,经过各种改变和修改将本发明应用于各种用途和条件,并不偏离本发明主体和范围。因此,除了本专利表述和讨论的各种修改外,本领域内的专业人员所做的不偏离本发明主题的修改也将落入本专利的权力要求范围内。
实施例1.耐旱基因的克隆和载体的构建
根据水稻激活标签突变体库初步的筛选结果和表2的基因id序列信息,设计引物,克隆了水稻耐旱基因osicdh1、osmtn3l、osdn-dtp6、osankl1、osmbd2、ostp1、osacoat1和osdn-dtp7。引物序列和扩增片段的长度如表3所示。
以中花11号水稻叶、茎和根混合的cdna库为模板克隆osicdh1、osacoat1和osdn-dtp7的cdna;以zh11水稻叶、茎和根混合的基因组dna库为模板克隆osmtn3l、osdn-dtp6、osankl1、osmbd2和ostp1的gdna。pcr反应混合液和pcr程序如表4和5所示。
表2.水稻基因名称、基因id(tigr)和载体的id
表3.克隆水稻耐旱性基因的引物
表4.克隆耐旱基因的pcr反应混合液
表5.克隆耐旱基因的pcr循环状况
pcr扩增产物在琼脂糖凝胶电泳分离后采用柱式试剂盒回收,并与ta克隆载体连接。通过测序确定pcr产物的核酸序列和在构建体中的方向,然后将基因克隆到植物双元载体dp0158(pcambia1300-dsred,seqidno:1)中。
dp0854载体中克隆的核苷酸序列和osicdh1的编码序列如seqidno:2和3所示,osicdh1的氨基酸序列如seqidno:4所示;dp0902载体中克隆的核苷酸序列和osmtn3l的编码序列如seqidno:5和6所示,osmtn3l的氨基酸序列如seqidno:7所示;dp0935载体中克隆的核苷酸序列和osdn-dtp6的编码序列如seqidno:8和9所示,osdn-dtp6的氨基酸序列如seqidno:10所示;dp0960载体中克隆的核苷酸序列和osankl1的编码序列如seqidno:11和12所示,osankl1的氨基酸序列如seqidno:13所示;dp0988载体中克隆的核苷酸序列和osmbd2的编码序列如seqidno:14和15所示,osmbd2的氨基酸序列如seqidno:16所示;dp1082载体中克隆的核苷酸序列和ostp1的编码序列如seqidno:17和18所示,ostp1的氨基酸序列如seqidno:19所示;dp1121载体中克隆的核苷酸序列和osacoat1的编码序列如seqidno:20和21所示,osacoat1的氨基酸序列如seqidno:22所示;dp1176载体中克隆的核苷酸序列和osdn-dtp7的编码序列如seqidno:23和24所示,osdn-dtp7的氨基酸序列如seqidno:25所示。
实施例2.转化获得转基因水稻
过表达载体和空载体(dp0158)采用林拥军和张启发((2005)plantcellrep.23:540-547)描述的农杆菌介导的方法转化入中花11号水稻。中花11号水稻为中国农业科学院作物研究所培育的品种,第一批种子由北京未名凯拓农业生物公司提供。转化实验室获得的t0代转基因幼苗移栽至田间水田中获得t1种子,t1和t2代种子储藏在4℃冷库中。过表达载体含有dsred和hyg基因,t1和t2代种子在绿色荧光灯下发红色荧光的为转基因种子,并用于下列性状验证试验。
实施例3.基因表达分析
采用标准的实时rt-pcr程序诸如源于
实时pcr分析测定了不同转基因水稻株系叶片中osicdh1基因的相对表达水平,zh11-tc叶片中基因的表达水平设置为1.00,其它转基因株系中osicdh1基因表达水平与对照zh11-tc相比,浮动范围在6-376倍之间。zh11-tc为组织培养获得的zh11,dp0158代表转空载体dp0158的zh11。osicdh1基因实时rt-pcr分析的引物如下:
dp0854-f1:5'-caaaaaggaggcgaaactag-3'(seqidno:42)
dp0854-r1:5'-catgtacaagaagagctagatcc-3'(seqidno:43)
实时pcr分析测定了不同转基因水稻株系叶片中osmtn3l基因的相对表达水平,zh11-tc叶片中基因的表达水平设置为1.00,其它转基因株系中osmtn3l基因表达水平与对照zh11-tc相比,浮动范围在134-446倍之间。osmtn3l在绝大多数测试的转基因株系中过量表达。实时rt-pcr的引物如下:
dp0902-f1:5'-tgaaggccaaatcgaccg-3'(seqidno:44)
dp0902-r1:5'-gaagtccataccacaggcag-3'(seqidno:45)
实时pcr分析测定了不同转基因水稻株系叶片中osdn-dtp6基因的相对表达水平,dp0158叶片中基因的表达水平设置为1.00,其它转基因株系中osdn-dtp6基因表达水平与对照dp0158相比,浮动范围在50-508倍之间。osdn-dtp6基因在除dp0935.13以外的大多数转基因株系中过量表达。
dp0935-f1:5'-ttgggacgcttcgagattg-3'(seqidno:46)
dp0935-r1:5'-atctggtccggagtaagataatttc-3'(seqidno:47)
实时pcr分析测定了不同转基因水稻株系叶片中osankl1基因的相对表达水平,dp0158叶片中基因的表达水平设置为1.00,其它转基因株系中osankl1基因表达水平与对照dp0158相比,浮动范围在1.3-168倍之间。大多数转基因株系中osankl1基因的表达水平稍高于zh11-tc植株。
dp0960-f1:5'-ggtatccctagctatcgcaaac-3'(seqidno:48)
dp0960-r1:5'-ttccacgtgaacaggagaac-3'(seqidno:49)
实时pcr分析测定了不同转基因水稻株系叶片中osmbd2基因的相对表达水平,dp0158叶片中基因的表达水平设置为1.00,其它转基因株系中osmbd2基因表达水平与对照dp0158相比,浮动范围在206-1226倍之间(图13)。
dp0988-f1:5'-cataccctgtagacttgcacc-3'(seqidno:50)
dp0988-r1:5'-cgccctcgtttcatagttttc-3'(seqidno:51)
实时pcr分析测定了不同转基因水稻株系叶片中ostp1基因的相对表达水平,dp1082.11叶片中基因的表达水平设置为1.00,其它转基因株系中ostp1基因表达量是dp1082.11中的1-4.8倍。ostp1基因在大多数的转基因株系中过量表达,而在zh11-tc和dp0158中的表达量较低。
dp1082-f1:5'-tgcagttccagaggtctttg-3'(seqidno:52)
dp1082-r1:5'-atcacacgtccatggctg-3'(seqidno:53)
实时pcr分析测定了不同转基因水稻株系叶片中osacoat1基因的相对表达水平,zh11-tc叶片中基因的表达水平设置为1.00,其它转基因株系中osacoat1基因表达水平与对照zh11-tc相比,浮动范围在6.26-33.47倍之间。
dp1121-f1:5'-acgcccatgtgaaagagatc-3'(seqidno:54)
dp1121-r1:5'-caagcacgcatcaaccaaag-3'(seqidno:55)
实时pcr分析测定了不同转基因水稻株系叶片中osdn-dtp7基因的相对表达水平,dp0158叶片中基因的表达水平设置为1.00,其它转基因株系中osdn-dtp7基因的表达量是dp0158中的10-44倍。
dp1176-f1:5'-gaaagggtatggctctggtaag-3'(seqidno:56)
dp1176-r1:5'-gatgtcagatagtgtgcgagg-3'(seqidno:57)
实施例4.转基因水稻在温室的干旱验证试验
水稻幼苗的耐旱性筛选试验在温室中进行。温室中设置比例为1:1的钠灯和金属卤化物灯作为光源,光照/黑暗时间为16h/8h,光源设置在苗床上部大约1.5m处,晴天时,高于苗床30cm处的光强度为10,000-20,000lx,阴天时为6,000-10,000lx;温室相对湿度为30%-90%,温度为20-35℃。
干旱验证方法:
t2转基因种子和对照种子首先采用800ppm的多菌灵在32℃消毒8h,然后使用蒸馏水冲洗3~5次,然后32℃浸种16h,在35-37℃培养箱中萌发18h。发芽的种子种植在装有有机质、蛭石和沙子(v:v:v=3:3:2)的托盘中。幼苗在正常的温室条件下生长,并浇灌改良的irri溶液,当幼苗生长到3-叶期时,停止浇水,并将水稻幼苗放置在干燥的位置,直至水稻叶片变干和卷曲(大约9~15天,根据季节不同),然后再将托盘放在水槽中使水稻苗复水5-7天,并计算恢复程度。应用以下的打分系统:大于半个绿茎=1,大于2/3的绿叶=1,大于1/3绿叶并低于2/3绿叶=0.5,低于1/3绿叶=0.2,零绿叶或绿茎=0。恢复度是绿色组织得分的总和,数据利用混合模型(mixedmodel)进行统计分析,表现显著优于对照的水稻株系被认为是阳性株系(p<0.05)。存活率指存活株数除以总株数的百分数,是干旱验证的一个指标。
随机区组设计用于从载体水平验证水稻的功能,同一载体的八个转基因株系种植在一个试验单元中,采用混合模型(mixedmodel)考虑载体、株系和环境效应来评估基因的功能。如果转基因水稻的存活率或恢复度显著高于对照(p<0.05),则认为测试的基因具有耐旱性的功能。
温室干旱试验结果:
1)osicdh1转基因水稻(dp0854)的温室干旱验证(drt)试验
第一次试验中,使用8个osicdh1转基因株系进行干旱验证,设置一个重复。当水稻长到三叶期时,将水稻植株在干旱的位置放置20天,然后在水中恢复6天,测定植物的存活率和恢复度。表6表明,在载体水平,osicdh1转基因水稻的恢复度和存活率高于zh11-tc和dp0158对照。转基因株系水平,5个转基因株系的存活率高于zh11-tc和dp0158对照。这些结果表明osicdh1转基因水稻提高了苗期的耐旱性。
表6.温室条件下osicdh1转基因水稻耐旱性分析(第一试验)
第二次试验中,采用同样的8个转基因株系进行验证,设置1个重复,当水稻幼苗长至三叶期时,将水稻植株放置在干燥的位置干旱胁迫15天,在水中恢复6天,计算恢复度。如表7所示,在载体水平,osicdh1转基因水稻的恢复度和存活率高于zh11-tc和dp0158对照;转基因株系水平,6个转基因株系的存活率高于zh11-tc和dp0158对照。这些结果表明osicdh1基因在提高植物幼苗的耐旱性发挥作用。
表7.温室条件下osicdh1转基因水稻耐旱性分析(第二试验)
2)osdn-dtp6转基因水稻(dp0935)的温室drt验证结果
8个osdn-dtp6转基因株系用于验证干旱试验,同时设置四个重复。当水稻长至三叶期,将水稻放置干燥的条件下胁迫20天,然后在水中恢复5~8天。载体水平,osdn-dtp6转基因水稻的恢复度和存活率与zh11-tc和dp0158相似。进一步在转基因株系水平的分析表明4个osdn-dtp6水稻株系的存活率高zh11-tc和dp0158对照,3个转基因株系的恢复度显著高于zh11-tc对照,2个转基因株系的恢复度显著高于dp0158对照(表8)。这些结果表明osdn-dtp6转基因水稻提高了苗期耐旱性。
表8.温室条件下osdn-dtp6转基因水稻耐旱性分析
3)osankl1转基因水稻(dp0960)的温室drt验证结果
8个osankl1转基因水稻用于干旱验证试验,设置三个重复。水稻植株在正常条件下生长至三叶期,然后水稻幼苗放置在干旱条件下,22天不浇水,然后在水中恢复5天,接着将水稻再次干旱胁迫22天,恢复7天后测定恢复度。如表9所示,载体水平和转基因株系水平,osankl1转基因水稻的存活率和恢复度与zh11-tc和dp0158对照相近。
表9.温室条件下osankl1转基因水稻耐旱性分析
4)osmbd2转基因水稻(dp0988)的温室drt验证结果
8个osmbd2转基因水稻株系用于干旱验证试验。当水稻植株生长至三叶期时,干旱胁迫22天,然后在水中恢复6天,96株osmbd2转基因水稻植株中,50株存活;而24株zh11-tc水稻中5株存活,24株dp0158水稻幼苗中2株存活。载体水平上,osmbd2转基因水稻的存活率高于zh11-tc和dp0158对照,恢复度显著高于zh11-tc和dp0158对照(表10)。转基因株系水平的分析表明所有8个转基因株系的存活率和恢复度高于zh11-tc和dp0158对照。这些结果表明与zh11-tc和dp0158对照相比,osmbd2转基因水稻提高了苗期耐旱性,osmbd2基因在提高转基因植物耐旱性中发挥作用。
表10.温室条件下osmbd2转基因水稻耐旱性分析
5)ostp1转基因水稻(dp1082)温室drt验证结果
8个ostp1转基因水稻株系进行了干旱验证,设置三次重复。当水稻生长到三叶期时,将水稻植株放置在干燥的条件下,24天不浇水,然后在水中恢复5天。其中1个重复再次干旱胁迫19天,在水中恢复5天后,测定恢复度。如表11所示,288株ostp1转基因水稻植株中,164株存活;72株zh11-tc水稻植株中,35株存活;72株dp0158水稻植株中,27株存活。载体水平上,ostp1转基因水稻的存活率高于zh11-tc和dp0158对照,恢复度显著高于zh11-tc和dp0158对照。转基因株系水平的分析表明所有8个ostp1转基因株系的存活率和恢复度高于zh11-tc和dp0158对照(表11)。这些结果表明ostp1基因在提高植物耐旱性中起作用。
表11.温室条件下ostp1转基因水稻耐旱性分析(第一次试验)
同样的8个ostp1转基因株系再次进行干旱验证,并设置三个重复。当生长至三叶期时,水稻植株干旱胁迫约18天,在水中恢复5天,288株ostp1转基因水稻中242株存活;而72株zh11-tc植株中,51株存活;72株dp0158植株中51株存活。载体水平上,ostp1转基因水稻的存活率高于zh11-tc和dp0158,平均恢复度显著高于zh11-tc和dp0158对照(表12)。转基因株系水平的分析表明8个转基因株系的存活率和平均恢复度均高于zh11-tc和dp0158对照。这些结果进一步表明ostp1转基因水稻提高了苗期的耐旱性,ostp1基因在提高转基因植物耐旱性中起作用。
表12.温室条件下ostp1转基因水稻的耐旱性分析(第二次试验)
6)osacoat1转基因水稻(dp1121)的温室drt验证结果
8个osacoat1转基因株系用于干旱验证试验,设置四个重复。当生长到三叶期时,首先将水稻植株干旱胁迫26天,恢复5天;然后将水稻植株再次干旱胁迫22天,再次在水中恢复5天后,测定恢复度。如表13所示,在载体和转基因株系水平,osacoat1转基因水稻的存活率和恢复度与zh11-tc和dp0158相近。
表13.温室条件下osacoat1转基因水稻的耐旱性分析
7)osdn-dtp7转基因水稻(dp1176)的温室drt验证结果
8个osdn-dtp7转基因水稻用于干旱验证试验,设置四个重复。水稻植株在正常条件下生长至三叶期后,放置在干燥的条件下,20~22天不浇水。在水中恢复6天后,384株osdn-dtp7转基因水稻植株中262株存活。如表14所示,载体水平上,osdn-dtp7转基因水稻的存活率和恢复度高于dp0158对照;4个osdn-dtp7转基因株系的恢复度显著高于dp0158对照,3个转基因的恢复度高于zh11-tc对照。这些结果表明osdn-dtp7转基因水稻可以提高苗期的耐旱性。
表14.温室条件下osdn-dtp7转基因水稻的耐旱性分析
实施例5.正常灌水条件下成熟转基因水稻植株的籽粒产量
过量表达转基因水稻植株和zh11-tc和dp0158水稻植株种植在水稻田中用于测定正常灌水条件下的籽粒产量。每个基因构建载体选择5个转基因株系进行测定。t2代种子按照实施例4的说明首先进行消毒,萌发的种子种植在田间苗床上。三叶期时,水稻幼苗移栽到试验田中,设置4个重复,每个转基因株系每重复40棵苗,4个重复种植在同一个地块中。zh11-tc和dp0158幼苗临近转基因株系种植,并用做统计分析的对照。
水稻植株正常管理,并施用相应的杀虫剂和化肥,观察并记录实验过程中植株的表型,表型包括抽穗期、卷叶程度、敏旱性和耐旱性。种植季结束时,选择每个株系每行中间的具有代表性的植株收获,测定单株产量,并采用混合线性模型(mixedlinearmodel)进行统计分析。
1)种植在正常灌水条件下的osicdh1转基因水稻植株(dp0854)
对5个osicdh1转基因水稻株系进行测试,种植期间,转基因水稻植株与对照水稻植株没有观察到不同的表型。如表15所示,载体水平上,osicdh1转基因水稻的单株产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有5个测试的转基因株系的单株产量高于两个对照。这些结果表明过量表达osicdh1基因可以提高正常灌水条件下的单株籽粒产量。
表15.osicdh1转基因水稻植株在正常灌水条件下的籽粒产量分析
2)正常灌水条件下的osankl1转基因水稻植株(dp0960)
对5个osankl1转基因水稻株系进行测试,种植期间,转基因水稻植株与对照水稻植株没有观察到不同的表型。如表16所示,载体水平上,osankl1转基因水稻的单株产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有5个测试的转基因株系的单株产量高于两个对照。这些结果表明过量表达osankl1基因可以提高正常灌水条件下的单株籽粒产量。
表16.osankl1转基因水稻植株在正常灌水条件下籽粒产量分析
3)正常灌水条件下osmbd2转基因水稻植株(dp0988)
对5个osmbd2转基因水稻株系进行测试,种植期间,转基因水稻植株与对照水稻植株没有观察到不同的表型。如表17所示,载体水平上,osmbd2转基因水稻的单株产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有5个测试的转基因株系的单株产量显著高于两个对照。这些结果表明过量表达osmbd2基因可以提高正常灌水条件下的单株籽粒产量。
表17.osmbd2转基因水稻植株在正常灌水条件下的籽粒产量分析
4)正常灌水条件下osacoat1转基因水稻植株(dp1121)
对5个osacoat1转基因水稻株系进行测试,种植期间,转基因水稻植株与对照水稻植株没有观察到不同的表型。如表18所示,载体水平上,osacoat1转基因水稻的单株产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有5个测试的转基因株系的单株产量显著高于两个对照。这些结果表明过量表达osacoat1基因可以提高正常灌水条件下的单株籽粒产量。
表18.osacoat1转基因水稻植株在正常灌水条件下的籽粒产量分析
5)正常灌水条件下osdn-dtp7转基因水稻植株(dp1176)
对5个osdn-dtp7转基因水稻株系进行测试,种植期间,转基因水稻植株与对照水稻植株没有观察到不同的表型。如表19所示,载体水平上,osdn-dtp7转基因水稻的单株产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有5个测试的转基因株系的单株产量高于两个对照。这些结果表明过量表达osdn-dtp7基因可以提高正常灌水条件下的单株籽粒产量。
表19.osdn-dtp7转基因水稻植株在正常灌水条件下的籽粒产量分析
实施例6.成熟转基因水稻植株的田间干旱试验
开花期干旱胁迫是农业生产中严重的问题。转基因水稻进一步在田间干旱条件下进行验证。田间干旱试验中,每个基因载体选择12个转基因株系。t2代种子首先参照实施例4描述的方法消毒,萌发的种子种植在田间苗床上,三叶期时,将水稻幼苗移栽到田间试验地,设置四个重复,每个重复每转基因株系10棵苗,并将四个重复种植在同一地块。同一地块中,邻近种植转基因株系种植的zh11-tc和dp0158用作统计分析中的对照。
水稻苗期正常管理,并使用相应的杀虫剂和化肥,幼穗期停止浇水,因此开花期时产生干旱胁迫,干旱时间长短取决于温度和湿度等天气条件。干旱期间,使用tdr30(spectrumtechnologies,inc.)在每块地的10个位点每四天测定土壤相对含水量。
试验过程中,观察并记录植株表型,植株的表型主要包括抽穗期、卷叶程度、敏旱性和抗旱性,尤其关注中午时植株的卷叶程度。收获时,每个株系在每行中间挑选约6株具有代表性的植株,并称量每株水稻籽粒的重量,利用混合线性模型(mixedlinearmodel)对籽粒重量进行统计分析。p<0.1时,认为转基因株系为阳性株系,基因具有提高耐旱性功能。
田间干旱试验结果:
1)osicdh1转基因水稻(dp0854)的田间drt验证结果
第一次田间试验中,使用12个osicdh1转基因水稻株系在海南省进行验证。主茎穗幼穗分化ii停止浇水,干旱胁迫一直持续到种子成熟,此过程中产生较重的干旱胁迫。抽穗期和成熟期期间,土壤含水量从38%下降到7%(图1)。断水后25天,主茎穗完全抽出,分蘖穗处于幼穗分化ⅶ-ⅷ期,且一些水稻植株展现出诸如卷叶的表型。转基因水稻株系dp0854.01、dp0854.02、dp0854.03、dp0854.04和dp0854.07表现出耐旱性的表型。收获时,转基因水稻株系dp0854.03、dp0854.04和dp0854.07表现出较好的结实率。单株籽粒产量如表20所示,载体水平上,osicdh1转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158对照;专转基因株系水平上,6个osicdh1转基因水稻株系的单株籽粒产量显著高于zh11-tc植株,7个转基因水稻株系的单株籽粒产量显著高于dp0158对照。这些结果表明osicdh1转基因水稻在干旱胁迫后,籽粒产量高于对照。
表20.田间干旱条件下osicdh1转基因水稻植株的籽粒产量分析(第一次试验)
第二次试验在宁夏田间进行,同样的12个osicdh1转基因株系进行了测试。当主茎穗处于幼穗分化ⅳ-ⅴ期,分蘖穗处于幼穗分化ⅱ-ⅲ期时停止灌水,抽穗过程中,土壤含水量从20%降至5%(图2)。断水52天后,主茎穗处于乳熟期,水稻植株开始出现叶片卷曲等胁迫表型。如表21所示,载体水平上,osicdh1转基因水稻的单株籽粒产量高于zh11-tc对照,并显著高于dp0158对照;转基因株系水平,8个转基因株系的单株籽粒产量显著高于dp0158对照,5个转基因株系的单株籽粒产量高于zh11-tc对照,这些结果进一步表明osicdh1转基因水稻具有耐旱性,过量表达osicdh1基因提高开花期和抽穗期干旱胁迫后单株籽粒产量。
表21.田间干旱条件下osicdh1转基因水稻植株的籽粒产量分析(第二次试验)
2)osmtn3l转基因水稻(dp0902)的田间drt验证结果
12个osmtn3l转基因水稻株系在海南省田间进行验证,邻近种种的zh11-tc和dp0158水稻植株用作对照。主茎穗幼穗分化ⅱ-ⅳ期,分蘖穗幼穗分化ⅰ期时停止灌水,开始干旱胁迫。抽穗和成熟过程中,田间土壤含水量从43%下降到5%(图3)。断水后19天,主茎穗处于幼穗分化ⅸ期,分蘖穗处于幼穗分化ⅵ-ⅶ期,水稻植株开始出现叶片卷曲等胁迫后的表型。一个转基因株系dp0902.03表现出卷叶程度低和比对照更绿的表型;成熟期,4个转基因株系dp0902.03、dp0902.07、dp0902.11和dp0902.12观察到了结实好等性状。
收获时,每个转基因株系每行选取位于中间的六株收获,并测定单株籽粒产量。载体水平,osmtn3l转基因水稻的单株籽粒产量高于zh11-tc对照,并显著高于dp0158对照;转基因株系水平上,3个osmtn3l转基因株系的单株籽粒产量显著高于zh11-tc对照,8个转基因株系的单株籽粒产量显著高于dp0158对照。4个观察到结实好性状的转基因株系的单株籽粒产量显著高于dp0158对照(表22)。这些结果表明osmtn3l转基因水稻能够耐受干旱胁迫,过量表达osmtn3l基因提高了苗期的耐旱性,并能够增加开花期干旱胁迫后单株籽粒产量。
表22.田间干旱条件下osmtn3l转基因水稻植株的籽粒产量分析(第一次试验)
第二次田间干旱试验在宁夏省进行,使用12个osmtn3l转基因株系验证。20%主茎穗幼穗分化ii期停止灌水,开始干旱胁迫,抽穗过程中土壤含水量从47%降到10%(图4)。断水后的降雨导致干旱胁迫过程中土壤含水量发生变化。断水后18天,主茎穗幼穗分化ⅳ-ⅴ期,分蘖穗幼穗分化ⅲ-ⅳ期,水稻植株开始出现叶片卷曲等胁表型。
如表23所示,载体水平上,osmtn3l转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158对照;转基因株系水平上,5个osmtn3l转基因株系的单株籽粒产量显著高于zh11-tc对照,9个转基因株系的单株籽粒产量显著高于dp0158对照。2个转基因株系dp0902.07和dp0902.11在两次田间干旱试验中均表现出最高的单株籽粒产量。这些结果进一步表明osmtn3l转基因水稻是耐旱的,过量表达osmtn3l基因提高了开花期和抽穗期干旱胁迫后单株籽粒产量。
表23.田间干旱条件下osmtn3l转基因水稻植株的籽粒产量分析(第二次试验)
3)osdn-dtp6转基因水稻(dp0935)的田间drt验证结果
第一次田间试验中,使用12个osdn-dtp6转基因水稻株系在海南田间进行干旱验证。在主茎穗幼穗分化ⅱ期,分蘖穗幼穗分化尚未开始时停止灌水,开始干旱胁迫,抽穗和成熟过程中,田间土壤含水量从35%降到5%(图5)。断水后25天,主茎穗抽出,分蘖穗幼穗分化ⅶ-ⅷ期,水稻植株开始出现叶片卷曲、叶片变黄等胁迫后的表型。6个osdn-dtp6转基因水稻株系dp0935.01、dp0935.03、dp0935.07、dp0935.14、dp0935.15和dp0935.17的卷叶程度较轻。如表24所示,载体水平上,osdn-dtp6转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158对照;转基因株系水平上,8个转基因株系的单株籽粒产量显著高于zh11-tc和dp0158对照。这些结果表明osdn-dtp6转基因水稻是耐旱的,过量表达osdn-dtp6基因提高了开花期干旱胁迫后单株籽粒产量。
表24.田间干旱条件下osdn-dtp6转基因水稻植株籽粒产量分析(第一次试验)
第二次田间干旱试验在海南省进行,使用同样的12个osdn-dtp6转基因株系验证。主茎穗幼穗分化ⅲ-ⅴ期,分蘖穗处于幼穗分化ii期,停止灌水,开始干旱胁迫,抽穗过程中,田间土壤含水量从24%降到6%(图6)。断水后52天,主茎穗处于乳熟期,水稻植株开始出现叶片卷曲等胁迫表型。两个转基因株系dp0935.13和dp0935.17在成熟时,表现出较好的结实表型。载体水平,osdn-dtp6转基因株系籽粒产量高于zh11-tc对照,并显著高于dp0158对照;转基因株系水平,10个osdn-dtp6转基因株系的单株籽粒产量显著高于dp0158对照,4个株系的单株籽粒产量显著高于zh11-tc对照(表25)。这些结果进一步表明osdn-dtp6过表达转基因水稻是耐旱的,过量表达osdn-dtp6提高了耐旱性并提高了单株籽粒产量。
表25.田间干旱条件下osdn-dtp6转基因水稻植株籽粒产量分析(第二次试验)
4)osankl1转基因水稻(dp0960)的田间drt验证结果
12个osankl1转基因水稻株系在海南田间进行验证,邻近种植的zh11-tc和dp0158水稻植株用作对照。在主茎穗幼穗分化ⅰ-ⅱ期,分蘖穗幼穗分化尚未开始时停止灌水,开始干旱胁迫。在抽穗和成熟过程中,田间土壤含水量从35%降到5%(图5)。断水后25天,主茎穗抽出,分蘖穗幼穗分化ⅵ-ⅶ期,水稻植株开始出现叶片卷曲等胁迫后的表型。干旱胁迫过程中,6个osankl1转基因株系dp0960.01、dp0960.02、dp0960.09、dp0960.10、dp0960.11和dp0960.12表现出耐旱表型,如与对照植株相比,卷叶程度低和较少的干叶。5个转基因株系dp0960.01、dp0960.02、dp0960.07、dp0960.09和dp0960.11在成熟期表现出较好的结实表型。
收获时,测定单株籽粒产量。载体水平上,osankl1转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158两个对照;转基因株系水平上,8个osankl1转基因株系的单株籽粒产量显著高于zh11-tc对照,9个转基因株系的单株籽粒产量显著高于dp0158对照(表26)。这些结果表明osankl1转基因水稻耐干旱胁迫,过量表达osankl1基因提高了植物苗期耐旱性,并能够提高开花期干旱胁迫后单株籽粒的产量。
表26.田间干旱条件下osankl1转基因水稻植株的籽粒产量分析(第一次试验)
同样的12个osankl1转基因株系再次在海南田间验证。主茎穗处于幼穗分化ⅲ-ⅴ期,分蘖穗处于幼穗分化ⅱ期,停止灌水,开始干旱胁迫。断水后52天,主茎穗处于乳熟期,水稻植株开始出现干旱胁迫表型。抽穗过程中,田间土壤含水量从24%降到6%(图6)。2个osankl1转基因株系dp0960.03和dp0960.07在成熟期表现出较好的结实表型。
籽粒产量分析表明,载体水平上,osankl1转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158对照;转基因株系水平上,6个osankl1转基因株系的单株籽粒产量显著高于dp0158(表27)。这些结果进一步表明osankl1转基因水稻获得了耐旱性,并能提高单株籽粒产量。
表27.田间干旱条件下osankl1转基因水稻植株的籽粒产量分析(第二次试验)
5)osmbd2转基因水稻(dp0988)的田间drt验证结果
12个osmbd2转基因水稻株系在海南田间进行验证,邻近种植的zh11-tc和dp0158水稻植株用作对照。在主茎穗幼穗分化ⅱ-ⅲ期,分蘖穗幼穗分化尚未开始时停止灌水,开始干旱胁迫。抽穗和成熟过程中,田间土壤含水量从27%降到4%(图7)。断水后24天,主茎穗抽出,分蘖穗幼穗分化ⅵ-ⅶ期,水稻植株开始出现叶片卷曲等胁迫后的表型。干旱胁迫过程中,osmbd2转基因水稻的卷叶程度和枯叶程度低于对照;成熟期,除转基因株系dp0988.03、dp0988.04和dp0988.05外,均表现出结实好的表型。
籽粒产量分析表明,载体水平上,osmbd2转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158两个对照;转基因株系水平上,10个osmbd2转基因株系的单株籽粒产量显著高于zh11-tc对照,12个转基因株系的单株籽粒产量显著高于dp0158对照(表28)。这些结果表明osmbd2转基因水稻是耐干旱的,过量表达osmbd2基因提高了苗期耐旱性,并提高了开花期干旱胁迫后单株籽粒产量。
表28.田间干旱条件下osmbd2转基因水稻植株籽粒产量分析(第一次试验)
同样的12个osmbd2转基因水稻株系再次在海南田间验证。主茎穗处于幼穗分化ⅳ-ⅴ期,分蘖穗处于幼穗分化ⅱ-ⅲ期时,停止灌水,开始干旱胁迫。断水后52天,主茎穗处于乳熟期,水稻植株开始出现干旱胁迫表型。抽穗过程中,田间土壤含水量从16%降到6%(图8)。
籽粒产量分析表明,载体水平上,osmbd2转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158对照;转基因株系水平上,所有osmbd2转基因株系的单株籽粒产量高于zh11-tc和dp0158对照(表29)。这些结果进一步表明osmbd2转基因水稻获得了耐旱性,并表现出较高的单株籽粒产量。
表29.田间干旱条件下osmbd2转基因水稻植株籽粒产量分析(第二次试验)
6)ostp1转基因水稻(dp1082)的田间drt验证结果
12个ostp1转基因水稻株系在海南田间进行验证。在主茎穗幼穗分化ii期时停止灌水,并产生较重的干旱胁迫。抽穗和成熟过程中,田间土壤含水量从35%降到5%(图9)。与邻近种植的zh11-tc和dp0158相比,2个ostp1转基因株系dp1082.03和dp1082.06叶片较绿和卷叶程度较低,4个转基因株系dp1082.04、dp1082.10、dp1082.11和dp1082.12在成熟期表现较好的结实率表型。
载体水平上,ostp1转基因水稻的单株籽粒产量高于zh11-tc,并显著高于dp0158对照;转基因株系水平上,5个ostp1转基因株系的单株籽粒产量显著高于zh11-tc对照,8个转基因株系的单株籽粒产量显著高于dp0158对照(表30)。这些结果表明,ostp1转基因水稻提高了苗期耐旱性,并提高了干旱胁迫后单株籽粒产量。
表30.田间干旱条件下ostp1转基因水稻植株籽粒产量分析(第一次试验)
同样的12个ostp1转基因株系再次在宁夏田间进行验证。10%主茎穗处于幼穗分化ii期停止灌水,开始干旱胁迫,抽穗过程中,田间土壤含水量从45%降到5%(图10)。断水后26天,降雨导致了田间土壤含水量的变化。3个转基因株系dp1082.03、dp1082.11和dp1082.12表现出较好的结实表型。籽粒产量分析表明,ostp1转基因水稻的单株籽粒产量低于zh11-tc对照,但是显著高于dp0158对照(表31)。这些结果表明ostp1过表达转基因水稻获得了苗期耐旱性,过量表达ostp1基因提高耐旱性。
表31.田间干旱条件下ostp1转基因水稻植株籽粒产量分析(第二次试验)
7)osacoat1转基因水稻(dp1121)的田间drt验证结果
第一次试验中,12个osacoat1转基因水稻株系在海南田间进行验证。在主茎穗幼穗分化ii期时停止灌水,开始干旱胁迫,直至种子成熟,产生较严重的干旱胁迫力度。抽穗和成熟过程中,田间土壤含水量从35%降到5%(图9)。断水后26天,主茎穗抽出,分蘖穗幼穗分化ⅶ-ⅷ期,一些水稻植株开始出现叶片卷曲等胁迫后的表型。转基因株系dp1121.06和dp1121.08表现出较好的耐旱表型,诸如较绿的叶片和较轻的卷叶程度。收获时,转基因水稻株系dp1121.01、dp1121.02、dp1121.04、dp1121.06、dp1121.08和dp1121.09表现出较好的结实表型。
表32为单株籽粒产量结果,载体水平上,osacoat1转基因水稻的单株籽粒产量显著高于zh11-tc和dp0158两个对照;转基因株系水平上,7个osacoat1转基因株系的单株籽粒产量显著高于zh11-tc和dp0158对照。这些结果表明osacoat1水稻在干旱胁迫后获得较高的单株籽粒产量。
表32.田间干旱条件下osacoat1转基因水稻植株籽粒产量分析(第一试验)
第二次干旱验证试验在宁夏省进行,同样的12个osacoat1转基因株系进行了验证。10%主茎穗幼穗分化ii期停止灌水,开始干旱胁迫,抽穗过程中,田间土壤含水量从45%降到10%(图10)。降雨导致了干旱胁迫过程中田间土壤含水量的变化。断水后16天,主茎穗幼穗分化ⅳ-ⅴ期,分蘖穗幼穗分化ⅲ-ⅳ期,一些水稻植株开始出现叶片卷曲等胁迫表型。1个转基因株系dp1121.06在干旱胁迫过程中表现出耐旱性表型,2个转基因株系dp1121.01和dp1121.04在成熟期表现出较好的结实表型。
如表33所示,载体水平上,osacoat1转基因水稻的单株籽粒产量显著高于dp0158对照,但低于zh11-tc对照;转基因株系水平上,5个转基因株系的单株籽粒产量显著高于dp0158对照,1个株系的单株籽粒产量显著高于zh11-tc对照。这些结果表明osacoat1转基因水稻是耐干旱的,过量表达osacoat1基因提高了开花期和抽穗期干旱胁迫后单株籽粒产量。
表33.田间干旱条件下osacoat1转基因水稻植株籽粒产量分析(第二次试验)
8)osdn-dtp7转基因水稻(dp1176)田间drt验证结果
第一试验中,12个osdn-dtp7转基因水稻株系在海南田间进行验证。在主茎穗幼穗分化ii期,分蘖穗幼穗分化1期时停止灌水,开始干旱胁迫直至种子成熟,产生较重的干旱胁迫。抽穗和成熟过程中,田间土壤含水量从40%降到7%(图11)。断水后19天,主茎穗幼穗分化ⅷ期,分蘖穗幼穗分化ⅴ-ⅵ期,一些水稻植株开始出现叶片卷曲等胁迫后的表型。3个转基因水稻株系dp1176.02、dp1176.04和dp1176.10表现出耐旱表型,诸如叶片较绿和卷叶程度较轻等。收获时,4个转基因水稻株系dp1176.02、dp1176.05、dp1176.06和dp1176.08表现出较好的结实表型。
表34为单株籽粒产量结果,在载体水平上,osdn-dtp7转基因水稻的单株籽粒产量高于zh11-tc对照并显著高于dp0158对照;在转基因株系水平上,5个osdn-dtp7转基因株系的单株籽粒产量显著高于zh11-tc对照,6个转基因株系的单株籽粒产量显著高于dp0158对照。这些结果表明,osdn-dtp7转基因水稻在干旱胁迫后获得比对照高的单株籽粒产量。
表34.田间干旱条件下osdn-dtp7转基因水稻植株籽粒产量分析(第一次试验)
第二试验在海南田间进行验证,对同样的12个osdn-dtp7转基因水稻株系进行测试。在主茎穗幼穗分化ⅴ-ⅵ期,分蘖穗幼穗分化ii-ⅲ期时停止灌水。断水后52天,主茎穗处于乳熟期,一些水稻植株开始出现干旱胁迫表型。抽穗和成熟过程中,田间土壤含水量从22%降到7%(图12)。收获时,2个转基因水稻株系dp1176.02和dp1176.08表现出较好的结实表型。
表35为单株籽粒产量结果,在载体水平上,osdn-dtp7转基因水稻的单株籽粒产量显著高于zh11-tc对照并高于dp0158对照;在转基因株系水平上,5个osdn-dtp7转基因株系的单株籽粒产量显著高于zh11-tc对照,4个转基因株系的单株籽粒产量显著高于dp0158对照。这些结果表明,osdn-dtp7转基因水稻在干旱胁迫后获得比对照高的单株籽粒产量。
表35.田间干旱条件下osdn-dtp7转基因水稻植株籽粒产量分析(第二次试验)
实施例7.转基因水稻植株的实验室百草枯试验
百草枯(1,1-二甲基-4,4-联吡啶二氯化物)是一类叶片喷施的非选择性的吡啶除草剂,是世界范围内广泛应用一种除草剂,能够控制大量作物如玉米、水稻、大豆等中生长的杂草。在植物细胞中,百草枯主要靶向叶绿体,百草枯通过接受光系统i的电子,然后与氧气发生化学反应生成过氧化物和过氧化氢,而过氧化物和过氧化氢可以导致产生光氧化胁迫。干旱胁迫通常导致植物中产生活性氧(ros),有时,植物的耐旱性与增强的抗活性氧能力相关。百草枯是强有力的氧化胁迫诱导剂,能够大大的增加活性氧(ros)的产生,同时抑制抗氧化系统活性所需的的还原物和化合物的再生。非生物胁迫增加了ros的产生,而植物响应耐性到死亡的范围取决于胁迫力度和与其相关的ros水平。相对较低水平的百草枯能够模拟胁迫相关的ros产生,并用作植物胁迫生物学中胁迫耐性的标记(hasaneenm.n.a.(2012)herbicide-properties,synthesisandcontrolofweedsbook)。因此,进一步采用百草枯验证耐旱性的转基因水稻。
百草枯试验方法:
每个载体的水稻选择10个转基因株系用于百草枯试验,组培中花11(zh11-tc)和转空载体对照dp0158用作对照。t2代种子参照实施例4描述的方法消毒和萌发。百草枯试验在温度28-30℃,湿度30%的生长室中进行。萌发的种子放置在底部有孔的离心管中,采用水稻水培方法,培养5天,至一叶一心期;然后选择高度大约3.5~4cm的一致的幼苗用于百草枯试验。本实验采用随机区组设计,在同一个筛选水槽内设置5个区组;区组内包含所有测试的10个转基因株系、zh11-tc和dp0158;区组的行列为16*12,每一行为一份测试材料,故每个转基因株系在区组内各12株,对照zh11-tc和dp0158在区组内各3行;区组内的所有转基因株系和对照均随机排布。幼苗用终浓度为0.8μm的百草枯溶液进行处理7天,光周期为10h黑暗/14h光照,每两天更换一次溶液,在处理和更换溶液后,保证幼苗首先进入光周期的黑暗时期。处理7天后,计算绿色的幼苗。绿色没有损伤的幼苗为百草枯耐性幼苗;叶片、茎部变白褪色的幼苗为非百草枯耐性的幼苗。
耐性率是百草枯试验的一个指标,指保持绿色并显示百草枯耐性表型的幼苗数除以总幼苗数的百分数。
试验数据在载体水平(所有的转基因幼苗与对照幼苗相比)和转基因株系水平(不同的转基因株系与对照相比)进行分析,采用的统计模型为“y~seg+line(seg)+rep+error”,随机效应为“rep”,统计方法是“
百草枯试验结果:
1)osicdh1转基因水稻(dp0854)的百草枯验证结果
第一次试验中,百草枯溶液处理7天后,600株osicdh1转基因幼苗中,398株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,72株具有百草枯耐性表型;180株dp0158幼苗中,105株显示百草枯耐性表型。载体水平上,所测试的osicdh1转基因幼苗的耐性率显著高于zh11-tc和dp0158对照。
进一步在转基因株系水平的分析表明9个osicdh1转基因株系的百草枯耐性率显著高于zh11-tc对照和3个株系的百草枯耐性率显著高于dp0158对照(表36)。这些结果表明,与zh11-tc和dp0158两个对照相比,osicdh1转基因水稻在载体水平和转基因株系水平均提高了幼苗的百草枯耐性,osicdh1在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表36.osicdh1转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,测试同样的10个osicdh1转基因株系,百草枯溶液处理7天后,600株osicdh1转基因幼苗中,424株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,104株具有百草枯耐性表型;180株dp0158幼苗中,114株显示百草枯耐性表型。载体水平上,所测试的osicdh1转基因幼苗的耐性率显著高于zh11-tc对照。
进一步在转基因株系水平的分析表明4个osicdh1转基因株系的百草枯耐性率显著高于zh11-tc对照和1个株系的百草枯耐性率显著高于dp0158对照(表37)。这些结果进一步表明,osicdh1在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表37.osicdh1转基因水稻植株的百草枯耐性试验(第二次试验)
2)osmtn3l转基因水稻(dp0902)的百草枯验证结果
第一次试验中,百草枯溶液处理7天后,600株osmtn3l转基因幼苗中,219株保持绿色并显示出百草枯耐性表型;而168株zh11-tc幼苗中,48株具有百草枯耐性表型;180株dp0158幼苗中,58株显示百草枯耐性表型。载体水平上,所测试的osmtn3l转基因幼苗的耐性率高于zh11-tc和dp0158对照。
进一步在转基因株系水平的分析表明4个osmtn3l转基因株系的百草枯耐性率显著高于zh11-tc和dp0158两个对照(表38)。这些结果表明,与zh11-tc和dp0158两个对照相比,osmtn3l转基因水稻在载体水平和转基因株系水平均提高了幼苗的百草枯耐性,osmtn3l在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表38.osmtn3l转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,测试同样的10个osmtn3l转基因株系,百草枯溶液处理7天后,600株osmtn3l转基因幼苗中,465株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,122株具有百草枯耐性表型;180株dp0158幼苗中,117株显示百草枯耐性表型。载体水平上,所测试的osmtn3l转基因幼苗的耐性率显著高于zh11-tc和dp0158对照。
进一步在转基因株系水平的分析表明所有10个osmtn3l转基因株系的百草枯耐性率高于zh11-tc和dp0158两个对照(表39)。这些结果表明,与zh11-tc和dp0158两个对照相比,osmtn3l转基因水稻在载体水平和转基因株系水平均提高了幼苗的百草枯耐性,osmtn3l在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表39.osmtn3l转基因水稻植株的百草枯耐性试验(第二次试验)
3)osdn-dtp6转基因水稻(dp0935)的百草枯验证结果
第一次试验中,0.8μm百草枯溶液处理7天后,600株osdn-dtp6转基因幼苗中,321株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,68株显示出百草枯耐性表型;180株dp0158幼苗中,80株显示出百草枯耐性。载体水平上,osdn-dtp6转基因幼苗的耐性率显著高于zh11-tc和dp0158两个对照。表40为转基因株系水平的分析结果,7个osdn-dtp6转基因株系的耐性率显著高于zh11-tc或dp0158对照,且其中4个转基因株系的耐性率高于60%。这些结果表明过量表达osdn-dtp6基因提高了转基因植物的百草枯耐性或抗氧化活性。
表40.osdn-dtp6转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,测试同样的10个osdn-dtp6转基因株系,0.8μm百草枯溶液处理7天后,600株osdn-dtp6转基因幼苗中,449株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,115株具有百草枯耐性表型;180株dp0158幼苗中,104株显示百草枯耐性表型。载体水平上,所测试的osdn-dtp6转基因幼苗的耐性率显著高于zh11-tc和dp0158对照。
表41为进一步在转基因株系水平的分析,4个osdn-dtp6转基因株系的百草枯耐性率显著高于zh11-tc和dp0158对照,其中9个转基因株系的百草枯耐性率大于70%。这些结果表明,过量表达osdn-dtp6基因提高百草枯耐性,osdn-dtp6在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表41.osdn-dtp6转基因水稻植株的百草枯耐性试验(第二次试验)
4)osankl1转基因水稻(dp0960)百草枯验证结果
第一次试验中,百草枯溶液处理后,600株osankl1转基因幼苗中,348株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,75株显示出百草枯耐性表型;180株dp0158幼苗中,94株显示出百草枯耐性表型。载体水平上,osankl1转基因水稻的耐性率显著高于zh11-tc对照,并高于dp0158对照。表42为转基因株系水平上的分析结果,7个转基因株系的百草枯耐性率高于zh11-tc和dp0158幼苗,这进一步表明osankl1转基因水稻在载体水平和转基因株系水平提高了苗期百草枯的耐性,过量表达osankl1基因提高了转基因植物的百草枯耐性。
表42.osankl1转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,百草枯溶液处理后,600株osankl1转基因幼苗中,418株保持绿色并显示出百草枯耐性表型;而180株zh11-tc幼苗中,114株显示出百草枯耐性表型;180株dp0158幼苗中,114株显示出百草枯耐性表型。载体水平上,osankl1转基因水稻的耐性率高于zh11-tc和dp0158对照。表43为转基因株系水平上的分析结果,7个转基因株系的百草枯耐性率高于zh11-tc和dp0158幼苗,这进一步表明osankl1转基因水稻在载体水平和转基因株系水平提高了苗期百草枯的耐性,过量表达osankl1基因提高了转基因植物的百草枯耐性。
表43.osankl1转基因水稻植株的百草枯耐性试验(第二次试验)
5)osmbd2转基因水稻(dp0988)的百草枯验证结果
第一次试验中,百草枯溶液处理后,600株osmbd2转基因水稻幼苗中,295株保持绿色并显示出百草枯耐性表型;180株zh11-tc幼苗中,99株显示出百草枯耐性表型;180株dp0158幼苗中,90株显示出百草枯耐性表型。载体水平上,osmbd2转基因水稻的耐性率低于zh11-tc和dp0158对照。表44为转基因株系水平上的分析,1个osmbd2转基因株系的耐性率显著高于zh11-tc对照,2个转基因株系的耐性率显著高于dp0158对照,osmbd2转基因水稻没有表现出较好的百草枯耐性。
表44.osmbd2转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,百草枯溶液处理后,600株osmbd2转基因幼苗中,263株保持绿色,并显示出百草枯耐性表型;180株zh11-tc幼苗中,80株显示出百草枯耐性表型;180株dp0158幼苗中,88株显示出百草枯耐性表型。载体水平上osmbd2转基因幼苗的耐性率与zh11-tc和dp0158对照相近。转基因株系水平的分析显示8个转基因株系的耐性率低于zh11-tc和dp0158对照,只有2个转基因株系的耐性率显著高于zh11-tc对照(表45)。这些结果表明osmbd2转基因水稻没有表现出较好的百草枯耐性或抗氧化活性。
表45.osmbd2转基因水稻植株的百草枯耐性试验(第二次试验)
6)ostp1转基因水稻(dp1082)的百草枯验证结果
第一次试验中,百草枯溶液处理7天后,600株ostp1转基因幼苗中,371株保持绿色并显示出百草枯耐性表型;180株zh11-tc幼苗中,103株显示出百草枯耐性表型;180株dp0158幼苗中,108株显示出百草枯耐性表型。载体水平上,ostp1转基因幼苗的耐性率高于zh11-tc和dp0158两个对照。表46为转基因株系水平的分析结果,5个ostp1转基因株系的百草枯耐性率高于zh11-tc和dp0158对照。这些结果表明ostp1过表达转基因水稻在载体水平和转基因株系水平提高了幼苗的百草枯耐性。
表46.ostp1转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,测试同样的10个ostp1转基因株系,百草枯溶液处理7天后,588株ostp1转基因幼苗中,465株保持绿色并显示出百草枯耐性表型;而192株zh11-tc幼苗中,135株具有百草枯耐性表型;180株dp0158幼苗中,119株显示百草枯耐性表型。载体水平上,所测试的ostp1转基因幼苗的耐性率显著高于zh11-tc和dp0158对照。
表47为进一步在转基因株系水平的分析,4个ostp1转基因株系的百草枯耐性率显著高于zh11-tc,6个株系的百草枯耐性率显著大于dp0158对照。这些结果表明,过量表达ostp1基因的转基因植物提高了构建体水平和转基因株系水平的百草枯耐性,ostp1在提高转基因植物百草枯耐性或抗氧化能力中起作用。
表47.ostp1转基因水稻植株的百草枯耐性试验(第二次试验)
7)osacoat1转基因水稻(dp1121)的百草枯验证结果
第一次试验中,百草枯溶液处理7天后,600株osacoat1过表达转基因幼苗中,441株保持绿色并表现出百草枯耐性表型;而180株zh11-tc幼苗中,113株显示出百草枯耐性表型;180株dp0158幼苗中,118株显示出百草枯耐性表型。载体水平上,osacoat1转基因水稻的百草枯耐性率显著高于zh11-tc和dp0158对照。表48的转基因株系水平的分析结果表明,7个osacoat1转基因水稻株系的百草枯耐性率高于zh11-tc和dp0158对照,其中3个转基因株系的百草枯耐性率显著高于zh11-tc和dp0158对照。这些结果表明osacoat1过表达转基因水稻提高了载体水平和转基因株系水平的百草枯耐性。
表48.osacoat1转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,百草枯溶液处理7天后,600株osacoat1过表达转基因幼苗中,453株保持绿色并表现出百草枯耐性表型;而180株zh11-tc幼苗中,115株显示出百草枯耐性表型;180株dp0158幼苗中,112株显示出百草枯耐性表型。载体水平上,osacoat1转基因水稻的百草枯耐性率显著高于zh11-tc和dp0158对照。表49的转基因株系水平的分析结果表明,7个osacoat1转基因水稻株系的百草枯耐性率高于zh11-tc和dp0158对照。这些结果表明osacoat1过表达转基因水稻提高了载体水平和转基因株系水平的百草枯耐性。osacoat1基因在提高转基因植物的百草枯耐性或抗氧化能力中起作用。
表49.osacoat1转基因水稻植株的百草枯耐性试验(第二次试验)
8)osdn-dtp7转基因水稻(dp1176)的百草枯验证结果
第一次试验中,百草枯溶液处理7天后,600株osdn-dtp7过表达转基因幼苗中,447株保持绿色并显示出百草枯耐性表型;180株zh11-tc幼苗中,113株显示出百草枯耐性表型;180株dp0158幼苗中,112显示出百草枯耐性表型。载体水平上,osdn-dtp7转基因幼苗的百草枯耐性率显著高于zh11-tc和dp0158两个对照。表50的转基因株系水平的分析结果表明8个osdn-dtp7转基因株系的百草枯耐性率高于zh11-tc对照,9个转基因株系的耐性率高于dp0158对照。这些结果表明osdn-dtp7过量表达转基因水稻提高了载体水平和转基因株系水平的苗期百草枯耐性。
表50.osdn-dtp7转基因水稻植株的百草枯耐性试验(第一次试验)
第二次试验中,百草枯溶液处理7天后,600株osdn-dtp7过表达转基因幼苗中,427株保持绿色并显示出百草枯耐性表型;180株zh11-tc幼苗中,100株显示出百草枯耐性表型;180株dp0158幼苗中,116显示出百草枯耐性表型。载体水平上,osdn-dtp7转基因幼苗的百草枯耐性率显著高于zh11-tc和dp0158对照。表51的转基因株系水平的分析结果表明9个osdn-dtp7转基因株系的百草枯耐性率高于zh11-tc和dp0158对照。这些结果表明osdn-dtp7过量表达转基因水稻提高了载体水平和转基因株系水平的苗期百草枯耐性,osdn-dtp7基因在提高转基因植物的百草枯耐性或抗氧化活性中起作用。
表51.osdn-dtp7转基因水稻植株的百草枯耐性试验(第二次试验)
实施例8.osmbd2转基因水稻的开花性状验证
t2代osmbd2转基因植株种植在宁夏田间(宁夏,38°36'n,106°23'e,海拔高1106.3m),并记录植株生长过程中的表型。
方法:
t2代转基因种子于32℃800ppm多菌灵中消毒8h后,蒸馏水冲洗3~5次,然后32℃浸种16h,再在35-37℃培养箱中萌发18h。萌发的种子种植在田间苗床上,三叶期时,将幼苗移栽到田间稻田中。每个转基因株系的10棵植株种植一行,zh11-wt(中花11号野生型)、zh11-tc和dp0158临近转基因水稻,种植在同一个小区,并用作对照。
水稻植株正常管理,并使用相应的杀虫剂和化肥,实验过程中观察并记录植株表型。
本试验记录抽穗期和成熟期,抽穗期是指一行中每棵植株50%的幼穗抽出剑叶叶鞘的日期,成熟期指每穗谷粒颖壳90%以上变黄或谷粒小穗轴及副护颖变黄的日期,此时为最佳收获时期。如果转基因水稻的抽穗期早于对照水稻植物(中花11号、zh11-tc或dp0158植株),那么转基因水稻被认为具有早抽穗的特征,转入的基因在控制植物开花时间上发挥作用。
本试验还测定了植株高度、有效穗数和单株产量。植株高度指从地表到最高穗或叶顶部的长度。收获时,将每个转基因株系的全部或位于中间位置的六株收获,先将稻穗剪下并存放在一个袋子中,然后将茎部贴地剪下放在另外的袋子中,测定每个单株的有效穗数和单株产量。植株高度、有效穗数和单株产量采用asreml混合线性模型分析。
结果:
12个osmbd2转基因水稻株系种植在宁夏田间进行测试,开始,水稻植株正常管理。当主茎穗处于幼穗分化ⅰ期,停止灌水。水稻植株遭受干旱胁迫,观察和记录了osmbd2转基因水稻的抽穗期。8月16日,8个转基因株系的50%的幼穗从剑叶的叶鞘中抽出,而对照zh11-tc和dp0158水稻的50%的幼穗抽出的时间为8月26日。一些osmbd2转基因株系的抽穗期比对照水稻早10天,这些早抽穗的水稻也能够比对照水稻提前10天成熟。除转基因株系dp0988.01、dp0988.07、dp0988.08和dp0988.12外,osmbd2转基因水稻植株表现出较好的结实表型;籽粒产量的分析表明,转基因株系水平,所有osmbd2转基因株系的单株籽粒产量高于dp0158对照,3个转基因株系的单株籽粒产量高于zh11-tc植株(表52),这些结果表明osmbd2基因调控转基因水稻的抽穗期和/或开花时间,osmbd2转基因水稻在干旱胁迫后显示出增加的单株籽粒产量。
表52.osmbd2转基因水稻植株在宁夏开花性状和籽粒产量分析(第一次试验)
选择5个osmbd2转基因水稻株系,再次在宁夏进行干旱测试,zh11-tc和dp0158幼苗用作对照。种子于4月14日播种,5月14日移栽到田间,开始,水稻植株正常管理。当主茎穗处于幼穗分化ⅰ期,停止灌水。水稻植株遭受干旱胁迫。4个osmbd2转基因水稻株系比zh11-tc和dp0158水稻植株早抽穗7-8天,并表现出较好的结实表型,早抽穗的水稻也比对照水稻提前成熟。籽粒产量的分析表明,4个转基因株系水平的单株籽粒产量高于zh11-tc和dp0158水稻植株(表53)。
表53.osmbd2转基因水稻植株在宁夏开花性状和籽粒产量分析(第二次试验)
5个osmbd2转基因水稻株系,种植宁夏田间水田中,zh11-tc和dp0158幼苗用作对照。种子于4月14日播种,5月14日移栽到田间。所有5个osmbd2转基因水稻株系比zh11-tc和dp0158水稻植株早抽穗6-11天,并表现出较好的结实表型,早抽穗的水稻也比对照水稻提前成熟。籽粒产量的分析表明,4个转基因株系水平的单株籽粒产量低于zh11-tc和dp0158水稻植株(表54)。
osmbd2基因在转基因水稻植株中的表达水平如图13所示,并在实施例3中描述dp0988.07、dp0988.08和dp0988.12的表达水平比其他水稻株系低,dp0988.07、dp0988.08和dp0988.12的抽穗天数也比其他水稻株系长,这些结果表明osmbd2基因表达水平影响转基因水稻的抽穗时间或者开花时间。
表54.osmbd2转基因水稻植株在宁夏开花性状和籽粒产量分析(第三次试验)
实施例9.成熟转基因水稻植株的田间低氮耐性试验
田间耐低氮试验在北京进行。设置一个氮素水平:n-0(不含氮的化肥)。种子萌发和幼苗培养如实施例4所述。萌发种子种植于田间苗床中,三叶期将幼苗移栽入试验田中,重复4次,每个转基因株系每个重复种植10株,4个重复种于同一个田块中。同一田块中临近转基因株系的zh11-tc和dp0158植株作为对照,并用于统计分析。
水稻植株正常管理,并使用相应的杀虫剂,n-0处理施用磷肥和钾肥。
生长季节结束时,从每个转基因株系中间收获六株代表植株,测量单株籽粒产量。利用asreml软件的混合线性模型统计分析粒重数据。基于分析选择阳性转基因株系(p<0.1)。
1)osmtn3l转基因水稻(dp0902)的田间nue验证结果
osmtn3l转基因水稻植株和相应的对照种植在田间水田中,整个生育期不施用氮肥,营养生长期,osmtn3转基因水稻植株比其相应的对照粗壮,测定osmtn3l转基因水稻植株的单株籽粒产量。如表55所示,低氮条件下,载体水平上osmtn3l转基因水稻的单株籽粒产量显著高于zh11-tc对照,并高于osmtn3l对照;转基因株系水平上,10个株系的单株籽粒产量高于zh11-tc和dp0158对照。这些结果表明osmtn3l转基因水稻获得了低氮耐性,过量表达osmtn3l基因提高了低氮条件下转基因植株的籽粒产量,osmtn3l基因在提高低氮耐性和/或nue中起作用。
表55.osmtn3l转基因水稻在田间低氮条件下的籽粒产量分析
2)osankl1转基因水稻(dp0960)的田间nue验证结果
osankl1转基因水稻植株和相应的对照种植在田间水田中,整个生育期不施用氮肥,营养生长期,osankl1转基因水稻植株比其相应的对照粗壮;种植季结束时,测定osankl1转基因水稻植株的单株籽粒产量。如表56所示,低氮条件下,载体水平上osankl1转基因水稻的单株籽粒产量高于zh11-tc对照和osankl1对照;转基因株系水平上,6个株系的单株籽粒产量高于zh11-tc和dp0158对照。这些结果表明osankl1转基因水稻获得了低氮耐性,过量表达osankl1基因提高了低氮条件下转基因植株的籽粒产量,osankl1基因在提高低氮耐性和/或nue中起作用。
表56.osankl1转基因水稻(dp0960)在田间低氮条件下的籽粒产量分析
实施例10.转基因水稻植株的实验室氯酸盐试验
硝酸盐是高等植物无机氮利用的主要来源。氯酸盐是硝酸盐类似物,植物通过氮素的吸收、转运系统吸收和转运氯酸盐,并通过硝酸还原酶(nr)还原为有毒化合物(亚硝酸盐)。为了进一步证明转基因水稻植物的氮素利用效率,用氯酸盐溶液处理水稻幼苗,对氯酸盐敏感的幼苗被认为具有增加的氮素利用效率或低氮耐受性。
实验室氯酸盐试验方法:
每个构建体选择大约10个株系进行氯酸盐试验。zh11-tc植株为对照。
如实施例4所描述对t2代转基因种子和对照种子的进行消毒和并使其萌发,本试验在温度为28-30℃、相对湿度约30%的培养室中进行。种子萌发后置于底部有孔的管中,30℃水培6天至一叶一心期。选取高度为5.5cm的一致性幼苗用于氯酸盐筛选。本试验选用随机区组设计。每个容器分为5个区域,每个区域中随机将每个转基因株系放1行(每列12株),zh11-tc幼苗放3行(3x12棵植株)。幼苗置于0.4mm的氯酸盐溶液中培养3-5天,光照/黑暗时间为10h/14h。处理幼苗首先进入黑暗时期进行氯酸盐吸收,并在第三天更换氯酸盐溶液。处理3-5天后,幼苗转入1/10hoagland营养液(表6)中恢复培养4天。整株叶片枯萎和叶片全部失绿的幼苗被视为敏感幼苗。仅叶片或茎部坏死的幼苗,或叶片褪色的幼苗视为非敏感幼苗。
本筛选以敏感率为参数,所述敏感率为敏感植株数除以总植株数的百分比。
构建体水平(所有转基因植物与对照相比)和转基因株系水平(不同转基因株系分别与对照相比)的数据分析均采用“y~seg+line(seg)+rep+error”统计模型,随机效应为“rep”,统计方法为“sasprocglimmix”。
氯酸盐试验结果:
osankl1转基因水稻(dp0960)的氯酸盐验证试验
第一次试验中,对osankl1转基因水稻用0.4mm的氯酸盐溶液进行处理5天后置于1/10的hoagland营养液中恢复培养4天,600株osankl1转基因幼苗中369株死亡,而180株zh11-tc幼苗死亡70株。osankl1转基因幼苗的敏感率显著高于zh11-tc对照。表明osankl1转基因幼苗显示增强的氯酸盐敏感性。
进一步在转基因株系水平的分析表明所有10个转基因株系的敏感率高于zh11-tc对照,其中7个株系的氯酸盐敏感率显著高于zh11-tc对照(表57)。这些结果表明osankl1转基因水稻植株的敏感率在载体水平和转基因株系水平高于zh11-tc幼苗。
表57.osankl1转基因水稻幼苗的氯酸盐敏感性试验(第一次试验)
第二次试验中,0.4mm的氯酸盐溶液进行处理5天后置于1/10的hoagland营养液中恢复培养4天,600株osankl1转基因幼苗中216株死亡,而180株zh11-tc幼苗死亡33株。osankl1转基因幼苗的敏感率显著高于zh11-tc对照,表明osankl1转基因幼苗显示增强的氯酸盐敏感性。
转基因株系水平的分析表明8个转基因株系的敏感率显著高于zh11-tc对照(表58)。这些结果表明osankl1转基因水稻植株在苗期获得了增加的氯酸盐敏感率,过量表达osankl1基因增加转基因植物的氯酸盐敏感性。
表58.osankl1转基因水稻幼苗的氯酸盐敏感试验(第二次试验)
实施例11.水稻耐旱基因转化玉米并评估玉米的抗旱性
将水稻耐旱性基因或者玉米、拟南芥或其它物种中的相应的同源基因转化玉米植株,以组成型的启动子如玉米泛素启动子(christensen等(1989)plantmol.biol.12:619-632和christensen等(1992)plantmol.biol.18:675-689)或诸如胁迫响应启动子或组织偏爱启动子的其它启动子驱动玉米转化载体中基因的表达,采用微粒轰击法(国际专利公布wo2009/006276)将构建的重组dna载体转化玉米,也可以以采用农杆菌介导法(zhao等,meth.mol.biol.318:315-323(2006)和zhao等,mol.breed.8:323-333(2001)和美国专利号5,981,840,1999年11月9日公布)转化玉米。农杆菌介导转化的过程包括细菌的接种、共培养、静止、筛选和植株的再生。
再生植株的子代如t1代植株可以种在土壤中进行干旱胁迫,采用图像分析,测量胁迫前和干旱胁迫过程中多个时期点植株面积、体积、生长速率和叶片颜色。与对照相比,明显延迟胁迫期间植株的萎蔫,叶面积的减小,黄色素积累的减少,和/或促进生长速率的增加被认为基因提高玉米的耐旱性能。
实施例12.水稻耐旱基因转化拟南芥的实验室干旱验证
为了验证水稻耐旱性基因是否提高双子叶植物的耐旱性或其它性状,采用农杆菌介导的花浸法将水稻耐旱性基因过表达载体转化拟南芥(cloumbia),并鉴定转基因拟南芥(clough,s.t.和bent,a.f.(1998)theplantjournal16,735–743;zhang,x.等(2006)natureprotocols1:641-646)。
一个称为pbc-yellow的16.8-kbt-dna双元载体用于本试验。该载体包含一个rd29a启动子驱动zs-yellow基因表达,zs-yellow基因赋予转化种子黄色荧光。按照实施例1描述的方法克隆水稻耐旱性基因,并构建gateway载体;随后采用invitrogentm的
t2种子用于实验室干旱试验,拟南芥的干旱试验是一种基于土壤的水分限制试验,在生长室中进行,光照强度为145μmol,温度22℃白天/20℃夜晚,湿度为60%。转基因种子采用copastm(复杂对象的参数分析和分选机、种子分选机,unionbiometrica)分离,然后成层放置在0.1%的琼脂溶液中,4℃放置3天。野生型拟南芥用作对照,按照上述方法打破休眠。36株过表达拟南芥和野生型等距种植。土壤的成分为3份泥煤苔、2份蛭石和1份珍珠岩,除此之外,肥料和杀真菌剂以下述的浓度添加在土壤中:npk(氮磷钾)-1m/kg土壤、微量营养物-0.5g/kg土壤、杀真菌剂-0.5g/kg土壤。间苗后,每盆中有9株拟南芥,一个平盘中有72株拟南芥,前12天正常浇水,最后以1l去离子水是拟南芥的土壤饱和30min,多余的水分完全流出。种子萌发后28天到36天,采用成像仪使植株成像,并分析成像数据。种子种植后的第二天开始每天转动平盘直至成像的最后一天。成像系统产生的文件转变成xls文件,并转换成stan’s格式,发送给esl产生每个试验株系的stan’s分数。干旱条件下损害或萎蔫的速率用作测试参数,截止点分数=1.5。
序列表
<110>未名生物农业集团有限公司
先锋海外公司
<120>非生物胁迫耐性提高的植物和方法
<130>rts22593c
<150>201610507365.2
<151>2016-06-30
<160>57
<170>patentinversion3.5
<210>1
<211>11934
<212>dna
<213>人工序列
<220>
<223>dp0158载体的核苷酸序列
<400>1
gaattctctagtcccgatctagtaacatagatgacaccgcgcgcgataatttatcctagt60
ttgcgcgctatattttgttttctatcgcgtattaaatgtataattgcgggactctaatca120
taaaaacccatctcataaataacgtcatgcattacatgttaattattacatgcttaacgt180
aattcaacagaaattatatgataatcatcgcaagaccggcaacaggattcaatcttaaga240
aacgcggccgcttcagttgtggcccagcttggaggtcgactcgcgaggatctctgcagag300
agatagatttgtagagagagactggtgatttcagcgtgtcctctccaaatgaaatgaact360
tccttatatagaggaagggtcttgcgaaggatagtgggattgtgcgtcatcccttacgtc420
agtggagatatcacatcaatccacttgctttgaagacgtggttggaacgtcttctttttc480
cacgatgctcctcgtgggtgggggtccatctttgggaccactgtcggcagaggcatcttg540
aacgatagcctttcctttatcgcaatgatggcatttgtaggtgccaccttccttttctac600
tgtccttttgatgaagtgacagatagctgggcaatggaatccgaggaggtttcccgatat660
taccctttgttgaaaagtctcaatagccctttggtcttctgagactgtatctttgatatt720
cttggagtagacgagagtgtcgtgctccaccatgttcacatcaatccacttgctttgaag780
acgtggttggaacgtcttctttttccacgatgctcctcgtgggtgggggtccatctttgg840
gaccactgtcggcagaggcatcttgaacgatagcctttcctttatcgcaatgatggcatt900
tgtaggtgccaccttccttttctactgtccttttgatgaagtgacagatagctgggcaat960
ggaatccgaggaggtttcccgatattaccctttgttgaaaagtctcaatagccctttggt1020
cttctgagactgtatctttgatattcttggagtagacgagagtgtcgtgctccaccatgt1080
tgccaagctgctctaagctttggcggccgcattcgcaaaacacacctagactagatttgt1140
tttgctaacccaattgatattaattatatatgattaatatttatatgtatatggatttgg1200
ttaatgaaatgcatctggttcatcaaagaattataaagacacgtgacattcatttaggat1260
aagaaatatggatgatctctttctcttttattcagataactagtaattacacataacaca1320
caactttgatgcccacattatagtgattagcatgtcactatgtgtgcatccttttatttc1380
atacattaattaagttggccaatccagaagatggacaagtctaggttaaccatgtggtac1440
ctacgcgttcgaatatccatgggccgctacaggaacaggtggtggcggccctcggtgcgc1500
tcgtactgctccacgatggtgtagtcctcgttgtgggaggtgatgtccagcttggcgtcc1560
acgtagtagtagccgggcagctgcacgggcttcttggccatgtagatggacttgaactcc1620
accaggtagtggccgccgtccttcagcttcagggccttgtgggtctcgcccttcagcacg1680
ccgtcgcgggggtacaggcgctcggtggaggcctcccagcccatggtcttcttctgcatc1740
acggggccgtcggaggggaagttcacgccgatgaacttcaccttgtagatgaagcagccg1800
tcctgcagggaggagtcctgggtcacggtcgccacgccgccgtcctcgaagttcatcacg1860
cgctcccacttgaagccctcggggaaggacagcttcttgtagtcggggatgtcggcgggg1920
tgcttcacgtacaccttggagccgtactggaactggggggacaggatgtcccaggcgaag1980
ggcagggggccgcccttcgtcaccttcagcttcacggtgttgtggccctcgtaggggcgg2040
ccctcgccctcgccctcgatctcgaactcgtggccgttcacggtgccctccatgcgcacc2100
ttgaagcgcatgaactcggtgatgacgttctcggaggaggccatggtggcgaggatctac2160
tcggctacactcacacgctcgctctcgcagttgcaggtgtaagtttctagctagggcact2220
cacggggtacgtatttgtagccagccacgcacggtctgagctcgccatgtgccgccatgc2280
atgcgggggcacgtcgccagcgtacgcggccatcgtcgctgacgaaggtagcgcattcaa2340
gtccggtcggtagaggtcagctgggtcgttcgccgatggtagttgccgcccggactcagt2400
gggcggtaggcgaaggctagcaagcagacgactccattcatgcgcatcatccaaaggtga2460
tgcaaagccttccaaacgcgattgtctcatgatgtttccgtctcttgttacgaggagtac2520
aattttttcttatacacgaacgttactttatgtcacatttccatgccatgaacaccttgg2580
cttcaaataagtgagtgttttttttcacattctgtggcataaacagaatttctagagtgg2640
catttgtgatacattgtgaaagctaagagtggtaaaagtaaaataaaattgttttgcttt2700
tgccgcggaatggaaattatttgtcaaaacctaagagtggcaaaactgaaatgtcaaaac2760
ctagagtgacataaacaaaatttacccatcactaaatgagcacaaaatatttcaccacaa2820
tggaggtatgtgaggtccgatgtactactagagctcatcggaaaagcatcctcttgatga2880
gtaaacctcttgaagtactgtaccaccacattttatttatcctcatcggcttatttttag2940
gccacggttattctcacgaagagacggttaacccttctcgtagactacacatcgagatcc3000
actagttctagagcggccagcttcgaagcttggcactggccgtcgttttacaacgtcgtg3060
actgggaaaaccctggcgttacccaacttaatcgccttgcagcacatccccctttcgcca3120
gctggcgtaatagcgaagaggcccgcaccgatcgcccttcccaacagttgcgcagcctga3180
atggcgaatgctagagcagcttgagcttggatcagattgtcgtttcccgccttcagttta3240
aactatcagtgtttgacaggatatattggcgggtaaacctaagagaaaagagcgtttatt3300
agaataatcggatatttaaaagggcgtgaaaaggtttatccgttcgtccatttgtatgtg3360
catgccaaccacagggttcccctcgggatcaaagtactttgatccaacccctccgctgct3420
atagtgcagtcggcttctgacgttcagtgcagccgtcttctgaaaacgacatgtcgcaca3480
agtcctaagttacgcgacaggctgccgccctgcccttttcctggcgttttcttgtcgcgt3540
gttttagtcgcataaagtagaatacttgcgactagaaccggagacattacgccatgaaca3600
agagcgccgccgctggcctgctgggctatgcccgcgtcagcaccgacgaccaggacttga3660
ccaaccaacgggccgaactgcacgcggccggctgcaccaagctgttttccgagaagatca3720
ccggcaccaggcgcgaccgcccggagctggccaggatgcttgaccacctacgccctggcg3780
acgttgtgacagtgaccaggctagaccgcctggcccgcagcacccgcgacctactggaca3840
ttgccgagcgcatccaggaggccggcgcgggcctgcgtagcctggcagagccgtgggccg3900
acaccaccacgccggccggccgcatggtgttgaccgtgttcgccggcattgccgagttcg3960
agcgttccctaatcatcgaccgcacccggagcgggcgcgaggccgccaaggcccgaggcg4020
tgaagtttggcccccgccctaccctcaccccggcacagatcgcgcacgcccgcgagctga4080
tcgaccaggaaggccgcaccgtgaaagaggcggctgcactgcttggcgtgcatcgctcga4140
ccctgtaccgcgcacttgagcgcagcgaggaagtgacgcccaccgaggccaggcggcgcg4200
gtgccttccgtgaggacgcattgaccgaggccgacgccctggcggccgccgagaatgaac4260
gccaagaggaacaagcatgaaaccgcaccaggacggccaggacgaaccgtttttcattac4320
cgaagagatcgaggcggagatgatcgcggccgggtacgtgttcgagccgcccgcgcacgt4380
ctcaaccgtgcggctgcatgaaatcctggccggtttgtctgatgccaagctggcggcctg4440
gccggccagcttggccgctgaagaaaccgagcgccgccgtctaaaaaggtgatgtgtatt4500
tgagtaaaacagcttgcgtcatgcggtcgctgcgtatatgatgcgatgagtaaataaaca4560
aatacgcaaggggaacgcatgaaggttatcgctgtacttaaccagaaaggcgggtcaggc4620
aagacgaccatcgcaacccatctagcccgcgccctgcaactcgccggggccgatgttctg4680
ttagtcgattccgatccccagggcagtgcccgcgattgggcggccgtgcgggaagatcaa4740
ccgctaaccgttgtcggcatcgaccgcccgacgattgaccgcgacgtgaaggccatcggc4800
cggcgcgacttcgtagtgatcgacggagcgccccaggcggcggacttggctgtgtccgcg4860
atcaaggcagccgacttcgtgctgattccggtgcagccaagcccttacgacatatgggcc4920
accgccgacctggtggagctggttaagcagcgcattgaggtcacggatggaaggctacaa4980
gcggcctttgtcgtgtcgcgggcgatcaaaggcacgcgcatcggcggtgaggttgccgag5040
gcgctggccgggtacgagctgcccattcttgagtcccgtatcacgcagcgcgtgagctac5100
ccaggcactgccgccgccggcacaaccgttcttgaatcagaacccgagggcgacgctgcc5160
cgcgaggtccaggcgctggccgctgaaattaaatcaaaactcatttgagttaatgaggta5220
aagagaaaatgagcaaaagcacaaacacgctaagtgccggccgtccgagcgcacgcagca5280
gcaaggctgcaacgttggccagcctggcagacacgccagccatgaagcgggtcaactttc5340
agttgccggcggaggatcacaccaagctgaagatgtacgcggtacgccaaggcaagacca5400
ttaccgagctgctatctgaatacatcgcgcagctaccagagtaaatgagcaaatgaataa5460
atgagtagatgaattttagcggctaaaggaggcggcatggaaaatcaagaacaaccaggc5520
accgacgccgtggaatgccccatgtgtggaggaacgggcggttggccaggcgtaagcggc5580
tgggttgtctgccggccctgcaatggcactggaacccccaagcccgaggaatcggcgtga5640
cggtcgcaaaccatccggcccggtacaaatcggcgcggcgctgggtgatgacctggtgga5700
gaagttgaaggccgcgcaggccgcccagcggcaacgcatcgaggcagaagcacgccccgg5760
tgaatcgtggcaagcggccgctgatcgaatccgcaaagaatcccggcaaccgccggcagc5820
cggtgcgccgtcgattaggaagccgcccaagggcgacgagcaaccagattttttcgttcc5880
gatgctctatgacgtgggcacccgcgatagtcgcagcatcatggacgtggccgttttccg5940
tctgtcgaagcgtgaccgacgagctggcgaggtgatccgctacgagcttccagacgggca6000
cgtagaggtttccgcagggccggccggcatggccagtgtgtgggattacgacctggtact6060
gatggcggtttcccatctaaccgaatccatgaaccgataccgggaagggaagggagacaa6120
gcccggccgcgtgttccgtccacacgttgcggacgtactcaagttctgccggcgagccga6180
tggcggaaagcagaaagacgacctggtagaaacctgcattcggttaaacaccacgcacgt6240
tgccatgcagcgtacgaagaaggccaagaacggccgcctggtgacggtatccgagggtga6300
agccttgattagccgctacaagatcgtaaagagcgaaaccgggcggccggagtacatcga6360
gatcgagctagctgattggatgtaccgcgagatcacagaaggcaagaacccggacgtgct6420
gacggttcaccccgattactttttgatcgatcccggcatcggccgttttctctaccgcct6480
ggcacgccgcgccgcaggcaaggcagaagccagatggttgttcaagacgatctacgaacg6540
cagtggcagcgccggagagttcaagaagttctgtttcaccgtgcgcaagctgatcgggtc6600
aaatgacctgccggagtacgatttgaaggaggaggcggggcaggctggcccgatcctagt6660
catgcgctaccgcaacctgatcgagggcgaagcatccgccggttcctaatgtacggagca6720
gatgctagggcaaattgccctagcaggggaaaaaggtcgaaaaggtctctttcctgtgga6780
tagcacgtacattgggaacccaaagccgtacattgggaaccggaacccgtacattgggaa6840
cccaaagccgtacattgggaaccggtcacacatgtaagtgactgatataaaagagaaaaa6900
aggcgatttttccgcctaaaactctttaaaacttattaaaactcttaaaacccgcctggc6960
ctgtgcataactgtctggccagcgcacagccgaagagctgcaaaaagcgcctacccttcg7020
gtcgctgcgctccctacgccccgccgcttcgcgtcggcctatcgcggccgctggccgctc7080
aaaaatggctggcctacggccaggcaatctaccagggcgcggacaagccgcgccgtcgcc7140
actcgaccgccggcgcccacatcaaggcaccctgcctcgcgcgtttcggtgatgacggtg7200
aaaacctctgacacatgcagctcccggagacggtcacagcttgtctgtaagcggatgccg7260
ggagcagacaagcccgtcagggcgcgtcagcgggtgttggcgggtgtcggggcgcagcca7320
tgacccagtcacgtagcgatagcggagtgtatactggcttaactatgcggcatcagagca7380
gattgtactgagagtgcaccatatgcggtgtgaaataccgcacagatgcgtaaggagaaa7440
ataccgcatcaggcgctcttccgcttcctcgctcactgactcgctgcgctcggtcgttcg7500
gctgcggcgagcggtatcagctcactcaaaggcggtaatacggttatccacagaatcagg7560
ggataacgcaggaaagaacatgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaa7620
ggccgcgttgctggcgtttttccataggctccgcccccctgacgagcatcacaaaaatcg7680
acgctcaagtcagaggtggcgaaacccgacaggactataaagataccaggcgtttccccc7740
tggaagctccctcgtgcgctctcctgttccgaccctgccgcttaccggatacctgtccgc7800
ctttctcccttcgggaagcgtggcgctttctcatagctcacgctgtaggtatctcagttc7860
ggtgtaggtcgttcgctccaagctgggctgtgtgcacgaaccccccgttcagcccgaccg7920
ctgcgccttatccggtaactatcgtcttgagtccaacccggtaagacacgacttatcgcc7980
actggcagcagccactggtaacaggattagcagagcgaggtatgtaggcggtgctacaga8040
gttcttgaagtggtggcctaactacggctacactagaaggacagtatttggtatctgcgc8100
tctgctgaagccagttaccttcggaaaaagagttggtagctcttgatccggcaaacaaac8160
caccgctggtagcggtggtttttttgtttgcaagcagcagattacgcgcagaaaaaaagg8220
atctcaagaagatcctttgatcttttctacggggtctgacgctcagtggaacgaaaactc8280
acgttaagggattttggtcatgcattctaggtactaaaacaattcatccagtaaaatata8340
atattttattttctcccaatcaggcttgatccccagtaagtcaaaaaatagctcgacata8400
ctgttcttccccgatatcctccctgatcgaccggacgcagaaggcaatgtcataccactt8460
gtccgccctgccgcttctcccaagatcaataaagccacttactttgccatctttcacaaa8520
gatgttgctgtctcccaggtcgccgtgggaaaagacaagttcctcttcgggcttttccgt8580
ctttaaaaaatcatacagctcgcgcggatctttaaatggagtgtcttcttcccagttttc8640
gcaatccacatcggccagatcgttattcagtaagtaatccaattcggctaagcggctgtc8700
taagctattcgtatagggacaatccgatatgtcgatggagtgaaagagcctgatgcactc8760
cgcatacagctcgataatcttttcagggctttgttcatcttcatactcttccgagcaaag8820
gacgccatcggcctcactcatgagcagattgctccagccatcatgccgttcaaagtgcag8880
gacctttggaacaggcagctttccttccagccatagcatcatgtccttttcccgttccac8940
atcataggtggtccctttataccggctgtccgtcatttttaaatataggttttcattttc9000
tcccaccagcttatataccttagcaggagacattccttccgtatcttttacgcagcggta9060
tttttcgatcagttttttcaattccggtgatattctcattttagccatttattatttcct9120
tcctcttttctacagtatttaaagataccccaagaagctaattataacaagacgaactcc9180
aattcactgttccttgcattctaaaaccttaaataccagaaaacagctttttcaaagttg9240
ttttcaaagttggcgtataacatagtatcgacggagccgattttgaaaccgcggtgatca9300
caggcagcaacgctctgtcatcgttacaatcaacatgctaccctccgcgagatcatccgt9360
gtttcaaacccggcagcttagttgccgttcttccgaatagcatcggtaacatgagcaaag9420
tctgccgccttacaacggctctcccgctgacgccgtcccggactgatgggctgcctgtat9480
cgagtggtgattttgtgccgagctgccggtcggggagctgttggctggctggtggcagga9540
tatattgtggtgtaaacaaattgacgcttagacaacttaataacacattgcggacgtttt9600
taatgtactgaattaacgccgaattaattcgggggatctggattttagtactggattttg9660
gttttaggaattagaaattttattgatagaagtattttacaaatacaaatacatactaag9720
ggtttcttatatgctcaacacatgagcgaaaccctataggaaccctaattcccttatctg9780
ggaactactcacacattattatggagaaactcgagcttgtcgatcgacagatccggtcgg9840
catctactctatttctttgccctcggacgagtgctggggcgtcggtttccactatcggcg9900
agtacttctacacagccatcggtccagacggccgcgcttctgcgggcgatttgtgtacgc9960
ccgacagtcccggctccggatcggacgattgcgtcgcatcgaccctgcgcccaagctgca10020
tcatcgaaattgccgtcaaccaagctctgatagagttggtcaagaccaatgcggagcata10080
tacgcccggagtcgtggcgatcctgcaagctccggatgcctccgctcgaagtagcgcgtc10140
tgctgctccatacaagccaaccacggcctccagaagaagatgttggcgacctcgtattgg10200
gaatccccgaacatcgcctcgctccagtcaatgaccgctgttatgcggccattgtccgtc10260
aggacattgttggagccgaaatccgcgtgcacgaggtgccggacttcggggcagtcctcg10320
gcccaaagcatcagctcatcgagagcctgcgcgacggacgcactgacggtgtcgtccatc10380
acagtttgccagtgatacacatggggatcagcaatcgcgcatatgaaatcacgccatgta10440
gtgtattgaccgattccttgcggtccgaatgggccgaacccgctcgtctggctaagatcg10500
gccgcagcgatcgcatccatagcctccgcgaccggttgtagaacagcgggcagttcggtt10560
tcaggcaggtcttgcaacgtgacaccctgtgcacggcgggagatgcaataggtcaggctc10620
tcgctaaactccccaatgtcaagcacttccggaatcgggagcgcggccgatgcaaagtgc10680
cgataaacataacgatctttgtagaaaccatcggcgcagctatttacccgcaggacatat10740
ccacgccctcctacatcgaagctgaaagcacgagattcttcgccctccgagagctgcatc10800
aggtcggagacgctgtcgaacttttcgatcagaaacttctcgacagacgtcgcggtgagt10860
tcaggctttttcatatctcattgccccccgggatctgcgaaagctcgagagagatagatt10920
tgtagagagagactggtgatttcagcgtgtcctctccaaatgaaatgaacttccttatat10980
agaggaaggtcttgcgaaggatagtgggattgtgcgtcatcccttacgtcagtggagata11040
tcacatcaatccacttgctttgaagacgtggttggaacgtcttctttttccacgatgctc11100
ctcgtgggtgggggtccatctttgggaccactgtcggcagaggcatcttgaacgatagcc11160
tttcctttatcgcaatgatggcatttgtaggtgccaccttccttttctactgtccttttg11220
atgaagtgacagatagctgggcaatggaatccgaggaggtttcccgatattaccctttgt11280
tgaaaagtctcaatagccctttggtcttctgagactgtatctttgatattcttggagtag11340
acgagagtgtcgtgctccaccatgttatcacatcaatccacttgctttgaagacgtggtt11400
ggaacgtcttctttttccacgatgctcctcgtgggtgggggtccatctttgggaccactg11460
tcggcagaggcatcttgaacgatagcctttcctttatcgcaatgatggcatttgtaggtg11520
ccaccttccttttctactgtccttttgatgaagtgacagatagctgggcaatggaatccg11580
aggaggtttcccgatattaccctttgttgaaaagtctcaatagccctttggtcttctgag11640
actgtatctttgatattcttggagtagacgagagtgtcgtgctccaccatgttggcaagc11700
tgctctagccaatacgcaaaccgcctctccccgcgcgttggccgattcattaatgcagct11760
ggcacgacaggtttcccgactggaaagcgggcagtgagcgcaacgcaattaatgtgagtt11820
agctcactcattaggcaccccaggctttacactttatgcttccggctcgtatgttgtgtg11880
gaattgtgagcggataacaatttcacacaggaaacagctatgaccatgattacg11934
<210>2
<211>1349
<212>dna
<213>水稻(oryzasativa)
<400>2
gactccgacgaccagaagctaccggcgacgcgagcggagaagcgcggaggggagggggcg60
cgcgccgccatggcgttcgagaagatcgtcgtggccaaccccgtcgtcgagatggacggt120
gatgagatgactcgagttatttggaaatggatcaaagataagcttatattccctttcttg180
gacttggacataaaatactatgacttaggtctacctaatcgtgacgctactggggacaaa240
gttacaatagagagtgcagaagcgaccctaaagtataatgtagccatcaaatgtgcaact300
ataactccagatgaaggacgcgtgaaagagtttaatttaagtgcaatgtggaagagtcca360
aatgggacaataaggaacattttgaatggaactgttttccgagaaccaatcatctgcaag420
aatattcctcggcttgtacctgggtggataaagcccatatgcattggacgacatgcattt480
ggtgatcaataccgagcaacagatacagttattaaaggtcctgggaagttgaagttagta540
tttgatggcagagaggaacaaatagagttggatgtgttcaactttactggtgctggtgga600
gtagccttgtctatgtataatactgacgagtctatttgggcattcgctgaagcttccatg660
aacatggcttaccagaaaagatggccactttatcttagcaccaaaaacacgatcctcaaa720
aaatacgatggaaggtttaaagatatatttcaggagaactatgaaacaaaatggagagcc780
aagtttgatgatgcaggaatatggtacgaacatcggctgattgatgatatggtggcctat840
gcccttaagagtgaaggtggctatgtttgggcttgcaagaactatgatggagatgtgcag900
agcgatctaattgctcaaggttttggatcgctaggtctaatgacatcagttctggtgtgc960
cctgatggtagaaccattgaagctgaagctgctcatggtacagtcacacgccattacaga1020
gttcaccaaaaaggaggcgaaactagtacaaatagcattgcttcaatatttgcttggaca1080
accggactaggacatagggcaaagctcgatgacaataaaagactgttagattttgtacaa1140
aaacttgaagctgcttgtgtgggaacagtggaatctggaaagatgacaaaggatctagct1200
cttcttgtacatgggccaaatgttagccgagataagtatcttaacactgttgagttcatc1260
gatgctgttgctgaggatttaagaacaagattgtcagtaacatccaagttatgaaggaat1320
gaatagggcaacggcaggggacatgttag1349
<210>3
<211>1245
<212>dna
<213>水稻(oryzasativa)
<400>3
atggcgttcgagaagatcgtcgtggccaaccccgtcgtcgagatggacggtgatgagatg60
actcgagttatttggaaatggatcaaagataagcttatattccctttcttggacttggac120
ataaaatactatgacttaggtctacctaatcgtgacgctactggggacaaagttacaata180
gagagtgcagaagcgaccctaaagtataatgtagccatcaaatgtgcaactataactcca240
gatgaaggacgcgtgaaagagtttaatttaagtgcaatgtggaagagtccaaatgggaca300
ataaggaacattttgaatggaactgttttccgagaaccaatcatctgcaagaatattcct360
cggcttgtacctgggtggataaagcccatatgcattggacgacatgcatttggtgatcaa420
taccgagcaacagatacagttattaaaggtcctgggaagttgaagttagtatttgatggc480
agagaggaacaaatagagttggatgtgttcaactttactggtgctggtggagtagccttg540
tctatgtataatactgacgagtctatttgggcattcgctgaagcttccatgaacatggct600
taccagaaaagatggccactttatcttagcaccaaaaacacgatcctcaaaaaatacgat660
ggaaggtttaaagatatatttcaggagaactatgaaacaaaatggagagccaagtttgat720
gatgcaggaatatggtacgaacatcggctgattgatgatatggtggcctatgcccttaag780
agtgaaggtggctatgtttgggcttgcaagaactatgatggagatgtgcagagcgatcta840
attgctcaaggttttggatcgctaggtctaatgacatcagttctggtgtgccctgatggt900
agaaccattgaagctgaagctgctcatggtacagtcacacgccattacagagttcaccaa960
aaaggaggcgaaactagtacaaatagcattgcttcaatatttgcttggacaaccggacta1020
ggacatagggcaaagctcgatgacaataaaagactgttagattttgtacaaaaacttgaa1080
gctgcttgtgtgggaacagtggaatctggaaagatgacaaaggatctagctcttcttgta1140
catgggccaaatgttagccgagataagtatcttaacactgttgagttcatcgatgctgtt1200
gctgaggatttaagaacaagattgtcagtaacatccaagttatga1245
<210>4
<211>414
<212>prt
<213>水稻(oryzasativa)
<400>4
metalapheglulysilevalvalalaasnprovalvalglumetasp
151015
glyaspglumetthrargvaliletrplystrpilelysasplysleu
202530
ilephepropheleuaspleuaspilelystyrtyraspleuglyleu
354045
proasnargaspalathrglyasplysvalthrilegluseralaglu
505560
alathrleulystyrasnvalalailelyscysalathrilethrpro
65707580
aspgluglyargvallysglupheasnleuseralamettrplysser
859095
proasnglythrileargasnileleuasnglythrvalpheargglu
100105110
proileilecyslysasnileproargleuvalproglytrpilelys
115120125
proilecysileglyarghisalapheglyaspglntyrargalathr
130135140
aspthrvalilelysglyproglylysleulysleuvalpheaspgly
145150155160
argglugluglnilegluleuaspvalpheasnphethrglyalagly
165170175
glyvalalaleusermettyrasnthraspgluseriletrpalaphe
180185190
alaglualasermetasnmetalatyrglnlysargtrpproleutyr
195200205
leuserthrlysasnthrileleulyslystyraspglyargphelys
210215220
aspilepheglngluasntyrgluthrlystrpargalalyspheasp
225230235240
aspalaglyiletrptyrgluhisargleuileaspaspmetvalala
245250255
tyralaleulyssergluglyglytyrvaltrpalacyslysasntyr
260265270
aspglyaspvalglnseraspleuilealaglnglypheglyserleu
275280285
glyleumetthrservalleuvalcysproaspglyargthrileglu
290295300
alaglualaalahisglythrvalthrarghistyrargvalhisgln
305310315320
lysglyglygluthrserthrasnserilealaserilephealatrp
325330335
thrthrglyleuglyhisargalalysleuaspaspasnlysargleu
340345350
leuaspphevalglnlysleuglualaalacysvalglythrvalglu
355360365
serglylysmetthrlysaspleualaleuleuvalhisglyproasn
370375380
valserargasplystyrleuasnthrvalglupheileaspalaval
385390395400
alagluaspleuargthrargleuservalthrserlysleu
405410
<210>5
<211>631
<212>dna
<213>水稻(oryzasativa)
<400>5
cactgacatgtggccttccttcttcctcccccctctctcccttctctctctttctctccc60
cttcttccccttctgtcgacacgcgggatggcaacaagagcgagcagcggcgggcggcga120
ggccggagctgcggcggtgggcgggagggcggcgagatcgagcggcggcgcgctgcgacg180
gcgagggagctgcgctcggcgggagagcaagcggcgaaggaaaaggacccccaccctaga240
caccgcccagcctggagctgcgctcggcggcgagagcgagcggcggcgacaagtgtaagc300
acacgttcggtttgttttttttcttgtttctttgattcgtagccgaagttcataactgaa360
ttttccttgtttcaggacgacgtttaagaggatcctgaaggccaaatcgaccgagcgatt420
cgatgggctcccttacctgttctcgctgctgaactgcctcatctgcctgtggtatggact480
tccctgggtcgccaatggcaggctgctcgtcaccaccgtcaacggcaccggagcggtgtt540
ccagctcgcctacatttgtctcttcatcttctacgcggacagcaagaagacgagcgtaat600
tcttcccatcctgcacctaattttgtcttag631
<210>6
<211>543
<212>dna
<213>水稻(oryzasativa)
<400>6
atgtggccttccttcttcctcccccctctctcccttctctctctttctctccccttcttc60
cccttctgtcgacacgcgggatggcaacaagagcgagcagcggcgggcggcgaggccgga120
gctgcggcggtgggcgggagggcggcgagatcgagcggcggcgcgctgcgacggcgaggg180
agctgcgctcggcgggagagcaagcggcgaaggaaaaggacccccaccctagacaccgcc240
cagcctggagctgcgctcggcggcgagagcgagcggcggcgacaagtgacgacgtttaag300
aggatcctgaaggccaaatcgaccgagcgattcgatgggctcccttacctgttctcgctg360
ctgaactgcctcatctgcctgtggtatggacttccctgggtcgccaatggcaggctgctc420
gtcaccaccgtcaacggcaccggagcggtgttccagctcgcctacatttgtctcttcatc480
ttctacgcggacagcaagaagacgagcgtaattcttcccatcctgcacctaattttgtct540
tag543
<210>7
<211>180
<212>prt
<213>水稻(oryzasativa)
<400>7
mettrpproserphepheleuproproleuserleuleuserleuser
151015
leuprophepheprophecysarghisalaglytrpglnglngluarg
202530
alaalaalaglyglyglualaglyalaalaalavalglyglyargala
354045
alaargserserglyglyalaleuargargargglysercysalaarg
505560
arggluserlysargargarglysargthrprothrleuaspthrala
65707580
glnproglyalaalaleuglyglyglusergluargargargglnval
859095
thrthrphelysargileleulysalalysserthrgluargpheasp
100105110
glyleuprotyrleupheserleuleuasncysleuilecysleutrp
115120125
tyrglyleuprotrpvalalaasnglyargleuleuvalthrthrval
130135140
asnglythrglyalavalpheglnleualatyrilecysleupheile
145150155160
phetyralaaspserlyslysthrservalileleuproileleuhis
165170175
leuileleuser
180
<210>8
<211>1459
<212>dna
<213>水稻(oryzasativa)
<400>8
gctgcaaaggaggaagagaaagagtgttgaccgggggagaagagaggagaagccgggaag60
agggccgggatggcgtcgagcaacggagagcaacctccagaatcgccgaaggaagcaaat120
gctgcgcctgtagcagccaagcttgccatggcgtcagggcgcgatggatgtgagaggctc180
aaggatctggtgagtagggaagacgatgctactacgatggttgtggccatggcgacaagc240
aagaacgtggacgacacaagacgacctccccctccccgtgtaattatggatccccagctc300
ctcatggctgcacggaatggagcttgccagtctcttgagagcctagcttctcctcggcca360
tgaggtgcgttgtcaaaccttgcctgaatcccatctaatcatctacttgcccgaaccaga420
tgagggagctccgaccacagatccaattgaagatcaggctgaaggaatctctgcgactga480
tcagcaagcctctggtgctgtatattcgctctcgctcctggagggactgactcttgactc540
ggaccagaactccgcgctgcatgtggtggcagcctccggtgacagccaggcgtatgtgga600
gtgtgcgcggatggtgtacgaccaggcgaggcacctactcggcgcggccaacaacaacgg660
cgacacgcccctgcactgcgccgctgcagctgggaaccactcctcgcgctggctgctgct720
gcgcctgcgcgtgacagcagcggtgcagcgagggtggatgaccaggagctcctagcaagg780
aggaagaacaaggtcggtgagacggctttgcacggggcggtccgtgctggccacagcaaa840
gtggtggaggtcctcatgaaggaggatccgggactggccggcgttgatcggcacgacggc900
acgtcaccgctgtaccttgccgtttcgttgggacgcttcgagattgcgtgggatctgctt960
gatatgagctctaggaaattatcttactccggaccagatggtcagaatgtcttgcacgtc1020
gcagtacaacacccccaaggtatgcgccacattaccgtactctctttgttgcccttccat1080
tacatcagtaataacaaaataattatgtttgttttaaaagatcaattaaatccaactcat1140
tatatattcaacaatgcacagcaccgttaaacaacctgtaaataaagctgtatcgaaaat1200
tatggttttgaggatatcatctaattaagattgtttgtataaaaaggaagagattcgtga1260
tccccctatatatatactgaggcaaagacttttttttttagtacagattaattaatttgg1320
gtctcttactccttttatgataataactctatttttatcttggatttgtatatacttttg1380
gatataaagagcataggtaggaagtagacctaatcgactatctcgtcaacatgtttgggt1440
atgggtttgttgcgatctc1459
<210>9
<211>699
<212>dna
<213>水稻(oryzasativa)
<400>9
atggcgtcgagcaacggagagcaacctccagaatcgccgaaggaagcaaatgctgcgcct60
gtagcagccaagcttgccatggcgtcagggcgcgatggatgtgagaggctcaaggatctg120
atgagggagctccgaccacagatccaattgaagatcaggctgaaggaatctctgcgactg180
atcagcaagcctctgccaggcgtatgtggagtgtgcgcggatggtgtacgaccaggcgag240
gcacctactcggcgcggccaacaacaacggcgacacgcccctgcactgcgccgctgcagc300
tgggaaccactcctcgcgctggctgctgctgcgcctgcgcgtgacagcagcggtgcagcg360
agggtggatgaccaggagctcctagcaaggaggaagaacaaggtcggtgagacggctttg420
cacggggcggtccgtgctggccacagcaaagtggtggaggtcctcatgaaggaggatccg480
ggactggccggcgttgatcggcacgacggcacgtcaccgctgtaccttgccgtttcgttg540
ggacgcttcgagattgcgtgggatctgcttgatatgagctctaggaaattatcttactcc600
ggaccagatggtcagaatgtcttgcacgtcgcagtacaacacccccaagatcaattaaat660
ccaactcattatatattcaacaatgcacagcaccgttaa699
<210>10
<211>232
<212>prt
<213>水稻(oryzasativa)
<400>10
metalaserserasnglygluglnproprogluserprolysgluala
151015
asnalaalaprovalalaalalysleualametalaserglyargasp
202530
glycysgluargleulysaspleumetarggluleuargproglnile
354045
glnleulysileargleulysgluserleuargleuileserlyspro
505560
leuproglyvalcysglyvalcysalaaspglyvalargproglyglu
65707580
alaprothrargargglyglnglnglnargarghisalaproalaleu
859095
argargcyssertrpgluproleuleualaleualaalaalaalapro
100105110
alaargaspserserglyalaalaargvalaspaspglngluleuleu
115120125
alaargarglysasnlysvalglygluthralaleuhisglyalaval
130135140
argalaglyhisserlysvalvalgluvalleumetlysgluasppro
145150155160
glyleualaglyvalasparghisaspglythrserproleutyrleu
165170175
alavalserleuglyargphegluilealatrpaspleuleuaspmet
180185190
serserarglysleusertyrserglyproaspglyglnasnvalleu
195200205
hisvalalavalglnhisproglnaspglnleuasnprothrhistyr
210215220
ilepheasnasnalaglnhisarg
225230
<210>11
<211>2829
<212>dna
<213>水稻(oryzasativa)
<400>11
caagtacctgtaaattgaaacctgcagacactctggaagtaggcagattcacattgtgtc60
agcggtcgccggtcggacgatgtccagggcaaacagtcgacagacaaaggcacaaagctg120
tgaagggctgaatgctgaagggatcgccggcggcccagcgtggggtgaacttgcagaggc180
gccgtggagggcgcagggcgatggaggcggattggcggagactgccgccggtagaggaga240
ggagacgagaagactggaactcggacggtcgcgtatgcccgacgattcgatcagaaactt300
gtatacatgtatatggtatacctaatacatacctaggtcttctgggcccctacttttcct360
gggccctaggccatcgcacttcttgccgtagcccagagccggccctgaaaaagaggagat420
acaagtgcacgtgagggagagaggagagggagggattacattctgtttataatctaatct480
aatctaatggtctaaaacaatgagcccaccaatttaaatgaaaaaaattgagggctagat540
gtttctaatctattagaatgccacttggtggtttagaagcgtttacagaaatgccacatt600
ggcatgagaacgcttgtagtatgtttaatggacttgttcaagataaagtggtataattca660
aatatcataaaatttggtgttatatatgaaaaatgtcatgtttaagagtgccatatgtta720
attttggcagtgcagttggatgctgaaagccgacagagagaaaattaaatcaaagatgac780
cttttgttagggatcgatgtggaagaacatgactgtattactgtaatagaagtttattaa840
ataatcttaacatcttagagatatttcattctagagatcttatgtctccagttggttgta900
ggtagattggacgtccagtatttctatgctaacatggccctccgctattgcaaggaaccc960
taggaagattgattaatttagttttctaaatttgattttggttccatttgccctttgggc1020
ttgcttaactgatggtatcacctgctgcagcactgtctgtactgttagagaagtgcaaga1080
atgtcgaggtcaatgtacagcaagaagaccaacaaaggagtattcctcttctcttgtacc1140
tcactagtcaaagtgacaaaaatgggagcactcctcttcactttgctgcatcgttgaaaa1200
ctagtattgaaggtttcacatcgagattatgtgaacattttcgcccgaaacaaagcccaa1260
caacattgcttcttggactcaatgaatctgctatatatcagccagacaacagaggatcat1320
atcctatacttgttgctgcctccaatgggatactcaaggttgtcataaccttgctcaaaa1380
ggtatccagattgtgccactttgcgtgacatccaaggaagaactttcttccatgttgccg1440
tggagaagaagcggcgtaatatcgttgcatatgtgtgtgaaaggccaggattttctccta1500
tcttgaatatgcaggacagccatggggatactgctctacaccttgctgtcaaagccggtg1560
ttttttcaattttcagctcgttgttccggaataggtaggtttgcctgaatttatcaaatg1620
aggatggtttgacacctcgtgatctttcgtggattatgatacctgcgaggttgtactcta1680
aaaaggtaatcgttctctgccttttctgaccataattatcccatataatacactacagag1740
gccgagtttagttccaaacttattctttaaacttccagcttttccatcacatcgaaactt1800
tcctacatacacaaactttcaacttttccgtcacatcgttccaatttcaatcaaacttcc1860
aattttagcgtgaactaaacacacctagagttgatgatactatacctatttaactaggct1920
accacatgttaatctttcagaacccacgatatatgataagtcaactattagcactttctg1980
gaggtactgttggttacagtcgccaggaccacttcttcgaaaaatacagcaagaaaagag2040
acgaagtaatcgattcaaacgatatgacaagtgcggcacaggttctcggcatttcttcag2100
cgcttatagcaaccgtgacatttgcagctgcattcacgttgcctggaggttacagagcag2160
atgaccacactgatggtggtacaccaacacttgctgggagctacccttttgacgcgttca2220
ttatctcgaacagcctagcattcatttgctctttgctagctaccgtgagtctgctctact2280
ctgggatacaatcgagggacatctccatccggcgcaggtactacgcgttctctatgctct2340
tgatgcagagttcaactacgagcttcactgttgccttcgccatgggaatgtacctggtgc2400
tagctcctgttactcttaatgcagcagtgtctgtctgcatcatagcatttgtttctctgc2460
tacctggaaatatggaaatcggggtatccctagctatcgcaaacacattgaggattagat2520
tgggaatttgggcagcgatgtcccaagcaaggccagttctcctgttcacgtggaaaagag2580
tttggtcctgcataattatctttggattgcctggattgatgaagattcatagaaccaaaa2640
tggcttgattggcgtgtcacctttttttctgcattgatgtactttcttaatatatccttt2700
attcggagtgttagaagtacatttacacaacatcgtgtatgacagctttgctgttacccc2760
tctgcattttaacgtagaatatacctacgatcttaggcagaagatgaaagaggggaattc2820
agagtgtgg2829
<210>12
<211>1389
<212>dna
<213>水稻(oryzasativa)
<400>12
atgtccagggcaaacagtcgacagacaaaggcacaaagctgtgaagggctgaatgctgaa60
gggatcgccggcggcccagcgtggggtgaacttgcagaggcgccgtggagggcgcagggc120
gatggaggcggattggcggagactgccgccgcactgtctgtactgttagagaagtgcaag180
aatgtcgaggtcaatgtacagcaagaagaccaacaaaggagtattcctcttctcttgtac240
ctcactagtcaaagtgacaaaaatgggagcactcctcttcactttgctgcatcgttgaaa300
actagtattgaaggtttcacatcgagattatgtgaacattttcgcccgaaacaaagccca360
acaacattgcttcttggactcaatgaatctgctatatatcagccagacaacagaggatca420
tatcctatacttgttgctgcctccaatgggatactcaaggttgtcataaccttgctcaaa480
aggtatccagattgtgccactttgcgtgacatccaaggaagaactttcttccatgttgcc540
gtggagaagaagcggcgtaatatcgttgcatatgtgtgtgaaaggccaggattttctcct600
atcttgaatatgcaggacagccatggggatactgctctacaccttgctgtcaaagccggc660
taccacatgttaatctttcagaacccacgatatatgataagtcaactattagcactttct720
ggaggtactgttggttacagtcgccaggaccacttcttcgaaaaatacagcaagaaaaga780
gacgaagtaatcgattcaaacgatatgacaagtgcggcacaggttctcggcatttcttca840
gcgcttatagcaaccgtgacatttgcagctgcattcacgttgcctggaggttacagagca900
gatgaccacactgatggtggtacaccaacacttgctgggagctacccttttgacgcgttc960
attatctcgaacagcctagcattcatttgctctttgctagctaccgtgagtctgctctac1020
tctgggatacaatcgagggacatctccatccggcgcaggtactacgcgttctctatgctc1080
ttgatgcagagttcaactacgagcttcactgttgccttcgccatgggaatgtacctggtg1140
ctagctcctgttactcttaatgcagcagtgtctgtctgcatcatagcatttgtttctctg1200
ctacctggaaatatggaaatcggggtatccctagctatcgcaaacacattgaggattaga1260
ttgggaatttgggcagcgatgtcccaagcaaggccagttctcctgttcacgtggaaaaga1320
gtttggtcctgcataattatctttggattgcctggattgatgaagattcatagaaccaaa1380
atggcttga1389
<210>13
<211>462
<212>prt
<213>水稻(oryzasativa)
<400>13
metserargalaasnserargglnthrlysalaglnsercysglugly
151015
leuasnalagluglyilealaglyglyproalatrpglygluleuala
202530
glualaprotrpargalaglnglyaspglyglyglyleualagluthr
354045
alaalaalaleuservalleuleuglulyscyslysasnvalgluval
505560
asnvalglnglngluaspglnglnargserileproleuleuleutyr
65707580
leuthrserglnserasplysasnglyserthrproleuhispheala
859095
alaserleulysthrserilegluglyphethrserargleucysglu
100105110
hispheargprolysglnserprothrthrleuleuleuglyleuasn
115120125
gluseralailetyrglnproaspasnargglysertyrproileleu
130135140
valalaalaserasnglyileleulysvalvalilethrleuleulys
145150155160
argtyrproaspcysalathrleuargaspileglnglyargthrphe
165170175
phehisvalalavalglulyslysargargasnilevalalatyrval
180185190
cysgluargproglypheserproileleuasnmetglnaspserhis
195200205
glyaspthralaleuhisleualavallysalaglytyrhismetleu
210215220
ilepheglnasnproargtyrmetileserglnleuleualaleuser
225230235240
glyglythrvalglytyrserargglnasphisphepheglulystyr
245250255
serlyslysargaspgluvalileaspserasnaspmetthrserala
260265270
alaglnvalleuglyileserseralaleuilealathrvalthrphe
275280285
alaalaalaphethrleuproglyglytyrargalaaspasphisthr
290295300
aspglyglythrprothrleualaglysertyrpropheaspalaphe
305310315320
ileileserasnserleualapheilecysserleuleualathrval
325330335
serleuleutyrserglyileglnserargaspileserileargarg
340345350
argtyrtyralaphesermetleuleumetglnserserthrthrser
355360365
phethrvalalaphealametglymettyrleuvalleualaproval
370375380
thrleuasnalaalavalservalcysileilealaphevalserleu
385390395400
leuproglyasnmetgluileglyvalserleualailealaasnthr
405410415
leuargileargleuglyiletrpalaalametserglnalaargpro
420425430
valleuleuphethrtrplysargvaltrpsercysileileilephe
435440445
glyleuproglyleumetlysilehisargthrlysmetala
450455460
<210>14
<211>4979
<212>dna
<213>水稻(oryzasativa)
<400>14
ccctgtttagaactccatcctatagatcgatcaaggtagttaagctagcttgcacagcga60
tgaggcggcgatcgaggagagaacgggacgtcggtcaccagcagcggtggaggagcttgc120
acgctggtgctgctgctccggggagtagaggccgtagcggcgagtgtcgccggcgtgggg180
gcggcggcggcgcgggtgggaggtcggggacgaacatggcggtctagggttttaattttg240
ttttggaatggggaatcgagaggttgacgacgtggggtggttggagttggagtaggaggg300
ggaacgcgaggtcgcgaaatatgggccgggccgggcccatctcctcctcctatatggatt360
ccgaatgggccttccagaatacaaatggatctggtggaaggcccagcacagcaccaaaat420
agaacacattgaccttttttttctctctttttggcaaaaacattgactttgttggagtta480
tggagtactatcatgttttgtcagattttaaattgggctcaaaatatttcaacttcgctc540
aaaaagaaaacggaacaggctataggcataggatggagcgtagtttgggccacaccgcaa600
ctcccctctttaagttagatttaggagttaaaactctactaaatgcactcttaaattaga660
ctttaatcgcacatatacgtactagcatatagtgcctagctagctactcgaggtttggtt720
tattttgggacgaatggcgtatatctagatttataatactattatgtgtcacatttcgta780
cgaggttgatttattttaaaaccatgggtgtaatatgcccagtaaagccaccacggctcg840
cccataactccatattctttatgaccggcctctgctatgataaacataatctagaggttc900
acctcaagatttggcttcatacagcatatccgggcatttgagatactctcttcggttttt960
aatagatgacactattaacttttgaacacatgtttgattattcgttttatttaaaaattt1020
tatacaaatatataagatactctctccgttttagattataagttgttttaactttggtaa1080
aggcaaattagtttaaatttgaccaagtttgtagaaaaagttaattaaattttcatctca1140
agaaaaaatattatgaaaacatatttaataaaactaatttagtgttgtaaatattactat1200
atttgtctacaaacttgatcaaacatgaagtagtttaagtttaaccaaagtcaaaataac1260
ttataatatgaaataaagggagcataaatcatgcttaaattactttgaatgataaaacaa1320
ctcacaacaaaaaaaatataattacgtaaattttcttaaataaaacgaatagtcaaacat1380
gtgacaaaataacaacgacatcatctattaaaaaaaatacttcagtacctttatataatt1440
atacaaaagttttaaattagtaaatgatcaaatgatttacaaaaaatctaacatcaacgt1500
ttattttaaaaggagtagtactctatagtagagcaagttcgatagtatagctaactgcta1560
agtctaaattatctatagttaatttaatagttaatatatatagtagttacctacaaatat1620
atacttatagtgtactgatacatggtctgatacttggggctcatagctagcgcgtgcgcg1680
cggctagcaagcacctccccttaagactttaactacatctattaatttattagttaatat1740
ttaacactaatgatactagtagtaattagaaaagattctgaagatttttaagagattttt1800
ttactaacagattgaaaatttaattttttaagttagatttgaaaacttttgatttaagtt1860
tcaaattttaaagtcaaattttgaaaactttcaaaatccagatttgaaaaatttgaaaac1920
ttttaacttgagatttaaaaattttcaactcgagatttaaaaactttcaacttaagattc1980
gaaaactttcaagtctagatttgaaaattttgaaaactttcaacttgagatttgaaaatt2040
ttcaactcgagatttgaaaactttcaactcgatattcgaaaactttcaagtccagatttg2100
aaaattttgaaaactttcaactaagattcgaaaattttcactcgagaattgaaattttca2160
aggctagatttgaaaattttcaagtcaatatttaaaaactttcaactcaaaacatttgaa2220
aaaagcacatttgctattggctaggaaatagtggggatggtggatgcatcgagtttgttg2280
ctagagtaaatatagtatgagagagttattagcttttcttggtcttagtgtacctatgaa2340
atatatagatcaatttggaatggagggagtacttaggtgcagaaggaaatgatataactt2400
ttccccccagattttctgttacttcatgttttaggtataccaatatttttgtgtacttgg2460
ttgatctgtgcactagcatataacaattatccggccatcaaaatggatgaattttccatt2520
tcatcatgtgaaagacataataactggaaccttatcaaagacatatacttattagttatt2580
gcatgaggtagttaacagaattgattgctttcattccccagctttcactggtgatgaact2640
tcaacttcttgtatataatctttttgatgctgcagttcatgtgatgtactgatttccttc2700
tggtcattgaattactatcatattgtattgttttctttaacggggaacagaaagggatac2760
cctttctgaaatcatgctgctgttattgaaccctgtgttccttattacaggtaaaagaca2820
gcatgtcatgaggactttccaaggtcataaccccaacactatgaaagttcattctcgcaa2880
gtctcaaccttcaaaaaagaagcctcgcgagttctatgacactgttgaggtacacattat2940
tgatgatgacagtgatggtgatgcgaatatccacaaggattactcaatggaagacacatc3000
aaagcaccttgtcatgtacaatcctgagatcacgtatgacaaacaaggggaagttgaagt3060
tactgagcctatagataactatacttcgctgaaccaaagatacatgaagccaagacacgg3120
atataacactgttttaccatctatcggtgcttacaccgtgcaatgtgctcactgcttcaa3180
atggaggatcatcccaacaaaagagaaatatgaagagatacgtgagaatatttgtcagga3240
tgtgtttgtatgtgaaagagctcgtgaatggaaccgtgttatatcttgcgttgatcctga3300
agatatatcacaggatggaagtagggtgtgggcgattgacaaggcaagcatttcccagac3360
tcctcctggctgggatagagaggtcagaatcagaggagagggttgcagcaagtttgcaga3420
tgtgtatgtcactgtttcctttcctggatttgcagtagtacgtgatatttggtaccatat3480
ttttggccgtttccatattcggatatcaaacttctttctttatgtgcctaatagcatctt3540
catgttaatcgtggcacactggtcctgtttatttatgccttgactagctgctaactgcta3600
agattagttaacactttctaaataattagtgcgatctcaagtttattcatactagtacca3660
cctatgatcatgaaacttctacagttttcagttaatactttgtaacaccatagcaaatat3720
ctcactaattccttatgatatgcctgcttgaataatgcaaacagccattattatgtttgc3780
cagtttcctcaggaactttttcattatatgatatttcgaacagcaacattgcttgtagga3840
cacacaagatactcgaggttgacatatgtgaaatacttttcttttttccttcaggtacta3900
cacttctccaactgggaagaagttgaggtcattggtagaagttgggaggtgggattcatc3960
catttgtgctctttttgttgcatattttgtgtggctagtgtattacttggggtacctagt4020
tctagttggtctggtacaacaaaacgaagctctggaaaagctctggaaagttcttgaaat4080
atttcccaggaaatgatgtaattattatgtccatattctaggtacttggcagagaaccca4140
cactacattaggcaaggggttaacttaacccagttctctttcgccacccccaagcctctg4200
caagaagactatgtcaggaagcatacatatgcagcaactcctgaattaccagagcttctt4260
gagactgcgcaaggtgtgttatcagtcctaggtttgttacatctattctacttttttttc4320
atttttaacttcttgttccccacattgttggaaaggaggcgtgcttcaaacaaggtaagt4380
tgctgcgcaaatggcacagttggaggtatctttgcaatgcaagcatgtctagttattttc4440
agtcaaatgagctgtgagaagtcatgtactcatgatgcccttgcattttgctactttaca4500
tcctttcttgatcttagcaccatctaaatatcatttagtgttgtgatattaactgaatat4560
attttcccccagtttcatcccatagctatatcctgtcaaaactacctgttcttgggagct4620
tttgctgatatatttgttattgagtatacattgttatttttcagtggatccactctgctg4680
ggcagcaccccctacacgcagcgaactgcttggtgagcttggtgcttcaacctcaagatc4740
tgttgatgtcagccagtctgaggtgtcataccctgtagacttgcaccagcccgaggagtc4800
tgcgccactacctccacgcacaaagaagaaaactatgaaacgagggcgagtgtcagcaac4860
ggggtctcaatcgactccacctggttcatccaaagatcagtctggaggatgcgtaagcga4920
cgtcgagtttgtttcgctgtgaactctggatgcagtgcctttgtctctgtcagtctgcc4979
<210>15
<211>1233
<212>dna
<213>水稻(oryzasativa)
<400>15
atgaggcggcgatcgaggagagaacgggacgtcggtcaccagcagcggtggaggagcttg60
cacgctggtgctgctgctccggggagtagaggccgtagcggcgagtgtcgccggcgtggg120
ggcggcggcggcgcgggtaaaagacagcatgtcatgaggactttccaaggtcataacccc180
aacactatgaaagttcattctcgcaagtctcaaccttcaaaaaagaagcctcgcgagttc240
tatgacactgttgaggtacacattattgatgatgacagtgatggtgatgcgaatatccac300
aaggattactcaatggaagacacatcaaagcaccttgtcatgtacaatcctgagatcacg360
tatgacaaacaaggggaagttgaagttactgagcctatagataactatacttcgctgaac420
caaagatacatgaagccaagacacggatataacactgttttaccatctatcggtgcttac480
accgtgcaatgtgctcactgcttcaaatggaggatcatcccaacaaaagagaaatatgaa540
gagatacgtgagaatatttgtcaggatgtgtttgtatgtgaaagagctcgtgaatggaac600
cgtgttatatcttgcgttgatcctgaagatatatcacaggatggaagtagggtgtgggcg660
attgacaaggcaagcatttcccagactcctcctggctgggatagagaggtcagaatcaga720
ggagagggttgcagcaagtttgcagatgtgtactacacttctccaactgggaagaagttg780
aggtcattggtagaagttgggaggtacttggcagagaacccacactacattaggcaaggg840
gttaacttaacccagttctctttcgccacccccaagcctctgcaagaagactatgtcagg900
aagcatacatatgcagcaactcctgaattaccagagcttcttgagactgcgcaagtggat960
ccactctgctgggcagcaccccctacacgcagcgaactgcttggtgagcttggtgcttca1020
acctcaagatctgttgatgtcagccagtctgaggtgtcataccctgtagacttgcaccag1080
cccgaggagtctgcgccactacctccacgcacaaagaagaaaactatgaaacgagggcga1140
gtgtcagcaacggggtctcaatcgactccacctggttcatccaaagatcagtctggagga1200
tgcgtaagcgacgtcgagtttgtttcgctgtga1233
<210>16
<211>410
<212>prt
<213>水稻(oryzasativa)
<400>16
metargargargserargarggluargaspvalglyhisglnglnarg
151015
trpargserleuhisalaglyalaalaalaproglyserargglyarg
202530
serglyglucysargargargglyglyglyglyglyalaglylysarg
354045
glnhisvalmetargthrpheglnglyhisasnproasnthrmetlys
505560
valhisserarglysserglnproserlyslyslysproarggluphe
65707580
tyraspthrvalgluvalhisileileaspaspaspseraspglyasp
859095
alaasnilehislysasptyrsermetgluaspthrserlyshisleu
100105110
valmettyrasnprogluilethrtyrasplysglnglygluvalglu
115120125
valthrgluproileaspasntyrthrserleuasnglnargtyrmet
130135140
lysproarghisglytyrasnthrvalleuproserileglyalatyr
145150155160
thrvalglncysalahiscysphelystrpargileileprothrlys
165170175
glulystyrglugluilearggluasnilecysglnaspvalpheval
180185190
cysgluargalaargglutrpasnargvalilesercysvalasppro
195200205
gluaspileserglnaspglyserargvaltrpalaileasplysala
210215220
serileserglnthrproproglytrpasparggluvalargilearg
225230235240
glygluglycysserlysphealaaspvaltyrtyrthrserprothr
245250255
glylyslysleuargserleuvalgluvalglyargtyrleualaglu
260265270
asnprohistyrileargglnglyvalasnleuthrglnpheserphe
275280285
alathrprolysproleuglngluasptyrvalarglyshisthrtyr
290295300
alaalathrprogluleuprogluleuleugluthralaglnvalasp
305310315320
proleucystrpalaalaproprothrargsergluleuleuglyglu
325330335
leuglyalaserthrserargservalaspvalserglnsergluval
340345350
sertyrprovalaspleuhisglnproglugluseralaproleupro
355360365
proargthrlyslyslysthrmetlysargglyargvalseralathr
370375380
glyserglnserthrproproglyserserlysaspglnserglygly
385390395400
cysvalseraspvalgluphevalserleu
405410
<210>17
<211>2044
<212>dna
<213>水稻(oryzasativa)
<400>17
gaagtagtactaaaatggatctcgcacactttctctttccgtatagccaagctgtgctgc60
aaagatcactgatcagagtgaacctggtacattttcacgggctggttcattttgtaggac120
tgtctgcttggcttggcctgtgcaatccagtgatcagcagtactgtactccactatagta180
atcagagcatgtgcgccagtttctcccttatgttgctactctaatacatttgggattaac240
taattatggtcactaatcagctattgacagtagcaattggccggcttggccaaatatgca300
agtgagttgggctgttactttgccttctttttcacaaaggaaaatgttactgtcaagata360
tactgggtaagatgaatgattaaaagttaaggtagtacattgtatgcgctctcttttttt420
ttaaccggttctggcaatgagaccactatgttctttttgctatgtcaaaacacctaattt480
aaatcatattagttttgaatcctcatctttttgttataggaattcaaaatctgtaggagt540
ctcctgtgatttttgtccacagtttttgtcagagttagaggtagagacggaactctttca600
cttcgtgtcatgattcctacgcgacagcgagaccgtatcagatgctcccacaacattacg660
caatagtgtcctgcaggtcttgactctcgagatttagattttgggatgtgcctatatgct720
ccgaaaaaaaaaacttgcacgaaagttaaagataatcagcggctgtgattagaagcgaga780
ttagcacttaattatcattagcgaggggtaatttcgtcaagattttcgagcccatatcca840
gcccagtccatctgaagcgatccccacccccaccggtccaccacggcgccgctcctcaac900
cgcaccgccacgccgcctccactccctcctcccctctcctctccgccgctgctccgctca960
atgccgccttgccgccccagcgctccgctctcttcttcccctaggctgaccgctcagccc1020
agcgccgcccggcttgctgcctctccatcctcttcgccccctcgtctcgccgcttccccg1080
cccctgcgccgtgcagcccggcgcacccccttcatccatctcacatgcgtagtttgcttg1140
atgagtggtttgctccatgttagaagcttggtattgcagcagaacatggctattttgtca1200
ggtaaacattttgagtacctttttttcttcttaaatctgtttttatttgcattttctcca1260
cacactattatgcaattgtacagttgaattattcaggagtataatgaactgtttatgttc1320
tctcagatggaacaaagcagctgaatgggaatcaagctatccaaaccatgattttgagtg1380
gaagcacatcgctgaaccgatcatgcaagtttacaccgaaacaattgatgggtcctccat1440
agagccaaaggaaagtgctctattatggcattatctggatgcagaccatgacttcggttc1500
ctgccaagcaaaggaactactgggtcatcttgaaagggtgctatcaaatgaacctgttgt1560
tgtgaagtgtggtcattacattgtagaagtcaaaccacaggtctataccatttgaattta1620
ttactacattttttcactttcaactcattatgtccagctcacactaaatgaaatacatta1680
cttgtgcagggagttagcaagggtcttgttgtggacaaggtaattcacagattgatgaac1740
aacgggaagacgctggatttcgtagtgtgcatcggtaacgatcgatctgatgaggacatg1800
ttcaaaagcatcgacagcatgacctccagctctgcattccctgcagttccagaggtcttt1860
gcctgttcagttggtcagaaacccagcaaagcaaaatactatgtcgacaatgttggtgaa1920
gtagtaagattgctcaagaatgtagccggcatttcgtcgcaccgggaggccgtcagccat1980
ggacgtgtgatcttcagggatgtgatcgattatgtggactgaaccacctaatctttgtaa2040
catc2044
<210>18
<211>819
<212>dna
<213>水稻(oryzasativa)
<400>18
atggatctcgcacactttctctttccgtatagccaagctgtgctgcaaagatcactgatc60
agagtgaacctgctattgacagtagcaattggccggcttggccaaatatgcaagaattca120
aaatctgtaggagtctcctgtgatttttgtccacagtttttgtcagagttagagaagctt180
ggtattgcagcagaacatggctattttgtcagatggaacaaagcagctgaatgggaatca240
agctatccaaaccatgattttgagtggaagcacatcgctgaaccgatcatgcaagtttac300
accgaaacaattgatgggtcctccatagagccaaaggaaagtgctctattatggcattat360
ctggatgcagaccatgacttcggttcctgccaagcaaaggaactactgggtcatcttgaa420
agggtgctatcaaatgaacctgttgttgtgaagtgtggtcattacattgtagaagtcaaa480
ccacagggagttagcaagggtcttgttgtggacaaggtaattcacagattgatgaacaac540
gggaagacgctggatttcgtagtgtgcatcggtaacgatcgatctgatgaggacatgttc600
aaaagcatcgacagcatgacctccagctctgcattccctgcagttccagaggtctttgcc660
tgttcagttggtcagaaacccagcaaagcaaaatactatgtcgacaatgttggtgaagta720
gtaagattgctcaagaatgtagccggcatttcgtcgcaccgggaggccgtcagccatgga780
cgtgtgatcttcagggatgtgatcgattatgtggactga819
<210>19
<211>272
<212>prt
<213>水稻(oryzasativa)
<400>19
metaspleualahispheleupheprotyrserglnalavalleugln
151015
argserleuileargvalasnleuleuleuthrvalalaileglyarg
202530
leuglyglnilecyslysasnserlysservalglyvalsercysasp
354045
phecysproglnpheleusergluleuglulysleuglyilealaala
505560
gluhisglytyrphevalargtrpasnlysalaalaglutrpgluser
65707580
sertyrproasnhisasppheglutrplyshisilealagluproile
859095
metglnvaltyrthrgluthrileaspglyserserilegluprolys
100105110
gluseralaleuleutrphistyrleuaspalaasphisaspphegly
115120125
sercysglnalalysgluleuleuglyhisleugluargvalleuser
130135140
asngluprovalvalvallyscysglyhistyrilevalgluvallys
145150155160
proglnglyvalserlysglyleuvalvalasplysvalilehisarg
165170175
leumetasnasnglylysthrleuaspphevalvalcysileglyasn
180185190
aspargseraspgluaspmetphelysserileaspsermetthrser
195200205
serseralapheproalavalprogluvalphealacysservalgly
210215220
glnlysproserlysalalystyrtyrvalaspasnvalglygluval
225230235240
valargleuleulysasnvalalaglyileserserhisarggluala
245250255
valserhisglyargvalilepheargaspvalileasptyrvalasp
260265270
<210>20
<211>1481
<212>dna
<213>水稻(oryzasativa)
<400>20
catgaactcgctccaatccttcctcgccctcaacccgcccgccgccgccgccgccctcgg60
cggcgcgcgcctccgcccgtcgcgcgtcaccgcgtgcctcgccacgcccacgcccacgcc120
gccgcccccgacctccgcgcccctcgcccccgccgccgcggccgcgcgccgcgagctgtc180
cgccgcgagccgcgccgtggtggaggacgaggcgaggtacatcgtcgggacgtacaaccg240
ctcccgcgtcgtcctcgtcgccggccgcggctgcaagctgtacgacgccgacgggcgcga300
gtacctcgacatggccgccggcatcgccgtcaacgcgctcggccacgccgaccccgactg360
ggtcgccgccgtctccgcccaggccgccaccctcgtccacgccagcaacgtccagtacac420
cgtcccccaggtggcgctcgccaagcgcctcgtggaggcctccttcgcggaccgcgtctt480
cttcgccaacaccggcacggaggccaacgaggcggccatcaagttcgccaggaagttcca540
gagggtggcgcgccccgacggcgacgcgcccaccgagttcatgtcgttcaccaactgctt600
ccatggcaggaccatggggtcgctcgcgctcaccagcaaggtccaataccgggagccatt660
cgcgccggtgatgcccggcgcgacgttcgccgagtacgggaacctagaggaggccaagaa720
ggtgatacagtctggcaaaattgctgccgtgttcgtcgagcccgtgcagggcgagggtgg780
gatccatagtgccaccaaggagttcttgcaggggctgcgggatgcctgcgatgaggctgg840
agctctcttggtctttgatgaggtgcaatgtggtctgggacgcacaggttacctctgggc900
gtatgaagcctatggagtactacctgacattatgaccttggcaaagccattggccggtgg960
tctccccattggtgtagtcttggtcaccgagaaggttgcttcagcaataaacttcggcga1020
ccacggtaccacattcgggggaggccctcttgtttgccaagctgcattgaccacattgga1080
taagatccagaaacctggcttcctagcagaggtggccaagaaaggagagaacttcaagca1140
gcttctcagtaccaagctgagcggaaacgcccatgtgaaagagatccgggggatcggtct1200
cattgtcggcatcgagctcgatgtcccagcgggtcctttggttgatgcgtgcttggatcg1260
cggtgtcatagtgctgacagctggtaaaggcaatgttgtgaggctggtgccgccactcat1320
catctcggaaaaggagcttgagcaggctgcagaggtgataagggactgtctacctgcact1380
tgatgcctctacctcttaatgtaagcaacatccttagcaatggaggctagaatgatcttc1440
cccttttgggaagacccttagcaatggatgttagaatgaac1481
<210>21
<211>1398
<212>dna
<213>水稻(oryzasativa)
<400>21
atgaactcgctccaatccttcctcgccctcaacccgcccgccgccgccgccgccctcggc60
ggcgcgcgcctccgcccgtcgcgcgtcaccgcgtgcctcgccacgcccacgcccacgccg120
ccgcccccgacctccgcgcccctcgcccccgccgccgcggccgcgcgccgcgagctgtcc180
gccgcgagccgcgccgtggtggaggacgaggcgaggtacatcgtcgggacgtacaaccgc240
tcccgcgtcgtcctcgtcgccggccgcggctgcaagctgtacgacgccgacgggcgcgag300
tacctcgacatggccgccggcatcgccgtcaacgcgctcggccacgccgaccccgactgg360
gtcgccgccgtctccgcccaggccgccaccctcgtccacgccagcaacgtccagtacacc420
gtcccccaggtggcgctcgccaagcgcctcgtggaggcctccttcgcggaccgcgtcttc480
ttcgccaacaccggcacggaggccaacgaggcggccatcaagttcgccaggaagttccag540
agggtggcgcgccccgacggcgacgcgcccaccgagttcatgtcgttcaccaactgcttc600
catggcaggaccatggggtcgctcgcgctcaccagcaaggtccaataccgggagccattc660
gcgccggtgatgcccggcgcgacgttcgccgagtacgggaacctagaggaggccaagaag720
gtgatacagtctggcaaaattgctgccgtgttcgtcgagcccgtgcagggcgagggtggg780
atccatagtgccaccaaggagttcttgcaggggctgcgggatgcctgcgatgaggctgga840
gctctcttggtctttgatgaggtgcaatgtggtctgggacgcacaggttacctctgggcg900
tatgaagcctatggagtactacctgacattatgaccttggcaaagccattggccggtggt960
ctccccattggtgtagtcttggtcaccgagaaggttgcttcagcaataaacttcggcgac1020
cacggtaccacattcgggggaggccctcttgtttgccaagctgcattgaccacattggat1080
aagatccagaaacctggcttcctagcagaggtggccaagaaaggagagaacttcaagcag1140
cttctcagtaccaagctgagcggaaacgcccatgtgaaagagatccgggggatcggtctc1200
attgtcggcatcgagctcgatgtcccagcgggtcctttggttgatgcgtgcttggatcgc1260
ggtgtcatagtgctgacagctggtaaaggcaatgttgtgaggctggtgccgccactcatc1320
atctcggaaaaggagcttgagcaggctgcagaggtgataagggactgtctacctgcactt1380
gatgcctctacctcttaa1398
<210>22
<211>465
<212>prt
<213>水稻(oryzasativa)
<400>22
metasnserleuglnserpheleualaleuasnproproalaalaala
151015
alaalaleuglyglyalaargleuargproserargvalthralacys
202530
leualathrprothrprothrproproproprothrseralaproleu
354045
alaproalaalaalaalaalaargarggluleuseralaalaserarg
505560
alavalvalgluaspglualaargtyrilevalglythrtyrasnarg
65707580
serargvalvalleuvalalaglyargglycyslysleutyraspala
859095
aspglyargglutyrleuaspmetalaalaglyilealavalasnala
100105110
leuglyhisalaaspproasptrpvalalaalavalseralaglnala
115120125
alathrleuvalhisalaserasnvalglntyrthrvalproglnval
130135140
alaleualalysargleuvalglualaserphealaaspargvalphe
145150155160
phealaasnthrglythrglualaasnglualaalailelyspheala
165170175
arglyspheglnargvalalaargproaspglyaspalaprothrglu
180185190
phemetserphethrasncysphehisglyargthrmetglyserleu
195200205
alaleuthrserlysvalglntyrarggluprophealaprovalmet
210215220
proglyalathrphealaglutyrglyasnleugluglualalyslys
225230235240
valileglnserglylysilealaalavalphevalgluprovalgln
245250255
glygluglyglyilehisseralathrlysglupheleuglnglyleu
260265270
argaspalacysaspglualaglyalaleuleuvalpheaspgluval
275280285
glncysglyleuglyargthrglytyrleutrpalatyrglualatyr
290295300
glyvalleuproaspilemetthrleualalysproleualaglygly
305310315320
leuproileglyvalvalleuvalthrglulysvalalaseralaile
325330335
asnpheglyasphisglythrthrpheglyglyglyproleuvalcys
340345350
glnalaalaleuthrthrleuasplysileglnlysproglypheleu
355360365
alagluvalalalyslysglygluasnphelysglnleuleuserthr
370375380
lysleuserglyasnalahisvallysgluileargglyileglyleu
385390395400
ilevalglyilegluleuaspvalproalaglyproleuvalaspala
405410415
cysleuaspargglyvalilevalleuthralaglylysglyasnval
420425430
valargleuvalproproleuileileserglulysgluleuglugln
435440445
alaalagluvalileargaspcysleuproalaleuaspalaserthr
450455460
ser
465
<210>23
<211>413
<212>dna
<213>水稻(oryzasativa)
<400>23
gcgaaaatcttatcattgtagttctttccatcgcagagggtgggctagggcgatccgggg60
ggcgcgaccatggcttacgtcgagagaggtgttgtgaaagacaaacggacaatttggcgg120
ctaagcataatttctgacttcttcagggctattgtgaacttcatcagaatgttcttcctt180
acaatgttctcgattgaaaaaacagacagctataggaaagggtatggctctggtaagaaa240
tgggatggtgggcctggtgggggaggtcctggtggtggcccttatgggggcggccgcggc300
ggtggaggcggcctccgaggtcctcgcacactatctgacatccgatctaacgatcagaat360
tctctccctgcttgtggatcctgctgcggttaacctgtacatctgtcattgtg413
<210>24
<211>324
<212>dna
<213>水稻(oryzasativa)
<400>24
atggcttacgtcgagagaggtgttgtgaaagacaaacggacaatttggcggctaagcata60
atttctgacttcttcagggctattgtgaacttcatcagaatgttcttccttacaatgttc120
tcgattgaaaaaacagacagctataggaaagggtatggctctggtaagaaatgggatggt180
gggcctggtgggggaggtcctggtggtggcccttatgggggcggccgcggcggtggaggc240
ggcctccgaggtcctcgcacactatctgacatccgatctaacgatcagaattctctccct300
gcttgtggatcctgctgcggttaa324
<210>25
<211>107
<212>prt
<213>水稻(oryzasativa)
<400>25
metalatyrvalgluargglyvalvallysasplysargthriletrp
151015
argleuserileileseraspphepheargalailevalasnpheile
202530
argmetphepheleuthrmetpheserileglulysthraspsertyr
354045
arglysglytyrglyserglylyslystrpaspglyglyproglygly
505560
glyglyproglyglyglyprotyrglyglyglyargglyglyglygly
65707580
glyleuargglyproargthrleuseraspileargserasnaspgln
859095
asnserleuproalacysglysercyscysgly
100105
<210>26
<211>23
<212>dna
<213>人工序列
<220>
<223>克隆osicdh1基因cdna的正向引物
<400>26
gactccgacgaccagaagctacc23
<210>27
<211>21
<212>dna
<213>人工序列
<220>
<223>克隆osicdh1基因cdna的反向引物
<400>27
ctaacatgtcccctgccgttg21
<210>28
<211>26
<212>dna
<213>人工序列
<220>
<223>克隆osmtn3l基因gdna的正向引物
<400>28
cactgacatgtggccttccttcttcc26
<210>29
<211>27
<212>dna
<213>人工序列
<220>
<223>克隆osmtn3l基因gdna的反向引物
<400>29
ctaagacaaaattaggtgcaggatggg27
<210>30
<211>29
<212>dna
<213>人工序列
<220>
<223>克隆osdn-dtp6基因gdna的正向引物
<400>30
gctgcaaaggaggaagagaaagagtgttg29
<210>31
<211>27
<212>dna
<213>人工序列
<220>
<223>克隆osdn-dtp6基因gdna的反向引物
<400>31
gagatcgcaacaaacccatacccaaac27
<210>32
<211>27
<212>dna
<213>人工序列
<220>
<223>克隆osankl1基因gdna的正向引物
<400>32
caagtacctgtaaattgaaacctgcag27
<210>33
<211>24
<212>dna
<213>人工序列
<220>
<223>克隆osankl1基因gdna的反向引物
<400>33
ccacactctgaattcccctctttc24
<210>34
<211>29
<212>dna
<213>人工序列
<220>
<223>克隆osmbd2基因gdna的正向引物
<400>34
ccctgtttagaactccatcctatagatcg29
<210>35
<211>25
<212>dna
<213>人工序列
<220>
<223>克隆osmbd2基因gdna的反向引物
<400>35
ggcagactgacagagacaaaggcac25
<210>36
<211>29
<212>dna
<213>人工序列
<220>
<223>克隆ostp1基因gdna的正向引物
<400>36
gaagtagtactaaaatggatctcgcacac29
<210>37
<211>28
<212>dna
<213>人工序列
<220>
<223>克隆ostp1基因gdna的反向引物
<400>37
gatgttacaaagattaggtggttcagtc28
<210>38
<211>25
<212>dna
<213>人工序列
<220>
<223>克隆osacoat1基因cdna的正向引物
<400>38
catgaactcgctccaatccttcctc25
<210>39
<211>27
<212>dna
<213>人工序列
<220>
<223>克隆osacoat1基因cdna的反向引物
<400>39
gttcattctaacatccattgctaaggg27
<210>40
<211>29
<212>dna
<213>人工序列
<220>
<223>克隆osdn-dtp7基因cdna的正向引物
<400>40
gcgaaaatcttatcattgtagttctttcc29
<210>41
<211>26
<212>dna
<213>人工序列
<220>
<223>克隆osdn-dtp7基因cdna的反向引物
<400>41
cacaatgacagatgtacaggttaacc26
<210>42
<211>20
<212>dna
<213>人工序列
<220>
<223>osicdh1基因实时rt-pcr分析的正向引物
<400>42
caaaaaggaggcgaaactag20
<210>43
<211>23
<212>dna
<213>人工序列
<220>
<223>osicdh1基因实时rt-pcr分析的反向引物
<400>43
catgtacaagaagagctagatcc23
<210>44
<211>18
<212>dna
<213>人工序列
<220>
<223>osmtn3l基因实时rt-pct分析的正向引物
<400>44
tgaaggccaaatcgaccg18
<210>45
<211>20
<212>dna
<213>人工序列
<220>
<223>osmtn3l基因实时rt-pct分析的反向引物
<400>45
gaagtccataccacaggcag20
<210>46
<211>19
<212>dna
<213>人工序列
<220>
<223>osdn-dtp6基因实时rt-pcr分析的正向引物
<400>46
ttgggacgcttcgagattg19
<210>47
<211>25
<212>dna
<213>人工序列
<220>
<223>osdn-dtp6基因实时rt-pcr分析的反向引物
<400>47
atctggtccggagtaagataatttc25
<210>48
<211>22
<212>dna
<213>人工序列
<220>
<223>osankl1基因实时rt-pcr分析的正向引物
<400>48
ggtatccctagctatcgcaaac22
<210>49
<211>20
<212>dna
<213>人工序列
<220>
<223>osankl1基因实时rt-pcr分析的反向引物
<400>49
ttccacgtgaacaggagaac20
<210>50
<211>21
<212>dna
<213>人工序列
<220>
<223>osmbd2基因实时rt-pcr分析的正向引物
<400>50
cataccctgtagacttgcacc21
<210>51
<211>21
<212>dna
<213>人工序列
<220>
<223>osmbd2基因实时rt-pcr分析的反向引物
<400>51
cgccctcgtttcatagttttc21
<210>52
<211>20
<212>dna
<213>人工序列
<220>
<223>ostp1基因实时rt-pcr分析的正向引物
<400>52
tgcagttccagaggtctttg20
<210>53
<211>18
<212>dna
<213>人工序列
<220>
<223>ostp1基因实时rt-pcr分析的反向引物
<400>53
atcacacgtccatggctg18
<210>54
<211>20
<212>dna
<213>人工序列
<220>
<223>osacoat1基因实时rt-pcr分析正向引物
<400>54
acgcccatgtgaaagagatc20
<210>55
<211>20
<212>dna
<213>人工序列
<220>
<223>osacoat1基因实时rt-pcr分析反向引物
<400>55
caagcacgcatcaaccaaag20
<210>56
<211>22
<212>dna
<213>人工序列
<220>
<223>osdn-dtp7基因实时rt-pcr分析的正向引物
<400>56
gaaagggtatggctctggtaag22
<210>57
<211>21
<212>dna
<213>人工序列
<220>
<223>osdn-dtp7基因实时rt-pcr分析的反向引物
<400>57
gatgtcagatagtgtgcgagg21
- 还没有人留言评论。精彩留言会获得点赞!