用于基因内植物转化的构建体和载体的制作方法
本发明涉及植物转化。具体而言,本发明涉及但不限于用于基因内植物转化的基因构建体,以及使用该构建体的方法。
背景技术:
与常规育种方法相比,用于产生新作物品种的基因技术提供了明显的优势,例如节省时间和成本、消除遗传累赘(geneticdrag)、以及避免作物与其具有部分生育力的野生亲缘植物之间的杂交。然而,通过基因技术进行作物遗传改良的主要障碍是缺乏公众对转基因品种的接受。这至少部分是由于认为遗传物质在属于分类学上遥远组的有机体之间转移是“不自然的”。
通常认为,相比于转基因作物,公众更接受使用涉及遗传物质在相同植物品种或其性相容亲缘植物之间转移的基因技术产生的植物。这些过程可以被认为是基因重组,其中一部分植物基因组(或其性相容亲缘植物的基因组)被部分重排和重组以产生遗传多样性。基因重组在自然界中是一个重要的过程,因此来自具有不同基因库的群体的个体能够适应不断变化的环境。基因重组的模拟可以利用分子生物学工具通过两种目前正在探讨的称为“顺式基因(cisgenic)”和“基因内(intragenic)”的方法来实现。
顺式基因方法是相对保守的,只允许基因的未修饰基因组版本转移,包括来自相同植物或其性相容亲缘植物的内含子和调节元件。相比之下,基因内方法通过转移包含衍生自植物基因组内多个区域和/或衍生自相同物种或其性相容亲缘植物的多个单个植物的序列的核酸来拓宽机会。
技术实现要素:
本发明广泛涉及使用植物衍生的核苷酸序列转化植物。
本发明的优选目的是提供其至少一个片段可插入至植物的遗传物质中的重组基因构建体,其中所述基因构建体的至少一个片段由衍生自一株或多株植物的一条或多条核苷酸序列组成。本发明还广泛涉及所述基因构建体用于产生遗传改良的植物的用途。
在第一方面,本发明提供了包含能插入至植物的遗传物质中的一个或多个核酸片段的重组基因构建体,其中所述一个或多个核酸片段包含衍生自一株或多株植物的长度为至少15个核苷酸或优选长度为至少20个核苷酸的多条核苷酸序列、由所述多条核苷酸序列组成、或基本上由所述多条核苷酸序列组成。
优选地,衍生自一株或多株植物的所述核苷酸序列衍生自一株植物。适当地,在所述核苷酸序列衍生自一个以上植物的实施方案中,所述植物是互交可育的和/或为相同物种的。
优选地,基因构建体的可插入至植物的遗传物质中的一个或多个核酸片段的总长度为至少100个碱基对、至少500个碱基对、至少1000个碱基对、至少2000个碱基对、至少2500个碱基对或至少3000个碱基对。
优选地,该方面的基因构建体的可插入至植物的遗传物质中的一个或多个核酸片段包含用于表达的一条或多条核苷酸序列。优选地,所述一条或多条核苷酸序列适合在植物中表达。
优选地,用于在植物中表达的所述一条或多条核苷酸序列适于在植物中表达,以改变或修饰植物的性状。
在某些优选的实施方案中,用于在植物中表达的一条或多条所述核苷酸序列包含蛋白编码核苷酸序列。在一个优选的实施方案中,所述蛋白编码核苷酸序列包含seqidnos:38-46、76、78或98所示的核苷酸序列,或其片段或变体。
在某些优选的实施方案中,适合在植物中表达的一条或多条所述核苷酸序列是非蛋白编码核苷酸序列。优选地,所述非蛋白编码核苷酸序列包含一条或多条小rna核苷酸序列。在一个优选的实施方案中,所述包含一条或多条小rna核苷酸序列的用于表达的所述核苷酸序列包含seqidnos:12-26、64-66、80-81、83-92、94或96-101所示的核苷酸序列,或其片段或变体。
在优选的实施方案中,用于在植物中表达的所述一条或多条核苷酸序列包含一条或多条选择标记核苷酸序列。在一个优选的实施方案中,所述选择标记核苷酸序列包含编码seqidnos:38-46所示氨基酸序列的核苷酸序列,或seqidno:119所示的核苷酸序列,或其片段或变体。
优选地,该方面的基因构建体的可插入至植物的遗传物质中的一个或多个核酸片段包含一条或多条调节核苷酸序列。
适当地,基因构建体的可插入至植物的遗传物质中的核酸片段的可表达核苷酸序列与一条或多条所述调节核苷酸序列可操作地连接。
优选地,所述调节核苷酸序列包含一条或多条启动子序列。在一个优选的实施方案中,所述启动子核苷酸序列包含seqidnos:4-7、67、73、74、76、78或98所示的核苷酸序列,或其片段或变体。
优选地,所述调节序列包含一条或多条终止子序列。在一个优选的实施方案中,所述终止子核苷酸序列包含seqidnos:8-11、106、108、111或112所示的核苷酸序列,或其片段或变体。
适当地,该方面的重组基因构建体可包含可插入至植物的遗传物质中的一个或多个片段的侧翼序列,或围绕可插入至植物的遗传物质中的一个或多个片段的侧翼序列。在一些优选的实施方案中,所述侧翼序列或其一部分衍生自一株或多株植物。
在一些优选的实施方案中,所述侧翼序列包含限制性消化位点。在某些特别优选的实施方案中,一条或多条侧翼序列包含seqidnos:102、103、109、110、115、116、117、118、120或121所示的核苷酸序列,或其片段或变体。
在某些特别优选的实施方案中,该方面的重组基因构建体包含seqidnos:1-35、49、51-56、66-68、71-92或94-101所示的核苷酸序列,或编码seqidnos:38-46所示氨基酸序列的核酸,或其片段或变体。
在第一方面的某些优选实施方案中,重组基因构建体的侧翼序列包含边界序列。在优选的此类实施方案中,重组基因构建体包含:
第一边界核苷酸序列;
第二边界核苷酸序列;和
位于所述第一边界核苷酸序列和所述第二边界核苷酸序列之间的一条或多条额外核苷酸序列(additionalnucleotidesequence),
其中所述额外核苷酸序列和与所述额外核苷酸序列相邻的至少一部分所述第一边界核苷酸序列衍生自一株或多株植物物种。
在这些实施方案中,任选地,与所述一条或多条额外核苷酸序列相邻的至少一部分所述第二边界核苷酸序列可以衍生自一株或多株植物。适当地,所述一个或个种植物是衍生所述额外核苷酸序列和至少一部分所述第一边界序列的相同植物。
在第一方面的这些优选实施方案中,优选地,由衍生自一株或多株植物的长度为至少15个核苷酸或优选长度为至少20个核苷酸的多条核苷酸序列组成的可插入至植物的遗传物质中的所述一个或多个核酸片段,由以下组成:
(i)衍生自一株或多株植物的至少一部分所述第一边界序列;
(ii)衍生自一株或多株植物的所述一条或多条额外核苷酸序列;和任选地
(iii)衍生自一株或多株植物的至少一部分所述第二边界序列。
在某些实施方案中,(i)和(ii)的额外核苷酸序列衍生自植物的长度为至少15个或优选至少20个核苷酸的相同核苷酸序列。
在某些实施方案中,(iii)和(ii)的额外核苷酸序列衍生自植物的长度为至少15个或优选至少20个核苷酸的相同核苷酸序列。
优选地,这些实施方案的基因构建体的第一边界核苷酸序列属于农杆菌(agrobacterium)右边界核苷酸序列。
优选地,这些实施方案的基因构建体的第二边界核苷酸序列属于农杆菌左边界核苷酸序列。
适当地,这些实施方案的额外核苷酸序列可包括用于表达的核苷酸序列和/或调节核苷酸序列。
在某些优选的实施方案中,包含调节序列的额外核苷酸序列包含与基因构建体的第二边界核苷酸序列相邻并与选择标记核苷酸序列可操作地连接的启动子序列。
在重组基因构建体包含边界序列的一些特别优选的实施方案中,基因构建体包含seqidnos:1-35、49、51-66、81、94或100所示的核苷酸序列,和/或编码seqidnos:38-46所示氨基酸序列的核苷酸序列,或其片段或变体。
在第二方面,本发明提供了用于产生重组基因构建体的方法,所述方法包括从一株或多株植物衍生可插入至植物的遗传物质中的一个或多个核酸片段,从而产生重组基因构建体的步骤,其中所述一个或多个核酸片段由长度为至少15个核苷酸,或优选长度为至少20个核苷酸的多条核苷酸序列组成。
在第二方面的某些优选实施方案中,所述方法包括将第一边界核苷酸序列和第二边界核苷酸序列添加到一条或多条额外核苷酸序列的各自末端的步骤,其中所述一条或多条额外核苷酸序列和至少一部分所述第一边界核苷酸序列衍生自一株或多株植物。
在第三方面,本发明提供了第二方面的方法产生的基因构建体。在特别优选的实施方案中,所述基因构建体包含seqidnos:1-35、49、51-56、66-68、71-92或94-101所示的核苷酸序列,或编码seqidnos:38-46所示氨基酸序列的核酸,或其片段或变体。
优选地,第一至第三方面的一株或多株植物是或包括单子叶植物或双子叶植物。
更优选地,所述一株或多株植物是或包括禾本科(poaceae)的草本植物(grass);谷物,包括高粱、稻、小麦、大麦、燕麦和玉米;豆科物种,包括豆类和花生;茄科物种,包括番茄和马铃薯;十字花科物种,包括卷心菜和东方芥末;葫芦科植物,包括南瓜和西葫芦;蔷薇科植物,包括玫瑰;紫菀科植物,包括莴苣和向日葵,或任何前述植物的亲缘植物。
在某些特别优选的实施方案中,所述一株或多株植物是或包括番茄或番茄的亲缘植物。在某些特别优选的实施方案中,所述一株或多株植物是或包括稻或稻的亲缘植物。在某些特别优选的实施方案中,所述一株或多株植物是或包括高粱或高粱的亲缘植物。
在第四方面,本发明提供了包含第一方面或第三方面的重组基因构建体的载体。适当地,所述载体进一步包含主链核苷酸序列。在一个优选的实施方案中,所述载体主链核苷酸序列包含seqidno:50,或其片段或变体。
优选地,该方面的载体的主链核苷酸序列包含主链插入标记核苷酸序列。在某些优选的实施方案中,主链插入标记核苷酸序列包含seqidno:36或seqidno:37,或其片段或变体。
在该方面的某些优选实施方案中,载体包含另外的基因构建体。
在某些实施方案中,所述另外的基因构建体包含不是植物的或不是衍生自植物的一条或多条核苷酸序列用于插入至植物的遗传物质中。在这些实施方案中,优选地,所述一条或多条核苷酸序列包含选择标记核苷酸序列。所述一条或多条核苷酸序列可包含调节核苷酸序列。在一个特别优选的此类实施方案中,所述另外的基因构建体包含seqidno:69所示的核苷酸序列,或其片段或变体。
在一些特别优选的实施方案中,第四方面的载体包含seqidnos:47、48、63、70、82、93或95所示的核苷酸序列。
在第五方面,本发明提供了包含第一方面或第三方面的重组基因构建体或第四方面的载体的宿主细胞。
在第六方面,本发明提供了对植物进行遗传改良的方法,包括将第一方面或第三方面的重组基因构建体的至少一个核酸片段插入至植物细胞或植物组织的遗传物质中的步骤。
在一些优选的实施方案中,通过细菌介导的植物细胞或植物组织转化,将所述基因构建体的所述至少一核酸片段插入至植物细胞或植物组织的遗传物质中。在所述实施方案中,优选通过农杆菌介导的植物细胞或植物组织转化,优选使用第四方面的载体,将所述基因构建体的所述至少一片段插入至植物细胞或植物组织的遗传物质中。
在一些优选的实施方案中,通过直接转化,例如粒子轰击,将所述基因构建体的所述至少一核酸片段插入至植物细胞或植物组织的遗传物质中。
根据该方面特别优选的是,导入植物细胞或植物组织的遗传物质中的第一或第三方面的基因构建体的至少一核酸片段是可插入至植物的遗传物质中的一个或多个核酸片段,其中所述一个或多个核酸片段由衍生自一株或多株植物的长度为至少15个核苷酸,或优选长度为至少20个核苷酸的多条核苷酸序列组成。
适当地,根据该方面进行遗传改良的植物属于与所述一株或多株植物互交可育的植物和/或属于与所述一株或多株植物相同物种的植物。
优选地,该方面的方法包括选择遗传改良的植物的另外的步骤,其中由于基因构建体的至少一片段插入至植物的遗传物质中,所述植物的一个或多个性状被改变。
优选地,在包括所述步骤的该方面方法的实施方案中,所述性状根据基因构建体的用于表达的一条或多条核苷酸序列在植物中的表达被改变。
在优选的实施方案中,所述一个或多个被改变的性状是相对增加的非生物胁迫耐受性。在优选的实施方案中,所述一个或多个被改变的性状是相对增加的抗病性。在优选的实施方案中,所述一个或多个被改变的性状是相对改善的营养和/或适口性特性。在优选的实施方案中,所述一个或多个被改变的性状是相对改善的形态学特性。
优选地,所述用于表达的一条或多条核苷酸序列的长度为至少15个核苷酸,或更优选至少20个核苷酸。
在一些优选的实施方案中,所述用于表达的一条或多条核苷酸序列包含蛋白编码核苷酸序列。
在一些优选的实施方案中,所述用于表达的一条或多条核苷酸序列包含小rna序列。
在该方面的一个特别优选的实施方案中,通过表达包含一条或多条小rna序列的所述一个或多个分离的核酸,植物的抗病性被相对改善或增加,其中所述分离的核酸能够改变植物病原体的一种或多种核酸的表达、翻译和/或复制。
优选地,植物病原体是病毒性植物病原体。
在第六方面的方法的某些实施方案中,所述方法包括下述另外步骤:
将另外的基因构建体的核酸片段插入至所述植物的遗传物质中;
从所述植物产生植物群,其中第一方面基因构建体的核酸片段和所述另外的基因构建体的核酸片段已插入至所述遗传物质中;和
从所述植物群中选择植物,其中所述植物的遗传物质包含第一方面基因构建体的核酸片段,但不包含所述另外的基因构建体的核酸片段。
优选地,插入至所述植物的遗传物质中的所述另外的基因构建体的核酸片段包含选择标记核苷酸序列。
在一些优选的此类实施方案中,第一方面的基因构建体和所述另外的基因构建体是第四方面的载体。
在另外的或可选的此类实施方案中,所述另外的基因构建体属于另外的载体。
在第七方面,本发明提供了根据第六方面的方法产生的遗传改良的植物或植物部分。
在优选的实施方案中,该方面的植物或植物部分具有相对改善的抗病性。优选地,所述相对改善的抗病性是或包括对病毒性病原体的抗性。
在优选的实施方案中,该方面的植物或植物部分具有相对改善的非生物胁迫耐受性。优选地,所述非生物胁迫耐受性是耐盐性。
在优选的实施方案中,该方面的植物或植物部分具有相对改善的营养和/或适口性特性。
在优选的实施方案中,该方面的植物或植物部分具有相对改善的形态学特性。
在第八方面,本发明提供了植物,其中重组基因构建体的至少一核酸片段已插入至所述植物的遗传物质中,其中所述重组基因构建体包含可插入至植物的遗传物质中的一个或多个核酸片段,其中所述一个或多个核酸片段由衍生自一株或多株植物的长度为至少15个核苷酸,或优选长度为至少20个核苷酸的多条核苷酸序列组成。
优选地,所述重组基因构建体的已插入至植物的遗传物质中的所述至少一核酸片段是由衍生自一株或多株植物的长度为至少15个核苷酸或优选长度至少20个核苷酸的多条核苷酸序列组成的一个或多个核苷酸片段。
适当地,插入了基因构建体的至少一核酸片段的植物属于与衍生所述一条或多条核苷酸序列的一株或多株植物相同的物种和/或与其互交可育。
优选地,第六至第八方面的植物是单子叶植物或双子叶植物。
更优选地,所述植物是或包括禾本科的草本植物;谷物,包括稻、高粱、小麦、大麦、燕麦和玉米;豆科物种,包括豆类和花生;茄科物种,包括番茄和马铃薯;十字花科物种,包括卷心菜和东方芥末;葫芦科植物,包括南瓜和西葫芦;蔷薇科植物,包括玫瑰;紫菀科植物,包括莴苣和向日葵,或任何前述植物的亲缘植物。
在某些特别优选的实施方案中,所述一株或多株植物是或包括番茄或番茄的亲缘植物。在某些特别优选的实施方案中,所述一株或多株植物是或包括稻或稻的亲缘植物。在某些特别优选的实施方案中,所述一株或多株植物是或包括高粱或高粱的亲缘植物。
应当理解,不定冠词“a(一)”和“an(一)”不应被理解为单数不定冠词或以其他方式排除多于一个或多于不定冠词所指的单一目标。例如,“a(一)”核苷酸序列包括一条核苷酸序列、一条或多条核苷酸序列或多条核苷酸序列。
如本文所使用的,除非上下文另有要求,否则词语“包括(comprise)”,“包含(comprises)”和“包含(comprising)”将理解为包括指出的整体或整体组但不排除任何其他整体或整体组。
附图说明
为了使本发明易于理解并付诸实效,现参考附图以举例的方式描述优选的实施方案,其中:
图1给出了本发明的基因构建体和本发明的包含所述基因构建体的载体(pintr2)的示意图。该基因构建体的核苷酸序列如seqidno:1所示。
图2给出了本发明的基因构建体和本发明的包含所述基因构建体的载体的示意图。
图3给出了本发明的基因构建体和本发明的包含所述基因构建体的载体的示意图。
图4给出了用prbcs3c:sgfp:trbcs3c构建体和作为对照的p35s:sgfp:tnos瞬时转化番茄叶肉原生质体的结果。
图5给出了prbcs3c:sgfp:trbcs3c在番茄叶维管组织和气孔中表达的结果。
图6给出了通过在农杆菌渗入的本氏烟草(nicotianabenthamiana)叶子中瞬时表达,由属于番茄actin(肌动蛋白)(act7)基因、cyclophilin(亲环素)(cyp40)基因和ubiquitin(泛素)(ubi3)基因的启动子-终止子对驱动的gfp表达的比较。
图7给出了通过在农杆菌渗入的本氏烟草叶子中瞬时表达,由属于番茄actin基因(act7;左栏)、camv35s基因(中栏)和rubisco(核酮糖-1,5-二磷酸羧化酶)亚基3c(rbcs3c)基因(右栏)的启动子-终止子对驱动的gfp表达的比较。
图8给出了在选择性1mg/lga培养基上用基因内prbcs3c:gs1g245c:trbcs3c构建体转化的番茄子叶再生2周的结果;左边的两个平板是对照的并发子叶,其未与携带构建体的农杆菌共温育。
图9给出了在1mg/l选择性ga培养基上由用基因内prbcs3c:gs1g245c:trbcs3c构建体转化的番茄子叶初始再生4周的结果。
图10给出了使用番茄衍生的amirna构建体靶向黄瓜花叶病毒(cucumbermosaicvirus)序列的结果。显示了本氏烟草叶子农杆菌渗入后的双重luc测定。n=6;误差条表示平均值的标准误差。
图11给出了五株野生型植物与五株ami10(seqidno:15)表达植物的cmv症状发展。a:cmv接种后3周,野生型(上图)与ami10(下图)植物的cmv症状发展。b:cmv接种后3周,野生型(左)与ami10(右)的cmv症状发展。
图12给出了在五株野生型植物与五株ami10(seqidno:15)表达植物中通过qrt-pcr的cmv病毒载量定量。基于两个参考基因actin和gapdh的相对比率的几何平均值计算相对表达比率。
图13显示了使用番茄(栽培品种moneymaker)序列设计具有seqidno:18所示核苷酸序列的rnai构建体的过程,使用所述番茄序列并利用生物信息学集合起来生成seqidno:18,其中每条植物衍生的序列的长度为至少20个核苷酸。
图14列出了使用番茄衍生的rnai构建体靶向黄瓜花叶病毒序列的结果。显示了本氏烟草叶子农杆菌渗入后双重luc测定的结果。n=6;误差条表示平均值的标准误差;t检验显示出非常显著的差异。
图15给出了基本基因内克隆载体pintr2所含的基因构建体的序列(seqidno:1),pintr2示意性地描绘在图1中,显示了:包含农杆菌rb和lb的第一和第二边界核苷酸序列(粗体);番茄rbcs3c启动子和终止子(下划线);用于插入基因和附加基因内表达盒的限制酶位点(粗体)。注意,第一边界核苷酸序列(rb序列)描绘在所述序列的5'末端,第二边界核苷酸序列(lb序列)描述在所述序列的3'末端。
图16给出了农杆菌介导的t-dna插入突变植物(med18)的抗病毒性(a);以及使用番茄衍生的amirna序列抑制番茄med18。
图17给出了fasta格式的seqidnos:1-66、68、71-72、75、77、80、83-89、93和95。
图18给出了本发明的优选克隆构建体pintra的核苷酸序列和结构(seqidno:67)。通过突出显示和/或下划线表示bbvci限制酶位点(seqidno:102);sphi限制酶位点(seqidno:103);rb(seqidno:104);lb(seqidno:105);hpai限制酶位点;pmli限制酶位点;添加以创建克隆位点的核苷酸;部分actin7启动子(seqidno:106)和部分actin7终止子(seqidno:107)。
图19给出了用于与本发明的基因构建体一起共转化的包含选择标记基因的构建体的核苷酸序列(seqidnono:69)和结构,所述选择标记基因(nptii)不是植物的或不是衍生自植物。通过突出显示和/或下划线表示rb;lb、nptii选择标记;双35s启动子;nos终止子、ant1智利番茄(solanumchilense)花色素苷基因;番茄actin7启动子;和番茄rbcs3c终止子。
图20给出了本发明载体的核苷酸序列(seqidno:70)和结构,所述本发明的载体包含本发明的优选基因构建体以及包含不是植物的或不是衍生自植物的选择标记基因(nptii)的另外的基因构建体,用于根据本发明的共转化。通过突出显示和/或下划线表示hpai限制酶位点;pmli限制酶位点;rb;lb、nptii选择标记;可视选择ant1标记和部分actin启动子和终止子。
图21给出了本发明优选的克隆构建体psbiubi1(seqidno:73),其包含来自高粱(sorghumbicolor)的ubi1启动子和终止子;ctgcagpsti限制酶位点和ggcgccsfoi限制酶位点。通过突出显示和/或下划线表示来自sobic004g050000的ubi1启动子和终止子(seqidno:108);ctgcagpsti限制酶位点(seqidno:109);和ggcgccsfoi限制酶位点(seqidno:110)。
图22给出了本发明优选的克隆构建体psbiubi2(seqidno:74),其包含来自高粱的ubi2启动子;来自高粱的ubi1终止子;ctgcagpsti限制酶位点和ggcgccsfoi限制酶位点。通过突出显示和/或下划线表示来自sobic.004g049900的ubi2启动子(seqidno:111)和来自sobic.004g050000的ubi1终止子;以及ctgcagpsti限制酶位点;ggcgccsfoi限制酶位点。
图23给出了本发明优选的克隆构建体posaapx(seqidno:76),其包含稻(oryzasativa)apx启动子和终止子;和由saci或eco53ki和平端限制酶(bluntcutter)psii组成的gagctccggattataa多克隆位点;gaacgt和cgattc:xmni限制酶位点。通过突出显示和/或下划线表示apx启动子(seqidno:112);apx终止子(seqidno:113);由saci或eco53ki和平端限制酶psii(seqidno:114)组成的gagctccggattataa多克隆位点;gaacgt(seqidno:115)和cgattc(seqidno:116);和xmni限制酶位点。
图24给出了表达seqidno:69的番茄植物,显示出增加的花色素苷水平(紫色茎、根、叶脉和部分叶)。
图25给出了用seqidno:69(左)所示的本发明载体共转化的番茄植物,与对照番茄植物(右)相比,显示出强烈的花色素苷产生。
图26给出了本发明的actin1:dreb1a:dreb1a基因构建体(seqidno:78),其包含稻dreb1a基因的核苷酸序列;稻actin1启动子和稻dreb1a终止子。所述基因构建体还包含用于切除和克隆的nhei和pmli限制性消化位点。利用突出显示和下划线表示nhei(seqidno:117)和pmli(seqidno:118)限制位点;dreb1a编码序列(seqidno:119);和在dreb1a编码序列的3’末端添加的gtgtt序列。
图27给出了本发明的nced3:dreb1a:nced3基因构建体(seqidno:79),其包含稻dreb1a基因的核苷酸序列;以及稻nced3启动子和终止子。利用突出显示和下划线表示附加tgc(seqidno:120)和gca(seqidno:121)核苷酸、nced3启动子(seqidno:122)、nced3终止子(seqidno:123)和dreb1a编码序列。
图28给出了在含有100mmnacl的培养基上用actin1:dreb1a:dreb1a(左)或nced3:dreb1a:nced3(右)转化的稻愈伤组织的再生。
图29给出了cmv接种的ami11-it1植物和cmv接种的野生型对照番茄植物。所有野生型植物在新生长(右侧)中都显示出“鞋带”症状。大多数ami11-i植物看起来没有症状(左侧)。
图30列出了wt(野生型)、不成对(azygous)t1和ami11-it1番茄植物中cmv载量的elisa评估。
图31给出了wt、t1不成对和ami11-iit1番茄植物中cmv载量的elisa评估。
图32给出了ami11-i和ami11-ii番茄植物中cmv严重性和植物高度的评估。
图33给出了用cmv感染的ami11-i和ami11-ii系的果实数量和示例性果实形态。
图34给出了具有番茄衍生的抗cmvami10和ami11的“双”抗cmvamirna插入物的核苷酸序列,以及插入物的rna靶向评估。
图35给出了包含cmvamirna10和amirna11的本发明优选的基因构建体的核苷酸序列(seqidno:81)和结构。通过突出显示和文本颜色表示lb;肌动蛋白启动子;sly-mir156b中的cmvamirna10;sly-mir156a中的amirna11;肌动蛋白终止子和rb。
图36给出了本发明的优选载体的核苷酸序列(seqidno:82)和结构,其包含图35中所示的基因构建体连同图19所示的含有选择标记的基因构建体。通过突出显示/文本颜色表示载体的组分。
图37给出了基因内tswv靶向的amirna7序列(seqidno:83);在本氏烟草(n.benthamiana)叶子的农杆菌渗入后使用双luc测定对该序列的rna靶向进行的评估(误差条表示平均值的标准误差);转化以表达该序列的番茄植物的示例性形态。
图38给出了靶向mdmv和scmv的保守区域的高粱衍生的amirna(amirna3和amirna6)的核苷酸序列(seqidnos:84-85)、这些序列的rna靶向评估,以及再生高粱植物。预计成功的转化体具有mdmv/scmv抗性表型。
图39给出了靶向jgmv的高粱衍生的amirna(amirna2、amirna4、amirna5和amirna7)的核苷酸序列(seqidnos:86-89)、这些序列的rna靶向评估,以及再生高粱植物。预计成功的转化体具有jgmv抗性表型。
图40给出了本发明的基因构建体的核苷酸序列(seqidno:90)和结构,其包含高粱ubi1启动子和终止子,以及靶向jgmv的三个高粱衍生的amirna(amirna4、amirna5和amirna2)。构建体的组分由文本颜色来表示。
图41给出了本发明的优选基因构建体的核苷酸序列(seqidno:91)和结构,其包含高粱ubi2启动子和高粱ubi1终止子,以及靶向jgmv的三个高粱衍生的amirna(amirna4、amirna5和amirna2)。构建体的组分由文本颜色来表示。
图42给出了稻衍生的rtsvamirna1的核苷酸序列(seqidno:92)。
图43给出了靶向tswv的番茄衍生的发夹rnai构建体(seqidno:94)的设计。rnai载体的完整核苷酸序列如seqidno:95所示。
图44给出了对图43所示构建体的rna靶向进行的评估;使用图43所示构建体转化的番茄植物的示例性表型;当用tswv攻击时,与野生型番茄植物相比,使用图43所示构建体转化的番茄植物中的tswv载量。
图45给出了番茄衍生的amirna27靶向med18;与枯萎型对照相比,转化的番茄植物中amirna27和med18的表达;与amirna27转化的植物(标记的med18)相比,wt中的cmv载量。
图46给出了与amirna27转化的(标记为a或med18)番茄植物相比,对照(标记为w或wt)中分离的叶丁香假单胞菌(p.syringae)测定的结果。如通过丁香假单胞菌促旋酶的qpcr测量的与amirna27转化系相比,对照中的丁香假单胞菌的丰度。
图47给出了在含有100mmnacl的培养基上用actin1:dreb1a:dreb1a转化的稻植物的再生和生长。
图48给出了在含有100mmnacl的培养基上用nced3:dreb1a:nced3转化的稻植株的再生和生长。
图49给出了与野生型对照系相比,用番茄衍生的amirna27转化的番茄植物的形态比较。
图50给出了靶向med25的番茄衍生的amirna6的核苷酸序列;对amirna6的med25靶向进行的评估;与野生型对照系相比,用amirna6转化的番茄系中amirna6和med25的表达。
图51给出了使用seqidno:69所示构建体转化的番茄系中花色素苷的表达(左);使用seqidno:98所示稻衍生的构建体转化的再生稻植物中花色素苷的表达(右)。
图52给出了番茄衍生的发夹rnai构建体的核苷酸序列(seqidno:100)和结构,所述构建体靶向编码b型异源三聚体g蛋白(ggb1)的γ-亚基的番茄基因;和用所述构建体和seqidno:69所示构建体共转化的示例性转化番茄植物,并表达花色素苷。
图53给出了使用靶向稻badh2的稻衍生的rnai构建体通过粒子轰击产生的发育中的稻植物。预计成功的转化体具有芳香表型。
序列表的简单描述
seqidno:1包含在本发明的基本基因内克隆载体pintr2中的本发明的基因构建体的核苷酸序列(图1中示意性地显示)。
seqidno:2本发明的某些优选的基因构建体中一部分第一边界序列的核苷酸序列。
seqidno:3本发明的某些优选的基因构建体中一部分第二边界序列的核苷酸序列。
seqidno:4栽培番茄(solanumlycopersicum)rubisco亚基3c(rbs3c)基因启动子的核苷酸序列。
seqidno:5栽培番茄肌动蛋白(actin)基因启动子的核苷酸序列。
seqidno:6栽培番茄ubiquitin(泛素)基因启动子的核苷酸序列。
seqidno:7栽培番茄cyclophilin(亲环素)基因启动子的核苷酸序列。
seqidno:8栽培番茄rubisco亚基3c(rbs3c)基因终止子的核苷酸序列。
seqidno:9栽培番茄肌动蛋白(actin)基因终止子的核苷酸序列。
seqidno:10栽培番茄ubiquitin(泛素)基因终止子的核苷酸序列。
seqidno:11栽培番茄cyclophilin(亲环素)基因终止子的核苷酸序列。
seqidno:12番茄mir156b基因的核苷酸序列。成熟的mirna大写。
seqidno:13基于靶向黄瓜花叶病毒(cmv)k节段1复制酶(核苷酸2665-2685)的seqidno:12的番茄衍生的amirna构建体的核苷酸序列。成熟的mirna大写。
seqidno:14基于靶向k节段2orf3(核苷酸198-218)的seqidno:12的番茄衍生的amirna构建体的核苷酸序列。成熟的mirna大写。
seqidno:15基于靶向cmvk节段3orf1(核苷酸56-76)的seqidno:12的番茄衍生的amirna构建体的核苷酸序列。成熟的mirna大写。
seqidno:16基于靶向cmvk节段1复制酶(核苷酸1437-1457)的seqidno:12的番茄衍生的amirna构建体的核苷酸序列。成熟的mirna大写。成熟的mirna大写。
seqidno:17基于靶向cmvk节段3orf1(核苷酸707-727)的seqidno:12的番茄衍生的amirna构建体的核苷酸序列。成熟的mirna大写。
seqidno:18靶向cmv的番茄衍生的rnai构建体的核苷酸序列。
seqidno:19与cmvk节段1复制酶核苷酸751-896高度相似的seqidno:18的片段的核苷酸序列。
seqidno:20与cmvk节段1复制酶核苷酸1235-1358高度相似的seqidno:18的片段的核苷酸序列。
seqidno:21与cmvk节段3orf2(外壳蛋白)核苷酸250-375高度相似的seqidno:18的片段的核苷酸序列。
seqidno:22靶向番茄斑萎病毒(tomatospottedwiltvirus)(tswv)的番茄衍生的rnai构建体的核苷酸序列。
seqidno:23与tswvqld1节段lrdrp核苷酸1918-2155高度相似的seqidno:22的片段的核苷酸序列。
seqidno:24与tswvqld1节段lrdrp核苷酸8429-8639高度相似的seqidno:22的片段的核苷酸序列。
seqidno:25与tswvqld1节段morf1核苷酸187-360高度相似的seqidno:22的片段的核苷酸序列。
seqidno:26与tswvqld1节段morf2核苷酸297-510高度相似的seqidno:22的片段的核苷酸序列。
seqidno:27番茄甜菜碱醛脱氢酶(badh)cdna的核苷酸序列(gi209362342)。
seqidno:28番茄山梨醇脱氢酶(sdh)cdna的核苷酸序列(gi78183415)。
seqidno:29番茄渗调蛋白(osmotin)cds的核苷酸序列(gi460400210)。
seqidno:30番茄谷氨酰胺合成酶(gts)cdna的核苷酸序列(gi460409535)。
seqidno:31番茄八氢番茄红素去饱和酶cdna的核苷酸序列(gi512772532)。
seqidno:32番茄5-烯醇式丙酮-3-磷酸莽草酸(5-enolpyruvyl-3-phosphoshikimate)cdna的核苷酸序列(gi822092668)。
seqidno:33番茄乙酰乳酸合酶cdna的核苷酸序列(gi723680771)。
seqidno:34番茄原卟啉原氧化酶cdna的核苷酸序列(gi723658549)。
seqidno:35智利番茄花色素苷1(ant1)cdna的核苷酸序列(gi126653934)。
seqidno:36番茄叶绿素合酶cdna的核苷酸序列(gi460401624)。
seqidno:37针对茄属(solanum)表达而密码子优化的芽胞杆菌rna酶自杀构建体的核苷酸序列,具有来自马铃薯st-ls1基因的内含子。
seqidno:38seqidno:27编码的甜菜碱醛脱氢酶的氨基酸序列。
seqidno:39seqidno:28编码的番茄山梨醇脱氢酶蛋白的氨基酸序列。
seqidno:40seqidno:29编码的番茄渗调蛋白(osmotin)的氨基酸序列。
seqidno:41seqidno:30编码的番茄谷氨酰胺合成酶蛋白的氨基酸序列。
seqidno:42seqidno:31编码的番茄八氢番茄红素去饱和酶蛋白的氨基酸序列。
seqidno:43seqidno:32编码的番茄5-烯醇式丙酮-3-磷酸莽草酸蛋白的氨基酸序列。
seqidno:44seqidno:33编码的番茄乙酰乳酸合酶蛋白的氨基酸序列。
seqidno:45seqidno:34编码的番茄protox蛋白的氨基酸序列。
seqidno:46seqidno:35编码的智利番茄花色素苷1蛋白的氨基酸序列。
seqidno:47图1中示意性描绘的基本基因内克隆载体pintr2的核苷酸序列。
seqidno:48本发明的载体'pintr2gs1g245ccml18'的核苷酸序列。
seqidno:49与天然gs1启动子和终止子序列可操作地连接的谷氨酰胺合成酶1(gs1)g245c标记基因的核苷酸序列。
seqidno:50本发明的修饰的part27主链的核苷酸序列。
eqidno:51编码g245c蛋白的番茄gs1g733t基因的cds的核苷酸序列。
seqidno:52编码h249y蛋白的番茄gs1c745tcds的cds核苷酸序列。
seqidno:53番茄gs1启动子的核苷酸序列。
seqidno:54番茄gs1终止子的核苷酸序列。
seqidno:55番茄八氢番茄红素去饱和酶启动子的核苷酸序列。
seqidno:56番茄八氢番茄红素去饱和酶终止子的核苷酸序列。
seqidno:57番茄乙酰乳酸合酶启动子的核苷酸序列。
seqidno:58番茄乙酰乳酸合酶终止子的核苷酸序列。
seqidno:59番茄5-烯醇式丙酮酰莽草酸-3-磷酸合酶(5-enolpyruvylshikimate-3-phosphatesynthase)启动子的核苷酸序列。
seqidno:60番茄5-烯醇式丙酮酰莽草酸-3-磷酸合酶终止子的核苷酸序列。
seqidno:61番茄protox启动子的核苷酸序列。
seqidno:62番茄protox终止子的核苷酸序列。
seqidno:63基因内克隆载体pintr2(seqidno:1)的核苷酸序列,其在用pmli和pcii限制酶消化后除去,并促进核苷酸序列连接到pintr2中。
seqidno:64来自番茄的med18基因的核苷酸序列(gi|723704094|ref|xm_010323502.1)。
seqidno:65靶向番茄med18的amirna序列(med18ami3)的核苷酸序列。
seqidno:66靶向番茄med18的amirna序列(med18ami27)的核苷酸序列。
seqidno:67pintra的基本基因内克隆构建体的核苷酸序列。
seqidno:68pintra的含有限制性消化位点的可去除序列的核苷酸序列。
seqidno:69用于与本发明的基因构建体一起共转化的包含选择标记基因(nptii)的构建体的核苷酸序列,所述选择标记基因(nptii)不是植物的或不是衍生自植物。
seqidno:70用于根据本发明共转化的本发明载体的核苷酸序列,其包含本发明的优选基因构建体以及包含选择标记基因(nptii)的另外的基因构建体,所述选择标记基因(nptii)不是植物的或者不是衍生自植物。
seqidno:71本发明的某些优选的基因构建体中一部分第一边界序列的核苷酸序列。
seqidno:72本发明的某些优选的基因构建体中一部分第二边界序列的核苷酸序列。
seqidno:73psbiubi1的核苷酸序列。
seqidno:74psbiubi2的核苷酸序列。
seqidno:75psbiubi1和psbiubi2克隆位点处间隔区的核苷酸序列。
seqidno:76posaapx构建体的核苷酸序列。
seqidno:77posaapx克隆位点处间隔区的核苷酸序列。
seqidno:78稻actin1:dreb1a:dreb1a构建体的核苷酸序列。
seqidno:79稻nced3:dreb1a:nced3构建体的核苷酸序列。
seqidno:80番茄衍生的双抗cmvamirna插入物的核苷酸序列。
seqidno:81包含seqidno:80的基因内番茄衍生的构建体的核苷酸序列。
seqidno:82包含seqidno:81的载体的核苷酸序列。
seqidno:83番茄衍生的抗tswvamirna7的核苷酸序列。
seqidno:84靶向mdmv和scmv的保守区域的高粱衍生的amirna3的核苷酸序列。
seqidno:85靶向mdmv和scmv的保守区域的高粱衍生的amirna6的核苷酸序列。
seqidno:86靶向jgmv的高粱衍生的amirna2的核苷酸序列。
seqidno:87靶向jgmv的高粱衍生的amirna4的核苷酸序列。
seqidno:88靶向jgmv的高粱衍生的amirna5的核苷酸序列。
seqidno:89靶向jgmv的高粱衍生的amirna7的核苷酸序列。
seqidno:90psbiubi1中的高粱衍生的三重抗jgmvamirna构建体的核苷酸序列。
seqidno:91psbiubi2中的高粱衍生的三重抗jgmvamirna构建体的核苷酸序列。
seqidno:92靶向rtsv的稻衍生的amirna1的核苷酸序列。
seqidno:93包含seqidno:92的载体的核苷酸序列。
seqidno:94靶向tswv的番茄衍生的发夹rnai的核苷酸序列。
seqidno:95包含seqidno:94的载体的核苷酸序列。
seqidno:96番茄med25基因的核苷酸序列。
seqidno:97靶向med25的番茄衍生的amirna6的核苷酸序列。
seqidno:98稻衍生的r1g1b:osb2:r1g1b构建体的核苷酸序列。
seqidno:99番茄ggb1基因的核苷酸序列。
seqidno:100番茄衍生的发夹rnai构建体的核苷酸序列,其靶向编码b型异源三聚体g蛋白(ggb1)的γ-亚基的番茄基因。
seqidno:101靶向badh2的稻衍生的rnai构建体的核苷酸序列。
seqidno:102bbvci限制酶位点的核苷酸序列。
seqidno:103sphi限制酶位点的核苷酸序列。
seqidno:104rb序列的核苷酸序列。
seqidno:105lb序列的核苷酸序列。
seqidno:106部分actin7启动子的核苷酸序列。
seqidno:107部分actin7终止子的核苷酸序列。
seqidno:108高粱ubi1启动子和终止子的核苷酸序列。
seqidno:109psti限制位点的核苷酸序列。
seqidno:110sfoi限制位点的核苷酸序列。
seqidno:111高粱ubi2启动子的核苷酸序列。
seqidno:112稻apx启动子的核苷酸序列。
seqidno:113稻apx终止子的核苷酸序列。
seqidno:114posaapx的多克隆位点的核苷酸序列。
seqidno:115xmni限制位点的核苷酸序列。
seqidno:116xmni限制位点的核苷酸序列。
seqidno:117nhei限制位点的核苷酸序列。
seqidno:118pmli限制位点的核苷酸序列。
seqidno:119添加了3'gtgtt的稻dreb1a编码序列的核苷酸序列。
seqidno:120fspi限制位点的核苷酸序列。
seqidno:121fspi限制位点的核苷酸序列。
seqidno:122稻nced3启动子的核苷酸序列。
seqidno:123稻nced3终止子的核苷酸序列。
seqidno:124sly-mir156b中番茄衍生的抗cmvamirna10的核苷酸序列。
seqidno:125sly-mir156a中番茄衍生的抗cmvamirna11的核苷酸序列。
seqidnos:126-152本说明书中提出的引物的核苷酸序列。
具体实施方式
本发明至少部分地基于认识到需要对植物进行遗传改良,其中避免将不是衍生自或不可衍生自植物的核苷酸序列引入至植物的遗传物质中。
因此,本发明广泛地提供了使用包含衍生自一株或多株植物的核苷酸序列的重组基因构建体产生遗传改良植物的方法。在一个优选的实施方案中,所述一条或多条核苷酸序列衍生自单一植物。适当地,在所述一条或多条核苷酸序列衍生自多于一株植物的实施方案中,所述植物是相同的物种和/或互交可育。
应当认识到,由于将本发明的优选基因构建体的核酸片段插入植物的遗传物质中而发生的基因改变可以与自然界中发生的基因重组相同或至少相似,例如用于增加植物群基因库的多样性,以增加其在不断变化的环境条件下的生存变化的天然基因重组。
如下文所述,还应当认识到,优选的是利用本发明的优选基因构建体插入植物中的核苷酸序列包含至少15个,或优选至少20个植物衍生的核苷酸。对于本发明,已经认识到,该核苷酸序列长度通常是被理解为在植物中发挥功能的核苷酸序列的最小长度。
如本文所用,术语“植物”应理解为包括:
“有胚植物(embryophyta)”或“陆生植物”,参考margulis,l(1971)evolution,25:242-245(通过引用并入本文),包括苔类、角苔类、苔藓以及维管植物;
“绿色植物亚界(viridiplantae)”或“绿色植物(greenplants)”,参考copeland,hf(1956)paloalto:pacificbooks,p.6(通过引用并入本文),包括陆生植物和绿藻;
“原始色素体生物(archaeplastida)”,参考cavalier-smith,t(1981)biosystems14:461–481(通过引用并入),包括陆生植物、绿色植物、红藻植物门(红藻)和灰藻门(褐藻(glaucophytealgae));以及
“植物界(vegetabilia)”,参考linnaeus,c(1751)philosophiabotanica,1sted,p.37(通过引用并入),包括陆生植物、绿色植物、原始色素体生物和多种多样的藻类和真菌,例如包括蘑菇在内的食用菌。
如本文所用,“基因构建体”应理解为是指包含一个或多个分离的核酸的人工产生的遗传物质节段。
如本文所用,由植物“衍生的”或“可衍生”的核苷酸序列应理解为是指与植物的天然或内源遗传物质中发现的核苷酸序列基本上相同的核苷酸序列。应当容易理解的是,包含衍生自植物或可衍生自植物的核苷酸序列的分离的核酸不必从植物获得,而可以参考下文提供的详情以任何合适的方式获得。
优选的是,“衍生”自或“可衍生”自植物的核苷酸序列与天然或内源植物核苷酸序列相同。适当地,至少其中植物衍生的或植物可衍生的核苷酸序列是蛋白质编码序列时,所述衍生的或可衍生的核苷酸序列将编码与相应的天然或内源氨基酸序列基本相同或优选相同的氨基酸序列。然而,应当理解,尽管优选植物衍生的或植物可衍生的核苷酸序列与天然或内源植物核苷酸序列相同,但在某些替代实施方案中,核苷酸序列可包含同义核苷酸替换,条件是由核苷酸序列编码的蛋白质与相应的天然或内源植物蛋白质基本相同或优选相同。
如本文在遗传物质(包括基因构建体)背景下所用,“重组”应当理解为是指衍生自多种来源的遗传物质。应当理解,尽管“重组”的遗传物质的局部(part)、部分(portion)或片段可以包含对应于生物有机体(例如植物)遗传物质的天然核苷酸序列的核苷酸序列,但是重组遗传物质中核苷酸序列的排列不会发生于所述生物有机体的遗传物质中。
应当认识到,本发明的重组基因构建体被设计为促进植物的遗传改良,其中将基因构建体的至少一个核酸片段插入植物的遗传物质中,所述核酸片段由一条或多条衍生自或可衍生自植物的核苷酸序列组成。
适当地,使用本发明的基因构建体避免或至少基本上最小化产生包含不是衍生自一株或多株植物的核苷酸序列的遗传改良植物。
适当地,根据本发明插入植物中的基因构建体的核酸片段由衍生自或可衍生自一株或多株植物的一条或多条长度为至少15个,或优选至少20个核苷酸的核苷酸序列组成,其中所述一株或多株植物与所述植物互交可育。
在实施方案中,衍生或可衍生本发明基因构建体的核苷酸序列的一株或多株植物是或包括如上所述分类为植物界的有机体。
在优选的实施方案中,衍生或可衍生本发明的基因构建体的核苷酸序列的一株或多株植物是或包括如上所述的分类为原始色素体生物的有机体。
更优选地,衍生或可衍生本发明的基因构建体的核苷酸序列的一株或多株植物是或包括如上所述的分类为绿色植物亚界的有机体。
甚至更优选地,衍生或可衍生本发明的基因构建体的核苷酸序列的一株或多株植物是或包括如上所述的分类为有胚植物的有机体。
在一些实施方案中,植物是包括微藻和大型藻类的藻类。
在一些实施方案中,植物是包括蘑菇在内的食用菌。
优选地,植物是单子叶植物或双子叶植物。
更优选地,所述一株或多株植物是或包括:禾本科(poaceae)的草本植物,例如甘蔗;棉属(gossypium)种,例如棉花;浆果,例如草莓;树的物种,包括诸如苹果、橙等果树和诸如扁桃等坚果树;观赏植物,例如观赏性开花植物,包括诸如玫瑰等蔷薇科植物;藤本植物,包括诸如葡萄等果藤;谷物,包括高粱、稻、小麦、大麦、燕麦和玉米;豆科物种,包括诸如大豆和花生等豆类;茄科物种,包括番茄和马铃薯;十字花科物种,包括卷心菜和东方芥末;葫芦科植物,包括南瓜和西葫芦;蔷薇科植物,包括玫瑰;紫菀科植物,包括莴苣、菊苣和向日葵,或任何前述植物的亲缘植物。
在一些具体的实施方案中,所述植物是或包括番茄。
在一些特别优选的实施方案中,所述植物是或包括高粱。
在一些特别优选的实施方案中,所述植物是或包括稻,包括野生稻。
分离的核酸和蛋白质
出于本发明的目的,“分离的”是指已从其天然状态移除或以其他方式经受人为操纵的材料。
分离的材料可以基本上或实质上不含在其天然状态下通常伴随它的组分,或者可以被操纵以便与通常在其天然状态下伴随它的组分一起处于人工状态。分离的材料可以是天然的、化学合成的或重组的形式。
本文所用的术语“核酸”表示单链或双链dna和rna。dna包括基因组dna和cdna。rna包括mrna、rna、srna、rnai、sirna、crna和自催化rna。核酸也可以是dna-rna杂合体。核酸包含核苷酸序列,所述核苷酸序列通常包括含有a、g、c、t或u碱基的核苷酸。然而,核苷酸序列可包括其他碱基,例如肌苷、甲基胞嘧啶、甲基肌苷、甲基腺苷和/或硫代尿苷,但不限于此。
“多核苷酸”是具有八十(80)个或更多个连续核苷酸的核酸,而“寡核苷酸”具有少于八十(80)个连续核苷酸。
“探针”可以是被合适标记的用于在例如northern或southern印迹中检测互补序列的单链或双链寡核苷酸或多核苷酸。
“引物”通常是优选具有15-50个连续核苷酸的单链寡核苷酸,其能够与互补核酸“模板”退火并通过诸如taq聚合酶、rna依赖性dna聚合酶或sequenasetm等dna聚合酶的作用以模板依赖性方式延伸。
如本文所用,“蛋白质”是指包含天然和/或非天然氨基酸的氨基酸聚合物,包括本领域熟知的l-和d-异构形式。
在某些实施方案中,本发明的基因构建体的分离的核酸或由本发明的基因构建体编码的分离的蛋白质分别是片段核酸或蛋白质。
在某些实施方案中,“片段”核酸包含构成seqidnos:1-35、49、51-56、66-68、71-92或94-101所示核苷酸序列的小于100%,但至少20%,优选至少30%,更优选至少80%或甚至更优选至少90%、95%、96%、97%、98%或99%的核苷酸序列。
在某些实施方案中,“片段”蛋白质包含构成seqidnos:38-46所示氨基酸序列的小于100%,但至少20%,优选至少30%,更优选至少80%,或甚至更优选至少90%、95%、96%、97%、98%或99%的氨基酸序列。
在一个优选的实施方案中,本发明的基因构建体的片段包含seqidnos:1、67、73-74、76、81、95、98、100或101所示核苷酸序列的不超过10、12、15、20、30、40、50、60、70、80、90、100、120、150、200、250、300、350、400、500、600、700、800、900、1000、1500、2000、2500或3000个连续核苷酸。
本发明基因构建体的分离的核酸,由本发明基因构建体编码的分离的蛋白质,或本发明基因构建体导致转录或翻译沉默或增强的核苷酸序列可以分别是“变异的”核酸或蛋白质,其中一个或多个核苷酸或氨基酸分别已经缺失或分别被不同的核苷酸或氨基酸替换。
变体包括天然存在的(例如,等位基因)变体、直系同源物(例如来自其他植物)和合成变体,例如使用诱变技术在体外产生的变体。
在一些实施方案中,核酸变体包括与seqidnos:1-35、49、51-56、66-68、71-92或94-101所示核苷酸序列具有至少75%、80%、85%、90%或95%、96%、97%、98%或99%的核苷酸序列同一性的分离的核酸。
在一些实施方案中,蛋白质变体包括与seqidnos:38-46所示氨基酸序列具有至少75%、80%、85%、90%或95%、96%、97%、98%或99%的氨基酸序列同一性的蛋白质。
在本文中通常用于描述各核苷酸序列和氨基酸序列之间的序列关系的术语包括“比较窗”、“序列同一性”、“序列同一性百分比”和“基本同一性”。因为各自的核酸/蛋白质可各自包含(1)核酸/蛋白质共有的完整核酸/蛋白质序列的仅一个或多个部分,和(2)在核酸/蛋白质之间不同的一个或多个部分,序列比较通常通过比较“比较窗”上的序列来进行,以鉴定和比较序列相似性的局部区域。“比较窗”是指通常6、9或12个连续残基的与参考序列进行比较的概念节段。
与用于各序列的最佳比对的参考序列相比,比较窗可包括约20%或更少的添加或缺失(即,缺口)。用于对齐比较窗的序列的最佳比对可以通过以下方式进行:算法的计算机化的执行(intelligenetics的geneworks程序;威斯康星遗传学软件包发布7.0(wisconsingeneticssoftwarepackagerelease7.0)中的gap、bestfit、fasta和tfasta,geneticscomputergroup,575sciencedrivemadison,wi,usa,通过引用并入本文),或检查和通过任何所选各种方法生成的最佳比对(即在比较窗上引起最高百分比同源性)。还可以参考例如altschuletal.,1997,nucl.acidsres.253389公开的blast程序家族(blastfamilyofprograms),该文献通过引用并入本文。序列分析的详细讨论见于currentprotocolsinmolecularbiologyeds.ausubeletal.(johnwiley&sonsincny,1995-1999)的19.3单元。
术语“序列同一性”在本文中以其最广泛的含义使用,包括考虑到使用标准算法进行适当比对的、考虑到序列在比较窗上相同的程度的精确核苷酸或氨基酸匹配的数量。因此,“序列同一性百分比”通过以下方式计算:在比较窗上比较两个最佳比对序列,确定两条序列中都存在相同核酸碱基(例如,a、t、c、g、u)的位置的数目以产生匹配位置的数目,将匹配位置的数量除以比较窗中的位置总数目(即窗大小),并将结果乘以100以产生序列同一性的百分比。例如,“序列同一性”可以理解为表示由dnasis计算机程序(用于windows的版本2.5;可从hitachisoftwareengineeringco.,ltd.,southsanfrancisco,california,usa获得)计算的“匹配百分比”。
序列分析的详细讨论可以在上文ausubel等人的第19.3章中找到。
应当认识到,可以通过诱变蛋白或编码核酸例如通过随机诱变或定点诱变来产生核酸和蛋白质变体,但不限于此。核酸诱变方法的实例在上文ausubel等人的currentprotocolsinmolecularbiology(现代分子生物学实验技术)的第9章提供,通过引用将该文献并入本文。诱变也可以通过化学方法诱导,例如甲基磺酸乙酯(ems)和/或辐射方法,例如本领域已知的种子的快中子辐照(carrolletal.,1985,proc.natl.acad.sci.usa824162;carrolletal.,1985,plantphysiol.7834;menetal.,2002,genomeletters3147)。
基因构建体
本发明的一个方面提供了包含可插入至植物的遗传物质中的一个或多个核酸片段的重组基因构建体,其中所述一个或多个核酸片段包含衍生自一株或多株植物的长度为至少15个核苷酸或优选长度为至少20个核苷酸的多条核苷酸序列,由所述多条核苷酸序列组成,或基本上由所述多条核苷酸序列组成。
如在本文中所用,“基本上由”衍生自或可衍生自一株或多株植物的核苷酸序列“组成”的核酸片段,应理解为包括不超过1、2、3或4个核苷酸不是衍生自或不可衍生自植物。
优选地,所述可插入至植物的遗传物质中的一个或多个核酸片段由植物衍生的或植物可衍生的核苷酸序列组成。
在某些优选的实施方案中,可插入至植物的遗传物质中的所述重组基因构建体的所述一个或多个核酸片段由以下长度的多条核苷酸序列组成:16、17、18、19、20、21、22、23、24、25、26、27、28、29、30、31、32、33、34、35、36、37、38、39、40、41、42、43、44、45、46、47、48、49、50、51、52、53、54、55、56、57、58、59、60、61、62、63、64、65、66、67、68、69、70、71、72、73、74、75、76、77、78、79、80、81、82、83、84、85、86、87、88、89、90、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、117、118、119、120、121、122、123、124、125、126、127、128、129、130、131、132、133、134、135、136、137、138、139或140个核苷酸。
在某些优选的实施方案中,所述多条核苷酸序列的长度为至少100、200、300、400、500、600、700、800、900或1000个核苷酸。
在一个优选的实施方案中,所述一条或多条核苷酸序列衍生自一株植物。
适当地,在所述一条或多条核苷酸序列衍生自多于一株植物的实施方案中,所述植物是互交可育的,例如性相容亲缘植物,和/或是相同物种。
优选地,可插入至植物的遗传物质中的基因构建体的一个或多个核酸片段的总长度为至少:100、200、300、400、500、600、700、800、900、1000、1500、2000、2500、3000或3500个碱基对。
参考实施例,应当认识到,该方面的优选重组基因构建体pintr2包含适于插入至植物的遗传物质中的1110个碱基对,并且该构建体是被设计为进一步接受植物衍生的用于插入或并入至植物的遗传物质中的核苷酸序列的克隆构建体。类似地,该方面的优选重组基因构建体pintra包含适于插入至植物的遗传物质中的1787个碱基对,并且该构建体是被设计为进一步接收植物衍生的用于插入或并入至植物的遗传物质中的核苷酸序列的克隆构建体。此外,seqidnos:78、79、81、98和100中所示的优选重组基因构建体分别包含2387、3369、2084、3304和3071个适于插入至植物的遗传物质中的碱基对。
基因构建体的序列
该方面的重组基因构建体会适当地包含可按如下分类的一条或多条核苷酸序列。
用于表达的序列
优选地,该方面的重组基因构建体包含一条或多条用于表达的核苷酸序列。适当地,所述用于表达的核苷酸序列是该方面的基因构建体的一个或多个可插入至植物的遗传物质中的核酸片段。
如本文在本发明的重组基因构建体的上下文中所用,“用于表达”的核苷酸序列应理解为是指基因构建体的能够在宿主细胞或宿主有机体例如植物中表达的核苷酸序列。优选地,用于表达的序列是用于在植物中表达的序列。
优选地,本发明的基因构建体包含一条或多条用于表达的额外核苷酸序列,其中所述核苷酸序列适合在植物中表达以改变或修饰植物的性状。参考实施例,应当认识到,已经证明某些优选核苷酸序列的表达改变或修饰植物的性状,包括非生物胁迫耐受性、营养特性和抗病性。
在某些优选的实施方案中,一条或多条所述用于在植物中表达的核苷酸序列包含蛋白质编码核苷酸序列。用于表达的蛋白质编码序列可以是任何合适的蛋白质编码序列。优选地,核苷酸序列编码与所需或有益的植物性状或特征相关的蛋白质,这是本领域所熟知的。举例来说,本文已经证明了编码包括dreb1a和ant1在内的蛋白质的核苷酸序列的表达,dreb1a与非生物胁迫耐受性,包括耐盐性相关,ant1与花色素苷产生相关。
在某些特别优选的实施方案中,所述蛋白质编码核苷酸序列包含seqidnos:38-46、76、78或98所示的核苷酸序列或其片段或变体。
在一些优选的实施方案中,基因构建体包含一条或多条序列,所述一条或多条序列包含一条或多条适合在植物中表达以改变或修饰所述植物的性状的非编码核苷酸序列。
优选地,所述非编码序列包含小rna序列。
如本文所用,“小rna”应理解为是指具有结合并调节其他核酸分子的表达、翻译和/或复制能力的小的非编码rna分子。对于小的非编码rna分子和植物中的这类分子,本领域技术人员分别被导向ipsaro,j.j.,&joshua-tor,l.,2015,naturestruc.&mol.biol.2220和axtell,j.m.,2013,ann.rev.plantbiol.64,137-159,将这两篇文献通过引用并入本文。
应当理解,如本文所用,术语小rna包括所有这样的分子,而不管科学界可以使用的具体名称如何。作为非限制性实例,技术人员应当容易地理解,如本文所用,术语小rna包括称为“mirna”和“sirna”的小的非编码rna分子。
还应当理解,小rna分子通常与其具有结合并调节其表达、翻译和/或复制的能力的核酸分子具有高度的核苷酸序列同一性,即对于所述核酸分子。然而,还应理解,小rna分子不必与这样的序列具有100%的同一性。
在某些实施方案中,本发明的小rna与其针对的具有结合并调节表达、翻译和/或复制的能力的核酸分子具有至少85%、至少90%或至少91%、92%、93%、94%、95%、96%、97%、98%或99%的同一性。
应当认识到,成熟的小rna的长度通常为18-40个核苷酸。通常,成熟的植物小rna的长度为19-26个核苷酸,特别是19-24个核苷酸。因此,基因构建体的用于表达的小rna核苷酸序列的核苷酸序列长度可以是19、20、21、22、23或24个核苷酸。
小rna序列可以是小rna前体序列。如本领域技术人员会容易理解的,小rna前体包含比成熟的小rna更长的核苷酸序列。当在植物中表达时,小rna前体被加工成成熟的小rna。通常,dicer样蛋白如dcl-1、dcl-2、dcl-4和/或argonaute蛋白-1(ago1)介导将小rna前体加工成成熟的小rna,但不限于此。
在某些优选的实施方案中,基因构建体的包含一条或多条微rna序列的用于表达的核苷酸序列包含mirna前体(前体mirna(pre-mirna))(例如seqidno:12),或包含修饰的前体mirna的人工mirna(amirna)构建体(例如seqidnos:13-17)。
本领域技术人员应当容易地理解,在植物中,前体mirna是从中产生成熟的小rna序列的非蛋白质编码序列。通常,前体mirna序列的长度在约60个核苷酸和约100个核苷酸之间,但应当认识到它们的长度可以大于几百个核苷酸。在加工成一个或多个成熟的mirna之前,这些前体mirna序列形成二级“茎环”结构;参见axtell,j.m.,同上。
适当地,可以使用包含修饰的前体mirna序列的amirna构建体,其中所述前体mirna序列的一条或多条小rna序列被一条或多条感兴趣的小rna序列(例如seqidnos:13-17)替换。
在某些其他优选的实施方案中,包含一条或多条小rna序列的用于表达的序列包含'双链rna'('dsrna')或'rnai'构建体(例如seqidno:18和seqidno:22)。
将容易理解的是,设计dsrna或rnai构建体以表达形成双链rna“发夹”结构的rna序列。举例来说,技术人员被导向miki,d,&shimamoto,k,2004,plantandcellphysiology45490。通常,所述发夹结构的长度高达数百个碱基对。将容易理解的是,当在植物中表达时,所述发夹结构被加工成上文所述的小rna。
在一些优选的实施方案中,基因构建体的用于表达的核苷酸序列的一条或多条小rna序列能够改变植物病原体的一种或多种核酸的表达、翻译和/或复制。
在某些特别优选的实施方案中,所述小rna能够抑制植物病毒核酸的复制。在其他特别优选的实施方案中,所述小rna能够抑制细菌植物病原体的感染和/或复制。另外或可选地,所述小rna能够抑制真菌植物病原体和/或植物感染性或侵染性卵菌、线虫和/或昆虫的感染和/或复制。
在特别优选的实施方案中,所述包含小rna序列的用于表达的非编码核苷酸序列包含seqidnos:12-26、80、81、83-92或94-101所示的核苷酸序列或其片段或变体。
该方面用于表达的基因构建体的一条或多条核苷酸序列可以另外地或可选地包含一条或多条选择标记核苷酸序列。
如本文所用,“选择标记”核苷酸序列是指适合在植物细胞、植物组织或植物中表达,并且适于促进植物细胞、植物组织或植物鉴定的核苷酸序列,其中已经将本发明的基因构建体或其片段插入所述植物细胞、植物组织或植物的遗传物质中。
在特别优选的实施方案中,所述一条或多条选择标记核苷酸序列包含seqidnos:27-35或119中的一个或多个,或其片段或变体,或一条或多条分别编码seqidnos:38-46中任何一个所示氨基酸序列的核苷酸序列,或其片段或变体。
作为非限制性实例,基因构建体的一条或多条用于表达的额外核苷酸序列的选择标记核苷酸序列可以属于这样的基因,所述基因当在植物中表达时,与相应的野生型植物相比,所述基因增加植物对有毒代谢物的耐受性,或增加植物利用替代营养源的能力。
在这方面,将认识到,编码seqidno:38所示氨基酸序列的seqidno:27所示的核苷酸序列属于甜菜碱醛脱氢酶基因。技术人员将认识到,包含甜菜碱醛脱氢酶基因核苷酸序列或其片段或变体的选择标记的表达可以增加植物对化学品甜菜碱醛的耐受性,从而促进通过应用外源甜菜碱醛进行选择。
作为另一个非限制性实例,一条或多条用于表达的额外核苷酸序列的选择标记核苷酸序列可以是赋予除草剂耐受性的基因的。作为非限制性实例,将认识到可以使用编码包含引入的赋予除草剂耐受性的突变之光合作用相关的或其他除草剂作用酶靶标的选择标记核苷酸序列。
在这方面,将认识到编码seqidno:41所示氨基酸序列的seqidno:30所示核苷酸序列属于谷氨酰胺合成酶基因。
技术人员将认识到,编码包含与相应的野生型蛋白质相比一个或多个突变的谷氨酰胺合成酶蛋白的选择标记核苷酸序列的表达,可以赋予植物对除草剂(例如草铵膦(glufosinateammonium))的耐受性,从而促进通过应用外源性除草剂进行选择。就此而言,技术人员被导向tischer,e.,dassarma,s.,&goodman,h.m.,1986,mol.gen.genet.203221;和pornprom,t.,prodmatee,n.,&chatchawankanphanich,o.,2009,pestmanagementsci.65216,通过引用将这两篇文献并入本文。
作为另一个非限制性实例,一条或多条用于表达的额外核苷酸序列的选择标记核苷酸序列可以是促进可视选择的基因。
在这方面,编码seqidno:46所示氨基酸序列的核苷酸序列seqidno:35属于花色素苷1基因。
技术人员将认识到,包含花色素苷1基因核苷酸序列或其片段或变体的选择标记表达可以促进用本发明的基因构建体或其片段转化的植物的可视选择。
将容易理解的是,根据本发明的该实施方案,可以使用本领域技术人员已知的一系列其他合适的选择标记。
将认识到,在一些实施方案中,本发明基因构建体的选择标记核苷酸序列也可以是适合在植物中表达以改变或修饰植物性状的核苷酸序列。
作为非限制性实例,技术人员将认识到,甜菜碱醛脱氢酶基因的(如上文所述)编码seqidno:38所示氨基酸序列的seqidno:27的表达可以赋予植物增加的干旱和/或盐胁迫耐受性。
参考实施还将认识到,本文已经证明dreb1a的表达可以赋予盐耐受性,这使得能够通过在含盐培养基上再生来选择基因内转化的植物的产生。
作为另一个非限制性实例,技术人员将认识到,花色素苷1基因(如上文所述)的编码seqidno:46所示氨基酸序列的seqidno:35的表达可以增加植物的胁迫耐受性,并增加植物供人食用的营养特性。
至少在某些实施方案中,使用既赋予所需性状又能作为选择标记的用于表达的核苷酸序列,可以是非常有利的。本文已经证明,该方法可以促进基因内转化体的有效选择,而无需使用其他选择标记。
调节序列
该方面的重组基因构建体优选包含一个或多个调节核苷酸。适当地,所述一条或多条调节序列是该方面基因构建体的可插入至植物的遗传物质中的核酸片段。适当地,基因构建体的用于表达的核苷酸序列与一条或多条所述调节核苷酸序列可操作地连接。
如本文所用,“调节序列”是能够控制或以其他方式促进、赋予或修饰与调节序列可操作地连接的一条或多条其他核苷酸序列的转录和/或翻译的核苷酸序列。
“可操作地连接”或“可操作地联接”是指所述调节核苷酸序列相对于所述一条或多条核苷酸序列适当定位,以实现所述转录和/或翻译的控制或修饰。
适当地,基因构建体的额外序列的调节序列能够控制或修饰与其可操作地连接的重组基因构建体的一条或多条用于表达的核苷酸序列的转录和/或翻译。
多种调节序列是本领域技术人员已知的,并且可包括但不限于:启动子序列;前导序列或信号序列;核糖体结合位点;转录起始和终止序列、翻译起始和终止序列;增强子或激活剂序列;以及终止子序列。
优选地,所述一个或多个调节核苷酸序列包含启动子序列。
优选地,所述一个或多个调节序列包含终止子序列。
将认识到,根据本发明,可以使用促进作为非限制性实例的组成型表达;组织特异性表达;发育阶段特异性表达或诱导型表达(例如响应于环境刺激)的调节序列。
在某些优选的实施方案中,可以基于植物基因或与所述基因可操作地连接的非编码序列的内源表达,选择一株或多株植物的天然调节元件或其片段或变体,用于本发明的基因构建体。
在优选的实施方案中,调节序列包含启动子,所述启动子包含seqidnos:4-7、53、55、57、59、61、67、73、74、76、78或98所示的核苷酸序列或其片段或变体。
在优选的实施方案中,调节序列包含终止子,所述终止子包含seqidnos:8-11、54、56、58、60、62、106、108、111或112所示的核苷酸序列或其片段或变体。
其他序列
该方面的基因构建体可包含如下所述的其他核苷酸序列。将认识到,所述其他序列可以是但不必是该方面重组基因构建体的一个或多个可插入至植物的遗传物质中的核酸片段。还将认识到,所述其他序列可以是重组基因构建体的一条或多条用于表达的核苷酸序列,和/或一个或多个调节序列。
优选地,基因构建体包含含有一个或多个限制性消化位点或限制酶位点的核苷酸序列。适当地,限制性消化位点促进添加和/或去除本发明的基因构建体的核苷酸序列。
在某些特别优选的实施方案中,该方面的重组基因构建体包含可插入至植物的遗传物质中的核酸片段的侧翼序列或包含围绕可插入至植物的遗传物质中的核酸片段的侧翼序列。在一些实施方案中,所述侧翼序列或其部分衍生自一株或多株植物。优选地,所述侧翼序列包含限制性消化位点。在某些特别优选的实施方案中,一条或多条所述侧翼序列包含seqidnos:102、103、109、110、115、116、117、118、120或121所示的核苷酸序列或其片段或变体。
适当地,包含限制性消化位点的侧翼序列促进去除或切除该方面重组基因构建体的一个或多个片段,所述片段由来自较大构建体和/或载体的植物衍生的序列组成。参考实施例将认识到,作为实例,seqidnos:73-74、78、79、98和101所示的优选基因构建体包含这类促进去除重组基因构建体的由植物衍生的序列组成的片段的侧翼序列。
对于使用涉及直接转化例如粒子轰击的该方面基因构建体的转化方法,这些实施方案可能是特别期望的。将认识到,去除或切除由植物衍生的核苷酸序列组成的片段促进应用这个片段转化植物,从而预期基因构建体的非植物衍生序列不会转移到植物的遗传物质中。
此外,在某些实施方案中,本发明的基因构建体可包含一条或多条“间隔”核苷酸序列。优选地,基因构建体的作为表达的核苷酸序列或调节核苷酸序列的核苷酸序列的功能不受所述间隔序列的影响或基本上不受其影响。
作为非限制性实例,所述一条或多条间隔核苷酸序列可包含延伸的调节序列、基因间序列和/或内含子序列。重组基因构建体在任何合适的位置包含间隔序列,例如在基因构建体的多个其他额外核苷酸序列之间,但不限于此。
边界序列
在该方面的某些优选实施方案中,重组基因构建体包含作为“边界”核苷酸序列的侧翼序列。
如在本文中所用,“边界”核苷酸序列将理解为指在细菌介导的植物、植物细胞或植物组织转化过程中识别的序列。具体而言,在本发明的重组基因构建体中,边界核苷酸序列通过细菌介导的转化促进基因构建体的至少一个片段转移到植物的遗传物质中。如本领域技术人员将理解的,细菌介导的植物转化通常使用农杆菌(agrobacterium)进行。就此而言,本领域技术人员被导向bantal.m.,montenegrom.,2008,“agrobacteriumandplantbiotechnology,”于agrobacterium:frombiologytobiotechnologyeds.tzfirat.,citovskyv.,(newyork,ny:springer)。
适当地,在重组基因构建体包含边界序列的实施方案中,构建体包含第一边界核苷酸序列、第二边界核苷酸序列和一条或多条位于第一边界核苷酸序列和第二边界核苷酸序列之间的额外核苷酸序列,其中所述额外核苷酸序列和与所述额外核苷酸序列相邻的至少一部分所述第一边界核苷酸序列衍生自或可衍生自一株或多株植物。
在一些实施方案中,与所述额外核苷酸序列相邻的至少一部分第二边界核苷酸序列衍生自一株或多株植物。优选地,所述一株或多株植物是衍生所述额外核苷酸序列和第一边界核苷酸序列的至少一个片段的相同植物。
将认识到,在植物的农杆菌转化过程中,边界序列,其通常称为“右边界”(rb)和“左边界”(lb)核苷酸序列,能够将位于rb和lb之间通常称为“t-dna”的核苷酸序列插入至植物的遗传物质中。通常,所述rb和lb序列的长度约为25个核苷酸,但不限于此。
优选地,本发明基因构建体的第一边界核苷酸序列包含农杆菌rb序列。优选地,本发明基因构建体的第二边界核苷酸序列包含农杆菌lb序列。将认识到,在这些优选的实施方案中,根据这些实施方案的重组基因构建体的一个或多个额外核苷酸序列在农杆菌介导的植物转化过程中可以作为t-dna发挥作用。
还将认识到,在农杆菌介导的植物转化过程中,通常将rb序列的与t-dna序列相邻的2或3核苷酸部分插入至植物的遗传物质中(thomasandjones,2007,moleculargeneticsandgenomics278411)。例如,在拟南芥属(arabidopsis)中,整合后的rb经常(36%)在距经典t-dna插入位点的第二和第五碱基之间截短,并且对于番茄而言,rb通常在整合后保留三个或更少的碱基。
因此,如上所述,在包含边界序列的该方面的优选基因构建体中,与一条或多条额外核苷酸序列相邻的至少一部分第一边界核苷酸序列将衍生自一株或多株植物。适当地,与额外核苷酸序列相邻的至少一部分第一边界核苷酸序列的长度为至少3个核苷酸。在某些优选的实施方案中,与额外核苷酸序列相邻的至少一部分第一边界核苷酸序列包含seqidno:2或seqidno:71所示的序列。将认识到,这些序列可以衍生自任何合适的植物,并且这些序列可以形成具有所需功能的相邻较大的植物衍生的t-dna序列的一部分。
还将认识到,在大多数情况下(例如拟南芥属中76%;番茄中100%),在农杆菌介导的植物转化过程中,部分或全部lb序列本身;并且在一些情况下,lb序列上游(即朝向rb序列)高达100个核苷酸或甚至更大的序列被截短,因此不插入至植物的遗传物质中(thomasandjones,同上;brunaudetal.,2002,emborep.31152)。在一些植物中,lb序列在t-dna整合后经常被完全截短(thomasandjones,同上;番茄中98%的情况)。因此,对于包含边界序列的该方面的优选基因构建体而言,第二边界核苷酸序列的一部分衍生自一株或多株植物不是必需的。
然而,还将认识到,在农杆菌介导的植物转化过程中,在某些情况下,lb序列的一部分仍然能够插入至植物的遗传物质中。至少在某些植物中,例如拟南芥属,当在农杆菌介导的转化过程中将lb序列的一部分插入至植物的遗传物质中时,所述部分的长度通常在1个核苷酸和22个核苷酸之间(参见brunaudetal.,同上)。因此,在某些实施方案中,与额外核苷酸序列相邻的至少一部分第二边界核苷酸序列衍生自一株或多株植物。优选地,所述一部分边界核苷酸序列的长度为至少2个核苷酸。在一些实施方案中,所述一部分第二边界核苷酸序列的长度为至少22个核苷酸。
所述衍生自植物的一部分第二边界核苷酸序列的存在,在将所述边界序列的一部分插入至植物的遗传物质中的情况下是有利的,因为这会降低基因构建体的非植物衍生的任何核苷酸序列在这些情况下被插入至植物的遗传物质中的可能性。在某些优选的实施方案中,与额外核苷酸序列相邻并且衍生自一株或多株植物的至少一部分第二边界核苷酸序列包含seqidno:3或seqidno:72所示的序列。将认识到,这些序列可以衍生自任何合适的植物,并且这些序列能形成具有所需功能的相邻较大的植物衍生的t-dna序列的一部分。
在重组基因构建体包含边界序列的特别优选的实施方案中,优选重组基因构建体的一个或多个可插入至植物的遗传物质中的核酸片段,由衍生自一株或多株植物的多条长度为至少15个核苷酸或优选长度为至少20个核苷酸的核苷酸序列组成,所述一个或多个可插入至植物的遗传物质中的核酸片段由以下组成:
(i)衍生自一株或多株植物的至少一部分第一边界序列;
(ii)衍生自一株或多株植物的一条或多条额外核苷酸序列;和,任选地
(iii)衍生自一株或多株植物的至少一部分第二边界序列。
将认识到,在某些所述优选实施方案中,单个至少15个或优选至少20个核苷酸序列可以形成包含衍生自植物的核苷酸序列的一部分第一边界序列和与所述第一边界序列相邻的额外核苷酸序列。类似地,将认识到,在某些所述优选实施方案中,单个至少15个或优选至少20个核苷酸序列可以形成包含衍生自植物的核苷酸序列的一部分第二边界序列,和与所述第二边界序列相邻的额外核苷酸序列。
举例来说,在图2所示的基因构建体中,将认识到,番茄rbcs3c终止子的单个植物衍生的核苷酸序列形成了第一边界序列的衍生自植物的3-核苷酸部分和与所述第一边界序列相邻的额外核苷酸序列;并且番茄rbcs3c启动子的单个植物衍生的核苷酸序列形成了第二边界序列的衍生自植物的3-核苷酸部分和与第二边界序列相邻的额外核苷酸序列。
在重组基因构建体包含边界核苷酸序列的该方面的一些优选实施方案中,基因构建体还包含如上所述与第二边界核苷酸序列相邻的间隔序列。
如上所述,当本发明的基因构建体或其片段通过农杆菌介导的转化插入至植物的遗传物质中时,通常基因构建体的第二边界序列和朝向第二边界序列的一条或多条额外序列的至少一部分被截断,并且未插入植物的遗传物质中。
因此,间隔序列的位置与第二边界核苷酸序列相邻是有利的,因为这可以导致包含基因构建体的所有其他额外核苷酸序列的一部分所述一条或多条额外核苷酸序列插入至植物的遗传物质中,其中由所述间隔序列组成的一部分所述一条或多条额外核苷酸序列发生截短。
在重组基因构建体包含边界序列的该方面的某些优选实施方案中,基因构建体包含作为启动子序列的调节序列,其与第二边界核苷酸序列相邻并与选择标记序列可操作地连接。
如上所述,将认识到,当基因构建体或其核酸片段通过农杆菌介导的转化插入至植物的遗传物质中时,通常基因构建体的第二边界序列和基本上朝向第二边界序列的一条或多条额外序列的至少一部分被截短并且未插入至植物的遗传物质中。然而,在某些情况下,至少一部分第二边界序列可以插入至植物的遗传物质中。
因此,与选择标记核苷酸序列可操作地连接的启动子序列的位置与第二边界核苷酸序列相邻是有利的,因为这可以促进鉴定本发明产生的遗传改良植物,其中本发明的基因构建体的第二边界核苷酸序列可能已经插入至植物的遗传物质中。
特别是在本发明的第二边界核苷酸序列不包含与一条或多条核苷酸序列相邻的一部分植物衍生的核苷酸序列的实施方案中,基因构建体的插入植物中的核苷酸序列可包含第二边界序列的不是衍生自一株或多株植物的至少一个片段,如上文所述,这是不希望的。
因此,包括与相邻于第二边界序列的启动子序列可操作地连接的选择标记序列是有利的,因为所述选择标记序列在植物中的表达会指示基因构建体的第二边界核苷酸序列可能已插入植物的遗传物质中。这可以表明植物对于本发明的进一步用途可能不是期望的,或者对植物进行进一步的分析以确定不是衍生自一株或多株植物的第二边界序列的核苷酸序列是否已插入植物的遗传物质中,可能是有益的。
在一个优选的实施方案中,所述与第二边界核苷酸序列相邻的启动子序列与编码seqidno:46所示氨基酸序列或其片段或变体的选择标记核苷酸序列可操作地连接,如上文所述,该序列属于花色素苷1编码基因。
载体
根据另一方面,本发明提供了载体,其中所述载体包含上文所述的本发明的重组基因构建体。包含本发明基因构建体的载体的核苷酸序列的某些优选实例在seqidnos:47、48、63、70、82、93和95中列出。
适当地,载体还包含载体主链序列。seqidno:50示出了本发明载体的载体主链序列的一个优选实例。然而,应当认识到,可以使用一系列包含一系列合适主链序列的合适载体,如本领域所熟知。
在重组基因构建体包含边界序列的优选实施方案中,本发明的载体适于通过细菌介导的植物转化用本发明的基因构建体或其核酸片段转化植物。优选地,所述细菌介导的转化是农杆菌介导的植物转化。
如技术人员将容易地认识,通常通过'二元'载体系统促进农杆菌介导的植物转化。关于用于农杆菌介导的植物转化的二元载体系统的概述,技术人员被导向gartland&davey,1995,agrobacteriumprotocols(农杆菌实验技术)(humanapressinc.njusa);和lee,l.y.,&gelvin,s.b.,2008,plantphysiol.,146325,两者通过引用并入本文。
简而言之,二元载体通常包含如上文所述侧翼为rb和lb序列的t-dna序列,以及位于载体主链序列上、促进载体在细菌的某些常见实验室菌株(例如e.coli菌株)和农杆菌中复制和选择的其他元件。
适当地,可以将二元载体转移至包含单独的载体(通常称为“辅助质粒”)的农杆菌菌株,所述单独的载体包含促进利用农杆菌菌株通过农杆菌介导的植物转化将t-dna序列转移到植物的遗传物质的元件(通常称为“毒力”元件)。
在这些实施方案中,载体优选是二元载体。
在某些优选的实施方案中,载体的主链序列包含主链插入标记。
如本文所用,术语“主链插入标记”将理解为是指这样的核苷酸序列,其促进区分以下:使用本发明的载体转化的植物细胞、组织或植物,其中载体主链已被导入植物的遗传物质中;和利用本发明的载体转化、但载体主链并未被导入植物的遗传物质中的植物细胞、组织或植物。
将认识到,在通常情况下,由于使用本发明的载体进行的农杆菌介导的植物转化,载体主链不会转移到植物的遗传物质中。然而,在某些情况下,例如由于本发明的基因构建体的不正确加工,主链可以插入至植物的遗传物质中。还将认识到,尽管本发明的适于直接转化植物的优选基因构建体被设计为允许切除由植物衍生序列组成的用于转化的片段,但是可能的是(例如由于技术错误),载体主链可能通过直接转化并入植物的遗传物质中。
这种情况对于本发明通常是不希望的;例如,载体主链序列可包含不是衍生自一株或多株植物,和/或对于表达一条或多条额外序列是不必要或不期望的序列,所述一条或多条额外序列是本发明的基因构建体的用于在植物中表达的序列。因此,包含主链插入标记是期望的,因为根据本发明这可以允许携带载体主链序列的植物得以鉴定并避免进一步发育。
本发明的主链插入标记可以采用任何合适的形式。在某些实施方案中,主链插入标记可以促进通过施用化学品或通过视觉筛选来筛选转化的植物,类似于上文针对本发明的基因构建体的选择标记所述。
在一个优选的实施方案中,主链插入标记包含能够抑制或降低编码例如seqidno:36所示的叶绿素合酶蛋白的基因的表达的小rna的核苷酸序列。
将认识到,本发明载体的主链插入标记对编码叶绿素合酶蛋白基因在植物中表达的抑制或降低,可以允许视觉筛选使用本发明的载体转化的植物,其中叶绿素色素沉着减少或没有叶绿素色素沉着表示载体主链已插入植物的遗传物质中的转化。适当地,根据本发明可以避免这种植物进一步发育。
在某些优选的实施方案中,主链插入标记是“致死”或“阴性选择”标记。适当地,在这些实施方案中,主链插入至植物的遗传物质中的转化导致转化的植物死亡或其生长和发育基本上受到抑制。
作为非限制性实例,阴性选择主链插入标记可包含seqidno:37所示序列,或其片段或变体,所述序列属于芽胞杆菌rna酶自杀基因。
作为另一个非限制性实例,本发明载体的阴性选择主链插入标记可包含能够抑制或减少对植物的存活和/或生长和发育重要的一株或多株植物基因或非蛋白编码序列的表达或翻译的小rna序列。
宿主细胞
本发明还提供了包含本发明的基因构建体或载体的宿主细胞或有机体。所述宿主细胞或有机体可以是原核的或真核的。
在某些优选的实施方案中,所述宿主细胞可以是能够繁殖本发明的基因构建体或载体的细菌细胞(例如大肠杆菌(e.coli)细胞)。
在一个优选的实施方案中,所述宿主细胞是能够使用本发明的载体转化植物细胞的农杆菌细胞,如上所述。
在一个优选的实施方案中,所述宿主细胞是能够瞬时测试转化构建体或本发明的基因内序列的rna结合能力的植物细胞或植物组织(例如本氏烟草),如上所述。
对植物进行遗传改良的方法
本发明的另一方面提供了遗传改良植物的方法,包括将本发明基因构建体的至少一个片段或其片段引入至植物细胞或植物组织的遗传物质中的步骤。
如上所述,本发明特别希望,根据该方面的方法引入至植物细胞或植物组织的遗传物质中的基因构建体的至少一个核酸片段是,或者属于由衍生自一株或多株植物的一条或多条核苷酸序列组成的基因构建体的片段。适当地,所述引入至植物的遗传物质中的基因构建体的至少一个核酸片段由基因构建体的一个或多个片段组成,所述片段由可插入至植物的遗传物质中的、长度至少为15个核苷酸的或者优选长度为至少20个碱基对的植物衍生的核苷酸序列组成。
根据该方面特别优选的是,遗传改良的植物属于与衍生所述一条或多条核苷酸序列的一株或多株植物物种相同和/或互交可育的物种。
在一个实施方案中,该方面的方法包括以下步骤:
(i)使用本发明的基因构建体或包含本发明基因构建体的本发明的载体转化植物细胞或植物组织;和
(ii)由步骤(i)转化的植物细胞或植物组织选择性地繁殖遗传改良的植物,其中基因构建体的至少一个片段已插入至所述植物细胞或植物组织的遗传物质中。
适当地,步骤(i)所用的植物细胞或植物组织可以是叶盘、愈伤组织、分生组织、下胚轴、根、叶轴或叶轮、叶片、茎、芽(shoot)、叶柄、腋芽、茎尖(shootapex)、节间、子叶节、花梗或花序组织,但不限于此。
适当地,对于步骤(ii),作为非限制性实例,可以将转化的植物材料培养在芽诱导培养基(shootinductionmedium)中,然后培养在芽伸长培养基(shootelongationmedium)中,如本领域所熟知的。可切割芽并将其插入根诱导培养基中以诱导根形成,如本领域所熟知的。
在该方面的某些优选实施方案中,根据步骤(i)的植物细胞或植物组织转化是细菌介导的转化。特别优选的是,根据步骤(i)的植物细胞或植物组织转化是农杆菌介导的转化。
优选地,在植物细胞或植物组织的转化是细菌介导的转化的实施方案中,转化所用的基因构建体包含边界序列。优选地,本发明的载体用于所述农杆菌介导的转化。优选地,载体是如上所述的二元载体。
在某些优选的实施方案中,根据步骤(i)的植物细胞或植物组织转化是直接转化,例如本领域熟知的粒子轰击转化。本领域技术人员将了解各种植物转化方法,包括微粒轰击(franks&birch,1991,aust.j.plant.physiol.,18471;boweretal.,1996,molecularbreeding,2239;nuttetal.,1999,proc.aust.soc.sugarcanetechnol.21171)、脂质体介导的(ahokasetal.,1987,heriditas106129)、激光介导的(guoetal.,1995,physiologiaplantarum9319)、碳化硅或钨晶须介导的(美国专利号5,302,523;kaeppleretal.,1992,theor.appl.genet.84560)、病毒介导的(brissonetal.,1987,nature310511)、聚乙二醇介导的(paszkowskietal.,1984,emboj.32717)以及微注射转化(neuhausetal.,1987,theor.appl.genet.7530)和原生质体电穿孔(frommetal.,1986,nature319791),通过引用将所有上述文献并入本文。在实施方案中,该方面根据步骤(i)的转化可以通过任何上述方法进行。
在根据步骤(i)的植物细胞或植物组织转化是直接转化的实施方案中,优选地,用于转化的基因构建体包含用于切除由植物衍生的序列组成的片段的侧翼序列,如上所述,在所述片段用于转化前。
在该方面的一个优选实施方案中,如上所述,作为选择标记的本发明基因构建体的额外核苷酸序列的表达促进根据步骤(ii)的遗传改良植物的选择性繁殖。
在某些优选的实施方案中,与相应的野生型植物相比,所述选择标记核苷酸序列通过增加遗传改善植物对有毒代谢物的耐受性或增加植物利用替代营养源的能力来促进选择。在一个优选的实施方案中,所述选择标记包含编码seqidno:38所示氨基酸序列的核苷酸序列,如上所述,所述核苷酸序列属于甜菜碱醛脱氢酶基因。
在某些其他优选的实施方案中,所述选择标记序列通过赋予遗传改良植物除草剂耐受性来促进选择。在一个优选的实施方案中,所述选择标记包含编码seqidno:41所示氨基酸序列的核苷酸序列,如上所述,所述核苷酸序列属于谷氨酰胺合成酶基因。
在某些其他优选的实施方案中,所述选择标记序列通过赋予遗传改良植物耐盐性来促进选择。在一个优选的实施方案中,所述选择标记包含seqidno:119所示的核苷酸序列,如上所述,所述核苷酸序列属于dreb1a基因。
在该方面方法的某些实施例中,所述方法包括以下另外的步骤:
将另外的基因构建体的核酸片段插入至植物的遗传物质中;
由遗传物质中已经插入了第一方面基因构建体的核酸片段和另外的基因构建体的核酸片段的植物产生植物群;和
从所述植物群体中选择植物,其中所述植物的遗传物质包含第一方面基因构建体的核酸片段,但不包含另外的基因构建体的核酸片段。
优选地,插入植物的遗传物质中的另外的基因构建体的核酸片段包含选择标记核苷酸序列。
参考实施例,将认识到,这些实施方案在将另外的基因构建体的选择标记并入植物的遗传物质中对促进转化植物的初始选择是期望的情况下是特别期望的,但最终产生植物的遗传物质不含有所述选择标记的植物是期望的。
作为实例,本文已经证明,seqidno:69所示的另外构建体可以有利于根据这些实施方案促进转化体选择的用途。然而,将认识到,所述构建体适于将核酸片段并入植物的遗传物质中,其中所述片段尤其包含不属于或不衍生自一株或多株植物的nptii选择标记基因。因此,希望从根据该方面的方法最终选择的转化植物中除去该片段。
在涉及使用另外的基因构建体的一些优选的此类实施方案中,第一方面的基因构建体和另外的基因构建体属于第四方面的载体。参考实施例,这类载体包含第一方面的基因构建体和另外的基因构建体,并在seqidno:70中举例说明并给出。
在另外的或替代的此类实施方案中,另外的基因构建体属于另外的载体。
该方面的方法还可以包括选择植物的遗传物质中并未插入载体主链的遗传改良的植物的步骤。适当地,如上所述,本发明载体主链插入标记的表达促进根据该步骤选择遗传改良的植物。
在某些实施方案中,所述主链插入标记是可视标记。适当地,在这些实施方案中,当载体主链已插入植物的遗传物质中时,所述植物相对于相应野生型植物显示出可视改变。适当地,在这些实施方案中,根据该步骤仅选择不显示可视标记的植物。
在包括该步骤的一个优选实施方案中,主链插入标记包含能够抑制或降低编码例如seqidno:36所示叶绿素合酶蛋白的基因表达的小rna的核苷酸序列。适当地,根据该实施方案,当载体主链已插入植物的遗传物质中时,与相应的野生型植物相比,所述植物显示出基本上改变的叶绿素表达。适当地,根据该实施方案,根据该步骤仅选择不显示出基本上改变的叶绿素表达的植物。
在包括该另外步骤的方法的某些其他实施方案中,主链插入标记是“致死”或“阴性选择”标记。适当地,根据这些实施方案,当载体主链已插入植物的遗传物质中时,与相应的野生型植物相比,所述植物不会存活,或会显示出基本上受阻的生长和发育。适当地,根据这些实施方案,根据该步骤仅选择存活的植物和/或未显示出基本上受阻的生长和发育的那些植物。
在包括该另外步骤的一个特别优选的实施方案中,通过表达包含seqidno:37所示核苷酸序列或其片段或变体的主链插入标记,促进了该根据步骤的遗传改良植物的选择,如上所述,所述核苷酸序列属于芽孢杆菌rna酶自杀基因。
该方面的方法还可以包括鉴定遗传改良的植物的步骤,其中基因构建体的至少一部分第二边界核苷酸序列已经并入植物的遗传物质中的可能性增加。
适当地,通过表达基因构建体的额外序列来促进该步骤鉴定遗传改良的植物,所述额外核苷酸序列是与作为启动子核苷酸序列的基因构建体的额外序列可操作地连接的选择标记核苷酸序列,其中,如上所述,所述启动子序列与基因构建体的第二边界相邻。
适当地,根据该实施方案,表达选择标记核苷酸序列的植物被鉴定为基因构建体的至少一部分第二边界核苷酸序列已并入植物的遗传物质中的可能性增加。
在包括所述另外步骤的该方面方法的一个特别优选的实施方案中,通过表达包含seqidno:46所示核苷酸序列或其片段或变体的选择标记核苷酸序列来促进根据该步骤选择遗传改良的植物,如上所述,所述核苷酸序列属于花色素苷1蛋白质。
适当地,根据这些实施方案,根据该步骤鉴定与相应的野生型植物相比显示出花色素苷水平实质增加的植物。
具有改良的性状的遗传改良植物
优选地,该方面的方法包括选择遗传改良的植物的另外的步骤,所述遗传改良的植物相对于相应的野生型植物包含一种或多种改变的、修饰的或改良的性状。
优选地,根据基因构建体的适合在植物中表达以改变或修饰植物的性状的一条或多条额外核苷酸序列的表达将一种或多种性状改变。
在该方面的某些优选实施方案中,所述一条或多条核苷酸序列包含小rna核苷酸序列。
在该方面的某些优选实施方案中,所述一条或多条核苷酸序列可包含蛋白编码核苷酸序列。
可以根据该方面的方法修饰的植物性状的某些非限制性实例包括:营养品质(包括种子或谷粒质量特性和/或植物营养部分的营养质量或适口性质量);胁迫耐受性,例如非生物胁迫耐受性,例如抗干旱性,或抗盐性;植物产量(包括种子或谷粒产量和/或植物营养部分的产量);活力;植物高度(plantstature);和种子或谷粒休眠;生物胁迫抗性,例如抗病性;和营养素用途和/或效率。抗病性可包括病毒、细菌、真菌、线虫和/或昆虫抗性。
还应当认识到,所述性状可以是形态学性状,例如改善的观赏特性,或果实、叶子或任何其他植物部分的期望形状。
将进一步认识到,植物的基因内转化以表达具体的所需物质,例如在药学和/或营养学生产的背景下,可能是考虑的性状改善。
在一个优选的实施方案中,性状是抗病性性状。
在一个优选的实施方案中,性状是非生物胁迫耐受性性状。
在一个优选的实施方案中,性状是营养和/或适口性质量性状。
在一个优选的实施方案中,性状是形态学性状。
在该方面的某些优选实施方案中,通过表达一个或多个蛋白编码基因相对改善或增加或以其他方式积极改变植物的性状。参考实施例,已经证明根据该方面方法表达dreb1a可以赋予非生物胁迫耐受性,特别是耐盐性。
在该方面的某些优选实施方案中,通过表达基因构建体的作为小rna序列的一条或多条额外核苷酸序列,相对改善或增加或以其他方式积极改变植物的性状。
在优选的此类实施方案中,改善或增加植物的抗病性,其中所述小rna序列能够改变植物病原体的一种或多种核酸的表达、翻译和/或复制。
将认识到,不限于此,能够改变植物病原体的一种或多种核酸表达和/或复制的一条或多条小rna序列的表达,可以通过减弱、抑制或消除促进植物感染的植物病原体的基因或非蛋白质编码序列的表达来相对改善或增强该方面的遗传改良植物的抗病性。
将进一步认识到,不限于此,能够改变植物病原体的一种或多种核酸表达和/或复制的一条或多条小rna序列的表达,可以通过减弱、抑制或消除植物中植物病原体的复制或繁殖来相对改善或增强该方面的遗传改良植物的抗病性。
在某些优选的实施方案中,植物病原体是病毒性植物病原体。
在一个优选的实施方案中,能够改变植物病毒的一种或多种核酸表达和/或复制的一条或多条小rna序列的表达,能够减弱、抑制或消除植物中植物病毒的复制。
在某些特别优选的实施方案中,病毒性植物病原体是番茄病毒,例如黄瓜花叶病毒(cmv)和/或番茄斑萎病毒(tswv)。在某些特别优选的实施方案中,病毒性植物病原体是谷物病毒,例如高粱病毒或稻病毒。这些实施方案的特别优选的谷物植物病毒包括玉米矮花叶病毒(mdmv)、甘蔗花叶病毒(scmv)和约翰逊草花叶病毒(jgmv)。
在某些优选的实施方案中,植物病原体是细菌性植物病原体。在特别优选的此类实施方案中,细菌性植物病原体是丁香假单胞菌(pseudomonassyringae)。
在某些实施方案中,植物病原体是真菌性植物病原体。真菌性植物病原体可以是活体营养型(biotrophic)、死体营养型(necrotrophic)或半活体营养型(hemibiotrophic)真菌性植物病原体。
在其他实施方案中,通过表达基因构建体的作为小rna序列的一条或多条额外核苷酸序列,可以改善、增加或以其他方式积极改变植物的性状,其中所述小rna序列减少、抑制或去除植物内源基因的表达。
在某些优选的此类实施方案中,性状是营养和/或适口性性状。参考实施例,应当认识到,正在研究使用该方面方法的策略生产香米。
在某些优选的此类实施方案中,性状是形态学性状。参考实施例,应当认识到,正在研究使用该方面方法的策略生产“心形”番茄。
替代选择方法
尽管在该方面方法的某些优选实施方案中,如上所述,作为选择标记的本发明基因构建体的额外核苷酸序列的表达促进了根据步骤(ii)的遗传改良植物的选择性繁殖,但将认识到,另外或替代地,可以在步骤(i)中包括包含单独的选择标记的单独的选择构建体。
作为非限制性实例,合适的此类选择标记可包括赋予下述抗性的新霉素磷酸转移酶ii:卡那霉素和遗传霉素/g418抗性(nptii;raynaertsetal.,in:plantmolecularbiologymanuala9:1-16.gelvin&schilperoorteds(kluwer,dordrecht,1988),双丙氨磷/膦丝菌素抗性(bar;thompsonetal.,1987,emboj.61589),链霉素抗性(aada;jonesetal.,1987,mol.gen.genet.21086),巴龙霉素抗性(mauroetal.,1995,plantsci.11297),β-葡糖醛酸糖苷酶(gus;vancanneytetal.,1990,mol.gen.genet.220245)和潮霉素抗性(hmrorhpt;waldronetal.,1985,plantmol.biol.5103;perletal.,1996,naturebiotechnol.14624)。
如上所述,在涉及使用包含不是衍生自或不可衍生自植物的核苷酸序列的单独的选择标记的优选实施方案中,该方法包括导致最终选择遗传物质中不包含所述核苷酸序列的植物的另外步骤。
另外,将理解,根据步骤(ii)选择遗传改良的植物不一定需要使用选择标记。
例如,根据该方面产生的遗传改良植物的选择,可以通过利用一系列本领域技术人员已知方法中的任何方法,在植物的遗传物质内筛选本发明基因构建体的核苷酸序列或其片段的存在来进行。作为非限制性实例,可以利用适当的核苷酸序列特异性引物,使用southern杂交和/或pcr来检测插入根据该方面遗传改良的植物的遗传物质中的基因构建体或其片段的dna。
此外,在基因构建体包含一条或多条蛋白质编码核苷酸序列的实施方案中,可以通过例如使用对所述蛋白质具有特异性的抗体,筛选植物中由所述核苷酸序列编码的蛋白质的表达来进行根据该方面产生的遗传改良植物的选择:
(i)在elisa中,例如currentprotocolsinmolecularbiology(现代分子生物学实验技术)eds.ausubeletal.(johnwiley&sonsinc.ny,1995)第11.2章所述,其通过引用并入本文;或
(ii)通过蛋白质印迹法和/或免疫沉淀,例如currentprotocolsinproteinscience(现代蛋白质科学实验技术)eds.coliganetal.(johnwiley&sonsinc.ny,1997)的第12章所述,其通过引用并入本文。
如上所述的基于蛋白质的技术也可以见于上文的plantmolecularbiology:alaboratorymanual(植物分子生物学:实验室手册)的第4.2章,其通过引用并入本文。
还将认识到,在基因构建体包含一条或多条用于表达的核苷酸序列的实施方案中,可以通过利用例如rt-pcr(包括定量rt-pcr)、northern杂交和/或微阵列分析筛选所述核酸的表达,来对根据该方面的方法产生的遗传改良植物进行选择。
对于rna分离和northern杂交方法的实例,请技术人员参考上文plantmolecularbiology:alaboratorymanual(植物分子生物学:实验室手册)的第3章,其通过引用并入本文。southern杂交描述于例如上文plantmolecularbiology:alaboratorymanual(植物分子生物学:实验室手册)的第1章,其通过引用并入本文。
将容易地理解,尽管如本文所述的选择标记可有利于在植物转化期间增加阳性转化体的数量,但通过pcr和其他高通量类型系统(例如,微阵列、高通量测序)鉴定遗传改良的植物,由于可以容易地测试大量样品,因此可以使得能够在不使用选择标记的情况下选择遗传改良的植物。
作为非限制性实例,可以使用对转基因或其部分具有特异性的引物对数千个样品进行pcr,扩增的pcr产物可以通过凝胶电泳分离,涂布在多孔板上和/或斑点印迹到膜上,并用合适的探针杂交,例如本文所述的探针,包括放射性和荧光探针,以鉴定遗传改良的植物。
本发明的相关方面提供了根据前述方面的方法产生的遗传改良的植物。优选地,相对于相应野生型植物,所述植物具有改变或修饰的性状。
在实施方案中,根据方面的植物或根据直接前述方面的遗传改良的植物是如上所述分类为植物界(vegetabilia)的有机体。
在优选的实施方案中,所述植物是如上所述分类为原始色素体生物(archaeplastida)的有机体。
更优选地,所述植物是如上所述分类为绿色植物亚界(viridiplantae)的有机体。
甚至更优选地,所述植物是如上所述分类为有胚植物(embryophyta)的有机体。
在一些实施方案中,植物是包括微藻和大型藻类的藻类。
在一些实施方案中,植物是包括蘑菇在内的食用菌。
优选地,植物是单子叶植物或双子叶植物。
更优选地,所述植物是:禾本科(poaceae)的草本植物,例如甘蔗;棉属(gossypium)种,例如棉花;浆果,例如草莓;树,包括诸如苹果、橙等果树和诸如扁桃等坚果树;观赏植物,例如观赏性开花植物,包括诸如玫瑰等蔷薇科植物;藤本植物,包括诸如葡萄等果藤;谷物,包括高粱、稻、小麦、大麦、燕麦和玉米;豆科物种,包括诸如大豆和花生等豆类;茄科物种,包括番茄和马铃薯;十字花科物种,包括卷心菜和东方芥末;葫芦科植物,包括南瓜和西葫芦;蔷薇科植物,包括玫瑰;紫菀科植物,包括莴苣、菊苣和向日葵,或任何前述植物的亲缘植物。
在一些具体实施方案中,所述植物是番茄或番茄的亲缘植物。
在一些特别优选的实施方案中,所述植物是高粱或高粱的亲缘植物。
在一些特别优选的实施方案中,所述植物是稻或稻的亲缘植物。
实施例
实施例1.优选的基因构建体和载体
该实施例阐述了为本发明设计的某些优选基因构建体和包含这些基因构建体的优选载体的详情。
这些优选的基因构建体和载体已被涉及为通过农杆菌介导的转化促进植物的基因修饰,其中将基因构建体的由衍生自一株或多株植物的多条核苷酸序列组成的片段插入到植物的遗传物质中。所述衍生自一株或多株植物的多条核苷酸序列中的每一条的长度均为至少20个核苷酸序列。然而,将认识到,使用本文所述的基因构建体和/或载体,直接基因转移(例如通过使用基因枪法)也可用于植物转化。
基本克隆构建体和载体:番茄
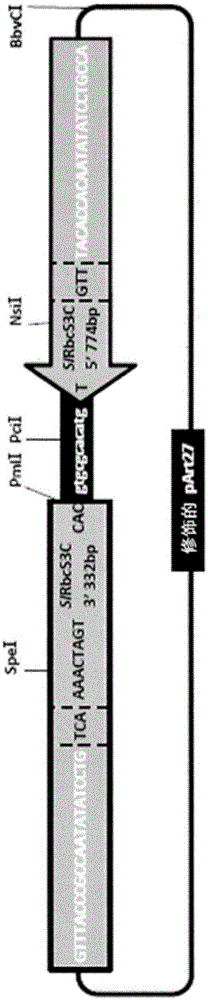
图1中显示了一个优选的基因构建体和包含该基因构建体的载体(pintr2)的示意图。该基因构建体的完整核苷酸序列示于seqidno:1所示。载体的完整核苷酸序列示于seqidno:47所示。
图1所示载体的主链序列是二元载体part27的主链序列。
所述基因构建体包含:第一边界序列,其属于农杆菌rb序列;第二边界序列,其属于农杆菌lb序列;以及多条位于rb序列和lb序列之间的额外序列。额外核苷酸序列以及rb序列和lb序列的各自部分均衍生自栽培番茄(番茄(solanumlycopersicum))。
rb序列的衍生自番茄的部分是与基因构建体的额外核苷酸序列相邻的rb序列的3个核苷酸,包含seqidno:2所示的序列。lb序列的衍生自番茄的部分是与额外核苷酸序列相邻的第二边界序列的3个核苷酸,包含seqidno:3所示的序列。
基因构建体的额外核苷酸序列包含:
(i)seqidno:4所示的调节序列,其属于番茄rbcs3c基因的启动子,与lb序列相邻;
(ii)seqidno:8所示的调节序列,其属于番茄rbcs3c基因的终止子,与于rb序列相邻;
(iii)间隔序列。
将认识到,lb序列的3核苷酸部分是(i)的番茄rbcs3c基因启动子序列的片段,使得lb序列的该部分和(i)属于单一植物衍生的核苷酸序列。
类似地,将认识到,rb序列的3核苷酸部分是(ii)的番茄rbcs3c基因终止子序列的片段,使得rb序列的该部分和(ii)属于单一植物衍生的核苷酸序列。
基因构建体的间隔序列采用与lb序列相邻的(i)的启动子核苷酸序列的“延伸”部分的形式。(i)的核苷酸序列已被设计为使得该间隔序列的截短不应实质上损害(i)的启动子功能。
该实施例的基因构建体进一步包含限制酶位点spei、pmii、pcii和nsii。限制酶位点spei属于rbcs3c终止子序列;限制酶位点nsii属于rbcs3c启动子序列。限制酶位点pcii属于核苷酸序列gtgcgcacatg(seqidno:63),位于rbcs3c启动子序列和rbcs3c终止子序列之间。限制酶位点pmli由rbcs3c终止子序列的核苷酸序列的3个碱基对(cac)和seqidno:63的三个碱基对(gtg)形成。
将理解,该基因构建体的seqidno:63非必须需要衍生自或可衍生自一株或多株植物。相反,根据本实施例的基因构建体,seqidno:63的序列和位置已被设计为通过使用上述pmli和pcii限制酶位点进行消化和连接,促进将衍生自番茄或番茄的亲缘植物的一条或多条核苷酸序列导入本实施例的基因构建体中。
将认识到,在使用pmli和pcii限制位点进行消化和连接,以及插入衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列后,从基因构建体中除去seqidno:63。
适当地,在将所述衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列导入基因构建体后,该实施例的基因构建体的片段由衍生自一株或多株植物的多条长度为至少15个或优选至少20个核苷酸的核苷酸序列组成,其中所述片段由以下组成:
(i)lb序列的3核苷酸部分,所述部分是番茄rbcs3c基因启动子序列的片段;
(ii)番茄rbcs3c基因的启动子,与lb序列相邻;
(iii)导入基因构建体的、衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列;
(iv)番茄rbcs3c基因的终止子,与rb序列相邻;和
(v)rb序列的一部分,所述部分是番茄rbcs3c基因终止子序列的片段。
图18中显示了本发明另一优选基因构建体的示意图。该基因构建体(pintra)的完整核苷酸序列如seqidno:67所示。
与pintr2类似,pintra的载体的主链序列是二元载体part27的主链序列。它是通过下述过程开发的:用asei酶从空part27去除rb和lb内的片段(以去除一些重复的限制酶位点),重新连接剩余部分并用合成序列取代bbvci和现在独特的sphi位点之间的片段,所述合成序列含有主链的除去的部分、rb、lb和番茄actin7启动子和终止子,以及它们之间的克隆位点hpai和pmli。包括被加入以在部分actin7启动子和部分actin7终止子之间产生克隆位点的核苷酸的合成片段的序列示于seqidno:67。
该基因构建体包含:第一边界序列,其属于农杆菌rb序列;第二边界序列,其属于农杆菌lb序列;以及多条位于所述rb序列和所述lb序列之间的额外序列。额外核苷酸序列以及rb序列和lb序列的各自部分都衍生自栽培番茄(番茄(solanumlycopersicum))。
rb序列的衍生自番茄的部分是与基因构建体的额外核苷酸序列相邻的rb序列的3个核苷酸,包含seqidno:2所示的序列。lb序列的衍生自番茄的部分是与额外核苷酸序列相邻的第二边界序列的5个核苷酸,包含seqidno:3所示的序列。
基因构建体的额外核苷酸序列包含:
(i)调节序列,其属于番茄actin7基因的启动子,与lb序列相邻;
(ii)调节序列,其属于番茄actin7基因的终止子,与rb序列相邻;
(iii)间隔序列。
将认识到,lb序列的5核苷酸部分是(i)的番茄actin7基因启动子序列的片段,使得lb序列的该部分和(i)属于单一植物衍生的核苷酸序列。
类似地,将认识到,rb序列的3核苷酸部分是(ii)的番茄actin7基因终止子序列的片段,使得rb序列的该部分和(ii)属于单一植物衍生的核苷酸序列。
基因构建体的间隔序列采用与lb序列相邻的(i)的启动子核苷酸序列的“延伸”部分的形式。(i)的核苷酸序列已被设计为使得该间隔序列的截短不应实质上损害(i)的启动子功能。
该实施例的基因构建体包含位于actin7启动子序列和actin7终止子序列之间的限制酶位点hpai和pmli。限制酶位点hpai由来自actin7启动子的3'碱基对(gtt)形成,并且添加三个碱基对(aac),所述三个碱基对在dna限制和插入所需dna后丢失。类似地,限制酶位点pmli由来自actin7终止子的5'碱基对(gtg)形成,并且添加三个碱基对(cac),所述三个碱基对在dna限制和插入所需dna后丢失。
将理解,根据本发明的基因构建体,seqidno:67中actin7启动子和终止子之间的seqidno:68非必须需要衍生自或可衍生自一株或多株植物。相反,根据本实施例的基因构建体,seqidno:68的序列和位置已被设计为通过使用上述hpai和pmli限制酶位点的消化和连接,促进将衍生自番茄或番茄的亲缘植物的一条或多条核苷酸序列导入本实施例的基因构建体中。
将认识到,在使用hpai和pmli限制位点进行消化和连接,以及插入衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列后,从基因构建体中除去seqidno:68。
适当地,在将所述衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列导入基因构建体后,该实施例的基因构建体的片段由衍生自一株或多株植物的多条长度为至少15个或优选至少20个核苷酸的核苷酸序列组成,其中所述片段由以下组成:
(i)lb序列的5核苷酸部分(seqidno:71),其是番茄actin7基因启动子序列的片段;
(ii)番茄actin7基因的启动子,与lb序列相邻;
(iii)导入基因构建体的、衍生自番茄或番茄的野生亲缘植物的一条或多条核苷酸序列;
(iv)番茄actin7基因的终止子,与rb序列相邻;和
(v)rb序列的一部分,其是番茄actin7基因的终止子序列的片段(seqidno:72)。
将序列克隆到pintra中使用必须与插入物互补的独特的平末端克隆限制酶位点,核苷酸连同用于扩增所述插入物的引物被添加至所述位点,并且这些引物也必须被5'磷酸化以能够进行平端连接,即:
正向引物:5'磷酸gattaaaa[起始插入序列]
反向引物:5'磷酸c[插入序列末端的反向互补序列)。
如上所述那些构建体(pinr2和pintra)一样,t-dna构建体在整合到植物基因组中时成为完全基因内的(植物基因组衍生的),当整合后仅剩下rb的5'末端的3个碱基,而lb在整合期间经常被截断(通常去除相邻序列的部分;thomasandjones,同上)。因此,相邻启动子序列经已选择为足够大,使得启动子功能不会受到损害,即使在整合期间5'末端的启动子部分被截短。
构建体和载体,具有用于表达的序列:番茄
图2示出了另一基因构建体和包含所述基因构建体的载体的示意图。
图2所示载体的主链序列是由二元载体part27的主链序列来修饰,并在seqidno:50中给出。修饰的part27主链序列包含与合适的启动子序列(例如,图2所示的camv35s启动子序列,尽管这可以根据需要改变)和合适的终止子序列可操作地连接的主链插入标记序列。
基因构建体包含:第一边界序列,其属于农杆菌rb序列;第二边界序列,其属于农杆菌lb序列;以及多条位于rb序列和lb序列之间的额外序列。额外核苷酸序列以及rb序列和lb序列的各自部分都衍生自栽培的番茄(番茄(solanumlycopersicum))或其野生亲缘植物智利番茄(solanumchilense)。
rb序列的衍生自番茄的部分是rb序列的与基因构建体的额外核苷酸序列相邻的3个核苷酸,包含seqidno:2所示的序列。lb序列的衍生自番茄的部分是与额外核苷酸序列相邻的第二边界序列的3个核苷酸,包含seqidno:3所示的序列。
基因构建体的额外核苷酸序列包含:
(i)seqidno:7所示的调节核苷酸序列,其属于番茄cyp40基因的启动子序列,与lb序列相邻并与(ii)可操作地连接;
(ii)seqidno:35所示的选择标记核苷酸序列,其属于智利番茄(solanumchilense)ant1花色素苷基因;
(iii)seqidno:11所示的调节序列,其属于番茄cyp40基因的终止子,与(ii)可操作地连接;
(iv)seqidno:5所示的调节核苷酸序列,其属于番茄actin基因的启动子序列,与(v)可操作地连接;
(v)seqidno:27所示的选择标记核苷酸序列,其属于番茄甜菜碱醛脱氢酶基因;
(vi)seqidno:9所示调节核苷酸序列,其属于番茄actin基因的终止子,与(v)可操作地连接;
(vii)seqidno:4所示的调节核苷酸序列,其属于番茄rbcs3c基因的启动子,与(viii)可操作地连接;
(viii)用于表达的核苷酸序列,其包含一条或多条够修饰植物病毒的一种或多种核酸表达和/或复制的小rna核苷酸序列;
(ix)seqidno:8所示的调节核苷酸序列,其属于番茄rbcs3c基因的终止子序列,与rb序列相邻并与(viii)可操作地连接。
将认识到,lb序列的3-核苷酸部分是(i)的番茄cyp40基因启动子序列的片段,使得lb序列部分的这部分和(i)属于单一植物衍生的核苷酸序列。
类似地,将认识到,rb序列的3-核苷酸部分是(ix)的番茄rbcs3c基因终止子序列的片段,使得rb序列的这部分和(ix)属于单一植物衍生的核苷酸序列。
(i)的序列已被设计使得cyp40启动子序列的实质性截短会消除或实质上损害(i)的启动子功能,从而(i)驱动选择标记序列(ii)表达的能力会被消除或大幅减少,所述选择标记序列(ii)属于智利番茄(solanumchilense)ant1花色素苷基因。
将理解,该实施例基因构建体的由lb和rb序列的上述3核苷酸部分和lb与rb序列之间的所有序列组成的片段,由衍生自番茄(solanumlycopersicum)或智利茄(solanumchilense)的长度为至少20个核苷酸序列的多条核苷酸序列组成。
具有用于表达的序列的构建体和载体:通用的
图3示出了另一优选的基因构建体和包含所述基因构建体的优选载体的示意图。
包含基因构建体的优选载体还包含主链序列。所述主链序列包含与合适的启动子序列(例如,图3中所示的camv35s启动子序列,尽管这可以根据需要改变)和合适的终止子序列(例如,图3中所示的ocs终止子,尽管这可以根据需要改变)可操作地连接的主链插入标记序列。如图3所示,主链插入标记是芽孢杆菌rna酶自杀基因,但是这可以根据需要而改变。
图3所示载体的基因构建体包含:第一边界序列,其属于农杆菌rb序列;第二边界序列,其属于农杆菌lb序列;以及位于rb序列和lb序列之间的多条额外序列。
额外核苷酸序列以及rb序列和lb序列的各自部分都衍生自一株或多株植物。所述植物可以是任何合适的植物。在额外序列衍生自多株植物的实施方案中,适当地,所述植物是互交可育的。
rb序列的衍生自植物的部分与基因构建体的额外核苷酸序列相邻。lb序列的衍生自植物的部分与基因构建体的额外核苷酸序列相邻。
基因构建体的额外核苷酸序列包含:
(i)调节核苷酸序列,其属于与(ii)可操作地连接的启动子;
(ii)选择标记序列。如图3所示,所述选择标记序列是花色素苷基因,但这可以根据需要而改变;
(iii)调节序列,其属于与(ii)可操作地连接的终止子;
(iv)另一种调节核苷酸序列,其属于启动子,与(v)可操作地连接;
(v)另一种选择标记序列,优选其中所述序列不同于(ii)的序列;
(vi)调节核苷酸序列,其属于终止子,与(v)可操作地连接;
(vii)调节核苷酸序列,其属于与(viii)可操作地连接的启动子;
(viii)一条或多条用于表达的核苷酸序列,其中所述核苷酸序列适合在植物中表达以改变或修饰植物的性状。
(ix)调节核苷酸序列,其属于与(viii)可操作地连接的终止子。
任选地,lb序列的衍生自一株或多株植物的部分是(i)的启动子序列的片段,使得lb序列的这部分和(i)属于单一植物衍生的核苷酸序列。
任选地,rb序列的衍生自一株或多株植物的部分是(ix)的终止子序列的片段,使得lb序列的这部分和(ii)属于单一植物衍生的核苷酸序列。
应设计(i)的序列,使得(i)的启动子序列的实质性截短会消除或实质上损害的(i)启动子功能,从而(i)驱动选择标记序列(ii)表达的能力会被消除或大幅减少。
适当地,本实施例基因构建体的由上述lb和rb序列的衍生自一株或多株植物的片段和lb序列与rb序列之间的所有序列组成的至少一片段由下述组成:衍生自(a)一株植物或(b)两个或多个互交可育植物的长度为至少20个核苷酸的多条核苷酸序列。
本实施例所述的基因构建体被设计用于转化植物,使得所述基因构建体的由衍生自一株或多株植物的多条长度为至少20个核苷酸的核苷酸序列组成的片段(或其一部分)插入至植物的遗传物质中,其中转化的植物与衍生基因构建体的所述片段的核苷酸序列的一株或多株植物相同或互交可育。
具有用于表达的序列的构建体和载体:高粱
用于高粱转化的优选方法是使用基因枪法的直接基因转移。为了确保仅使用高粱基因组衍生的序列,使用用于直接基因转移的线性dna片段在基因枪法之前可以容易地被切除的载体。这种优选的基因构建体(psbiubi1)的示意图示于图21。该基因构建体的完整核苷酸序列示于seqidno:73中。
该载体的主链序列是载体pkannibal的主链序列。其含有高粱(sorghumbiocolor)ubiquitin1基因(sobic.004g049900)的启动子和高粱ubiquitin2基因(sobic.004g050000)的终止子。其是通过利用ubi1启动子3’末端的天然psti位点开发的,所述ubi1启动子是由高粱gdna用引物f5’磷酸cctcacgtgttacacagctcaattacagactactcacc(seqidno:126)(向所述启动子的起点添加3个核苷酸,以产生能在直接基因转移之前切除基因内盒的平端切割位点(blunt-cuttersite)pmli)和rtccctgcagaagtcaccaaaataatgggt(seqidno:125)扩增的。将所述片段用psti消化,并连接到用stui和psti打开的载体pkannibal中。用引物ftccctgcagcgctaggcgccataggtcgtttaagctgctg(seqidno:127)(向所述终止子的起点添加3个核苷酸,以产生平端切割克隆位点sfoi)和rtcccactagtcacgtgtatagcacaatgcatgatcttgct(seqidno:128)(向终止子的末端添加3个核苷酸,以产生用于切除基因内盒的平端切割位点pmli和用于插入以前的载体中的spei位点)扩增终止子ubi1。将所述片段用psti和spei消化,并连接到用相同的酶打开的两个以前获得的中间载体中。
该载体(psbiubi1)适合通过用引物fctgcag[插入序列的起点]和r5'磷酸[插入序列末端的反向互补物]扩增插入物来在高粱中表达感兴趣的序列。然后将所述片段用psti消化,并连接到用psti和sfoi限制酶打开的psbiubi1中。
将认识到,在切除后,用于直接基因转移的序列由植物衍生的核苷酸序列组成。
将认识到,在使用psti和sfoi限制位点消化和连接,以及插入衍生自高粱或高粱的野生亲缘植物的一条或多条核苷酸序列后,从基因构建体中除去间隔区seqidno:75。
适当地,在将所述衍生自高粱或高粱的野生亲缘植物的一条或多条核苷酸序列导入基因构建体后,该实施例基因构建体的片段由衍生自一株或多株植物的长度为至少15个或优选至少20个核苷酸的多条核苷酸序列组成,其中所述片段由以下组成:
(i)高粱ubiquitin1基因的启动子,
(ii)导入基因构建体中的衍生自高粱或高粱野生亲缘植物的一条或多条核苷酸序列;
(iii)高粱ubiquitin1基因的终止子。
图22示出了另一种优选的基因构建体(psbiubi2)的示意图。该基因构建体的完整核苷酸序列如seqidno:74所示。
该载体的主链序列是载体pkannibal的主链序列。其含有高粱ubiquitin2基因(sobic.004g050000)的启动子和终止子序列。其是通过使用ubi2启动子3’末端的天然psti位点开发的,所述ubi2启动子是从高粱gdna用引物f5phos/cctcacgtgaggcccgtatagatgtagttaaatagctaaa(seqidno:129)(向启动子的起点添加3个核苷酸,以产生能切除基因内盒的平端切割位点pmli)和rtccctgcagaagagtcaccgaactaaagg(seqidno:130)扩增的。将所述片段用psti消化,并连接到用stui和psti消化的载体pkannibal中。如上文针对psbiubi1所述,扩增和克隆终止子ubi1。
该载体(psbiubi2)适合通过用引物fctgcag[插入序列的起点]和r5'磷酸[插入序列末端的反向互补物]扩增插入物来在高粱中表达感兴趣的序列。然后将所述片段psti消化,并连接到用psti和sfoi限制酶打开的psbiubi1中。
将认识到,在切除后,用于直接基因转移的序列由植物衍生的核苷酸序列组成。
将认识到,在使用psti和sfoi限制位点消化和连接,以及插入衍生自高粱或高粱的野生亲缘植物的一条或多条核苷酸序列后,从基因构建体中除去间隔区seqidno:75。
适当地,在将所述衍生自高粱或高粱的野生亲缘植物的一条或多条核苷酸序列导入基因构建体后,该实施例基因构建体的片段由衍生自一株或多株植物的多条长度为至少15个或优选至少20个核苷酸的多个核苷酸序列组成,其中所述片段由以下组成:
(i)高粱ubiquitin2基因的启动子,
(ii)导入基因构建体的衍生自高粱或高粱野生亲缘植物的一条或多条核苷酸序列;
(iii)高粱ubiquitin1基因的终止子。
具有用于表达的序列的构建体和载体:稻
用于稻转化的优选方法是通过利用基因枪法的直接基因转移。为了确保仅使用稻基因组衍生的序列,使用可以在基因枪法之前容易地切除用于直接基因转移的线性dna片段的载体。这种优选基因构建体(posaapx)的示意图如图23所示。该基因构建体的完整核苷酸序列如seqidno:76所示。
该载体的主链序列是载体puc57-kan的主链序列。其含有水稻(oryzasativa)apx基因的启动子和终止子序列。它是通过将合成序列seqidno:76连接到puc57-kan的切割eco53ki位点而开发的。选择apx基因启动子是因为其在整个植物中的组成型表达及其在叶中的强表达。
该载体(posaapx)适合通过用引物fgagctc[插入序列的起点]和r5'磷酸[插入序列末端的反向互补物]扩增插入物来在稻中表达感兴趣的序列。然后将所述片段用saci(或eco53ki)消化,并连接到用saci(或eco53ki)和psiii限制酶打开的posaapx1中。
将认识到,在切除后,用于直接基因转移的序列由植物衍生的核苷酸序列组成。
将认识到,在使用saci(或eco53ki)和psii限制位点消化和连接,以及插入衍生自稻或稻的野生亲缘植物的一条或多条核苷酸序列后,从基因构建体中除去间隔区seqidno:77。
适当地,在将所述衍生自稻或稻的野生亲缘植物的一条或多条核苷酸序列导入基因构建体后,该实施例基因构建体的片段由衍生自一株或多株植物的长度为至少15个或优选至少20个核苷酸的多条核苷酸序列组成,其中所述片段由以下组成:
(i)稻apx基因的启动子,
(ii)导入基因构建体的衍生自稻或稻野生亲缘植物的一条或多条核苷酸序列;
(iii)稻apx基因的终止子。
实施例2.用于本发明基因构建体中的调节序列的评估
使用基因内调节序列,例如启动子和终止子,对于在植物中实现所需的表达是重要的。例如,这可以在整个植物中实现强组成型表达,在各种植物器官或细胞类型中表达,在某些发育阶段中表达,和/或在用信号化合物(例如植物激素)诱导后表达。
除了整个植物的特异性和表达模式之外,在本发明构建体的优选实施方案中,选择来自与使用构建体的用于表达的序列相同或相关物种的基因内调节序列,例如启动子和终止子。
此外,在构建体包含边界序列并且针对农杆菌介导的转化进行优化的优选实施方案中,使用含有部分lb或rb序列的调节序列。另外,在其中针对不是农杆菌介导的转化的转化(例如直接基因转移方法)优化构建体的优选实施方案中,使用含有至少部分限制位点的调节序列,以促进在没有任何周围的非植物衍生的序列的情况下,切除待转移到植物的遗传物质中的植物衍生的片段。
对于本发明,分离了几条番茄调节序列并用报道基因例如绿色荧光蛋白(gfp)编码基因进行了测试,以研究它们作为本发明基因构建体的调节核苷酸序列的潜力。
通过gfp在番茄叶肉原生质体中的瞬时表达和稳定的农杆菌介导的番茄植物转化,测试番茄rubisco亚基3c(rbcs3c)基因启动子的示于seqidno:4的核苷酸序列以及属于相同基因的终止子的示于seqidno:8的的核苷酸序列。
在原生质体中获得了与广泛使用的花椰菜花叶病毒(camv)35s启动子驱动的gfp表达相当的强gfp表达,这证实了rbcs3c终止子的功能性(图4)。稳定转化实验的目的之一是建立rbcs3c驱动的表达模式。虽然假设由rbcs3c调节元件调节的报道基因的表达会限于植物的绿色部分,但是在根中以及叶中的一些细胞类型中观察到gfp荧光(图5)。这可以通过仅使用rbcs3c启动子的763个核苷酸这一事实来解释。
为了鉴定用于本发明基因构建体中的其他候选调节元件,有关常见番茄管家基因表达水平的信息从mascia,t.etal.,2010,molecularplantpathology,11805获得,其通过引用并入本文。
在芽和根两者中表达最高和最稳定的那些基因中actin(gi460378622)ubiquitin(gi19396)和cyclophilin(gi225312116)基因特别突出。然后在农杆菌渗入的本氏烟草叶中进行由这些调节基因驱动的gfp的瞬时表达,以评估它们调节表达的能力。
使用特异性引物通过聚合酶链反应(pcr)从番茄基因组dna(栽培品种moneymaker)扩增基因起始密码子上游约1000个核苷酸和基因终止密码子下游几个至几百个核苷酸的序列,并用作gfp构建体中的启动子和终止子。然后将gfp表达盒插入二元载体part27中,并通过包括携带phelper质粒的e.coli在内的三亲交配导入根癌农杆菌(a.tumifaciens)菌株gv3101中。将携带二元载体的过夜根癌农杆菌培养物以4000×g离心15min(分钟),并将沉淀重悬于补充有200mm乙酰丁香酮的10mm氯化镁中至od600为1.0。将悬浮液在室温下温育4小时,并使用无针注射器渗入4-6周龄的本氏烟草的幼叶中。
在渗入后3天,使用荧光显微镜观察gfp表达。所有三个启动子-终止子对都能够驱动gfp在本氏烟草中在瞬时叶农杆菌渗入测定中的表达。对于actin启动子,观察到最佳水平的gfp表达,无论是表达的亮度还是含有表达细胞的叶面积的大小(图6)。在另一个农杆菌渗入测试中,将番茄actin启动子-终止子组合的活性与番茄rbcs3c和camv35s的活性进行比较,其中actin基因调控元件在亮度和均匀性方面也表现良好,或者可能比传统使用的启动子表现得更好(图7)。
为了测试番茄actin启动子和rbcs3c终止子在稳定转化的植物中是否也表现良好,构建了插入part27中的启动子-报道基因-终止子盒。该盒含有actin7启动子、ant1基因和rbcs3c终止子(part27act:ant1:rbcs3c35s:nptii:nos)。该盒及其载体的构建已在实施例1中描述,并在图19中给出。该报道基因构建体的序列如seqidno:69所示。
接下来,通过农杆菌介导的转化(遵循(subramaniametal.,2016,plantphysiology,1701117的方法),用part27act:ant1:rbcs3c35s:nptii:nos产生番茄植物。通过定量实时pcr(qpcr)确认它们的转化状态,并通过定量实时逆转录酶pcr(qrt-pcr)确认它们的ant1表达。
如图24所示,与相应的野生型番茄植物相比,这些表达seqidno:69的植物显示出花色素苷水平增加(紫色茎、根、叶脉和部分叶)。这证明了番茄actin7启动子和rbcs3c终止子对于近组成型基因表达和图24和seqidno:69中包括的基因内盒的功能。
类似地,还确立了其他基因内植物启动子和终止子的功能。这包括稻actin1启动子与稻dreb1a终止子(见实施例7)、脱落酸(aba)生物合成基因nced3的aba诱导型启动子和终止子、r1g1b启动子和终止子,以及apx启动子和终止子(图23;seqidno:76)。测试了所有启动子和终止子在基因内构建体与稻dreb1a基因的组合(参见实施例7),所述稻dreb1a基因也作为选择标记(参见实施例3)。
稻actin1启动子被充分确立为稻的功能性组成型启动子(mcelroyetal.,1991,molecularandgeneralgenetics,231150)。对于诱导型调节序列,选择稻nced3启动子和终止子作为实例,因为相应的nced3基因是aba可诱导的。之所以选择稻r1g1b启动子和终止子,是因为预期它们在整个植物中高表达,特别是在胚乳中(parketal.,2010,journalofexperimentalbotany,612459),因此用于表达在稻粒中表达的性状(例如香米;参见实施例9,和花色素苷产生)。基于稻中预期的强组成型表达,选择稻apx启动子和终止子。对于apx、actin1/dreb1a/nced3和r1g1b,这些基因内dna片段的构建体及其序列分别如实施例2、3和9所示。
产生稻愈伤组织(水稻栽培品种reiziq),并用于对切除的线性dna进行直接基因转移(关于水稻体细胞胚发生和转化的详情,参见实施例7)。与dreb1a终止子组合的稻actin1组成型启动子的功能被证实为,9%的转化的稻愈伤组织在再生过程中在高盐度(100mmnacl)培养基上存活(参见实施例3和图28)。
稻nced3aba诱导型启动子和终止子的功能性和诱导性被证实为,19%的转化稻愈伤组织在再生过程中在高盐度(100mmnacl)培养基上存活(参见实施例3和图28)。
稻r1g1b启动子和终止子的功能性和诱导性被证实为,21%的转化稻愈伤组织在再生过程中在高盐度(100mmnacl)培养基中存活。
此外,确立了高粱基因内植物启动子和终止子的功能。这包括先前未测试的来自sobic.004g050000的高粱ubiquitin1(ubi1)启动子和终止子,以及先前使用的ubiquitin2启动子(ref),其也用ubiquitin1终止子进行了测试。这两个克隆盒的构建已在实施例2中做了描述,并分别在图21和22以及seqidno:74和seqidno:77中给出。
实施例3.天然基因作为用于转化的选择标记的用途
在植物转化过程中使用选择标记有助于有效选择转化的植物。为此目的,有利的是,本发明的基因构建体包含衍生自一株或多株植物的、为选择标记核苷酸序列的一条或多条附加核苷酸序列。
对于本发明,评估了几种天然番茄基因在本发明的基因构建体中作为选择标记核苷酸序列的潜力。
鉴定了番茄中与甜菜碱醛脱氢酶具有同源性的基因(gi209362342),其包含seqidno:27所示的核苷酸序列,并用包含该基因于35s或番茄rbcs3c启动子控制下的转基因盒,通过稳定的农杆菌介导的转化进行测试。在含有5mmba的选择性培养基上再生的芽中,18%的芽含有整合的p35s:badh盒。未获得prbcs3c:badh再生体(regenerant)。p35s:badh转化体在体外正常发育,并将其种植在土壤中,在那里它们健康生长并产生形态正常的花。
另外,在番茄中鉴定了与苜蓿和大豆细胞质谷氨酰胺合成酶1(gs1)同源的基因(gi460409536),其包含seqidno:30所示的核苷酸序列,所述核苷酸序列在氨基酸序列上与两者具有超过90%的相似性,且在编码序列上与两者具有超过80%的同一性。通过定点诱变,导入已经描述为在苜蓿(tischer,e.,etal.,同上;uspatent4975374a)中和在大豆(pornprom,t.,etal.,同上)中赋予除草剂耐受性的番茄gs1的突变。
具体而言,所产生的两个突变体是g245c(由seqidno:51所示的核苷酸序列编码)和h249y(由seqidno:52所示的核苷酸序列编码)。如上所述,将番茄gs1变体首先克隆到转基因载体,其后克隆到基因内二元载体中,在番茄rbcs3c启动子和终止子的控制下。举例来说,编码g245c变体的基因内二元载体的完整核苷酸序列如seqidno:48所示。
将用携带载体的农杆菌处理的番茄子叶外植体培养于含有1mg/l草铵膦(ga)的芽再生培养基(shoot-regeneratingmedium)上。用gs1g245c进行转基因转化再生的86%丛生芽(multipleshoots)对于标记是pcr阳性的。来自gs1h249y转化的芽再生的效率大大减少。
使用含有两个表达盒的基因内载体进行的测试转化产生旺盛的芽生长,所述表达盒的prbc3c:gs1g245c位于与左边界紧邻的启动子的起点处,与在相同的再生培养基上没有由非农杆菌共培养的对照外植体获得芽生长形成对照(图8和9)。然而,对于prbcs3c:gs1g245c盒的整合,只有一小部分芽是pcr阳性的,有更多的仅第二表达盒整合的情况。
在迄今为止转基因和基因内测试转化的两种情况下,主要由于生长模式不均匀,将最初快速形成的含有整合的prbc3c:gs1g245c的旺盛的芽再生到准备在土壤中种植的阶段存在一些困难。作为潜在的解决方案,可以测试明显重要的位置245处的不同氨基酸替代物,例如,类比天然存在于ga耐受性苜蓿中的那些的g245s或g245r,以及使用替代的天然启动子,例如,来自许多已测试过的那些启动子。
出于本发明的目的,还测试了花色素苷作为可视选择标记的有用性。如图24所示,与相应的野生型番茄植物相比,表达seqidno:69的番茄植物显示出花色素苷水平增加(紫色茎、根、叶脉和部分叶)。这原则上证明了ant1基因作为合适的可视标记基因和图24和seqidno:69中包括的基因内盒的功能。
然而,使用花色素苷作为唯一的选择标记可能是费力的,并且可能需要许多转化事件,因为对于非转化的细胞仅存在可视选择但非生理活性选择。因此,存在用转基因选择标记基因单独转化的常规选择,例如赋予庆大霉素或卡那霉素抗性的nptii基因。这种单独转化的基因盒会在随后可被删除(例如通过回交)的不同基因座上独立整合到植物的基因组中。花色素苷作为可视标记的使用在此可以极大地帮助快速筛选那些推定已经去除了选择标记的植物。为了评估该方法,如实施例1所述制备用于两种选择的构建体。在选择1中,提供具有ant1的选择标记盒作为利用共转化的单独载体(图19;seqidno:69)。在选择2中,将具有ant1的选择标记盒包括在相同质粒上,但通过提供其自身的lb和rb序列独立整合(图20;seqidno:70)。
接下来,通过农杆菌介导的转化(遵循subramaniametal.,2016,plantphysiology,1701117的方法),用与赋予期望的心形番茄性状的构建体共转化的part27act:ant1:rbcs3c35s:nptii:nos(图19),产生番茄植物(详见实施例9;图x)。通过定量实时pcr(qpcr)确认它们的转化状态,并通过定量实时逆转录酶pcr(qrt-pcr)确认它们的ant1表达。
如图25所示,用part27act:ant1:rbcs3c35s:nptii:nos共转化的番茄植物显示出强烈的花色素苷产生(左),而没有该构建体的比较植物(右)没有显示出提高花色素苷产生的可视迹象。这表明产生花色素苷的基因在与选择标记共转化时是有用的,作为用于选择标记盒在f1代中被去除的可视工具。还产生了用于在稻和高粱中产生花色素苷的基因构建体(参见实施例8),其可以用作可视选择标记。
为了开发用于基因内植物转化的另一种内源(基因内)选择标记,测试了稻dreb1a基因。为了实施这种方法,首先在稻愈伤组织上建立杀灭曲线。产生水稻栽培品种reiziq和ir64的稻愈伤组织,并如实施例7所述再生植物。ms基础培养基补充有gamborgeb5维生素、1mg.l-1naa、2mg.l-1bap、2mg.l-1激动素、3%蔗糖和7%琼脂7%,经测定其最适合作为稻再生培养基。向培养基中加入六种浓度的氯化钠(100mm、150mm、200mm、250mm、300mm和350mm)。4周后的结果显示,100mmnacl提供了充分抑制两个栽培品种(reiziq和ir64)再生的最适合的选择条件。因此,100mmnacl被认为是产生转化的稻植物的有效选择。
接下来,通过提供耐盐性,测试稻dreb1a基因与稻actin1启动子和dreb1a终止子或稻nced3启动子和终止子组合作为用于稻转化的合适选择标记。
这些完全基因内构建体通过以下方式产生:首先合成表达盒,然后由制造商(genscript)将它们插入亚克隆载体puc57-kan的ecorv限制酶位点,尽管任何其他具有平末端克隆位点的e.coli质粒会是合适的。actin1:dreb1a:dreb1a盒如图26和seqidno:78所示。nced3:dreb1a:nced3盒如图27和seqidno:79所示。在通过粒子轰击转化稻愈伤组织之前,对于actin1:dreb1a:dreb1a,使用独特的限制酶位点nhei/pmli,对nced3:dreb1a:nced3,使用独特的限制酶位点fspi,切除所述盒。
如图28所示,在用actin1:dreb1a:dreb1a盒转化的180个愈伤组织中,有9%的愈伤组织在15天后在含有100mmnacl的培养基上存活,并且在用nced3:dreb1a:nced3盒转化的300个愈伤组织中,有19%的愈伤组织在15天后在含有100mmnacl的培养基上存活,并且大部分这些愈伤组织在1个月后存活。相比之下,没有未转化的对照愈伤组织在含有100mm的培养基中存活。
这些百分比是可接受的转化效率,因此认为dreb1a基因和相应的基因内盒适合作为完全的基因内选择标记,用于本发明的构建体中以产生转化的基因内植物。
实施例4.共转化策略
与独立载体共转化
在至少某些情况下,可优选在“双载体双农杆菌菌株”共转化策略中使用如同本文所述那些的基因内构建体。在这里,构建体可以与单独的含有选择标记基因的t-dna构建体结合使用,所述选择标记基因会在不同的基因座上整合,并且可以在f1或f2代中被去除,从而留下基因组中不含外源序列的植物。
包含所述适合作为选择标记的基因构建体的这种单独的t-dna构建体和载体的示意图显示在图19中。该选择标记构建体的序列如seqidno:69所示,并且之前上文已经做了描述。应当理解,由于存在被设计成并入植物的遗传物质中的非植物衍生的调节序列和选择标记序列,该构建体本身不是本发明的优选构建体,尽管它确实与这类优选的构建体共享某些组分。
图19所示载体的主链序列是二元载体part27的主链序列。如上所述,除了选择标记基因(nptii)之外,还包括用于花色素苷生物合成的可视标记基因(ant1)以使得能够容易地去除。基因构建体包含农杆菌rb序列的序列;农杆菌lb序列的序列。位于rb和lb序列之间的是:
(i)seqidno:5所示的核苷酸序列,其属于番茄actin7基因的启动子序列,与rb序列相邻并与(ii)可操作地连接;
(ii)seqidno:35所示的核苷酸序列,其属于智利番茄ant1花色素苷基因;
(iii)seqidno:8所示的核苷酸序列,其属于番茄rbcs3c基因的终止子,与(ii)可操作地连接;
(iv)花椰菜花叶病毒双35s启动子序列的核苷酸序列,与(v)可操作地连接;
(v)新霉素磷酸转移酶ii(nptii)基因的核苷酸序列;
(vi)属于农杆菌nos基因终止子的核苷酸序列,其与(v)可操作地连接,与lb序列相邻。
(i)的序列已设计为使得actin启动子序列的实质性截短会消除或实质上损害(i)的启动子功能,从而(i)驱动选择标记序列表达的能力会被消除或实质性降低,所述选择标记序列是智利番茄ant1花色素苷基因。
或者,产生该载体的另一种形式,其中actin启动子被rbcs3c启动子(part27rbcs3c:ant1:rbcs3c35s:nptii:nos)替换。
用单个载体上的独立构建体共转化
虽然用两个载体进行的共转化是可以实现的,但在某些情况下,共转化效率可能非常低。为了避免这个问题,像本文所述的那些的基因内t-dna构建体的另一个优选用途是单载体农杆菌共转化策略。在此,两种t-dna构建体可以共同位于同一载体上。然而,由于它们各自含有它们自己的lb和rb序列,它们也产生在不同基因座整合的单独的t-dna。因此,含有选择标记基因的t-dna插入物可以在f1或f2代中被删除,从而留下在基因组中不含外源序列的植物。
包含所述基因构建体的这种双t-dna载体的示意图如图20所示。该载体的序列如seqidno:70所示。
除了选择标记基因(nptii)之外,还包括用于花色素苷生物合成的可视标记基因(ant1)以使得能够容易地去除。载体的构建如下:使用空pintra克隆载体作为模板用下述引物扩增含有番茄部分actin启动子和终止子的t-dna:正向引物(bsiwi)cgtacggaatgccagcactcc(seqidno:131)和反向引物(bsrgi)tgtacaatcgtcaacgttcacttctaaagaaatagc(seqidno:132),并通过用bsiwi酶消化插入单t-dna质粒(part27rbcs3c:ant1:rbcs3c35s:nptii:nos)中。
然后可以用5'磷酸化引物扩增所需的插入物:正向引物5'磷酸gattaaaa[起始插入序列]和反向引物5'磷酸c[插入物序列末端的反向互补物],并插入用hpai和pmli限制酶打开的所得载体中,所述酶的位点在克隆载体序列中是独特的。
实施例5.用于表达的序列,包含用于改善植物病毒抗性的小rna序列
如上所述,在某些优选的实施方案中,本发明的基因构建体包含一条或多条用于表达的核苷酸序列,其含有一条或多条小rna核苷酸序列,其中所述小rna序列能够修饰或改变植物病原体的一种或多种核酸的表达、翻译和/或复制。使用所述基因构建体进行遗传改良的植物可以表现出对诸如植物病毒等植物病原体的相对改善或增强的抗病性。
过去,开发对病毒性病原体具有改善的抗病性的遗传改良植物的方法已经使用了病毒序列衍生的抗病毒序列。这些先前的方法呈现了在感染期间与病毒基因组重组的风险,从而产生新菌株形成的可能性。实际上,这已通过实验证明,例如greene,a.e.,1993,mol.biol.22367,并被视为是一项可能导致毒力增加的病毒毒株的真正风险。
本实施例证明,衍生自植物的小rna核苷酸序列可用于改变或修饰病毒性病原体核酸的表达和/或复制。
在本实施例所述的本发明的优选实施方案中,衍生自植物的小rna序列与病毒靶标不完美匹配,并且不编码病毒功能所需的氨基酸,因此不适合病毒基因组内的有效(viable)重组事件。然而,这些小rna序列仍然能够有效地沉默这些病毒靶标的表达。
对于本实施例,产生并测试了衍生自植物序列的几条amirna序列。此外,已经产生并测试了包含衍生自植物序列的小rna序列的较长的rnai构建体。
本实施例证明,适合抑制植物病原体核酸表达和/或复制的构建体可以衍生自植物序列。预期包含此类序列的本发明的基因构建体可用于产生具有改善的抗病性的遗传改良植物。举例来说,用本发明的这种构建体转化的番茄植物表现出对cmv的改善的抗性,如实施例6所述。
amirna方法
设计并克隆天然(番茄栽培品种moneymaker)基因组衍生的人工微rna(amirna)核苷酸序列,以靶向黄瓜花叶病毒(cmv)。使用天然mirna156b(seqidno:12),向其中导入了几条番茄基因组衍生的成熟微rna序列,所述微rna序列与cmv分离株k(cmv-k)序列在对于cmv的各种分离株保守的区域中部分匹配。
使用双luc测定法测试这些amirna构建体。测试的大约25%的设计的amirna有效地起作用,例如具有seqidnos:13-18所示核苷酸序列的构建体,引起含有互补病毒靶序列的萤火虫荧光素酶的表达敲低(knockdown)(图10)。
作为另一概念验证,通过农杆菌介导的转化(遵循subramaniametal.,2016,plantphysiology,1701117的方法),使用标准二元载体(含有camv35s启动子和农杆菌ocs终止子的part27)产生表达这些amirna核苷酸序列之一(seqidno:15所示的amirna10)的番茄植物。如图11所示,这些表达seqidno:15的植物显示出对cmv的改善的抗性,显示与相应的野生型番茄植物相比cmv疾病症状降低。此外,如图12所示,与野生型植物相比,如同通过qrt-pcr评估,平均cmv病毒载量显著降低。
为了测试是否还可以靶向病毒的其他部分,以及抗性性状是否可遗传,通过农杆菌介导的转化,使用其他与上文相同的条件产生表达不同基因内amirna核苷酸序列(seqidno:16所示的amirna11)的番茄植物。在植物转化之前,如图10所示,通过本氏烟草叶子的农杆菌渗入使用瞬时荧光素酶测定。这导致cmv靶序列的显著下调,从而表明amirna11也适合在稳定转化植物中沉默这种病毒。如上所述产生t0植物,并通过定量pcr和定量逆转录酶pcr测试获得的品系,以确保转化的构建体的分别存在和表达。然后使来自两个品系(ami11-i和ami-11-ii)的植物生长至成熟,并收集来自初级转化体的种子。通过定量pcr鉴定表达纯合或杂合amirna11序列或不表达amirna11序列(不成对的)的幼苗。
当生长至2-3叶期(发芽后3周)并用cmv攻击时,两个品系的携带amirna11的纯合和杂合植物都显示出病毒抗性,而不含有amirna11的不成对植物显示出与野生型植物相似的cmv症状。这些植物的实例绘示在图29中。这与使用昆士兰农业和渔业部(departmentofagricultureandfisheries,daf)为cmv检测开发的酶联免疫吸附测定(elisa)获得的结果一致。如图30和31所示(ami11-i和ami-11-iit1后代病毒攻击测试),对于大多数测试的植物,野生型和不成对植物显示出强烈的cmv存在,而几乎所有携带amirna11构建体的植物显示存在很少或不存在elisa可检测的cmv。此外,通过daf在两个时间点(接种后3周和15周)进行cmv接种植物症状的常规严重性评分。这些数据在图32中列出,并进一步证明ami11-i和ami-11-iit1后代植物与不含ami11构建体的野生型和不成对植物相比在早期和晚期时间点都显示出抗性。另外,cmv接种的野生型植物比模拟接种的植物矮,但ami11-i和ami11-ii植物平均不比模拟接种的野生型植物矮(图32)。
果实质量和数量看起来正常,与野生型或不成对植物无法区分(图33)。来自cmv攻击的植物的果实在野生型植物中受到严重影响,但对于amirna11转化的植物显示很少或没有症状。
综合起来,这表明,植物基因组衍生的基因内小rna序列可以成功地用于产生具有正常产量和果实质量的抗病毒植物,并且这种性状可以传递给新一代。
为了进一步改善病毒抗性的耐久性,一起测试两个证明了的基于amirna的方法(amirna10和amirna11)。出于此目的,两种amirna必须由两种不同的天然番茄微rna表达。因此,分别用基因内抗cmvami10和ami11替换天然sly-mir156a和sly-mir156b微rna中的核苷酸。这种基因内双ami序列示于图34和seqidno:80中。为了测试所述构建体是否能够抑制相应病毒序列,如上所述使用利用本氏烟草植物农杆菌渗入的双荧光素酶测定。出于该测定的目的,将序列克隆到侧翼为camv35s启动子和ocs终止子的part27质粒中。如图34所示,所述构建体显著(p<0.001;学生t检验)抑制相应的cmv靶序列。
为了转化植物,在一个完全基因内构建体中组合两个已证明的基于amirna的方法(amirna10和amirna11)。如图35和seqidno:81所示,以可以与单独的选择标记构建体组合使用的作为主链的part27,产生“双载体双农杆菌菌株共转化策略”载体,所述单独的选择标记在商业化之前的后期阶段可以被去除。出于此目的,将序列(seqidno:81)插入pintra(图18;seqidno:67)中。首先合成seqidno:81,接着用f引物5’磷酸gattaaaagagcaggaaagtattgggtgagatattg(seqidno:133)和r引物5’磷酸ccgaaagaggtgaaggtgatgatca(seqidno:134)扩增,以补充actin启动子和终止子的缺失末端,随后与用hpai和pmli打开的pintra连接。通过测序测试插入物的方向。
用如上所述的这种构建体(图35)与图19和seqidno:69所示、作为单独载体的选择标记构建体一起转化番茄植物,所述单独载体也携带用于可视地识别转化的植物的番茄ant1基因。再生的植物显示出紫色的根,证实了它们的转化状态。目前正在进行双amirna表达和cmv抗性的进一步测试。
或者,使用图20和seqidno:70所示的双盒(一个载体含有两个t-dna盒的)方法。出于此目的,将双amirnat-dna盒(seqidno:81)插入part27rbcs3c:ant1:rbcs3c35s:nptii:nos(图19;seqidno:69)中。首先,使用引物:正向引物(bsiwi)cgtacggaatgccagcactcc(seqidno:135)和反向引物(bsrgi)tgtacaatcgtcaacgttcacttctaaagaaatagc(seqidno:136),由图35所示的载体扩增双amirnat-dna,然后插入用bsiwi酶打开的单t-dna质粒part27rbcs3c:ant1:rbcs3c35s:nptii:nos(图19;seqidno:69)中。目前正在进行用于该方法的番茄植物转化、再生和cmv攻击实验。用于持久的基因内cmv抗性的该双t-dna载体的基因结构和完整序列示于图36和seqidno:82中。
为了对其他病毒也应用基因内amirna方法,在番茄中产生了用于番茄斑点枯萎病毒(tswv)抗性的基因内构建体,番茄斑点枯萎病毒是另一种在全世界引起严重产量损失的病毒。与cmv类似,首先鉴定了与tswv序列匹配的足够长度的基因内序列。然后,用内源性抗tswvamirna7序列替换天然sly-mir156b微rna中的核苷酸,产生图37和seqidno:83所示的基因内序列。
为了测试构建体是否能够抑制相应病毒序列,如上所述使用利用本氏烟草植物农杆菌渗入的双荧光素酶测定。为了该测定的目的,将序列克隆到侧翼为camv35s启动子和ocs终止子的part27质粒中。如图37所示,构建体显著(p<0.001;学生t检验)抑制相应的tswv靶序列。
为了转化植物,将序列(seqidno:83)插入pintra(图18;seqidno:67)中。首先合成seqidno:83,然后用f引物5'磷酸gattaaaagagcaggaaagtattgggtgagatattg(seqidno:137)和r引物5'磷酸ccgaaagaggtgaaggtgatgatca(seqidno:138)扩增,以补充actin启动子和终止子的缺失末端。随后与用hpai和pmli打开的pintra连接。通过测序测试插入物的方向。
用如上所述的该构建体(图37)与图19和seqidno:69所示的、作为单独的载体的选择标记构建体一起转化番茄植物,所述单独的载体也携带用于可视地识别转化的植物的番茄ant1基因。对amirna7存在和表达进行的测试对于7个品系是阳性的,并且收获番茄用于收集种子。这些植物具有正常的表型,尽管它们比平时长得更高,它们以与wt相当的速率结果实并产生种子。目前正在对t1幼苗(野生型、不成对的、纯合的和杂合的)进行tswv抗性测试。
为了测试这种方法对其他作物是否也有效,还为高粱中的约翰逊草花叶病毒(jgmv)抗性、甘蔗花叶病毒(scmv)抗性和玉米矮花叶病毒(mdmv)抗性,以及稻中的水稻东格鲁杆状病毒(ricetungrobacilliformvirus,rtbv)抗性产生了基因内amirna构建体。
为了开发这种针对高粱多重病毒抗性的方法,设计了amirna,使得它们靶向多种病毒或同一病毒的多种病毒分离株。首先鉴定了与jgmv、scmv和/或mdmv在保守区域中匹配的足够长度的基因内序列。然后,用各种基因内抗病毒amirna序列替换天然高粱微rnasbi-mir156b中的核苷酸。其中一些在图38-39和seqidno:83-89中列出。合成amirna并用引物ftccctgcaggcactttgcctgaagagaggacg(seqidno:139)和r5'磷酸gctccaaatcggacagagagatgagc(seqidno:140)扩增,用psti消化并插入用psti和sfoi酶打开的载体psbiubi1(图21;seqidno:73)或psbiubi2(图22;seqidno:74)中。用pmli切割所得质粒,以获得最小的基因内转化盒。
然而,在植物转化之前,使用本氏烟草叶的农杆菌渗入测试amirna构建体。图38显示使用双荧光素酶测定成功测试了两种抗mdmv-scmvamirna构建体,所述测定导致显著(p<0.05;学生t检验)敲低mdmv-scmv靶序列。图39显示使用双荧光素酶测定成功测试了四种抗jgmvamirna构建体,所述测定导致显著(p<0.01;学生t检验)敲低jgmv靶序列。
接下来,使用用于表达的基因内psbiubi1和psbiubi2盒用上述amirna转化高粱植物(高粱(sorhumbicolor)栽培品种tx430)。切下线性基因内dna盒并用于对高粱未成熟胚进行粒子轰击。使用liu等人2014年描述的高粱转化方案(in:cerealgenomics:methodsandprotocols,methodsinmolecularbiology,r.j.henry&a.furtado(eds.),springer,newyork)。植物目前正在再生并准备用于scmv和jgmv病毒攻击。
为了在高粱中提供对jgmv的多种基因内抗性,使用三重amirna方法。如图40和seqidno:90所示,这些构建体之一在psbiubi1(seqidno:73)中含有amirna2(seqidno:86)、amirna4(seqidno:87)和amirna5(seqidno:88)。如图41和seqidno:91所示,这些构建体中的另一个在psbiubi2(seqidno:74)中含有amirna2(seqidno:86)、amirna4(seqidno:87)和amirna5(seqidno:88)。
这些构建体的克隆策略如下:用引物ftccctgcaggcactttgcctgaagagaggacg(seqidno:141)(向5'末端添加psti位点)和rgtgcactccaaatcggacagagagatgagcc(seqidno:142)(向3'末端添加apali位点)扩增amirna4。用引物fgtgcactttgcctgaagagaggacg(seqidno:143)(向5'末端添加apali位点)和raacccctaggctccaaatcggacagagagatgag(seqidno:144)(向3'末端添加avrii位点)扩增amirna5。用引物fcctaggggttttgcactttgcctg(seqidno:145)(向5'末端添加avrii位点)和r5'磷酸gctccaaatcggacagagagatgagc(seqidno:146)扩增amirna2。将所述片段用各自的酶消化,并在一个反应中连接到用psti和sfoi打开的载体psbiubi1或psbiubi2中。
为了开发用于水稻中病毒抗性的基因内amirna方法,设计了amirna,使得它们靶向水稻东格鲁球状病毒(ricetungrosphericalvirus,rtsv),一种介导由rtbv引起的症状严重性的辅助病毒。出于此目的,用各种基因内抗病毒amirna序列替换天然稻微rnaosa-mir156a中的核苷酸。这些序列之一(amirna1)如图42和seqidno:93所示。为了产生基因内稻转化盒,合成amirna1序列,并用引物fgagctcaaatgtatgtctaaccatgcacatatgg(seqidno:147)(向5’末端导入核苷酸以补足saci位点)和r5'磷酸tagtcaggaattacgaagggtgtagttatgttattc(seqidno:148)扩增。它由saci限制酶切并插入用saci和平末端切割酶psii打开的posaapx(图23;seqidno:76)中,其最后三个核苷酸有助于天然osa-mir156a折回的“延续”,与其数据库中全部序列相同。目前正在进行再稻植物中的进一步测试。
rnai方法
为了测试包含产生dsrna的核苷酸序列rna的“传统”发夹rnai构建体是否可以使用本发明的植物衍生的序列产生,设计了包含靶向cmv-k的rna序列的、跨越数百个核苷酸的长rnai构建体(seqidno:18)。通过下述方式产生基因内rnai序列:将cmv-k节段序列与番茄基因组进行比对(blasting),选择长度≥20nt(核苷酸)的最佳匹配片段并在可能的情况下将它们以小重叠排列在一起。图13显示了如何使用番茄(栽培品种moneymaker)序列并将它们集合在一起来产生seqidno:18,其中每条植物衍生的序列的长度为至少20nt(核苷酸)。该序列显示与cmv-k序列(seqidno:19-21)的总体匹配为90%。
当与三种不同的相应cmv靶序列接触时(使用双luc测定)测试该序列的rnai沉默能力。如图14所示,相对于对照,cmvrnai构建体对所有三种cmv靶标都引起强烈的表达敲低。
对于番茄转化,首先通过在有义方向上包括cmv-krnai序列(seqidno:18),其后跟随作为间隔区的pdk内含子序列和反义cmv-krnai序列,在pkannibal中建立基因内rnai构建体。然后使用saci和spei位点将所述盒转移到part27中。相应载体的完整序列如seqidno:93所示。再生植物并确认14个品系含有所述基因内构建体。这些品系具有正常表型(图14)并且目前正在进行cmv抗性测试。
为了测试是否可以使用本发明的植物衍生的序列产生其他发夹rnai构建体,设计了包含靶向tswv的rnai序列的、跨越数百个核苷酸的长rnai构建体(seqidno:94)。基因内rnai序列通过以下方式产生:将tswv-qld1节段序列与番茄基因组进行比对(blasting),选择长度≥20nt的最佳匹配片段并在可能的情况下将它们以小重叠排列在一起。图43显示了如何使用番茄(栽培品种moneymaker)序列并将它们集合在一起来产生seqidno:94,其中每条植物衍生的序列的长度为至少20nt。序列显示与tswv序列的总体匹配为91%。
当与四种不同的相应tswv靶序列接触时(使用本文所述的双luc测定),测试该序列的rnai沉默能力。如图44所示,相对于对照,tswvrnai构建体对四种靶标中的两种引起强烈的表达敲低(p<0.001;学生t检验)。
对于番茄转化,首先通过在有义方向上包括tswvrnai序列(seqidno:94),其后跟随作为间隔区的pdk内含子序列和反义tswvrnai序列,在pkannibal中建立基因内rnai构建体。然后使用saci和spei位点将所述盒转移到part27中。相应载体的完整序列如seqidno:95所示。再生植物并确认14个品系含有基因内构建体。这些品系具有正常表型,并收集番茄种子用于t1幼苗的tswv攻击测试(图44)。当通过qrt-pcr测试时,来自这些品系之一(l4)的t1幼苗显示出略微降低的tswv感染水平(图44),并且正在进行其他品系的进一步测试。
实施例6.开发快速基因内策略以在其他种类的作物植物中提供有用性状。
如本文所述,本发明的基因内构建体可适合改善作物植物的性状,例如,使用amirnas来开发番茄的抗病性。此外,本发明的构建体可以促进一种植物的性状改善,基于在另一种植物中获得的信息。举例来说,参考图16,本文描述了使用模式植物拟南芥属开发的基因内策略用于作物番茄中的评估。
在开发该策略时,假设植物病毒抗性可以通过激活植物的水杨酸(sa)途径来实现。这种途径被激活后能快速识别活体营养型病原体,通过产生活性氧发动(mount)氧化爆发,然后活性氧在感染部位导致局部过敏反应和局部程序性细胞死亡(muretal.,1997,plantj.121113)。结果,依赖活细胞的活体营养型病原体不能增殖并且植物具有抗性。然而,sa信号传导受到通常会拮抗sa途径的茉莉酸(ja)信号传导的损害,并且许多植物病原体似乎劫持并激活一种途径以危害另一种途径,并促进疾病进展(thatcheretal.,2009,plantj.58927)。因此,开发了抑制ja途径以上调sa途径的新策略,来试图诱导植物对活体营养型病原体例如病毒的抗性。
中介体(mediator)亚基控制植物的各种生理途径,本文在拟南芥属中提供的实施例表明,通过突变med18mediator(中介体)亚基基因可以实现ja信号传导的抑制和sa信号传导的同时上调。在该实施例中表明,当用芜菁花叶病毒(tumv;图16a)攻击时,具有功能失调的中介体18亚基的农杆菌介导的t-dna插入突变植物(med18)显示出病毒抗性。内源med18基因表达的改变降低ja介导的但增加sa介导的防御信号传导增加,从而导致显著的(p<0.05)病毒抗性。
应当认识到,med18或许多其他基因的突变可以以基因内的方式实现,例如通过导入仅含有内源的(基因组衍生的序列)的根癌农杆菌t-dna,如实施例3中所示。或者,如实施例4所示,可以以基因内方式使用rnai或amirna方法以抑制基因或蛋白质表达。
为了测试是否可以针对其他植物快速开发用于该有用性状(通过修饰防御信号传导的植物的病毒抗性)的策略,针对med18直系同源物(seqidno:64)的存在搜索番茄基因组。然后如实施例4中所述,通过在本氏烟草中使用农杆菌渗入,使用荧光素酶报道基因构建体瞬时基因表达测定测试两条番茄衍生的amirna序列(seqidnos:65-66)对番茄med18的抑制。如图16b所示,两种构建体均导致番茄med18的抑制,从而验证了该策略并提供了使用本发明的基因构建体来改善作物植物的抗病性(和可能的其他性状)的替代策略(参见实施例7)。
对拟南芥属med18突变体的进一步测试显示出对其他三种病毒的抗性。如图16c所示,这些包括cmv、camv和莲子草花叶病毒(alternantheramosaicvirus,altmv)。与tumv一起,这包括四种不同的病毒家族,其抗性可潜在地通过基因内方法实现。这证明了使用经过充分研究的模式植物例如拟南芥(arabidopsisthaliana)来快速开发用于作物性状的新基因内策略的强大方法。
实施例7.调节生理途径以改善对作物植物病毒的抗性
如实施例6中所述,经过充分研究的模式植物,例如拟南芥可用于在作物中开发新的基因内性状发育。植物病原体可分为两类:依赖活细胞摄取营养物质(活体营养型和半活体营养型)的植物病原体和以死亡细胞的营养物质(死体营养型)生活的植物病原体。植物病毒是专性活体营养型病原体。如实施例6中对拟南芥属所证明,感染病毒的细胞的局部程序性细胞死亡是植物防止活体营养型病原体例如不同类型的病毒对植物全身感染的合适反应(图16)。植物处理病原体的一种方法是通过在预期的感染之前调节植物防御途径来使植物做好准备。中介体亚基控制植物的各种生理途径,本文在拟南芥属和番茄植物中提供的实施例表明,通过突变或下调med18亚基基因可以实现对ja信号传导的抑制和对sa信号传导的同时上调。此外,他们证明这种方法可以导致直系同源基因的快速鉴定。
选择amirna27(seqidno:66)靶向的在番茄中针对med18鉴定的潜在直系同源物(seqidno:64),用于进一步开发用于病毒抗性的基因内性状。首先,重复对荧光素酶测定获得的实验(图16b),以进一步增加根据该方法的信心。如图45所示,amirna27显著(p<0.001;学生t检验)下调med18靶序列,这证实了先前的数据。接下来,使用subramaniam等人同上的方法,用标准二元载体(含有camv35s启动子、amirna27和农杆菌ocs终止子的part27)转化番茄植物以过表达amirna27。克隆繁殖pcr阳性系,并用qrt-pcr测试克隆的amirna27表达和med18抑制。
如图45所示,在这些植物中实现了高amirna27表达(是gapdh转录物表达的高达60倍),因此med18表达在植物中显著(p<0.05;学生t检验)下调。它们的表型外观包括伴有植物高度增加和叶子更宽的更旺盛生长(图45),同时果实大小正常但种子数量减少(参见实施例9和11)。由于模式植物(拟南芥属)中的结果预测了病毒抗性,因此开发了分离的芽测定以测试cmv抗性。从植物(野生型和med18-受损植物)分离高约15cm和处于相当发育阶段的芽。如上所述将它们用cmv机械接种,随后保存在保水装置中。在接种后2周,通过qrt-pcr定量新发育的叶中cmv存在。如图45所示,med18下调的植物显示出的cmv繁殖显著低于野生型植物,表明这些植物确实具有抗病毒性。
应当认识到,可以以基因内方式实现med18或许多其他基因的下调,例如通过导入仅含有内源(基因组衍生的序列)的根癌农杆菌t-dna,如实施例3所示。或者,可以如实施例4所示以基因内方式使用rnai方法,以抑制基因或蛋白质表达。
实施例8.使用基因内方法赋予对非病毒性病原体的抗病性
由于在实施例4-6中已经证实了各种基因内方法(amirna、rnai、途径调节)对各种病毒性病原体的抗性,因此本发明的目的是,该方法是否可以用来赋予对其他非病毒性病原体的抗性。已经使用模式植物拟南芥属阐述了这些策略之一,其中可以证明生理途径的调节可以使植物能够发展对活体营养型病原体的快速抗性。
特别是,ja防御途径的下调可以导致sa途径的上调,在一些方面,sa途径以拮抗ja信号传导的方式起作用。据信,途径之间的这种决策使得植物能够发动赋予抗性的适当途径(即抗活体营养型/半活体营养型病原体的sa途径和抗死体营养型病原体和吸吮昆虫的ja途径)。然而,似乎许多病原体通过故意诱导不适当的途径劫持这种用于植物防御信号传导的硬接线(hardwiring)。例如,半活体营养型细菌性病原体丁香假单胞菌番茄致病变种(pseudomonassyringaepvtomato)产生ja模拟物冠菌素(coronatine),其可在拟南芥属和其他植物中诱导ja防御信号途径。该途径防止或减少通常是对活体营养型病原体的最有效反应的活性氧、过敏反应和程序化细胞的产生。
因此,出于本发明的目的,测试了以基因内方式下调ja信号传导(和相关的上调的sa信号传导)是否可以赋予对除植物病毒之外的活体营养型病原体的抗性。首先,在番茄中使用注射器渗入,为丁香假单胞菌番茄致病变种开发了分离叶测定。在接种后5天,通过使用定量pcr的症状评分和病原体定量可以成功评估抗病性。接下来,将来自实施例6的野生型番茄植物和ja信号传导降低的med18-受损的番茄植物用于丁香假单胞菌番茄致病变种接种实验。
如图46所示,具有注射器渗入的丁香假单胞菌番茄致病变种的叶子在接种后5天显示出明显的病斑和黄化症状,而模拟接种的叶子没有显示黄化,尽管可能观察到一些创伤诱导的病斑。注意到在med18下调的植物中,创伤诱导的病斑明显更突出,证实这些植物具有产生更强的过敏反应的能力,导致程序性细胞死亡。这与这些植物的sa信号传导能力提高的预测性状一致。
用针对丁香假单胞菌番茄致病变种中的促旋酶编码基因的引物,通过定量pcr相对于番茄gapdh基因组序列,实现丁香假单胞菌番茄致病变种的定量。如图46所示,所有接种的叶子都增殖了丁香假单胞菌番茄致病变种,但模拟接种的叶子不含有这些细菌的可计量的量。值得注意的是,来自med18下调的植物的叶子与野生型植物相比显示出显著(p=0.011;学生t检验)减少的每植物细胞细菌,表明这种基因内方法也提供了赋予作物植物细菌抗性的有效策略。可以预期对其他活体营养型和半活体营养型病原体(例如真菌病原体镰刀菌(fusariumsp))的抗性,并且正在进行针对这些病原体的测试。
实施例9.使用基因内方法在作物植物中提供非生物胁迫耐受性
诸如盐度、干旱、高温、寒冷和洪水等作物系统中的非生物胁迫每年造成数十亿美元的产量损失。这种胁迫也严重限制了土地用于作物种植的用途,这是粮食安全和世界人口增长的一个主要问题。例如,越来越多的耕地面积受到高土壤盐度的影响,这通常是由过度灌溉做法造成的。仅在澳大利亚,据估计有12%的土地受到盐度的影响,而受干旱影响的土地甚至更多。因此迫切需要开发具有增加的非生物胁迫耐受性的作物栽培种。
稻是为数十亿人提供食物主要作物。因此,在本实施例中,开发了耐盐性稻栽培品种,其仅使用内源(基因内)基因组序列而没有外源序列。可以认识到,本实施例描述的这种完全基因内方法可以应用于其他稻栽培品种和其他重要作物。
首先确定了一种在澳大利亚和其他地方广泛用作商业作物的稻品种。在具有高产潜力的生长物中栽培品种粳稻(oryzajaponica)reiziq很受欢迎,但其缺乏对非生物胁迫,特别是低温和盐度的耐受性。因此,该品种被认为是基因内导入非生物胁迫耐受性的理想候选物,包括耐盐性的性状。通过将世界上许多受盐度影响的地区包括在内,其商业化可能会带来更广泛的种植。
出于本实施例的目的,首先必须为reiziq品种建立新的转化方案。培养基如下:愈伤组织诱导培养基包括ls基础培养基、ls维生素、500mg.l-1谷氨酰胺、50mg.l-1色氨酸、3%蔗糖、2.5mg.l-12,4-d和5%植物凝胶(phytagel)。再生培养基包括ms基础培养基、gamborgeb5维生素、1mg.l-1naa、3mg.l-1bap、1mg.l-1激动素、3%蔗糖和5%植物凝胶。选择培养基(1)包括具有200mmnacl的再生培养基。选择培养基(2)包括具有100mmnacl的再生培养基。选择培养基(3)包括具有25mmnacl的再生培养基。
种子表面灭菌方法包括将种子去壳、将去壳的种子浸泡在70%乙醇中并摇动30s(秒),然后在含有三滴tween20的4%(m/v)的次氯酸钠溶液中浸泡和摇动种子20min(分钟),然后用无菌蒸馏水漂洗种子5次以洗去漂白剂。
体细胞胚性愈伤组织诱导方法包括在层流气流中的每个皮氏培养皿中放置15-20粒种子,在愈伤组织诱导培养基中轻微推动种子,并将皮氏培养皿置于暗室中3-4周以产生体细胞胚性愈伤组织。然后将体细胞胚性愈伤组织直接用于转化或在愈伤组织诱导培养基中传代培养。发现使用14-20日龄的胚性愈伤组织进行转化是有利的。
粒子轰击和转化步骤包括通过用相应的侧翼限制位点(其剩余的核苷酸形成基因内序列的一部分)切割纯化的质粒dna,然后从进行电泳的琼脂糖凝胶中纯化片段来制备基因内dna片段。或者,可以直接使用合成的dna。使用10μl的1μg/μl线性纯化的dna用金颗粒(0.6μm直径)对胚性愈伤组织进行粒子轰击。对于用两个dna片段的共轰击,每个片段使用5μg。将至少10个微愈伤组织置于含有选择培养基(1)的平板的中心,并用基因内dna片段轰击。
选择步骤包括将平板置于黑暗中3天并将愈伤组织传代培养至选择培养基(1)。然后在10天后将健康的愈伤组织传代培养至选择培养基(2)。然后将绿色(存活的)愈伤组织传代培养至选择培养基(3)直至出现叶子。在充分形成根之后,将植物小心地转移到土壤中,并通过在植物顶部放置透明塑料容器来使之茁壮。
为了赋予稻植物耐盐性,在用限制酶nhei和pml1切割后,用seqidno:78所示的基因内dna片段actin1:dreb1a:dreb1a转化reiziq胚性愈伤组织。然后如上所述,产生并再生基因内耐盐性稻植物。如图47所示,这些稻植物能够在含有100mmnacl的培养基中生长,而没有一种对照植物在这些条件下存活。100mm的盐浓度对应于6ppt盐含量(或17%海水浓度)。目前正在对这种新的水稻品种进行试验,以测定耐盐性的最大范围以及这可以如何影响产量和谷物质量。也可以预期这些植物的其他非生物胁迫耐受性,并且为此目的计划了另外的试验。
上述新的水稻品种携带由相对强的近组成型启动子(actin1)介导的耐盐性。对于本领域技术人员而言,可能出现的问题是,由于植物需要分配额外的资源来赋予耐盐性,因此稻中dreb1a介导的途径的连续激活是否可能导致一些产量受损。为了克服这个潜在的问题,用另一种构建体试验稻转化,所述构建体包括稻aba诱导型启动子nced3。aba信号传导通常在植物的非生物胁迫期间被激活,因此,可以预期在不存在非生物胁迫的情况下植物在生长期间不使用或使用很少的资源。因此,当在无胁迫条件下使具有nced3启动子介导的aba诱导型胁迫耐受性的植物生长时,预期不会有产量受损。
为了赋予稻植物aba诱导型耐盐性,在用限制酶fspi切割后,用seqidno:79所示的基因内dna片段nced3:dreb1a:nced3转化reiziq胚性愈伤组织。然后如上所述产生并再生基因内耐盐性稻植物。如图48所示,这些稻植物也能够在含有100mmnacl的培养基中生长,而没有一种对照植物在这些条件下存活。目前计划了用这种新稻品种的试验,来测定耐盐性的最大范围。预计产量和谷物质量不会受损。也可以预期这些植物的其他非生物胁迫耐受性,并且会为此目的进行另外的试验。
可以认识到,通过使用上文的实施例所述的基因内策略,可以在水稻和其他作物植物中以基因内方式赋予耐盐性和其他非生物胁迫耐受性。
实施例10.使用基因内方法来修饰作物植物的植物结构和外观
植物结构和外观的改变是作物植物的理想性状。例如,谷物的矮化品种使更高的产量和早期收获成为可能,并成为“绿色革命”的一部分。对于许多果树而言,为了能便于收获,矮化品种也是期望的,而较高、较浓密的品种是对于诸如蓝莓等其他植物是期望的。产生多产产叶的饲料植物是期望的,较强健、较强壮的茎可以为香蕉植物提供优势以实现旋风抗性。在果实中,需要许多改良,例如果实大小增加、风味和种子减少。
如本文所述,基因内技术可提供修饰作物植物的植物结构和外观的选项。为了探索这种可能性,通过基因内amirna技术研究了一套植物中介体亚基。植物中介体提供了与植物启动子的tata盒结合的rna聚合酶ii和与通常位于tata盒上游的启动子中的其他顺式作用元件结合的转录因子之间的联系。中介体复合物由大约30个亚基组成,其中一些亚基与各种转录因子结合。因此,不同的中介体亚基为植物的各种生理途径提供信号传导和调节控制单元。在实施例6中已经针对med18-受损植物探索了该特征,所述med18-受损植物显示出ja信号传导减少和抗病毒性和细菌性病原体的生物胁迫耐受性增加。
对其表型外观进行的评估显示,如图49所示,这些植物显示出伴有植物高度增加和叶片更宽的更旺盛生长。植物产生正常大小的果实但种子数减少。在这个阶段这些植物是否显示出果实产量变化仍未经过测试,但是可以认识到,植物高度增加、叶片更宽(更繁茂)和种子负荷减少的植物可以为农民或消费者提供一些优势。
雄性细胞质不育是应该使用基因内方法调节中介体亚基的另一性状,因为这一性状对于那些可以将这些植物用作亲本系并且不希望得到的后代真实而被分型的种子公司具有商业价值。这是商业番茄品种的共同特征,要求种植者从种子公司购买种子。
为了测试番茄中其他中介体亚基的调节是否可以产生期望的植物结构性状,在番茄中鉴定了推定的med25直系同源物(seqidno:96),并且设计了用于其下调的基因内amirna(seqidno:97;图50)。如图50所示,当在本氏烟草中使用如上所述的双荧光素酶测定时,amirna6能够显著(p<0.001;学生t检验)下调番茄med25序列。将amirna9插入pintra中,并如上所述转化番茄植物。用qrt-pcr测试9个pcr阳性转化体(品系)的amirna6表达和med25抑制。如图50所示,所有9个品系都表达amirna6,并且与野生型植物相比,所有所产生的品系的med25表达均显著降低(p<0.05)。
这些植物的表型外观与野生型植物显著不同,包括矮小的植株高度、较浓密的植物、卷曲的更宽的叶子和叶子的黄色斑点。这表明本发明所述的基因内方法可用于改变植物结构和外观。可以理解,通过使用上述实施例所述的基因内策略,可以在番茄和其他作物植物中以基因内方式赋予改变的植物结构和外观。
实施例10.改善作物植物的营养价值
作为食物来源的植物的营养价值无疑是获得消费者高度赞赏的性状。具有改善的营养价值的基因内植物因此提供直接的消费者益处并且可能容易被接受。营养增强的植物可包括具有较高蛋白质、维生素、矿物质、抗氧化剂、多不饱和脂肪酸水平的植物。被强调有益于消费者健康的一个特定营养方面是水果和蔬菜中的花色素苷含量。这些花色素苷含量增加的“超级食品”中的一些包括蓝莓、紫萝卜、甜菜根和超级李子(queengarnetplum)。值得注意的是,消费食物中较高的花色素苷含量已导致血压降低和其他心血管和预防癌症的益处。
为了本发明的目的并且为了增加粮食作物的营养价值,产生了所含的花色素苷水平比野生型植物高的番茄和稻植物。如前所述,用如seqidno:69所示的构建体转化番茄植物,所述构建体包括侧翼为天然actin启动子和rbcs3c终止子的番茄ant1基因。使植物在温室中生长直至座果期并评估其果实颜色。如图51所示,新出现的番茄果实具有明显的紫色外观,表明其花色素苷含量高。
此外,为了改善商业上广泛消费的主要粮食作物的营养价值,产生了新的reiziq稻栽培品种的植物,其携带增加稻粒花色素苷水平的完全基因内盒。如上所述,用seqidno:98所示的基因内构建体转化稻栽培品种reiziq植物,所述基因内构建体除了actin1:dreb1a:dreb1a盒以外还包括侧翼为天然r1g1b启动子和终止子的稻osb2基因。在粒子轰击之前,通过用fspi和apali限制酶切割来切除基因内osb2盒并纯化。选择稻r1g1b启动子和终止子盒,因为相应基因在成熟稻粒的胚乳中表达最强。如图51所示成功产生了植物,并且目前使之生长至成熟以测量稻粒中的花色素苷水平。预计消费者对这些植物的接受度会很高,因为这些植物提供直接的消费者益处并且是完全基因内的。此外,它们可能显示出由基因内dreb1a盒介导的改善的非生物胁迫耐受性,这可有益于该品种的种植者。随着这些植物被整合到育种计划中,可以预期未来与其他品种的杂交。
可以认识到,通过使用上述实施例所述的基因内策略,可以在番茄、稻和其他作物植物中以基因内方式赋予较高的花色素苷水平和其他改善的营养价值。
实施例11.其他对消费者友好性状
如果消费者体验了对现有植物产品的改善,那么新作物品种的益处可能会受到消费者的最好领会。出于本发明的目的并且证明通过为消费者体验提供直接益处使本专利中提出的基因内技术有用,产生了两种改良的作物品种。这些包括心形西红柿和香米。
心形西红柿可能会基于其颜色和原始形状而受到消费者的欢迎。由于它们有增强消费者体验的潜力,因此该产品有潜在的市场。香(茉莉)米已经受到消费者的欢迎,他们根据烹饪后释放的挥发物准备为这种米饭支付更高的价格。因此,选择这些对消费者友好的性状作为本发明描述的基因内技术的实例。
通过rnai介导的编码b型异源三聚g蛋白(ggb1)的γ-亚基的番茄基因的下调生成产生心形番茄的植物。最近已经针对microtom番茄描述了以转基因方式下调该基因,其中它导致尖果(subramaniametal.,同上)。该基因的转录物序列如seqidno:99所示。
为了在actin启动子-终止子表达盒(pintraa)中产生基因内rnai构建体,首先用补充actin启动子末端的f引物5'磷酸gattaaaatacaaatcgatctccatttcctccatc(seqidno:149)和添加三个核苷酸以产生临时mfei限制酶位点的r引物tcccaattgtcaagttgaaacaattttttgtgcatataac(seqidno:150)扩增长的“正向”片段。用添加三个核苷酸以产生临时mfei限制酶位点的f引物tcccaattgggaagtgtatgagttacaaaacatacttacct(seqidno:151)和补充actin终止子的起点的r引物5'磷酸ctacaaatcgatctccatttcctccatc(seqidno:152),扩增较短的“反向”片段。用mfei限制酶切所述片段,并在一次连接中与用hpai和pmli打开的pintra组装。由于mfei位点连接在长片段和短片段之间,其中一半属于长片段,另一半属于短片段。验证用于与启动子和终止子互补的插入物的方向。包括lb和rb片段的完整基因内构建体如seqidno:100和图52所示。
如上文实施例所述,通过将seqidno:100中的构建体与含有用于选择转化的植物的ant1和nptii基因的标记基因盒共转化,进行番茄转化(栽培品种moneymaker)。选择紫色植物(表明它们的阳性转化状态)并通过qrt-pcr进一步测试其基因表达。还选择了不表达ant1基因的其他植物。由这些植物产生的番茄果实预期具有尖的心形的外观,分别具有紫色或红色果色。
稻是主要的粮食作物。为了以基因内方式开发消费者友好的,从目前不具有该性状的流行的澳大利亚品种(reiziq)开发了高香味稻栽培品种。可认识到,本发明中描述的实现该性状的基因内方法可以应用于其他稻栽培种以及可能的其他重要作物。
栽培品种粳稻reiziq在具有高产潜力的生长物中很受欢迎,但缺乏通常见于茉莉(芳香)稻中的香味。通过破坏稻badh2基因的表达可以实现稻的香味。因此,构建了在稻胚乳中表达的具有内源r1g1b启动子和终止子的badh2rnai盒。完整的盒如seqidno:101所示。使用fspi限制酶和琼脂糖凝胶电泳大小片段化,在粒子轰击稻愈伤组织之前切除该dna盒。图53展示了正在发育的具有潜在香味的基因内稻植物。
在整个说明书中,目的是描述本发明的优选实施方案,而不是将本发明限制于任何一个实施方案或特征的具体集合。在不脱离本发明的情况下,可以对所描述和示出的实施方案进行各种改变和修改。
本说明书中提到的每一专利和科学文献、计算机程序和算法的公开内容通过引用整体并入。
- 还没有人留言评论。精彩留言会获得点赞!