癌症的联合治疗的制作方法
本申请要求2016年8月12日提交的美国临时专利申请第62/374,621的优先权和权益,通过引用将该申请的内容纳入本文。
发明领域
本发明总体涉及用于治疗癌症的联合疗法,尤其以下所述的联合:(i)双功能分子,其包含tgfβrii或其能够结合tgfβ的片段和结合免疫哨点蛋白例如程序性死亡配体1(pd-l1)的抗体或其抗原结合片段和(ii)至少一种其他抗癌治疗剂。抗癌治疗剂包括例如辐照、化疗药、生物制剂或疫苗。在本发明的某些实施方式中,联合治疗提供协效性抗癌效果。
背景
在癌症治疗中,一直认为化疗与高毒性相关联,并且会导致抗性癌细胞变体的出现。大多数化疗药有副作用,包括心脏毒性和肾毒性,脱发,恶心和呕吐。癌症治疗还包括放射疗法。此类治疗采用高能粒子或波例如x射线、γ射线、电子束或质子来摧毁或破坏癌细胞。不同于全身暴露于抗癌药物的化疗,放射疗法更常见的是局部治疗。然而,很难仅对异常组织选择性地施以治疗性辐照,因此,在整个治疗过程中,异常组织附近的正常组织也暴露于潜在的破坏性剂量的辐照。
癌症的免疫疗法是癌症治疗中的新典范,它不是靶向癌细胞而是聚焦于免疫系统的激活。其原理是重新激活宿主的免疫反应,尤其是过继性t细胞反应,提供免疫监控来杀死癌细胞,特别是逃脱了其他形式治疗的微小残留病变,由此实现持久的保护性免疫。
2011年,fda批准了抗ctla-4抗体伊匹单抗(ipilimumab)用于治疗黑素瘤,由此引领了癌症免疫治疗的新纪元。临床试验中抗pd-1或抗pd-l1疗法显示在黑素瘤、肾癌和肺癌中诱导持久反应,这进一步表明属于它们的时代的到来(pardoll,d.m.,natimmunol.2012;13:1129-32)。然而,伊匹单抗治疗受其毒性特征的限制,可能是因为抗-ctla-4治疗通过干扰初始t细胞抑制性哨点可能引起新自身反应性t细胞的产生。尽管抑制pd-l1/pd-1相互作用会将耗竭性t细胞(大多数本身是抗病毒或抗癌的)中的现存慢性免疫应答去抑制(wherry,e.j.,natimmunol.2011;12:492-9),但抗pd-1治疗有时会导致可能致命的肺相关自身免疫不良事件。且不论目前抗-pd1和抗-pd-l1颇具临床前景的活性,提高治疗指数,不论是通过提高治疗活性还是降低毒性或两者兼具,仍然是免疫治疗剂开发的中心目标。
发明概述
本发明是基于一项癌症联合治疗的发现,其包括给予双功能蛋白,所述双功能蛋白包含至少一部分tgfβ受体ii(tgfβrii)和抗体或抗原结合片段,这至少一部分tgfβ受体ii能够结合tgfβ,而所述抗体结合免疫哨点蛋白,如人类蛋白质程序性死亡配体1(pd-l1)。联合疗法还包括给予抗癌治疗剂,例如辐照、化疗药、生物制剂和/或疫苗。与各个药剂分别施用的效果相比,联合疗法表现出协同效应。
因此,在第一方面中,本发明包括一种治疗对象癌症的方法,其包括(i)施用包含人tgfβrii或其能够结合tgfβ的片段(如可溶性片段)和结合pd-l1的抗体或其抗原结合片段(如本文所述的任何抗体或抗体片段)的双功能分子;和(ii)给予至少一种其他的第二种抗癌治疗剂。
某些实施方式中,本发明的联合治疗方法包括以下所述多肽与至少一种其他抗癌治疗剂的联用,所述多肽包括(a)至少pd-l1结合性抗体的重链可变域(如seqidno:2的氨基酸1-120);和(b)人tgfβrii或其能够结合tgfβ的可溶性片段(如人tgfβrii胞外域(ecd),seqidno:9的氨基酸24-159,或任何本文所述的片段)。所述多肽可以进一步包括连接可变结构域c端与人tgfβrii或其能够结合tgfβ的可溶性片段的n末端的氨基酸接头。所述多肽可包含seqidno:3所示氨基酸序列或与seqidno:3基本相同的氨基酸序列。抗体片段可以是scfv、fab、f(ab’)2或fv片段。
某些实施方式中,蛋白质或多肽包含含有seqidno:2的抗体或其抗原结合片段和人tgfβrii。所述抗体可任选地包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可任选地包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中氨基酸1-120的抗体或其抗原结合片段和人tgfβrii。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中氨基酸1-120的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中氨基酸1-120的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中超变区的抗体或其抗原结合片段和人tgfβrii。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中超变区的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:2中超变区的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含:含有seqidno:12的抗体或其抗原结合片段和人tgfβrii。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:12的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:12的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:12中超变区的抗体或其抗原结合片段和人tgfβrii。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:12中超变区的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包含c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:12中超变区的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含:含有seqidno:14的抗体或其抗原结合片段和人tgfβrii。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:14的抗体或其抗原结合片段和能够结合tgfβ的人tgfβrii的片段(例如可溶性片段)。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
某些实施方式中,所述蛋白质或多肽包含含有seqidno:14的抗体或其抗原结合片段和人tgfβriiecd。所述抗体可包含经修饰的恒定区(例如本文所述的各种,包括c末端lys→ala取代,leu-ser-leu-ser(seqidno:19)突变为ala-thr-ala-thr(seqidno:20),或包含igg1铰链区和igg2ch2结构域的杂交恒定区)。
本发明还提供用于本发明所述联合疗法的如下所述蛋白质,该蛋白质包括前文所述的多肽和至少一个抗体轻链可变域,所述抗体轻链可变域与所述多肽组合时形成pd-l1结合性抗原结合位点。所述蛋白质可包含:(a)两个多肽,各自具有由seqidno:3的氨基酸序列组成的氨基酸序列,和(b)两个其他多肽,各自具有由seqidno:1的氨基酸序列组成的氨基酸序列。
本发明的特征包括一种癌症的联合治疗,包括给予前文所述蛋白质并联合给予一种或多种其他抗癌治疗剂来用于治疗癌症或用于抑制肿瘤生长。所述一种或多种其他抗癌治疗剂包括辐照、化疗药、生物制剂和/或疫苗。
所述癌症或肿瘤可选自:结直肠癌、乳腺癌、卵巢癌、胰腺癌、胃癌、前列腺癌、肾癌、宫颈癌、骨髓瘤、淋巴瘤、白血病、甲状腺癌、子宫内膜癌、子宫癌、膀胱癌、神经内分泌癌、头颈部癌、肝癌、鼻咽癌、睾丸癌、小细胞肺癌、非小细胞肺癌、黑素瘤、基底细胞皮肤癌、鳞状细胞皮肤癌、隆突性皮肤纤维肉瘤、梅克尔细胞癌、成胶质细胞瘤、胶质瘤、肉瘤、间皮瘤,和骨髓增生异常综合征。
本发明的特点还包括抑制肿瘤生长或治疗癌症的联合疗法。所述方法包括使肿瘤暴露于如上前文所述蛋白质。所述方法还可包括使肿瘤暴露于辐照或化疗药、生物制品或疫苗的施用。在某些实施方式中,所述肿瘤或癌症选自:结直肠癌、乳腺癌、卵巢癌、胰腺癌、胃癌、前列腺癌、肾癌、宫颈癌、骨髓瘤、淋巴瘤、白血病、甲状腺癌、子宫内膜癌、子宫癌、膀胱癌、神经内分泌癌、头颈部癌、肝癌、鼻咽癌、睾丸癌、小细胞肺癌、非小细胞肺癌、黑素瘤、基底细胞皮肤癌、鳞状细胞皮肤癌、隆突性皮肤纤维肉瘤、梅克尔细胞癌、成胶质细胞瘤、胶质瘤、肉瘤、间皮瘤,和骨髓增生异常综合征。
“tgfβrii”或“tgfβ受体ii”表示:具有野生型人2型tgfβ受体同种型a序列(如ncbi参考序列(refseq)登记号np_001020018的氨基酸序列(seqidno:8))的多肽,或具有野生型人2型tgfβ受体同种型b序列(如ncbirefseq登记号np_003233的氨基酸序列(seqidno:9))或具有与seqidno:8或seqidno:9的氨基酸序列基本相同序列的多肽。tgfβrii可保留野生型序列tgfβ结合活性的至少0.1%、0.5%、1%、5%、10%、25%、35%、50%、75%、90%、95%或99%。表达的tgfβrii多肽没有信号序列。
“能够结合tgfβ的tgfβrii片段”表示ncbirefseq登记号np_001020018(seqidno:8)或ncbirefseq登记号np_003233(seqidno:9)或与seqidno:8或seqidno:9基本相同序列的任意部分,片段长度至少20(例如至少30、40、50、60、70、80、90、100、110、120、130、140、150、160、175或200)个氨基酸且保留野生型受体或其野生型片段的至少部分tgfβ结合活性(例如至少0.1%、0.5%、1%、5%、10%、25%、35%、50%、75%、90%、95%或99%)。通常,这些片段是可溶性片段。示例性片段之一是具有seqidno:10序列的tgfβrii胞外域。
“基本相同”表示多肽与参照氨基酸序列显示至少50%,优选60%、70%、75%或80%,更优选85%、90%或95%,且最优选99%的氨基酸序列相同性。比较序列的长度一般至少10个氨基酸,优选至少15个连续氨基酸,至少20、25、50、75、90、100、150、200、250、300或350个连续氨基酸更好,最好是全长氨基酸序列。
“患者”表示人或非人动物(例如哺乳动物)。
“治疗”患者的疾病、异常或症候(如癌症)表示通过给予患者治疗剂来减轻或减少疾病、异常或症候的至少一种症状。
“癌症”指异常增殖的细胞集群。
本发明的其他实施方式和详细内容可见后文。
附图简要说明
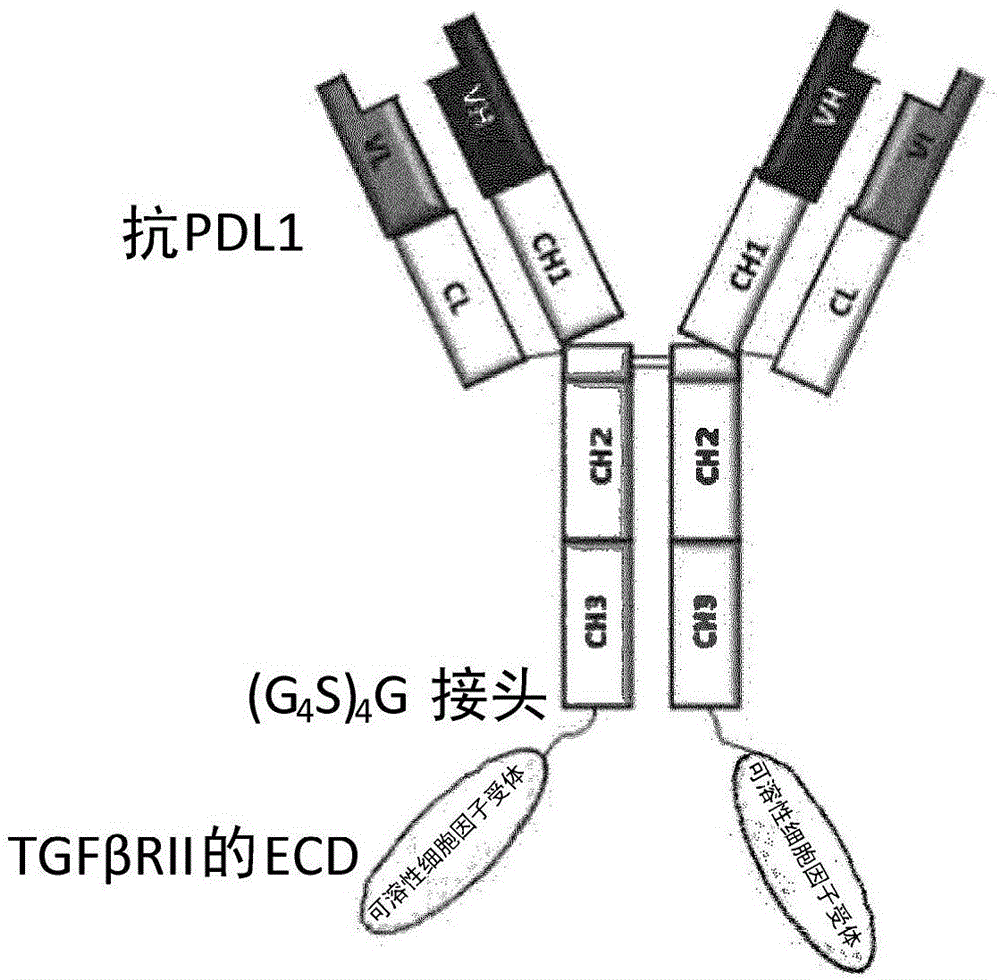
图1是抗pd-l1/tgfβ阱(tgfβtrap)分子的示意图,该分子包含一个抗pd-l1抗体与两个tgfβii型受体胞外域(ecd)通过(gly4ser)4gly接头融合。
图2:表中汇总了“ti13-027:c57b/l6野生型小鼠mc38肿瘤模型中抗pd-l1/tgfβ阱与5-fu和奥沙利铂的联合治疗”的研究设计,其中分组处理是n=10个小鼠/组。
图3:表中汇总了“ti14-012:b细胞缺陷型小鼠mc38肿瘤模型中抗pd-l1/tgfβ阱与5-fu和奥沙利铂的联合治疗”的研究设计,其中分组处理是n=10个小鼠/组。
图4a-4d的一系列图表显示奥沙利铂/5-fu与抗pd-l1/tgfβ阱联合增强肿瘤生长抑制和肿瘤反应性cd8+t细胞反应(c57bl/6小鼠,研究ti13-027)。图4a和图4d:整个研究期间每周两次测定肿瘤体积。将肿瘤体积数据转换为对数(log)并进行双向重复anova分析:图4b。对肿瘤重量数据进行单向anova分析:图4c。用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。对elispot数据进行单向anova分析。所有anova包括tukey多重比较修正以测定处理组之间的统计学差异。p<0.05认为具有统计学显著性。
图5a-5d的一系列图表显示奥沙利铂/5-fu与抗pd-l1/tgfβ阱联合增强肿瘤生长抑制和肿瘤反应性cd8+t细胞反应(b6.129s2-ighmtm1cgn/j鼠,研究ti14-012)。图5a和4d:将肿瘤体积数据转换为对数(log)并进行双向重复anova分析。图5b:对肿瘤重量数据进行单向anova分析。图5c:用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。对elispot数据进行单向anova分析。所有anova包括tukey多重比较修正以测定处理组之间的统计学差异。p<0.05认为具有统计学显著性。
图6a-6c的一系列图表显示辐照与抗pd-l1/tgfβ阱联合诱导肿瘤生长抑制和肿瘤反应性cd8+t细胞反应的协效性效果(ti13-109)。图6a:每周两次测定肿瘤体积,平均肿瘤体积表示为平均值±平均值标准误差(sem)。图6b:第14天测定肿瘤重量。图6c:在第14天用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。抗pd-l1/tgfβ阱在164μg剂量水平的数据与55μg剂量水平的数据相似,不论是作为单一疗法还是联合疗法。
图7a-7c的一系列图表显示辐照与抗pd-l1/tgfβ阱联合诱导肿瘤生长抑制和肿瘤反应性cd8+t细胞反应的协效性效果(重复研究)(ti14-013)。图7a:每周两次测定肿瘤体积,平均肿瘤体积表示为平均值±平均值标准误差(sem)。图7b:第14天测定肿瘤重量。图7c:在第14天用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。
图8a-8d的一系列图表显示辐照与抗pd-l1/tgfβ阱促进肿瘤浸润性cd8+t细胞和nk细胞(ti14-013)图8a:肿瘤浸润性cd8+tils。图8b:肿瘤浸润性nk1.1+tils。图8c:cd8+tileomes表达。图8d:cd8+til脱粒。
图9a是示意图,显示对带有原发和继发肿瘤的小鼠施以辐照,为的是检测远位效应(abscopaleffect)。
图9b的折线图显示自治疗开始按日测量的小鼠内原发肿瘤体积。
图9c是折线图显示自治疗开始按日测量的小鼠内继发肿瘤体积(mm3)。(●=同型对照400μg;◆=抗pdl1-tgfβ阱μg;■=辐照500拉德(rad);▼=辐照+抗pdl1-tgfβ阱)。
具体说明
本发明总体涉及用于治疗癌症的联合疗法,尤其以下所述的联合:(i)双功能分子,其包含tgfβrii或其能够结合tgfβ的片段和结合免疫哨点蛋白例如程序性死亡配体1(pd-l1)的抗体或其抗原结合片段和(ii)至少一种其他抗癌治疗剂。抗癌治疗剂包括例如辐照、化疗药、生物制剂和或疫苗。本发明的某些实施方式中,联合治疗提供协效性抗癌效果。
本发明的联合疗法特别有利,因为与每种物质各自单用的效果相比,不仅抗癌作用增强,而且联合疗法中的一种或多种物质相比各自单一疗法剂量减少但仍能达到总体抗癌效果。由于协同效应,可以有利地减少给予患者的药物总量,从而减少副作用。
本发明的联合疗法能够实现肿瘤微环境中tgfβ的局部减少,这是通过用与抗体部分相连的可溶性细胞因子受体(tgfβrii)来捕获tgfβ,所述抗体部分靶向某些肿瘤细胞或免疫细胞外表面上的细胞免疫哨点受体。本发明抗体部分的一个例子是针对免疫哨点蛋白的如抗pd-l1。该双功能分子,有时在本文件中亦称“抗体-细胞因子阱”,是确实有效的,因为抗受体抗体与细胞因子阱物理连接。由此产生的优势(例如相对于所述抗体和所述受体分开给予)部分归因于细胞因子通过自泌和旁泌作用成为局部环境中的主要作用。抗体部分将细胞因子阱引向肿瘤微环境,细胞因子阱在此可通过中和局部免疫抑制性自泌或旁泌作用而表现得最为有效。此外,当抗体的靶标一旦被抗体结合即被内化,则由此提供了清除细胞因子/细胞因子受体复合物的有效机制。已观察到pd-l1有抗体介导的靶标内化。这是相比使用抗tgfβ抗体的独特优势,因为首先,抗tgfβ抗体的中和可能不完全;第二,抗体可以作为载体延长细胞因子的半衰期,且抗体/细胞因子复合物通常有循环槽(circulatingsink)的作用累积并最终将细胞因子解离释放回循环(montero-julian等,blood.1995;85:917-24)。采用细胞因子阱来中和配体也会是比用抗体封阻受体更好的策略,例如对csf-1来说。因为csf-1从循环中清除是通过受体介导的内吞作用,所以抗csf-1受体抗体阻断引起循环csf-1浓度的显著增加(hume等,blood.2012;119:1810-20)。
如后文所述,用抗pd-l1/tgfβ阱治疗联合至少一种其他抗癌治疗剂引起协效性抗肿瘤效果,这是因为同时发生的肿瘤细胞上pd-l1与免疫细胞上pd-1之间的相互作用被阻断、肿瘤微环境中tgfβ被中和以及抗癌药物的治疗作用。不受理论束缚,推定这是因为同时发生的由单个分子体阻断两种主要免疫逃逸机制产生的协同效应并定向清除肿瘤微环境中的tgfβ,加上其他抗癌药物的抗肿瘤作用。所述清除是通过(1)抗pd-l1靶向肿瘤细胞;(2)肿瘤微环境中的tgfβ自泌/旁泌被tgfβ阱所结合和(3)被结合的tgfβ被pd-l1受体介导的内吞作用所破坏。上述作用机制无法通过各自独立的抗pd-l1、tgfβ阱和其他抗癌治疗剂的合并疗法来实现。并且,tgfβrii融合于fc(igg的结晶片段)的c末端比将tgfβrii置于fc的n末端的tgfβrii-fc强数倍。用抗pdl1/tgfβ阱获得的有益功效还消除了tgfβrii不捕获tgfβ2的一些问题。yang等,trendsimmunol.2010;31:220–227指出,尽管某些肿瘤类型最初确实分泌tgfβ2,随着肿瘤进展,肿瘤微环境中的tgfβ主要由髓源抑制细胞分泌,这些细胞分泌tgfβ1。在显示出作为有效免疫-肿瘤治疗剂的巨大前景之外,采用可溶tgfβrii的治疗能潜在地减少tgfβ靶向治疗尤其是tgfβri激酶抑制剂的心脏毒性问题。这是因为tgfβ2在心脏的胚胎发育中、在缺血和再灌注损伤后的心肌损伤修复中起重要作用(roberts等,jclininvest.1992;90:2056-62)。
以tgfβ为癌症靶标
tgfβ作为癌症免疫治疗的靶标曾一度遭到质疑,因为其双重作用好比癌症的分子性双重人格(jekyllandhyde)(bierie等,natrevcancer.2006;6:506-20)。就像其他有些细胞因子,tgfβ活性具有发育阶段和背景依赖性。实际上,tgfβ既可以作为肿瘤促进因子也可以作为肿瘤抑制因子影响肿瘤的发生、进展和转移。tgfβ这种双重作用的潜在机制仍不清楚(yang等,trendsimmunol.2010;31:220-227)。虽然曾假定smad依赖性信号传导介导tgfβ信号传导的生长抑制而非smad依赖性途径提供肿瘤促进作用,但也有数据显示smad依赖性途径参与肿瘤进展(yang等,cancerres.2008;68:9107-11)。
人们已对tgfβ配体及其受体作为治疗靶标都进行了深入的研究。有三种配体同种型:tgfβ1、2和3,都是均二聚体。tgfβ受体(tgfβr)也有三种,它们被称为i型、ii型和iii型tgfβr(lópez-casillas等,jcellbiol.1994;124:557-68)。tgfβri是信号传导链但不结合配体。tgfβrii高亲合力地结合配体tgfβ1和3,对tgfβ2则不然。tgfβrii/tgfβ复合物募集tgfβri形成信号传导复合物(won等,cancerres.1999;59:1273-7)。tgfβriii是tgfβ与其信号传导受体结合的正向调节物,并以高亲和力结合全部3种tgfβ同种型。在细胞表面上,tgfβ/tgfβriii复合物结合tgfβrii,然后募集tgfβri取代tgfβriii形成信号传导复合物。
尽管三种不同的tgfβ同种型均通过相同的受体发出信号,但已知它们在体内具有差异表达模式且功能不重叠。敲除了三种不同tgf-β同种型的小鼠具有不同的表型,表明它们有许多非代偿行的功能(bujak等,cardiovascres.2007;74:184-95)。tgfβ1缺失小鼠具有造血功能和血管形成缺陷,tgfβ3缺失小鼠表现出肺发育和腭形成缺陷,tgfβ2缺失小鼠表现出各种发育异常,最突出的是多种心脏畸形(bartram等,circulation.2001;103:2745-52;yamagishi等,anatrec.2012;295:257-67)。此外,tgfβ还在缺血和再灌注损伤后的心肌损伤修复中起重要作用。在成人心脏中,心肌细胞分泌tgfβ作为自泌物维持自发搏动速率。重要的是,心肌细胞分泌的tgfβ中70-85%是tgfβ2(roberts等,jclininvest.1992;90:2056-62)。总之,鉴于tgfβ1和tgfβ2各自在肿瘤微环境和心脏生理学中的重要作用,中和tgfβ1而不中和tgfβ2的治疗剂可通过尽可能降低心脏毒性而不折损抗肿瘤活性来提供最佳的治疗指数。这与本发明发明人的发现一致,本发明发明人观察到抗pd-l1/tgfβ阱在猴子中没有毒性(包括心脏毒性)。
中和tgfβ的治疗方法包括采用tgfβ受体的胞外域作为可溶性受体阱和中和抗体。在受体捕获方法中,可溶性tgfβriii似乎是显然的选择,因为它结合所有三种tgfβ配体。然而,tgfβriii的天然形式是280-330kd的葡糖胺聚糖(gag)-糖蛋白,具有762个氨基酸残基的胞外域,对于生物治疗开发来说是非常复杂的蛋白质。可在昆虫细胞中产生去gag的可溶性tgfβriii并显示是有效的tgfβ中和剂(vilchis-landeros等,biochemj.355:215,2001)。tgfβriii两个分开的结合域(内皮糖蛋白(endoglin)相关和尿调节素相关)可以各自独立表达,但是它们的亲和力比可溶性tgfβriii低20至100倍,中和活性也低得多(mendoza等,biochemistry.2009;48:11755-65)。另一方面中,tgfβrii的胞外域长度仅为136个氨基酸残基,可以25-35kd糖蛋白质的形式生产。还显示,重组可溶性tgfβrii以200pm的kd结合tgfβ1,这与细胞上全长tgfβrii50pm的kd非常相近(lin等,jbiolchem.1995;270:2747-54)。对可溶性tgfβrii-fc进行了作为抗癌剂的测试,在肿瘤模型中表现为能够抑制已建立鼠恶性间皮瘤生长(suzuki等,clincancerres.2004;10:5907-18)。由于tgfβrii不结合tgfβ2,tgfβriii结合tgfβ1和3的亲和力低于tgfβrii,于是在细菌中生产tgfβriii的内皮糖蛋白结构域与tgfβrii胞外域的融合蛋白,该蛋白在细胞实验中表现为抑制tgfβ1和2的信号传导比tgfβrii或riii更有效(verona等,proteinengdessel.2008;21:463-73)。尽管在肿瘤模型中发现了一些令人鼓舞的抗肿瘤活性,但据我们所知,尚无tgfβ受体阱重组蛋白进入临床测试。
中和tgfβ配体全部三种同种型的另一种方法是筛选泛中和性抗tgfβ抗体或阻断受体与tgfβ1、2和3结合的抗受体抗体。gc1008是对全部tgfβ同种型特异的人抗体,已进入晚期恶性黑素瘤或肾细胞癌患者的i/ii期研究(morris等,jclinoncol2008;26:9028(会议摘要))。虽然发现该治疗安全且耐受良好,但仅观察到有限的临床效果,因此难以在不进一步鉴定免疫作用的情况下解释抗tgfβ治疗的重要性(flavell等,natrevimmunol.2010;10:554-67)。也有tgfβ同种型特异性抗体进入临床试验。tgfβ1特异性的美替木单抗(metelimumab)进行过用于预防青光眼术后过度术后瘢痕形成的2期临床试验;tgfβ2特异性的乐德木单抗(lerdelimumab)在3期研究中被发现是安全的但不能有效改善眼科手术后的瘢痕形成(khaw等,ophthalmology2007;114:1822-1830)。阻断受体与全部三种tgfβ同种型结合的抗tgfβrii抗体,例如抗人tgfβrii抗体tr1和抗小鼠tgfβrii抗体mt1,也显示出对小鼠原发性肿瘤生长和转移的一些治疗功效。迄今为止,绝大多数对tgfβ靶向抗癌治疗的研究,包括通常具有相当毒性的tgfβ信号传导小分子抑制剂,大多数处于临床前阶段,并且获得的抗肿瘤效果非常有限(calone等,exponcol.2012;34:9-16;connolly等,intjbiolsci.2012;8:964-78)。
本发明的抗体-tgfβ阱是双功能蛋白质,可用于本发明的联合治疗,其包含能够结合tgfβ的人tgfβ受体ii(tgfβrii)的至少部分。实施方式之一中,tgfβ阱多肽是能够结合tgfβ的2型人tgfβ受体同种型a(seqidno:8)的可溶性部分。另一个实施方式中,tgfβ阱多肽包含至少seqidno:8的氨基酸73-184。另一个实施方式中,tgfβ阱多肽包含seqidno:8的氨基酸24-184。另一个实施方式中,tgfβ阱多肽是能够结合tgfβ的2型人tgfβ受体同种型b(seqidno:9)的可溶性部分。另一个实施方式中,tgfβ阱多肽包含至少seqidno:9的氨基酸48-159。另一个实施方式中,tgfβ阱多肽包含seqidno:9的氨基酸24-159。另一个实施方式中,tgfβ阱多肽包含seqidno:9的氨基酸24-105。
免疫哨点去抑制
用治疗性抗体靶向t细胞抑制哨点来去抑制是一个具有深入研究的领域(综述可见pardoll,natrevcancer.2012;12:253-264)。方案之一中,抗体部分或其抗原结合片段靶向t细胞上的t细胞抑制哨点受体蛋白,例如:ctla-4、pd-1、btla、lag-3、tim-3和lair1。另一方案中,抗体部分靶向抗原呈递细胞和肿瘤细胞(它们也选择这些反受体中的某些用于它们自身的免疫逃逸)上的反受体(counter-receptor),例如:pd-l1(b7-h1)、b7-dc、hvem、tim-4、b7-h3或b7-h4。
本发明考虑采用通过其抗体部分或其抗原结合片段靶向t细胞抑制哨点的抗体tgfβ阱来去抑制。为此,发明人测试了tgfβ阱与靶向多种t细胞抑制哨点受体蛋白的抗体(例如抗pd-1、抗pd-l1、抗tim-3和抗lag3)联合的抗肿瘤效果。发明人发现,tgfβ阱联合抗pd-l1抗体显示出超过单一疗法的显著的抗肿瘤活性。相比之下,靶向上述目标的抗体的其他组合均未显示出任何优异的效果。尤其,人们可能预期tgfβ阱与抗pd-1抗体联合治疗将表现出与用抗pd-l1所得相似的活性,因为pd-1/pd-l1是通过彼此结合实现免疫哨点抑制的同源受体。然而,本发明的发明人发现并非如此。
抗pd-l1抗体
本发明可包括采用本领域述及的任何抗pd-l1抗体或其抗原结合片段。抗pd-l1抗体是可购得的,例如29e2a3抗体(biolegend,cat.no.329701)。抗体可以是单克隆抗体、嵌合抗体、人源化抗体或人抗体。抗体片段包括fab、f(ab’)2、scfv和fv片段,详见后文。
示例性抗体可见pct公开文本wo2013/079174。这些抗体可包含含有hvr-h1、hvr-h2和hvr-h3序列的重链可变区多肽,其中:
(a)hvr-h1序列是x1yx2mx3;
(b)hvr-h2序列是siypsggx4tfyadx5vkg;
(c)hvr-h3序列是iklgtvttvx6y;
并且其中:x1是k、r、t、q、g、a、w、m、i或s;x2是v、r、k、l、m或i;x3是h、t、n、q、a、v、y、w、f或m;x4是f或i;x5是s或t;x6是e或d。
实施方式之一中,x1是m、i或s;x2是r、k、l、m或i;x3是f或m;
x4是f或i;x5是s或t;x6是e或d。
另一个实施方式中x1是m、i或s;x2是l、m或i;x3是f或m;x4是i;x5是s或t;x6是d。
另一个实施方式中,x1是s;x2是i;x3是m;x4是i;x5是t;x6是d。
另一方面中,所述多肽还包含位于hvr之间的可变区重链框架序列,如下所示:(hc-fr1)-(hvr-h1)-(hc-fr2)-(hvr-h2)-(hc-fr3)-(hvr-h3)-(hc-fr4)。
另一个方面,所述框架序列源自人共有框架序列或人种系框架序列。
另一方面中,所述框架序列至少其一如下所述:
hc-fr1是evqllesggglvqpggslrlscaasgftfs;
hc-fr2是wvrqapgkglewvs;
hc-fr3是rftisrdnskntlylqmnslraedtavyycar;
hc-fr4是wgqgtlvtvss。
另一方面中,重链多肽进一步与包括hvr-l1、hvr-l2和hvr-l3的可变区轻链组合,其中:
(a)hvr-l1序列是tgtx7x8dvgx9ynyvs;
(b)hvr-l2序列是x10vx11x12rps;
(c)hvr-l3序列是ssx13tx14x15x16x17rv;
并且其中:x7是n或s;x8是t、r或s;x9是a或g;x10是e或d;x11是i、n或s;x12是d、h或n;x13是f或y;x14是n或s;x15是r、t或s;x16是g或s;x17是i或t。
另一实施方式中,x7是n或s;x8是t、r或s;x9是a或g;x10是e或d;x11是n或s;x12是n;x13是f或y;x14是s;x15是s;x16是g或s;x17是t。
另一实施方式中,x7是s;x8是s;x9是g;x10是d;x11是s;x12是n;x13是y;x14是s;x15是s;x16是s;x17是t。
另一方面中,所述轻链还包含位于hvr之间的可变区轻链框架序列,如下所示:(lc-fr1)-(hvr-l1)-(lc-fr2)-(hvr-l2)-(lc-fr3)-(hvr-l3)-(lc-fr4)。
另一方面中,所述轻链框架序列源自人共有框架序列或人种系框架序列。
另一方面中,所述轻链框架序列是λ轻链序列。
另一方面中,所述框架序列至少其一如下所述:
lc-fr1是qsaltqpasvsgspgqsitisc;
lc-fr2是wyqqhpgkapklmiy;
lc-fr3是gvsnrfsgsksgntasltisglqaedeadyyc;
lc-fr4是fgtgtkvtvl。
另一实施方式中,本发明提供抗pd-l1抗体或包含重链和轻链可变区序列的抗原结合片段,其中:
(a)所述重链包括hvr-h1、hvr-h2和hvr-h3,并且其中:(i)hvr-h1序列是x1yx2mx3;(ii)hvr-h2序列是siypsggx4tfyadx5vkg;(iii)hvr-h3序列是iklgtvttvx6y,且;
(b)所述轻链包括hvr-l1、hvr-l2和hvr-l3,并且其中:(iv)hvr-l1序列是tgtx7x8dvgx9ynyvs;(v)hvr-l2序列是x10vx11x12rps;(vi)hvr-l3序列是ssx13tx14x15x16x17rv;其中:x1是k、r、t、q、g、a、w、m、i或s;x2是v、r、k、l、m或i;x3是h、t、n、q、a、v、y、w、f或m;x4是f或i;x5是s或t;x6是e或d;x7是n或s;x8是t,r或s;x9是a或g;x10是e或d;x11是i,n或s;x12是d、h或n;x13是f或y;x14是n或s;x15是r、t或s;x16是g或s;x17是i或t。
实施方式之一中,x1是m、i或s;x2是r、k、l、m或i;x3是f或m;
x4是f或i;x5是s或t;x6是e或d;x7是n或s;x8是t,r或s;x9是a或g;x10是e或d;x11是n或s;x12是n;x13是f或y;x14是s;x15是s;x16是g或s;x17是t。
另一实施方式中,x1是m、i或s;x2是l、m或i;x3是f或m;x4是i;x5是s或t;x6是d;x7是n或s;x8是t、r或s;x9是a或g;x10是e或d;x11是n或s;x12是n;x13是f或y;x14是s;x15是s;x16是g或s;x17是t。
另一实施方式中,x1是s;x2是i;x3是m;x4是i;x5是t;x6是d;x7是s;x8是s;x9是g;x10是d;x11是s;x12是n;x13是y;x14是s;x15是s;x16是s;x17是t。
另一方面中,所述重链可变区包含如下所示位于hvr之间的一个或多个框架序列:(hc-fr1)-(hvr-h1)-(hc-fr2)-(hvr-h2)-(hc-fr3)-(hvr-h3)-(hc-fr4),并且所述轻链可变区包含如下所示位于hvr之间的一个或多个框架序列:(lc-fr1
mhvr-l1)-(lc-fr2)-(hvr-l2)-(lc-fr3)-(hvr-l3)-(lc-fr4)。
另一方面中,所述框架序列源自人共有框架序列或人种系序列。
另一方面中,所述重链框架序列中一个或多个如下所述:
hc-fr1是evqllesggglvqpggslrlscaasgftfs;
hc-fr2是wvrqapgkglewvs;
hc-fr3是rftisrdnskntlylqmnslraedtavyycar;
hc-fr4是wgqgtlvtvss。
另一方面中,所述轻链框架序列是λ轻链序列。
另一方面中,所述轻链框架序列中一个或多个如下所述:
lc-fr1是qsaltqpasvsgspgqsitisc;
lc-fr2是wyqqhpgkapklmiy;
lc-fr3是gvsnrfsgsksgntasltisglqaedeadyyc;
lc-fr4是fgtgtkvtvl。
另一方面中,重链可变区多肽、抗体或抗体片段还包含至少ch1结构域。
在一更具体的方面中,重链可变区多肽、抗体或抗体片段还包含ch1、ch2和ch3结构域。
另一方面中,可变区轻链、抗体或抗体片段还包含cl结构域。
另一方面中,抗体还包括ch1、ch2、ch3和cl结构域。
另一方面中,抗体还包含人或鼠恒定区。
另一方面中,人恒定区选自igg1、igg2、igg2、igg3和igg4。
在一更具体的方面中,人或鼠恒定区是lgg1。
另一实施方式中,本发明的特征包括一种抗pd-l1抗体,其包含重链和轻链可变区序列,其中:
(a)所述重链包括hvr-h1、hvr-h2和hvr-h3,它们分别与syimm、siypsggitfyadtvkg和iklgtvttvdy具有至少80%总体序列相同性,且
(b)所述轻链包括hvr-l1、hvr-l2和hvr-l3,它们分别与tgtssdvggynyvs、dvsnrps和ssytssstrv具有至少80%总体序列相同性。
在一具体的方面中,所述序列相同性是81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
另一实施方式中,本发明的特征包括一种抗pd-l1抗体,其包含重链和轻链可变区序列,其中:
(a)所述重链包括hvr-h1、hvr-h2和hvr-h3,它们分别与mymmm、siypsggitfyadsvkg和iklgtvttvdy具有至少80%总体序列相同性,且
(b)所述轻链包括hvr-l1、hvr-l2和hvr-l3,它们分别与
tgtssdvgaynyvs、dvsnrps和ssytssstrv具有至少80%总体序列相同性。
在一具体的方面中,所述序列相同性是81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
另一方面中,本发明的抗体或抗体片段中,相比hvr-h1、hvr-h2和hvr-h3的序列,至少如下所示以下划线突出显示的那些氨基酸保持不变:
(a)hvr-h1中:syimm,
(b)hvr-h2中:siypsggitfyadtvkg,
(c)hvr-h3中:iklgtvttvdy;
并且其中,相比hvr-l1、hvr-l2和hvr-l3的序列,至少如下所示以下划线突出显示的那些氨基酸保持不变:
(a)hvr-l1tgtssdvggynyvs
(b)hvr-l2dvsnrps
(c)hvr-l3ssytssstrv。
另一方面中,所述重链可变区包含如下所示位于hvr之间的一个或多个框架序列:(hc-fr1)-(hvr-h1)-(hc-fr2)-(hvr-h2)-(hc-fr3)-(hvr-h3)-(hc-fr4),并且所述轻链可变区包含如下所示位于hvr之间的一个或多个框架序列:
(lc-fr1)-(hvr-l1)-(lc-fr2)-(hvr-l2)-(lc-fr3)-(hvr-l3)-(lc-fr4)。
另一方面中,所述框架序列源自人种系序列。
另一方面中,所述重链框架序列中一个或多个如下所述:
hc-fr1是evqllesggglvqpggslrlscaasgftfs;
hc-fr2是wvrqapgkglewvs;
hc-fr3是rftisrdnskntlylqmnslraedtavyycar;
hc-fr4是wgqgtlvtvss。
另一方面中,所述轻链框架序列源自λ轻链序列。
另一方面中,所述轻链框架序列中一个或多个如下所述:
lc-fr1是qsaltqpasvsgspgqsitisc;
lc-fr2是wyqqhpgkapklmiy;
lc-fr3是gvsnrfsgsksgntasltisglqaedeadyyc;
lc-fr4是fgtgtkvtvl。
另一更具体的方面中,抗体还包含人或鼠恒定区。
另一方面中,人恒定区选自igg1、igg2、igg2、igg3和igg4。
另一个实施方式中,本发明的特征包括一种抗pd-l1抗体,其包含重链和轻链可变区序列,其中:
(a)所述重链序列与以下重链序列具有至少85%序列相同性:
evqllesggglvqpggslrlscaasgftfssyimmvwrqapgkglewvssiypsggitfyadwkgrftisrdnskntlylqmnslraedtavyycariklgtvttvdywgqgtlvtvss,并且
(b)所述轻链序列与以下轻链序列具有至少85%序列相同性:
qsaltqpasvsgspgqsitisctgtssdvggynyvswyqqhpgkapklmiydvsnrpsgvsnrfsgsksgntasltisglqaedeadyycssytssstrvfgtgtkvtvl。
在一具体的方面中,所述序列相同性是86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
另一实施方式中,本发明提供一种抗pd-l1抗体,其包含重链和轻链可变区序列,其中:
(a)所述重链序列与以下重链序列具有至少85%序列相同性:
evqllesggglvqpggslrlscaasgftfsmymmmwvrqapgkglevwssiypsggitfyadsvkgrftisrdnskntlylqmnslraedtaiyycariklgtvttvdywgqgtlvtvss,并且
(b)所述轻链序列与以下轻链序列具有至少85%序列相同性:
qsaltqpasvsgspgqsitisctgtssdvgaynyvswyqqhpgkapklmiydvsnrpsgvsnrfsgsksgntasltisglqaedeadyycssytssstrvfgtgtkvtvl。
在一具体方面中,所述序列相同性是86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。另一实施方式中,所述抗体结合人、小鼠或猕猴pd-l1。在一具体方面中,抗体能够阻断人、小鼠或猕猴pd-l1与相应人、小鼠或猕猴pd-1受体之间的相互作用。
另一实施方式中,抗体以5x10-9m或更低kd、优选以2x10-9m或更低kd、甚至更优选以1x10-9m或更低kd结合人pd-l1。
另一实施方式中,本发明涉及抗pd-l1抗体或其抗原结合片段,其结合包含人pd-l1残基y56和d61的功能性表位。
在一具体方面中,所述功能性表位还包含人pd-l1的e58、e60、q66、r113和m115。
在一更具体的方面中,所述抗体结合包含人pd-l1残基54-66和112-122的构象表位。
另一实施方式中,本发明涉及抗pd-l1抗体或其抗原结合片段的用途,所述抗pd-l1抗体或其抗原结合片段与本文所述的本发明抗体交叉竞争结合pd-l1。
另一实施方式中,本发明的特征包括包含任何上述抗pd-l1抗体的蛋白质和多肽与至少一种药学上可接受的运载体组合用于本发明的联合疗法。
另一个实施方式中,本发明的特征包括一种分离的核酸,其编码本文所述的抗pd-l1抗体或其抗原结合片段的多肽、或轻链或重链可变区序列。另一实施方式中,本发明提供一种分离的核酸,其编码抗pd-l1抗体的轻链或重链可变区序列,其中:
(a)所述重链包括hvr-h1、hvr-h2和hvr-h3序列,它们分别与syimm、siypsggitfyadtvkg和iklgtvttvdy具有至少80%序列相同性,或者
(b)所述轻链包括hvr-l1、hvr-l2和hvr-l3序列,它们分别与tgtssdvggynyvs、dvsnrps和ssytssstrv具有至少80%序列相同性。
在一个具体方面,所述序列相同性是81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%。
另一方面,重链的核酸序列是:
轻链的核酸序列是:
进一步,可用于抗pd-l1/tgfβ阱的示例性抗-pd-l1抗体可见美国专利申请公开文本us2010/0203056。本发明实施方式之一中,抗体部分是yw243.55s70。本发明另一实施方式中,抗体部分是mpdl3280a。
另一实施方式中,本发明的特征包括包含重链和轻链可变区序列的抗pd-l1抗体部分的用途,其中:
(a)所述重链序列与以下重链序列具有至少85%序列相同性:
evqlvesggglvqpggslrlscaasgftfsdswihwvrqapgkglewvawispyggstyyadsvkgrftisadtskntaylqmnslraedtavyycarrhwpggfdywgqgtlvtvss(seqidno:12),并且
(b)所述轻链序列与以下轻链序列具有至少85%序列相同性:
diqmtqspsslsasvgdrvtitcrasqdvstavawyqqkpgkapklliysasflysgvpsrfsgsgsgtdftltisslqpedfatyycqqylyhpatfgqgtkveikr(seqidno:13)。
在一具体方面中,所述序列相同性是86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%或99%。
另一实施方式中,本发明的特征包括包含重链和轻链可变区序列的抗pd-l1抗体部分的用途,其中:
(a)所述重链可变区序列是:
evqlvesggglvqpggslrlscaasgftfsdswihwvrqapgkglewvawispyggstyyadsvkgrftisadtskntaylqmnslraedtavyycarrhwpggfdywgqgtlvtvss(seqidno:12),并且
(b)所述轻链可变区序列是:
diqmtqspsslsasvgdrvtitcrasqdvstavawyqqkpgkapklliysasflysgvpsrfsgsgsgtdftltisslqpedfatyycqqylyhpatfgqgtkveikr(seqidno:13)。
另一实施方式中,本发明的特征包括一种包含重链和轻链可变区序列的抗pd-l1抗体部分,其中:
(a)所述重链可变区序列是:
evqlvesggglvqpggslrlscaasgftfsdswihwvrqapgkglewvawispyggstyyadsvkgrftisadtskntaylqmnslraedtavyycarrhwpggfdywgqgtlvtvsa(seqidno:14),并且
(b)所述轻链可变区序列是:
diqmtqspsslsasvgdrvtitcrasqdvstavawyqqkpgkapklliysasflysgvpsrfsgsgsgtdftltisslqpedfatyycqqylyhpatfgqgtkveikr(seqidno:13)。
进一步,可用于抗pd-l1/tgfβ阱的示例性抗-pd-l1抗体可见美国专利公开文本us7,943,743。
本发明实施方式之一中,所述抗pd-l1抗体是mdx-1105。
另一实施方式中,所述抗pd-l1抗体是medi-4736。
恒定区
本发明联合疗法中所用蛋白质和肽可包含免疫球蛋白恒定区或所述恒定区的片段、类似物、变体、突变体或衍生物。优选实施方式中,恒定区源自人免疫球蛋白重链,例如igg1、igg2、igg3、igg4或其他类。实施方式之一中,恒定区包括ch2结构域。另一实施方式中,恒定区包括ch2和ch3结合域或包括铰链-ch2-ch3。或者,恒定区可包括全部或部分铰链区、ch2结构域和/或ch3结构域。
实施方式之一中,恒定区包含降低对fc受体亲和力或降低fc效应功能的突变。例如,恒定区可包含消除igg重链恒定区中糖基化位点的突变。一些实施方式中,恒定区含有氨基酸位置对应于igg1的leu234、leu235、gly236、gly237、asn297或pro331的突变、缺失或插入(氨基酸根据eu命名法编号)。具体实施方式之一中,恒定区含有氨基酸位置对应于igg1asn297的突变。另一实施方式中,恒定区含有氨基酸位置对应于igg1的leu281、leu282、gly283、gly284,、asn344或pro378的突变、缺失或插入。
一些实施方式中,恒定区含有源自人igg2或igg4重链的ch2结构域。优选地,ch2结构域包含消除ch2结构域中糖基化位点的突变。实施方式之一中,突变改变igg2或igg4重链ch2结构域内gln-phe-asn-ser(seqidno:15)氨基酸序列内的天冬酰胺。优选地,突变将天冬酰胺变成谷氨酰胺。或者,突变改变gln-phe-asn-ser(seqidno:15)氨基酸序列内苯丙氨酸和天冬酰胺两者。在实施方式之一中,gln-phe-asn-ser(seqidno:15)氨基酸序列被gln-ala-gln-ser(seqidno:16)氨基酸序列取代。
gln-phe-asn-ser(seqidno:15)氨基酸序列内的天冬酰胺对应于igg1的asn297。
另一实施方式中,恒定区包括ch2结构域和至少部分铰链区。铰链区可以是源自免疫球蛋白重链例如igg1、igg2、igg3、igg4或其他类的。优选地,铰链区源自人igg1、igg2、igg3、igg4或其他类。更优选地,铰链区源自人igg1重链。实施方式之一中,igg1铰链区pro-lys-ser-cys-asp-lys(seqidno:17)氨基酸序列中的半胱氨酸被改变。一优选实施方式中,pro-lys-ser-cys-asp-lys(seqidno:17)氨基酸序列被pro-lys-ser-ser-asp-lys(seqidno:18)氨基酸序列取代。实施方式之一中,恒定区包括源自第一抗体同种型的ch2结构域和源自第二抗体同种型的铰链区。具体实施方式之一中,ch2结构域源自人igg2或igg4重链,而铰链区源自经改变的人igg1重链。
fc部分与非fc部分连接处附近的氨基酸改变会显著提高fc融合蛋白的血清半衰期(pct公开文本wo01/58957,其公开内容通过引用纳入本文)。因此,本发明蛋白质或多肽的连接区可含有相对于免疫球蛋白重链和促红细胞生成素天然序列的改变,优选距连接点约10个氨基酸以内。这些氨基酸改变会引起疏水性增强。实施方式之一中,恒定区源自igg序列,其中c末端的赖氨酸残基被替换。优选地,igg序列的c末端赖氨酸被替换成非赖氨酸的氨基酸(例如丙氨酸或亮氨酸)以进一步提高血清半衰期。另一实施方式中,恒定区源自igg序列,其中,恒定区c末端附近的leu-ser-leu-ser(seqidno:19)氨基酸序列具有消除潜在接合性t细胞表位的改变。例如,实施方式之一中,leu-ser-leu-ser氨基酸序列被ala-thr-ala-thr(seqidno:20)氨基酸序列取代。其他实施方式中,leu-ser-leu-ser(seqidno:19)段内的氨基酸被例如甘氨酸或脯氨酸等其他氨基酸替换。在igg1、igg2、igg3、igg4或他类免疫球蛋白分子的c-末端附近产生leu-ser-leu-ser(seqidno:19)段氨基酸取代的方法可详见美国专利公开文本2003/0166877,其公开内容通过引用纳入本文。
本发明的适宜铰链区可源自igg1、igg2、igg3、igg4及其他免疫球蛋白类别。igg1铰链区有三个半胱氨酸,其中两个参与免疫球蛋白两重链之间的二硫键。这些相同的半胱氨酸使得fc部分之间二硫键的形成高效且一致。所以,本发明的优选铰链区源自igg1,更优选源自人igg1。优选实施方式中,人igg1铰链区内的第一个半胱氨酸突变为其他氨基酸,优选丝氨酸。igg2同种型铰链区有四个二硫键,它们倾向于促成重组系统分泌期间的寡聚化和可能不正确的二硫键。适宜的铰链区可源自igg2铰链,其中前两个半胱氨酸各自突变成其他氨基酸。已知igg4的铰链区形成链间二硫键不大有效。然而,本发明的合适铰链区可以源自igg4铰链区,优选含有增强重链衍生部分之间正确形成二硫键的突变(angals等,(1993)mol.immunol.,30:105-8)。
根据本发明,恒定区可包含源自不同抗体同种型的ch2和/或ch3结构域和铰链区,即杂交(hybrid)恒定区。例如,实施方式之一中,恒定区含有源自igg2或igg4的ch2和/或ch3结构域和源自igg1的突变铰链区。或者,杂交恒定区中可采用源自其他igg亚类的突变铰链区。例如,可以采用igg4铰链的突变形式来有效形成两条重链之间的二硫键结合。突变型铰链也可源自igg2铰链,其中前两个半胱氨酸各自突变成其他氨基酸。杂交恒定区的组装可见美国专利公开2003/0044423,其公开内容通过引用纳入本文。
根据本发明,恒定区可包含一处或多处本文所述的突变。fc部分中突变的组合对延长血清半衰期和提高双功能分子体内效力具有累加或协同效应。因此,示例性实施方式之一中,恒定区可包含(i)源自igg序列的区域,其中leu-ser-leu-ser(seqidno:19)氨基酸序列被ala-thr-ala-thr(seqidno:20)氨基酸序列取代;(ii)c-末端为丙氨酸而非赖氨酸;(iii)源自不同抗体同种型的ch2结构域和铰链区,例如igg2的ch2结构域和改变的igg1铰链区;和(iv)消除igg2源ch2结构域内糖基化位点的突变,例如,igg2源ch2结构域内gln-ala-gln-ser(seqidno:16)氨基酸序列取代gln-phe-asn-ser(seqidno:15)的氨基酸序列。
抗体片段
用于本发明联合疗法的本发明蛋白质和多肽还可以包括抗体的抗原结合片段。示例性抗体片段包括scfv、fv、fab、f(ab’)2和单域vhh片段,例如来自骆驼科动物的那些。
单链抗体片段,也称为单链抗体(scfv),是重组多肽,它们通常结合抗原或受体;这些片段包含用或不用一个或多个互连接头相连的至少一个抗体可变轻链序列(vl)片段和至少一个抗体可变重链氨基酸序列(vh)片段。这样的接头可以是短的柔性肽,选来确保vl和vh结构域相连后正确的三维折叠,从而保留作为单链抗体片段来源的全抗体的靶分子结合特异性。通常,vl或vh序列的羧基末端通过这样的肽接头共价连接至互补vl和vh序列的氨基酸末端。单链抗体片段可通过分子克隆、抗体噬菌体文库展示或类似技术来产生。这些蛋白质可以在真核细胞中也可以在原核细胞(包括细菌)中生产。
单链抗体片段包含具有本文所述完整抗体可变区或cdr中至少其一的氨基酸序列,但缺失那些抗体的全部或部分恒定域。这些恒定域对于抗原结合来说不是必需的,但是构成完整抗体结构的主要部分。因此,单链抗体片段可以克服采用包含部分或全部恒定区的抗体相关的一些问题。例如,单链抗体片段倾向于没有生物分子与重链恒定区之间不希望有的相互作用或其他不希望有的生物活性。此外,单链抗体片段比完整抗体小得多,因此可具有比完整抗体更高的毛细血管通透性,这令单链抗体片段能够更高效地寻址并结合至靶抗原结合位点。并且,抗体片段可以在原核细胞中以相对大的规模产生,从而促进其生产。此外,单链抗体片段相对较小使得它们比完整抗体更不会在接受者中引发免疫反应。
还可以存在与完整抗体具有相同或相当的结合特征的抗体片段。这样的片段可含有一个或两个fab片段或f(ab’)2片段。抗体片段可包含完整抗体的全部六个cdr,但包含少于全部这些区域、例如含三、四或五个cdr的片段同样具有功能。
蛋白质生产
抗体-细胞因子阱蛋白质一般采用包含经工程改造以表达所述蛋白质的核酸的哺乳动物细胞通过重组技术来生产。尽管在实施例1和2中描述了合适的细胞系和蛋白质生产方法的一个例子,但是已有大量合适的载体、细胞系和蛋白质生产方法用来生产基于抗体的生物药物并可以用于合成本文的抗体-细胞因子阱蛋白。
治疗适应症
本发明涉及用于治疗癌症或抑制肿瘤生长的联合疗法,尤其以下所述的联合:(i)双功能分子,其包含tgfβrii或其能够结合tgfβ的片段和结合免疫哨点蛋白例如程序性死亡配体1(pd-l1)的抗体或其抗原结合片段和(ii)至少一种其他抗癌治疗剂。抗癌治疗剂包括例如辐照、化疗药、生物制剂或疫苗。本发明的某些实施方式中,联合治疗提供协效性抗癌效果。
示例性的癌症包括:结直肠癌、乳腺癌、卵巢癌、胰腺癌、胃癌、前列腺癌、肾癌、宫颈癌、骨髓瘤、淋巴瘤、白血病、甲状腺癌、子宫内膜癌、子宫癌、膀胱癌、神经内分泌癌、头颈部癌、肝癌、鼻咽癌、睾丸癌、小细胞肺癌、非小细胞肺癌、黑素瘤、基底细胞皮肤癌、鳞状细胞皮肤癌、隆突性皮肤纤维肉瘤、梅克尔细胞癌、成胶质细胞瘤、胶质瘤、肉瘤、间皮瘤,和骨髓增生异常综合征。
待用抗pd-l1/tgfβ阱联合一种或多种其他抗癌治疗剂(如化疗和/或放疗)治疗的癌症或肿瘤可根据肿瘤中pd-l1和tgfβ的表达或表达升高、表达水平与预后或疾病进展的相关性、以及关于肿瘤对于pd-l1和tgfβ靶向治疗敏感性的临床前和临床经验来选择。这些癌症或肿瘤包括但不限于,结直肠癌、乳腺癌、卵巢癌、胰腺癌、胃癌、前列腺癌、肾癌、宫颈癌、膀胱癌、头颈部癌、肝癌、非小细胞肺癌、黑素瘤、梅克尔细胞癌和间皮瘤。
药物组合物
本发明的特征还包括含有治疗有效量的本文所述蛋白质的药物组合物,其用于本发明的治疗方法。所述组合物可配制成适用于多种药物递送系统。组合物中还可包含一种或多种生理学上可接受的赋形剂或运载体以制成合适的制剂。可用于本发明的合适制剂可见《雷明顿药物科学》第17版,宾夕法尼亚州伊斯顿的马克出版公司(mackpublishingcompany),1985。关于药物递送方法的综述可见例如langer(science249:1527-1533,1990)。
药物组合物意在通过胃肠外、鼻内、局部(topical)、经口或本地(local)(例如通过透皮方式)给药用于治疗。药物组合物可肠胃外给药(例如通过静脉内、肌肉内或皮下注射)、或口服摄入、或在受血管或癌症影响的部位局部施用或关节内注射给药。其他给药途径包括血管内、动脉内、肿瘤内、腹膜内、心室内、脑膜内,以及鼻、眼、巩膜内、眶内、直肠、局部或气溶胶吸入给药。因此,本发明提供用于胃肠外给药的组合物,其包含前文所述物质溶解或悬浮于可接受的运载体中、优选水性运载体例如水、带缓冲的水、盐水、pbs等。组合物可含有模拟生理条件所需的药学上可接受的辅料,例如ph调节剂和缓冲剂、张力调节剂、润湿剂,清洁剂(detergent)等。本发明还提供用于经口递送的组合物,其可包含惰性成分例如粘合剂或填充剂用于配制片剂、胶囊等。此外,本发明提供用于本地(local)给药的组合物,其可含有惰性成分如溶剂或乳化剂用于配制乳膏、软膏等。
这些组合物可用常规灭菌技术灭菌或可进行无菌过滤。所得水性溶液可包装成原样使用(“useas-is”)型产品或冻干,冻干制剂在给予之前与无菌水性运载体合并。制剂的ph通常在3至11之间,5至9之间或6至8之间更好,7至8之间例如7至7.5之间最好。得到的固体形式的组合物可以包装成多个单剂量单元,各个单元包含定量的一种或多种前文所述物质,例如以密封包装的多个片剂或胶囊的形式。固体形式的组合物还可以包装在给量灵活的容器内,例如用于局部施用乳膏或软膏的可挤压管中。
治疗
根据本发明任一方面内容确定剂量和疗程是本领域专业技术人员所能胜任的。本领域技术人员完全能够通过检测患者来确定是否启动、继续、中止(或终止)或恢复治疗。用于实施本发明联合疗法的抗体-tgfβ阱的量、抗癌治疗剂的量或辐照量将根据诸如所治疗的病症、患者整体健康状况以及给药方法、途径和剂量等而不同。
根据某些实施方式,抗体-tgfβ阱和至少一种其他抗癌药物各自的给药量是各自已知用于治疗特定类型癌症的治疗量。根据其他实施方式,由于观察到的本发明联合疗法相关协同效应,抗体-tgfβ阱和至少一种其他抗癌药物各自的给药量可低于各自已知作为单一疗法用于治疗癌症的治疗量。
抗体-tgfβ阱的最佳剂量取决于要实现最大治疗效果的抗体部分对受体占位百分比,因为细胞因子阱是大量过量使用的。例如,将靶向细胞受体的单克隆抗体的治疗剂量确定为波谷浓度约10至100μg/ml,即60至600nm(对于解离常数kd为6nm的抗体来说,这样的波谷浓度将确保细胞上90-99%的靶受体被抗体占据)。这相对于细胞因子大量过量,因为细胞因子在循环中通常是pg至ng/ml的水平。
用于本发明治疗方法的抗体-tgfβ阱最佳剂量取决于所治疗的疾病、疾病的严重程度和是否有副作用。最佳剂量可通过常规试验来确定。就胃肠外给药而言,给予0.1mg/kg至100mg/kg之间、或者0.5mg/kg至50mg/kg之间、或者1mg/kg至25mg/kg之间、或者10mg/kg至25mg/kg之间、或者5mg/kg至20mg/kg之间、或者2mg/kg至10mg/kg之间、或者5mg/kg至10mg/kg之间的剂量,并可按照例如每轮治疗每周一次、隔周一次、每三周一次或每月一次来给药。本发明的一些实施方式中,抗体-tgfβ阱在组合疗法中实现治疗效果所需的有效剂量将小于抗体-tgfβ阱单一疗法中实现类似治疗效果所需的有效剂量。
本发明的一些实施方式中,有效剂量将比抗体-tgfβ阱单一疗法实现类似治疗效果所需有效剂量小约2-10倍。本发明的一些实施方式中,有效剂量将比抗体-tgfβ阱单一疗法实现类似治疗效果所需有效剂量小约2-5倍。
与抗体-tgfβ阱联用治疗癌症的其他化疗药或辐照的有效剂量可以根据所用的具体化合物或药物组合物、给药方式、所治病症以及所治病情严重程度而不同。普通医技人员或临床医生能够确定治疗或预防癌症进展所必需的各其他化疗药或辐照的有效量。本发明一些实施方式中,其他化疗药或放疗在本发明联合治疗中达到治疗效果所需的有效剂量将小于所述化疗药或所述放疗作为单一疗法达到类似治疗效果所需的有效剂量。
根据本发明的方法,化疗药可以与抗体-细胞因子阱分子联合给药来治疗癌症或减少肿瘤生长。这些化疗药包括例如烷化剂、抗代谢物、蒽环霉素、植物生物碱、拓扑异构酶抑制剂、抗瘤抗生素、激素类药、抗血管生成剂、分化诱导剂、细胞生长停滞诱导剂、细胞凋亡诱导剂、细胞毒性类药物和其他抗肿瘤药物。这些药物会以某种方式影响细胞分裂或dna合成和功能。代表性的化疗药包括但不限于烷化剂(如顺铂、卡铂、奥沙利铂、二氯甲基二乙胺、环磷酰胺、苯丁酸氮芥、达卡巴嗪、洛莫司汀、卡莫司汀、丙卡巴肼、苯丁酸氮芥和异环磷酰胺),抗代谢类(如氟尿嘧啶(5-fu)、吉西他滨、甲氨蝶呤、阿糖胞苷、氟达拉滨和氟尿嘧啶),抗有丝分裂类(包括紫杉烷类(如紫杉醇和多西他赛(decetaxel))和长春花生物碱类(如长春新碱、长春花碱、长春瑞滨和长春地辛),蒽环类(包括多柔比星、柔红霉素、戊柔比星(valrubicin)、伊达比星和表柔比星,以及放线菌素d等放线菌素类),细胞毒性抗生素(包括丝裂霉素、普鲁卡霉素和博来霉素)和拓扑异构酶抑制剂(包括喜树碱类(如伊立替康和拓扑替康)和表鬼臼毒素衍生物(如安吖啶、依托泊苷、磷酸依托泊苷和替尼泊苷)。
某些实施方式中,可用铂基类治疗剂例如顺铂、卡铂、奥沙利铂。治疗与效果可从与抗pd-l1/tgfβ阱分子联合中受益的其他抗癌药物包括抗代谢物类药物,例如干扰dna合成的氟尿嘧啶(5-fu)。某些实施方式中,一种或多种化疗药可组合与抗pd-l1/tgfβ阱分子联合给药。某些实施方式中,一种或多种化疗药可组合与放疗和抗pd-l1/tgfβ阱分子联合给药。
本发明一具体实施方式中,可给予以下剂量的奥沙利铂:20mg/m2至200mg/m2之间、或者40mg/m2至160mg/m2之间、或者60mg/m2至145mg/m2之间、或者85mg/m2至135mg/m2之间、或者40mg/m2至65mg/m2之间。
本发明一具体实施方式中,可给予以下剂量的5-fu:100mg/m2至3000mg/m2之间、或者250mg/m2至2400mg/m2之间、或者400mg/m2至1500mg/m2之间、或者200mg/m2至600mg/m2之间。本发明实施方式之一中,5-fu剂量可通过例如一段时间的输注来给予。
本发明一具体实施方式中,可给予亚叶酸(leucovorin)来增强5-fu的效果或降低化疗副作用。
本发明一具体实施方式中,提供以下化疗方案作为与pd-l1/tgfβ阱分子联合使用的例子。第一天,给予85mg/m2奥沙利铂和200mg/m2亚叶酸,2小时后给予400mg/m2推注5-fu和600mg/m2输注5-fu。第二天,给予200mg/m2亚叶酸,2小时后给予400mg/m2推注5-fu和600mg/m2输注5-fu。
本发明另一实施方式中,化疗方案包括例如第一天85mg/m2剂量的奥沙利铂、400mg/m2剂量亚叶酸、400mg/m2静脉推注5-fu和600mg/m2输注5-fu继以1200mg/m2/天x2天(总共2400mg/ml2、历时46-48小时)静脉连续输注。这样的治疗每2周重复一次。
另一实施方式中,2小时400mg/m2亚叶酸输注继以46小时2400mg/m2的5-fu输注。在第一天还包括2小时130mg/m2奥沙利铂输注。这样的治疗每两周重复一次。
根据本发明的方法,辐照可以与抗体-细胞因子阱分子联合给药来治疗癌症。放疗即辐照治疗通常采用高能粒子束或波例如x射线和γ射线通过诱导细胞dna突变来杀灭癌细胞。癌细胞分裂比正常细胞快,这使得肿瘤组织比正常组织更容易受到辐照作用。可以对患者施用各种类型的辐照只要辐照剂量是患者能够耐受的且没有不可接受的不良副作用。合适的放疗类型包括例如电离辐照(例如x射线、γ射线或高能线性辐照(highlinearenergyradiation))。电离辐照定义为包含粒子或光子的辐照,所述粒子或光子具有足够的能量以产生电离,即得电子或失电子。辐照的效果可至少部分地由临床医生控制。对辐照的剂量以进行划分为宜,为的是令尽可能多的靶细胞受到辐照同时降低毒性。辐照可与辐照增敏剂或辐照防护剂(如il-1或il-6)同时施用,辐照增敏剂增强肿瘤细胞杀伤效果,辐照防护剂保护健康组织免受辐照伤害。类似地,升温即体温过高或化疗也会令组织对辐照敏感。
辐照源可以在患者外部或内部。外部放疗是最常见的,一般包括使用例如线性加速器将一束高能辐射(粒子束)透过皮肤引导到肿瘤部位。虽然辐射束局限于肿瘤部位但几乎不可能避免触及正常健康组织。然而,外部辐照通常患者耐受良好。
另一个例子中,辐照是从患者外施以伽马(γ)射线。伽马射线是放射性同位素例如钴60分解产生的。采用称为立体定向放射治疗(sbrt)的治疗方法可以将伽马射线密集聚焦而仅靶向肿瘤组织,由此使得对健康组织损伤非常小。sbrt可用于局灶性肿瘤患者。另一方面,粒子加速器产生的x射线可用于对身体更大区域进行辐照。
内部放疗涉及在肿瘤部位处或附近体内植入辐照发射源(如珠、线、丸、胶囊等)。所用辐照来自放射性同位素,例如但不限于碘、锶、磷、钯、铯、铱、磷酸盐或钴。这些植入物可在治疗后取出或以非活性状态留在体内。内部放疗的类型包括但不限于近距离放疗、间质辐照和腔内辐照。目前较少见的内部放疗形式涉及放射性同位素生物载体,例如放射免疫疗法中使用的,其中,给予患者与放射性物质结合的肿瘤特异性抗体。抗体结合肿瘤抗原从而有效地将辐照剂量施用于相关组织。
放疗可用作方案的组成之一来控制原发性肿瘤的生长(见,例如comphausen等,(2001)“对原发肿瘤的放疗在小鼠中加速转移性生长”,cancerres.61:2207-2211)。尽管放疗单独治疗癌症可能不太有效,但是将辐照与本文所述抗pd-l1/tgfβ阱分子联合能够增强放疗的局部和全身性功效。
由于辐照杀伤免疫效应细胞,辐照的剂量和时间非常重要。辐照后肿瘤中的t细胞和树突细胞在辐照后立即降低,但t细胞水平反弹高出基线水平。无论施用方法如何,完整的辐照日剂量可以在一天内有疗程地给予。较好的是,总剂量划分成数日来给予。因此,辐照日剂量可包含大约1-50gy/天,例如至少1、至少2、至少3、1-4、1-10、1-20、1-50、2-4、2-10、2-20、2-25、2-50、3-4、3-10、3-20、3-25、3-50gy/天。
日剂量可单剂施用,也可在按日疗程期间分两份或更多个份以“微量划分”剂量给予。采用内部辐照源时(例如近距离放疗或放射免疫疗法),暴露时间通常会增加,辐照强度相应降低。
根据本发明的一些实施方式,抗体-tgfβ阱与至少一种其他抗癌药物同时施用。根据本发明的一些实施方式,抗体-tgfβ阱与至少一种其他抗癌药物依序施用。
抗体-tgfβ阱和至少一种其他抗癌药物的给药频率可在疗程期间根据用药医师的判断进行调整。
实施例
以上是对本发明的一般性说明,参考以下实施例将更容易理解,所述实施例仅用于说明本发明的某些方面和实施方案,无意于以任何方式限制本发明的范围。
实施例1–dna构建和蛋白质表达
抗pd-l1/tgfβ阱即抗pd-l1抗体-tgfβii型受体融合蛋白。该分子的轻链与抗pd-l1抗体的轻链(seqidno:1)相同。该分子的重链(seqidno:3)是融合蛋白,包含抗pd-l1抗体的重链(seqidno:2)通过柔性的(gly4ser)4gly接头(seqidno:11)在基因水平上与可溶性tgfβii型受体(seqidno:10)的n末端融合。在融合连接处,抗体重链的c末端赖氨酸残基突变为丙氨酸来减少蛋白酶裂解。为了表达抗pd-l1/tgfβ阱,用处于同一表达载体上或分处不同表达载体上的编码抗pd-l1轻链的dna(seqidno:4)和编码抗pd-l1/tgfβii型受体的dna(seqidno:5)采用瞬时或稳定转染标准方案转染哺乳动物细胞。收获条件培养基并通过标准蛋白a琼脂糖凝胶色谱法纯化抗pd-l1/tgfβ阱融合蛋白。纯化蛋白包含一分子抗pd-l1抗体和两分子可溶性tgfβii型受体(图1),在尺寸排阻色谱和非还原sds-聚丙烯酰胺电泳上测得的估计分子量(mw)约190千道尔顿。还原条件下轻链和重链的表观mw分别是28和75千道尔顿。
如此制备抗pd-l1(突变)/tgfβ阱融合蛋白,其含有相似的重链融合多肽(seqidno:7)和具有可变区内a31g、d52e、r99y突变的轻链,上述突变消除pd-l1(seqidno:6)结合性。该融合蛋白在后序实验中用作tgfβ阱的对照。
实施例2-生物治疗剂抗pd-l1/tgfβ阱的生产
通过瞬时转染人胚肾293(hek)细胞产生的抗pd-l1/tgfβ阱被发现含有不同程度的剪切产物,在还原sds-page显示为表观mw约60kd的一条弱带。该条带被证实是抗pd-l1/tgfβ阱重链在tgfβriin-末端部分靠近融合连接点的位置的剪切产物。
在cho-s宿主细胞系中产生表达抗pd-l1/tgfβ阱的稳定克隆,其经预适应能在无血清培养基中悬浮培养生长。用含有编码抗pd-l1-tgfβrii蛋白的基因和谷氨酰胺合成酶选择标记的表达载体转染细胞。随后用l-甲硫氨酸砜亚胺(msx)挑选稳定的整合子。使用小池(minipool)法产生抗pd-l1/tgfβ阱表达细胞系,然后用beckton-dickinson荧光激活细胞分选仪(facsariaii)将单细胞沉积到384孔板中。在通用平台补料分批测试中评估生长情况、生产力和蛋白质质量。基于这些分析,挑选了14个克隆作为先导候选者接受进一步研究。对这些克隆进行了稳定性研究,从克隆放大期间建立的研究细胞库进行至约90pdl(群体倍增水平)。小池研发结束时发现重链-接头-tgfβrii亚基发生了剪切,就如瞬时表达中所见。稳定性研究中的所有克隆产生了剪切产物,尽管经蛋白a纯化的材料中显示相对于完整亚基的剪切产物百分比各克隆各不相同。此外,开发了一种改进的纯化方法,包括蛋白a色谱法后继以强阳离子交换,用以减少剪切产物的共纯化。即使采用改进工艺,也只有采用发生低水平剪切的克隆才能得到最终剪切产物水平按要求<5%的纯化材料。基于这些组合分析,挑选克隆02b15作为最终候选克隆。对该克隆在0pdl、30pdl、60pdl和90pdl时表达的抗pd-l1/tgfβ阱经分析显示剪切百分比没有随着群体倍增水平而升高。
实施例3-皮下mc38肿瘤小鼠模型中化疗与抗pd-l1/tgfβ阱的联合治疗
结直肠癌(crc)是男性中第三大、女性中第二大最常见的癌症,全世界有超过120万新发病患。尽管过去十年中治疗取得了重大进展,但crc仍是第四大最常见的癌症相关死因。因此,需要新的治疗方式。在以下实施例中,研究了抗pd-l1/tgfβ阱分子联合基于奥沙利铂(ox)和5-氟尿嘧啶(5-fu)的治疗在结直肠癌小鼠模型中的功效。
在有皮下mc38肿瘤的小鼠中,抗pd-l1/tgfβ阱与化疗药ox/5-fu联合治疗显著抑制肿瘤生长。这些临床前数据支持将化学疗法(ox/5-fu)与抗pd-l1/tgfβ阱免疫疗法联合用于临床治疗结直肠癌的策略。
材料和方法
mc38肿瘤细胞系获自美国典型培养物保藏中心(atcc)。mc38细胞系经测试证实无外来病毒无支原体。8-12周龄的c57bl/6小鼠获自查尔斯河实验室(charlesriverlaboratories)。8-12周龄的b6.129s2-ighmtm1cgn/j小鼠获自杰克逊实验室(jacksonlaboratories)。
试验品剂量如下:抗pd-l1/tgfβ阱:24.6mg/kg;492μg/鼠;2.46mg/ml;0.2ml剂量体积静脉给药。氟尿嘧啶(5-fu):60.0mg/kg;120μg/鼠;6.00mg/ml;0.02剂量体积静脉给药。奥沙利铂:5.0mg/kg;10μg/鼠;0.500mg/ml;0.02ml腹膜内给药。
以“mg/kg”表示的是假设平均体重为每只小鼠20克的近似值。
阴性对照是无活性同型对照(抗pd-l1(突变))按400μg/小鼠的试验浓度给药。细胞培养.mc38细胞于无菌条件下培养在达尔伯克必需基本培养基(dmem)中,培养基含10%热灭活胎牛血清,维持37℃和5%co2。待长到50-70%融合度时按1:5的比例进行细胞传代,在体内植入之前共传代2次。通过胰蛋白酶消化收获细胞,用血细胞计数器和台盼蓝排除染色来测定活细胞数。全部细胞培养所用试剂购自生命技术(lifetechnologies)(盖瑟斯堡,md)。
mc38肿瘤模型.ti13-027研究中,将mc38肿瘤细胞(1×106细胞/鼠)在100μl无菌pbs中制成悬液,然后植入c57bl/6小鼠右侧腹。待肿瘤尺寸达到~45mm3的平均值,将小鼠随机分成4组(n=10小鼠/组)启动治疗。治疗的实施按照图2和图3中的给药方案进行。用数字卡尺测量肿瘤的长(l)、宽(w)、高(h),并用winwedge软件自动记录到计算机上,每周两次。且每周两次记录体重评估耐受性。用ellipsoid体积公式计算肿瘤体积:体积=π/6*(lxwxh);其中,l=肿瘤长度,w=肿瘤宽度,h=肿瘤高度。通过如下所述在体内研究期间全程检测肿瘤体积并在研究终点测量肿瘤重量来确定功效。在第17天处死全部动物,切取肿瘤并称重。采集脾脏用于ifn-γelispot分析。
ti14-012研究中,如上所述将mc38肿瘤细胞注射到b6.129s2-ighmtm1cgn/j小鼠中。用于评估肿瘤生长和治疗功效的所有其他程序也如上所述。
ifn-γelispot检测.用酶联免疫吸附斑点(elispot)检测来测定针对p15e抗原的细胞毒t淋巴细胞(ctl)反应,已知p15e抗原是mc38肿瘤内的t细胞排斥表位(yang和perry-lalley,jimmunotherapy2000;23:177-183)。elispot检测测定与载有p15e表位kpswfttl的抗原呈递细胞(apc)共培养后产ifn-γcd8+t细胞的频率。载有无关肽siinfekl(来自鸡卵清蛋白(ova))的apc作为阴性对照。用pma和离子霉素刺激阳性对照样品,引发细胞毒性t淋巴细胞的非特异性活化。elispot检测用bd生物科学(bdbiosciences)的小鼠ifn-γelispot试剂盒按照制造商的说明来进行。ti13-027研究的第17天,收获脾脏(n=5鼠/组)加工成单细胞悬液,用终浓1μg/ml的p15e肽刺激,然后37℃培养7天。在体外刺激后,用cd8+t细胞分离试剂盒(美天旎生物技术(miltenyibiotech))和automacspro分离器通过磁激活细胞分选分离cd8+t细胞。为了建立用于体外刺激elispot检测的共培养系统,来自从未接受过处理的(naive)小鼠脾细胞的apc用kpswfttl肽或不相关的siinfekl肽冲击1小时,然后在gammacell40辐照仪中接受2gy辐照。分离自实验小鼠的cd8+t细胞(1x105个细胞/孔)与经肽冲击和辐照的apc(2.5x105细胞/孔)在elispot检测板(抗ifn-γ抗体包被)中培养,一式三份。
ti14-012研究的第18天,建立离体elispot检测,其中收获脾脏(n=5鼠/组)加工成单细胞悬液,用cd8+t细胞分离试剂盒(美天旎生物技术(miltenyibiotech))和automacspro分离器通过磁激活细胞分选分离cd8+t细胞。为了建立用于离体elispot检测的共培养系统,来自从未接受过处理的小鼠脾细胞的apc用kpswfttl肽或不相关的siinfekl肽冲击1小时,然后在gammacell40辐照仪中接受2gy辐照。。分离自实验小鼠的cd8+t细胞(5x105个细胞/孔)与经肽冲击和辐照的apc(5x105细胞/孔)在elispot检测板(抗ifn-γ抗体包被)中培养,一式三份。
两个实验中,实验cd8+t细胞都与经肽冲击的apc在37℃共培养19-20小时后才从测定板中移出。将生物素化的抗ifn-γ抗体加入板的各个孔中,洗涤后加入链霉亲和素-hrp检测缀合物。再次洗涤后,将板与生色底物溶液一起温育,监测反应,然后通过用水洗板终止反应。用ctl-免疫斑点s5uv分析仪(细胞技术有限公司(cellulartechnologylimited))计数检测板各孔中的ifn-γ阳性斑点数。数据表示为平均斑点数/孔±sem。
在研究期间每天检查死亡率。临床观察.使用此前所述的身体状况(bc)评分系统,在研究期间每天一次记录所有动物的临床体征(例如健康不良和行为改变)(ullman-cullere和foltz,labanimsci.1999;49:319-23)。通过co2窒息对濒死小鼠施以人道安乐死。研究中所有动物每周两次记录体重,包括每组的终止日。实验期间用数字卡尺测量肿瘤的三维尺寸并用winwedge软件自动记录到计算机上,每周两次。用ellipsoid体积公式计算肿瘤体积:体积=0.5236(lxwxh);其中,l=肿瘤长度,w=肿瘤宽度,h=肿瘤高度。在处死时切取原发性肿瘤并称重作为次要效果终点。用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。小鼠fn-γelispot检测用小鼠fn-γelispot试剂盒(bd生物科学)按照制造商的说明进行。
统计学分析.在整个研究期间每周两次测量肿瘤体积。肿瘤体积数据表示为平均值±平均值标准误差(sem)。肿瘤生长抑制%t/c比的计算为:治疗组的肿瘤体积除以对照组的肿瘤体积再乘以100。将肿瘤体积数据转换为对数(log),并进行带多重比较tukey修正的双向重复anova来测定处理组之间的统计学差异。t/c比的计算为:治疗组的肿瘤体积除以对照组的肿瘤体积。研究结束时测量肿瘤重量。数据表示为平均值±sem。%t/c比的计算为:治疗组的肿瘤重量除以对照组的肿瘤重量再乘以100。用带多重比较tukey修正的单向anova分析肿瘤重量数据以测定处理组之间的统计学差异。ifn-γelispot数据表示为平均值±sem。用graphpadprism软件通过带多重比较tukey修正的单向anova进行统计学分析。将p<0.05确定为具有统计学显著性。
结果
ti13-027研究中,由于完全人源化抗体在b细胞感受态小鼠中引起免疫原性,c57bl/6野生型小鼠在一周内只能接受三次抗pd-l1/tgfβ阱分子给药。因此,抗pd-l1/tgfβ阱单一疗法未观察到显著的抗肿瘤活性(据肿瘤体积的%t/c=91%,见图4)。与同型对照相比,奥沙利铂/5-fu治疗在mc38皮下肿瘤模型中引起显著的肿瘤生长抑制(据肿瘤体积的%t/c=53.2%;p<0.0001)。与对照组相比,抗pd-l1/tgfβ阱与奥沙利铂/5-fu联合治疗显著抑制mc38肿瘤生长(据肿瘤体积的%t/c=33.2%;p<0.0001)。并且,相比奥沙利铂/5-fu单用,抗pd-l1/tgfβ阱联合奥沙利铂/5-fu显著提高肿瘤生长控制(439.6mm3与703.7mm3(肿瘤体积)相比;p<0.0001)。观察到相同的趋势即与抗pd-l1/tgfβ阱单用相比联合治疗统计学显著性地提高肿瘤生长控制(439.6mm3与1204.0mm3相比;p<0.0001)(见图4a-d和表1)。
最后,据elispot检测,与同型对照组相比,观察到抗pd-l1/tgfβ阱单一疗法或奥沙利铂/5-fu单一疗法显著提高ifn-γ生产性cd8+t细胞的频率(分别为p<0.05和p<0.05)。与各个单一疗法组相比,抗pd-l1/tgfβ阱联合奥沙利铂/5-fu显著提高p15e特异性ifn-γ生产性cd8+t细胞的频率(p<0.05,见图4c)。
ti14-012研究中,用b细胞缺陷型小鼠来避免小鼠抗体抗人抗体(maha)反应,实验动物接受5次抗pdl1/tgfβ阱处理。不出所料,与ti13-027研究(其中野生型小鼠仅接受三次处理,据肿瘤体积的%t/c=91%)相比,抗pdl1/tgfβ阱单一疗法表现出更高的抗肿瘤活性(据肿瘤体积的%t/c=57.3%)。t114-012研究中,与两种单一疗法相比(p<0.0001)或与同型对照相比(p<0.0001),抗pd-l1/tgfβ阱与奥沙利铂/5-fu联合治疗显著地更为有效(见图5d和表2)。
类似地,与同型对照相比,抗pd-l1/tgfβ阱单一疗法显著提高ifn-γ生产性cd8+t细胞的频率(见图5c,p<0.05)。与单一疗法组或同种型对照组相比,抗pd-l1/tgfβ阱与奥沙利铂/5-fu联合治疗引起p15特异性ifn-γ生产性cd8+t细胞频率协效性提高(见图5c;p<0.05)。
抗pd-l1/tgfβ阱是一种双功能抗体-细胞因子受体融合蛋白,旨在通过对pd-1/pd-l1轴和tgfβ信号传导的双重靶向来逆转肿瘤微环境中的细胞内在和外在免疫抑制。在本文所述的研究中,在带有皮下mc38肿瘤的小鼠中观察到抗pdl1/tgfβ阱与ox/5-fu联合治疗显著的mc38肿瘤生长抑制和对p15e特异性cd8+t细胞ifn-γ生产的协效性诱导。在野生型小鼠中观察到的这些对抗肿瘤效力和免疫应答的影响在b细胞缺陷型小鼠中更为突出。据信,这样的差异主要是由于在b细胞缺陷型小鼠中没有maha(鼠抗人抗体)反应而给予了更多剂次的抗pd-l1/tgfβ阱。综合起来,这些数据表明,在小鼠结直肠癌模型中,化疗成分(ox/5-fu)能与抗pdl1/tgfβ阱治疗有效联合增强肿瘤生长抑制和肿瘤反应性cd8+t细胞反应。综上,以上临床前结果支持联合疗法临床用于治疗结直肠癌。
实施例4-肌内mc38肿瘤小鼠模型中放疗与抗pd-l1/tgfβ阱的联合治疗
抗pd-l1/tgfβ阱分子中人tgfβrii的胞外域(tgfβ阱)与人抗pd-l1抗体重链c-末端共价连接。抗pd-l1/tgfβ阱单一疗法在多种临床前模型中显示出优异的抗肿瘤效力。在此报道的本研究中,我们在带有肌内mc38结直肠肿瘤的b6.129s2-ighmtmlcgn/j小鼠中研究了抗pd-l1/tgfβ阱联合分次局部放疗的抗肿瘤活性。数据显示,四次分量局部辐照(360拉德/剂)联合一次抗pd-l1/tgfβ阱给药(55μg)具有显著的协效性抗肿瘤效果,在100%的小鼠中获得肿瘤缓解。此外,四次分量局部辐照(500拉德/剂)联合一次抗pd-l1/tgfβ阱(164μg)给药激发对距离被照射肿瘤远处肿瘤的抗癌作用,这是远位效应的表现并提示这样的治疗可用于治疗转移。相比之下,单独放疗或单独抗pd-l1/tgfβ阱治疗获得中等的肿瘤负荷降低。并且,在接受联合治疗的小鼠中观察到p15e特异性ifn-γ生产性cd8+t细胞频率显著升高。最后,联合治疗与效应cd8+t细胞和nk细胞对mc38肿瘤浸润改善有关联。这些结果表明抗pd-l1/tgfβ阱处理与放疗以协同作用方式促进t细胞介导的抗肿瘤反应。下文描述的结果支持这一认定即这一联合策略具有临床应用前景。
材料和方法
细胞系:mc38鼠结肠癌细胞系由斯克利普斯研究所(scrippsresearchinstitute)惠赠。细胞系经测试证实无鼠病毒无支原体。8-12周龄的b6.129s2-ighmtm1cgn/j小鼠(c57bl/6)获自杰克逊实验室(jacksonlaboratories)。
试验材料剂量如下所述:抗pd-l1/tgfβ阱:2.75mg/kg;55μg/小鼠;13.75mg/ml;0.2ml剂量体积静脉给药和抗pd-l1/tgfβ阱:8.25mg/kg;164μg/小鼠;41.25mg/ml;0.2ml剂量体积静脉给药。
阴性对照如下:无活性同型对照(抗pd-l1(突变)a11-121-6)按133μg/小鼠或45μg/小鼠的试验浓度给药。
mc38细胞于无菌条件下培养在达尔伯克必需基本培养基(dmem)中,培养基含10%热灭活胎牛血清,维持37℃和5%co2。待长到50-70%融合度时按1:5的比例进行细胞传代,在体内植入之前共传代2次。通过胰蛋白酶消化收获细胞,用血细胞计数器和台盼蓝排除染色来测定活细胞数。
c38肿瘤模型:在第-8天,在c57bl/6.129s2-ighmtm1cgn/j小鼠右侧大腿肌内植入0.1mlpbs中的0.5x106活mc38肿瘤细胞。当肿瘤达到~128mm3的平均体积时,将小鼠随机分成不同处理组。处理从第0天(肿瘤细胞接种后8天)开始。
本灶放疗可在远处部位引发抗癌效果,这种现象称为“远位”效应。为了测试抗pd-l1/tgfβ阱对放疗远位效应的影响,在治疗前7天,给小鼠在右侧大腿接种0.5x106个活mc38肿瘤细胞形成原发性肌内mc38肿瘤,左侧腹皮下注射1×106个mc38细胞形成继发性皮下mc38肿瘤(图9a)。处理从第7天开始。
放疗:小鼠在专用有机玻璃托盘上定位,除了要照射的肿瘤区域外,全身用铅屏蔽保护。用gammacell40辐照仪对肿瘤区域进行放疗。
酶联免疫吸附斑点(elispot)检测:用elispot检测来测定针对p15e抗原的细胞毒t淋巴细胞(ctl)反应,已知p15e抗原是mc38肿瘤内的t细胞排斥表位(yang和perry-lalley,2000)elispot检测测定与载有p15e表位kpswfttl的抗原呈递细胞(apc)共培养后ifn-γ生产性cd8+t细胞的频率。载有来自鸡卵清蛋白的无关肽(siinfekl)的apc作为阴性对照。用pma和离子霉素刺激阳性对照样品,引发ctl的非特异性活化。elispot检测用bd生物科学(bdbiosciences)的试剂盒进行。研究第14天,收获每组一鼠的脾脏加工成单细胞悬液。用cd8+t细胞分离试剂盒(美天旎生物技术(miltenyibiotech))和automacspro分离器通过磁激活细胞分选分离cd8+t细胞。然后将cd8+t细胞接种到elispot检测板(抗ifn-γ抗体包被)中与经kpswfttl肽冲击1小时并经gammacell40辐照仪2gy照射的源自从未接受过处理的小鼠脾细胞的apc共培养。37℃培养16-20小时后,将细胞从检测板中移出。将生物素化的抗ifn-γ抗体加入板的各个孔中,洗涤后加入链霉亲和素-hrp检测缀合物。再次洗涤后,将板与生色底物溶液一起温育,监测反应,然后通过用水洗板终止反应。用免疫斑点elispot读板仪系统测定检测板各孔中的ifn-γ阳性斑点数。
免疫表型:通过机械破碎然后裂解红细胞从脾脏制备细胞悬浮液。通过对精细研磨的肿瘤浆液进行酶消化来制备肿瘤细胞悬浮液。浆液在iv型胶原酶(400单位/ml)和dnase1(100μg/ml)的溶液中于37℃温育1小时,在此期间频繁搅拌。肿瘤降解后,沉降分离碎片,悬浮液过40μm尼龙细胞过滤器过滤。按照制造商(如e生物科学(ebioscience)或bd生物科学)的推荐进行脾细胞悬浮液和肿瘤细胞悬浮液的抗体染色以便facs分析。
对于脾脏样品的分析,围绕由前向和侧向散射特征鉴定的淋巴细胞群设定亲本门(parentalgate)。由此淋巴细胞门在点图上鉴定出免疫细胞亚群:辅助t细胞(cd4+),细胞毒性t淋巴细胞(cd8+),nk细胞(nk1.1+),效应记忆cd8+t细胞(cd8+/cd44高/cd62l低),中枢记忆cd8+t细胞(cd8+/cd44高/cd62l高)和调节性t细胞(cd4+/cd25+/foxp3+)。为了评估脱粒作为裂解活性的量度,检测淋巴细胞表面上的cd107a。细胞表面蛋白染色之后,固定样品并透化以便t盒转录因子(eomes和t-bet)和效应细胞因子(ifn-γ和粒酶b)的细胞内染色。由白细胞门在点图上鉴定出免疫细胞亚群:树突细胞(cd11c+/i-ab+),中性粒细胞(cd11b+/ly6g+),巨噬细胞(cd11b+/ly-6c高)和mdscs(gr-1+/cd11b+)。
用类似的门控策略分析肿瘤样品,不同的是首先围绕cd45+细胞群设定亲本门来鉴定来自其他肿瘤细胞和基质组分的肿瘤浸润性白细胞。
研究设计:ti13-109放疗联合抗pd-l1/tgfβ阱在b细胞缺陷型小鼠mc38模型中。分组处理(n=10)。
第1部分:效力
第2部分:elispot检测:在第14天,处死全部小鼠并通过elispot检测分析亚组(n=5只小鼠/组)的功能性反应。如前文所述收获脾脏并加工用于elispot检测。用免疫斑点elispot读板仪系统测定检测板各孔中的ifn-γ阳性斑点数。
研究设计:ti14-013:放疗联合抗pd-l1/tgfβ阱在b细胞缺陷型小鼠mc38模型中。分组处理(n=10)。
第1部分:效力
第2部分:elispot检测和免疫表型:在第14天,处死全部小鼠对亚组(n=5只小鼠/组)进行通过elispot检测的脾功能性反应分析和til免疫表型分析。如前文所述收获脾脏并加工用于elispot检测。用免疫斑点elispot读板系统测定检测板各孔中的ifn-γ阳性斑点数。如前文所述收获肿瘤组织进行加工。通过facs分析%cd8+til、%nk1.1+til、cd8+tileomes表达和cd8+til脱粒来分析til表型。
使用此前所述的身体状况(bc)评分系统,在研究期间每天一次记录所有动物的临床体征(例如健康不良和行为改变)(ullman-cullere和foltz,labanimsci.1999;49:319-23)。通过co2窒息对濒死小鼠施以人道安乐死。研究中所有动物每周两次记录体重,包括每项研究的终止日。实验期间用数字卡尺测量肿瘤的三维尺寸。用以下公式计算肿瘤体积:体积=0.5236(lxwxh);其中,l=肿瘤长度,w=肿瘤宽度,h=肿瘤高度。生成kaplan-meier存活曲线以量化从肿瘤接种到处死的时间间期并计算每个处理组的中值存活时间。
用elispot检测对ifn-γ生产性p15e-特异性cd8+t细胞的频率进行定量检测。通过facs(荧光激活细胞分选)分析脾细胞和肿瘤浸润淋巴细胞(til)的免疫表型。
统计学分析:在整个研究期间每周两次测量肿瘤体积。肿瘤体积数据表示为平均值±平均值标准误差(sem)。将肿瘤体积数据转换为对数(log),并进行带多重比较tukey修正的双向重复anova来测定处理组之间的统计学差异。研究结束时采集肿瘤重量。数据表示为平均值±sem。t/c比的计算为:治疗组的肿瘤体积(或肿瘤重量)除以对照组的肿瘤体积(或肿瘤重量)。用带多重比较tukey修正的单向anova对肿瘤重量数据进行评测来测定处理组之间的统计学差异。用elispot检测对ifn-γ生产性cd8+t细胞的频率进行定量测定并表示为每孔斑点数平均值(平均值±sem)。用graphpadprism软件通过带多重比较tukey修正的单向anova进行统计学分析。将p<0.05确定为具有统计学显著性。
研究设计:放疗与抗pd-l1/tgfβ阱联合治疗在b细胞缺陷型小鼠mc38模型中的远位效应测试。分组处理(n=6)。处理自第0天开始:同型对照(400μg,第0、2、4天),辐照(500拉德/天,第0、1、2、3天),抗pd-l1/tgfβ阱(164μg,第0天),抗pd-l1/tgfβ阱(164μg,第0天)+辐照(500拉德/天,第0、1、2、3天)。只对原发肿瘤进行照射,如图9a所示。每周两次测量原发肿瘤体积和继发肿瘤体积。肿瘤体积表示为平均值±sem。
结果
辐照联合抗pd-l1/tgfβ阱表现出协效性抗肿瘤功效。在mc38肌内肿瘤模型(ti13-109)中,辐照(360拉德/天,第0-3天)或抗pd-l1/tgfβ阱单一疗法(55或164μg,第2天)诱导显著的肿瘤生长抑制(各自p<0.0001,相对于同型对照),但是在第10天,辐照联合抗pd-l1/tgfβ阱相比辐照(p<0.0001)或抗pd-l1/tgfβ阱(p<0.0001)单一疗法诱导显著的疗效性协同效应(见图6a)。放疗的第14天肿瘤重量t/c比是0.45,55μg和164μg抗pd-l1/tgfβ阱的分别是0.50和0.36,辐照联合抗pd-l1/tgfβ阱(分别55μg比164μg)组的则是0.04比0.01(见图6b)。早在pd-l1/tgfβ阱处理后4天在50%(20只中的10只)接受抗pd-l1/tgfβ阱单一治疗的小鼠中,100%(20只中的20只)接受联合治疗的小鼠中,仅10%(10只中的1只)接受辐照单一治疗的小鼠中观察到肿瘤消退。并且,由于抗pd-l1/tgfβ阱仅给予了一剂,抗pd-l1/tgfβ阱单一治疗组中10个消退肿瘤中有4个再度生长,而联合组中全部消退肿瘤都持续缩小直至第14天小鼠被处死接受免疫功能评估的时候。
免疫激活与抗肿瘤功效相关。在第14天,处死小鼠并使用离体elispot检测对ifn-γ生产性肿瘤反应性(p15e)cd8+t细胞的频率进行定量检测(见图6c)。在辐照和抗pd-l1/tgfβ阱单一治疗组中仅观察到ifn-γ生产性肿瘤反应性cd8+t细胞的中度诱导(与同型对照相比分别为p>0.05和p<0.05)。与观察到的抗肿瘤效果一致;然而,用联合疗法处理的小鼠发生了p15e特异性ifn-γ生产性cd8+t细胞频率的协效性诱导(图6c)。联合治疗诱导的cd8+t细胞ifn-γ生产比同型对照高7倍,比单一疗法高至少5倍(与两单一疗法相比均p<0.001)。该项研究(ti13-109)中,联合治疗中将抗pd-l1/tgfβ阱的剂量从55μg提高至164μg没有进一步加速肿瘤消退。由于高剂量组中cd8+t细胞的低产量,因此无法对ifn-γ生产性肿瘤反应性(p15e)cd8+t细胞的频率进行充分评估。因此,进行了重复研究以确保结果一致性。
用低剂量的55μg抗pd-l1/tgfβ阱重复实验(ti14-013),联合治疗在肿瘤生长抑制和p15特异性cd8+t细胞ifn-γ生产诱导上得到了近乎相同的协同效应(见图7a-c)。
对ti14-013研究中接受治疗小鼠的肿瘤浸润性淋巴细胞分析显示辐照和抗pd-l1/tgfβ阱联合处理后cd8+til和nk细胞的频率相比单一疗法组或同型对照组升高(见图8a-b)。另外的分析显示,辐照联合抗pd-l1/tgfβ阱的治疗促进肿瘤浸润性cd8+t细胞上t-box转录因子eomes的表达和脱粒(cd107a)(见图8c-d)。
相比单独的抗pd-l1/tgfβ阱或辐照,辐照与单剂抗pd-l1/tgfβ阱联合治疗减小原发肿瘤的体积(两项比较都p<0.0001,第14天)(图9b)。然而,相比单独的抗pd-l1/tgfβ阱或辐照,联合治疗还减小继发肿瘤的体积(分别为p=0.0066和p=0.0006,第14天)(图9c)。值得注意的是,单独的辐照和单剂低剂量抗pd-l1/tgfβ阱都没有相比同型对照处理显著抑制继发性肿瘤生长,这表明抗pd-l1/tgfβ阱与辐照协同作用诱导了远位效应。
与观察到的抗肿瘤功效和肿瘤反应性ifn-γ生产性cd8+t细胞频率上的协同效果以及cd8+t细胞和nk细胞浸润增强一致的是辐照与抗pd-l1/tgfβ阱分子联合治疗深广的先天性和过继性抗肿瘤免疫应答诱导。因此,这种联合治疗具有临床相关应用,即用于改善癌症患者的放疗。
此处的数据表明,在mc38结直肠癌模型中,作为标准护理的外放射线治疗(ebrt)可以与抗pd-l1/tgfβ阱治疗联合实现协效性肿瘤生长抑制和肿瘤反应性cd8+t细胞应答。由于p15e是mc38肿瘤细胞系表达的内源性逆转录病毒抗原(zeh等,jimmunol.1999;162:989-94),观察到的p15e特异性ifn-γ生产性cd8+t细胞增加是联合治疗后的肿瘤反应性而非泛化t细胞反应。该协效性免疫反应与该治疗方案中观察到的抗肿瘤效力增强一致,这表明用辐照与抗pd-l1/tgfβ阱联合的治疗促进cd8+t细胞介导的抗肿瘤应答。放疗和抗pd-l1/tgfβ阱治疗联合还表现为促进cd8+t细胞和nk细胞的mc38肿瘤浸润。此外,据转录因子eomes和脱粒标记物cd107a表达水平升高可见,联合治疗诱导效应cd8+til表型。
观察到的结果支持这一认定即该联合策略的潜在临床应用具有协同效应。放疗和抗pd-l1/tgfβ阱的依序治疗对于放疗后循环tgfβ水平升高的患者将是有益的。此外,由于单剂低量抗pd-l1/tgfβ阱也观察到强烈的协同效应,预期这种联合疗法在临床中具有良好的安全性。
序列
seqidno:1
分泌的抗pd-l1λ轻链肽序列
qsaltqpasvsgspgqsitisctgtssdvggynyvswyqqhpgkapklmiydvsnrpsgvsnrfsgsksgntasltisglqaedeadyycssytssstrvfgtgtkvtvlgqpkanptvtlfppsseelqankatlvclisdfypgavtvawkadgspvkagvettkpskqsnnkyaassylsltpeqwkshrsyscqvthegstvektvaptecs
seqidno:2
分泌的抗pdl1h链的肽序列
evqllesggglvqpggslrlscaasgftfssyimmwvrqapgkglewvssiypsggitfyadtvkgrftisrdnskntlylqmnslraedtavyycariklgtvttvdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk
seqidno:3
分泌的抗pdl1/tgfβ阱h链的肽序列
evqllesggglvqpggslrlscaasgftfssyimmwvrqapgkglewvssiypsggitfyadtvkgrftisrdnskntlylqmnslraedtavyycariklgtvttvdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgaggggsggggsggggsggggsgipphvqksvnndmivtdnngavkfpqlckfcdvrfstcdnqkscmsncsitsicekpqevcvavwrkndenitletvchdpklpyhdfiledaaspkcimkekkkpgetffmcscssdecndniifseeyntsnpd
seqidno:4
抗pd-l1λ轻链从翻译起始密码子到翻译终止密码子的dna序列(vl之前的前导序列是来自尿激酶纤溶酶原激活物的信号肽)
atgagggccctgctggctagactgctgctgtgcgtgctggtcgtgtccgacagcaagggccagtccgccctgacccagcctgcctccgtgtctggctcccctggccagtccatcaccatcagctgcaccggcacctccagcgacgtgggcggctacaactacgtgtcctggtatcagcagcaccccggcaaggcccccaagctgatgatctacgacgtgtccaaccggccctccggcgtgtccaacagattctccggctccaagtccggcaacaccgcctccctgaccatcagcggactgcaggcagaggacgaggccgactactactgctcctcctacacctcctccagcaccagagtgttcggcaccggcacaaaagtgaccgtgctgggccagcccaaggccaacccaaccgtgacactgttccccccatcctccgaggaactgcaggccaacaaggccaccctggtctgcctgatctcagatttctatccaggcgccgtgaccgtggcctggaaggctgatggctccccagtgaaggccggcgtggaaaccaccaagccctccaagcagtccaacaacaaatacgccgcctcctcctacctgtccctgacccccgagcagtggaagtcccaccggtcctacagctgccaggtcacacacgagggctccaccgtggaaaagaccgtcgcccccaccgagtgctcatga
seqidno:5
从翻译起始密码子至翻译终止密码子的dna序列(mvksp前导序列:小写下划线;vh:大写;含k至a突变的igg1m3:小写;(g4s)x4-g接头:粗体大写;tgfβrii:粗体下划线小写;两个终止密码子:粗体下划线大写)
atggaaacagacaccctgctgctgtgggtgctgctgctgtgggtgcccggctccacaggcgaggtgcagctgctggaatccggcggaggactggtgcagcctggcggctccctgagactgtcttgcgccgcctccggcttcaccttctccagctacatcatgatgtgggtgcgacaggcccctggcaagggcctggaatgggtgtcctccatctacccctccggcggcatcaccttctacgccgacaccgtgaagggccggttcaccatctcccgggacaactccaagaacaccctgtacctgcagatgaactccctgcgggccgaggacaccgccgtgtactactgcgcccggatcaagctgggcaccgtgaccaccgtggactactggggccagggcaccctggtgacagtgtcctccgctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagagagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaggagcagtacaacagcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctatagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtccccgggtgctggcggcggaggaagcggaggaggtggcagcggtggcggtggctccggcggaggtggctccggaatccctccccacgtgcagaagtccgtgaacaacgacatgatcgtgaccgacaacaacggcgccgtgaagttccctcagctgtgcaagttctgcgacgtgaggttcagcacctgcgacaaccagaagtcctgcatgagcaactgcagcatcacaagcatctgcgagaagccccaggaggtgtgtgtggccgtgtggaggaagaacgacgaaaacatcaccctcgagaccgtgtgccatgaccccaagctgccctaccacgacttcatcctggaagacgccgcctcccccaagtgcatcatgaaggagaagaagaagcccggcgagaccttcttcatgtgcagctgcagcagcgacgagtgcaatgacaacatcatctttagcgaggagtacaacaccagcaaccccgactgataa
seqidno:6
分泌的抗pd-l1(突变)/tgfβ阱λ轻链的多肽序列,含a31g、d52e、r99y突变qsaltqpasvsgspgqsitisctgtssdvggynyvswyqqhpgkapklmiyevsnrpsgvsnrfsgsksgntasltisglqaedeadyycssytssstyvfgtgtkvtvlgqpkanptvtlfppsseelqankatlvclisdfypgavtvawkadgspvkagvettkpskqsnnkyaassylsltpeqwkshrsyscqvthegstvektvaptecs
seqidno:7
分泌的抗pd-l1(突变)/tgfβ阱重链的多肽序列
evqllesggglvqpggslrlscaasgftfsmymmmwvrqapgkglewvssiypsggitfyadsvkgrftisrdnskntlylqmnslraedtaiyycariklgtvttvdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkrvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgaggggsggggsggggsggggsgipphvqksvnndmivtdnngavkfpqlckfcdvrfstcdnqkscmsncsitsicekpqevcvavwrkndenitletvchdpklpyhdfiledaaspkcimkekkkpgetffmcscssdecndniifseeyntsnpd
seqidno:8
人tgfβrii同种型a前体多肽(ncbirefseq登记号:np_001020018)
mgrgllrglwplhivlwtriastipphvqksdvemeaqkdeiicpscnrtahplrhinndmivtdnngavkfpqlckfcdvrfstcdnqkscmsncsitsicekpqevcvavwrkndenitletvchdpklpyhdfiledaaspkcimkekkkpgetffmcscssdecndniifseeyntsnpdlllvifqvtgisllpplgvaisviiifycyrvnrqqklsstwetgktrklmefsehcaiileddrsdisstcanninhntellpieldtlvgkgrfaevykaklkqntseqfetvavkifpyeeyaswktekdifsdinlkhenilqfltaeerktelgkqywlitafhakgnlqeyltrhviswedlrklgsslargiahlhsdhtpcgrpkmpivhrdlkssnilvkndltcclcdfglslrldptlsvddlansgqvgtarymapevlesrmnlenvesfkqtdvysmalvlwemtsrcnavgevkdyeppfgskvrehpcvesmkdnvlrdrgrpeipsfwlnhqgiqmvcetltecwdhdpearltaqcvaerfselehldrlsgrscseekipedgslnttk
seqidno:9
人tgfβrii同种型b前体多肽(ncbirefseq登记号:np_003233)
mgrgllrglwplhivlwtriastipphvqksvnndmivtdnngavkfpqlckfcdvrfstcdnqkscmsncsitsicekpqevcvavwrkndenitletvchdpklpyhdfiledaaspkcimkekkkpgetffmcscssdecndniifseeyntsnpdlllvifqvtgisllpplgvaisviiifycyrvnrqqklsstwetgktrklmefsehcaiileddrsdisstcanninhntellpieldtlvgkgrfaevykaklkqntseqfetvavkifpyeeyaswktekdifsdinlkhenilqfltaeerktelgkqywlitafhakgnlqeyltrhviswedlrklgsslargiahlhsdhtpcgrpkmpivhrdlkssnilvkndltcclcdfglslrldptlsvddlansgqvgtarymapevlesrmnlenvesfkqtdvysmalvlwemtsrcnavgevkdyeppfgskvrehpcvesmkdnvlrdrgrpeipsfwlnhqgiqmvcetltecwdhdpearltaqcvaerfselehldrlsgrscseekipedgslnttk
seqidno:10
人tgfβrii同种型b胞外域多肽
ipphvqksvnndmivtdnngavkfpqlckfcdvrfstcdnqkscmsncsitsicekpqevcvavwrkndenitletvchdpklpyhdfiledaaspkcimkekkkpgetffmcscssdecndniifseeyntsnpd
seqidno:11
(gly4ser)4gly接头
ggggsggggsggggsggggsg
seqidno:12
分泌的抗pd-l1抗体mpdl3280a重链可变区的多肽序列
evqlvesggglvqpggslrlscaasgftfsdswihwvrqapgkglewvawispyggstyyadsvkgrftisadtskntaylqmnslraedtavyycarrhwpggfdywgqgtlvtvss
seqidno:13
分泌的抗pd-l1抗体mpdl3280a轻链可变区和抗pd-l1抗体yw243.55s70的轻链可变区多肽序列
diqmtqspsslsasvgdrvtitcrasqdvstavawyqqkpgkapklliysasflysgvpsrfsgsgsgtdftltisslqpedfatyycqqylyhpatfgqgtkveikr
seqidno:14
分泌的抗pd-l1抗体yw243.55s70重链可变区的多肽序列
evqlvesggglvqpggslrlscaasgftfsdswihwvrqapgkglewvawispyggstyyadsvkgrftisadtskntaylqmnslraedtavyycarrhwpggfdywgqgtlvtvsa
援引纳入
本文提及的每篇专利文献和科学论文其全部公开内容通过援引纳入本文用于所有目的。美国专利申请14/618,454的全部公开内容通过援引整体纳入本文用于所有目的。
等同形式
在不脱离本发明的精神或基本特征的情况下,本发明可以其他特定形式实施。因此,以上所述实施例从各方面讲都是说明性的而不是对本文所述发明的限制。不同实施例中的各种结构元件和所述各种方法步骤可以任意组合排列而所有这些变型都应视为本发明的方式。因此,本发明的范围由所附权利要求而非以上说明来指示,并且涵盖权利要求含义和等同范围内的所有改换形式。
- 还没有人留言评论。精彩留言会获得点赞!