一种提高高山被孢霉产不饱和脂肪酸能力的方法与流程
本发明涉及一种提高高山被孢霉产不饱和脂肪酸能力的方法,属于微生物以及基因工程技术领域。
背景技术
多不饱和脂肪酸是指含有两个或两个以上不饱和键的脂肪酸(pufas),依据从甲基端开始的第一个双键的位置差异,可以将pufas分为ω-3、ω-6和ω-9等类别。pufas具有十分重要的生理功能,其中二十二碳六烯酸(dha)和二十碳五烯酸(epa)具有降低血液中胆固醇和甘油三酯的功能,能将人体的血脂水平维持在正常的水平,预防心血管疾病和改善内分泌失调。
植物油和深海鱼油是pufas目前的主要来源,但面临着需求量增加,环境污染等问题,pufas资源日益短缺,传统的生产方法已经不能满足人们的需要,利用微生物生产油脂的新兴方法的优势越来越明显。相比于传统工艺,微生物生产油脂具有周期短,大批量生产,可以通过定向筛选和改造的方法增加油脂产量等优点。
由于高山被孢霉具有安全性好、脂质产量高、不饱和脂肪酸种类丰富及含量较高等特点,现已成为工业化生产aa的主要菌株,也被认为是生产epa的极具潜力的候选菌株之一。
乙酰辅酶a和nadph在脂肪合成的关键因素。乙酰辅酶a是脂肪酸合成时的重要底物和前体,而nadph是脂肪酸合成时所需还原力的唯一来源,因此nadph是脂质生成中的限制因素之一。高山被孢霉合成脂肪酸所需nadph的来源可由戊糖磷酸途径(ppp)中的葡萄糖-6-磷酸脱氢酶(g6pd)和6-磷酸葡萄糖酸脱氢酶(6pgd)、苹果酸酶(me)、异柠檬酸脱氢酶(idh)、乙醛脱氢酶(aldh)及氨基酸代谢,如丝氨酸合成(serinesynthesis)以及甘氨酸裂解系统(glycinecleavagesystem)等提供。因此,通过解析nadph的来源,可进一步明确脂质合成机理。
近年来,有研究表明在老鼠肾脏细胞中ppp和me提供约三分之一的nadph,而叶酸代谢途径提供近一半的nadph,而且大部分通过叶酸代谢途径产生的还原力nadph用于合成脂质,但过去人们并不将叶酸代谢途径视为nadph的重要来源之一。亚甲基四氢叶酸脱氢酶(mthfdl)是叶酸代谢途径中的关键酶。
技术实现要素:
本发明的目的是提供一种产不饱和脂肪酸能力提升的重组高山被孢霉工程菌,即一株过表达亚甲基四氢叶酸脱氢酶mthfdl基因的重组高山被孢霉工程菌株,以及在高山被孢霉(m.alpina)中过表达mthfdl基因的方法。
所述过表达mthfdl基因的高山被孢霉重组菌株,是以含mthfdl基因的根癌农杆菌转化高山被孢霉尿嘧啶营养缺陷型菌株构建而成的。
具体地,所述过表达mthfdl基因的高山被孢霉重组菌株是用含有mthfdl基因的重组质粒pbig2-ura5s-mthfdl转化根癌农杆菌后,再用含有转化质粒pbig2-ura5s-mthfdl的根癌农杆菌转化高山被孢霉尿嘧啶营养缺陷型菌株构建而成的。
过表达mthfdl基因时,通过基因工程技术将mthfdl基因插入高山被孢霉通用载体pbig2-ura5s-its中,构建二元表达载体pbig2-ura5s-mthfdl。将该二元表达载体pbig2-ura5s-mthfdl通过根癌农杆菌转化高山被孢霉,得到过表达mthfdl基因的高山被孢霉重组菌株。
所述载体pbig2-ura5s-its记载于公开号为cn103571762a,公开日为2014年2月12日的专利中。
所述亚甲基四氢叶酸脱氢酶(mthfdl)基因来源于高山被孢霉(mortierellaalpina)atcc#32222。
所述表达宿主为高山被孢霉尿嘧啶营养缺陷型菌株是已公开的重组高山被孢霉ccfm501,保藏编号为cgmccno.8414,该菌株已在公开号为cn103468581a,公开日为的2013年12月25日的中国专利申请中公开。
本发明还提供了一种构建过表达亚甲基四氢叶酸脱氢酶mthfdl基因的重组高山被孢霉菌株的方法,该方法包括如下步骤:
(1)以高山被孢霉cdna为模板,用引物p1/p2进行pcr扩增,获得编码mthfdl脱氢酶的基因mthfdl;
(2)借助高山被孢霉基因操作通用载体pbig2-ura5s-its构建重组质粒pbig2-ura5s-mthfdl;
(3)用构建获得的重组质粒pbig2-ura5s-mthfdl转化根癌土壤杆菌;
(4)用含重组质粒pbig2-ura5s-mthfdl的根癌土壤杆菌转化高山被孢霉尿嘧啶营养缺陷型菌株;
(5)筛选鉴定转化菌株,获得过表达mthfdl基因的重组高山被孢霉菌株ma-mthfdl。
在本发明中,应用于转化高山被孢霉的根癌农杆菌可以选用根癌农杆菌agrobacteriumtumefaciensccfm834,为本领域技术人员可以公开获得的菌株。
本发明还提供应用所述过表达亚甲基四氢叶酸脱氢酶mthfdl基因的重组高山被孢霉工程菌株生产脂质的方法。例如,将重组菌接种于broth培养基中,于适宜温度下培养,使重组菌合成脂质。例如,在28℃,200r/min培养7天。
本发明在现有的高山被孢霉转化系统的基础上,利用根癌土壤杆菌介导的基因转化方法,构建了在高山被孢霉中过表达mthfdl基因的工程菌株。所得基因工程菌株经过多次传代,经验证mthfdl基因片段仍稳定存在基因组中,且重组菌的生长特性与野生型菌株无明显差别,重组菌株中mthfdl基因的相对表达水提高了约6.8倍,重组菌株的脂肪酸含量能够明显得到提高,其总脂含量比原养型高山被孢霉提高了35.6%,同时nadph的含量提高了26.2%。这表明mthfdl在脂质合成过程中具有重要作用,很可能是脂质合成所需nadph的重要调控因素。本发明为深入解析高山被孢霉脂质合成机理,使之成为生产功能性脂质的细胞工厂提供理论依据。
附图说明
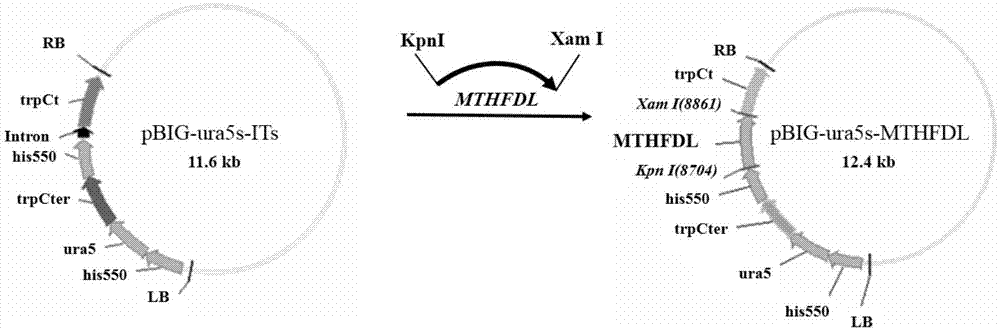
图1为二元表达载体pbig2-ura5s-mthfdl的构建示意图;
图2为二元表达载体pbig2-ura5s-mthfdlsilent的构建示意图;
图3为过表达mthfdl基因的高山被孢霉重组菌株鉴定的琼脂糖凝胶电泳图;
其中,m为marker,1为pbig2-ura5s-mthfdl重组菌株,n为野生型高山被孢霉菌株。
图4为干扰mthfdl基因的高山被孢霉重组菌株鉴定的琼脂糖凝胶电泳图;
其中,m为marker,1为pbig2-ura5s-mthfdlsilent重组菌株,2为原养型高山被孢霉菌株。
图5为重组菌株与原养型高山被孢霉菌株中mthfdl基因转录水平;
图6为重组菌株与原养型高山被孢霉菌株中nadph相对含量;
图7为干扰重组菌株ma-mthfdlsilent与原养型高山被孢霉菌株脂肪酸相对组成;
图8为过表达重组菌株ma-mthfdl与原养型高山被孢霉菌株脂肪酸相对组成。
具体实施方式
以下实施例用于非限制性地解释本发明的技术方案。
在本发明中,如无特殊说明,用于解释浓度的“%”均为重量百分比、“份”均为重量份。
在本发明中,涉及以下培养基:
broth培养基:20g/l葡萄糖,5g/l酵母提取物,1g/lkh2po4,0.25g/lmgso4·7h2o,10g/lkno3,ph6.0。
mm培养基:1.74g/lk2hpo4,1.37g/lkh2po4,0.146g/lnacl,0.49g/lmgso4·7h2o,0.078g/lcacl2,0.0025g/lfeso4·7h2o,0.53g/l(nh4)2so4,7.8g/lmes,1.8g/l葡萄糖,5g/l甘油,ph6.8。
im培养基是在mm培养基的基础上添加了200μm的乙酰丁香酮(as)构成的。
yep培养基:10g/l酵母提取物,10g/l胰蛋白胨,5g/l氯化钠,余量为水。
当采用固体培养基时,在原有培养基组成的基础上添加20g/l琼脂。
sc固体培养基:20g/l葡萄糖,5g/l酵母氮源无氨基酸和硫酸铵,1.7g/l硫酸铵,60mg/l异亮氨酸,60mg/l亮氨酸,60mg/l苯丙氨酸,50mg/l苏氨酸,40mg/l赖氨酸,30mg/l酪氨酸,20mg/l腺嘌呤,20mg/l精氨酸,20mg/l组氨酸,10mg/l甲硫氨酸,20g/l琼脂,余量为水,ph6.8。
sc-cs培养基是在sc固体培养基的基础上添加浓度为100μg/ml壮观霉素奇霉素(spectinomycin)和浓度为100μg/ml头孢噻肟抗生素(cefotaximesodium)。
gy固体培养基:20g/l葡萄糖,10g/l酵母提取物,2g/l硝酸钾,1g/l磷酸二氢钠,3g/l七水硫酸镁,20g/l琼脂,余量为水,ph6.8。
gy-u培养基是在gy固体培养基的基础上添加0.1g/l的尿嘧啶。
gy-cs培养基是在gy固体培养基的基础上添加浓度为100μg/ml壮观霉素奇霉素(spectinomycin)和浓度为100μg/ml头孢噻肟抗生素(cefotaximesodium)。
soc复苏培养基:20g/l胰蛋白胨,5g/l酵母粉,0.5g/l氯化钠,2.5mm氯化钾,10mm氯化镁,20mm葡萄糖,余量为水。
lb固体培养基:10g/l胰蛋白胨,5g/l酵母粉,10g/l氯化钠,20g/l琼脂,余量为水。
本发明涉及的检测方法如下:
琼脂凝胶电泳分析:取1μgrna在1.2%的变性胶(含1.5%甲醛)中电泳,观察rna完整性。
rt-qpcr检测:取0.5-1μg总rna为模板,根据primescriptrtreagentkit(takara,otsu,shiga,japan)试剂盒说明进行操作,获得重组菌株的cdna;使用abi-prism7900sequencedetectionsystem(appliedbiosystems,ca)按照sybrgreenpcrmastermix(appliedbiosystems,ca)的说明进行rt-qpcr反应,反应体系为20μl,其中含power
脂肪酸甲酯检测:采用gc-2010(shimadzuco.,japan),色谱柱为db-waxetr(30m×0.32mm,0.22μm),氢火焰离子检测器检测,气化室和检测器温度分别为240℃和260℃,分流方式进样1μl,分流比10:1,载气为氮气。程序升温:初始温度120℃保持3min,以5℃/min升到190℃,再以4℃/min升到220℃,保持20min。通过与商业化的脂肪酸甲酯标准品(37种脂肪酸甲酯混标,supelco,usa)和加入内标c15:0的质量比较,定性、定量分析样品中脂肪酸组分。
实施例1:高山被孢霉mthfdl编码基因的克隆
根据高山被孢霉(m.alpina)atcc#32222的mthfdl基因的序列信息,设计引物对pf/pr、p1/p2和p3/p4,下划线部分分别为酶切位点kpni、xami、kpni、nhei、smai和xmai,以mthfdl编码基因为模板,分别用引物pf/pr、p1/p2和p3/p4、kod高保真聚合酶,进行pcr扩增,得到mthfdl(其核酸序列及氨基酸序列如seqidno.1和seqidno.2所示)、mthfdlsf和mthfdlsr。
pcr程序为:94℃3min,94℃30s,55℃30s,68℃1.5min,30个循环,68℃5min;再对pcr产物进行纯化,纯化产物用1.2%的琼脂糖凝胶电泳验证。
pf:cggggtaccgcatgtcctgcaaggttgttctc
pr:tccccccgggttaagcgcttgcggc
p1:cggggtacccaaggcatctcggcctc
p2:ctagctagctgattccgtgcactccagcgtcctg
p3:tcccccgggcaaggcatctcggcctc
p4:tccccccgggtgattccgtgcactccagcgtcctg
实施例2:二元表达载体的构建
用限制性内切酶kpni、xami过夜酶切实施例1克隆的基因片段mthfdl及载体pbig2-ura5s-its,酶切体系(100μl)为:2μlkpni,2μlxami,30μl质粒或pcr产物,10μlcutsmartbuffer,56μl去离子水,37℃金属浴酶切12h后纯化;用限制性内切酶、kpni、nhei过夜酶切基因片段mthfdlsf及载体pbig2-ura5s-its,酶切体系(100μl)为:2μlkpni,2μlnhei,30μl质粒或pcr产物,10μlcutsmartbuffer,56μl去离子水,37℃金属浴酶切12h后纯化。
用t4连接酶将酶切纯化后的基因片段mthfdl、mthfdlsf分别与载体pbig2-ura5s-its连接,4℃下12h,得到重组表达载体pbig2-ura5s-mthfdl和pbig2-ura5s-itmthfdlsf,连接体系为(100μl):10μl目的基因酶切后片段,30μl载体酶切后片段,10μl连接酶buffer,5μlt4连接酶,45μl无菌水,4℃连接16h。
用限制性内切酶smai和xmai过夜酶切基因片段mthfdlsr及载体pbig2-ura5s-itmthfdlsf,酶切体系(100μl)为:2μlsmai,2μlxmai,30μl质粒或pcr产物,10μlcutsmartbuffer,56μl去离子水,37℃金属浴酶切12h后纯化。
用t4连接酶将酶切纯化后的基因片段mthfdlsr与载体pbig2-ura5s-itmthfdlsf连接,4℃连接16h,得到重组表达载体为pbig2-ura5s-mthfdlsilent。
连接体系为(10μl):2μl目的基因酶切后片段,6μl载体酶切后片段,1μl连接酶buffer,1μlt4连接酶,4℃连接16h(二元表达载体pbig2-ura5s-mthfdl的构建如图1,二元表达载体pbig2-ura5s-mthfdlsilent的构建见图2)
实施例3:二元表达载体的验证
将实施例2所得的重组表达载体pbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent电转化大肠杆菌top10感受态细胞,方法如下:
1)无菌状态下取80μl感受态细胞,加入1-2μl上述连接产物pbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent,吹吸混匀;
2)将上述混匀的感受态细胞移入预冷过的电转杯中,避免产生气泡;
3)将电转杯放入bio-rad电转仪,调到合适预设程序档位,根据仪器说明电转,电压条件为1.8kv;
4)电转后的感受态细胞移入含有1mlsoc复苏培养基的1.5ml无菌离心管中,37℃,150rpm孵育1小时;
5)取200μl涂布含有100μg/ml卡那霉素的lb固体培养基平板,倒置37℃培养过夜;
6)挑取阳性转化子,提取质粒,测序验证结果表明连接成功,获得二元表达载体pbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent。
实施例4:制备含二元表达载体的根癌农杆菌
将实施例3所得的二元表达载体pbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent电击转化根癌农杆菌的方法参照转化大肠杆菌top10的方法,电击电压为2.5kv,分别得到含有质粒pbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent的根癌农杆菌ccfm834。
实施例5:含二元表达载体的根癌农杆菌的培养
步骤如下:
1)将实施例4所得的含有质粒ppbig2-ura5s-mthfdl和pbig2-ura5s-mthfdlsilent的根癌农杆菌ccfm834在含有100μg/ml利福平和100μg/ml卡那霉素的yep固体培养基平板上划线,28℃倒置避光培养48小时;
2)挑取单克隆接种至20ml含有100μg/ml利福平和100μg/ml卡那霉素的液体yep培养基中28℃,200rpm避光培养48小时;
3)取200μl液体到20mlim培养基中,30℃200rpm避光培养24-48h;
4)用im培养基调整菌浓度至od600为0.3,置于28℃,200rpm摇床避光培养至od600到0.8;
5)用灭菌的broth-u冲刷在gy-u斜面培养1个月以上的高山被孢霉尿嘧啶营养缺陷性菌株ccfm501,收集孢子;
6)取100μl根癌农杆菌与100μl孢子混合,均匀涂布于铺有玻璃纸的im固体培养基上。16℃避光正置培养24h后再放于23℃避光正置培养24-36h;
7)将玻璃纸转移到含有100μg/ml壮观霉素和100μg/ml头孢噻肟抗生素的sc平板(sc-cs)上,16℃避光正置培养2d后再置于23℃避光正置培养;
8)持续观察菌落在sc-cs平板上的生长情况,若长出明显菌落,及时用尖头镊子将菌落外沿挖出(大约2mm2),接种于sc-cs平板上,继续正置于23℃培养箱中培养;
9)待sc-cs平板上的转化子长出后,挑菌丝转接于sc-cs平板,重复筛选3次,排除阴性转化子;
10)将筛选3次后生长的菌落接种至gy平板,28℃培养至产生大量孢子,保藏在4℃。
实施例6:重组高山被孢霉工程菌的构建
取实施例5中所得的孢子液构建重组高山被孢霉工程菌,步骤如下:
1)以4mlbroth-u培养基来冲刷gy平皿表面,收集实施例5所得的孢子液于一个无菌1.5ml离心管中,过25μm滤膜;
2)调整根癌农杆菌的浓度,od600为0.3、0.7、1.0左右,各取100μl高山被孢霉菌液与根癌农杆菌混匀涂布于im固体培养基上,23℃避光培养2-3天;
3)随时用无菌镊子挑出生长的真菌菌丝sc-cs平板上,25℃培养2-3天;
4)观察高山被孢霉在平板上的生长情况,挑出在sc-cs平板上生长的菌丝接种于gy斜面上;
5)将上述步骤4)中平板上的高山被孢霉菌株孢子在gy斜面上传代三次;
6)将稳定遗传的菌株鉴定为干扰和过表达mthfdl基因的重组高山被孢霉菌株,保藏于gy斜面上;
7)提取鉴定正确的重组高山被孢霉菌株的基因组dna,用一对与启动子和终止子特异性结合的引物进行pcr验证:
p5(sense):cacacacaaacctctctcccact
p6(antisense):caaatgaacgtatcttatcgagatcc;
通过琼脂凝胶电泳分析来鉴定重组菌株,结果如图3,ma-mthfdl菌株产物条带有ura5(818bp)表达单元和mthfdl(mthfdl936bp加载体上164bp共1100bp)单元,野生型高山被孢霉无条带,电泳结果表明过表达载体成功整合到高山被孢霉基因组中;ma-mthfdlsilent菌株产物条带为ura5(818bp)表达单元,原养型高山被孢霉产物条带有ura5(818bp)表达单元和its(200bp)单元,电泳结果表明干扰载体成功整合到高山被孢霉基因组中。
实施例7:重组高山被孢霉工程菌中mthfdl基因转录水平的rt-qpcr检测
根据mthfdl序列和内参18srdna序列设计如下引物:
p7(sense):cggggtacccaaggcatctcggcctc
p8(antisense):ctagctagctgattccgtgcactccagcgtcctg
p9(sense):cgtactaccgattgaatggcttag
p10(antisense):cctacggaaaccttgttacgact
取实施例6所得的重组高山被孢霉工程菌的总rna进行rna完整性检测,步骤如下:
1)将实施例6中得到的重组高山被孢霉工程菌进行培养离心,取约0.1g上清液中的菌体于预冷的灭酶研钵中,加液氮充分研磨;
2)加入1mltrizol(invitrogen,carlsbad,ca,usa)继续研磨成粉末状后,室温溶解;
3)将溶解后液体用无酶枪头吸出约1ml于无酶离心管中,添加200μl三氯甲烷颠倒混匀,室温放置2min;
4)12000rpm,4℃离心15min,吸取上层水相于新的无酶离心管中;
5)添加等体积异丙醇,颠倒混匀,室温放置10min,12000rpm,4℃,离心15min;
6)用无酶枪头吸去异丙醇;
7)添加1ml新配75%乙醇,12000rpm,4℃,离心15min后,去上清;
8)室温放置干燥,用100μl无酶水溶解rna沉淀,-80℃保存;
9)浓度测定:取1μlrna用nanodrop2000测定浓度;
10)变性凝胶电泳检测rna完整性。
反转录合成重组高山被孢霉工程菌的cdna进行rt-qpcr检测。
rt-qpcr结果如图5所示,m.alpina为原养型高山被孢霉菌株对照,ma-mthfdl为过表达重组菌株,与对照组相比,在培养7d后,过表达菌株中的mthfdl基因的转录水平约是对照菌株的6.8倍,说明转化的mthfdl基因表达原件成功实现了转录及过量表达;ma-mthfdlsilent为干扰重组菌株,rna干扰菌株中的mthfdl基因的转录水平下降了0.539倍,说明转化的mthfdl基因表达原件成功抑制了转录。
实施例8:重组高山被孢霉工程菌中nadph的提取与检测
取实施例6所得的重组高山被孢霉工程菌中nadph进行检测,步骤如下:
1)将实施例6中得到的重组高山被孢霉工程菌进行培养离心,取约0.1g上清液中的菌体于2mlep管中,加入1mlnadp/nadphextrationbuffer,组织破碎仪破碎后,置于冰上10min,离心(1000xg,10min);
2)转移上清至10kda超滤管中,过滤除去酶;
3)剩余步骤参考说明书进行;
计算出nadph质量后,结合高山被孢霉酶菌体干湿重比算出nadph重量/菌体重量(mg/g),结果如图6所示,过表达重组菌株ma-mthfdl中的nadph水平相对原养型高山被孢霉提高了26.21%,干扰重组菌株ma-mthfdlsilent中的nadph水平下降了19.0%,进一步证明了干扰mthfdl对细胞内的nadph水平产生了影响。
实施例9:重组高山被孢霉工程菌产脂肪能力的检测
1)采用盐酸反复冻融破壁的方式,用有机溶剂提取高山被孢霉干菌体中油脂;
2)将高山被孢霉野生型菌株与实施例6获得的重组高山被孢霉工程菌接种于broth培养基中,28℃,200r/min摇床培养7天;
3)收集菌体后真空冷冻干燥,并称重计算生物量;
4)用研钵将菌体研磨成粉末,精确称取50mg菌粉于提脂瓶,加入2ml4mol/l盐酸;
4)80℃水浴1h,-80℃冷却15min,重复一次且震荡混匀,于80℃水浴1h;
5)冷却至室温,加入1ml甲醇,震荡混匀;
6)加入1ml氯仿,震荡10min,6000g离心3min,小心吸取下部氯仿于新提脂瓶中;
7)重复步骤(6)两次;
8)合并氯仿(约3ml),加入1ml饱和氯化钠,震荡混匀,3000g离心3分钟。收集氯仿层于新提脂瓶中。再继续加入1ml氯仿于原瓶,3000g离心3分钟。合并氯仿(约4ml);
9)氮吹干燥得到粗脂;
10)向上述粗脂中分别加入100μl2.02mg/ml内标c15:0和1ml10%盐酸甲醇溶液,60℃水浴3h,每隔半小时振荡1min;
11)冷却至室温后加入2ml正己烷和1ml饱和nacl溶液,震荡混匀,4000rpm离心3min;
12)吸出200μl正己烷层于新瓶,加入800μl正己烷混匀后转入气相瓶,得到脂肪酸甲酯溶液;
13)进行脂肪酸甲酯检测。
其中总脂肪酸含量用单位菌体中总脂肪酸的质量表示。(表1、2是原养型高山被孢霉与重组菌株脂肪酸的组成)
表1高山被孢霉菌株与重组菌株脂肪酸组成比较
表2高山被孢霉菌株与重组菌株脂肪酸组成比较
表1、2和图7表明,本发明得到的重组菌株ma-mthfdlsilent完成了干扰,ma-mthfdlsilent的总脂肪酸含量由35.0%降到了21.2%,证明了高山被孢霉脂质合成和mthfdl的相关性;表1和图8表明,本发明得到的重组菌株ma-mthfdl完成了过表达,ma-mthfdl的总脂肪酸产量由35.0%升高到了47.5%,花生四烯酸含量提高了80.8%,表明过表达mthfdl基因还增强了高山被孢霉脂肪酸的去饱和作用,这对高山被孢霉分子改造来解析脂质合成机理,为后续工业化生产功能性脂质提供了理论依据。
虽然本发明已以较佳实施例公开如上,但其并非用以限定本发明,任何熟悉此技术的人,在不脱离本发明的精神和范围内,都可做各种的改动与修饰,因此本发明的保护范围应该以权利要求书所界定的为准。
sequencelisting
<110>江南大学
<120>一种提高高山被孢霉产不饱和脂肪酸能力的方法
<160>14
<170>patentinversion3.3
<210>1
<211>936
<212>dna
<213>高山被孢霉
<400>1
atgtcctgcaaggttgttctcgctcaaggcatctcggcctcttacagagaggagctcaag60
aacacgatcaagagccgcaacatccgcccgaagctggtcggtttcctcgccaacgaggat120
cctgctgctgcaaagtatgccgagtggactgcaaagacctgcgccgagacaggtgtcgag180
tttgagctcagaacctgcgccagaaacgatctcgaggacaagatcatcgaggccaaccag240
gacgctggagtgcacggaatcatggtctactatcccgtttttggcgatcgccaggatcaa300
tacctgcaaaacgtcgtcgatgtcagcaaggatgtggaaggtctgcaccacaagtatgtc360
tacaacatgtaccacaacatccgcttcctcgacgaggccaagaccaagaagtgcattatt420
ccctgcacccccctgggattggtgaaggtgatggagtatgtcggagtttacaacggcatc480
cttccccacggcaaccgcctgcacggaagggtcgtcaccatcatcaaccgctccgagatt540
gtcggtcgtcccttggccgctctcctcgctaacgacggtgccagggtttactcggtcgac600
atcaatggcatccaggaatttcaccgtggcgttggtcttcagctcaagaagcacgaggtt660
accgagacgacgctgaaggttgaggacgttttgcccctctccgacgttgtcattaccggt720
gtgccctcgccctcgtacaaggtcccctcgaacctgatccgcgatggtgccgtctgcatc780
aacttctcgaccttcaagaactttgatggcgaagagatcaagaagaaggctgccatctac840
gtccccgccgtcggcaaggtcactgtcgccatgctcgagcgcaacctggtccgtctctac900
gactaccagaaccaagaccgcgccgcaagcgcttaa936
<210>2
<211>311
<212>prt
<213>高山被孢霉
<400>2
metsercyslysvalvalleualaglnglyileseralasertyrarg
151015
glugluleulysasnthrilelysserargasnileargprolysleu
202530
valglypheleualaasngluaspproalaalaalalystyralaglu
354045
trpthralalysthrcysalagluthrglyvalgluphegluleuarg
505560
thrcysalaargasnaspleugluasplysileileglualaasngln
65707580
aspalaglyvalhisglyilemetvaltyrtyrprovalpheglyasp
859095
argglnaspglntyrleuglnasnvalvalaspvalserlysaspval
100105110
gluglyleuhishislystyrvaltyrasnmettyrhisasnilearg
115120125
pheleuaspglualalysthrlyslyscysileileprocysthrpro
130135140
leuglyleuvallysvalmetglutyrvalglyvaltyrasnglyile
145150155160
leuprohisglyasnargleuhisglyargvalvalthrileileasn
165170175
argsergluilevalglyargproleualaalaleuleualaasnasp
180185190
glyalaargvaltyrservalaspileasnglyileglngluphehis
195200205
argglyvalglyleuglnleulyslyshisgluvalthrgluthrthr
210215220
leulysvalgluaspvalleuproleuseraspvalvalilethrgly
225230235240
valproserprosertyrlysvalproserasnleuileargaspgly
245250255
alavalcysileasnpheserthrphelysasnpheaspglygluglu
260265270
ilelyslyslysalaalailetyrvalproalavalglylysvalthr
275280285
valalametleugluargasnleuvalargleutyrasptyrglnasn
290295300
glnaspargalaalaserala
305310
<210>3
<211>32
<212>dna
<213>人工序列
<400>3
cggggtaccgcatgtcctgcaaggttgttctc32
<210>4
<211>25
<212>dna
<213>人工序列
<400>4
tccccccgggttaagcgcttgcggc25
<210>5
<211>26
<212>dna
<213>人工序列
<400>5
cggggtacccaaggcatctcggcctc26
<210>6
<211>34
<212>dna
<213>人工序列
<400>6
ctagctagctgattccgtgcactccagcgtcctg34
<210>7
<211>26
<212>dna
<213>人工序列
<400>7
tcccccgggcaaggcatctcggcctc26
<210>8
<211>35
<212>dna
<213>人工序列
<400>8
tccccccgggtgattccgtgcactccagcgtcctg35
<210>9
<211>23
<212>dna
<213>人工序列
<400>9
cacacacaaacctctctcccact23
<210>10
<211>26
<212>dna
<213>人工序列
<400>10
caaatgaacgtatcttatcgagatcc26
<210>11
<211>26
<212>dna
<213>人工序列
<400>11
cggggtacccaaggcatctcggcctc26
<210>12
<211>34
<212>dna
<213>人工序列
<400>12
ctagctagctgattccgtgcactccagcgtcctg34
<210>13
<211>24
<212>dna
<213>人工序列
<400>13
cgtactaccgattgaatggcttag24
<210>14
<211>23
<212>dna
<213>人工序列
<400>14
cctacggaaaccttgttacgact23
- 还没有人留言评论。精彩留言会获得点赞!