基于基因编辑技术的ALS突变型蛋白及其基因在植物育种中的应用的制作方法
本发明属于作物遗传育种、作物抗除草剂新资源创新领域,具体涉及基于基因编辑技术的als突变型蛋白及其基因在植物育种中的应用。
背景技术
随着我国新型城镇化和现代农业的发展,水稻生产的轻简栽培越来越受到青睐,机插秧、直播等方式已成为发展趋势。然而,直播稻田容易滋生杂草及杂草稻,严重影响水稻生长、产量及稻谷品质。人工除草和机械除草的成本极高,制约了水稻生产朝着高产、高效、低成本方向发展,不利于发展现代农业。喷施除草剂是防治杂草和杂草稻危害的有效手段。
植物和微生物支链氨基酸(缬氨酸、亮氨酸和异亮氨酸)的生物合成需要4种酶共同催化作用,乙酰乳酸合成酶(acetolactatesynthase,als)、酮醇还原异构酶(ketol-acidreductoisomerase)、二羟基酸脱水酶(dihydroxyaviddehydratase)、支链氨基酸转氨酶(branched-chainaminoacidtransaminase)。乙酰乳酸合成酶是生物合成过程中第一阶段的关键酶,在缬氨酸和亮氨酸的合成中催化2分子丙酮酸生成乙酰乳酸和二氧化碳,在异亮氨酸的合成中催化1分子丙酮酸与1分子α丁酮酸生成2-乙醛基-2-羟基丁酸和二氧化碳。als抑制剂类除草剂通过抑制植物体内的als酶活性,从而阻止支链氨基酸的合成,导致蛋白质的合成受到破坏,阻碍细胞分裂期的dna合成,从而使植物细胞的有丝分裂停止在gl阶段的s期(dna合成期)和g2阶段的m期,干扰了dna的合成,细胞因此不能完成有丝分裂,进而使植物停止生长,最终导致植物个体死亡。
乙酰乳酸合成酶(als)(也称乙酰羟酸合成酶,ahas;ec4.1.3.18)抑制剂类除草剂以als作为靶标而导致杂草死亡,主要包括磺酰脲类(sulfonylureas,su)、咪唑啉酮类(imidazolinones,imi)、三唑嘧啶类(triazolopyrimidines,tp)、嘧啶氧(硫)苯甲酸类[pyrimidinylthio(oroxy)–benzoates,ptb;pyrimidinyl-carboxyherbicides;pcs]和磺酰胺基羰基三唑啉酮类(sulfonylamino-carbonyltriazolinones,sct)等13类化合物。乙酰乳酸合成酶,存在于植物生长过程中,它能以高度专一性和极高的催化效率催化丙酮酸为乙酰乳酸,从而导致支链氨基酸的生物合成。
咪唑啉酮类除草剂是由美国氰胺公司开发的一类高效广谱低毒的除草剂,目前已经有6种商品化产品,包括:咪唑烟酸、咪唑乙烟酸、咪草酸、咪唑喹啉酸、甲氧咪草烟和甲基咪草烟。咪唑乙烟酸,又称为咪草烟,是一种常用于大豆田的高效除草剂,能有效防除一年生禾本科杂草和阔叶杂草。但这些除草剂对一般不具有抗(耐)除草剂特性的农作物本身也产生药害,极大限制了其使用时间和使用空间,如需要在农作物播种前一段时间使用除草剂才能避免农作物遭受药害。培育抗(耐)除草剂作物品种可减少作物药害、拓宽除草剂的使用范围。
目前,水稻已知的als基因抗除草剂突变位点包括gln25、gly95、ala96、gln113、ala122、ser160、pro171、ala179、ala237、asn350、his367、lys390、trp548、ser627和leu636。als突变体抗除草剂水平与als氨基酸突变的位置有关,还与突变后的氨基酸种类及突变氨基酸的数目有关。因此,创制和筛选新的抗除草剂突变类型,有利于丰富抗除草剂基因遗传多样性,为培育水稻新品种提供基因资源。
目前,筛选抗除草剂新基因资源主要是通过化学诱变。众所周知,化学突变频率低,前期投入大,筛选获得一个抗性稳定材料需要2-3年,同时化学诱变可能导致野生型材料多个基因突变,应用于生产还需要通过杂交改良,剔除不良基因突变。将目标基因导入到优良品种背景中,利用常规育种手段,主要是通过杂交、回交、复交、阶梯杂交等将目标基因导入到背景亲本中。对于质量性状,回交转育是常用的方法,一般需要回交4-6代,加上自交纯合等至少需要3-5代,要改良1个性状并能在生产上发挥作用,至少需要3-5年时间。
相对于传统育种,基因编辑育种效率高。由于粳稻品种农杆菌介导的遗传转化效率较高,我们发现,10个t0代转化子一般能筛选到3-5个纯合的目标等位基因同时被编辑的转化子,即在t0代就能观察到表型,如香味、低直链淀粉含量导致的暗胚乳、抽穗期提前等性状,利用t0代单株所结种子在t1代,一般在100株中就能筛选到剔除潮霉素和cas9基因的单株5株左右,繁殖5株苗,一般每株能收种子500粒种子,一株就能繁殖400株,可以繁殖常规粳稻种子20斤左右,足够进行品质分析,甚至参加试验、示范等各种试验。即使在t0代出现组织培养过程中无性系变异,利用基因组编辑的材料与野生型杂交,也可以剔除不良变异。
利用基因组编辑技术可以精准创制预期的目标基因,常规育种手段做不到。常规育种只能从品种资源中筛选具有目标基因的资源,然后通过杂交回交等手段改良现有品种,一般来说,品种资源材料农艺性状较差,因此遗传改良周期长,而基因组编辑技术可以直接利用农艺性状优良并在生产上大面积推广的水稻品种为背景材料编辑目标基因。目前,江苏省农业科学院通过构建抽穗期相关基因多基因编辑载体,利用江苏大面积推广的武运粳24、南粳9108为遗传转化背景材料,从中获得了抽穗期60多天、70多天、80多天抽穗的剔除外源标记基因的新材料,相对于野生型,抽穗期提早并呈梯度分布,可以扩大优质品种的种植范围,加快优质品种改良进程,极大节约育种成本,凸显了基因编辑分子育种的生命力、创造力。
借助分子标记辅助选择育种可以进行基因型选择,能够对目标性状基因杂合和纯合基因型进行筛选,可以加快目标基因纯合进程,因此,开发基因功能标记有利于加快育种进程。als基因功能突变多为单碱基突变,可以针对性地设计酶切靶点标记,但其过程相对繁琐,开发等位基因特异pcr,通过两次pcr就可以区分抗感基因型。
根据报道,trp548、ser627的变异可以使蛋白质对als抑制剂类除草剂抗性较好,但这些位点的不同变异类型仍然会有不同的效应。已有研究者利用基因替换技术成功实现了548位氨基酸由色氨酸到亮氨酸和627位氨基酸由丝氨酸到异亮氨酸的变异,但该方法基于已经报道的碱基变异,不能创制新的变异类型。
目前,科技人员想获得抗除草剂的水稻新材料或新基因,需要通过化学或辐射诱变等方式,工作量很大、效果很不理想,且获得的抗除草剂als蛋白大多数为已经报道的变异类型。
然而采用常规的化学诱变和常规转育育种,其育种年限至少要4~6年,目前并没有相关报道利用基因编辑技术对水稻品种als基因进行定点突变创制新的抗除草剂新的等位基因的相关研究。
技术实现要素:
发明目的:本发明所要解决的技术问题是提供了一种水稻als突变型蛋白及其核酸或基因。
本发明还要解决的技术问题是提供了表达盒、重组载体或细胞。
本发明还要解决的技术问题是提供了水稻als突变型蛋白、核酸或基因,所述的表达盒、重组载体或细胞在植物抗除草剂方面的应用。
本发明还要解决的技术问题是提供了一种利用基因编辑创制抗除草剂水稻的育种方法。本发明首次利用基因编辑技术对水稻品种als基因进行定点突变,创制新的抗除草剂新等位基因,并剔除t-dna外源序列,获得抗性稳定遗传的株系,一般只需要2年左右时间,相对于化学诱变和常规转育育种,育种年限至少提早2-4年。因此,基因编辑分子育种具有精准、效率高等常规育种不具备的优势,具有广泛的应用前景。
本发明还要解决的技术问题是提供了用于鉴定所述的基因或核酸的引物对。
本发明还要解决的技术问题是提供了所述的基因或核酸、所述的引物对在抗除草剂品系鉴定和选育中的应用。
技术方案:为了解决上述技术问题,本发明采用的技术方案如下:一种水稻als突变型蛋白,所述als突变型蛋白的氨基酸序列存在以下突变:其对应于水稻als的氨基酸序列的第628位氨基酸发生突变。
具体的,本发明首次报道了该628位氨基酸由甘氨酸突变为色氨酸并具有除草剂抗性。本发明的第628位氨基酸的突变还可以包括谷氨酸、天冬氨酸、色氨酸、丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、酪氨酸、丝氨酸、苏氨酸、半胱氨酸、蛋氨酸、天冬酰胺、谷氨酰胺、赖氨酸、精氨酸、组氨酸以及终止密码子等21种变异类型。而关于上述的氨基酸的其他变异或提前终止是否影响乙酰乳酸合成酶活性、生理功能以及是否具有除草剂抗性,还有待进一步研究证实。
本发明所述的水稻als突变型蛋白,包括:
(a)其氨基酸序列如seqidno:2所示;或
(b)在(a)中的氨基酸序列经过取代和/或缺失和/或添加一个或几个氨基酸且具有乙酰乳酸合成酶活性的由(a)衍生的蛋白质。
本发明内容还包括核酸或基因,其编码所述的突变型蛋白。
其中,所述的核酸或基因,包括:
(a)其编码所述的突变型蛋白;或
(b)在严格条件下与(a)限定的核苷酸序列杂交且编码具有乙酰乳酸合成酶活性的蛋白质的核苷酸序列;或
(c)其核苷酸序列如seqidno:1所示。
本发明内容还包括表达盒、重组载体或细胞,其含有所述的核酸或基因。
本发明内容还包括所述的水稻als突变型蛋白、核酸或基因,所述的表达盒、重组载体或细胞在植物抗除草剂方面的应用。
本发明内容还包括获得具有除草剂抗性植物的方法,包括如下步骤:
1)使植物包含所述的核酸或基因;或
2)使植物表达所述的水稻als突变型蛋白。
本发明内容还包括一种利用基因编辑创制抗除草剂水稻的育种方法,包括以下步骤:
1)als基因的克隆及基因编辑的靶位点设计;
2)含有目的片段的crispr/cas9基因编辑载体的构建;
3)具备所述的als突变型蛋白、或具备所述的核酸或基因的抗除草剂水稻的获得。
其中,所述步骤2)的含有目的片段的crispr/cas9基因编辑载体的构建方法如下:
a)靶点接头制备:采用接头引物用te溶解成母液,母液稀释后90℃30s,移至室温冷却完成退火,即获得靶点接头;
b)sgrna连接产物制备:采用pylsgrna-osu3中间载体、靶点接头、dna连接酶、bsai进行pcr扩增获得sgrna连接产物;
c)扩增sgrna表达盒:以引物组合u-f、grna-r对sgrna连接产物进行第一轮pcr扩增获得第一轮pcr产物,然后以uctcg-b1和grcggt-bl为扩增引物对稀释后的第一轮pcr产物进行第二轮pcr获得的pcr产物即为sgrna表达盒;
d)将sgrna表达盒连接到crispr/cas9表达载体上获得连接产物;
e)将步骤d)的连接产物进行热激发转化大肠杆菌获得重组菌,提取通过验证的含目的条带的菌液的阳性质粒即得。
其中,所述步骤3)的抗除草剂水稻获得方法如下:将步骤2)获得的含有目的片段的crispr/cas9基因编辑载体转入农杆菌eha105,获得t0代转基因植株,以引物alst-f和alst-r对t0代转基因植株进行扩增并测序鉴定获得具备所述的突变型蛋白、所述的核酸或基因的植株。
其中,所述育种方法还包括将步骤3)中所述具备除草剂抗性的t0代转基因植株的含有目标等位基因双突变的t1代植株的t-dna载体的剔除,所述t-dna载体包括hpt基因和cas9核酸酶基因。
其中,所述t-dna载体的剔除通过对含有目标等位基因双突变的t1代植株的hpt基因和cas9基因同时检测,重复多次,筛选得到不携带这两个基因的t1代单株即为目标植株。
其中,所述hpt基因检测方法通过以目标等位基因双突变的t1代植株的基因组dna为模板,以hyg283-f和hyg283-r为引物进行pcr扩增,同时,所述cas9基因检测方法通过以目标等位基因双突变的t1代植株的基因组dna为模板,以cas9t-f和cas9t-r为引物进行pcr扩增,当均未同时检测到hpt基因和cas9基因,表明这成功剔除了t-dna。
本发明内容还包括一种用于鉴定所述的基因或核酸的引物对,所述引物对为als4和/或als6,所述引物对als4序列如seqidno:6和seqidno:7所示,所述引物对als6序列如seqidno:8和seqidno:9所示。
本发明内容还包括所述的als突变型基因或核酸、所述的引物对在抗除草剂品系鉴定和选育中的应用。
有益效果:相对于现有技术,本发明具备以下优点:
1)本发明首次利用crispr/cas9基因编辑技术,对als基因进行编辑,通过后代的筛选,在t2代就可以获得剔除t-dna、抗除草剂特性稳定遗传的新材料,而新材料的基本农艺性状无明显改变。相对于化学诱变、杂交转育等育种,基因编辑定向改良分子育种技术具有快速、精准、高效等优点,利用基因功能标记进行基因型选择,将会极大提高育种效率,大大加快育种进程。
2)本发明根据野生型与突变体在als基因第1882位点的碱基变异,开发了能特异区分野生型(als基因1882位碱基为g)和突变体(als基因1882位碱基为t)的分子标记als4和als6,可以用于分子标记辅助选择育种中。
3)本发明的基因编辑技术育种得到的水稻品种的1-2叶幼苗在施用210g(a.i.)hm-2“咪草烟”(相当于3倍推荐使用浓度)后,植株仍然正常生长发育和结实,而野生型水稻1-2叶幼苗在施用210g(a.i.)hm-2“咪草烟”(相当于3倍推荐使用浓度,70g(a.i.)hm-2)14天后表现为整株死亡。
4)本发明的基因编辑技术育种得到的水稻品种的1-2叶幼苗在施用240g(a.i.)hm-2“百垄通”(相当于1倍推荐使用浓度,240g(a.i.)hm-2)后,植株仍然正常生长发育和结实,而野生型水稻1-2叶幼苗在施用240g(a.i.)hm-2“百垄通”(相当于1倍推荐使用浓度,240g(a.i.)hm-2)14天后表现为整株死亡。
附图说明
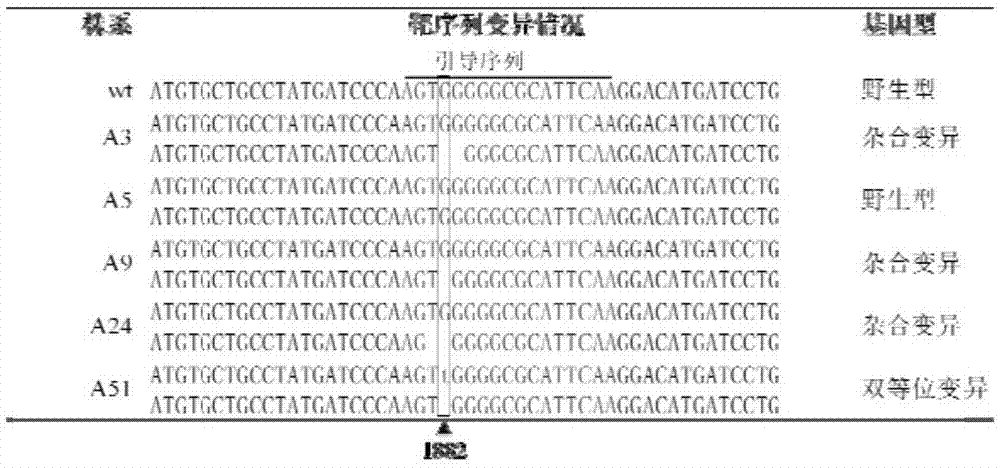
图1转基因植株的碱基变异;
图2除草剂筛选获得的抗性水稻突变体;wt为南粳9108,a3、a5、a9、a24和a51为t1代转基因株系;
图3t1代植株hpt基因和cas9基因检测结果;a:hpt基因;b:cas9基因。m为dl2000分子标记,1-18为t1代转基因植株,19为以质粒为模板的阳性对照,20为南粳9108模板的阴性对照;
图4突变体材料咪草烟处理结果;a为野生型,b为突变体,1~4分别为0、210、700、1400g(a.i.)hm-2浓度咪草烟进行喷施;
图5突变体材料百垄通处理结果;a为野生型,b为突变体,1~4分别为0、240、2400、4800g(a.i.)hm-2浓度百垄通进行喷施;
图6突变体与野生型农艺性状比较;a~f分别表示株高、有效穗、穗长、每穗粒数、结实率和千粒重;
图7als628w功能标记的开发;a:野生型;b:突变体。m为dl2000分子标记,1-13分别为分子标记als1~als13;
图8als628w功能标记检测品种;a:als4;b:als6。m为dl2000分子标记,1~27分别为南粳9108突变体、南粳9108野生型、日本晴、黄华占、9311、连粳7号、苏秀867、镇稻88、镇稻99、淮稻5号、常农粳8号、南粳44、南粳45、南粳46、南粳49、南粳51、南粳47、南粳5055、苏垦118、武运粳24号、武运粳27号、武运粳29号、徐稻8号、徐稻9号、扬育粳2号、华粳5号、盐稻16号;
图9als628w功能标记检测f2分离群体单株(部分);a:als4;b:als6。m为dl2000分子标记,1为徐稻9号,2为南粳9108突变体,3为徐稻9号/南粳9108突变体,1~21为徐稻9号/南粳9108突变体的f2单株。r为抗除草剂,s为感除草剂。
具体实施方式
下面将结合实施例对本发明的实施方案进行详细描述,但是本领域技术人员将会理解,下列实施例仅用于说明本发明,而不应视为限定本发明的范围。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市购获得的常规产品。
本发明选取的背景材料为南粳9108(购买于江苏高科种业有限公司),该品种是由江苏省农业科学院粮食作物研究所选育的迟熟中粳新品种,全生育期150天左右,适宜江苏省苏中及宁镇扬丘陵地区种植,具有优良的综合农艺性状,已在生产上大面积推广应用,深受市场欢迎。南粳9108株型较紧凑,分蘖力较强,抗倒性较强,熟相好,直链淀粉含量为10%左右,稻米外观呈云雾状,有香味,对咪唑啉酮类除草剂不具有抗性。本发明通过crispr/cas9基因编辑技术对南粳9108als基因进行定点编辑,获得抗咪唑啉酮类除草剂的突变体,以满足轻简栽培生产的迫切需要。
实施例1:水稻抗咪唑啉酮类除草剂突变体获取过程(咪草烟)
1、南粳9108als基因克隆及靶位点设计
参考murray等的ctab方法,提取南粳9108的基因组dna(murraymg,etal.,nucleicacidsresearch,1980,8(19):4321-4326)。用引物als5-f:tcgcccaaacccagaaaccc,als5-r:ctctttatgggtcattcaggtc对基因组dna进行pcr扩增,将扩增产物送往英潍捷基(上海)贸易有限公司进行测序。测序结果在ncbi(https://blast.ncbi.nlm.nih.gov/blast.cgi)数据库进行blast比对分析,发现南粳9108的als编码区序列与参考基因组水稻日本晴相同。
根据南粳9108的als基因序列,用crispr-ge网站(http://skl.scau.edu.cn/targetdesign/)预测,选取5’-tccttgaatgcgcccccact-3’作为基因编辑的靶位点。该靶位点造成的cas9切割位点位于1881与1882位碱基之间,临近碱基的变异有望造成627或628位氨基酸的变异,获得新的抗除草剂基因型。
2、crispr/cas9基因编辑载体构建
基因编辑载体构建参考mao等(maoy,etal.,molplant,2013,6(6):2008-2011.)报道方法,按以下步骤进行:
(1)靶点接头制备
将接头引物(als-u3-f:5’-ggcatccttgaatgcgcccccact-3’;als-u3-r:5’-aaacagtgggggcgcattcaagga-3’)用1xte(ph8.0)溶解成100μm母液,各取1μl加入到98μl0.5xte混合稀释到1μm。约90℃30s,移至室温冷却完成退火,即获得靶点接头。
(2)sgrna表达盒制备
按以下反应体系进行pcr扩增:
注:t4dnaligase及10xdnaligasebuffer购于takara,bsai购于neb。
pcr反应程序为37℃5min,20℃5min,5个循环。获得的pcr产物即为sgrna连接产物。
pylsgrna-osu3为中间载体,为sgrna表达盒提供启动子和引导序列骨架,由华南农业大学刘耀光教授团队开发(max,zhangq,zhuq,etal.arobustcrispr/cas9systemforconvenient,high-efficiencymultiplexgenomeeditinginmonocotanddicotplants.molplant,2015,8(8):1274-1284.)。
(3)扩增sgrna表达盒
以引物组合正向引物u-f:5’-ctccgttttacctgtggaatcg-3’和反向引物grna-r:5’–cggaggaaaattccatccac-3’,按照以下反应体系进行pcr扩增:
其中,primestarhsdnapolymerase、dntpmix和2×primestargcbuffer均购于takara。在eppendorfmastercycle热循环仪中进行pcr。pcr反应程序:95℃1min;95℃10s,60℃15s,68℃20s,10个循环;95℃10s,60℃15s,68℃30s,22个循环;4℃保存。
以uctcg-b1’:ttcagaggtctctctcgcactggaatcggcagcaaagg-3;grcggt-bl:agcgtgggtctcgaccgggtccatccactccaagctc-3为扩增引物,按以下反应体系进行pcr扩增:
在eppendorfmastercycle热循环仪中进行pcr。pcr反应程序:95℃10s,60℃15s,68℃20s,25个循环;4℃保存。获得的pcr产物即为sgrna表达盒。
(4)sgrna表达盒连接pylcrispr/cas9p35s-h载体
按以下反应体系和过程,在eppendorfmastercycle热循环仪中,将sgrna表达盒连接pylcrispr/cas9p35s-h载体,获得连接产物。
反应体系及过程:
pylcrispr/cas9p35s-h载体为植物双元表达载体,由华南农业大学刘耀光教授团队开发(max,zhangq,zhuq,etal.arobustcrispr/cas9systemforconvenient,high-efficiencymultiplexgenomeeditinginmonocotanddicotplants.molplant,2015,8(8):1274-1284.)
(5)转化大肠杆菌dh5α及验证
将连接产物用热激法(42℃)转化大肠杆菌dh5α,菌液涂布于含有50mg/l卡那霉素的lb平板上,培养约12h。挑取平板上长出的单菌落,摇菌扩繁。以菌液为模板进行pcr验证。
pcr反应体系为:
taqdna聚合酶购于北京鼎国昌盛生物技术有限公司。
在eppendorfmastercycle热循环仪中进行pcr。pcr反应程序:95℃10min;95℃30s,51℃30s,72℃45s,28个循环;72℃5min;4℃保存。扩增产物经琼脂糖凝胶电泳分离,用凝胶成像仪拍照并记录结果。提取pcr检测含目的条带的菌液的质粒,送往英潍捷基(上海)贸易有限公司测序。
(6)抗除草剂突变体的获得
将上述阳性质粒转入农杆菌eha105。采用常规农杆菌介导法转化水稻南粳9108(购自江苏高科种业有限公司)。为提高获得抗性植株的概率,尽量多获得转化植株,本发明共获得了58个转基因植株(t0代)。
将t0代植株以单株收种,收获的种子催芽后,按450kghm-2密度播种。在水稻长至二叶一心时,排干田里的水,将咪草烟(水剂,购自南京艾津农化有限责任公司)以210g(a.i.)hm-2喷施,喷洒24h后复水,14d后调查抗性。植株叶片全部枯萎或死亡为感,植株健康存活为抗。在58个株系(a1~a58)中,只有a51株系中有18个单株存活,表现为抗除草剂,其它57个株系植株全部死亡(图2),因此,本研究获得抗除草剂株系的频率为1.72%。根据报道,利用化学诱变的方法获得抗除草剂水稻株系的突变频率为0.00003~0.006%。因此,本发明采用的基因编辑育种方法获得抗除草剂株系的效率是化学诱变方法的285倍以上,显著优于化学诱变方法。
为了鉴定编辑位点的碱基变异,以引物alst-f:cgcatacatacttgggcaac和alst-r:acaaacatcataggcataccac对部分t0代转基因株系进行扩增并测序。结果如图1所示,a5未发生突变;a3、a9和a24为杂合突变,这些单株的一个等位基因未突变,另一个等位基因缺失1或2个碱基而发生移码突变;a51为双等位基因变异,其中一个等位基因为第1882位点碱基发生由g到t变异,另一个等位基因为缺失了1882位的g碱基(图1)。
实施例2:抗咪唑啉酮类除草剂水稻突变体als基因克隆
对上述实施例1的a51株系的t1代18个单株进行编号,取它们的植株叶片,提取基因组dna,以als基因全长的特异引物als-f5’-tcgcccaaacccagaaaccc-3’和als-r5’-ctctttatgggtcattcaggtc-3’进行pcr扩增。扩增产物送英潍捷基(上海)贸易有限公司进行测序。测序结果与南粳9108野生型als基因比较,发现编号1~9、11、14、16和17的单株为第1882位点碱基发生由g到t的纯合突变;编号10、12、13和15的单株发生双等位变异,其中一个等位基因为第1882位点碱基发生由g到t变异,另一个等位基因为缺失了1882位的g碱基;没有获得1882位碱基缺失的纯合单株,推测碱基缺失的移码突变水稻不能存活。综合1882位点g到t的纯合突变或杂合突变都对除草剂产生抗性,推测该突变为产生除草剂抗性的关键突变。
进一步分析抗除草剂水稻突变体在als基因第1882位点碱基发生由g到t的突变,导致第628位氨基酸由甘氨酸突变为色氨酸。抗除草剂突变体的als基因的核苷酸序列如seqidno.1所示,其编码的als蛋白的氨基酸序列如seqidno:2所示,将克隆得到的新基因命名为als-nj。
本发明鉴定到的als-nj基因第1882位碱基由野生型g到t的变异,以及因此引起的第628位氨基酸由甘氨酸突到色氨酸的变异均为首次报道。
实施例3抗咪唑啉酮类除草剂水稻突变体t-dna剔除
本发明构建的定向编辑als基因的双元t-dna载体,本发明涉及的t-dna主要包含了潮霉素磷酸转移酶hpt基因和cas9核酸酶基因,由于潮霉素磷酸转移酶hpt基因和cas9基因的主要作用是完成对目标基因的定点突变,并且这两个基因相对于水稻基因组来说是外源基因,一方面潮霉素是抗生素,需要剔除,cas9基因如果保留还可能导致继续编辑等功能;另一方面,t-dna随机插入还可能导致非预期的基因突变,所以在其完成基因编辑任务后,需要清除。通过农杆菌介导转化南粳9108,在转基因过程中,t-dna序列会随机插入水稻的染色体中,可能以单拷贝或多拷贝插入。由于t-dna插入位点与其靶位点一般不连锁,因此有望通过转基因植株的后代分离获得不携带t-dna的植株,即使连锁也可以通过遗传交换重组筛选到不携带t-dna的材料。因此,为了获得不含上述t-dna的植株,本发明人通过对目标基因双突变的t1代植株hpt基因和cas9基因的同时检测,重复3次,筛选不携带这两个基因,即为剔除t-dna的t1代单株。
取上述实施例2的18个单株的基因组dna,以引物hyg283-f:
tccggaagtgcttgacatt和hyg283-r:gtcgtccatcacagtttgc对hpt基因进行pcr扩增;以引物cas9t-f:agcggcaagactatcctcgact和cas9t-r:
tcaatcctcttcatgcgctccc对cas9基因进行pcr扩增。结果如图3所示,编号为1、12和18单株未检测到hpt基因和cas9基因,表明这3个单株成功剔除了外源t-dna。本例中18个单株中有3株已通过重组剔除t-dna,获得剔除t-dna植株的比例为六分之一,推测t-dna是以多拷贝方式插入到水稻基因组中。
实施例4突变体a51对咪草烟(咪唑乙烟酸,咪唑啉酮类除草剂)的抗性鉴定
取实施例2和3中鉴定到的剔除t-dna的纯合突变t1单株进行繁种,于南京江苏省农业科学院人工气候室收获种子,即为t2代;将t2代继续繁殖,获得t3代种子。将收获的t3种子催芽后,按450kghm-2密度播种。在水稻长至二叶一心时,排干田间水,分别以210、700、1400g(a.i.)hm-2浓度咪草烟(水剂,购自南京艾津农化有限责任公司)喷施,以水为对照组。喷洒24h后复水,14d后调查抗性。如图4所示,无论野生型还是突变体,在喷施水的对照组中都能正常生长;野生型在210、700、1400g(a.i.)hm-2浓度咪草烟处理下均死亡;突变体在210、700、1400g(a.i.)hm-2浓度咪草烟处理均能存活。以上结果表明,突变体能抗咪草烟除草剂,而且能稳定遗传给下一代。
实施例5突变体a51对百垄通(甲咪唑烟酸,咪唑啉酮类除草剂)的抗性鉴定
将实施例4中的将t3代种子催芽后,按450kghm-2密度播种。在水稻长至二叶一心时,排干田间水,分别以240、2400、4800g(a.i.)hm-2浓度百垄通(水剂,购自南京艾津农化有限责任公司)喷施,以水为对照组。喷洒24h后复水,14d后调查抗性。如图5所示,无论野生型还是突变体,在喷施水的对照组中都能正常生长;野生型在240、2400、4800g(a.i.)hm-2浓度百垄通处理后均死亡;突变体在240和2400g(a.i.)hm-2浓度咪草烟处理均能存活,在4800g(a.i.)hm-2浓度百垄通处理后死亡。以上结果表明,突变体能抗浓度为2400g(a.i.)hm-2甲咪唑烟酸,而且其抗性能稳定遗传给下一代。
实施例6:突变体的农艺性状调查
将野生型与取实施例2和3中鉴定到的剔除t-dna的纯合突变体种植于海南省三亚市试验基地,野生型与突变体种植小区,每小区200苗,三次重复。对农艺性状进行分析发现,野生型和突变体的株高、有效穗、穗长、每穗粒数、结实率、千粒重等6个产量构成性状进行比较,经t测验,差异不显著(p<0.05)(图6),其他如抽穗期、株叶形态、叶片颜色、稻米香味、稻米外观(呈云雾状)等农艺性状均无显著差异。因此,编辑获得的抗除草剂新材料保留了野生型材料的丰产、优质等重要农艺性状。
实施例7:als-nj基因遗传特性及其功能标记开发和应用
分子标记辅助选择有利于加快育种进程。本发明的als-nj基因为单碱基突变,可以针对性地设计酶切靶点标记,但其过程相对繁琐,开发等位基因特异pcr,通过两次pcr就可以区分抗感基因型,操作简便快速。本发明针对野生型与突变体在als基因第1882位点的碱基变异,利用等位基因特异pcr原理,设计了13组引物als1~als13(表1)。als1~als7共用上游引物als-1f,下游引物分别为als-1r、als-2r、als-3r、als-4r、als-5r、als-6r和als-7r。als7~als13共用下游引物als-1r,上游引物分别为als-2f、als-3f、als-4f、als-5f、als-6f和als-7f。为了进一步提高引物的特异性,在部分引物的3’端引入了碱基错配,als-3f和als-6f引物的3’到5’第3个碱基由g错配为a,als-4f和als-7f引物的3’到5’第3个碱基由g错配为c,als-3r和als-6r引物的3’到5’第3个碱基由c错配为t,als-4r和als-7r引物的3’到5’第3个碱基由c错配为a。
经多轮筛选和优化pcr反应条件,发现引物对als4和als6均具有很好的扩增效率和特异性,可以分别作为区分野生型、突变体基因型以及杂合基因型的引物对(图7)。pcr的最佳反应体系为:野生型或突变体dna模板2μl,10×pcrbuffer2μl,mgcl2(5mmol/l)2μl,dntp(2mmol/l)2μl,上游引物2μl,下游引物2μl,taq酶(2.5u/μl)0.2μl,ddh2o7.8μl。pcr反应程序为95℃10min;95℃30s,60℃30s,72℃45s,35个循环;72℃5min;4℃保存。将als4和als6组成的分子标记命名为als628w。
表1检测突变体基因的分子标记
注:小写字母标出的碱基为错配碱基。
利用als628w标记检测水稻品种,发现在检测品种中,只有南粳9108突变体能被als6扩增出条带,其余粳稻或籼稻品种(南粳9108野生型、日本晴、黄华占、9311、连粳7号、苏秀867、镇稻88、镇稻99、淮稻5号、常农粳8号、南粳44、南粳45、南粳46、南粳49、南粳51、南粳47、南粳5055、苏垦118、武运粳24号、武运粳27号、武运粳29号、徐稻8号、徐稻9号、扬育粳2号、华粳5号、盐稻16号)均只能被als4扩增出条带(图8),表明als628w标记能特异检测出als-nj基因1882位碱基由g到t的变异,该标记可以用于分子标记辅助选择育种。
为了验证als628w标记在抗除草剂品系选育中的应用,进一步利用als628w标记检测徐稻9号、南粳9108突变体、“徐稻9号/南粳9108突变体”杂交种及它们的132个f2单株,并进行表型鉴定。“徐稻9号/南粳9108突变体”杂交种表现为抗除草剂,f2单株中,102个单株表现为抗除草剂,30个单株表现为感除草剂,经卡方检验,分离比例符合3:1(χ2=0.2525,p>0.05)。
综合抗感亲本、杂交种f1以及f2的抗感分离比例,als-nj基因的除草剂抗性是由单基因控制的显性性状。结合标记检测结果发现,所有携带突变型基因的f2群体单株都表现为抗除草剂,而所有不携带突变型基因的f2群体单株均表现为感除草剂(图9)。基因型检测结果与表型鉴定结果完全对应。als628w标记与抗/感除草剂表型完全共分离,同时还能检测到抗性杂合基因型,表明我们开发的als628w标记可用于抗咪草烟等除草剂育种的精准选育,在f2代筛选抗除草剂的纯合基因型,可以进行早代选择。
通过上述具体实施例表明,利用crispr/cas9基因编辑技术,对als基因进行编辑,通过后代的筛选,在t2代就可以获得剔除t-dna、抗除草剂特性稳定遗传的新材料,而新材料的基本农艺性状无明显改变。相对于化学诱变、杂交转育等育种,基因编辑定向改良分子育种技术具有快速、精准、高效等优点,结合基因功能标记基因型选择,将会极大提高育种效率,大大加快育种进程(表2)。
表2基因编辑育种方法与传统育种方法的比较
尽管本发明的具体实施方式已经得到详细描述,本领域技术人员将会理解。根据已经公开的所有教导,可以对那些细节进行各种修改和替换,这些均在本发明的保护范围内。本发明的全部范围由所附专利要求极其任何等同物给出。
序列表
<110>江苏省农业科学院
<120>基于基因编辑技术的als突变型蛋白及其基因在植物育种中的应用
<160>31
<170>siposequencelisting1.0
<210>1
<211>1935
<212>dna
<213>als-nj突变型基因(acetolactatesynthase)
<400>1
atggctacgaccgccgcggccgcggccgccgccctgtccgccgccgcgacggccaagacc60
ggccgtaagaaccaccagcgacaccacgtccttcccgctcgaggccgggtgggggcggcg120
gcggtcaggtgctcggcggtgtccccggtcaccccgccgtccccggcgccgccggccacg180
ccgctccggccgtgggggccggccgagccccgcaagggcgcggacatcctcgtggaggcg240
ctggagcggtgcggcgtcagcgacgtgttcgcctacccgggcggcgcgtccatggagatc300
caccaggcgctgacgcgctccccggtcatcaccaaccacctcttccgccacgagcagggc360
gaggcgttcgcggcgtccgggtacgcgcgcgcgtccggccgcgtcggggtctgcgtcgcc420
acctccggccccggggcaaccaacctcgtgtccgcgctcgccgacgcgctgctcgactcc480
gtcccgatggtcgccatcacgggccaggtcccccgccgcatgatcggcaccgacgccttc540
caggagacgcccatagtcgaggtcacccgctccatcaccaagcacaattaccttgtcctt600
gatgtggaggacatcccccgcgtcatacaggaagccttcttcctcgcgtcctcgggccgt660
cctggcccggtgctggtcgacatccccaaggacatccagcagcagatggccgtgccggtc720
tgggacacctcgatgaatctaccagggtacatcgcacgcctgcccaagccacccgcgaca780
gaattgcttgagcaggtcttgcgtctggttggcgagtcacggcgcccgattctctatgtc840
ggtggtggctgctctgcatctggtgacgaattgcgctggtttgttgagctgactggtatc900
ccagttacaaccactctgatgggcctcggcaatttccccagtgacgacccgttgtccctg960
cgcatgcttgggatgcatggcacggtgtacgcaaattatgccgtggataaggctgacctg1020
ttgcttgcgtttggtgtgcggtttgatgatcgtgtgacagggaaaattgaggcttttgca1080
agcagggccaagattgtgcacattgacattgatccagcagagattggaaagaacaagcaa1140
ccacatgtgtcaatttgcgcagatgttaagcttgctttacagggcttgaatgctctgcta1200
caacagagcacaacaaagacaagttctgattttagtgcatggcacaatgagttggaccag1260
cagaagagggagtttcctctggggtacaaaacttttggtgaagagatcccaccgcaatat1320
gccattcaggtgctggatgagctgacgaaaggtgaggcaatcatcgctactggtgttggg1380
cagcaccagatgtgggcggcacaatattacacctacaagcggccacggcagtggctgtct1440
tcggctggtctgggcgcaatgggatttgggctgcctgctgcagctggtgcttctgtggct1500
aacccaggtgtcacagttgttgatattgatggggatggtagcttcctcatgaacattcag1560
gagctggcattgatccgcattgagaacctccctgtgaaggtgatggtgttgaacaaccaa1620
catttgggtatggtggtgcaatgggaggataggttttacaaggcgaatagggcgcataca1680
tacttgggcaacccggaatgtgagagcgagatatatccagattttgtgactattgctaag1740
gggttcaatattcctgcagtccgtgtaacaaagaagagtgaagtccgtgccgccatcaag1800
aagatgctcgagactccagggccatacttgttggatatcatcgtcccgcaccaggagcat1860
gtgctgcctatgatcccaagttggggcgcattcaaggacatgatcctggatggtgatggc1920
aggactgtgtattaa1935
<210>2
<211>644
<212>prt
<213>als突变型蛋白(acetolactatesynthase)
<400>2
metalathrthralaalaalaalaalaalaalaleuseralaalaala
151015
thralalysthrglyarglysasnhisglnarghishisvalleupro
202530
alaargglyargvalglyalaalaalavalargcysseralavalser
354045
provalthrproproserproalaproproalathrproleuargpro
505560
trpglyproalagluproarglysglyalaaspileleuvalgluala
65707580
leugluargcysglyvalseraspvalphealatyrproglyglyala
859095
sermetgluilehisglnalaleuthrargserprovalilethrasn
100105110
hisleuphearghisgluglnglyglualaphealaalaserglytyr
115120125
alaargalaserglyargvalglyvalcysvalalathrserglypro
130135140
glyalathrasnleuvalseralaleualaaspalaleuleuaspser
145150155160
valprometvalalailethrglyglnvalproargargmetilegly
165170175
thraspalapheglngluthrproilevalgluvalthrargserile
180185190
thrlyshisasntyrleuvalleuaspvalgluaspileproargval
195200205
ileglnglualaphepheleualaserserglyargproglyproval
210215220
leuvalaspileprolysaspileglnglnglnmetalavalproval
225230235240
trpaspthrsermetasnleuproglytyrilealaargleuprolys
245250255
proproalathrgluleuleugluglnvalleuargleuvalglyglu
260265270
serargargproileleutyrvalglyglyglycysseralasergly
275280285
aspgluleuargtrpphevalgluleuthrglyileprovalthrthr
290295300
thrleumetglyleuglyasnpheproseraspaspproleuserleu
305310315320
argmetleuglymethisglythrvaltyralaasntyralavalasp
325330335
lysalaaspleuleuleualapheglyvalargpheaspaspargval
340345350
thrglylysileglualaphealaserargalalysilevalhisile
355360365
aspileaspproalagluileglylysasnlysglnprohisvalser
370375380
ilecysalaaspvallysleualaleuglnglyleuasnalaleuleu
385390395400
glnglnserthrthrlysthrserserasppheseralatrphisasn
405410415
gluleuaspglnglnlysargglupheproleuglytyrlysthrphe
420425430
glyglugluileproproglntyralaileglnvalleuaspgluleu
435440445
thrlysglyglualaileilealathrglyvalglyglnhisglnmet
450455460
trpalaalaglntyrtyrthrtyrlysargproargglntrpleuser
465470475480
seralaglyleuglyalametglypheglyleuproalaalaalagly
485490495
alaservalalaasnproglyvalthrvalvalaspileaspglyasp
500505510
glyserpheleumetasnileglngluleualaleuileargileglu
515520525
asnleuprovallysvalmetvalleuasnasnglnhisleuglymet
530535540
valvalglntrpgluaspargphetyrlysalaasnargalahisthr
545550555560
tyrleuglyasnproglucysglusergluiletyrproasppheval
565570575
thrilealalysglypheasnileproalavalargvalthrlyslys
580585590
sergluvalargalaalailelyslysmetleugluthrproglypro
595600605
tyrleuleuaspileilevalprohisglngluhisvalleupromet
610615620
ileprosertrpglyalaphelysaspmetileleuaspglyaspgly
625630635640
argthrvaltyr
<210>3
<211>1935
<212>dna
<213>als野生型基因(acetolactatesynthase)
<400>3
atggctacgaccgccgcggccgcggccgccgccctgtccgccgccgcgacggccaagacc60
ggccgtaagaaccaccagcgacaccacgtccttcccgctcgaggccgggtgggggcggcg120
gcggtcaggtgctcggcggtgtccccggtcaccccgccgtccccggcgccgccggccacg180
ccgctccggccgtgggggccggccgagccccgcaagggcgcggacatcctcgtggaggcg240
ctggagcggtgcggcgtcagcgacgtgttcgcctacccgggcggcgcgtccatggagatc300
caccaggcgctgacgcgctccccggtcatcaccaaccacctcttccgccacgagcagggc360
gaggcgttcgcggcgtccgggtacgcgcgcgcgtccggccgcgtcggggtctgcgtcgcc420
acctccggccccggggcaaccaacctcgtgtccgcgctcgccgacgcgctgctcgactcc480
gtcccgatggtcgccatcacgggccaggtcccccgccgcatgatcggcaccgacgccttc540
caggagacgcccatagtcgaggtcacccgctccatcaccaagcacaattaccttgtcctt600
gatgtggaggacatcccccgcgtcatacaggaagccttcttcctcgcgtcctcgggccgt660
cctggcccggtgctggtcgacatccccaaggacatccagcagcagatggccgtgccggtc720
tgggacacctcgatgaatctaccagggtacatcgcacgcctgcccaagccacccgcgaca780
gaattgcttgagcaggtcttgcgtctggttggcgagtcacggcgcccgattctctatgtc840
ggtggtggctgctctgcatctggtgacgaattgcgctggtttgttgagctgactggtatc900
ccagttacaaccactctgatgggcctcggcaatttccccagtgacgacccgttgtccctg960
cgcatgcttgggatgcatggcacggtgtacgcaaattatgccgtggataaggctgacctg1020
ttgcttgcgtttggtgtgcggtttgatgatcgtgtgacagggaaaattgaggcttttgca1080
agcagggccaagattgtgcacattgacattgatccagcagagattggaaagaacaagcaa1140
ccacatgtgtcaatttgcgcagatgttaagcttgctttacagggcttgaatgctctgcta1200
caacagagcacaacaaagacaagttctgattttagtgcatggcacaatgagttggaccag1260
cagaagagggagtttcctctggggtacaaaacttttggtgaagagatcccaccgcaatat1320
gccattcaggtgctggatgagctgacgaaaggtgaggcaatcatcgctactggtgttggg1380
cagcaccagatgtgggcggcacaatattacacctacaagcggccacggcagtggctgtct1440
tcggctggtctgggcgcaatgggatttgggctgcctgctgcagctggtgcttctgtggct1500
aacccaggtgtcacagttgttgatattgatggggatggtagcttcctcatgaacattcag1560
gagctggcattgatccgcattgagaacctccctgtgaaggtgatggtgttgaacaaccaa1620
catttgggtatggtggtgcaatgggaggataggttttacaaggcgaatagggcgcataca1680
tacttgggcaacccggaatgtgagagcgagatatatccagattttgtgactattgctaag1740
gggttcaatattcctgcagtccgtgtaacaaagaagagtgaagtccgtgccgccatcaag1800
aagatgctcgagactccagggccatacttgttggatatcatcgtcccgcaccaggagcat1860
gtgctgcctatgatcccaagtgggggcgcattcaaggacatgatcctggatggtgatggc1920
aggactgtgtattaa1935
<210>4
<211>644
<212>prt
<213>als野生型蛋白(acetolactatesynthase)
<400>4
metalathrthralaalaalaalaalaalaalaleuseralaalaala
151015
thralalysthrglyarglysasnhisglnarghishisvalleupro
202530
alaargglyargvalglyalaalaalavalargcysseralavalser
354045
provalthrproproserproalaproproalathrproleuargpro
505560
trpglyproalagluproarglysglyalaaspileleuvalgluala
65707580
leugluargcysglyvalseraspvalphealatyrproglyglyala
859095
sermetgluilehisglnalaleuthrargserprovalilethrasn
100105110
hisleuphearghisgluglnglyglualaphealaalaserglytyr
115120125
alaargalaserglyargvalglyvalcysvalalathrserglypro
130135140
glyalathrasnleuvalseralaleualaaspalaleuleuaspser
145150155160
valprometvalalailethrglyglnvalproargargmetilegly
165170175
thraspalapheglngluthrproilevalgluvalthrargserile
180185190
thrlyshisasntyrleuvalleuaspvalgluaspileproargval
195200205
ileglnglualaphepheleualaserserglyargproglyproval
210215220
leuvalaspileprolysaspileglnglnglnmetalavalproval
225230235240
trpaspthrsermetasnleuproglytyrilealaargleuprolys
245250255
proproalathrgluleuleugluglnvalleuargleuvalglyglu
260265270
serargargproileleutyrvalglyglyglycysseralasergly
275280285
aspgluleuargtrpphevalgluleuthrglyileprovalthrthr
290295300
thrleumetglyleuglyasnpheproseraspaspproleuserleu
305310315320
argmetleuglymethisglythrvaltyralaasntyralavalasp
325330335
lysalaaspleuleuleualapheglyvalargpheaspaspargval
340345350
thrglylysileglualaphealaserargalalysilevalhisile
355360365
aspileaspproalagluileglylysasnlysglnprohisvalser
370375380
ilecysalaaspvallysleualaleuglnglyleuasnalaleuleu
385390395400
glnglnserthrthrlysthrserserasppheseralatrphisasn
405410415
gluleuaspglnglnlysargglupheproleuglytyrlysthrphe
420425430
glyglugluileproproglntyralaileglnvalleuaspgluleu
435440445
thrlysglyglualaileilealathrglyvalglyglnhisglnmet
450455460
trpalaalaglntyrtyrthrtyrlysargproargglntrpleuser
465470475480
seralaglyleuglyalametglypheglyleuproalaalaalagly
485490495
alaservalalaasnproglyvalthrvalvalaspileaspglyasp
500505510
glyserpheleumetasnileglngluleualaleuileargileglu
515520525
asnleuprovallysvalmetvalleuasnasnglnhisleuglymet
530535540
valvalglntrpgluaspargphetyrlysalaasnargalahisthr
545550555560
tyrleuglyasnproglucysglusergluiletyrproasppheval
565570575
thrilealalysglypheasnileproalavalargvalthrlyslys
580585590
sergluvalargalaalailelyslysmetleugluthrproglypro
595600605
tyrleuleuaspileilevalprohisglngluhisvalleupromet
610615620
ileproserglyglyalaphelysaspmetileleuaspglyaspgly
625630635640
argthrvaltyr
<210>5
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>5
tccttgaatgcgcccccact20
<210>6
<211>19
<212>dna
<213>als-1f(artificialsequence)
<400>6
atccgcattgagaacctcc19
<210>7
<211>20
<212>dna
<213>als-4r(artificialsequence)
<400>7
atgtccttgaatgcgccacc20
<210>9
<211>20
<212>dna
<213>als-6r(artificialsequence)
<400>9
atgtccttgaatgcgcctca20
<210>10
<211>20
<212>dna
<213>als5-f(artificialsequence)
<400>10
tcgcccaaacccagaaaccc20
<210>11
<211>22
<212>dna
<213>als5-r(artificialsequence)
<400>11
ctctttatgggtcattcaggtc22
<210>12
<211>24
<212>dna
<213>als-u3-f(artificialsequence)
<400>12
ggcatccttgaatgcgcccccact24
<210>13
<211>24
<212>dna
<213>als-u3-r(artificialsequence)
<400>13
aaacagtgggggcgcattcaagga24
<210>14
<211>22
<212>dna
<213>u-f(artificialsequence)
<400>14
ctccgttttacctgtggaatcg22
<210>15
<211>20
<212>dna
<213>grna-r(artificialsequence)
<400>15
cggaggaaaattccatccac20
<210>16
<211>38
<212>dna
<213>uctcg-b1(artificialsequence)
<400>16
ttcagaggtctctctcgcactggaatcggcagcaaagg38
<210>17
<211>37
<212>dna
<213>grcggt-bl(artificialsequence)
<400>17
agcgtgggtctcgaccgggtccatccactccaagctc37
<210>18
<211>20
<212>dna
<213>alst-f(artificialsequence)
<400>18
cgcatacatacttgggcaac20
<210>19
<211>22
<212>dna
<213>alst-r(artificialsequence)
<400>19
acaaacatcataggcataccac22
<210>20
<211>20
<212>dna
<213>als-f(artificialsequence)
<400>20
tcgcccaaacccagaaaccc20
<210>21
<211>22
<212>dna
<213>als-r(artificialsequence)
<400>21
ctctttatgggtcattcaggtc22
<210>22
<211>19
<212>dna
<213>hyg283-f(artificialsequence)
<400>22
tccggaagtgcttgacatt19
<210>23
<211>19
<212>dna
<213>hyg283-r(artificialsequence)
<400>23
gtcgtccatcacagtttgc19
<210>24
<211>22
<212>dna
<213>cas9t-f(artificialsequence)
<400>24
agcggcaagactatcctcgact22
<210>25
<211>22
<212>dna
<213>cas9t-r(artificialsequence)
<400>25
tcaatcctcttcatgcgctccc22
<210>27
<211>21
<212>dna
<213>als-1r(artificialsequence)
<400>27
taggattaccatgccaagcac21
<210>29
<211>20
<212>dna
<213>als-2r(artificialsequence)
<400>29
atgtccttgaatgcgccccc20
<210>28
<211>20
<212>dna
<213>als-3r(artificialsequence)
<400>28
atgtccttgaatgcgcctcc20
<210>28
<211>20
<212>dna
<213>als-4r(artificialsequence)
<400>28
atgtccttgaatgcgccacc20
<210>29
<211>20
<212>dna
<213>als-5r(artificialsequence)
<400>29
atgtccttgaatgcgcccca20
<210>30
<211>20
<212>dna
<213>als-6r(artificialsequence)
<400>30
atgtccttgaatgcgcctca20
<210>31
<211>20
<212>dna
<213>als-7r(artificialsequence)
<400>31
atgtccttgaatgcgccaca20
- 还没有人留言评论。精彩留言会获得点赞!