利用Bac-to-Bac真核表达系统获得甜菜夜蛾脱饱和酶SexiDes11的方法与流程
本发明属于生物技术领域,具体涉及一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法。
背景技术
在鳞翅目昆虫的交流通讯系统中,雌虫通过信息素腺体产生并释放具有种特异的性信息素组分来吸引同种雄虫进行交配。绝大多数蛾类的性信息素是一些含c10-c18的不饱和酯类、醇类和醛类,这些物质间的不同比例组合具有种特异性,因此性信息素在蛾类昆虫生殖隔离和物种形成过程中有着重要影响。近些年来,国内外针对鳞翅目昆虫性信息素合成途径开展了较多的研究。
酰基coa脱饱和酶(acyl-coadesaturase)是鳞翅目昆虫性信息素合成过程中的一类关键酶,它能够催化饱和脂肪酸形成一个或多个不饱和键,其对底物的区域和立体专一性使得性信息素结构具有多样性。根据系统发育关系和酶活性的特征,roelofs等学者将蛾类脱饱和酶分为四个组:δ9(16>18),δ9(18>16),δ10,11和δ14,随着研究的深入,将会有更多具有新功能的脱饱和酶加入到这个家族,这表明这类基因家族有着复杂的进化模式。因此开展鳞翅目昆虫脱饱和酶的研究不仅有助于阐明性信息素的合成途径,还将有利于构建出鳞翅目昆虫的依赖性信息素的进化模式。
董双林等学者鉴定了我国甜菜夜蛾的性信息素成分并在田间试验中得到成功应用,但是,对于甜菜夜蛾性信息素合成途径中所涉及的关键酶却鲜有文献报道。目前,仅有jurenka在1997年通过同位素标记的体外试验认为甜菜夜蛾体内存在δ12脱饱和酶,但没有该酶基因克隆的报道。
因此,现有技术存在的问题是,缺乏一种可成功表达并获得甜菜夜蛾脱饱和酶sexides11的方法。
技术实现要素:
本发明提供的一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,可成功表达并获得甜菜夜蛾脱饱和酶sexides11。
本发明的是提供一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,包括以下步骤:
s1,提取甜菜夜蛾总rna,合成甜菜夜蛾cdna第一链;
s2,连接反应,获得重组质粒peasy-sexides
分别将peasy-t3载体和甜菜夜蛾cdna第一链经sali和hindiii双酶切,分别得到peasy-t3酶切产物和甜菜夜蛾cdna酶切产物,将二者连接,得到重组质粒peasy-sexides;
s3,构建甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides
分别将重组质粒peasy-sexides和pfastbac1载体经sali和hindiii双酶切,分别得到peasy-sexides酶切产物和pfastbac1酶切产物,将二者连接,得到甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides;
s4,利用甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides和昆虫细胞构建bac-to-bac真核表达系统,利用bac-to-bac真核表达系统表达甜菜夜蛾脱饱和酶sexides11。
优选的,上述的利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,s3中,双酶切反应体系如下:
优选的,上述的利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,s4的步骤包括:
s41,培养昆虫细胞,获得昆虫细胞株;
s42,构建携带sexides11基因的重组病毒
s421,制备dh10bac感受态细胞;
s422,转座:将甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides转入dh10bac感受态细胞中进行转座,获得重组病毒bacmiddna的细胞;该重组病毒bacmiddna上携带sexides11基因;
s423,从含有重组病毒bacmiddna的细胞中提取重组病毒bacmiddna;
s424,利用重组病毒bacmiddna对昆虫细胞进行转染及感染,提取能表达甜菜夜蛾脱饱和酶sexides11的病毒体,然后利用该病毒体转染再次感染昆虫细胞并表达出甜菜夜蛾脱饱和酶sexides11。
优选的,上述的利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,用非感受态大肠杆菌dh5α制备dh10bac感受态细胞。
优选的,上述的利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,所述昆虫细胞为high5细胞。
与现有技术相比,本发明的利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶(sexides11)的方法,具有以下有益效果:
本发明采用race技术克隆得到了甜菜夜蛾饱和酶sexides11基因的全长序列,利用rt-pcr技术进行了组织表达谱研究,并通过bac-to-bac真核表达系统对这个基因进行体外蛋白表达,结合westernblot对表达蛋白进行验证。
本方法基于昆虫真核细胞表达系统,主要优点为:1)表达基因和所用细胞均来自鳞翅目夜蛾科,因此表达出的甜菜夜蛾脱饱和酶sexides11更具真实性;2)表达过程中对甜菜夜蛾脱饱和酶sexides11的结构和特性能够进行转录后修饰,更贴近原始功能,优于大肠杆菌等原核表达系统;3)细胞繁殖快,培养周期短,仅为1-2个月,培养条件要求不高,为科学研究或规模化生产提供助力。
附图说明
图1为利用rt-pcr技术研究甜菜夜蛾脱饱和酶基因的组织表达谱;
图2为重组病毒bacmiddna的pcr产物电泳结果;
其中m泳道为dl4000marker;另一泳道为sexides11基因;
图3为用不同病毒转染high5细胞2天后显微镜观察图,其中图3a是正常high5细胞,图3b是p1代病毒感染high5细胞,图3c是重组的绿色荧光蛋白基因(gfp)病毒感染的high5细胞;
图4为甜菜夜蛾脱饱和酶蛋白的westernblot免疫分析结果。
具体实施方式
下面结合具体实施例对本发明进行详细说明,但不应理解为本发明的限制。下列实施例中未注明具体条件的试验方法,通常按照常规条件操作,由于不涉及发明点,故不对其步骤进行详细描述。
本发明提供的一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,包括以下实施例。
下述实施例中所用材料和试剂如下:
供试昆虫:甜菜夜蛾为室内人工饲料饲养,饲料配方采用常规配方。饲养温度为26℃,湿度为70%-80%,光周期14l:10d。
主要试剂:svtotalrnaisolation(promega,usa)、m-mlv反转录酶、rnase-freednaseⅰ、oligo(dt)18随机引物、dntpmixture(2.5mm和10mm)和rnase抑制剂等均购自takara公司;pcr引物由上海生工生物工程公司合成。l-glutamine、cellfectin为invitrogen公司产品。限制性内切酶salⅰ、hindⅲ、t4dna连接酶均购自fermentas公司。碱性磷酸酶购自promega公司。卡那霉素(kanamycin)(50mg/ml),四环素(tetracycline)(10mg/ml),庆大霉素(genlamycin)(7mg/ml)为国产试剂。
实施例1
一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法,包括以下步骤:
s1,提取甜菜夜蛾总rna,合成甜菜夜蛾cdna第一链;
s11,提取甜菜夜蛾总rna:用svtotalrnaisolation试剂盒进行总rna提取,获得甜菜夜蛾总rna。
s12,合成甜菜夜蛾cdna
s121,合成cdna第一链
依据takaratmm-mlv反转录试剂盒进行cdna第一链的合成,合成的甜菜夜蛾cdna第一链置于-20℃保存备用。其中合成甜菜夜蛾cdna第一链所用的引物序列是oligodt:tttttttttttttttttt,如seqidno.1所示。
s122,race扩增基因的全长cdna
(1)5′-racecdna第一链的合成
以甜菜夜蛾总rna为模板,依据clontechtmsmarter反转录试剂盒的操作步骤,进行5’-racecdna第一链的合成。其中该步骤使用的扩增引物序列是5'-cdsprimeraagcagtggtatcaacgcagagt,seqidno.2所示;
(2)3′-racecdna第一链的合成
甜菜夜蛾总rna为模板,依据invitrogentmsuperscriptiii反转录试剂盒完成3’-racecdna第一链的合成。其中该步骤使用的扩增引物序列是gene-specificprimerctaatacgactcactatagggc,seqidno.3所示。
需要说明的是,通过s121合成cdna第一链,并利用pcr扩增后,发现1个脱饱和酶基因片段。为了对该基因片段进行分析,我们利用race技术(s122步骤)得到了它的全长序列,这个基因命名为sexides11,它的开放阅读框1059bp,编码353个氨基酸,预测成熟蛋白分子量为40.53kda,等电点为8.95;序列比对和进化分析结果表明,sexides11的氨基酸序列如seqidno.4所示:
mapnisedvngvlfesdaatpdlalarppvqkadnkpkqlvwrniilfaylhlaalyggylflfsakwqtdifayilyvisglgitagahrlwahksykakwplkviliifntvafqdaamdwardhrmhhkysetdadphnatrgfffshigwllvrkhpdlkekgkgldmsdlladpllrfqkkyylvlmplacfvmptmipvylwgetwtnaffvaamfryafilnvtwlvnsaahkwgdkpydksikpsenmsvamfalgegfhnyhhtfpwdyktaelgnnklnfatafinffakigwaydmktvsddivknrvkrtgdgshhlwgwgdknqpkeeieaairinpkdd,sexides11的核苷酸序列如seqidno.5所示:
atggctccaaatatatcggaagacgtgaatggagtgctctttgagagcgacgctgctactccagatctggcgcttgccagacctccagtccaaaaagccgacaacaaaccgaagcaactggtgtggaggaatatcatattgtttgcatatcttcacttagcagctctgtacggtggttacttgtttctcttctccgcgaagtggcaaacagatatttttgcctacatcttgtatgtgatatcgggactgggaatcacggctggagcacatcgcttgtgggcccacaagtcttacaaagctaaatggccactgaaggtgatcctgatcattttcaacactgtcgcctttcaggatgcggctatggactgggcacgcgaccaccgcatgcatcacaagtactcggagactgacgctgatcctcacaacgcgacccgtggcttcttcttctcccacatcggctggctgctcgtcaggaagcatccagacctcaaggaaaagggcaaaggacttgacatgagcgacttgcttgctgatcctctacttagattccagaaaaagtactatctggtcctgatgcctctagcatgcttcgttatgcctacaatgattcctgtgtacctgtggggtgaaacctggaccaatgctttcttcgtggctgccatgttccgttacgcattcatcctgaacgtgacgtggctcgtcaactccgctgcccacaagtggggagacaaaccttacgacaaaagcatcaagccctctgagaacatgtcggtagcgatgttcgcgctcggtgaaggtttccataactaccaccacaccttcccctgggactataaaactgctgaactcggtaacaacaaactcaacttcgccacagccttcataaatttctttgccaaaatcggctgggcctacgacatgaaaactgtgtctgatgacattgtcaagaacagggtgaagcgcactggcgatggctcgcatcatctgtggggctggggtgacaagaaccaacccaaagaagaaattgaggctgctattaggattaatcctaaggatgatta,与报道的鳞翅目昆虫脱饱和酶具有较高的同源性。
s2,连接反应,获得重组质粒peasy-sexides,并进行克隆测序分析
s21,分别将peasy-t3载体和s121的甜菜夜蛾cdna第一链经sali和hindiii双酶切,分别得到peasy-t3酶切产物(双酶切后的长序列片段)和甜菜夜蛾cdna酶切产物(含有sexides11基因的一段序列),将peasy-t3酶切产物和甜菜夜蛾cdna酶切产物连接,得到重组质粒peasy-sexides。
采用北京全式金生物技术有限公司的peasy-t3载体进行快速连接反应依载体自带的说明书加样,总体系为5μl,混匀后置常温连接反应10-15min。
s22,克隆测序分析
s221,转化反应;
(1)从-70℃冰箱中取出经适量分装的感受态细胞transt1(50μl)悬液,冰浴解冻,轻弹管壁以混匀细胞。
(2)短暂离心含有重组质粒peasy-sexides的连接反应管14000g,30s,取s21连接反应物重组质粒peasy-sexides(5μl)于解冻后的感受态细胞中,轻弹均匀,冰水溶30min。
(3)取出离心管,于42℃水浴30s(关键步骤,确保水温和时间的正确性)。
(4)冰水溶2min。
(5)离心管中加入500μl(不含amp)的lb液体培养基(此步骤在超净台操作)。
(6)37℃,200rpm/min振摇1.5h-2h,使细菌复苏,并使质粒编码的抗性基因表达(此时可以在37℃培养箱进行平板回温,并准备超净台灭菌)。
(7)1000g,常温(22℃)离心10min,(从此在超净台内操作),倒去绝大部分上清液(吸350μl)
(8)取5μliptg和40μl5μg/ml的x-gal于第(7)步中的离心管中,吸打混匀,然后将混合液涂于平板培养基上,做好标记。
(9)将第(8)步中的平板正放入37℃培养箱中,待1h-2h待菌液完全被培养基吸收后,再倒置培养皿,16hr左右观察蓝、白斑菌落的生长情况。
s222,挑斑
(1)选取中号,边缘圆润的白斑,不要选得太集中,上下左右5个或4个;
(2)靠近酒精灯倒培养基(含0.1%amp)3ml-4ml,倒完培养基倾斜刻度试管(防止灰尘);
(3)用镊子夹住灭过菌的牙签或小白枪头,点一下选取的白斑,垂直仍入试管中,试管盖子不要盖的太紧,倾斜放入摇床中,250rpm、37℃、振摇12hr-16hr。
s223,小量质粒dna的提取
采用omega质粒提取试剂盒提取质粒dna。
s224,序列分析
序列分析通过genedoc、mega(4.0)等软件完成,并通过blast程序(http://www.ncbi.nlm.nih.gov/blast/)与genebank中的序列进行同源性比对,利用http://web.expasy.org网站对蛋白分子量、等电点和疏水性等进行预测。
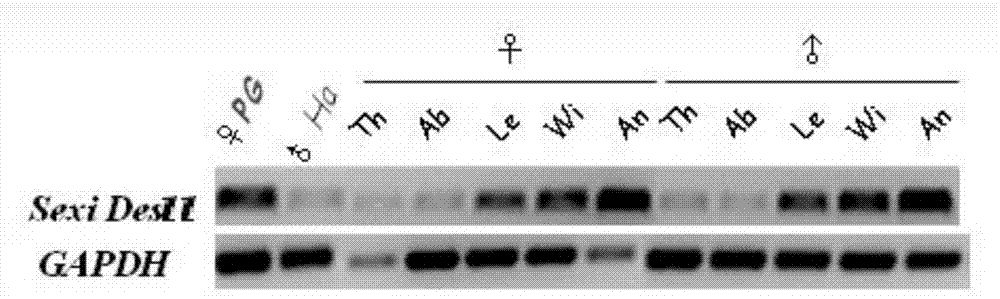
利用tmhmmserverv.2.0、http://web.expasy.org网站和genedoc软件对脱饱和酶进行蛋白结构预测,结果表明,甜菜夜蛾脱饱和酶sexides11基因具有其他鳞翅目昆虫该类基因的保守特点:三个组氨酸盒子结构和四个跨膜域。图1是利用rt-pcr技术研究甜菜夜蛾脱饱和酶基因的组织表达谱,结果表明,这个脱饱和酶基因在性信息素腺体中有表达,且有着较广的组织表达范围。
s3,构建甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides
分别将重组质粒peasy-sexides和pfastbac1载体经sali和hindiii双酶切,分别得到peasy-sexides酶切产物(含有sexides11基因的一段序列)和pfastbac1酶切产物(双酶切后的长序列片段),将peasy-sexides酶切产物和pfastbac1酶切产物连接,得到甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides。
其中,双酶切反应体系如下:
37℃温育3h。
连接反应体系如下:
22℃温育1h
s4,利用甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides和昆虫细胞(high5细胞)构建bac-to-bac真核表达系统,利用bac-to-bac真核表达系统表达甜菜夜蛾脱饱和酶sexides11,然后按照常规方法提取甜菜夜蛾脱饱和酶sexides11。具体步骤如下:
s41,培养昆虫细胞,获得昆虫细胞株
high5细胞27℃贴壁培养,培养基为expressfive昆虫细胞培养基,平均每2天传代,细胞形态呈梭形,得到high5细胞株。
s42,构建携带sexides11基因的重组病毒
s421,制备dh10bac感受态细胞
用非感受态大肠杆菌dh5α制备dh10bac感受态细胞,具体方法如下:
将保存于-70℃,30%甘油中的非感受态大肠杆菌dh5α用灭菌牙签涂划amp-平板,37℃培养12-16h;从平板中用无菌牙签挑取单菌落于含3mllb培养基(amp-)的灭菌试管中,37℃,250r/min振摇过夜;取培养的菌液1ml于100mllb培养基(amp-)的灭菌锥形瓶中,37℃,250r/min振摇培养约2-3h,至od600值达到0.3-0.4时停止培养,立即将锥形瓶置冰浴10-15min;自该步骤起均须无菌操作。将100ml细菌转移到冰预冷的50ml经灭菌的离心管中;4000g,4℃,10min离心,回收细胞;弃去培养液,将管倒置于滤纸上1min左右,使残留的培养液流尽;10ml冰预冷的0.1mol·l-1cacl2重悬菌体(可用枪头轻轻吸打),冰水浴30min;4000g,4℃,10min离心,弃去上清液,倒置于滤纸上1min,使残留的培养液流尽;再加4ml冰预冷的0.1mol·l-1cacl2重悬菌体(可用枪头轻轻吸打);置4℃冰箱12-24h,加入30%体积甘油(约1.7ml甘油于4ml感受态细胞中),50μl左右分装后获得dh10bac感受态细胞,置-70℃保存备用。
s422,转座:将甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides转入dh10bac感受态细胞中进行转座,获得重组病毒bacmiddna的细胞。具体方法如下:
主要参照bac-to-bac表达系统的操作手册进行。取制备好的dh10bac感受态细胞,分别加入1μl的甜菜夜蛾脱饱和酶基因真核表达供体质粒pfastbac-sexides,混匀,冰浴30min,42℃热激60sec后,加入800μl无抗性的lb液体培养基,37℃,200rpm振荡培养4h;取不同稀释倍数的培养物100μl涂布于含kan、tet和gen抗生素以及x-gal和iptg的lb固体培养基平板上;37℃倒置培养36-48h,观察菌落生长情况,挑选大而圆的白色单菌落接种到含kan、tet和gen抗生素的lb液体培养中培养,获得运载重组病毒bacmiddna的细胞平板,重组病毒bacmiddna即为携带sexides11基因的重组病毒,以备下一步实验使用。
s423,从含有重组病毒bacmiddna的细胞中提取重组病毒bacmiddna,并进行pcr鉴定
(1)提取重组病毒bacmiddna
主要参照bac-to-bac表达系统的操作手册进行。简单的操作过程如下:
①在含有重组病毒bacmiddna的细胞平板上挑选大而圆的白色单菌落接种到含kan、tet和gen;抗生素的lb液体培养中,37℃剧烈振摇(250-300rpm)培养至少24h;室温下,12000rpm离心1min收菌,弃上清;加入0.3ml溶液i,充分重悬菌体沉淀物;加入0.3ml溶液ii,轻轻混匀,室温放置5min;加入0.3ml溶液iii,轻轻混匀,冰上放置10min;室温下,13000rpm离心10min;转移上清至另一盛有800μl异丙醇的ep管中,轻轻颠转混匀,-20℃放置10min;室温下,13000rpm离心15min;弃上清,加入70%乙醇500μl,清洗沉淀;除尽上清,在超净台上风干dna5-10min,加40μl的灭菌ddh2o溶解dna,得到重组病毒bacmiddna,-20℃保存备用。该步骤使用的各溶液均按照bac-to-bac表达系统的操作手册制备。
(2)重组病毒bacmiddna的pcr鉴定:
用m13上下游引物m13f和m13r进行pcr,序列如下:m13f:tgtaaaacgacggccagt(seqidno.6),m13r:caggaaacagctatgacc(seqidno.7)。
pcr反应体系如下:
按以下条件进行pcr反应:
94℃3min,1个循环;94℃45sec,58℃45sec,72℃5min,共32个循环;72℃延伸7min;最后4℃保存。
重组病毒bacmiddna的pcr产物电泳结果如图2所示,其中m泳道为dl4000marker;另一泳道为sexides11基因。
s424,利用重组病毒bacmiddna对昆虫细胞进行转染及感染,获得能表达甜菜夜蛾脱饱和酶sexides11的病毒体,提取能表达甜菜夜蛾脱饱和酶sexides11的病毒体,然后利用该病毒体转染再次感染昆虫细胞(病毒体转染再次感染昆虫细胞后可构建出高效表达甜菜夜蛾脱饱和酶sexides11的bac-to-bac真核表达系统)并表达出甜菜夜蛾脱饱和酶sexides11。
主要参照bac-to-bac表达系统的操作手册进行。简单的操作过程如下:
(1)将2×106个处于对数生长期的high5细胞接种于直径为35mm的细胞培养皿中,置27℃培养箱中培养至少1h使其贴壁形成细胞单层;
(2)准备dna-cellfectin混合液:
a.稀释2μgpfastbac-sexides于100μl无血清、无抗生素的expressfive培养基中,充分混匀,得到pfastbac-sexides稀释液;
b.从冰箱中取出cellfectin试剂,颠倒混匀5-10次,然后取出5μlcellfectin试剂稀释于100μl无血清、无抗生素的expressfive培养基中,充分混匀,得到cellfectin稀释液;
c.将pfastbac-sexides稀释液与cellfectin稀释液混合到一起,轻轻混匀,得到dna-cellfectin混合液,室温下温育45min,每隔15min轻轻混匀一次;
(3)吸出已完成细胞贴壁的培养皿中的培养基,向培养皿中加入2ml无血清、无抗生素的expressfive培养基,轻晃培养皿以洗涤细胞;吸出培养基,向培养皿中加入2ml无血清、无抗生素的expressfive培养基,温育30min;吸出培养基;加0.8ml无血清、无抗生素的high5培养基到dna-cellfectin混合液中,混匀,然后加入到吸出了培养基的培养皿中;
(4)置27℃培养箱中温育5-6h;吸出培养皿中液体,加入2ml含抗生素的expressfive培养基;置27℃培养箱中培养,进行病毒转染;病毒转染后6-7天后,吸出培养基,室温下,800g离心5min,转移上清,即为含有病毒粒子的病毒上清液,分装到ep管中,4℃保存备用,此为p1代病毒;
(5)用收集的p1代病毒感染high5细胞进行病毒扩增,6-7天后收集p2代病毒。室温下,800g离心5min,转移上清,即为含有病毒粒子的病毒上清液,分装到ep管中,4℃保存备用;
(6)重复步骤(5)得到p3-p4代病毒,其中,p3-p4代病毒均是携带有sexides11基因的病毒体,其能在high5细胞中存活并表达,且多次细胞传代后,其增殖能力更强,能够在high5细胞中表达出所需的甜菜夜蛾脱饱和酶sexides11,我们只需要按照常规方法提取出甜菜夜蛾脱饱和酶sexides11,即可用于大量的蛋白表达实验。
图3是用不同病毒转染high5细胞2天后显微镜观察图,其中图3a是正常high5细胞,图3b是p1代病毒感染high5细胞,图3c是重组的绿色荧光蛋白基因(gfp)病毒感染的high5细胞,结果显示,用p1代病毒转染high5细胞2天后显微镜观察,发现细胞出现了典型的转染症状,细胞由正常的梭形变成圆形且细胞核扩大,体积变大,说明转染成功(图3b)。同时,重组的绿色荧光蛋白基因(gfp)病毒感染的high5细胞在激发光源下发出绿色荧光(图3c),说明供体质粒和受体质粒的序列都是正确的。
s425,表达产物的sds-page分析
sds-page广泛应用于蛋白质的分离、鉴定和纯化,目前使用最广泛的sds-page为laemmli的tris-甘氨酸系统。
sds聚丙烯酰胺凝胶电泳所用溶液:
①30%丙烯酰胺:将29g丙烯酰胺和1gn,n-亚甲双丙烯酰胺溶于总体积60ml水中,加热至37℃溶解,补加水至体积100ml,缓慢搅拌完全溶解,用0.45μm孔径滤器过滤除菌,棕色瓶保存。
②1.3mol/ltris(ph8.8):18.171gtris溶于100ml去离子水中,用盐酸调ph至8.8。
③1.0mol/ltris(ph6.8):12.114gtris溶于100ml去离子水中,用盐酸调ph至6.8。
④过硫酸铵(10%,m/v):1g过硫酸铵溶解于10ml去离子水中,贮存于4℃,2-3周重新配制。
⑤10×tris-甘氨酸电泳缓冲液:800ml去离子水中溶解30.2gtris碱、188g甘氨酸、10gsds,然后加去离子水至1000ml,调ph至8.3。
⑥1×sds凝胶加样缓冲液:50mmol/ltris·c1(ph6.8)、100mmo1/ldtt、2%sds、0.1%溴酚蓝和10%甘油。
⑦固定液:500ml无水乙醇,100ml冰乙酸,加ddh2o补足至1000ml。
⑧脱色液:250ml95%乙醇,80ml冰醋酸,加ddh2o补足至1000ml。
⑨染色液:0.29g考马斯亮兰r-250溶于250ml脱色液中,用whatman滤纸过滤,以除去颗粒状物质。
取s424(4)中病毒转染4天的high5细胞,5000g/min离心2min,弃上清,50μ1pbs缓冲液洗涤3次。用20μlph7.5,0.1m的pbs将沉淀重新悬浮,加入20μl1×上样缓冲液,混匀,煮沸5-7min,10000g/min离心5min后取上清15μ1与标准蛋白分子量同时电泳点样。浓缩胶浓度为5%,分离胶浓度为12%。用30v恒压电泳30min后80v恒压进行电泳,当溴酚蓝指示剂到达凝胶底部时停止电泳。凝胶于固定液中固定30~60min,考马斯亮蓝g-250染色液进行染色40~60min,然后用脱色液脱色数次至条带清晰背景无色。
利用构建的重组病毒载体上所带有的6×his标签,对high5细胞表达的甜菜夜蛾脱饱和酶蛋白进行westernblot免疫分析。图4是甜菜夜蛾脱饱和酶蛋白的westernblot免疫分析结果,结果显示,sexides11在44kda附近出现条带(图4中箭头所示),与软件预测大小一致。
需要说明的是,当本发明给出数值范围时,应理解,除非本发明另有说明,每个数值范围的两个端点以及两个端点之间任何一个数值均可选用。除非另外定义,本发明中使用的所有技术和科学术语与本技术领域技术人员通常理解的意义相同。尽管已描述了本发明的优选实施例,但本领域内的技术人员一旦得知了基本创造性概念,则可对这些实施例作出另外的变更和修改。所以,所附权利要求意欲解释为包括优选实施例以及落入本发明范围的所有变更和修改。
显然,本领域的技术人员可以对本发明进行各种改动和变型而不脱离本发明的精神和范围。这样,倘若本发明的这些修改和变型属于本发明权利要求及其等同技术的范围之内,则本发明也意图包含这些改动和变型在内。
序列表
<120>一种利用bac-to-bac真核表达系统获得甜菜夜蛾脱饱和酶sexides11的方法
<160>7
<170>siposequencelisting1.0
<210>1
<211>18
<212>dna
<213>人工序列
<400>1
tttttttttttttttttt18
<210>2
<211>23
<212>dna
<213>人工序列
<400>2
aagcagtggtatcaacgcagagt23
<210>3
<211>22
<212>dna
<213>人工序列
<400>3
ctaatacgactcactatagggc22
<210>4
<211>353
<212>prt
<213>甜菜夜蛾
<400>4
metalaproasnilesergluaspvalasnglyvalleuphegluser
151015
aspalaalathrproaspleualaleualaargproprovalglnlys
202530
alaaspasnlysprolysglnleuvaltrpargasnileileleuphe
354045
alatyrleuhisleualaalaleutyrglyglytyrleupheleuphe
505560
seralalystrpglnthraspilephealatyrileleutyrvalile
65707580
serglyleuglyilethralaglyalahisargleutrpalahislys
859095
sertyrlysalalystrpproleulysvalileleuileilepheasn
100105110
thrvalalapheglnaspalaalametasptrpalaargasphisarg
115120125
methishislystyrsergluthraspalaaspprohisasnalathr
130135140
argglyphephepheserhisileglytrpleuleuvalarglyshis
145150155160
proaspleulysglulysglylysglyleuaspmetseraspleuleu
165170175
alaaspproleuleuargpheglnlyslystyrtyrleuvalleumet
180185190
proleualacysphevalmetprothrmetileprovaltyrleutrp
195200205
glygluthrtrpthrasnalaphephevalalaalametpheargtyr
210215220
alapheileleuasnvalthrtrpleuvalasnseralaalahislys
225230235240
trpglyasplysprotyrasplysserilelysprosergluasnmet
245250255
servalalametphealaleuglygluglyphehisasntyrhishis
260265270
thrpheprotrpasptyrlysthralagluleuglyasnasnlysleu
275280285
asnphealathralapheileasnphephealalysileglytrpala
290295300
tyraspmetlysthrvalseraspaspilevallysasnargvallys
305310315320
argthrglyaspglyserhishisleutrpglytrpglyasplysasn
325330335
glnprolysglugluileglualaalaileargileasnprolysasp
340345350
asp
<210>5
<211>1061
<212>dna
<213>甜菜夜蛾
<400>5
atggctccaaatatatcggaagacgtgaatggagtgctctttgagagcgacgctgctact60
ccagatctggcgcttgccagacctccagtccaaaaagccgacaacaaaccgaagcaactg120
gtgtggaggaatatcatattgtttgcatatcttcacttagcagctctgtacggtggttac180
ttgtttctcttctccgcgaagtggcaaacagatatttttgcctacatcttgtatgtgata240
tcgggactgggaatcacggctggagcacatcgcttgtgggcccacaagtcttacaaagct300
aaatggccactgaaggtgatcctgatcattttcaacactgtcgcctttcaggatgcggct360
atggactgggcacgcgaccaccgcatgcatcacaagtactcggagactgacgctgatcct420
cacaacgcgacccgtggcttcttcttctcccacatcggctggctgctcgtcaggaagcat480
ccagacctcaaggaaaagggcaaaggacttgacatgagcgacttgcttgctgatcctcta540
cttagattccagaaaaagtactatctggtcctgatgcctctagcatgcttcgttatgcct600
acaatgattcctgtgtacctgtggggtgaaacctggaccaatgctttcttcgtggctgcc660
atgttccgttacgcattcatcctgaacgtgacgtggctcgtcaactccgctgcccacaag720
tggggagacaaaccttacgacaaaagcatcaagccctctgagaacatgtcggtagcgatg780
ttcgcgctcggtgaaggtttccataactaccaccacaccttcccctgggactataaaact840
gctgaactcggtaacaacaaactcaacttcgccacagccttcataaatttctttgccaaa900
atcggctgggcctacgacatgaaaactgtgtctgatgacattgtcaagaacagggtgaag960
cgcactggcgatggctcgcatcatctgtggggctggggtgacaagaaccaacccaaagaa1020
gaaattgaggctgctattaggattaatcctaaggatgatta1061
<210>6
<211>18
<212>dna
<213>人工序列
<400>6
tgtaaaacgacggccagt18
<210>7
<211>18
<212>dna
<213>人工序列
<400>7
caggaaacagctatgacc18
- 还没有人留言评论。精彩留言会获得点赞!