一种提高动物繁殖力的AMH-INH-GNIH三表达基因疫苗、其制备方法与应用与流程
本发明属于生物技术领域,具体涉及一种提高动物繁殖力的amh-inh-gnih三表达基因疫苗及其制备方法与应用。
背景技术:
卵泡发育是一个周期性的过程,受多种激素的综合调控,即有促进卵泡发育的激素,如促性腺激素、促性腺激素释放激素,也有抑制卵泡发育的激素,如抗苗勒管素(anti-müllerianhormone,amh)、卵泡抑制素(inhibin,inh)和促性腺激素释放抑制激素(gonadotropininhibitinghormone,gnih)等。
amh是参与调节生长和分化的糖蛋白转化生长因子-β(tgf-β)超家族成员,对卵泡发育有抑制作用。敲除amh能够解除其对原始卵泡的募集作用,使有腔卵泡数量增加。amh能削弱生长因子kit配体(kl)、碱性成纤维细胞生长因子(bfgf)和角化细胞生长因子(kgf)对原始卵泡发育的刺激作用,提示amh能减少原始卵泡向初级卵泡的转变,抑制原始卵泡的基础性和刺激性发育。对山羊卵巢进行免疫组化分析,发现amh定位于卵母细胞和颗粒细胞(原始卵泡到有腔卵泡阶段)。与阴性对照相比,阳性对照(在培养基中培养或加入kit配体)的原始卵泡百分率降低,生长卵泡的百分率升高,原始卵泡被激活。但是,在培养基中添加amh表现出与阴性对照相似的原始卵泡和生长卵泡百分比。这些结果表明,amh阻碍原始卵泡的启动。
inh由性腺分泌,是转化生长因子-β(tgf-β)家族成员,为分子量31~34kd的异二聚体糖蛋白,通过α亚基(18kd)密切相关的β亚基两个中的一个(βa和βb,大约14kd)组成一种异二聚体糖蛋白激素。亚基α与βa或βb亚基通过二硫键连接,分别形成抑制素a和抑制素b。inh是下丘脑-垂体-性腺轴调控系统中重要的激素之一,通过负反馈调控机体fsh的合成和分泌,进而调控卵泡发育。研究表明,被动或主动免疫inh,能够促进卵泡发育和排卵,提高产仔数,甚至提高胚胎质量,从而提高动物繁殖力。inh免疫动物能够促进卵泡发育,且有剂量依赖关系。han等(2008)分别用10、50和100μg的pcis质粒以20天的间隔免疫大鼠3次,并用50μgpcdna3.1和100μl0.85%盐水作对照,发现在第2次和第3次免疫后,试验组的平均成熟卵泡数比对照组多3.6和4.9个(p<0.05),平均产仔数和胎盘数也显著提高。特别是高剂量组(100μgpcis)的成熟卵泡数显著高于其他试验组,能显著增加成熟卵泡数,但对卵巢大小和重量无影响。wang等(2012)分别用10、50、100μg/100μl的pcisi质粒免疫小鼠,并用100μgpcmv-s和100μl生理盐水作对照,发现免疫组小鼠的血浆fsh、雌二醇浓度均高于对照组(p<0.05),特别在高剂量组的效果最好(p<0.05)。与对照组相比,所有免疫组的卵巢重量(p<0.05)、长度和宽度(p>0.05)均有变化,特别是高剂量组的成熟卵泡数高于其他各组(p<0.05),产仔数增加。mao等(2016)将120只鸡分为4组,用0、25、75或125μg的pcisi肌肉注射母鸡,20天后进行加强免疫。结果表明,各免疫组的优势卵泡和大白卵泡数均高于对照组(p<0.05)。特别是高剂量组,小黄卵泡数增加(p<0.05),产蛋性能显著提高。
gnih是日本学者于2000年最先从鹌鹑脑内成功分离、纯化得到的rf(精氨酸-苯丙氨酸)酰胺肽(rfrp),生物学作用与gnrh相反,主要抑制垂体前叶分泌fsh(促卵泡素)和lh(促黄体素)。随后,从哺乳动物脑中鉴定的gnih直系同源物命名为rfamide(rfrp)相关肽,包含:rfrp-1,rfrp-2,rfrp-3。其中,rfrp-3是生殖的主要调节因子,类似于体内抑制素的生理功能。gnih及其类似物不仅能通过gpr147作用于下丘脑的垂体和gnrh神经元,而且还能抑制促性腺激素的合成和释放以及性腺的发育和维持。在哺乳动物生殖的不同发育阶段,包括初情期前、初情期、发情周期、妊娠、泌乳、绝经和卵巢疾病中,rfrps已被证实是卵巢发育的关键介质、gnrh释放的潜在抑制调节因子,可能通过gnrh的上游调节因子间接发挥其对卵泡发育的影响,或直接通过gnrh神经元的子集发挥作用。除了调节促性腺激素分泌外,gnih通过改变脑中神经类固醇生物合成进一步调节生殖行为。gnih能抑制鸡和鹌鹑脑垂体释放lh和fsh。静脉给予rfrp-3可降低性腺切除雄性大鼠外周血促性腺激素水平,抑制成年小鼠睾丸类固醇激素生成和精子发生,绵羊lh脉冲幅度,抑制lh和fsh分泌。此外,也有研究表明,gnih抑制鸡的卵泡发育和类固醇生成。以上研究提示,gnih可能直接或间接抑制卵泡发育和排卵。
技术实现要素:
本发明的目的,是提供一种具有提高动物繁殖力作用的amh-inh-gnih三表达基因疫苗的工程菌。本发明的另一个目的是提供一种非抗性筛选的抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素的共表达质粒;该质粒可作为dna疫苗免疫动物,提高动物的繁殖能力,克服了一些技术难题。
为实现上述目的,本发明采用了以下技术方案:
一种具有提高动物繁殖力作用的amh-inh-gnih三表达基因疫苗的工程菌,该工程菌于2018年8月15日保藏在中国典型培养物保藏中心(地址:中国武汉),分类命名为猪霍乱沙门氏菌,保藏编号:cctccno:m2018544。
由上述工程菌株制备的amh-inh-gnih三表达基因疫苗工程菌,通过sds碱裂解法抽提即可得到amh-inh-gnih三表达基因质粒(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd质粒)。
一种提高动物繁殖力的amh-inh-gnih三表达基因疫苗,其特征在于,其依次包括tpa-samh基因、tpa-sinh基因和tpa-srfrp基因。
如上所述的amh-inh-gnih三表达基因疫苗,所述tpa-sinh的基因序列如seqidno.1所示,所述tpa-srfrp的基因序列如seqidno.2所示,所述tpa-samh的基因序列如seqidno.3所示。
如上所述的amh-inh-gnih三表达基因疫苗,所述tpa-samh基因和tpa-sinh基因之间均连接有2a肽,所述tpa-sinh基因和tpa-srfrp基因之间连接有2a'肽,所述2a肽的基因序列如seqidno.4所示;2a'肽的基因序列如seqidno.5所示。
一种如上所述提高动物繁殖力的amh-inh-gnih三表达基因疫苗的制备方法,其包括如下步骤:
s1、构建双表达pvax-tpa-samh-2a-tpa-srfrp-asd和pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒;
s2、将步骤s1中获得的质粒分别进行hindiii和bamhi酶切,之后连接,获得pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd三表达质粒。
如上所的方法,所述pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒的构建采用如下方法:
s1101、以pvax-tpa-sinh-asd为模板扩增tpa-sinh基因片段;所述tpa-sinhpcr的基因序列如seqidno.1所示,
以pvax-tpa-srfrp-asd为模板扩增tpa-srfrp基因片段;所述tpa-srfrp基因片段的序列如seqidno.2所示;
s1102、将pvax-asd和tpa-srfrfpcr产物酶切,进行ecorⅰ和xhoi酶切,连接,获得质粒pvax-tpa-srfrp-asd;
s1103、将pvax-tpa-srfrp-asd和tpa-sinhpcr产物酶切,进行bamhi和ecorⅰ酶切,连接,获得质粒pvax-tpa-sinh-tpa-srfrp-asd;
s1104、将pvax-tpa-sinh-tpa-srfrp-asd和puc57-2a'-2a'质粒酶切,进行ecorⅰ酶切,连接,获得质粒pvax-tpa-sinh-2a'-tpa-srfrp-asd;其中,所述2a'肽的基因序列如seqidno.5所示,
如上所的方法,所述pvax-tpa-samh-2a-tpa-srfrp-asd质粒的构建采用如下方法:
s1201、将pvax-tpa-srfrp-asd和tpa-samhpcr产物酶切,进行hindiii和kpni酶切,连接,获得质粒pvax-tpa-samh-tpa-srfrp-asd;所述tpa-samhpcr产物的基因序列如seqidno.3所示;
s1202、将pvax-tpa-samh-tpa-srfrp-asd和puc57-2a'-2a质粒酶切,进行kpni和bamhi酶切,连接,获得质粒pvax-tpa-samh-2a-tpa-srfrp-asd;所述2a肽的基因序列如seqidno.4所示;
s1203、将pvax-tpa-samh-2a-tpa-srfrp-asd和pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒酶切,进行hindiii和bamhi酶切,连接,获得质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd。
如上所述的提高动物繁殖力的amh-inh-gnih三表达基因疫苗或上述所述的制备方法制备的疫苗在制备提高动物繁殖力药品中的应用。
将含有提高动物繁殖力的amh-inh-gnih三表达基因疫苗的工程菌,(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd)直接免疫动物或与dna疫苗佐剂混合后免疫动物,能够提高动物繁殖力。
本发明的有益效果在于:
本发明提供的一种非抗性筛选的抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素的共表达质粒,具有如下优点:
1、抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素的三表达非抗性dna质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd,能够表达免疫原性较强的抗穆勒氏管激素(amh)、抑制素(inh)和促性腺激素释放抑制激素(rfrp)三种蛋白,刺激小鼠产生较高的抗体水平。
2、含有提高动物繁殖力的amh-inh-gnih三表达基因疫苗的工程菌c500(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd)通过灌胃免疫小鼠后,产生抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素抗体,中和内源性激素,减弱这三种激素对促性腺激素的抑制作用,进而促进小鼠繁殖,其产仔数(15.9±1.90)显著高于pbs对照组(13.6±1.72)和空质粒对照组(13.7±2.30),也高于amh单表达组(14.35±2.43)、inh单表达组(14.94±2.19)、rfrp单表达组(14.26±1.37)、双表达组(pvax-tpa-samh-2a-tpa-srfrp-asd,15.50±3.14)和双表达组(pvax-tpa-sinh-2a'-tpa-srfrp-asd,15.44±2.1),促繁殖效果明显。
3、含抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素的三表达非抗性dna质粒的工程菌c500(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd),可以直接免疫动物,通过喷鼻、口服、拌饲等方式,经粘膜免疫产生抗体。由于不含抗性基因,不须引入外源抗生素进行筛选,所以不产生抗生素残留。相对于其它基因疫苗需要质粒提取、纯化,生产成本较高,肌肉注射麻烦,并使动物产生应激反应,该疫苗生产成本低,使用方便,而且无抗性和注射应激反应。
附图说明
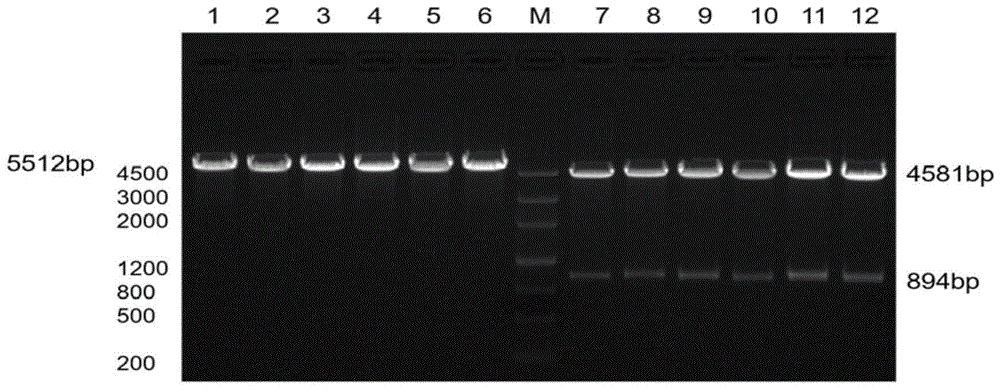
图1为pvax-tpa-sinh-2a'-tpa-srfrp-asd和pvax-tpa-samh-2a-tpa-srfrp-asd质粒酶切图;其中,m:markeriii;泳道1-6:pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒的酶切产物;泳道7-12:pvax-tpa-samh-2a-tpa-srfrp-asd质粒的酶切产物。
图2为pvax-samh-2a-sinh-2a'-srfrp-asd质粒酶切后电泳图;其中,m:markeriii;泳道1:hindiii/bamhi;泳道2:hindiii/ecori;泳道3:hindiii/xhoi;泳道4:质粒;m:1kbdnaladdermarker。
图3为pvax-samh-2a-sinh-2a'-srfrp-asd质粒图谱。
图4为pvax-samh-2a-sinh-2a'-srfrp-asdc500的pcr鉴定,m:markeriii;泳道1-2:inva阴性;泳道3-4:crp阴性;泳道5-6:asd阴性;泳道7-8:amh-inh阴性;泳道9-10:inh-gnih阴性;泳道11-12:inva;泳道13-14:crp;泳道15-16:asd;泳道17-18:amh-inh片段;泳道19-20:inh-gnih片段。
图5为pvax-samh-2a-sinh-2a'-srfrp-asd质粒纯化后酶切图谱;其中,m:markeriii;泳道1-4:hindiii/xhoi。
图6为pvax-samh-2a-sinh-2a'-srfrp-asd转染hela细胞转录水平图谱;其中,m:markeriii;泳道1:amh-inh阴性;泳道2:inh-rfrp阴性;泳道3:amh-inh无处理;泳道4:inh-rfrp无处理;泳道5:amh-inh转染空载;泳道6:inh-rfrp转染空载;泳道7:amh-inh转染三表达;泳道8:inh-rfrp转染三表达。
图7为不同疫苗免疫小鼠后抗amh抗体水平。
图8为不同疫苗免疫小鼠后抗inh抗体水平。
图9为不同疫苗免疫小鼠后抗rfrp抗体水平。
具体实施方式
现有的研究表明,gnih可能直接或间接抑制卵泡发育和排卵。而本发明人经过大量实验用gnih基因疫苗免疫绵羊和小鼠,发现均可刺激卵泡发育和排卵,并提高产仔数与产羔数。inh是下丘脑-垂体-性腺轴调控系统中重要的激素之一,通过反馈作用抑制机体fsh的合成和分泌,对卵泡发育具有调控作用。gnih在hpg轴的最上游水平通过抑制gnrh神经元的活性来降低促性腺激素(lh和fsh)和性腺性类固醇的循环水平。amh能降低原始卵泡向初级卵泡的转变,抑制原始卵泡的基础和刺激发育(nilssonetal.,2007),敲除amh的雌鼠卵巢会显示出原始卵泡存量耗尽(durlingeretal.,1999)。amh、inh和gnih对卵泡发育的抑制作用机制不同,前者直接抑制小卵泡发育,后二者通过抑制垂体fsh的分泌而抑制大卵泡发育;从上可看出如果三者结合使用会抑制卵泡发育,降低生殖能力,但本发明的发明人研究发现这三种抑制卵泡发育的激素中,如果仅免疫中和其中一种或两种激素,余下的两种或一种激素的抑制作用加强。因此,同时中和三种内源性激素,可以解除上述三种激素的抑制作用,进而提高促卵泡发育效果,认为inh,gnih和amh三者之间可能存在代偿作用,因而推测同时中和内源性inh、gnih和amh,对提高动物繁殖力可能有更好的效果。当需要构建表达多个基因的质粒时,可使用双顺反子或多顺反子表达载体,在构建多表达基因的各种策略中,核糖体内部进入位点(ires)被广泛使用。然而,由于ires体积较大,通常超过500个核苷酸,且前后基因间表达水平的较大差异(下游基因的表达量约为上游的10%-50%),需要一种新的策略来替代ires。本发明将大量研究发现采用自切割的2a肽取代ires,因为它体积小(18-22个氨基酸),在2a肽的上游和下游的基因之间有很高的切割效率(上下游几乎等摩尔表达)。因此,为了使多表达基因能够高效表达,本研究采用2a肽为连接子,优化了2a肽的基因序列,构建质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd。其中,经大量实验确定能有效表达所用的各基因的序列如下:tpa-sinh确定所用的基因序列如seqidno.1所示,tpa-srfrp确定所用的基因序列如seqidno.2所示,tpa-samh所用的基因序列如seqidno.3所示;2a肽所用的基因序列如seqidno.4所示;2a'肽所用的基因序列如seqidno.5所示。
以下实施例用于进一步说明本发明,但不应理解为对本发明的限制。在不背离本发明精神和实质的前提下,对本发明所作的修饰或者替换,均属于本发明的范畴。
若未特别指明,实施例中所用的技术手段为本领域技术人员所熟知的常规手段。所述试剂或原料,如未特别说明,均来源于商业渠道。
实施例1真核三表达抗穆勒氏管激素、抑制素、促性腺激素释放抑制激素质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的构建
1、构建双表达pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒;
1)以pvax-tpa-sinh-asd为模板(华中农业大学构建,具体操作见附录1)扩增tpa-sinh基因片段;其中所用的引物如表1中所示,扩增tpa-sinh基因片段如seqidno.1所示,以pvax-tpa-srfrp-asd为模板(华中农业大学构建,具体操作见附录2)扩增tpa-srfrp基因片段;其中所用的引物如表1中所示,扩增tpa-srfrp基因片段如seqidno.2所示,
2)将pvax-asd和tpa-srfrppcr产物酶切,进行ecorⅰ和xhoi酶切,连接,获得质粒pvax-tpa-srfrp-asd;
3)将pvax-tpa-srfrp-asd和tpa-sinhpcr产物酶切,进行bamhi和ecorⅰ酶切,连接,获得质粒pvax-tpa-sinh-tpa-srfrp-asd;
4)将pvax-tpa-sinh-tpa-srfrp-asd和puc57-2a'-2a(将目的片段连接至puc57载体,由上海生工合成)质粒酶切,进行ecorⅰ酶切,连接,获得质粒pvax-tpa-sinh-2a'-tpa-srfrp-asd;puc57-2a'-2a中2a'的基因序列如seqidno.5所示;
表1.tpa-sinh和tpa-srfrp引物序列如下:
2、构建pvax-tpa-samh-2a-tpa-srfrp-asd质粒;
以pvax-tpa-samh-asd为模板(华中农业大学构建,具体操作见附录3)扩增tpa-samh基因片段;其中所用的引物如表2中所示,扩增tpa-samh基因片段如seqidno.3所示,
5)将pvax-tpa-srfrp-asd和tpa-samhpcr产物酶切,进行hindiii和kpni酶切,连接,获得质粒pvax-tpa-samh-tpa-srfrp-asd;
6)将pvax-tpa-samh-tpa-srfrp-asd和puc57-2a'-2a质粒(将目的片段连接至puc57载体,上海生工合成)酶切,进行kpni和bamhi酶切,连接,获得质粒pvax-tpa-samh-2a-tpa-srfrp-asd;puc57-2a'-2a中2a的基因序列如seqidno.4所示;
7)将pvax-tpa-samh-2a-tpa-srfrp-asd和pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒,进行hindiii和bamhi酶切,连接,获得质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd;
表2.tpa-samh引物序列如下:
3、将pvax-tpa-sinh-2a'-tpa-srfrp-asd和pvax-tpa-samh-2a-tpa-srfrp-asd质粒分别进行hindiii和bamhi酶切,pvax-tpa-samh-2a-tpa-srfrp-asd质粒酶切获得片段tpa-samh-2a(894bp),(pvax-tpa-sinh-2a'-tpa-srfrp-asd是5512bp,pvax-tpa-samh-2a-tpa-srfrp-asd是4581bp),其中,反应体系总体积20μl,其中含:
37℃水浴锅反应1h;
最后,将酶切产物用1%琼脂糖凝胶电泳,以markeriii作为分子量标准,观察电泳结果,结果如图1所示,其中,m:markeriii;泳道1-6:pvax-tpa-sinh-2a'-tpa-srfrp-asd质粒的酶切产物;泳道7-12:pvax-tpa-samh-2a-tpa-srfrp-asd质粒的酶切产物。
3)连接:用胶回收试剂盒将酶切产物进行回收(参照omegagelextractionkit使用说明书)。回收后的产物,用takara连接酶,连接目的片段和载体,连接体系为总体积10μl:
16℃过夜。连接产物pvax-tpa-samh-2a-tpa-srfrp-asd,直接用于转化或者-20℃保存备用。
3)重组质粒pvax-tpa-samh-2a-tpa-srfrp-asd转化感受态细菌:用热休克法将连接产物转化感受态细菌χ6097,方法如下:
(1)取100μl冰浴上融化的感受态细胞,加入10μl重组dna(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd),轻轻混匀,在冰浴中放置30min。
(2)42℃水浴中热90sec,然后快速将管转移到冰浴中2min,该过程不要摇动离心管。
(3)向每个离心管中加入900μl无菌的lb培养基(不含抗生素),混匀后置于37℃,220r/min培养45min,使细菌复苏。
(4)1000rpm,5min离心,吸弃上清液,留适量上清液。
(5)将沉淀轻轻吹起,混匀,转移至lb固体培养基(不含抗生素),将细胞均匀涂开。将平板置于37℃恒温箱,30min后倒置平板,37℃培养过夜。
4)pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd阳性克隆的筛选、鉴定与测序:从上步的平板中挑取阳性克隆(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd),接种于lb液体培养基培养,用质粒小量试剂盒抽提质粒,用限制性内切酶hindiii/bamhi、hindiii/ecori和hindiii/xhoi酶切,37℃反应1h,1.0%琼脂糖凝胶电泳观察酶切结果。结果如图2所示,其中,m:markeriii;泳道1:hindiii/bamhi双酶切;泳道2:hindiii/ecori双酶切;泳道3:hindiii/xhoi双酶切;泳道4:质粒;m:1kbdnaladdermarker,表明电泳条带与目的条带(其中,894bp为tpa-samh-2a、1755bp为tpa-samh-2a-tpa-sinh、2683bp为tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp)大小相符,测序结果正确,获得真核表达质粒载体pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd(如图3所示)。
实施例2:真核三表达抗穆勒氏管激素抑制素促性腺激素释放抑制激素质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asdc500感受态工程菌的制备
1.感受态细胞的制备
用无菌接种环挑取冻存的菌种猪霍乱沙门氏菌c500于含有dap(50μg/ml)的lb琼脂平板上划线,同时在不加dap的lb琼脂平板上划线做对照,37℃培养过夜。次日挑取生长良好的单个菌落于5ml含有dap(50μg/ml)的lb液体培养基中,37℃振摇培养过夜。取3ml活化后的培养物接种到300ml含有dap(50μg/ml)的lb液体培养基中,37℃振摇培养2.5-3h,使od600值达到0.5左右。在无菌条件下将细菌培养物倒入预冷的无菌离心瓶,冰浴30min,4℃,5000rpm/min离心10min,弃尽上清,再用10ml冰预冷的10%甘油溶液温和悬起细菌沉淀,冰浴10min,4℃,5000rpm/min离心10min,弃上清,重复用10ml冰预冷的10%甘油溶液温和悬起细菌沉淀,冰浴10min,离心10min后,弃上清,最后用1.5ml冰预冷的10%甘油重悬,悬浮的感受态细胞按80μl分装,立刻用于电转化或置于-80℃冰箱中保存备用。
2.质粒电转化c500
取20μl小提的质粒,加入到新鲜的80μl感受态细胞中,混合后放冰上预冷30min。设置电转仪参数:电压1.8kv,时间4ms-6ms。将冷却的混合物加入电转杯中,把杯外面的水擦干净后电击,立刻加入soc培养基于37℃摇床复苏45min。8000rpm/min,离心1min,吸弃上清液,剩余的涂布于预先在培养箱中预热的麦康凯固体平板,37℃培养过夜。
3.pcr鉴定
从麦康凯平板中挑取单菌落于不添加任何抗生素的lb培养基中,采用pcr法鉴定筛选阳性克隆,主要是针对crp、asd、inva、inh-rfrp、amh-inh基因进行检测,inva、crp、asd检验是否为目的菌;amh-inh、inh-rfrp检测目的菌中是否转染目的质粒进去。采用的20μlpcr反应体系中含上、下游引物各1μl(引物序列见表3),2×taqpcrmix10μl,模板2μl,ddh2o6μl。反应程序为:94℃预变性4min,然后94℃变性40sec,58℃退火30sec,72℃延伸50sec,共35个循环,最后72℃延伸10min。结果见图4,其中,m:markeriii;泳道1-2:inva阴性对照;泳道3-4:crp阴性对照;泳道5-6:asd阴性对照;泳道7-8:引物amh-inh阴性对照;泳道9-10:引物inh-rfrp阴性对照;泳道11-12:菌液inva扩增;泳道13-14:菌液crp扩增;泳道15-16:菌液asd扩增;泳道17-18:菌液amh-inh片段扩增;泳道19-20:菌液inh-rfrp片段扩增。
表3
4.碱裂解法提取质粒(1)从麦康凯平板中挑取单菌落于5ml不添加任何抗生素的lb培养基中,37℃220rpm/min震荡培养过夜。
(2)取2ml菌液转入2mlep管中,13000rpm离心lmin,弃上清。
(3)将沉淀重悬于100μl冰预冷的solutionⅰ,涡旋震荡至菌体充分悬浮。
(4)加入200μl新配制的solutionⅱ,立即颠倒混匀,冰浴5min。加入150μl冰预冷的solutioniii,轻轻的上下颠倒混匀后,冰浴5min。
(5)13000rpm离心5min,将上清转移至新的ep管中。
(6)加入等体积的苯酚/氯仿/异戊醇混合液(按体积比为25:24:l),充分混匀。
(7)13000rpm离心5min,小心吸取上层水相,转移至新的ep管中,加入2倍体积无水乙醇,-20℃沉淀30min。
(8)13000rpm离心10min,弃上清,用75%乙醇洗沉淀一次。
(9)13000rpm离心5min,弃上清。
(10)将ep管室温静置几分钟,加入适量的te,重悬沉淀,56℃作用30min。
5.dna产物纯化(参照天根说明书操作)
(1)向吸附柱cb2中加入500μl的平衡液bl,13,000rpm离心1min,倒掉收集管中的废液,将吸附柱cb2重新放回收集管中。
(2)加入所需纯化的质粒,向其中加入5倍体积的结合液pb,充分混匀。
(3)将上一步所得溶液加入一个吸附柱cb2中(吸附柱放入收集管中),室温放置2min,13,000rpm离心1min,倒掉收集管中的废液,将吸附柱cb2放入收集管中。注意:吸附柱容积为800μl,若样品体积大于800μl可分批加入。
(4)向吸附柱cb2中加入600μl漂洗液pw(使用前请先检查是否已加入无水乙醇),静置2-5min再离心,13,000rpm离心1min,倒掉收集管中的废液,将吸附柱cb2放入收集管中。
(5)重复操作步骤4。
(6)将吸附柱cb2放回收集管中,13,000rpm离心2min,尽量除去漂洗液。将吸附柱cb2置于室温放置数min,彻底地晾干,以防止残留的漂洗液影响下一步的实验。
(7)将吸附柱cb2放入一个干净的离心管中,向吸附膜中间位置悬空滴加30-50μl洗脱缓冲液eb,室温放置2min。13,000rpm离心2min收集dna溶液。
6.dna产物纯化后酶切
限制性内切酶hindiii/xhoi酶切,37℃,反应1h,1.0%琼脂糖凝胶电泳,观察酶切结果,结果如图5所示,其中,m:markeriii;泳道1-4:hindiii/xhoi。结果说明菌液中含有目的片段,质粒成功转染c500中。
结果说明制备的质粒包含抗穆勒氏管激素、抑制素、促性腺激素释放抑制激素三表达重组质粒(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd),于2018年8月15日送至中国典型培养物保藏中心保藏,分类命名:猪霍乱沙门氏菌c500/pvax-samh-2a-sinh-2a-srfrp-asd(salmonellaentericac500/pvax-samh-2a-sinh-2a-srfrp-asd),保藏编号:cctccno:m2018544。该菌株通过sds碱裂解法抽提即可得到pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd质粒。
实施例3:真核三表达抗穆勒氏管激素、抑制素、促性腺激素释放抑制激素质粒pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd质粒在体外表达的检测
pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd质粒转染细胞后转录水平的检测:用质粒提取试剂盒抽提质粒,待hela细胞(人子宫颈癌细胞的细胞系)单层长至60%-70%时按lipofectamine3000脂质体转染试剂盒转染hela细胞,48h后用胰酶消化并收集细胞。用trizol提取细胞的mrna,反转录获得cdna,根据抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素引物(见表3)进行扩增。
以反转录得到的cdna为模板,用amh-inh和inh-rfrp的primers(见表3)进行amh-inh和inh-rfrp两个目的片段的扩增。经1%琼脂糖电泳检测可发现1010bp(amh-inh)和1027bp(inh-rfrp)左右的片段,如图6所示,其中,m:markeriii;泳道1:引物amh-inh阴性扩增;泳道2:引物inh-rfrp扩增阴性;泳道3:引物amh-inh扩增无处理组;泳道4:引物inh-rfrp扩增无处理组;泳道5:引物amh-inh扩增转染空载组;泳道6:引物inh-rfrp扩增转染空载组;泳道7:引物amh-inh扩增转染三表达组;泳道8:引物inh-rfrp扩增转染三表达组。
实施例4:抗穆勒氏管激素、抑制素、促性腺激素释放抑制激素的真核三表达非抗性dna质粒在促进动物繁殖中的应用:
本实验室用已经构建好的非抗性筛选疫苗(pvax-tpa-samh-asd、pvax-tpa-sinh-asd、pvax-tpa-srfrp-asd)免疫小鼠,证明其能够提高小鼠的产仔数。以各自的非抗性筛选抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素基因疫苗c500(pvax-tpa-samh-asd、pvax-tpa-sinh-asd、pvax-tpa-srfrp-asd)为阳性对照,比较同时抗抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素的新型三基因疫苗pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的免疫效果,鉴定疫苗的免疫效果,试图推动新型的三表达抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素基因疫苗在生产中的应用。具体操作如下:
1材料和方法
1.1质粒和菌株
质粒pvax-tpa-samh-asd、pvax-tpa-sinh-asd、pvax-tpa-srfrp-asd、pvax-tpa-samh-2a-tpa-srfrp-asd、pvax-tpa-sinh-2a'-tpa-srfrp-asd均由本实验室构建与保存;同时抗抗穆勒氏管激素、抑制素和促性腺激素释放抑制激素菌株猪霍乱沙门氏菌(salmonellaentericasv.choleraesuis)c500(pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd)鉴定后-80℃保存。
1.2实验动物饲养管理
spf级5周龄雌性昆明小鼠购自湖北省疾控中心,预饲1周后,随机分组,进入试验期,饲养于实验室专门的动物房,控制饲养温度在25℃左右,标准饲料和常规饮水饲养。笼养,每笼5只,每周进行一次卫生清洁,每天观察采食饮水情况和健康状况。
1.3试验分组
将预饲1周后的昆明雌鼠随机分成如下6组如表4,20只/组。
表4各组免疫的疫苗种类和剂量
1.4免疫方式
免疫前4h移除小鼠的水和鼠料,先灌胃200μl碳酸氢钠(7.5%),30min后,免疫组小鼠灌胃200μl疫苗(种类详见表3)。2周后按同样方式加强免疫一次,免疫后连续一周观察小鼠的精神和身体状况。
1.5小鼠的称重和血液采集
在免疫前、免疫后1和2周同一时间段对小鼠进行称重并记录体重数据。在免疫当天和免疫后第8周尾静脉采血,将血液收集于有20μl肝素钠抗凝剂/管的1.5mlep管中,3000r/min离心10min,小心吸取上层血浆,置于-20℃保存备用。
1.6小鼠交配后的产仔数、产仔窝重统计
疫苗加强免疫后2周,将所有母鼠分笼(每笼2只),标记,投入健康雄性小鼠,直至雌鼠全部怀孕。记录小鼠产仔数、产仔窝重和仔鼠重。
1.7amh/inh/rfrp抗体的检测
用间接elisa方法检测免疫后小鼠体内amh、inh和rfrp抗体的生成情况,具体步骤如下:
(1)96孔板酶标板每孔包被amh、inh或rfrp抗原50ng/100μl,4℃孵育过夜。
(2)弃反应液,pbst洗涤3次,300μl/孔,每次3min。
(3)加封闭液(即1%bsa溶液)200μl/孔,37℃孵育1h。
(4)弃反应液,pbst洗涤3次,300μl/孔,每次3min。
(5)加稀释待测血浆,100μl/孔,同时设置阴性对照孔、非特异性吸附孔(及pbst代替血浆)以及零调控,37℃孵育90min。
(6)弃反应液,pbst洗涤5次,300μl/孔,每次3min。
(7)加山羊抗鼠igg-hrp(谷歌,1:3000稀释)100μl/空,37℃反应1h。
(8)弃反应液,pbst洗涤5次,300μl/孔,每次3min。
(9)加tmb底物显色液150μl/孔,避光反应15min。
(10)加2mol/lh2so4终止液50μl/孔,终止反应,15min内在450nm波长处测定各孔的od值。
2结果与分析
2.1免疫反应
2.1.1不同疫苗免疫小鼠后抗amh抗体水平
将分别转化质粒pvax-asd、pvax-tpa-samh-asd、pvax-tpa-samh-2a-tpa-srfrp-asd或pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的减毒沙门氏菌c500菌液,按照1010cfu/ml的剂量免疫小鼠,初次免疫8周后,采集血液,检测抗amh抗体,发现实验组均产生了抗amh抗体,且各实验组之间差异不显著(p>0.05,结果如图7所示)。
2.1.2不同疫苗免疫小鼠后抗inh抗体水平
将分别转化质粒pvax-asd、pvax-tpa-sinh-asd、pvax-tpa-sinh-2a'-tpa-srfrp-asd或pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的减毒沙门氏菌c500菌液,按照1010cfu/ml的剂量免疫小鼠,初免免疫8周采集血液检测抗inh抗体,发现实验组均产生了抗inh抗体,且各实验组之间差异不显著(p>0.05,如图8所示)。
2.1.3不同疫苗免疫小鼠后抗rfrp抗体水平
将分别转化质粒pvax-asd、pvax-tpa-srfrp-asd、pvax-tpa-samh-2a-tpa-srfrp-asd、pvax-tpa-sinh-2a'-tpa-srfrp-asd或pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的减毒沙门氏菌c500菌液,按照1010cfu/ml的剂量免疫小鼠,初免免疫8w采集血液检测抗rfrp抗体,发现实验组均产生了抗rfrp抗体,且各实验组之间差异不显著(p>0.05,如图9所示)。
2.2不同疫苗免疫鼠后产仔数及出生重的比较
统计不同疫苗免疫小鼠后产仔数及出生重,发现pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd组产仔数显著高于pbs组、pvax-asd组、pvax-tpa-samh-asd组和pvax-tpa-srfrp-asd组(表5),也高于pvax-tpa-sinh-asd组、pvax-tpa-samh-2a-tpa-srfrp-asd组和pvax-tpa-sinh-2a'-tpa-srfrp-asd组。统计各组的出生重,发现基因免疫母鼠不影响仔鼠的出生重,各组之间差异不显著(p>0.05)。
表5不同疫苗免疫鼠后的产仔数及窝重比较
注:同一列中数据上标有的小写字母完全不相同表示差异显著(p<0.05),反之,表示差异不显著(p>0.05),同一列中数据上标有的大写字母完全不相同表示差异极显著(p<0.01),所有数据为均值±标准差表示。
结果说明本发明构建的pvax-tpa-samh-2a-tpa-sinh-2a'-tpa-srfrp-asd的dna疫苗可有效提高动物的繁殖能力。
附录1载体pvax-tpa-sinh-asd的构建
1.1.1tpa-sinh片段的扩增
以pires-tpa-sinh-tpa-srfrp质粒为模板进行pcr扩增,20μlpcr反应体系中模板1.5μl,tpa-sinh片段上、下游引物各1μl,2×taqpcrmix10μl,ddh2o6.5μl。pcr反应程序为94℃预变性4min,然后94℃变性40s,66℃退火30s,72℃延伸1min,共35个循环,最后72℃延伸10min,4℃10min。tpa-sinh上游引物带有kpni酶切位点,下游引物带有ecori酶切位点,引物序列如下:
1.1.2tpa-sinh片段与pmd19t(simple)连接
电泳结束后在紫外灯下从凝胶上切下tpa-sinh片段的琼脂糖凝胶,并按照takaraminibestagarosegeldnaextractionkitver.4.0试剂盒说明书操作规程进行回收,具体步骤如下:1%琼脂糖凝胶电泳后,在紫外灯下从琼脂糖胶上切取单一的目的dna条带,放入一个干净的1.5ml的ep管中,加入3倍质量体积(100mg=100μl)buffergm,混合均匀后于室温溶解胶块,其间轻弹ep管,使凝胶完全融化。然后将融化的液体转至一吸附柱,12000rpm离心1min,倒掉收集管中的废液,加入500μlbufferwb,12000rpm离心30s,洗涤两次;最后12000rpm离心1min,除去吸附柱内痕量液体。将吸附柱转移至一干净的1.5mlep管中,加入30μlelutionbuffer,于室温放置2min,12000rpm离心1min洗脱dna。电泳检测回收纯度,并测浓度,收集的dna溶液用于下一步实验或-20℃保存备用。
将tpa-sinh片段与pmd19t-simple载体连接,具体步骤如下:根据回收后的dna浓度计算出载体和目的片段的连接体积比,使用solutioni连接pmd19t和tpa-sinh片段,连接体系如下:
混匀,短暂离心,16℃水浴连接过夜。
1.1.3细菌的转化
(1)将感受态细胞dh5α从-80℃中取出冰浴解冻,加入10μl连接产物,轻轻混匀,冰浴30min。
(2)转至42℃热激90sec,然后迅速取出放在冰上2min,该过程不能摇动离心管。
(3)加入400μl不含抗生素的lb液体培养基,混匀后置于37℃,200r/min振荡培养1h。使细菌复苏。
(4)3000r/min离心5min,弃去400μl上清。
(5)轻轻混匀剩余液体,使用涂布器均匀涂于含amp抗生素(50μg/ml)的lb固体培养基上,37℃恒温培养12-14h,观察是否有转化菌落长出。
1.1.4阳性克隆的筛选与鉴定
挑取单菌落于含amp抗生素(50μg/ml)的lb液体培养基中,37℃200r/min振荡培养12h左右,使用天根试剂盒进行质粒小提,pmd19t-tpa-sinh质粒经过kpni和ecori进行双酶切鉴定。酶切体系如下:
混匀,短暂离心,37℃水浴反应过夜。
将10μl酶切产物进行1%琼脂糖凝胶电泳检测,筛选出疑似pmd19t-tpa-sinh的质粒,送到武汉擎科创新生物科技有限公司测序,将序列比对正确的质粒所对应的菌液扩大培养,提取质粒,-20℃保存备用。
1.1.5载体pvax-asd和质粒pmd19t-tpa-sinh的酶切与回收
参照thermo说明书稍作改动,具体步骤如下:用限制性内切酶kpni和ecori双酶切pvax-asd质粒和pmd19t-tpa-sinh质粒,露出两端的粘性末端,酶切体系分别如下:
混匀,短暂离心,37℃水浴反应过夜。
酶切后经1%琼脂糖凝胶电泳分离出目的条带,使用takaraminibestagarosegeldnaextractionkitver.4.0试剂盒回收tpa-sinh和线性pvax-asd片段,回收后电泳检测回收纯度,并测浓度。
1.1.6tpa-sinh片段与载体pvax-asd连接
按照thermot4连接酶说明书操作规范进行,具体步骤如下:根据回收后检测的dna浓度计算出载体和目的片段的连接比,使用t4dnaligase连接pvax-asd和tpa-sinh片段,连接体系如下:
混匀,短暂离心,16℃水浴连接过夜。
1.1.7χ6097感受态细胞的制备(氯化钙法)
用无菌接种环取冻存的菌种χ6097于含有dap(50μg/ml)的lb平板上划线,同时在不加dap的lb平板上划线做对照,37℃培养过夜。次日挑取生长良好的单菌落于5ml含有dap(50μg/ml)的lb液体培养基中37℃振摇培养过夜。取1ml活化后的培养物接种到100ml含有dap(50μg/ml)的lb液体培养基中,37℃振摇培养2.5-3h,使od600值达到0.5左右。在无菌条件将细菌培养物倒入预冷的无菌离心管中,冰浴30min,4℃5000r/min离心10min,弃尽上清。再用10ml冰预冷的0.1mcacl2温和悬起细菌沉淀,冰浴30min,4℃5000r/min离心10min,弃尽上清,最后用冰预冷的0.1mcacl2再次重悬沉淀,加入终浓度为15%的灭菌甘油,混匀分装为100μl/管,直接用于转化或置于-80℃冰箱中保存备用。
1.1.8细菌的转化
(1)将感受态细胞χ6097从-80℃中取出冰浴解冻,加入10μl连接产物,轻轻混匀,冰浴30min。
(2)转至42℃热激90sec,然后迅速取出放在冰上2min,该过程不能摇动离心管。
(3)加入400μl不含抗生素的lb液体培养基,混匀后置于37℃,200r/min振荡培养1h。使细菌复苏。
(4)3000r/min离心5min,弃去400μl上清。
(5)轻轻混匀剩余液体,使用涂布器均匀涂布不含任何外源物质的lb平板,37℃恒温培养18-20h,观察是否有转化菌落长出。
1.1.9阳性克隆的筛选与鉴定
挑取单菌落于不含任何外源物质的lb液体培养基中,37℃200r/min振荡培养12h左右,使用天根试剂盒进行质粒小提,pvax-tpa-sinh-asd质粒经过kpni和ecori进行双酶切鉴定。酶切体系如下:
混匀,短暂离心,37℃水浴反应过夜。
将10μl酶切产物进行1%琼脂糖凝胶电泳检测,筛选出疑似pvax-tpa-sinh-asd的质粒,送到武汉擎科创新生物科技有限公司测序,将序列比对正确的质粒所对应的菌液扩大培养,提取质粒,-20℃保存备用。
附录2载体pvax-tpa-srfrp-asd的构建
1.2.1tpa-srfrp片段的扩增
具体pcr体系和程序如1.1.2。tpa-srfrp上游引物带有kpni酶切位点,下游引物带有ecori酶切位点,引物序列如下:
1.2.2tpa-sinh片段与pmd19t(simple)连接
具体步骤参照1.1.2。
1.2.3细菌的转化
具体步骤参照1.1.3。
1.2.4阳性克隆筛选与鉴定
具体步骤参照1.1.4。
1.2.5载体pvax-asd和质粒pmd19t-tpa-sinh的酶切与回收
具体步骤参照1.1.5。
1.2.6目的基因连接
按照thermot4连接酶说明书操作规范进行,具体步骤如下:根据回收后检测的dna浓度计算出载体和目的片段的链接比,使用t4dnaligase连接pvax-asd和tpa-srfrp片段,连接体系如下:
混匀,短暂离心,16℃水浴连接过夜。
1.2.7细菌的转化
具体过程参照1.1.8。
1.2.8阳性克隆的筛选与鉴定
具体过程参照1.1.9。
附录3载体pvax-tpa-samh-asd的构建
1.3.1tpa-samh片段合成
在ncbi上查找筛选得到的对应的amh表位抗原编码基因的碱基序列,将其插入乙肝表面抗原的5’端,在乙肝表面抗原3’端插入tpa信号肽,并在整个片段的上游和下游分别添加bamhi和ecori两个酶切位点,构成tpa-samh片段,送至生工公司合成。
1.3.2载体pvax-asd和质粒puc-tpa-samh酶切与回收
具体过程参照1.1.5。
1.3.3目的基因连接
具体步骤参照1.1.2。
1.3.4细菌转化
具体步骤参照1.1.8。
1.3.5阳性克隆筛选与鉴定
具体过程参照1.1.9。
序列表
<110>华中农业大学
<120>一种提高动物繁殖力的amh-inh-gnih三表达基因疫苗、其制备方法与应用
<160>25
<170>siposequencelisting1.0
<210>1
<211>867
<212>dna
<213>人工序列(artificialsequence)
<400>1
ggatccgccaccatggatgcaatgaagagagggctctgctgtgtgctgctgctgtgtgga60
gcagtcttcgtttcgcccagcgctagcatggagagcacaacatcaggattcctaggaccc120
ctgctcgtgttacaggcggggtttttcttgttgacaagaatcctcacaataccacagagt180
ctagactcgtggtggacttctctcaattttctagggggagcacccacgtgtcctggccaa240
aattcgcagtccccaacctccaatcactcaccaacctcttgtcctccaatttgtcctggc300
tatcgctggatgtgtctgcggcgttttatcatattcctcttcatcctgctgctatgcctc360
atcttcttgttggttcttctggactaccaaggtatgttgcccgtttgtcctctacttcca420
ggaacatcaactaccagcacgggaccatgcaagacctgcacgattcctgctcaaggaacc480
tctatgtttccctcttgctgctgtacaaaaccttcggacggaaactgcacttgtattccc540
atcccatcatcctgggctttcgcaagattcctatgggagtgggcctcagtccgtttctcc600
tggctcagtttactagtgccatttgttcagtggttcgtagggctttcccccactgtttgg660
ctttcagttatatggatgatgtggtattgggggccaagtctgtacaacatcttgagtccc720
tttttacctctattaccaattttcttttgtctttggcatatgtccaccgcccctctgccc780
tggccttggtcccccgccgcgctgcgcctgctgcagaggcccccggaggaacccgctgtg840
cacgccgactgccacagacaagaattc867
<210>2
<211>864
<212>dna
<213>人工序列(artificialsequence)
<400>2
gaattcgccaccatggatgcaatgaagagagggctctgctgtgtgctgctgctgtgtgga60
gcagtcttcgtttcgcccagcgtcgacatggagagcacaacatcaggattcctaggaccc120
ctgctcgtgttacaggcggggtttttcttgttgacaagaatcctcacaataccacagagt180
ctagactcgtggtggacttctctcaattttctagggggagcacccacgtgtcctggccaa240
aattcgcagtccccaacctccaatcactcaccaacctcttgtcctccaatttgtcctggc300
tatcgctggatgtgtctgcggcgttttatcatattcctcttcatcctgctgctatgcctc360
atcttcttgttggttcttctggactaccaaggtatgttgcccgtttgtcctctacttcca420
ggaacatcaactaccagcacgggaccatgcaagacctgcacgattcctgctcaaggaacc480
tctatgtttccctcttgctgctgtacaaaaccttcggacggaaactgcacttgtattccc540
atcccatcatcctgggctttcgcaagattcctatgggagtgggcctcagtccgtttctcc600
tggctcagtttactagtgccatttgttcagtggttcgtagggctttcccccactgtttgg660
ctttcagttatatggatgatgtggtattgggggccaagtctgtacaacatcttgagtccc720
tttttacctctattaccaattttcttttgtctttggcatatggcgatggcccacctgcct780
ctgagactcggaaaaaatagagaggacagcctctccagatgggtcccaaatctgccccag840
aggtttgtatacatttaactcgag864
<210>3
<211>837
<212>dna
<213>人工序列(artificialsequence)
<400>3
aagcttgccaccatggatgcaatgaagagagggctctgctgtgtgctgctgctgtgtgga60
gcagtcttcgtttcgcccagcatggagagcacaacatcaggattcctaggacccctgctc120
gtgttacaggcggggtttttcttgttgacaagaatcctcacaataccacagagtctagac180
tcgtggtggacttctctcaattttctagggggagcacccacgtgtcctggccaaaattcg240
cagtccccaacctccaatcactcaccaacctcttgtcctccaatttgtcctggctatcgc300
tggatgtgtctgcggcgttttatcatattcctcttcatcctgctgctatgcctcatcttc360
ttgttggttcttctggactaccaaggtatgttgcccgtttgtcctctacttccaggaaca420
tcaactaccagcacgggaccatgcaagacctgcacgattcctgctcaaggaacctctatg480
tttccctcttgctgctgtacaaaaccttcggacggaaactgcacttgtattcccatccca540
tcatcctgggctttcgcaagattcctatgggagtgggcctcagtccgtttctcctggctc600
agtttactagtgccatttgttcagtggttcgtagggctttcccccactgtttggctttca660
gttatatggatgatgtggtattgggggccaagtctgtacaacatcttgagtcccttttta720
cctctattaccaattttcttttgtctttggcatatgagggaagaggtctccaatacctca780
gcctcgcccagggagcaggccacaggcagcgggacactcatctttcagcaaggtacc837
<210>4
<211>75
<212>dna
<213>人工序列(artificialsequence)
<400>4
ggtaccggcagtggagagggcagaggaagtctgctaacatgcggtgacgtcgaggagaat60
cctggcccaggatcc75
<210>5
<211>75
<212>dna
<213>人工序列(artificialsequence)
<400>5
gaattcggcagtggagagggcagaggaagtctgctaacatgcggtgacgtcgaggagaat60
cctggcccagaattc75
<210>6
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>6
cgcggatccgccaccatggatgcaatgaagagagggc37
<210>7
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>7
ccggaattcttgtctgtggcagtcggcg28
<210>8
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>8
ccggaattcgccaccatggatgcaatgaagagagggc37
<210>9
<211>34
<212>dna
<213>人工序列(artificialsequence)
<400>9
ccgctcgagttaaatgtatacaaacctctggggc34
<210>10
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>10
ccgaagcttgccaccatggatgcaatgaagagagggc37
<210>11
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>11
ccgggtaccttgctgaaagatgagtgtcccg31
<210>12
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>12
tacgcgcatacaacaaaagtcgc23
<210>13
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>13
gccattctgacggaattaacggg23
<210>14
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>14
ttgctttccaactgctgagc20
<210>15
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>15
tcctatctgcgtcgtcctac20
<210>16
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>16
caggatacctatagtgctgc20
<210>17
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>17
cgcaccgtcaaaggaaccgt20
<210>18
<211>19
<212>dna
<213>人工序列(artificialsequence)
<400>18
tatgtccaccgcccctctg19
<210>19
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>19
aatgtatacaaacctctggggca23
<210>20
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>20
atgagggaagaggtctccaata22
<210>21
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>21
tcttgtctgtggcagtcggc20
<210>22
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>22
cggtaccccgatggatgcaatgaagaga28
<210>23
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>23
cggcggaattcttaaatgtatactctgtggc31
<210>24
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>24
cggtaccccgatggatgcaatgaagaga28
<210>25
<211>31
<212>dna
<213>人工序列(artificialsequence)
<400>25
gaattcgcggccgcttaaatgtatacaaacc31
- 还没有人留言评论。精彩留言会获得点赞!