抗人CTLA4单克隆抗体及其应用的制作方法
发明领域
本发明属于肿瘤疗法和分子免疫学领域。本发明涉及抗ctla4抗体,药物组合物及其用途。
发明背景
适应性免疫应答的关键环节是t细胞特异性的识别抗原并被激活。完全激活t细胞需要两个信号,第一个信号是t细胞表面的tcr/cd3与抗原递呈细胞表面(apc)的多肽-mhc复合物的结合,这是一种抗原特异性信号。第二个信号,也称为共刺激信号,是t细胞和apc表面的共刺激分子对的作用(lenschow等,(1996)annurevimmunol.14:233)。t细胞在这两个信号作用后,通过plc-pkc、ip3及pi-3k-ras-mapk等信号途径,激活各种转录因子,诱导促进t细胞生长和分化的细胞因子(il-2等)及相应的受体表达。t细胞和apc表面可同时表达多种共刺激分子对及共抑制分子对,它们统称为共信号分子。共刺激分子介导t细胞激活和增殖,共抑制分子则发挥免疫负调节作用,维持机体免疫自稳。已识别特异性抗原的t细胞如果没有第二信号的激活,那么t细胞可能出现失能(anergy)或者克隆丢失(clonaldeletion)。
由apc(例如树突状细胞(dc),激活的单核细胞和b细胞)表达的b7-1(也称为b7,b7.1或cd80)和b7-2(也称为b7.2或cd86)是激活和维持t细胞免疫应答的重要的共刺激分子。在炎性状态下,apc会诱导表达b7-1和b7-2。b7-1/b7-2和t细胞的共刺激分子结合后给初始t细胞(
t细胞上的cd28分子是b7-1/b7-2的配体之一,属于组成型表达的免疫球蛋白家族,在激活后表达上调。当cd28与apc上的配体b7-1/b7-2结合后,共刺激信号通过cd28传导,诱导t细胞增殖并分泌白介素2(il-2)和抗凋亡因子等。t细胞活化需要双信号,cd28与b7-1/b7-2结合提供了关键的第二信号。只有tcr/mhc第一信号并不能诱导t细胞的免疫应答,缺乏cd28提供的第二信号,t细胞将进入无反应状态或免疫耐受,甚至引起细胞凋亡。因此,选择性的激活b7-cd28途径具有控制t细胞命运的巨大治疗潜力。
b7-1/b7-2结合的第二个配体是细胞毒性t淋巴细胞抗原4(ctla-4,也称为cd152),它也属于免疫球蛋白家族,在结构上与cd28同源,包含单个胞外跨膜区的ig结构域,但在静止t细胞上不表达,t细胞活化后表达上调(waterhouse等,(1995)science270:985)。cd28和ctla-4的天然配体都是b7-1和b7-2,ctla-4和b7相互作用的亲和力要比cd28-b7相互作用的亲和力高10倍以上(peach等,(1994)jexpmed.180:2049-2058)。尽管ctla-4和cd28是t细胞上表达的一对同源受体,具有相同的配体,但是它们具有完全相反的功能。ctla-4属于共抑制分子,其与b7-1/b7-2结合后,启动抑制信号,对t细胞激活发挥负调节作用。ctla-4与其他共抑制分子(如pd-1等)不同,胞内结构缺乏经典的免疫受体酪氨酸相关抑制基序(itim)。封闭ctla-4可以解除t细胞的抑制信号,使用ctla4/ig融合蛋白竞争性抑制cd28与b7结合,可发挥强的免疫抑制作用(allison和krummel(1995).science270:932)。ctla-4主要以两种方式抑制免疫应答,即细胞内在和细胞外在的方式。对于内在机制而言,ctla-4募集并激活磷酸酶,从而导致tcr和cd28信号传导途径中的关键蛋白去磷酸化,阻碍胞内激动性信号向下游信号分子转导(chuang等,(2000)immunity.13:313-322;parry等,(2005)molcellbiol.25:9543–9553;marengere等,(1996)science.272:1170-1173);与cd28竞争结合b7配体,阻断cd28-b7提供的共刺激信号(thompson和allison,(1997)immunity7:445–450);形成非配体结合的ctla-4剪接变体(vijayakrishnan等,(2004)immunity20:563-575;araki等(2009)jimmunol.183:5146-5157;chikuma等,(2005)jimmunol.175:177–181);抑制脂筏形成,脂筏有助于t细胞和apc间形成免疫突触,保证各种信号有序转导(rudd等,(2002)science’sstke.128:pe18.);加快t细胞的迁移,抑制tcr启动的终止信号(stopsignal),阻止t细胞和apc之间形成有效的免疫突触(schneider等,(2006)science313:1972–1975;schneider等,(2005)procnatlacadsciusa.102:12861–12866)。对于外在机制,ctla-4通过反式内吞作用选择性地吞噬apc表面的配体b7-1/b7-2,然后将其在表达ctla-4细胞的内吞泡中降解,从而导致cd28-b7形成的共刺激信号受损(qureshi等,(2011)science332:600–603);通过与apc上的b7-1/b7-2分子相互作用,ctla-4诱导吲哚胺2,3-二加氧酶(ido)的表达,该酶催化色氨酸向犬尿氨酸(kynurenine)的转化,导致局部色氨酸缺乏,对t细胞增殖和活化产生抑制作用(grohmann等,(2002)natimmunol.3:1097-1110;fallarino等,(2003)natimmunol.4:1206-1212;munndh等,(1999)jexpmed.189:1363-1372;mellor等,(2004)intimmunol16:1391-1401);ctla-4信号刺激分泌tgf-β和il-10等抑制性细胞因子(chen等,(1998)jexpmed.188:1849-1857;anneo'garra等,(2004)jclininvest.114:1372-1378)。因此,阻断ctla-4可能为治疗癌症和传染病等提供一种新思路。
ctla-4单克隆抗体能够特异性阻断ctla-4和b7-1/b7-2的相互作用,减弱或者封闭ctla-4对t细胞负调节信号的转导,增强机体抗肿瘤效应。体内和体外实验都表明,阻断ctla-4可以增强t细胞免疫应答,消除肿瘤。在结肠癌(saha等,(2010)scandjimmunol.71:70-82;kocak等,(2006)cancerres.66:7276-7284)、纤维肉瘤(leach等,(1996)science271:1734-1736)、前列腺癌(kwon等,(1997)procnatlacadsciusa.94:8099-8103)、乳腺癌(dewan等,(2009)clincancerres.15:5379-5388;demaria等,(2005)clincancerres.11:728-734)、黑色素瘤(curran等,(2010)procnatlacadsciusa.107:4275-4280;sorensen等,(2010)vaccine28:6757-6764)、卵巢癌(yang等,(1997)cancerres.57:4036-4041)和淋巴瘤(fevery等,(2007)leukemia21:1451-1459)等小鼠模型中都证明ctla-4单克隆抗体可以激发机体的免疫应答,增强抗肿瘤效应,有效杀伤肿瘤细胞。ctla-4单克隆抗体和手术(kwon等,(1999)procnatlacadsciusa.96:15074-15079)、放射疗法(dewan等,(2009)clincancerres.15:5379-5388;demaria等,(2005)clincancerres.11:728-734)、化学疗法(mokyr等,(1998)cancerres.58:5301-5304),冷冻消融(waitz等,(2012)cancerres.72:430-439)、射频消融(denbrok等,(2006)brjcancer.95:896-905)等癌症治疗方法联合使用,能进一步减缓或防止大量继发性肿瘤的生长。
ctla-4单克隆抗体进入临床试验的有伊匹单抗(ipilimumab,之前称为mdx-010或bms-734016;商品名yervoytm,百时美施贵宝公司)和曲美木单抗(tremelimumab,cp-675,206,辉瑞公司)。伊匹单抗是全人源化igg1单克隆抗体,能够阻断ctla-4和b7相互作用,解除其对t细胞活化的抑制作用。同时伊匹单抗的fc段与非经典(cd14+cd16++)单核细胞表达的fcγriiia受体结合,利用抗体依赖的细胞介导的细胞毒作用(adcc)杀伤调节性t细胞,增强抗肿瘤效应(romano等,(2015)procnatlacadsciusa.112:6140-6145)。通过临床试验,伊匹单抗已被批准治疗黑色素瘤、晚期肾细胞癌和转移性结直肠癌等(wolchok等,(2010)lancetoncol.11:155-164;hodi等,(2010)nengljmed.363:711-723)。曲美木单抗是人igg2单克隆抗体,在治疗黑色素瘤患者的临床iii期试验中常规化疗比较治疗效果不显著而终止。
伊匹单抗治疗会激活免疫系统,在发挥抗肿瘤效应的同时也会造成机体的病理损伤,即免疫相关的不良事件(iraes)。由伊匹单抗引起的副作用通常包括结肠炎(defelice等,(2015)melanomares.25:321-327)、皮肤毒性(uemura等,(2016)jimmunothercancer.4:55),肝炎(robert等,(2011)nengljmed.364:2517-2526)以及内分泌病变(faje,(2016)pitutary19:82-92)。另外,伊匹单抗的治疗费用比较高。因此,急需开发新的具有更高亲和力,特异性更强的针对ctla-4的低副作用的抗体。
本发明描述了一种具有增强抗肿瘤效应的抗ctla-4单克隆抗体,此抗体具有较低的毒副作用。本发明的抗ctla-4单抗还可以和抗pd-l1、pd-1抗体、抗4-1bb抗体等其他免疫检查点抗体联合使用,增强抗肿瘤作用。
技术实现要素:
本发明提供了一种ctla-4的单克隆抗体,其能够高亲和力的结合人ctla-4,用于抗肿瘤治疗,并且具有较低的毒副作用。特别地,本发明提供的抗体能特异性结合于人的ctla-4蛋白,阻断ctla-4与b7-1和b7-2配体的结合,其至少能阻断约27%,62%,85%,93%或95%的ctla-4和b7结合。本发明提供特异性结合人ctla-4的单克隆抗体在体外能增强t细胞的活化,促进ifn-γ的分泌,其分泌量能提高1.4到2.3倍。
本发明提供的抗ctla-4单克隆抗体具备以下一种或多种特性:
(a)特异性结合人ctla-4
(b)阻断人ctla-4与人b7-1和/或人b7-2的结合
(c)体外增强t细胞活化,促进ifn-γ的分泌
该抗体包含具有seqidnos:1、2和3所示的轻链可变区cdr序列,具有seqidnos:4、5和6所示的重链可变区cdr序列。抗体可以特异性结合人ctla-4。该抗体还可抑制人ctla-4与b7-1或b7-2的结合。
本发明进一步提供了本文所述抗体的抗原结合片段。
本发明还提供了包含所述抗体有效治疗剂量的药物组合物。所述药物组合物可以包含生理上可接受的载体或赋形剂。
在一些实施例中,抗ctla-4抗体特异性结合于ctla-4蛋白上的表位和配体b7-1和/或b7-2结合的表位相似。在一些实施例中,该抗ctla-4抗体可阻断b7-1和/或b7-2与人ctla-4的结合。在一些实施例中,该抗ctla-4抗体和伊匹单抗比较,阻断b7-1和/或b7-2与ctla-4结合的ic50值更低。在一些实施例中,该ctla-4抗体对b7-ctla4结合的阻断ic50是伊匹单抗的10%。
本发明的抗体可以在人混合淋巴细胞反应(mlr)中增强t细胞应答。t细胞应答增强可以检测很多指标,包括但不限于增加抗炎性细胞因子的分泌。增强的t细胞应答可以使用本领域已知的任何方法来检测。在一些实施例中,建立了人混合淋巴细胞实验模型,通过检测激活t细胞分泌的ifn-γ来确定t细胞活性的增加。在一些实施例中,t细胞经本发明的抗ctla-4抗体活化后,刺激生成的ifn-γ的量至少提高了1.4倍到2.3。
附图说明
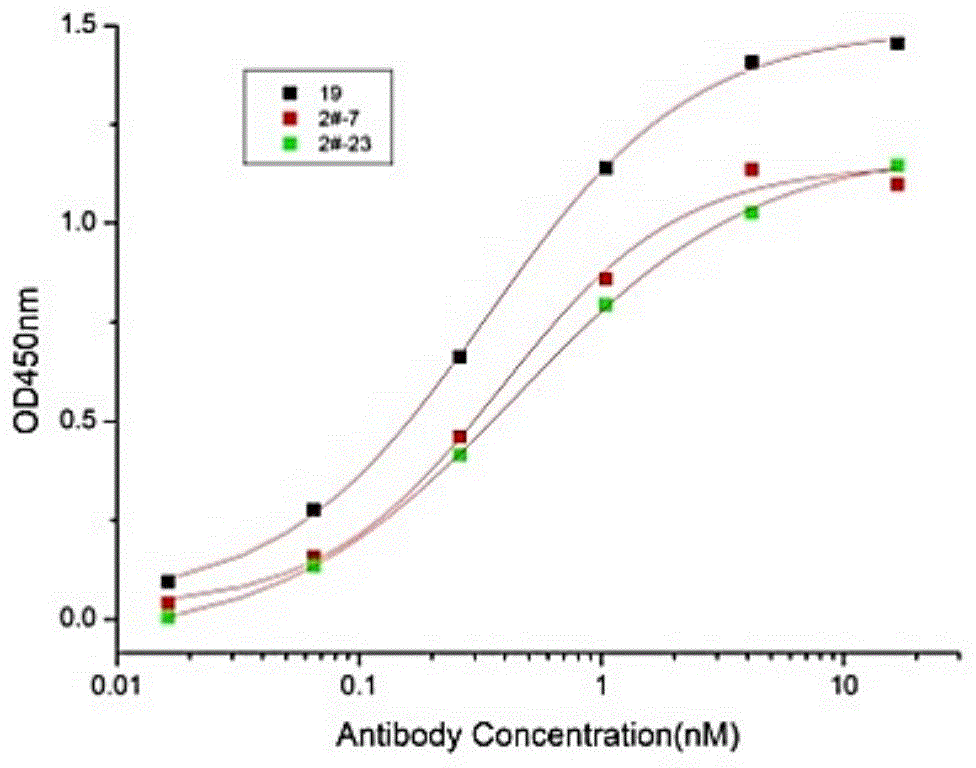
图1显示抗ctla-4抗体能特异性结合人ctla-4重组蛋白。编号为#2-7和#2-23的抗体以剂量依赖性和饱和动力学结合至纯化的人ctla-4重组蛋白上。
图2显示抗ctla-4抗体能阻断人ctla-4和人b7-1蛋白的结合。#2-7和#2-23两种抗体均结合至固定化的ctla4-fc,有效地阻断b7-1和ctla-4的相互作用。在配体竞争elisa实验中,将100ng/ml的ctla-4-fc蛋白包被在elisa板上。洗涤并封闭后,加入如图所示浓度的抗体,以及16ng/ml生物素化的b7-1-fc蛋白。用hrp标记的链霉亲和素检测结合的b7-1的量。结果显示#2-7和#2-23均以剂量依赖性方式阻断ctla-4和b7-1的相互作用,而19号抗体只有在较高浓度才能抑制ctla-4和b7-1的结合。而且#2-7和#2-23号抗体达到50%抑制率的抗体浓度(定义为ic50)分别为0.15nm和0.30nm均小于市售的伊匹单抗(1.5nm)。
图3显示抗ctla-4抗体能够阻断人ctla-4与人b7-2蛋白的结合。#2-7和#2-23两种抗体均结合至固定化的ctla4-fc,有效地阻断b7-2和ctla-4的相互作用。在配体竞争elisa实验中,将100ng/mlctla-4-fc蛋白包被在elisa板上。洗涤并封闭后,加入如图所示浓度的抗体,以及80ng/ml的生物素化的b7-2-fc蛋白。用hrp标记的链霉亲和素检测b7-2的量。结果显示#2-7和#2-23均以剂量依赖性方式阻断ctla-4和b7-2的相互作用,而19号抗体仅在高浓度时才有阻断活性,且阻断作用较弱。而且#2-7ic50为0.25nm小于市售的伊匹单抗(0.75nm)。
图4显示抗ctla-4抗体能够阻断ctla-4介导的免疫负调节,活化t细胞分泌ifn-γ。在混合淋巴细胞实验中,-19,#2-7和#2-23三株抗体都能在所测试的20nm,5nm和1.25nm浓度下,和同型对照抗体相比,显著激活t细胞,提高ifn-γ的分泌量。市售的伊匹单抗未观察到对t细胞的活化作用。
实施例
以下实施例不应被认为是对本发明范围的限制。这些图和表格与以下示例有关,如下所示:
图1:抗ctla-4抗体能特异性结合人ctla-4重组蛋白
图2:抗ctla-4抗体能够阻断ctla-4和配体b7-1的结合
图3:抗ctla-4抗体能够阻断ctla-4和配体b7-2的结合
图4:抗ctla-4抗体能够解除ctla-4介导的免疫负调节,刺激t细胞分泌ifn-γ
定义
“抗体”是指包含通过二硫键互相连接在一起的至少两条重(h)链和两条轻(l)链的糖蛋白,或其抗原结合部分。抗体包括完整抗体及任何可与抗原结合的片段或单链抗体。
“重链”由重链可变区(vh)和重链恒定区组成。
“重链恒定区”由ch1、ch2和ch3三个结构域组成。
“轻链”由轻链可变区(vl)和轻链恒定区(cl)组成。
“轻链恒定区”由cl一个结构域组成。
“重链可变区”和“轻链可变区”可进一步细分为互补决定区(cdr),也被称为高变区,这部分序列对抗原的结合作用最大,特别是重链和轻链的第三个cdr。cdr位于被称为“骨架区”(fr)之间,骨架区序列相对保守。每个vh和vl均由三个cdr和四个骨架区组成。
“恒定区”可以介导抗体与机体组织或因子的结合,该机体组织或因子包括免疫系统的各种细胞(如效应细胞)和经典补体系统的第一成分(c1q)。
“抗原结合片段”是指保留与抗原(如ctla-4)特异性结合的能力的抗体的一个或多个片段。已证明抗体的抗原结合功能可由全长抗体的片段来行使。“抗原结合片段”中所包括的结合片段包括:(1)fab片段,即由vl、vh、cl和ch1结构域组成的单价片段;(2)f(ab’)2片段,即包含在铰链区(重链ch1和ch2功能区之间的区域)通过二硫键连接的两个fab片段的双价片段;(3)fab'-sh,即二硫苏糖醇对抗体(fab')2片段进行切割和分离,生成的片段为c末端带有巯基的fab'片段,可以利用链间的硫醇-二硫键试剂将识别不同靶点的抗体片段重组为一类f(ab')2形式的双特异抗体;(4)由vh和ch1结构域组成的fd片段;(5)由单个抗体结构域vl和vh结构域组成的fv片段;(6)由vh结构域组成的dab片段;(7)分离的互补决定区(cdr)。此外,尽管fv片段的两个结构域vl和vh由单独的基因编码,但利用重组方法可以将二者连接起来,形成一条蛋白质链,其中vl和vh区配对构成单价分子称为单链fv(即scfv),这种单链抗体也包括在术语“抗原结合片段”内。这些抗体片段用本领域技术人员公知的常规技术获得,并用与完整抗体相同的方法对这些片段的实用性进行筛选。
“嵌合抗体”是指其中可变区序列来源于一个物种而恒定区序列来源于另一个物种的抗体(如其中可变区序列来源于小鼠抗体而恒定区序列来源于人抗体的抗体)。
“人源化抗体”是指其中来源于另外一种哺乳动物种系(如小鼠种系)的cdr序列被移植到人构架序列上的抗体,在人构架序列内也可以进行其它的构架区修饰。
“全人源抗体”包括具有如下可变区的抗体,在该可变区中,框架区和cdr区都源自人种系免疫球蛋白序列。而且,如果该抗体含有恒定区,则恒定区也源自人种系免疫球蛋白序列。
本发明的能特异识别ctla-4的单克隆抗体或抗体片段能够用于制备免疫抑制药物,该免疫抑制药物作为抗肿瘤的药物,该肿瘤包括:恶性淋巴瘤、黑色素瘤、肾癌、前列腺癌、卵巢癌、结直肠癌、纤维肉瘤、乳腺癌、肺癌中的任意一种或两种以上。其中肺癌包括小细胞肺癌和非小细胞肺癌。
正常的免疫微环境中,组织感染或损伤时,t细胞可被抗原提呈细胞(antigenpresentationcell,apc)上的抗原激活,诱导增殖,分泌细胞毒素,杀伤被感染的细胞。但是,在血液肿瘤(如恶性淋巴瘤)中,apc细胞受到表达ctla-4的调节性t细胞的调节作用,抗原提呈减弱,形成适宜肿瘤细胞增长的肿瘤微环境。
实施例1.抗人ctla-4的小鼠单克隆抗体的产生
为了产生针对人ctla-4的抗体,通过pcr扩增人ctla-4胞外区的cdna编码序列,c端融合人抗体g1亚型的fc区域(hctla4-hfc,seqidno:53),克隆到表达载体pcdna3.4(invitrogen,货号:17493)上,大量制备该质粒dna,按照invitrogen公司的expi293tmexpressionsystemkit(expi293tm表达系统试剂盒,货号:a14635)所附操作流程,在expi-293细胞中瞬时表达重组的hctla4-hfc蛋白,用hitraprproteinaff(gehealthcare,货号:17-5080-01)纯化hctla4-hfc融合蛋白,电泳检测后定量备用。
为了免疫产生杂交瘤细胞系所必需的小鼠,将60μg人ctla4-fc融合蛋白和100μl完全弗氏佐剂混合,将此混合物通过皮下注射免疫四只6-7周龄的雌性balb/c小鼠。以后每两周通过皮下注射对小鼠进行加强免疫,免疫原为30μg蛋白抗原与不完全弗氏佐剂混合而成。5周后,进行最后一次加强免疫。加强免疫三天后,取血制备血清。然后用pbs以1/1000稀释血清,进行elisa分析,测定识别人ctla-4抗体的效价,选择高效价小鼠进行细胞融合。
在细胞融合实验前三天,通过腹膜内注射向每只小鼠注射50μg重组人ctla4-fc融合蛋白。细胞融合时,使用peg1500(roche,货号78364)将获得的免疫小鼠脾细胞悬液与骨髓瘤细胞(sp2/0)混合。将混合的细胞以每孔100μl加入到提前铺有饲养细胞的96孔板中,并在37℃,5%co2的培养箱中温育24小时后进行hat(sigma,货号:h0262-10vl)选择(littlefield(1964)science145:709-710)。
通过elisa鉴定了分泌抗ctla4抗体的杂交瘤细胞。将96孔板(costar,货号:9018)加入100μl的2μg/mlctla4-fc的溶液,4℃包被过夜。用含0.05%(v/v)tween-20tm(sigma,货号:p1379)的磷酸盐缓冲液(pbst)洗板后,加入300μl的1%(w/v)bsa(roche,货号:738328)封闭液,室温封闭2h。洗板后,加入杂交瘤细胞上清液,每孔100μl,37℃孵育2h。洗板后每孔加入100μl偶联hrp的山羊抗小鼠igg(abcam,货号:ab6789)孵育1h。洗板后,加入100μl/孔的底物溶液tmb(ebioscience,货号:00-4201-56),并将板在室温下孵育5分钟。然后,加入50μl/孔的终止溶液(2nh2so4)以终止反应。然后测量450nm处的吸光度,并记录为光密度(od450nm)。
将筛选的阳性杂交瘤细胞,经3次亚克隆及elisa检测完全阳性后,扩大培养并建株。对抗体的亚型和效价检测后,大量制备单克隆抗体。杂交瘤细胞注射到降植烷(pristane,2,6,10,14-四甲基五癸烷)预致敏的balb/c小鼠腹腔。接种细胞7-10天后待腹水形成收集腹水。室温3000r/min离心10min收集腹水上清。利用proteina柱(gehealthcare)纯化腹水上清中的单克隆抗体,并冻存。
对总共二十种抗-hctla4杂交瘤抗体进行了抗体纯化。将杂交瘤细胞在含有10%胎牛血清,1%青霉素/链霉素,2%l-谷氨酰胺和1%调节nahco3溶液的dulbecco'smodifiedeagle's培养基(gibco,货号:11995065)中培养。然后将所选的杂交瘤细胞适应于无血清培养基中,并使用proteina柱(gehealthcare)从上清液中纯化抗体。用pbs洗涤后,用0.1m甘氨酸ph3.0洗脱结合的抗体,然后用2.0mtris中和ph。ultra-15离心浓缩器(amicon,货号:ufc905008)用于缓冲液交换和抗体浓缩。
实施例2:elisa实验测定鼠抗人ctla-4抗体与ctla-4抗原的结合能力
为了评估纯化的杂交瘤抗体结合人ctla-4的能力,使用elisa来测量结合亲和力。简而言之,将96孔板每孔加入100μl的2μg/ml人ctla-4重组蛋白,4℃包被过夜。用pbst洗涤3次,并用300μl/孔的1%bsa的封闭液在37℃封闭2h。洗板后,每孔加入100μl梯度稀释的纯化抗体,37℃孵育2小时。洗板后,每孔加入100μl偶联hrp的山羊抗小鼠igg孵育1h。洗板后,加入100μl/孔的底物溶液tmb,并将板在室温下孵育5分钟。然后,加入50μl/孔的终止溶液(2nh2so4)以终止反应。测量450nm处的吸光度,并记录为光密度(od450nm)。
实施例3.抗ctla4抗体阻断ctla-4与b7-1/b7-2结合
通过elisa测试抗体阻断ctla-4与其配体b7-1和b7-2结合的能力。将重组人ctla4-fc(acrobiosystems,ct4-h5255)在ph9.6的碳酸盐缓冲溶液中稀释至100ng/ml,96孔板每孔100μl4℃包被过夜。用pbst洗板后,每孔加入300μl的1%(w/v)bsa的封闭液37℃孵育2小时。洗板后,每孔加入50μl生物素化人b7.1-fc(32ng/ml)或生物素化人b7.2-fc(160ng/ml)和50μl梯度稀释的抗ctla-4抗体(3倍系列稀释度,终浓度范围20nm至0.027nm),37℃下孵育1小时。洗板后,每孔加入100μl偶联hrp的链霉亲和素(1:40000),37℃下孵育1小时。洗板后,每孔加入100μl的tmb显色底物,室温下孵育10分钟,然后加入50μl终止溶液(2nh2so4)终止反应。如图2和3所示,抗体-19,#2-7和#2-23均能阻断ctla-4和b7的结合。以ic50作为抑制ctla-4及其配体结合活性的指标时(ic50为达到最大抑制率一半时抗体的浓度),#2-7,#2-23两株抗体的阻断活性明显高于-19号抗体,亦优于伊匹单抗。#2-7抗体可在0.027nm的浓度下阻断ctla4-fc:b7.1和ctla4-fc:b7.2的体外结合,浓度为0.15nm即可达到50%的抑制效果,伊匹单抗阻断结合的ic50(1.5nm)约是该抗体的10倍。
实施例4.抗ctla-4单克隆抗体的细胞功能验证
为了制备由单核细胞诱导生成的树突状细胞(dc),将外周血单个核细胞(pbmc,中国tpcs)以1×106细胞/ml的密度接种在t75细胞培养瓶(greiner)中。将pbmc在37℃培养箱里静置2小时,以保证单核细胞贴壁。细胞贴壁后,用冷的pbs漂洗细胞两次,收集上清中未贴壁的细胞,另用。贴壁的单核细胞在添加gm-csf(500iu/ml,r&dsystems)和il-4(500iu/ml,r&dsystems)的rpmi1640培养基(gibco)中培养5天。加入tnf-α(10ng/ml),il-1β(10ng/ml),il-6(15ng/ml),pge2(1μg/ml)和gm-csf(500iu/ml)和il-4(500iu/ml)等细胞因子,继续培养2天以诱导dc的成熟(jonuleit等,(1997)eurjimmunol.27:3135-3142.)。第8天,收集诱导成熟的dc,计数,并使用台盼蓝染色检测细胞活力。
在开始混合淋巴细胞反应(mlr)实验之前,将dc以1×105个细胞/ml重悬于rpmi1640培养基中,每孔100μl加入到96孔板中。在同一天,制备来自其他供体的t细胞悬液,加入到已铺dc的孔中,每孔1×105个细胞。最后每孔分别加入抗ctla4抗体、同型对照抗体,抗体终浓度为20nm、5nm、1.25nm及0nm。将96孔板在37℃的培养箱中孵育5天后,离心,收上清。使用elisa试剂盒(r&dsystems,货号dy285)检测上清中ifn-γ的分泌量。同型对照抗体用作阴性对照。elisa检测如下:酶标板每孔包被100μl的ifn-γ捕获抗体,在4℃包被过夜。洗板后,每孔加入300μl的1%bsa封闭2小时。洗板后,加入稀释好的细胞上清,室温下孵育2小时。洗板后,每孔加入100μl检测抗体,并在室温下孵育2小时。洗板后,每孔加入100μl链霉亲和素-hrp(1:40),并在室温下孵育20分钟。洗板后,每孔加入100μltmb显色底物并在室温下孵育15分钟,然后用50μl终止溶液(2nh2so4)终止反应。测定结果见图4,结果表明#2-7、#2-23和19号三株抗ctla-4单克隆抗体都能增强t细胞分泌ifn-γ;而伊匹单抗不能消除ctla-4对t细胞的抑制,ifn-γ的分泌量和对照相比没有增加。和同型对照抗体相比,抗体19和#2-23在三个测试浓度下都能促进t细胞分泌ifn-γ,而且-19对t细胞的激活能力更强。#2-7只在20nm对t细胞有激活作用。
实施例5.抗体#2-7的重(vh)和轻(vk)可变区的核苷酸和氨基酸序列
对抗ctla-4单克隆抗体#2-7进行可变区(v区)序列分析。按照trizol试剂的说明书(invitrogen,货号:15596018)从3×106个杂交瘤细胞中提取总rna,将总rna逆转录成cdna。使用smarterrace5’kit(takara,日本)对鼠免疫球蛋白重链和κ轻链可变区片段进行pcr扩增。pcr产物分离后回收纯化目的片段,克隆到pmd-19t载体系统(takara,日本)中,并使用载体特异性引物m13primers对插入片段进行测序。
尽管本发明的内容已经通过上述优选实施例作了详细介绍,但应当认识到上述的描述不应被认为是对本发明的限制。在本领域技术人员阅读了上述内容后,对于本发明的多种修改和替代都将是显而易见的。因此,本发明的保护范围应由所附的权利要求来限定。
序列表
<110>北京华大蛋白质研发中心有限公司
<120>抗人ctla4单克隆抗体及其应用
<130>1
<160>6
<170>siposequencelisting1.0
<210>1
<211>16
<212>prt
<213>musmusculus
<400>1
lysserserglnthrileleuhisserasnglyilethrtyrleuglu
151015
<210>2
<211>7
<212>prt
<213>musmusculus
<400>2
lysvalserasnargpheser
15
<210>3
<211>9
<212>prt
<213>musmusculus
<400>3
pheglnglyserhisvalprophethr
15
<210>4
<211>5
<212>prt
<213>musmusculus
<400>4
thrtyrglymetser
15
<210>5
<211>17
<212>prt
<213>musmusculus
<400>5
thrileserthraspasnsertyrthrtyrtyrproaspasnvallys
151015
gly
<210>6
<211>10
<212>prt
<213>musmusculus
<400>6
hisalatyralaasptyrprovalasptyr
1510
- 还没有人留言评论。精彩留言会获得点赞!