具有人源化免疫球蛋白基因座的经遗传修饰的非人动物的制作方法
具有人源化免疫球蛋白基因座的经遗传修饰的非人动物
[0001]
优先权声明
[0002]
本申请要求2019年2月18日提交的pct/cn2019/075406和2019年9月18日提交的pct/cn2019/106320的优先权。前述的全部内容通过引用并入本文。
技术领域
[0003]
本公开涉及具有人源化重链免疫球蛋白基因座和/或人源化轻链免疫球蛋白基因座的经遗传修饰的动物和细胞。
[0004]
发明背景
[0005]
治疗性抗体是增长最快的治疗性化合物类别之一,其增长速度快于小分子药物的增长。这些治疗性抗体通常是人抗体或人源化抗体。人抗体或人源化抗体可通过啮齿动物抗体(例如,小鼠抗体)的人源化或通过使用噬菌体文库来产生。通过这些方法产生的抗体通常具有次优的结合亲和力和生物物理属性,导致制造困难和较差的药代动力学。特别地,人源化过程可能不利地影响结合亲和力,并将免疫原性表位引入抗体,并且使用噬菌体文库发现的抗体显示免疫球蛋白重链和轻链的有限多样性和非天然配对。经常需要迭代和耗时的实验来改善所述性质。在一些情况下,这些抗体在患者体内也可具有免疫原性,从而导致其效力随着时间的推移而减弱。
[0006]
产生完全人抗体的一种可能的方法是使用经工程化以表达人抗体谱系的转基因动物。转基因动物(诸如具有不同免疫球蛋白基因座的小鼠)的产生已允许在各种研究和开发应用中,例如在药物发现和对各种生物系统的基础研究中使用此类转基因动物。许多早期的转基因动物具有不完整的人抗体谱系,由于较低的v(d)j重组效率,抗体产量低于正常水平,具有可能引入免疫原性表位的内源抗体谱系,以及各种其它问题。需要高效且成本效益好的生产人抗体的方法,并且需要包含人源化免疫球蛋白基因座的非人动物,所述非人动物具有响应抗原以产生人源化抗体的能力。
技术实现要素:
[0007]
本公开涉及具有人源化重链和轻链免疫球蛋白基因座的经遗传修饰的动物和细胞。
[0008]
在一些方面,本公开涉及经遗传修饰的非人动物,其在内源重链免疫球蛋白基因座上包含一个或多个人ighv基因、一个或多个人ighd基因和一个或多个人ighj基因。在一些实施方案中,人ighv基因、人ighd基因和人ighj基因可操作地连接,并且可以进行vdj重排。
[0009]
在一些实施方案中,所述动物包含约或至少150个、151个、152个、153个、154个、155个、156个、157个、158个、159个、160个或161个选自表1的人ighv基因,约或至少20个、21个、22个、23个、24个、25个、26个或27个选自表2的人ighd基因,以及约或至少5个、6个、7个、8个或9个选自表3的人ighj基因。在一些实施方案中,所述动物包含表1中除ighv2-10、ighv3-9和ighv1-8外的全部人ighv基因,表2中的全部人ighd基因,以及表3中的全部人
ighj基因。在一些实施方案中,所述动物包含表1中除ighv5-10-1和ighv3-64d外的全部人ighv基因、表2中的全部人ighd基因和表3中的全部人ighj基因。在一些实施方案中,所述动物包含在人14号染色体上的内源重链免疫球蛋白基因座上的全部人ighv基因、全部人ighd基因和全部人ighj基因。在一些实施方案中,所述动物包含在人细胞(例如,体细胞、培养细胞、非免疫细胞、没有任何v(d)j重排的细胞)的人14号染色体的内源重链免疫球蛋白基因座上的全部人ighv基因、全部人ighd基因和全部人ighj基因。
[0010]
在一些实施方案中,所述动物包含在其内源重链免疫球蛋白基因座中的破坏。
[0011]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源重链免疫球蛋白基因座中的破坏包括表4中的一个或多个小鼠ighv基因、表5中的一个或多个小鼠ighd基因和/或表6中的一个或多个小鼠ighj基因的缺失。
[0012]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源重链免疫球蛋白基因座中的破坏包括从小鼠ighv1-85基因开始至小鼠ighj4基因的连续序列的缺失。
[0013]
在一些实施方案中,所述动物包含一个或多个内源ighm、ighδ、ighg3、ighg1、ighg2b、ighg2a、ighe和igha基因。
[0014]
在一些实施方案中,所述动物包含源自人重链免疫球蛋白基因座的未修饰的人序列。
[0015]
在一些实施方案中,所述未修饰的人序列约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb或1000kb。
[0016]
在一些实施方案中,所述动物包含源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人igv1-2的未修饰的人序列。在一些实施方案中,所述动物包含源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighv6-1的未修饰的人序列。在一些实施方案中,所述动物包含源自人重链免疫球蛋白基因座的从人ighd1-1开始至人ighj6的未修饰的人序列。
[0017]
在一些实施方案中,所述动物包含源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighj6的未修饰的人序列。
[0018]
在一些实施方案中,所述动物对于所述重链免疫球蛋白基因座是纯合的。在一些实施方案中,所述动物对于所述重链免疫球蛋白基因座是杂合的。
[0019]
在一些实施方案中,所述动物还包括在内源轻链免疫球蛋白基因座上包含一个或多个人igkv基因和一个或多个人igkj基因。
[0020]
在一些实施方案中,所述动物包含在其内源λ轻链免疫球蛋白基因座中的破坏。
[0021]
在一些实施方案中,所述动物是啮齿动物(例如,小鼠)。
[0022]
在一些方面,本公开涉及遗传修饰动物,所述遗传修饰动物在内源重链免疫球蛋白基因座上包含第一序列,所述第一序列包含一个或多个人ighv基因;第二序列,其包含内源序列;以及第三序列,其包含一个或多个人ighd基因和一个或多个人ighj基因,其中所述第一序列、第二序列和第三序列可操作地连接。
[0023]
在一些实施方案中,所述第一序列包含选自表1的约或至少150个、151个、152个、153个、154个、155个、156个、157个、158个、159个、160个或161个人ighv基因。在一些实施方案中,所述第一序列包含选自表2的约或至少20个、21个、22个、23个、24个、25个、26个或27
个人ighd基因。
[0024]
在一些实施方案中,所述第一序列是源自人重链免疫球蛋白基因座的未修饰的序列。在一些实施方案中,所述第一序列约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb或1000kb。
[0025]
在一些实施方案中,所述第二序列包含约为或至少为1kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、10kb、11kb、12kb、13kb、14kb、15kb、16kb、17kb、18kb、19kb或20kb的内源序列。
[0026]
在一些实施方案中,所述第三序列包含选自表2的约或至少20个、21个、22个、23个、24个、25个、26个或27个人ighd基因。在一些实施方案中,所述第三序列包含选自表3的约或至少5个、6个、7个、8个或9个人ighj基因。在一些实施方案中,所述第三序列包含表2中的全部人ighd基因和表3中的全部人ighj基因。
[0027]
在一些实施方案中,所述第三序列是源自人重链免疫球蛋白基因座的未修饰序列。在一些实施方案中,所述第三序列约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb或100kb。
[0028]
在一些实施方案中,所述动物包含在其内源重链免疫球蛋白基因座中的破坏。
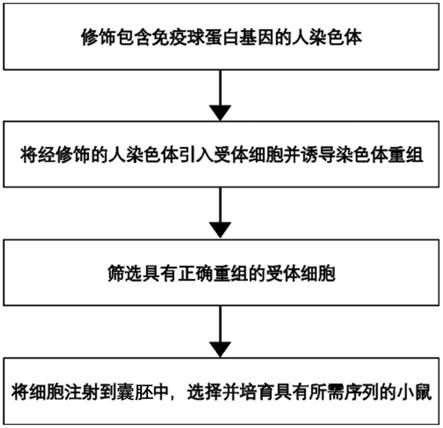
[0029]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源重链免疫球蛋白基因座中的破坏包括表4中的一个或多个小鼠ighv基因、表5中的一个或多个小鼠ighd基因和表6中的一个或多个小鼠ighj基因的缺失。
[0030]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源重链免疫球蛋白基因座中的破坏包括从小鼠ighv1-85开始至小鼠ighj4的序列的缺失。
[0031]
在一些实施方案中,所述动物包含选自ighm、ighδ、ighg3、ighg1、ighg2b、ighg2a、ighe和igha基因的一个或多个内源基因。
[0032]
在一些实施方案中,所述动物对于所述重链免疫球蛋白基因座是纯合的。在一些实施方案中,所述动物对于所述重链免疫球蛋白基因座是杂合的。
[0033]
在一些实施方案中,所述动物还在内源轻链免疫球蛋白基因座上包含一个或多个人igkv基因和一个或多个人igkj基因。
[0034]
在一些实施方案中,所述动物包含在其内源λ轻链免疫球蛋白基因座中的破坏。
[0035]
在一些实施方案中,所述动物是啮齿动物(例如,小鼠)。
[0036]
在一些方面,本公开涉及经遗传修饰的非人动物,其在内源轻链免疫球蛋白基因座上包含一个或多个人igkv基因和一个或多个人igkj基因。
[0037]
在一些实施方案中,所述动物包含表7中的约或至少65个、66个、67个、68个、69个、70个、71个、72个、73个、74个、75个或76个人igkv基因,和/或包含表8中的约或至少1个、2个、3个、4个或5个人igkj基因。
[0038]
在一些实施方案中,所述动物包含源自人轻链免疫球蛋白基因座的从人igkv3d-7开始至人igkj5的未修饰的人序列。
[0039]
在一些实施方案中,所述动物包含在其内源轻链免疫球蛋白基因座中的破坏。
[0040]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源轻链免疫球蛋白基因座中的破坏包括表9中的一个或多个小鼠igkv基因和表10中的一个或多个小鼠igkj基因的缺失。在一些实施方案中,所述动物包含人2号染色体的内源κ链免疫球蛋白基因座上的全
部人igkv基因以及全部人igkj基因。在一些实施方案中,所述动物包含在人细胞(例如,体细胞、培养细胞、非免疫细胞、无任何v(d)j重排的细胞)的人2号染色体的内源重链免疫球蛋白基因座上的全部人igkv基因和全部人igkj基因。
[0041]
在一些实施方案中,所述动物是小鼠,并且所述动物的内源轻链免疫球蛋白基因座中的破坏包括从小鼠igkv2-137开始至小鼠igkj5的序列的缺失。
[0042]
在一些实施方案中,所述动物包含内源igkc。
[0043]
在一些实施方案中,所述动物对于所述轻链免疫球蛋白基因座是纯合的。在一些实施方案中,所述动物对于所述轻链免疫球蛋白基因座是杂合的。
[0044]
在一些实施方案中,所述动物在内源重链免疫球蛋白基因座上还包含一个或多个人ighv基因、一个或多个人ighd基因和一个或多个人ighj基因。
[0045]
在一些实施方案中,所述动物包含在其内源λ轻链免疫球蛋白基因座中的破坏。
[0046]
在一些实施方案中,所述动物是啮齿动物(例如,小鼠)。
[0047]
在一些方面,本公开涉及经遗传修饰的非人动物,其基因组包含内源重链免疫球蛋白基因座,该内源重链免疫球蛋白基因座包含:将一个或多个内源ighv、内源ighd和内源ighj基因替换为一个或多个人ighv、人ighd和人ighj基因。在一些实施方案中,人ighv、人ighd和人ighj基因可操作地连接至选自ighm、ighδ、ighg、ighe和igha基因的一个或多个内源基因。
[0048]
在一些实施方案中,一个或多个内源ighv、内源ighd和内源ighj基因被替换为表1中的约或至少150个、151个、152个、153个、154个、155个、156个、157个、158个、159个、160个或161个人ighv基因、表2中的约或至少20个、21个、22个、23个、24个、25个、26个或27个人ighd基因和表3中的约或至少5个、6个、7个人ighj基因。
[0049]
在一些实施方案中,所述动物是小鼠,并且表4中的约或至少180个小鼠ighv基因、表5中的全部小鼠ighd基因和表6中的全部小鼠ighj基因被替换。
[0050]
在一些方面,本公开涉及经遗传修饰的非人动物,其基因组包含内源轻链免疫球蛋白基因座,该内源轻链免疫球蛋白基因座包含:一个或多个内源igkv和内源igkj基因被替换为一个或多个人igkv和人igkj基因。在一些实施方案中,所述人igkv和人igkj基因可操作地连接至内源igkc基因。
[0051]
在一些实施方案中,一个或多个内源igkv和内源igkj基因被替换为表7中的约或至少65个、66个、67个、68个、69个、70个、71个、72个、73个、74个、75个或76个人igkv基因和表8中的约或至少1个、2个、3个、4个或5个人igkj基因。
[0052]
在一些实施方案中,所述动物是小鼠,并且表9中的全部小鼠igkv基因和表10中的全部小鼠igkj基因被替换。
[0053]
在一些实施方案中,所述动物缺乏能够重排和形成编码内源重链可变结构域(例如,小鼠重链可变结构域)的核酸序列的内源免疫球蛋白重链可变结构域基因座。
[0054]
在一些实施方案中,所述动物缺乏能够重排和形成编码内源轻链可变结构域(例如,小鼠轻链可变结构域)的核酸序列的内源免疫球蛋白轻链可变结构域基因座。
[0055]
在一些实施方案中,所述动物可产生人源化抗体。
[0056]
在一些方面,本公开涉及从如本文所述动物获得的细胞。
[0057]
在一些实施方案中,所述细胞是表达嵌合免疫球蛋白重链的b细胞,所述嵌合免疫
球蛋白重链包含免疫球蛋白重链可变结构域,所述免疫球蛋白重链可变结构域源自一个或多个人ighv基因、一个或多个人ighd基因和一个或多个人ighj基因的重排。在一些实施方案中,所述免疫球蛋白重链可变结构域可操作地连接至非人重链恒定区。
[0058]
在一些实施方案中,所述细胞是表达嵌合免疫球蛋白轻链的b细胞,所述嵌合免疫球蛋白轻链包含免疫球蛋白轻链可变结构域,所述免疫球蛋白轻链可变结构域源自一个或多个人igkv基因和一个或多个人igkj基因的重排,并且其中所述免疫球蛋白轻链可变结构域可操作地连接至非人轻链恒定区。
[0059]
在一些实施方案中,所述细胞是胚胎干(es)细胞。
[0060]
在一些方面,本公开涉及制备与抗原特异性结合的嵌合抗体的方法,该方法包括将本文所述的动物暴露于抗原;由从所述动物收集的细胞产生杂交瘤;以及收集所述杂交瘤产生的所述嵌合抗体。在一些实施方案中,分离所述目标细胞并进行测序以确定重排的重链可变区和轻链可变区的序列。
[0061]
在一些实施方案中,该方法还包括对所述杂交瘤的基因组进行测序。
[0062]
在一些方面,本公开涉及修饰细胞的基因组的方法,该方法包括修饰人染色体;将经修饰的人染色体引入动物细胞;以及诱导经修饰的人染色体与内源染色体之间的重组,从而用一个或多个人基因替换一个或多个内源基因。
[0063]
在一些实施方案中,所述经修饰的人染色体包含两个或更多个外源重组位点。
[0064]
在一些实施方案中,所述内源染色体包含两个或更多个外源重组位点。
[0065]
在一些实施方案中,通过重组将选自表1的约或至少150个、151个、152个、153个、154个、155个、156个、157个、158个、159个、160个或161个人ighv基因、选自表2的约或至少20个、21个、22个、23个、24个、25个、26个或27个人ighd基因以及选自表3的约或至少5、6、7、8或9个人ighj基因整合到所述内源染色体中。
[0066]
在一些实施方案中,通过重组将表7中的约或至少65个、66个、67个、68个、69个、70个、71个、72个、73个、74个、75个或76个人igkv基因和表8中的约或至少1个、2个、3个、4个或5个人igkj基因整合到所述内源染色体中。
[0067]
在一些实施方案中,通过重组将人序列整合到所述内源染色体中,并且所述人序列约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb或1000kb。
[0068]
一方面,本公开提供了制备与抗原特异性结合的抗体的方法。该方法包括在细胞中获得编码人重链和轻链免疫球蛋白可变区的核酸序列,所述细胞表达与所述抗原特异性结合的嵌合抗体(hybrid antibody),其中所述细胞通过将本文所述的动物暴露于所述抗原而获得;将编码人重链免疫球蛋白可变区的核酸与编码人重链免疫球蛋白恒定区的核酸以及将编码人轻链免疫球蛋白可变区的核酸与编码人轻链免疫球蛋白恒定区的核酸可操作地连接;以及在细胞中表达所述核酸,从而获得所述抗体。
[0069]
一方面,本公开提供了获得编码与抗原特异性结合的抗体结合结构域的核酸的方法。该方法包括将本文所述的动物暴露于所述抗原;以及对细胞中编码人重链和轻链免疫球蛋白可变区的核酸进行测序,所述细胞表达与所述抗原特异性结合的嵌合抗体。
[0070]
一方面,本公开提供了获得样品的方法,该方法包括将本文所述的动物暴露于所述抗原;以及从所述动物收集样品。在一些实施方案中,所述样品是脾组织、脾细胞或b细
胞。
[0071]
一方面,本公开提供了制备与抗原特异性结合的抗体的方法。该方法包括将本文所述的动物暴露于所述抗原;在表达与所述抗原特异性结合的嵌合抗体的细胞中获得(例如通过测序)编码人重链和轻链免疫球蛋白可变区的核酸序列;以及在细胞中将编码人重链免疫球蛋白可变区的核酸与编码人重链免疫球蛋白恒定区的核酸以及将编码人轻链免疫球蛋白可变区的核酸与编码人轻链免疫球蛋白恒定区的核酸可操作地连接。
[0072]
本公开还涉及所述非人哺乳动物的后代。
[0073]
在一些实施方案中,所述非人哺乳动物是啮齿动物。在一些实施方案中,所述非人哺乳动物是小鼠。
[0074]
本公开还提供了包含本文所述的靶向载体的细胞。
[0075]
本公开还涉及源自所述非人哺乳动物或其后代的细胞(例如,干细胞、胚胎干细胞、免疫细胞、b细胞、t细胞或杂交瘤)或细胞系或其原代细胞培养物。本公开还涉及源自所述非人哺乳动物或其后代的组织、器官或其培养物。
[0076]
本公开还涉及所述非人哺乳动物或其后代、通过本文所述的方法产生的动物模型在开发与免疫过程相关的产品、制造人抗体或用于药理学、免疫学、微生物学和医学研究的模型系统方面的用途。
[0077]
除非另有定义,否则本文使用的所有技术和科学术语具有与本发明所属领域的普通技术人员通常理解的含义相同的含义。本文描述了用于本发明的方法和材料;还可使用本领域已知的其它合适的方法和材料。材料、方法和实例仅是说明性的,而不旨在是限制性的。本文提及的所有出版物、专利申请、专利、序列、数据库条目和其它参考文献通过引用整体并入本文。如有冲突,以本说明书(包括定义)为准。
[0078]
根据下面的详细描述和附图以及权利要求,本发明的其它特征和有利方面将变得明显。
附图说明
[0079]
图1a是将人免疫球蛋白基因引入小鼠基因组的方法的流程图。
[0080]
图1b是用人免疫球蛋白重链可变区替换小鼠免疫球蛋白重链可变区的概述。
[0081]
图1c是用人免疫球蛋白轻链可变区替换小鼠免疫球蛋白轻链可变区的概述。
[0082]
图2是显示小鼠重链免疫球蛋白基因座的示意图。
[0083]
图3a是显示在向基因组中引入两个重组位点后小鼠重链免疫球蛋白基因座的示意图。
[0084]
图3b是显示在用靶向载体重组后小鼠重链免疫球蛋白基因座的示意图。
[0085]
图4显示了用于修饰小鼠重链免疫球蛋白基因座的打靶策略。
[0086]
图5a显示了使用mighv-5’loxp-l-gt-f/mighv-005-l-gt-r2引物对的pcr检测的结果。
[0087]
图5b显示了使用mighv-005-5’loxp-r-gt-f2/mighv-5’loxp-r-gt-r引物对的pcr检测的结果。
[0088]
图6a显示了使用mighv-3’lox-l-gt-f2/mighv-3’lox-l-gt-r2引物对的pcr检测的结果。
[0089]
图6b显示了使用mighv3’lox-r-gt-f2/mighv-3’lox-r-gt-r1引物对的pcr检测的结果。
[0090]
图7是突出了重链免疫球蛋白基因座(未按比例绘制)的人14号染色体的示意图。重链免疫球蛋白基因座有可变区(v
h
,d
h
,j
h
)和恒定区(c
h
)。v
h
代表ighv基因簇的区段,d
h
代表ighd基因簇的区段,jh代表ighj基因簇的区段,c
h
代表表达恒定结构域的基因簇。
[0091]
图8是显示修饰后的人14号染色体的示意图。
[0092]
图9显示了用两个载体对人14号染色体进行的修饰。
[0093]
图10显示了经修饰的人14号染色体。
[0094]
图11显示了使用highv-5’loxp-l-gt-f1和highv-5’loxp-r-gt-r引物对对染色体hchr14-mut3上loxp位点301的pcr检测的结果。8-d7为阳性对照克隆。
[0095]
图12显示了染色体hchr14-mut3上loxp位点302的pcr检测的结果。8-d7是阴性对照克隆。
[0096]
图13是人14号染色体被修饰前细胞的荧光原位杂交(fish)图像。
[0097]
图14是人14号染色体被修饰后的细胞的fish图像。
[0098]
图15是显示经修饰的小鼠12号染色体的示意图。
[0099]
图16是显示cre介导的重组的示意图,所述重组用相应的人基因组dna序列替换小鼠重链可变区基因座。
[0100]
图17显示了使用m5-l引物对和m5-r引物对的pcr检测的结果。
[0101]
图18显示了使用m3引物对的pcr检测的结果。
[0102]
图19显示了使用h5-l引物对和h5-r引物对的pcr检测的结果。
[0103]
图20显示了使用h3-l引物对和h3-r引物对的pcr检测的结果。
[0104]
图21是fish图像。白色箭头(1)和(2)表示小鼠12号染色体。白色箭头(3)表示由人特异性igh断裂分离探针标记的人染色体片段。
[0105]
图22是显示flp介导的重组的示意图。
[0106]
图23是显示小鼠轻链免疫球蛋白基因座的示意图。
[0107]
图24a是显示在基因组中引入两个重组位点后小鼠轻链免疫球蛋白基因座的示意图。
[0108]
图24b是显示用靶向载体重组后小鼠轻链免疫球蛋白基因座的示意图。
[0109]
图25是显示小鼠6号染色体的基因打靶策略的示意图。
[0110]
图26显示了使用igkv-005-c-5g-l-gt-f/igkv-005-c-5g-l-gt-r2引物对的pcr检测的结果。
[0111]
图27显示了使用igkv-005-c-5g-r-gt-f1/igkv-005-c-5g-r-gt-r引物对的pcr检测的结果。
[0112]
图28是突出显示轻链免疫球蛋白基因座的人2号染色体的示意图(未按比例绘制)。v
hk
代表igkv基因簇的区段,j
hk
代表igkj基因簇的区段,c
hk
代表igkc基因。
[0113]
图29是显示经修饰的人2号染色体的示意图。
[0114]
图30是显示人2号染色体的基因打靶策略的示意图。
[0115]
图31显示了第一次重组(引入载体2702)后的pcr测定的结果。wt是野生型h9细胞。
[0116]
图32是fish图像结果。白色箭头表示正确重组的经修饰的人2号染色体。
[0117]
图33显示了第二次重组(引入载体2701)后的pcr检测的结果。
[0118]
图34是显示cre介导的重组的示意图,其中人轻链可变区基因组dna序列被添加到相应的小鼠基因座。
[0119]
图35是荧光原位杂交(fish)图像。箭头(1)和(3)表示小鼠6号染色体。箭头(2)表示由人特异性igk断裂分离探针标记的人染色体片段。
[0120]
图36是显示flp介导的重组的示意图。
[0121]
图37是显示14号染色体(14q32.33)上的人免疫球蛋白重链(igh)基因座的示意图。
[0122]
图38是显示12号染色体(12f2)(品系c57bl/6)上的小鼠(mus musculus)igh基因座的示意图。
[0123]
图39是显示2号染色体(2p11.2)上的人免疫球蛋白κ链(igk)基因座的示意图。
[0124]
图40是显示6号染色体(6c1)上的小鼠(mus musculus)igk基因座的示意图。
[0125]
图41列出了人重链免疫球蛋白基因座(igh)的imgt谱系。
[0126]
图42列出了小鼠igh的imgt谱系。
[0127]
图43列出了人κ链免疫球蛋白基因座(igk)的imgt谱系。
[0128]
图44列出了小鼠igk的imgt谱系。
[0129]
图45显示了在外周血中检测到的白细胞的百分比。
[0130]
图46显示了在脾细胞中检测到的白细胞的百分比。
[0131]
图47显示了在淋巴结中检测到的白细胞的百分比。
[0132]
图48显示了处于不同发育阶段的脾b细胞的百分比。
[0133]
图49显示了处于不同发育阶段的淋巴结b细胞的百分比。
[0134]
图50显示脾边缘区(mz-b)和滤泡区(fo-b)的脾b细胞的百分比。
[0135]
图51a显示了从野生型小鼠获得的骨髓中处于不同发育阶段的b细胞的流式细胞术分析结果。1区表示原b细胞,2区表示前b细胞,3区表示未成熟b细胞。
[0136]
图51b显示从人源化重链杂合小鼠获得的骨髓中处于不同发育阶段的b细胞的流式细胞术分析结果。1区表示原前b细胞,2区表示前b细胞,3区表示未成熟b细胞。
[0137]
图51c显示从人源化重链纯合小鼠获得的骨髓中处于不同发育阶段的b细胞的流式细胞术分析结果。1区表示原b细胞,2区表示前b细胞,3区表示未成熟b细胞。
[0138]
图52显示了梯度稀释的小鼠血清中的iga同种型水平。
[0139]
图53显示了梯度稀释的小鼠血清中的igg1同种型水平。
[0140]
图54显示了梯度稀释的小鼠血清中的igg2b同种型水平。
[0141]
图55显示了梯度稀释的小鼠血清中的igg2c同种型水平。
[0142]
图56显示了梯度稀释的小鼠血清中的igg3同种型水平。
[0143]
图57显示了梯度稀释的小鼠血清中的igm同种型水平。
[0144]
图58显示了vj重组后检测到的igkv基因表达在个体小鼠中的分布。
[0145]
图59显示了用人btla免疫后野生型小鼠的流式细胞术结果。
[0146]
图60显示了人源化重链纯合小鼠经人btla免疫后的流式细胞术结果。
[0147]
图61显示了用犬pd-1(dpd-1)免疫后野生型小鼠的流式细胞术结果。
[0148]
图62显示了用犬pd-1(dpd-1)免疫后人源化重链纯合小鼠的流式细胞术结果。
[0149]
图63显示在用卵清蛋白(ova)免疫后野生型小鼠(黑色柱;小鼠以1-5标记)和人源化重链纯合小鼠(灰色柱;小鼠以6-10标记)的elisa结果概述。
[0150]
图64显示了人远端vκ簇igkv基因的列表和人近端vκ簇igkv基因的列表。
[0151]
图65a显示了未免疫的野生型小鼠和hvh/hvl小鼠的体重。
[0152]
图65b显示了未免疫的野生型小鼠和hvh/hvl的脾脏重量。
[0153]
图66显示了未免疫的野生型小鼠和hvh/hvl小鼠脾脏中免疫细胞的百分比。
[0154]
图67a显示了脾b细胞中过渡1型(t1,b220
+
igm
+
igd-),过渡2型(t2,b220
+
igm
+
igd
+
)和成熟型(m,b220
+
igm
低
igd
+
)b细胞群的百分比。脾b细胞来自未免疫的野生型或hvh/hvl小鼠。
[0155]
图67b显示脾b细胞中边缘区(mz)和滤泡(fo)b细胞群的百分比。脾b细胞来自未免疫的野生型或hvh/hvl小鼠。
[0156]
图68a显示了骨髓b细胞中原b细胞(b220
低
cd43
高
igm
低
)、前b细胞(b220
低
cd43
中
igm
低
)和未成熟b细胞(b220
高
cd43
低
igm
高
)群的百分比。骨髓b细胞来自未免疫的野生型或hvh/hvl小鼠。
[0157]
图68b显示了骨髓b细胞中浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)群的百分比。骨髓b细胞来自未免疫的野生型或hvh/hvl小鼠。
[0158]
图68c显示脾b细胞中浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)群的百分比。脾b细胞来自未免疫的野生型或hvh/hvl小鼠。
[0159]
图69显示了未免疫的野生型或hvh/hvl小鼠中血清免疫球蛋白(ig)亚型的浓度。ig亚型浓度通过elisa来定量测定。
[0160]
图70a显示了未免疫的hvh/hvl小鼠中的ighv利用率(频率>1%)。
[0161]
图70b显示了未免疫的hvh/hvl小鼠中的ighv利用率((频率<1%)。
[0162]
图70c显示了未免疫的hvh/hvl小鼠中的ighd利用率。
[0163]
图70d显示了未免疫的hvh/hvl小鼠中的ighj利用率。
[0164]
图71a显示了未免疫的hvh/hvl小鼠中的igkv利用率(频率>1%)。
[0165]
图71b显示了未免疫的hvh/hvl小鼠中的igkv利用率(频率<1%)。
[0166]
图71c显示了未免疫的hvh/hvl小鼠中的igkj利用率。
[0167]
图72是显示来自未免疫的hvh/hvl小鼠的重链cdr3氨基酸长度分布的直方图。
[0168]
图73显示了未免疫的hvh/hvl小鼠中重链cdr3上的氨基酸频率。
[0169]
图74显示了hvh/hvl小鼠hcdr3中半胱氨酸残基的的频率。
[0170]
图75a是来自未免疫的野生型小鼠的脾脏的组织学图像。
[0171]
图75b是来自未免疫的野生型小鼠的腹股沟淋巴结的组织学图像。
[0172]
图75c是来自未免疫的野生型小鼠的派尔集合淋巴结(peyer
′
spatch)的组织学图像。
[0173]
图75d是来自未免疫的hvh/hvl小鼠的脾脏的组织学图像。
[0174]
图75e是来自未免疫的hvh/hvl小鼠的腹股沟淋巴结的组织学图像。
[0175]
图75f是来自未免疫的hvh/hvl小鼠的派尔集合淋巴结(peyer
′
spatch)的组织学图像。
[0176]
图76a显示了在野生型和hvh/hvl小鼠中使用人bcma(b细胞成熟抗原)作为抗原的
第二次和第三次免疫后的bcma特异性抗体的滴度。
[0177]
图76b显示了在野生型和hvh/hvl小鼠中使用人il4r(白细胞介素-4受体)作为抗原的第二次和第三次免疫后的il4r特异性抗体的滴度。
[0178]
图76c显示了在野生型和hvh/hvl小鼠中使用人pd-1(程序性细胞死亡蛋白1)作为抗原的第二次和第三次免疫后的pd-1特异性抗体的滴度。
[0179]
图76d显示了在野生型和hvh/hvl小鼠中使用人siglec15(唾液酸结合ig样凝集素15)作为抗原的第二次和第三次免疫后siglec15特异性抗体的滴度。
[0180]
图76e显示了在野生型和hvh/hvl小鼠中使用人sirpα(信号调节蛋白α)作为抗原的第二次和第三次免疫后的sirpα特异性抗体的滴度。
[0181]
图77a显示了免疫后野生型小鼠和hvh/hvl小鼠的体重。
[0182]
图77b显示了免疫后野生型小鼠和hvh/hvl小鼠的脾脏重量。
[0183]
图78显示了免疫后野生型小鼠和hvh/hvl小鼠的脾脏中免疫细胞的百分比。
[0184]
图79a显示了脾b细胞中过渡1型(t1,b220-igm
+
igd-)、过渡2型(t2,b220
+
igm
+
igd
+
)和成熟型(m,b220
+
igm
低
igd
+
)b细胞群的百分比。脾b细胞来自免疫后的野生型或hvh/hvl小鼠。
[0185]
图79b显示脾b细胞中边缘区(mz)和滤泡(fo)b细胞群的百分比。脾b细胞来自免疫后的野生型或hvh/hvl小鼠。
[0186]
图80a显示了骨髓b细胞中原b细胞(b220
低
cd43
高
igm
低
)、前b细胞(b220
低
cd43
中
igm
低
)和未成熟b细胞(b220
高
cd43
低
igm
高
)群的百分比。骨髓b细胞来自免疫后的野生型或hvh/hvl小鼠。
[0187]
图80b显示了骨髓b细胞中浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)群的百分比。骨髓b细胞来自免疫后的野生型或hvh/hvl小鼠。
[0188]
图80c显示脾b细胞中浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)群的百分比。脾b细胞来自免疫后的野生型或hvh/hvl小鼠。
[0189]
图81显示了未免疫的或第三次免疫后的野生型或hvh/hvl小鼠中的血清免疫球蛋白(ig)亚型的浓度。ig亚型浓度通过用elisa测定。
[0190]
图82显示了未免疫的或免疫后的野生型或hvh/hvl小鼠中的血清总igg浓度。igg浓度通过elisa测定。
具体实施方式
[0191]
本公开涉及具有人源化重链免疫球蛋白基因座和/或人源化轻链免疫球蛋白基因座(例如κ链基因座)的经遗传修饰的动物和细胞。
[0192]
可通过将人免疫球蛋白基因引入非人动物的基因组来产生经遗传修饰的动物,所述动物可表达人源化抗体或嵌合抗体的动物。图1a显示了产生人源化小鼠的方法。在一些实施方案中,该方法首先涉及修饰人染色体上的人免疫球蛋白区域。然后将该经修饰的人染色体引入小鼠受体细胞。然后通过直接替换(例如,在一步替换中)将人免疫球蛋白可变区引入小鼠基因组的相应区域。然后筛选受体细胞,优选不含人染色体的细胞。然后将细胞注射到囊胚中以制备嵌合动物(例如,小鼠)。可以进行后续育种以获得含有完整人源化免疫球蛋白基因座的动物。
[0193]
本文描述的转基因动物具有各种优势。例如,在一些情况下,本文所述的经遗传修饰的动物具有完整的人抗体谱系。因此,由这些动物产生的可变结构域可具有与人中的可变结构域的多样性非常相似的多样性。此外,因为人免疫球蛋白基因座上的整个序列(没有修饰或具有有限的修饰)被引入动物基因组,这些基因可以以与人中发生的非常相似的方式进行v(d)j重组。另外,由于有效的v(d)j重组,抗体可以非常有效的产生,并且具有与正常速率相似的速率。另外,因为内源ighv、ighd、ighj、igkv和igkj基因与人基因之间可发生v(d)j重组,如果内源ighv、ighd、ighj、igkv和igkj基因被整合到重排的重链vdj区段或重排的轻链vj区段中,则由该抗体谱系产生的抗体很可能在人中具有免疫原性表位。免疫原性可导致抗药物抗体的产生且可具有活性。此处,内源ighv、ighd、ighj、igkv和igkj基因被有效删除。由该抗体谱系产生的抗体在人体内不太可能具有免疫原性。因此,抗体更适合用作人的治疗剂。因此,经遗传修饰的动物为产生人源化抗体提供了有利的平台。
[0194]
如本文中所用,术语“抗体”是指包含通过二硫键相互连接的四条多肽链(两条重链和两条轻链)的免疫球蛋白分子。每条重链包含重链可变(vh)结构域和重链恒定区(ch)。每条轻链包含轻链可变(vl)结构域和轻链恒定区(cl)。vh和vl结构域可进一步细分为高变区,称为互补决定区(cdr),散布有更保守的区域,称为框架区(fr)。每个vh和vl包含从氨基端至羧基端按以下顺序排列的三个cdr和四个fr:fr1、cdr1、fr2、cdr2、fr3、cdr3、fr4(重链cdr可缩写为hcdr1、hcdr2和hcdr3;轻链cdr可缩写为lcdr1、lcdr2和lcdr3)。术语“高亲和力”抗体是指相对于其靶表位的k
d
约为10-9
m或更低(例如,约为或低于1
×
10-9
m、1
×
10-10
m、1
×
10-11
m或1
×
10-12
m)的抗体。在一些实施方案中,k
d
可通过表面等离子体共振(例如biacore
tm
)或elisa来测量。
[0195]
如本文中所用,术语“抗原结合片段”是指全长抗体的一部分,其中该抗体的部分能够与抗原特异性结合。在一些实施方案中,抗原结合片段包含至少一个可变结构域(例如,重链的可变结构域或轻链的可变结构域)。抗体片段的非限制性实例包括例如fab、fab’、f(ab’)2和fv片段。
[0196]
如本文中所用,术语“人抗体”是指由存在于人体内的核酸(例如,重排的人免疫球蛋白重链或轻链基因座)编码的抗体。在一些实施方案中,从人收集人抗体或在人细胞培养物(例如,人杂交瘤细胞)中产生人抗体。在一些实施方案中,在非人细胞(例如,小鼠或仓鼠细胞系)中产生人抗体。在一些实施方案中,在细菌或酵母细胞中产生人抗体。在一些实施方案中,在含有未重排或重排的人免疫球蛋白基因座(例如,重链或轻链人免疫球蛋白基因座)的转基因非人动物(例如,小鼠)中产生人抗体。
[0197]
如本文中所用,术语“嵌合抗体”是指包含存在于至少两种不同抗体中的序列的抗体(例如,来自两种不同哺乳动物物种的抗体,例如人和小鼠的抗体)。嵌合抗体的非限制性实例是包含人抗体的可变结构域序列(例如,轻链和/或重链可变结构域序列的全部或部分)和非人抗体的恒定结构域的抗体。本文描述了嵌合抗体的其它实例,并且所述其它实例是本领域已知的。
[0198]
如本文中所用,术语“人源化抗体”是指包含源自非人(例如,小鼠)免疫球蛋白的序列,并且包含源自人免疫球蛋白的序列的非人抗体。
[0199]
如本文中所用,术语“单链抗体”是指能够与抗原特异性结合的包含至少两个免疫球蛋白可变结构域(例如,哺乳动物免疫球蛋白重链或轻链的可变结构域)的单个多肽。
[0200]
如本文中所用,术语“受试者”和“患者”在整个说明书中可互换使用,并用于描述动物、人或非人。本发明考虑了兽医和非兽医应用。人患者可以是成人或青少年(例如,18岁以下的人)。除了人以外,患者还包括但不限于小鼠、大鼠、仓鼠、豚鼠、兔子、雪貂、猫、狗和灵长类动物。包括例如非人灵长类动物(例如,猴子、黑猩猩、大猩猩等)、啮齿动物(例如,大鼠、小鼠、沙鼠、仓鼠、雪貂、兔子)、兔子形目动物、猪(例如,猪、小型猪)、马科动物、犬科动物、猫科动物、牛类动物以及其它家养、农场和动物园动物。
[0201]
如本文中所用,当提及抗体时,短语“特异性结合(specifically binding)”和“特异性结合(specifically binds)”意指抗体与其靶分子相互作用优于与其它分子相互作用,因为所述相互作用取决于靶分子上特定结构(即,抗原决定簇或表位)的存在;换句话说,试剂识别包含特定结构的分子并与其结合,而不是一般的所有分子。与靶分子特异性结合的抗体可称为靶特异性抗体。
[0202]
如本文中所用,术语“多肽”、“肽”和“蛋白质”可互换地用于指具有至少为两个氨基酸的任意长度的氨基酸聚合物。
[0203]
如本文中所用,术语“多核苷酸”、“核酸分子”和“核酸序列”在本文中可互换用于指具有至少两个核苷酸的任意长度的核苷酸的聚合物,包括但不限于dna、rna、dna/rna杂交体及其修饰形式。
[0204]
如本文中所用,术语“未修饰的人序列”是指源自人受试者、人细胞、培养的人细胞或人细胞系的序列,其中该序列与人受试者、人细胞、培养的人细胞或人细胞系的遗传序列相同。
[0205]
经遗传修饰的重链免疫球蛋白基因座
[0206]
重链免疫球蛋白基因座(也称为igh或免疫球蛋白重链基因座)是染色体(例如,人14号染色体)上包含人抗体(或免疫球蛋白)重链基因的区域。
[0207]
这个区域代表重链基因座的种系组织。基因座包括v(可变的)、d(多样性)、j(连接)和c(恒定)区段。v区中的基因形成v基因簇(也称为ighv基因簇)。d区中的基因形成d基因簇(也称为ighd基因簇)。j区中的基因形成j基因簇(也称为ighj基因簇)。
[0208]
在b细胞发育过程中,dna水平上的重组事件将单个d区段(也称为ighd基因)与j区段(也称为ighj基因)连接起来;该部分重排的d-j区的融合的d-j外显子然后被连接至v区段(也称为ighv基因)。然后含有融合的v-d-j外显子的重排的v-d-j区被转录并在rna水平上融合至ighm恒定区;该转录物编码μ重链。在发育的后期,b细胞产生v-d-j-cμ-cδ前信使rna,该rna被选择性剪接以编码μ或δ重链。淋巴结中的成熟b细胞经历转换重组,使得融合的v-d-j基因区段接近ighg、igha或ighe基因区段之一,并且每个细胞表达γ、α或ε重链。许多不同的ighv基因与几个ighj基因的潜在重组提供了广泛的抗原识别。额外的多样性是通过连接多样性获得的,连接多样性是由末端脱氧核苷酸转移酶随机添加核苷酸和体细胞过度突变产生的,这发生在脾和淋巴结的b细胞成熟过程中。已知几个v、d、j和c区段不能编码蛋白,被认为是假基因区段(通常简称为假基因)。
[0209]
人重链免疫球蛋白基因座位于人14号染色体上。表1列出了ighv基因及其在该基因座中的相对顺序。
[0210]
表1.人14号染色体上的ighv基因列表
[0211][0212][0213]
rps8p1、adam6和kiaa0125也位于该基因座。rps8p1的相对顺序为160,adam6的相对顺序为161,kiaa0125的相对顺序为164。表2列出了人14号染色体上的全部ighd基因及其相对顺序。表3列出了人14号染色体上全部的ighj基因及其相对顺序。免疫球蛋白恒定结构域的基因位于ighv、ighd和ighj基因之后。这些基因包括(如以下顺序所示):免疫球蛋白重链恒定μ(ighm)、免疫球蛋白重链恒定δ(ighδ)、免疫球蛋白重链恒定γ3(ighg3)、免疫球蛋白重链恒定γ1(ighg1)、免疫球蛋白重链恒定εp1(假基因)(ighep1)、免疫球蛋白重链恒定α1(igha1)、免疫球蛋白重链恒定γp(非功能性)(ighgp)、免疫球蛋白重链恒定γ2(ighg2)、免疫球蛋白重链恒定γ4(ighg4)、免疫球蛋白重链恒定ε(ighe)和免疫球蛋白重链恒定α2(igha2)。这些基因和这些基因的顺序也显示在图37和图41中。
[0214]
表2.人14号染色体上的ighd基因列表
[0215]
基因名称顺序基因名称顺序基因名称顺序基因名称顺序ighd1-1165ighd2-8172ighd2-15179ighd3-22186ighd2-2166ighd3-9173ighd3-16180ighd4-23187ighd3-3167ighd3-10174ighd4-17181ighd5-24188ighd4-4168ighd4-11175ighd5-18182ighd6-25189ighd5-5169ighd5-12176ighd6-19183ighd1-26190ighd6-6170ighd6-13177ighd1-20184 *ighd1-7171ighd1-14178ighd2-21185ighd7-27192
[0216]
表3.人14号染色体上的ighj基因列表
[0217]
基因名称顺序基因名称顺序ighj1p191ighj4197ighj1193ighj5198ighj2194ighj3p199ighj2p195ighj6200ighj3196
ꢀꢀ
[0218]
小鼠重链免疫球蛋白基因座位于小鼠12号染色体上。表4列出了ighv基因及其在该基因座中的相对顺序。
[0219]
表4.小鼠12号染色体上的ighv基因列表
[0220]
[0221][0222]
表5列出了全部ighd基因及其在小鼠12号染色体上的相对顺序。表6列出了全部ighj基因及其在小鼠12号染色体上的相对顺序。免疫球蛋白恒定结构域的基因位于ighv、ighd和ighj基因之后。这些基因包括(如以下顺序所示):免疫球蛋白重链恒定μ(ighm)、免疫球蛋白重链恒定δ(ighδ)、免疫球蛋白重链恒定γ3(ighg3)、免疫球蛋白重链恒定γ1(ighg1)、免疫球蛋白重链恒定γ2b(ighg2b)、免疫球蛋白重链恒定γ2a(ighg2a)、免疫球蛋白重链恒定ε(ighe)和免疫球蛋白重链恒定α(igha)基因。这些基因和这些基因的顺序也示于图38和图42中。
[0223]
表5.小鼠12号染色体上的ighd基因列表
[0224][0225][0226]
表6.小鼠12号染色体上的ighj基因列表
[0227]
基因名称顺序基因名称顺序
ighj1203ighj3205ighj2204ighj4206
[0228]
本公开提供了经遗传修饰的非人动物,其包含一个或多个人ighv基因、一个或多个人ighd基因和/或一个或多个人ighj基因。在一些实施方案中,所述人ighv基因、人ighd基因和人ighj基因可操作地连接在一起,并且可进行vdj重排。在一些实施方案中,所述人ighv基因、人ighd基因和人ighj基因位于内源重链免疫球蛋白基因座上。
[0229]
在一些实施方案中,所述动物包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、20个、30个、40个、50个、60个、70个、80个、90个、100个、110个、120个、130个、140个、150个、151个、152个、153个、154个、155个、156个、157个、158个、159个、160个或161人ighv基因(例如,如表1所示的基因)。
[0230]
在一些实施方案中,所述动物包含1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv(iii)-82、ighv7-81、ighv4-80、ighv3-79、ighv(ii)-78-1、ighv5-78、ighv7-77、ighv(iii)-76-1、ighv3-76和ighv3-75的基因。
[0231]
在一些实施方案中,所述动物包含1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv(iii)-5-2、ighv(iii)-5-1、ighv2-5、ighv7-4-1、ighv4-4、ighv1-3、ighv(iii)-2-1、ighv1-2、ighv(ii)-1-1和ighv6-1的基因。
[0232]
在一些实施方案中,所述动物包含未修饰的人序列,所述未修饰的人序列包含起始于选自ighv(iii)-82、ighv7-81、ighv4-80、ighv3-79、ighv(ii)-78-1、ighv5-78、ighv7-77、ighv(iii)-76-1、ighv3-76和ighv3-75的基因并终止于选自ighv(iii)-5-2、ighv(iii)-5-1、ighv2-5、ighv7-4-1、ighv4-4、ighv1-3、ighv(iii)-2-1、ighv1-2、ighv(ii)-1-1和ighv6-1的基因的序列。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighv1-2的部分。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighv(ii)-1-1的部分。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighv-6-1的部分。
[0233]
在一些实施方案中,所述动物包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个、20个、21个、22个、23个、24个、25个、26个或27个人ighd基因(例如,如表2所示的基因)。在一些实施方案中,所述动物包含1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighd1-1、ighd2-2、ighd3-3、ighd4-4、ighd5-5、ighd4-23、ighd5-24、ighd6-25、ighd1-26和ighd7-27的基因。
[0234]
在一些实施方案中,所述动物包含约或至少1个、2个、3个、4个、5个、6个、7个、8个或9个人ighj基因(例如,表3所示的基因)。在一些实施方案中,所述动物包括1个、2个、3个、4个、5个、6个、7个、8个或9个选自ighj1p、ighj1、ighj2、ighj2p、ighj3、ighj4、ighj5、ighj3p和ighj6的人ighj基因。
[0235]
在一些实施方案中,所述动物包含未修饰的人序列,所述未修饰的人序列包含始于选自ighd1-1、ighd2-2、ighd3-3、ighd4-4、ighd5-5、ighd4-23、ighd5-24、ighd6-25、ighd1-26和ighd7-27的基因并终止于选自ighj1p、ighj1、ighj2、ighj2p、ighj3、ighj4、ighj5、ighj3p和ighj6的基因的序列。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighd1-1开始至人ighj6的部分。
[0236]
在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从ighd1-1开始至人ighd7-27的部分。
[0237]
在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighj1p开始至人ighj6的部分。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighj1开始至人ighj6的部分。
[0238]
在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv(iii)-82开始至人ighj6的部分。
[0239]
在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv1-2开始至人ighj6的部分。在一些实施方案中,所述未修饰的人序列源自人重链免疫球蛋白基因座的从人ighv(ii)-1-1开始至人ighj6的部分。在一些实施方案中,所述未修饰的人序列源自始人重链免疫球蛋白基因座的从人ighv6-1至人ighj6的部分。
[0240]
在一些实施方案中,所述动物可具有1个、2个、3个、4个、5个、6个、7个、8个、9个或10个未修饰的人序列。在一些实施方案中,所述未修饰的人序列具有约或至少10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb或1000kb的长度。
[0241]
在一些实施方案中,所述动物包含一个或多个选自免疫球蛋白重链恒定μ(ighm)、免疫球蛋白重链恒定δ(ighδ)、免疫球蛋白重链恒定γ3(ighg3)、免疫球蛋白重链恒定γ1(ighg1)、免疫球蛋白重链恒定γ2b(ighg2b)、免疫球蛋白重链恒定γ2a(ighg2a)、免疫球蛋白重链恒定ε(ighe)和免疫球蛋白重链恒定α(igha)基因的内源基因。在一些实施方案中,这些内源基因可操作地连接在一起。在一些实施方案中,这些内源基因具有与在野生型动物中相同的顺序。在一些实施方案中,同种型转换(免疫球蛋白类转换)可在所述动物中发生。
[0242]
在一些实施方案中,所述ighv基因、ighd基因和/或ighj基因可操作地连接在一起。vdj重组可发生在这些基因之间,并产生功能性抗体。在一些实施方案中,这些基因以与人重链免疫球蛋白基因座中的顺序相似的顺序排列。这种排列提供了各种有利方面,例如,这些基因的排列允许产生具有与人中的重链可变结构域的多样性非常相似的多样性的重链可变结构域。由于一些随机序列可在vdj重组过程中被插入到序列中,因此在一些实施方案中,无修饰或只有最小修饰的完整人抗体谱系可以降低非人序列在vdj重组过程中被插入的可能性。
[0243]
在一些实施方案中,所述ighv基因、ighd基因和/或ighj基因与一个或多个选自ighm、ighδ、ighg3、ighg1、ighg2b、ighg2a、ighe和igha基因的基因(例如,全部基因)可操作地连接在一起。
[0244]
在一些实施方案中,所述动物包含在其内源重链免疫球蛋白基因座中的破坏。在一些实施方案中,所述动物的内源重链免疫球蛋白基因座中的破坏包括一个或多个内源ighv基因、一个或多个内源ighd基因和一个或多个内源ighj基因的缺失。
[0245]
在一些实施方案中,所述动物是小鼠。所述动物的内源重链免疫球蛋白基因座中的破坏包括至少或约1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、20个、30个、40个、50个、60个、70个、80个、90个、100个、110个、120个、130个、140个、150个、160个、170个、171个、172个、173个、174个、175个、176个、177个、178个、179个、180个、181个或182个小鼠ighv基
因(例如,如表4中所示的基因)的缺失。在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv1-86,ighv1-85,ighv1-84,ighv1-83,ighv1-82,ighv1-81,ighv1-80,ighv1-79,ighv1-78和ighv1-77的小鼠ighv基因的缺失。在一些实施方案中,所述小鼠仍然包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv1-86、ighv1-85、ighv1-84、ighv1-83、ighv1-82、ighv1-81、ighv1-80、ighv1-79、ighv1-78和ighv1-77(例如,ighv1-86)的小鼠ighv基因。
[0246]
在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv5-6、ighv5-5、ighv2-3、ighv6-1、ighv5-4、ighv5-3、ighv2-2、ighv5-2、ighv2-1和ighv5-1的小鼠ighv基因的缺失。在一些实施方案中,所述小鼠仍然包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighv5-6、ighv5-5、ighv2-3、ighv6-1、ighv5-4、ighv5-3、ighv2-2、ighv5-2、ighv2-1和ighv5-1的小鼠ighv基因的缺失。
[0247]
在一些实施方案中,所述动物的内源重链免疫球蛋白基因座中的破坏包括至少或约1个、2个、3个、4个、5个、6个、7个、8个、9个、10个、11个、12个、13个、14个、15个、16个、17个、18个、19个或20个小鼠ighd基因(例如,如表5所示的基因)的缺失。在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9或10个,所述基因选自ighd5-1、ighd3-1、ighd1-1、ighd6-1、ighd2-3、ighd2-7、ighd2-8、ighd5-6、ighd3-2和ighd4-1的小鼠ighd基因的缺失。在一些实施方案中,所述小鼠仍然包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自ighd5-1、ighd3-1、ighd1-1、ighd6-1、ighd2-3、ighd2-7、ighd2-8、ighd5-6、ighd3-2和ighd4-1的小鼠ighd基因。
[0248]
在一些实施方案中,所述破坏包括约或至少1、2、3或4个选自ighj1、ighj2、ighj3和ighj4的小鼠ighj基因的缺失。在一些实施方案中,所述小鼠仍包含约或至少1个、2个、3个或4个选自ighj1、ighj2、ighj3和ighj4的小鼠ighj基因。
[0249]
在一些实施方案中,所述动物的内源重链免疫球蛋白基因座中的破坏包括约或至少500kb、600kb、700kb、800kb、900kb、1000kb、1500kb、2000kb、2500kb或3000kb的内源序列的缺失。
[0250]
在一些实施方案中,所述删除的序列始于ighv1-86至ighj4,始于ghv1-85至ighj4,始于ighv1-84至ighj4,始于ighv1-83至ighj4,或者始于ighv1-82至ighj4(例如,始于ighv1-85至ighj4)。
[0251]
在一些实施方案中,所述动物包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个序列,所述序列与人重链免疫球蛋白基因座中的序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性。在一些实施方案中,所述序列的长度约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb、1000kb、1500kb、2000kb、2500kb、3000kb或3500kb。在一些实施方案中,该序列从人ighv(iii)-82开始至ighv1-2。在一些实施方案中,该序列从人ighv7-81开始至ighv1-2。在一些实施方案中,该序列从人ighv(ii)-1-1开始至ighvj6。在一些实施方案中,该序列从人ighv6-1开始至ighvj6。
[0252]
所述人ighv基因、人ighd基因和人ighj基因可操作地连接在一起,并且可进行vdj重排。在一些实施方案中,所述经修饰的小鼠具有完整的人ighv、ighd和ighj基因谱系(例
如,包括全部人ighv、ighd和ighj基因中的非假(non-pseudo)基因)。因此,所述经修饰的小鼠可产生完整的人抗体谱系。在一些实施方案中,在vdj重组后,表15中的一个ighv基因(例如ighv3-21或ighv3-74)构成了编码抗体重链可变区的序列。表15中的一个ighd基因构成了编码抗体重链可变区的序列。表15中的一个ighj基因构成了编码抗体重链可变区的序列。在一些实施方案中,ighv基因是ighv3-21或ighv3-74。
[0253]
在一些实施方案中,图70a和图70b中的一个ighv基因(例如,ighv3-30、ighv3-33、ighv4-39或ighv4-34)构成了编码抗体重链可变区的序列。图70c中的一个ighd基因(例如,ighd6-19)构成了编码抗体重链可变区的序列。图70d中的一个ighj基因(例如,ighj4或ighj6)构成了编码抗体重链可变区的序列。在一些实施方案中,图71a和图71b中的一个igkv基因(例如,igkv4-1、igkv1-33、igkv2-30)构成了编码抗体轻链可变区的序列。图71c中的一个igkj基因(例如,igkj1、igkj2或igkj4)构成了编码抗体轻链可变区的序列。
[0254]
此外,在一些情况下,整个小鼠ighv基因、ighd基因和ighj基因(例如,包括全部非假基因)被敲除,并且重链可变区将不具有由源自小鼠的序列编码的任何序列,从而使人中的免疫原性最小化。
[0255]
经遗传修饰的κ轻链免疫球蛋白基因座
[0256]
κ链免疫球蛋白基因座(也称为igk或免疫球蛋白κ基因座)是染色体(例如,人2号染色体)上包含人抗体(或免疫球蛋白)轻链基因的区域。类似地,免疫球蛋白轻链基因也可以进行一系列重排,导致成熟免疫球蛋白轻链核酸(例如,κ链)的产生。
[0257]
v区段(也称为igkv基因)和j区段(也称为igkj基因)的连接产生了连续的外显子,其编码整个轻链可变结构域。在未重排的dna中,v基因区段(或igkv基因簇)位于离c区相对较远的位置。j基因区段(或igkj基因簇)位于c区附近。v区段与j基因区段的连接也使v基因接近c区序列。重排的v区的j基因区段仅通过一个内含子与c区序列分开。为了产生完整的免疫球蛋白轻链信使rna,转录后通过rna剪接将v区外显子连接至c区序列。
[0258]
人轻链免疫球蛋白基因座位于人2号染色体上。表7列出了igkv基因及其在该基因座中的相对顺序。人的igkv基因有几个不同的组,包括igkv1基因(包括始于igkv1的全部igkv基因,也称为vκi)、igkv2基因(包括始于igkv2的全部igkv基因,也称为vκii)、igkv3基因(包括始于igkv3的全部igkv基因,也称为vκiii)、igkv4基因(包括始于igkv4的全部igkv基因,也称为vκiv)、igkv5基因(包括始于igkv5的全部igkv基因,也称为vκv)、igkv6基因(包括始于igkv6的全部igkv基因,也称为vκvi)和igkv7基因(包括始于igkv7的全部igkv基因,也称为vκvii)。
[0259]
人2号染色体中的这些igkv基因也形成两个簇,近端的vκ簇和远端的vκ簇(图28)。两个簇中的序列相似,但不完全相同。这种序列的大区段复制发生在人谱系从与其它大型类人猿最近的共同祖先开始分化的时候。图64概述了每个簇中的相关igvk基因。
[0260]
表7.人2号染色体上的igkv基因列表
[0261][0262][0263]
表8列出了全部igkj基因及其在人2号染色体上的相对顺序。编码轻链免疫球蛋白恒定结构域的免疫球蛋白κ恒定区(igkc)基因位于igkv和igkj基因之后。这些基因和这些基因的顺序也示于图39和图43中。
[0264]
表8.人2号染色体上的igkj基因列表
[0265]
基因名称顺序基因名称顺序igkj177igkj480igkj278igkj581igkj379
ꢀꢀ
[0266]
小鼠轻链免疫球蛋白基因座位于小鼠6号染色体上。表9列出了igkv基因及其在该基因座中的相对顺序。
[0267]
表9.小鼠染色体6上的igkv基因列表
[0268][0269][0270]
gm9728和amd-ps2也位于该基因座中。gm9728的相对顺序是4,amd-ps2的相对顺序是134。表10列出了全部igkj基因及其在小鼠6号染色体上的相对顺序。编码轻链免疫球蛋白恒定结构域的igkc基因位于igkv和igkj基因之后。这些基因和这些基因的顺序也示于图40和图44中。
[0271]
表10.小鼠6号染色体上的igkj基因列表
133、igkv1-132、igkv1-131、igkv14-130和igkv9-129的小鼠igkv基因。
[0282]
在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自igkv3-10、igkv3-9、igkv3-8、igkv3-7、igkv3-6、igkv3-5、igkv3-4、igkv3-3、igkv3-2和igkv3-1的小鼠igkv基因的缺失。在一些实施方案中,所述小鼠仍然包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个选自igkv3-10、igkv3-9、igkv3-8、igkv3-7、igkv3-6、igkv3-5、igkv3-4、igkv3-3、igkv3-2和igkv3-1的小鼠igkv基因。
[0283]
在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个或5个选自igkj1、igkj2、igkj3、igkj4和igkj5的小鼠igkj基因的缺失。在一些实施方案中,所述小鼠仍然包含约或至少1个、2个、3个、4个或5个选自igkj1,igkj2,igkj3,igkj4和igkj5的小鼠igkj基因(例如,igkj5)。
[0284]
在一些实施方案中,所述动物的内源κ轻链免疫球蛋白基因座中的破坏包含约或至少500kb、600kb、700kb、800kb、900kb、1000kb、1500kb、2000kb、2500kb、3000kb或3500kb的内源序列的缺失。
[0285]
在一些实施方案中,所述删除的序列从igkv2-137开始至igkj4,从igkv1-136开始至igkj4,从igkv1-135开始至igkj4,从igkv2-137开始至igkj5,从igkv1-136开始至igkj5,或者从igkv1-135开始至igkj5(例如,从igkv2-137至igkj5)。
[0286]
在一些实施方案中,所述动物包含约或至少1个、2个、3个、4个、5个、6个、7个、8个、9个或10个序列,所述序列与人轻链免疫球蛋白基因座中的序列具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性。在一些实施方案中,所述序列的长度约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb、1000kb、1500kb、2000kb、2500kb、3000kb或3500kb。
[0287]
在一些实施方案中,所述动物可具有一个、两个、三个、四个、五个、六个、七个、八个、九个或十个未修饰的人序列。在一些实施方案中,所述未修饰的人序列的长度约为或至少为10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、200kb、300kb、400kb、500kb、600kb、700kb、800kb、900kb、1000kb、1500kb、2000kb、2500kb、3000kb或3500kb。
[0288]
在一些实施方案中,所述序列从人igkv3d-7开始至igkj5。在一些实施方案中,所述序列从人igkv3d-7开始至igkj4。在一些实施方案中,所述序列从人igkv1d-8开始至igkj5。在一些实施方案中,所述序列从人igkv1d-8开始至igkj4。
[0289]
所述人igkv基因和人igkj基因可操作地连接在一起,并且可进行vj重排。在一些实施方案中,所述经修饰的小鼠具有完整的人igkv和igkj基因谱系(例如,包括全部人igkv和igkj基因中的非假(non-pseudo)基因)。因此,所述经修饰的小鼠可产生完整的人抗体谱系。在一些实施方案中,在vj重组后,表16中的一个igkv基因(例如,igkv1d-43、igkv1d-13、igkv1d-16或igkv1d-12)构成编码抗体轻链可变区的序列。一个人igkj基因构成编码抗体轻链可变区的序列。在一些实施方案中,igkv基因是igkv1d-43、igkv1d-13、igkv1d-16或igkv1d-12。此外,在一些情况下,整个小鼠igkv基因和igkj基因(均为非假基因)被敲除,轻链可变区将不具有任何由源自小鼠的序列编码的序列,从而使人中的免疫原性最小化。
[0290]
在一些实施方案中,所述人近侧vκ簇igkv基因包含在经修饰的染色体中。在一些实施方案中,所述人远端vκ簇igkv基因包含在经修饰的染色体中。在一些实施方案中,所述
人近侧vκ簇igkv基因和人远侧vκ簇igkv基因都包含在修饰的染色体中。
[0291]
经遗传修饰的λ轻链免疫球蛋白基因座
[0292]
λ链免疫球蛋白基因座(也称为igl或免疫球蛋白λ基因座)是染色体(例如,人22号染色体)上包含人抗体(或免疫球蛋白)轻链基因的区域。类似地,免疫球蛋白轻链基因也可进行一系列重排,导致成熟免疫球蛋白轻链核酸(例如,λ链)的产生。在健康的人个体中,血清中的总κ与λ比率约为2∶1(测量完整的完全抗体),或者如果测量游离轻链,所述比率则为1∶1.5。在小鼠中,总κ与λ的比率大致为9∶1。
[0293]
在一些实施方案中,所述动物包含人λ链免疫球蛋白基因座。
[0294]
在一些实施方案中,所述动物包含在其内源λ轻链免疫球蛋白基因座中的破坏。在一些实施方案中,所述动物的内源轻链免疫球蛋白基因座中的破坏包括一个或多个内源iglv基因、一个或多个内源iglj基因和/或一个或多个免疫球蛋白λ恒定(iglc)基因(例如,iglc1、iglc2、iglc3和iglc4)的缺失。
[0295]
所述小鼠λ轻链免疫球蛋白基因座(igl基因座)位于小鼠16号染色体上。表11列出了iglv、iglj和iglc基因及其在该基因座上的相对顺序。
[0296]
表11.小鼠igl基因座上的基因列表
[0297][0298]
所述动物的内源λ轻链免疫球蛋白基因座中的破坏包括至少或约1,2,3,4,5,6,7,8,9,10,11或12个小鼠iglv、iglj和iglc基因(例如,如表11所示的基因)的缺失。在一些实施方案中,所述缺失包括约或至少1个、2个、3个或4个选自iglc1、iglc2、iglc3和iglc4的小鼠igkc基因。在一些实施方案中,所述破坏包括约或至少1个、2个或3个选自iglv1、iglv2和iglv3的小鼠iglv基因的缺失。在一些实施方案中,所述破坏包括约或至少1个、2个、3个、4个或5个选自iglj1、iglj2、iglj3、iglj3p和iglj4的小鼠iglj基因的缺失。
[0299]
在一些实施方案中,所述动物的内源λ轻链免疫球蛋白基因座中的破坏包括约或至少10kb、20kb、30kb、40kb、50kb、60kb、70kb、80kb、90kb、100kb、110kb、120kb、130kb、140kb、150kb、160kb、170kb、180kb、190kb、200kb、210kb、220kb、230kb、240kb、250kb、260kb、270kb、280kb、290kb、300kb、350kb、400kb、450kb、500kb或1000kb的核苷酸的缺失。在一些实施方案中,所述动物的内源λ轻链免疫球蛋白基因中没有破坏。
[0300]
在一些实施方案中,所述删除的序列从iglv2开始至iglc1、从iglv3开始至iglc1或从iglj2开始至iglc1。
[0301]
经遗传修饰的动物
[0302]
一方面,本公开提供了经遗传修饰的非人动物,其包含人源化重链免疫球蛋白基
因座和/或人源化轻链免疫球蛋白基因座。在一些实施方案中,所述动物包含一个或多个人ighv基因、一个或多个人ighd基因、一个或多个人ighj基因、一个或多个人igkv基因和/或一个或多个人igkj基因。在一些实施方案中,这些基因位于内源免疫球蛋白基因座中。
[0303]
在一些实施方案中,所述动物包含人λ链免疫球蛋白基因座。在一些实施方案中,所述动物包含在动物的内源λ轻链免疫球蛋白基因座中的破坏。在一些实施方案中,所述动物在动物的内源λ轻链免疫球蛋白基因座中没有破坏。
[0304]
所述经遗传修饰的非人动物可以是各种动物,例如小鼠、大鼠、兔子、猪、牛(例如,母牛、公牛、水牛)、鹿、绵羊、山羊、鸡、猫、狗、雪貂、灵长类动物(例如,狨猴、恒河猴)。对于其中不容易获得合适的可经遗传修饰的胚胎干(es)细胞的非人动物,可采用其它方法制备包含遗传修饰的非人动物。此类方法包括,例如,修饰非es细胞基因组(例如,成纤维细胞或诱导多能细胞),并采用核移植将经修饰的基因组转移到合适的细胞,例如卵母细胞,并在合适的条件下在非人动物中孕育经修饰的细胞(例如,经修饰的卵母细胞),以形成胚胎。这些方法在本领域中是已知的,并且在例如nagy等人“manipulating the mouse embryo:a laboratory manual(第3版),”cold spring harbor laboratory press,2003(其通过引用整体并入本文)中进行了描述。因此,在各种实施方案中,人v、d和/或j区段可以可操作地连接至非人动物(例如,啮齿动物,小鼠、大鼠、仓鼠)恒定区基因序列。在b细胞发育过程中,这些重排的人v、d和/或j区段连接至非人动物免疫球蛋白恒定区。
[0305]
在一个方面,该动物是哺乳动物,例如,双足总科或鼠总科(muroidea)的哺乳动物。在一些实施方案中,所述转基因动物是啮齿动物。啮齿动物可选自小鼠、大鼠和仓鼠。在一些实施方案中,所述经遗传修饰的动物来自选自丽仓鼠科(calomyscidae)(例如,鼠样仓鼠)、仓鼠科(cricetidae)(例如,仓鼠、新世界大鼠和小鼠、田鼠)、鼠科(小鼠和大鼠、沙鼠、多刺小鼠(spiny mice)、鬚毛大鼠(crested rats))、马岛鼠科(nesomyidae)(攀爬小鼠(climbing mice),岩石小鼠(rock mice)、白尾大鼠、马达加斯加大鼠和小鼠)、刺山鼠科(platacanthomyidae)(例如多刺睡鼠(spiny dormice)),和齡鼠科(spalacidae)(例如,鼹鼠、竹鼠和鼢鼠)的科。在一些实施方案中,所述经遗传修饰的啮齿动物选自小鼠或大鼠(鼠科)、沙鼠、多刺小鼠和鬚毛大鼠。在一些实施方案中,所述非人动物是小鼠。
[0306]
在一些实施方案中,所述动物是c57背景的小鼠(例如,选自c57bl/a,c57bl/an,c57bl/grfa,c57bl/kalwn,c57bl/6,c57bl/6j,c57bl/6byj,c57bl/6nj,c57bl/10,c57bl/10scsn,c57bl/10cr和c57bl/ola的c57bl品系)。在一些实施方案中,所述小鼠是选自为129p1,129p2,129p3,129x1,129s1(例如,129s1/sv,129s1/svim),129s2,129s4,129s5,129s9/svevh,129s6(129/svevtac),129s7,129s8,129t1,129t2的品系的129品系。这些小鼠描述于,例如,festing等人,revised nomenclature for strain 129mice,mammalian genome 10:836(1999);auerbach等人,establishment and chimera analysis of 129/svev-and c57bl/6-derived mouse embryonic stem cell lines(2000),这两篇文献通过引用整体并入本文。在一些实施方案中,所述经遗传修饰的小鼠是129品系与c57bl/6品系的杂交品系。在一些实施方案中,所述经遗传修饰的小鼠是129品系的杂交品系或bl/6品系的杂交品系。在一些实施方案中,所述小鼠是balb品系,例如balb/c品系。在一些实施方案中,所述小鼠是balb品系和另一种品系的杂交品系。在一些实施方案中,小鼠来自杂交品系(例如,50%balb/c-50%12954/sv;或50%c57bl/6-50%129)。
[0307]
在一些实施方案中,所述动物是大鼠。大鼠可选自wistar大鼠、lea品系、sprague dawley品系、fischer品系、f344、f6和dark agouti。在一些实施方案中,所述大鼠品系是选自wistar、lea、sprague dawley、fischer、f344、f6和dark agouti的两种或更多种品系的杂交品系。
[0308]
所述动物可具有一种或多种其它遗传修饰和/或其它修饰,其适合于制造人源化动物的特定目的。
[0309]
包含内源非人免疫球蛋白基因座的修饰的经遗传修饰的非人动物。在一些实施方案中,所述修饰可包含编码人蛋白质的至少一部分(例如,与人重链可变结构域或轻链可变结构域序列具有至少10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、96%、97%、98%或99%同一性)的人核酸序列。尽管还提供了可包含本文所述修饰的经遗传修饰的细胞(例如,es细胞、体细胞),但在许多实施方案中,所述经遗传修饰的非人动物包含在动物种系中的内源基因座的修饰。
[0310]
经遗传修饰的动物可表达来自内源小鼠基因座的人源化抗体和/或嵌合抗体,其中一个或多个内源小鼠免疫球蛋白基因已经被人免疫球蛋白基因和/或核苷酸序列替换,所述核苷酸序列与人免疫球蛋白基因序列(例如,ighv、ighd、ighj、igkv和/或igkj基因)具有至少10%、20%、30%、40%、50%、60%、70%、80%、90%、95%、96%、97%、98%或99%同一性。在各种实施方案中,内源非人免疫球蛋白基因座被完整地或部分地修饰以包含人核酸序列。
[0311]
可对上述非人哺乳动物进行遗传、分子和行为分析。本公开还涉及由本公开提供的非人哺乳动物与相同基因型或其它基因型交配产生的后代。非人哺乳动物可以是本领域已知的任何非人动物,并且可用于本文所述的方法中。优选的非人哺乳动物是哺乳动物(例如,啮齿动物)。在一些实施方案中,非人哺乳动物是小鼠。
[0312]
本公开还提供了源自非人哺乳动物或其后代的细胞系或原代细胞培养物。基于细胞培养的模型可以通过例如以下方法制备。细胞培养物可通过从非人哺乳动物中分离而获得,或者细胞可从使用相同构建体和标准细胞转染技术建立的细胞培养物中获得。包含编码人或人源化免疫球蛋白的dna序列的遗传构建体的整合情况可通过多种方法检测。
[0313]
有许多分析方法可用于检测外源dna或对基因组dna的修饰,包括核酸水平上的方法(包括使用逆转录聚合酶链式反应(rt-pcr)或southern印迹和原位杂交的mrna定量方法)和蛋白质水平上的方法(包括组织化学、免疫印迹分析和体外结合研究)。另外,目标基因的表达水平可通过本领域技术人员熟知的elisa技术进行定量。许多标准分析方法可用于完成定量测量。例如,转录水平可使用rt-pcr和杂交方法来测量,所述杂交方法包括rna酶保护、southern印迹分析、rna点分析(rna dot analysis)(rnadot)分析。免疫组织化学染色、流式细胞术、蛋白质印迹分析也可用于评估人或人源化蛋白质的存在。
[0314]
抗体和抗原结合片段
[0315]
本公开提供了通过本文所述方法产生的抗体及其抗原结合片段(例如,人源化抗体或嵌合抗体)。
[0316]
一般来说,抗体(也称为免疫球蛋白)由两类多肽链(轻链和重链)组成。本公开的非限制性抗体可以是完整的、包含两条重链和两条轻链的四链免疫球蛋白抗体。抗体的重链可以是任何同种型(包括igm、igg、ige、iga或igd)或亚类(包括igg1、igg2、igg2a、igg2b、
igg3、igg4、ige1、ige2)等。轻链可以是κ轻链或λ轻链。抗体可包含两条相同的轻链拷贝和两条相同的重链拷贝。重链各自包含一个可变结构域(或可变区,v
h
)和多个恒定结构域(或恒定区),通过它们的恒定区内的二硫键彼此结合,形成抗体的“茎”。轻链各自包含一个可变结构域(或可变区,v
l
)和一个恒定结构域(或恒定区),每条轻链通过二硫键与一条重链结合。每条轻链的可变区与其所结合的重链的可变区对齐。轻链和重链的可变区均包含三个夹在更保守的框架区(fr)之间的高变区。
[0317]
这些高变区(被称为互补决定区(cdr)形成包含抗体的主要抗原结合表面的环。四个框架区主要采用β-折叠构象,cdr形成连接β-折叠结构的环,在一些情况下形成β-折叠结构的一部分。每条链中的cdr被框架区域保持在非常接近的位置,并且与来自另一条链的cdr一起,促成抗原结合区域的形成。
[0318]
通过分析抗体的氨基酸序列来鉴定抗体的cdr区的方法是众所周知的,并且通常使用多种cdr的定义方式。kabat定义基于序列可变性,而chothia定义基于结构环区域的位置。这些方法和定义描述于,例如,martin,
″
protein sequence and structure analysis of antibody variable domains,
″
antibody engineering,springer berlin heidelberg,2001.422-439;abhinandan等人
″
analysis and improvements to kabat and structurally correct numbering of antibody variable domains,
″
molecular immunology 45.14(2008):3832-3839;wu,t.t.和kabat,e.a.(1970)j.exp.med.132:211-250;martin等人,methods enzymol.203:121-53(1991);morea等人,biophys chem.68(1-3):9-16(oct.1997);morea等人,j mol biol.275(2):269-94(jan.1998);chothia等人,nature 342(6252):877-83(dec.1989);ponomarenko and bourne,bmc structural biology 7:64(2007);;其中每一篇都通过引用整体并入本文。
[0319]
cdr对识别抗原的表位很重要。如本文中所用,“表位”是能够被抗体的抗原结合结构域特异性结合的靶分子的最小部分。表位的最小尺寸可以是约三个、四个、五个、六个或七个氨基酸,但这些氨基酸不需要在抗原一级结构的连续线性序列中,因为表位可能取决于基于抗原的二级和三级结构的抗原三维构型。
[0320]
在一些实施方案中,抗体是完整的免疫球蛋白分子(例如,igg1、igg2a、igg2b、igg3、igg4、igm、igd、ige、iga)。igg亚类(igg1、igg2、igg3和igg4)是高度保守的,相异在于它们的恒定区,特别是它们的铰链和上部ch2结构域。igg亚类的序列和差异在本领域中是已知的,并且描述于,例如,vidarsson等人,
″
igg subclasses and allotypes:from structure to effector functions.
″
frontiers in immunology 5(2014);irani等人
″
molecular properties of human igg subclasses and their implications for designing therapeutic monoclonal antibodies against infectious diseases.
″
molecular immunology 67.2(2015):171-182;shakib,farouk,编辑the human igg subclasses:molecular analysis of structure,function and regulation.elsevier,2016;其中每一篇都通过引用整体并入本文。
[0321]
抗体还可以是源自任何物种(例如,人、啮齿动物、小鼠、大鼠、骆驼)的免疫球蛋白分子。本文公开的抗体还包括但不限于多克隆抗体、单克隆抗体、单特异性抗体、多特异性抗体以及包含与另一种多肽融合的免疫球蛋白结合结构域的嵌合抗体。术语“抗原结合结构域”或“抗原结合片段”是保留完整抗体的特异性结合活性的抗体的一部分,即,能够与完
整抗体的靶分子上的表位特异性结合的抗体的任何部分。其包括例如fab、fab’、f(ab’)2和这些片段的变体。因此,在一些实施方案中,抗体或其抗原结合片段可以是,例如,scfv、fv、fd、dab、双特异性抗体、双特异性scfv、双抗体、线性抗体、单链抗体分子、由抗体片段形成的多特异性抗体,以及包括为抗体结合结构域或与抗体结合结构域同源的结合结构域的任何多肽。抗原结合结构域的非限制性实例包括,例如,完整抗体的重链和/或轻链cdr、完整抗体的重链和/或轻链可变区、完整抗体的全长重链或轻链、或来自完整抗体的重链或轻链的单个cdr。
[0322]
在一些实施方案中,抗原结合片段可形成嵌合抗原受体(car)的一部分。在一些实施方案中,嵌合抗原受体是如本文所述的单链可变区片段(scfv)与cd3-ζ的跨膜结构域和胞内域融合的融合体。
[0323]
在一些实施方案中,scfv具有一个重链可变结构域和一个轻链可变结构域。在一些实施方案中,scfv具有两个重链可变结构域和两个轻链可变结构域。在一些实施方案中,scfv具有两个抗原结合区,所述两个抗原结合区可与各自的靶抗原结合。
[0324]
通过本文所述方法产生的抗体及其抗原结合片段(例如,人源化抗体或嵌合抗体)具有各种有利方面。在一些实施方案中,不需要进一步优化来获得期望的性质(例如,结合亲和力、热稳定性和/或有限聚集)。
[0325]
在一些实施方式中,所述抗体(或其抗原结合片段)以小于0.1s-1
、小于0.01s-1
、小于0.001s-1
、小于0.0001s-1
或小于0.00001s-1
的解离速率(koff)与靶特异性结合。在一些实施方案中,解离速率(koff)大于0.01s-1
、大于0.001s-1
、大于0.0001s-1
、大于0.00001s-1
或大于0.000001s-1
。
[0326]
在一些实施方案中,动力学缔合速率(kon)大于1 x 102/ms、大于1 x 103/ms、大于1 x 104/ms、大于1 x 105/ms或大于1 x 106/ms。在一些实施方案中,动力学缔合速率(kon)小于1 x 105/ms、小于1 x 106/ms或小于1 x 107/ms。
[0327]
亲和力可从动力学速率常数的商(kd=koff/kon)中推导出来。在一些实施方案中,kd小于1 x 10-6
m、小于1 x 10-7
m、小于1 x 10-8
m、小于1 x 10-9
m或小于1 x 10-10
m。在一些实施方案中,kd小于50nm、40nm、30nm、20nm、15nm、10nm、9nm、8nm、7nm、6nm、5nm、4nm、3nm、2nm或1nm。在一些实施方案中,kd大于1 x 10-7
m、大于1 x 10-8
m、大于1 x 10-9
m、大于1 x 10-10
m、大于1 x 10-11
m或大于1 x 10-12
m。在一些实施方案中,所述抗体以小于或等于约0.9nm、0.8nm、0.7nm、0.6nm、0.5nm、0.4nm、0.3nm、0.2nm或0.1nm的kd与靶标结合。
[0328]
在一些实施方案中,测定了热稳定性。本文所述的抗体或抗原结合片段的tm可高于60℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃。
[0329]
由于igg可被描述为多结构域蛋白质,所以熔解曲线有时显示两个转变,或三个转变,以及第一变性温度tm d1、第二变性温度tm d2和任选的第三变性温度tm d3。
[0330]
在一些实施方案中,本文所述的抗体或抗原结合片段的tm d1高于60℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃。在一些实施方案中,本文所述的抗体或抗原结合片段的tm d2高于60
℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃。在一些实施方案中,本文所述的抗体或抗原结合片段的tm d3高于60℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃。
[0331]
在一些实施方案中,tm、tm d1、tm d2、tm d3低于60℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃。
[0332]
在一些实施方案中,当温度低于60℃、61℃、62℃、63℃、64℃、65℃、66℃、67℃、68℃、69℃、70℃、71℃、72℃、73℃、74℃、75℃、76℃、77℃、78℃、79℃、80℃、81℃、82℃、83℃、84℃、85℃、86℃、87℃、88℃、89℃、90℃、91℃、92℃、93℃、94℃或95℃时,本文所述的抗体或抗原结合片段不形成聚集。
[0333]
产生经遗传修饰的动物的方法
[0334]
所述经遗传修饰的动物可通过将人免疫球蛋白基因引入非人动物的基因组中以生产能够表达人源化抗体或嵌合抗体的动物来产生。图1a显示了产生人源化动物的方法。在一些实施方案中,所述方法首先涉及修饰人染色体上的人免疫球蛋白基因座。然后将经修饰的人染色体引入小鼠受体细胞。然后通过直接替换将人免疫球蛋白可变区引入小鼠基因组的相应区域。然后,筛选受体细胞。在一些实施方案中,细胞不包含人染色体。然后将细胞注射到囊胚中,以制备嵌合小鼠。可以进行后续育种以获得含有完整人源化免疫球蛋白基因座的小鼠。
[0335]
其它几种技术也可用于产生经遗传修饰的动物,包括例如非同源末端接合(nhej)、同源重组(hr)、锌指核酸酶(zfn)、基于转录激活因子样效应子的核酸酶(talen)和规律成族间隔短回文重复序列(crispr)-cas系统。在一些实施方案中,使用同源重组。在一些实施方案中,将crispr-cas9基因组编辑用于产生经遗传修饰的动物。这些基因组编辑技术中的许多基因组编辑技术在本领域中是已知的,并且描述于例如yin等人,
″
delivery technologies for genome editing,
″
nature reviews drug discovery 16.6(2017):387-399,该文献通过引用整体并入本文。还提供了许多其它方法,所述方法可用于基因组编辑,例如,将经遗传修饰的细胞核微注射到去核卵母细胞中,并将去核卵母细胞与另一种经遗传修饰的细胞融合。
[0336]
遗传修饰过程可涉及通过同源重组用人序列替换内源序列。在一些实施方案中,靶位点上游和下游的切割(例如,通过锌指核酸酶、talen或crispr)可导致dna双链断裂,并将同源重组用于将人序列替换内源序列。
[0337]
在一些实施方案中,用于产生经遗传修饰的人源化动物的方法可包括在内源基因座(或位点)处用人序列的相应区域替换核酸(例如,v、d、j区域或v、j区域)的步骤。该序列可包括ighv、ighd、ighj、igkv和/或igkj基因的区域(例如,一部分或整个区域)。在一些实施方案中,替换由同源重组介导。在一些实施方案中,替换由cre重组酶介导。
[0338]
图9显示了用于将功能性遗传元件添加到人染色体中的打靶策略。可在v区的上
游、j区与c区之间插入这些载体。
[0339]
在一些实施方案中,第一载体从5’至3’具有一个或多个下列序列:位于插入位点上游的dna同源臂序列、pgk启动子、红色荧光蛋白报告基因(tdtomato)、fmdv(口蹄疫病毒)自裂解肽(2a)、zeomycin抗性基因(zeo)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、loxp识别序列、潮霉素抗性基因(潮霉素磷酸转移酶的部分序列;“3’hygr”)和flp识别靶标(“frt”)、下游dna同源臂序列和dta基因。
[0340]
第二载体从5’至3’具有一个或多个下列序列:位于插入位点上游的dna同源臂序列、loxp识别序列、pgk启动子、嘌呤霉素抗性基因的部分序列(5’puror)、来自人延伸因子1α的哺乳动物表达启动子(ef-1a)、piggybac转座酶基因序列(pbase)、内部核糖体进入位点(ires)、卡那霉素抗性基因序列(neo)、转录终止/多聚腺苷酸化信号序列、位于插入位点下游的dna同源臂序列和dta。
[0341]
可将这些载体整合到细胞的基因组中,并且可通过药物抗性标志物或其组合(例如,博莱霉素(zeocin)、g418和/或嘌呤霉素)来选择细胞。在一些实施方案中,表达了转座酶,并且转座酶靶序列之间的遗传元件可以被删除。
[0342]
在一些实施方案中,这些载体被整合到已被修饰的人染色体中。在第一载体和第二载体被整合到基因组中之前,可以首先修改人染色体。在一些实施方案中,可根据需要在染色体的不同位置添加一个或多个另外的载体。在一些实施方案中,在c区与着丝粒(动粒)之间添加载体。第三载体从5’至3’可具有以下的一个或多个:插入位点上游的dna同源臂序列、pgk启动子、嘌呤霉素抗性基因序列(puror)、胸苷激酶基因序列(tk)、loxp识别序列、pgk启动子、嘌呤霉素抗性基因部分序列(5’puror)、哺乳动物表达启动子(ef-1a)、pbase、ires、neo、转录终止/多腺苷酸化信号序列、插入位点下游的dna同源臂序列和dta。在一些实施方案中,可将这些载体插入可变基因区或恒定区。在一些实施方案中,可删除内源可变基因区或内源恒定区的一部分。在一些实施方案中,可删除染色体的大片段(例如,在恒定区与着丝粒之间)。也可以用cre酶处理细胞,导致loxp位点的重组,从而去除人14号染色体上j区与着丝粒之间或人14号染色体上c区与着丝粒之间的基因组dna序列。在一些实施方案中,可发生自发的染色体断裂。可选择具有所需染色体断裂的经修饰的人染色体进行实验。
[0343]
可从人细胞系、癌细胞、原代细胞培养物和/或人成纤维细胞中获得人染色体。在一些实施方案中,将第一载体引入人细胞,然后将所述细胞与受体细胞融合。然后将经修饰的染色体分离并引入到另一种合适的受体细胞中。选择具有所需抗性的细胞以获得仅包含一条人染色体的细胞。然后,将第二个载体引入细胞,通过抗性选择细胞。然后,如果需要,可以引入第三载体和/或第四载体。受体细胞可以是哺乳动物细胞、人细胞或小鼠细胞。在一些实施方案中,受体细胞是cho细胞,或优选为a9细胞。在一些实施方案中,用荧光标记经修饰的染色体并分离所述染色体。并且通过染色体显微注射将经修饰的染色体注射到受体细胞中。在一些实施方案中,诱导供体细胞多核化它们的染色体。然后,迫使这些细胞核穿过细胞膜,以形成微细胞,可将所述微细胞与受体细胞融合。在一些实施方案中,还可使用微细胞介导的染色体转移。染色体操作技术描述于例如cn1200014a、cn109837307a、us20120093785a1和us2009253902;kuroiwa等人
″
manipulation of human minichromosomes to carry greater than megabase-sized chromosome inserts.
″
pb转座子序列(pb3’)、嘌呤霉素抗性基因部分序列(3’puror)、fmdv自裂解肽(2a)、dt受体(dtr)、cre重组酶的loxp识别序列、插入位点下游的dna同源臂序列以及dta。
[0350]
小鼠免疫球蛋白可变区可通过置换(例如,同源重组或cre介导的重组)被人免疫球蛋白可变区替换。在一些实施方案中,cre重组可用于介导置换。在一些实施方案中,载体可将loxp识别序列添加到人染色体中。可对小鼠染色体进行类似的修饰,其中可将两个loxp识别序列添加到染色体上。例如,cre重组酶可介导用人染色体上的v、j区置换小鼠染色体上的v、j区,或者用人染色体上的v、d、j区置换小鼠染色体上的v、d、j区。
[0351]
可进一步筛选细胞中不具有人染色体的细胞(例如,通过dt)。在一些情况下,未经dt筛选的细胞可包含重组人染色体片段,但这些片段很小,在小鼠细胞中不稳定(例如,shinohara等人(2000)chromosome research,8:713-725),并且在细胞增殖过程中会自然消失。在一些实施方案中,经修饰的人染色体的大片段被删除,例如通过cre介导的删除或通过自发的染色体断裂。
[0352]5’
末端同源臂和/或3’末端同源臂可具有期望的长度以促进同源重组。在一些实施方案中,同源臂约为或至少为1kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、10kb、20kb、30kb、40kb或50kb(例如,约3kb)。在一些实施方案中,同源臂小于1kb、2kb、3kb、4kb、5kb、6kb、7kb、8kb、9kb、10kb、20kb、30kb、40kb或50kb。
[0353]
在一些实施方案中,载体还可任选地包括报告蛋白,例如萤光素酶(例如,gluc)或荧光蛋白(例如,egfp、bfp等)。
[0354]
这些修饰可在不同的细胞中进行。在一些实施方案中,细胞是干细胞、胚胎干细胞或受精卵细胞。
[0355]
本公开还提供了建立人源化动物模型的方法,其包括以下步骤:
[0356]
(a)基于本文所述的方法提供细胞(例如受精卵细胞);
[0357]
(b)在液体培养基中培养细胞;
[0358]
(c)将培养的细胞移植到受者雌性非人哺乳动物的输卵管或子宫中,使细胞在雌性非人哺乳动物的子宫中发育;
[0359]
(d)在步骤(c)中鉴定怀孕雌性的后代经遗传修饰的人源化非人哺乳动物中的种系传播。
[0360]
在一些实施方案中,前述方法中的非人哺乳动物是小鼠(例如,c57小鼠、balb/c小鼠或c57bl/6小鼠)。
[0361]
在一些实施方案中,步骤(c)中的非人哺乳动物是伪怀孕(或假怀孕)的雌性动物。
[0362]
在一些实施方案中,用于上述方法的受精卵是c57bl/6受精卵。也可用于本文所述方法的其它受精卵包括但不限于fvb/n受精卵、balb/c受精卵、dba/1受精卵和dba/2受精卵。
[0363]
受精卵可来自任何非人动物,例如本文所述的任何非人动物。在一些实施方案中,受精卵细胞来自啮齿动物。可通过dna的显微注射将基因构建体引入受精卵。例如,通过在显微注射后培养受精卵,可将培养的受精卵转移到假怀孕的非人动物中,然后所述动物生产非人哺乳动物,从而产生上述方法中提及的非人哺乳动物。
[0364]
还提供了包含本文所述核苷酸序列的细胞、组织和动物(例如,小鼠),以及表达来自内源非人基因座的人源化或嵌合抗体的细胞、组织和动物(例如,小鼠)。
[0365]
本公开还提供了各种靶向载体(例如,用于产生经遗传修饰动物的载体)。在一些实施方案中,载体可包含:a)与待改变区域的5’末端同源的dna片段(5’同源臂);b)包含所需遗传元件(例如,loxp识别位点、抗药性基因和/或报告基因等)的序列;和c)与待改变的区域的3’末端同源的第二dna片段(3’同源臂)。本公开还涉及包含本文所述靶向载体的细胞。
[0366]
在一些实施方案中,所述细胞中的基因是杂合的。在一些实施方案中,所述细胞中的基因是纯合的。
[0367]
在一些实施方案中,所述非人哺乳动物细胞是小鼠细胞。在一些实施方案中,所述细胞是受精卵细胞。
[0368]
本公开还涉及产生具有两个或更多个人或嵌合基因的经遗传修饰的动物模型的方法。所述动物可包含一个或多个人或人源化免疫球蛋白基因座和编码另外的人或嵌合蛋白的序列。在一些实施方案中,所述另外的人或嵌合蛋白可以是程序性细胞死亡蛋白1(pd-1)、细胞毒性t淋巴细胞相关蛋白4(ctla-4)、淋巴细胞活化蛋白3(lag-3)、b和t淋巴细胞相关蛋白(btla)、程序性细胞死亡蛋白1配体1(pd-l1)、cd27、cd28、cd47、cd137、cd154、具有ig和itim结构域的t细胞免疫受体(tigit)、t细胞免疫球蛋白粘蛋白结构域3(t-cell immunoglobulin and mucin-domain containing-3)(tim-3)、糖皮质激素诱导的tnfr相关蛋白(gitr)或tnf受体超家族成员4(tnfrsf4或ox40)。
[0369]
产生具有另外的人或嵌合基因(例如,人源化基因)的经遗传修饰动的物模型的方法可包括以下步骤:
[0370]
(a)使用本文所述的方法获得经遗传修饰的非人动物;
[0371]
(b)将所述经遗传修饰的非人动物与另一种经遗传修饰的非人动物交配,然后筛选后代以获得具有两个或更多个人或嵌合基因的经遗传修饰的非人动物。
[0372]
在一些实施方案中,在该方法的步骤(b)中,可将所述经遗传修饰的动物与具有人或嵌合pd-1、ctla-4、lag-3、btla、pd-l1、cd27、cd28、cd47、cd137、cd154、tigit、tim-3、gitr、sirpa或ox40的经遗传修饰的非人动物交配。例如,在pct/cn2017/090320、pct/cn2017/099577、pct/cn2017/099575、pct/cn2017/099576、pct/cn2017/099574、pct/cn2017/106024、pct/cn2017/110494、pct/cn2017/110435、pct/cn2017/120388、pct/cn2018/081628、pct/cn2018/081629(其中每一篇都通过引用整体并入本文)中描述了这些经遗传修饰的非人动物中的一些。
[0373]
在一些实施方案中,所述经遗传修饰的动物可具有人adam6基因、内源adam6基因或经修饰的adam6基因。adam6蛋白是adam蛋白家族的一员,其中adam是解整联蛋白和金属蛋白酶的缩写。通常在人ighv基因ighv 1-2与ighv 6-1之间发现的人adam6基因是假基因(图37)。在小鼠中,存在两个adam6基因,adam6a和adam6b。它们位于小鼠ighv与ighd基因簇之间的基因间区域。小鼠adam6a位于小鼠ighv5-1与小鼠ighd5-1之间。小鼠adam6b位于小鼠ighd3-1与小鼠ighd1-1之间。因此,在一些实施方案中,所述经遗传修饰的动物可具有人adam6基因。在一些实施方案中,所述经遗传修饰的动物不具有内源adam6基因。
[0374]
在一些实施方案中,所述所述经遗传修饰的动物是小鼠。在一些实施方案中,所述小鼠被修饰成包括编码adam6蛋白(例如,adam6a或adam6b)的核苷酸序列。在一些实施方案中,所述序列被放置在任何合适的位置。其可被置于基因间区域,或者基因组中任何合适的
位置。在一些实施方案中,所述核酸编码与小鼠adam6a基因(例如,nc_000078.6的113539230-113547024;seq id no:53)或小鼠adam6b基因(例如,nc_000078.6的113486188-113492125;seq id no:54)具有至少80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或100%同一性的序列。在一些实施方案中,所述核酸另外包括adam6a基因和adam6b基因的调控元件(例如,启动子)。
[0375]
在一些实施方案中,功能性小鼠adam6基因座可置于人ighv基因簇的中间。在一些实施方案中,小鼠adam6基因座位于两个人ighv基因之间。在一些实施方案中,人vh1-2与人vh(ii)-1-1之间的人adam6假基因被小鼠adam6基因座替换。在一些实施方案中,在动物基因组中,adam6a基因和adam6b基因位于人ighv1-2与人vh(ii)-1-1之间。在一些实施方案中,小鼠adam6序列在人基因序列中的位置可接近人adam6假基因的位置,或者可接近小鼠adam6序列的位置(例如,在v-d基因间区域内)。在一些实施方案中,经遗传修饰的小鼠具有人源化重链免疫球蛋白基因座。在一些实施方案中,小鼠adam6a和小鼠adam6b位于人ighv1-2与ighv6-1基因之间。将小鼠adam6a和小鼠adam6b置于人ighv1-2与ighv6-1基因之间可有多种有利方面。例如,因为这些基因在相同的基因座替换了人adam6基因,所以人adam6基因的置换对vdj重组的影响有限,并且小鼠adam6a和小鼠adam6b基因也可以正常发挥作用(如在与内源基因座相似的位置中)。
[0376]
因此,在一个方面,本公开提供了经遗传修饰的动物,其在内源重链免疫球蛋白基因座包含含有一个或多个人ighv基因的第一序列;包含adam6基因的第二序列;以及包含一个或多个人ighd基因和一个或多个人ighj基因的第三序列。在一些实施方案中,所述第一序列、第二序列和第三序列是可操作连接的。
[0377]
在一些实施方案中,所述第一序列包含表1中除ighv2-10、ighv3-9、ighv1-8、ighv(ii)-1-1和ighv6-1外的全部人ighv基因。在一些实施方案中,所述第一序列包含表1中除ighv5-10-1和ighv3-64d、ighv(ii)-1-1和ighv6-1外的全部人ighv基因。在一些实施方案中,所述第一序列是源自人重链免疫球蛋白基因座的未修饰的序列。
[0378]
在一些实施方案中,所述第二序列包含小鼠adam6a基因和小鼠adam6b基因之一或两者。在一些实施方案中,所述动物是能育的雄性小鼠。在一些实施方案中,所述第二序列不具有小鼠adam6a基因或小鼠adam6b基因。
[0379]
在一些实施方案中,所述第三序列包含表2中的全部人ighd基因和表3中的全部人ighj基因。在一些实施方案中,所述第三序列包含人ighv6-1。在一些实施方案中,所述第三序列包含人ighv(ii)-1-1。在一些实施方案中,所述第三序列是源自人重链免疫球蛋白基因座的未修饰的序列。
[0380]
在一些实施方案中,amam6a和/或adam6b是内源序列。在一些实施方案中,amam6a和/或adam6b不被替换,和/或位于其内源或天然位置。在一些实施方案中,重链可变区基因座中小鼠ighv1-2之前的小鼠ighv基因被人ighv基因替换。在一些实施方案中,重链可变区基因座中小鼠ighv6-1之后的小鼠ighv、ighd和ighj基因被一个或多个人ighv基因、ighd和/或ighj基因替换。
[0381]
因此,在一些实施方案中,可用人ighv、ighd和ighj基因替换小鼠ighv、ighd和ighj基因一次以上。在第一步中,用人ighv基因替换在adam6a的5’侧选定数量的小鼠ighv
基因(例如,表4中的全部小鼠ighv基因)。在第二步中,用人ighd和人ighj基因替换adam6b的3’侧的选定数量的小鼠ighd和ighj基因(例如,表5中除ighd5-1和ighd3-1外的全部小鼠ighd基因和表6中的全部ighj基因)。置换可通过同源重组或cre介导的重组来进行。
[0382]
在一些实施方案中,所述小鼠不具有小鼠adam6a或adam6b基因。在一些实施方案中,所述小鼠具有人adam6基因。
[0383]
可使用各种方法来增强小鼠的生育力。在一些实施方案中,可将具有超数排卵的雌性小鼠用于交配。在一些实施方案中,可使用体外受精。可通过将血清促性腺激素和绒毛膜促性腺激素(例如,人或小鼠cg)注射到成熟的雌性小鼠中来诱导超数排卵。可处死成熟的雄性小鼠,分离其附睾尾。将附睾尾导的管切开以释放精子。接下来,可以处死超数排卵的成熟雌性小鼠并分离输卵管。卵丘-卵母细胞复合体(coc)可以从输卵管释放出来。接下来,可将精子悬浮液加入到coc并孵育以用于授精。可除去只含有一个原核的致病卵母细胞。孵育后,可将2细胞期的胚胎转移到受体雌性。增强小鼠生育力的方法是本领域已知的。
[0384]
本公开还提供了与本文所述的任何核苷酸序列具有至少1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%同一性的核酸序列,以及与本文所述任何氨基酸序列具有至少1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%同一性的氨基酸序列。
[0385]
在一些实施方案中,本公开涉及编码本文所述任何肽的核苷酸序列,或由本文所述任何核苷酸序列编码的任何氨基酸序列。在一些实施方案中,所述核酸序列少于10个、20个、30个、40个、50个、60个、70个、80个、90个、100个、110个、120个、130个、150个、200个、250个、300个、350个、400个、500个或600个核苷酸。在一些实施方案中,所述氨基酸序列少于5个、6个、7个、8个、9个、10个、20个、30个、40个、50个、60个、70个、80个、90个、100个、110个、120个、130个、140个、150个、160个、170个、180个、190个、200个、250个、300个、350个或400个氨基酸残基。
[0386]
在一些实施方案中,所述氨基酸序列(i)包含氨基酸序列;或者(ii)由氨基酸序列组成,其中所述氨基酸序列是本文所述的任一种序列。
[0387]
在一些实施方案中,所述核酸序列(i)包含核酸序列;或者(ii)由核酸序列组成,其中所述核酸序列是本文所述的任一种序列。
[0388]
为了确定两个氨基酸序列或两个核酸序列的同一性百分比,为了最佳比较的目的,对序列进行比对(例如,为了最佳比对,可以在第一和第二氨基酸或核酸序列之一或两者中引入缺口,并且为了比较的目的,可以忽略非同源序列)。出于比较目的而比对的参考序列的长度至少为参考序列长度的80%,在一些实施方案中至少为90%、95%或100%。然后比较相应氨基酸位置或核苷酸位置处的氨基酸残基或核苷酸。当第一序列中的位置被与第二序列中相应位置相同的氨基酸残基或核苷酸占据时,则该位置上的分子是相同的(如本文中所用,氨基酸或核酸的“同一性”相当于氨基酸或核酸的“同源性”)。在考虑了为了两个序列的最佳比对而需要引入的缺口的数量和每个缺口的长度的情况下,两个序列之间的同一性百分比是所述序列共有的相同位置的数量的函数。为了本发明的目的,序列的比较
和两个序列之间同一性百分比的确定可使用blossum 62评分矩阵,利用缺口罚分为12、缺口延伸罚分为4和移码缺口罚分为5来完成。
[0389]
具有相似物理化学性质的保守残基(例如亮氨酸和异亮氨酸)的百分比(同源性百分比)也可用于测量序列相似性。本领域已经定义了具有相似物理化学性质的氨基酸残基家族。这些家族包括例如具有碱性侧链(例如,赖氨酸、精氨酸、组氨酸)、酸性侧链(例如,天冬氨酸、谷氨酸)、不带电荷的极性侧链(例如,甘氨酸、天冬酰胺、谷氨酰胺、丝氨酸、苏氨酸、酪氨酸、半胱氨酸)、非极性侧链(例如,丙氨酸、缬氨酸、亮氨酸、异亮氨酸、脯氨酸、苯丙氨酸、甲硫氨酸、色氨酸)、β-支链侧链(例如,苏氨酸、缬氨酸、异亮氨酸)和芳香侧链(例如,酪氨酸、苯丙氨酸、色氨酸、组氨酸)的氨基酸。在许多情况下,同源性百分比高于同一性百分比。因此,本公开还提供了与本文所述的任何氨基酸序列具有1%、2%、3%、4%、5%、6%、7%、8%、9%、10%、15%、20%、25%、30%、35%、40%、45%、50%、55%、60%、65%、70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%同源性百分比的氨基酸序列,或编码这些氨基酸序列的核酸。
[0390]
使用经遗传修饰的动物的方法
[0391]
所述经遗传修饰的动物可用于产生能与靶标特异性结合的人源化或嵌合抗体。在一些实施方案中,可使用制备多克隆和单克隆抗体的标准技术,将靶标(例如,蛋白质或蛋白质的片段)用作免疫原以在这些动物中产生抗体。在一些实施方案中,将所述经遗传修饰的动物在允许所述动物产生特异于选定的抗原的抗体的条件下暴露于所述抗原一段时间。
[0392]
可通过多次注射(例如,皮下或腹膜内注射)抗原肽或蛋白质在动物体内产生多克隆抗体。在一些实施方案中,将抗原性肽或蛋白质与至少一种佐剂一起注射。在一些实施方案中,可将抗原性肽或蛋白质与在待免疫的物种中具有免疫原性的试剂缀合。可用抗原性肽或蛋白质注射动物超过一次(例如,两次、三次或四次)。
[0393]
可使用全长多肽或蛋白质,或者,可选地可将其抗原性肽片段用作免疫原。蛋白质的抗原性肽包含氨基酸序列的至少8个(例如,至少10个、15个、20个或30个)氨基酸残基,并含有蛋白质的表位,使得针对所述肽产生的抗体与蛋白质形成特异性免疫复合物。
[0394]
免疫原通常用于通过免疫合适的受试者(例如,本文所述的经遗传修饰动物)来制备抗体。合适的免疫原性制剂可包含例如重组表达的或化学合成的多肽(例如,蛋白质的片段)。所述制剂还可包含佐剂,诸如弗氏完全或不完全佐剂,或类似的免疫刺激剂。
[0395]
多克隆抗体可以如上所述通过用多肽或其抗原性肽(例如,蛋白质的部分)作为免疫原免疫合适的受试者来制备。可通过标准技术,诸如利用使用固定化多肽或肽的酶联免疫吸附测定(elisa)随时间推移监测免疫受试者中的抗体滴度。如果需要,可从哺乳动物(例如,从血液)中分离抗体分子,并通过公知的技术(诸如蛋白g色谱的蛋白a)进一步纯化以获得igg组分。在免疫后的适当时间,例如当特异性抗体滴度最高时,可从受试者获得产生抗体的细胞,并通过标准技术诸如最初由kohler等人(nature 256:495-497,1975)描述的杂交瘤技术、人b细胞杂交瘤技术(kozbor等人,immunol.today 4:72,1983)、ebv-杂交瘤技术(cole等人,monoclonal antibodies and cancer therapy,alan r.liss,inc.,第77-96页,1985)或三源杂交瘤技术将其用于制备单克隆抗体。产生杂交瘤的技术是公知的(通常参见,current protocols in immunology,1994,coligan等人(编辑),john wiley&sons,inc.,new york,ny)。产生单克隆抗体的杂交瘤细胞通过例如使用标准elisa测定法
筛选杂交瘤培养上清液的结合目标多肽或表位的抗体的方式来检测。
[0396]
一方面,本公开提供了包含内源免疫球蛋白重链基因座的修饰的小鼠,其中该小鼠产生包含可操作地连接至重链恒定区基因序列的重排免疫球蛋白序列的b细胞。在一些实施方案中,可操作地连接至所述重链恒定区基因序列的所述重排免疫球蛋白序列包含人重链v、d和/或j序列。在一些实施方案中,所述重链恒定区基因序列包含选自ch1、铰链、ch2、ch3及其组合的人或小鼠重链序列。
[0397]
一方面,本公开提供了包含内源免疫球蛋白轻链(例如,κ或λ)基因座的修饰的小鼠,其中该小鼠产生包含可操作地连接到轻链恒定区基因序列的重排免疫球蛋白序列的b细胞。在一些实施方案中,可操作地连接至所述轻链恒定区基因序列的所述重排免疫球蛋白序列包含人轻链v和/或j序列。在一些实施方案中,所述轻链恒定区基因序列包含人或小鼠轻链恒定区。
[0398]
所述小鼠b细胞或脾细胞可包含例如可操作地连接至小鼠免疫球蛋白恒定区基因的重排非小鼠免疫球蛋白可变基因序列。确定编码人重链可变区和人轻链可变区的序列。所述序列可通过例如对目标杂交瘤或b细胞进行测序来确定。在一些实施方案中,使用单个b细胞筛选。其可筛选天然抗体谱系而无需杂交瘤融合和组合展示。例如,可将b细胞与一小组dna条形编码的抗原混合,使得能够通过单细胞测序方案回收单个b细胞的一个或多个抗原条形码和b细胞受体(bcr)序列。
[0399]
可以例如通过将编码人重链可变区的序列可操作地连接至编码人重链恒定区的序列,和/或将编码人轻链可变区的序列可操作地连接至编码人轻链恒定区的序列,进一步修饰抗体以获得人源化抗体或人抗体。
[0400]
在一些实施方案中,如果小鼠表达与目标抗原非常相似的蛋白质,则可能难以在小鼠中引发免疫反应。这是因为在免疫细胞发育过程中,识别与自身来源肽结合的mhc分子的b细胞和t细胞被从免疫细胞的谱系中删除。在这些情况下,可进一步修饰人源化小鼠。可敲除小鼠中相应的基因,然后将小鼠暴露于目标抗原。因为小鼠对基因产物不进行阴性选择,所以小鼠可以产生容易与靶标特异性结合的抗体。
[0401]
本公开还提供了制备抗体、核酸、细胞、组织(例如,脾组织)的方法。在一些实施方案中,所述方法包括将本文所述的动物暴露于抗原。可从动物获得抗体(例如,嵌合抗体)、编码抗体的核酸、细胞和/或组织(例如,脾组织)。在一些实施方案中,编码人重链和轻链免疫球蛋白可变区的核酸可以例如通过测序来确定。在一些实施方案中,可将编码人重链免疫球蛋白可变区的核酸与编码人重链免疫球蛋白恒定区的核酸可操作地连接。在一些实施方案中,可将编码人轻链免疫球蛋白可变区的核酸与编码人轻链免疫球蛋白恒定区的核酸可操作地连接。在一些实施方案中,培养含有本文所述的核酸的细胞并收集抗体。
[0402]
在一些实施方案中,小鼠免疫球蛋白v、d、j基因(例如,小鼠ighv、ighd、ighj、igkv或igkj基因)对重链和/或轻链可变区序列没有贡献。在一些实施方案中,由动物产生的重链和/或轻链可变区序列完全是人的,并且完全由人免疫球蛋白v、d、j基因(例如,人ighv、ighd、ighj、igkv和igkj基因)贡献。
[0403]
本文所述的抗体或抗原结合片段的变体可通过将适当的核苷酸变化引入编码本文所述人抗体、人源化抗体或嵌合抗体或其抗原结合片段的dna中,或通过肽合成来制备。此类变体包括,例如,组成抗体的抗原结合部位或抗原结合结构域的氨基酸序列内的残基
的缺失、插入或取代。在此类变体群中,一些抗体或抗原结合片段对靶蛋白的亲和力增加。可进行缺失、插入和/或组合的任何组合,以获得对靶标具有增加的结合亲和力的抗体或其抗原结合片段。引入抗体或抗原结合片段的氨基酸变化也可改变抗体或抗原结合片段的翻译后修饰或将新的翻译后修饰引入抗体或抗原结合片段,诸如改变(例如,增加或减少)糖基化位点的数量,改变糖基化位点的类型(例如,改变氨基酸序列,使得在细胞内酶的作用下不同的糖被连接到糖基化位点),或引入新的糖基化位点。
[0404]
本文公开的抗体可源自任何动物物种,包括哺乳动物。天然抗体的非限制性实例包括源自人、灵长类动物(例如,猴子和猿)、牛、猪、马、绵羊、骆驼科动物(例如,骆驼和美洲驼)、鸡、山羊和啮齿动物(例如,大鼠、小鼠、仓鼠和兔子)(包括经遗传工程化以产生人抗体的转基因啮齿动物)的抗体。
[0405]
人抗体和人源化抗体包括具有源自人种系免疫球蛋白序列的可变区和恒定区或者具有与源自人种系免疫球蛋白序列的可变区和恒定区相同的氨基酸序列的抗体。人抗体可以例如在cdr中包括不由人种系免疫球蛋白序列编码的氨基酸残基(例如,通过体外随机或位点特异性诱变或体内体细胞突变引入的突变)。
[0406]
可对抗体或抗原结合片段进行另外的修饰。例如,可将一个或多个半胱氨酸残基引入fc区,从而允许在该区域中形成链间二硫键。如此产生的同二聚体抗体可在体外和/或体内具有任何延长的半衰期。还可使用异双功能交联剂制备在体外和/或体内具有延长的半衰期的同二聚体抗体,所述用异双功能交联剂制备抗体的方法描述于例如wolff等人(cancer res.53:2560-2565,1993)中。或者,抗体可被工程化成具有双fc区(参见,例如,stevenson等人,anti-cancerdrug design3:219-230,1989)。
[0407]
在一些实施方案中,可对抗体或其抗原结合片段进行共价修饰。这些共价修饰可通过化学或酶促合成,或者通过酶促或化学裂解来产生。通过使抗体或抗体片段的目标氨基酸残基与能够与选定的侧链或n或c末端残基反应的有机衍生剂反应,将抗体或抗体片段的其它类型的共价修饰引入分子。
[0408]
实施例
[0409]
在以下实施例中进一步描述了本发明,所述实施例不限制权利要求中描述的本发明的范围。
[0410]
实施例1:概述
[0411]
进行了将人免疫球蛋白基因引入小鼠基因组中以产生表达人源化抗体的小鼠的实验。图1a显示了产生人源化小鼠的方法。所述方法首先涉及修饰人染色体上的人免疫球蛋白区域。然后将经修饰的人染色体引入小鼠受体细胞。
[0412]
通过直接置换(例如,同源重组或cre介导的重组)用人免疫球蛋白可变区替换了小鼠免疫球蛋白可变区。在一些情况下,可通过逐步的方法将人免疫球蛋白可变区引入小鼠基因组。然后,筛选正确替换的受体细胞。然后将细胞注射到囊胚中以制备嵌合小鼠。随后进行育种以获得含有完整的人免疫球蛋白可变区的小鼠。
[0413]
因为小鼠重链基因和两个轻链基因分别位于12号、6号和16号染色体上,所以可以分别制备含有人重链可变区或人轻链可变区的小鼠(图1b和图1c)。然后可将这些小鼠相互交配以获得既能表达人重链可变区又能表达人轻链可变区的小鼠。
[0414]
实施例2:小鼠重链免疫球蛋白基因座的修饰
[0415]
重链免疫球蛋白基因座位于小鼠12号染色体上。图2是显示小鼠重链免疫球蛋白基因座的示意图。在重链免疫球蛋白基因座的可变区两侧引入两个重组位点(1301,1302),所得的经修饰的染色体如图3a-3b所示。其中一个是野生型loxp位点,另一个是异种特异性突变型lox位点(lox2272)。野生型loxp位点与异种特异性突变型lox位点之间不能发生重组。在小鼠胚胎干细胞中进行该修饰。打靶策略的概述如图4所示。载体(v1401)从5’至3’具有:插入位点上游的dna同源臂序列、flp识别靶标(frt)、cag启动子,潮霉素抗性基因(潮霉素磷酸转移酶的部分序列;“5’hygr”)、loxp(1301)、frt、5’pb转座子序列(pb5’)、pgk启动子、蓝色荧光蛋白报告基因(bfp)、fmdv自裂解肽(2a)、潮霉素抗性基因(潮霉素磷酸转移酶;hygr)、3’pb转座子序列(pb3’)、插入位点下游的dna同源臂序列和dta。
[0416]
载体(v1402)从5’至3’具有:插入位点上游的dna同源臂序列、5’pb转座子序列(pb5’)、pgk启动子、绿色荧光蛋白报告基因序列(egfp)、fmdv自裂解肽(2a)、嘌呤霉素抗性基因序列(puror)、3’pb转座子序列(pb3’)、flp识别靶标(frt)、嘌呤霉素抗性基因部分序列(3’puror)、fmdv自裂解肽(2a)、dt受体(dtr)、loxp识别序列(1302)、插入位点下游的dna同源臂序列和dta。
[0417]
将载体(v1401和v1402)引入小鼠胚胎干细胞。然后用潮霉素b和嘌呤霉素筛选细胞。通过pcr确认了外源基因整合到小鼠基因组中。结果如图5a-5b和图6a-6b所示。编号为030、035、036和037的克隆被确认为阳性。
[0418]
使用以下引物进行pcr测定:
[0419]
mighv-5’loxp-l-gt-f:5
’-
gccaaggaatttaaaaggggattgaaagcaa-3’(seq id no:1),
[0420]
mighv-005-l-gt-r2:5
’-
gccctccatgtacagcttcatgtgc-3’(seq id no:2);
[0421]
mighv-005-5’loxp-r-gt-f2∶5
’-
actgggcttgtcgagacagagaaag-3’(seq id no:3),
[0422]
mighv-5’loxp-r-gt-r:5
’-
ccacagcccgatctacttggctttt-3’(seq id no:4);
[0423]
mighv-3’lox-l-gt-f2:5
’-
gcaaggttttgactaagcggagcac-3’(seq id no:5);
[0424]
mighv-3’lox-l-gt-r2:5
’-
tgacgcatgtgttttatcggtctgt-3’(seq id no:6);
[0425]
mighv3’lox-r-gt-f2:5
’-
gtgcctgacacgtgctacgagattt-3’(seq id no:7);
[0426]
mighv-3’lox-r-gt-r1:5
’-
ttcaacaataagcagggccagaggg-3’(seq id no:8);
[0427]
在这些引物当中,mighv-5’loxp-l-gt-f和mighv-5’loxp-r-gt-r位于小鼠染色体上,mighv-005-l-gt-r2和mighv-005-5’loxp-r-gt-f2位于载体1401上,mighv-3’lox-l-gt-f2和mighv-3’lox-r-gt-r1位于小鼠染色体上,mighv-3’lox-l-gt-r2和mighv3’lox-r-gt-f2位于载体1402上。
[0428]
实施例3:修饰人14号染色体
[0429]
本实验的目的是产生至少有两个重组位点的经修饰的人染色体。这两个重组位点被引入重链免疫球蛋白基因座可变区的两侧。
[0430]
重链免疫球蛋白基因座位于人14号染色体上。图7是突出显示重链免疫球蛋白基因座的人14号染色体的示意图。
[0431]
经修饰的人可变区如图8所示。图9显示了打靶策略的概述。如图9所示,301和302位点是重组位点。重组位点1301和重组位点301是相同的。重组位点1302和重组位点302是
相同的。
[0432]
进行了将载体插入人14号染色体的v区上游以及j区与c区之间的实验。第一个靶向载体(v401)从5’至3’具有:插入位点上游的dna同源臂序列、pgk启动子、红色荧光蛋白报告基因(tdtomato)、来自fmdv(口蹄疫病毒)的自裂解肽(2a)、zeocin抗性基因序列(zeo)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、cre重组酶的loxp识别序列(301)、潮霉素抗性基因(潮霉素磷酸转移酶的部分序列;“3’hygr”)和flp识别靶标(“frt”)、下游dna同源臂序列和白喉毒素a亚单位(dta)基因。
[0433]
第二载体(v402)从5’到3’具有以下序列:插入位点上游的dna同源臂序列、cre重组酶的loxp识别序列(302)、pgk启动子、嘌呤霉素抗性基因的部分序列(5’puror)、来自人延伸因子1α的哺乳动物表达启动子(ef-1a)、piggybac转座酶基因序列(pbase)、内部核糖体进入位点(ires)、卡那霉素抗性基因序列(neo)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、插入位点下游的dna同源臂序列和dta。
[0434]
在一些实验中,将载体(v401,v402)引入细胞,并通过合适的药物抗性标记或其组合(zeocin,g418)选择细胞。
[0435]
有许多方法可以将目标载体引入人染色体。人染色体可获自人细胞系、癌细胞、原代细胞培养物和/或人成纤维细胞。在一个实验中,将第一个载体引入染色体。可将经修饰的染色体添加到受体细胞中,然后可将第二载体插入到经修饰的染色体中。在一些实验中,首先将v401引入人细胞,然后用荧光标记染色体,随后分离染色体,然后通过显微注射将经修饰的染色体注射到受体细胞中。然后将v402引入细胞。在另一个实验中,选择人成纤维细胞并引入载体402。然后将人成纤维细胞与受体细胞(a9细胞或cho细胞)融合。
[0436]
在一些实验中,可通过同源重组例如同时将一个或多个载体在所需位置插入人14号染色体。载体可包含抗药性标记(zeocin,g418),然后筛选细胞。标记染色体,然后分离,然后通过染色体显微注射将经修饰的染色体注射到受体细胞中。
[0437]
在一些实验中,插入一个或多个另外的载体。可以根据需要将这些另外的载体插入在人14号染色体的不同位置。在一个实验中,第三个载体从5’至3’可具有下列序列:插入位点上游的dna同源臂序列、pgk启动子、嘌呤霉素抗性基因序列(puror)、胸苷激酶基因序列(tk)、loxp识别序列(302)、pgk启动子、嘌呤霉素抗性基因部分序列(5’puror)、来自人延伸因子1α的哺乳动物表达启动子(ef-1a)、piggybac转座酶基因序列(pbase)、内部核糖体进入位点(ires)、卡那霉素抗性基因序列(neo)、转录终止/多聚腺苷酸化信号(polya;“pa”)、插入位点下游的dna同源臂序列和dta。将载体插入到c区内。
[0438]
在一个实验中,将第三个载体(v403)插入在c区与动粒之间。载体从5’至3’具有以下序列:插入位点上游的dna同源臂序列、pgk启动子、嘌呤霉素抗性基因序列(puror)、胸苷激酶基因序列(tk)、loxp识别序列(302)、pgk启动子、嘌呤霉素抗性基因部分序列(5’puror)、来自人延伸因子1α的哺乳动物表达启动子(ef-1a)、piggybac转座酶基因序列(pbase)、内部核糖体进入位点(ires)、卡那霉素抗性基因序列(neo)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、插入位点下游的dna同源臂序列和dta。
[0439]
在一个实验中,选择人成纤维细胞并引入载体402。然后将人成纤维细胞与受体细胞(a9细胞或cho细胞)融合。将经修饰的染色体分离并引入到另一个合适的受体细胞中。然后通过g418抗性选择细胞以获得仅包含一条人染色体的细胞。然后,将载体401引入细胞,
并通过对zeocin的抗性来选择细胞。之后,将载体403引入细胞中,并通过对嘌呤霉素的抗性来选择细胞。筛选后选择的阳性克隆用cre酶处理。染色体技术描述于例如kuroiwa等人
″
manipulation of human minichromosomes to carry greater than megabase-sized chromosome inserts.
″
nature biotechnology 18.10(2000)):1086-1090;cn1200014a;cn109837307a;us20120093785a1;us2009253902;cn1717483a;paulis,marianna.
″
chromosome transfer via cell fusion.
″
methods in molecular biology738(2011):57;genes,chromosomes&cancer 14:126127(1995);tomizuka等人
″
functional expression and germline atransmission of a human chromosome fragment in chimaeric mice.
″
nature genetics 16.2(1997):133-143;和somatic cell and molecular genetics,第13卷,第3期,1987,第279-284页,其中每一篇均通过引用整体并入本文。
[0440]
进行pcr以确认5
’-
末端重组位点301和3
’-
末端重组位点302存在于染色体上。没有随机插入的细胞通过southern印迹证实,并通过荧光原位杂交(fish)进行分析。
[0441]
图10显示了经修饰的人14号染色体。图11显示了染色体hchr14-mut3上loxp位点301的pcr鉴定结果。图12显示了染色体hchr14-mut3上loxp位点302的pcr鉴定结果。如图所示,12个克隆(编号为1-b2、1-b8、1-d6、1-d10、1-f11、1-g11、2-a2、3-e5、3-g5、3-h4、5-c3和6-f11)为阳性克隆。
[0442]
实验中使用了以下pcr引物:
[0443]
highv-5’loxp-l-gt-f1:5
’-
tcaaagtcaatttcctcagcgaggct-3’(seq id no:9),
[0444]
highv-5’loxp-r-gt-r:5
’-
agggagggaatggaatgagggtgat-3’(seq id no:10);
[0445]
highv-3’loxp-l-gt-f1:5
’-
ccatgtgacccattcgagtgtcctg-3’(seq id no:11),
[0446]
highv-3’loxp-r-gt-r:5
’-
ttgtgagggctcaagttcagtgcat-3’(seq id no:12)。
[0447]
使用阳性克隆和ccp14fish探针(cytotest inc.,rockville,md,目录号ct-ccp014)进行fish分析。克隆1-d10的表示性fish图像如图13和图14所示。在图13中,白色箭头表示人14号染色体(在修饰之前)的全长。在图14中,白色箭头表示经修饰的人14号染色体片段。
[0448]
实施例4:将人染色体或片段引入小鼠es细胞
[0449]
通过前述方法将实施例3中获得的经修饰的染色体引入实施例2中获得的细胞。然后通过g418筛选细胞。只选择仅含有一条人染色体的细胞。图15显示了经修饰的小鼠12号染色体。
[0450]
然后cre重组酶介导人染色体hchr14-mut3上的v、d、j区对小鼠染色体mchr12-mut2上的v、d、j区的替换(图16)。人染色体dna序列被重组位点1301与1302之间的序列替换。将潮霉素和嘌呤霉素用于筛选阳性细胞。在注射到小鼠囊胚中之前,通过dt进一步筛选细胞以获得不含人染色体的小鼠细胞。在一些情况下,将细胞直接注射到囊胚中,而无需进行dt筛选。
[0451]
对cre重组后的细胞进行测试,以确认人基因序列已被整合到小鼠基因组中。pcr测定的结果如图17、图18、图19和图20所示。所有的pcr测定的结果显示,编号为1-b4、1-b10和2-a7的细胞具有正确的重组,并且1-b10细胞中的人染色体消失。将鼠染色体全涂染探针(murine whole chromosome painting probes)(cytocell ltd,cambridge,uk;目录号
amp12r)和人特异性igh断裂分离探针(cytocell ltd,cambridge,uk;目录号lph 014)用于通过fish测试1-b10细胞。结果如图21所示,证实了人染色体片段存在于小鼠染色体中。这些引物如下表所示。
[0452]
表12
[0453][0454][0455]
实施例5:产生包含人源化重链免疫球蛋白基因座的小鼠
[0456]
通过显微注射将阳性克隆细胞注射到balb/c小鼠的囊胚中。根据例如a.nagy等人,“manipulating the mouse embryo:a laboratory manual(第3版),”cold spring harbor laboratory press,2003中描述的方法进行胚胎显微注射。然后将注射的受精卵转移到培养基中进行短时间培养,然后移植到受体小鼠的输卵管中以产生经遗传修饰的人源化小鼠(f0代)。然后将小鼠与背景为c57bl/6的小鼠交配。选择黑色后代与flp工具小鼠交配(图22)。对从小鼠尾部获得的dna进行pcr分析。将小鼠进一步与具有balb/c背景的小鼠杂交数次(例如,至少5次),以获得具有balb/c背景的人源化重链免疫球蛋白基因座杂合小鼠。
[0457]
为了证实小鼠表达人抗体重链,从嵌合鼠(f0代)和黑鼠(f1代)的小鼠中收集血液。提取rna,并将其反转录以获得cdna。使用下列pcr引物扩增序列,并对序列进行进一步测序。
[0458]
表13.pcr引物
[0459][0460]
实施例6:小鼠轻链免疫球蛋白基因座的修饰
[0461]
轻链免疫球蛋白基因座位于小鼠6号染色体上。图23是显示小鼠轻链免疫球蛋白基因座的示意图。在轻链免疫球蛋白基因座的可变区的两侧上引入两个重组位点,所得的经修饰的染色体如图24a-24b所示。详细的打靶策略如图25所示。
[0462]
在小鼠胚胎干细胞中进行该修饰。载体(v3901)从5’至3’具有:插入位点上游的dna同源臂序列、flp识别靶标(frt)、来自人延伸因子1α的哺乳动物表达启动子(ef-1a)、潮霉素抗性基因(潮霉素磷酸转移酶的部分序列;“5’hygr”)、cre重组酶的loxp识别序列(1101)、5’pb转座子序列(pb5’)、蓝色荧光蛋白报告基因(bfp)、dt受体(dtr)、fmdv自裂解肽(2a)、卡那霉素抗性基因序列(neo)、转录终止/多腺苷酸化信号序列(polya;“pa”)、3’pb转座子序列(pb3’)、嘌呤霉素抗性基因部分序列(3’puror)、fmdv自裂解肽(2a)、dt受体(dtr)、cre重组酶的loxp识别序列(1102)、插入位点下游的dna同源臂序列和dta。
[0463]
将载体(v3901)引入小鼠胚胎干细胞。通过相应的抗生素抗性基因标记或其组合来筛选细胞。通过pcr证实了载体v3901整合到小鼠基因组的正确位点。结果如图26-27所示。两个组合的pcr结果证实了编号为208、209、215、217和269的细胞是阳性克隆。
[0464]
使用以下引物进行pcr测定:
[0465]
igkv-005-c-5g-l-gt-f:5
′-
tcacacactacagcttccaccacaa-3
′
(seq id no:34);
[0466]
igkv-005-c-5g-l-gt-r2:5
′-
cggggaaaagtcgactctagaacgg-3
′
(seq id no:35);
[0467]
igkv-005-c-5g-r-gt-f1:5
′-
actgcattctagttgtggtttgtcca-3
′
(seq id no:36);
[0468]
igkv-005-c-5g-r-gt-r:5
′-
ggcctggaaaactcagctatccttt-3
′
(seq id no:37)。
[0469]
在这些引物中,igkv-005-c-5g-l-gt-f and igkv-005-c-5g-r-gt-r位于小鼠染色体上,igkv-005-c-5g-l-gt-r2 and igkv-005-c-5g-r-gt-f1位于载体v3901上。
[0470]
因此,两个重组位点被引入小鼠胚胎干细胞中的小鼠6号染色体中。
[0471]
实施例7:修饰人2号染色体
[0472]
人轻链免疫球蛋白基因座位于人2号染色体中。图28是突出显示轻链免疫球蛋白基因座的人2号染色体的示意图。
[0473]
在轻链免疫球蛋白基因座的可变区两侧引入了两个重组位点。将v
hk
与着丝粒之间的区域删除,以获得更短的人工染色体用于后续实验。在6号染色体上的小鼠免疫球蛋白基因座的可变区中引入了相似的重组位点。然后将人染色体引入小鼠受体细胞,获得人源化
轻链免疫球蛋白基因座。
[0474]
经修饰的人染色体2如图29所示。打靶策略如图30所示。载体(v2701)从5’至3’具有:插入位点上游的dna同源臂序列、pgk启动子、红色荧光蛋白报告基因序列(tdtomato)、fmdv自裂解肽(2a)、来自土曲霉(aspergillus terreus)的杀稻瘟菌素s脱氨酶(bsr)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、loxp识别序列2601、潮霉素抗性基因(潮霉素磷酸转移酶部分序列;“3’hygr”、flp识别靶标(frt)、插入位点下游的dna同源臂序列和白喉毒素a亚单位(dta)。
[0475]
载体(v2702)从5’至3’具有:插入位点上游的dna同源臂序列、loxp识别序列2602、pgk启动子、嘌呤霉素抗性基因序列的一部分(5’puror)、ef-1a、pbase、ires、卡那霉素抗性基因序列(neo)、转录终止/多聚腺苷酸化信号序列(polya;“pa”)、插入位点下游的dna同源臂序列和dta。
[0476]
通过测序验证了载体(v2702)的序列。通过转染将载体引入人h9细胞。然后通过g418和乌本苷抗性筛选细胞。通过pcr证实了基因整合到人基因组中。结果如图31所示。编号为01、02、03和04的克隆被确认为阳性克隆。
[0477]
使用以下引物进行pcr测定:
[0478]3’
l-l-gt-f:5
’-
aaggtgactctgcaatcagcctctg-3’(seq id no:38),
[0479]3’
l-l-gt-r1:5
’-
tcatctacagccacaacgtgagcag-3’(seq id no:39);
[0480]3’
l-r-gt-f1:5
’-
cccatgtacaggttccgcatgaact-3’(seq id no:40),
[0481]3’
l-r-gt-r:5
’-
ctccgtccgcttttatttcccctgt-3’(seq id no:41)。
[0482]
选择具有适于进一步实验的经修饰的染色体的细胞。通过染色体技术将经修饰的人染色体引入受体细胞。通过g418抗性对受体细胞a9细胞进行筛选。选择只包含一条人染色体的细胞用于进一步的基因编辑。
[0483]
在筛选过程中,图31中所示的编号为03的克隆具有正确的重组,并由人2号染色体计数探针(ccp2 fish探针)(cytotest inc.,rockville,md,目录号ct-ccp002)标记。结果证实在细胞中存在经修饰的人2号染色体(图32)。
[0484]
然后将载体(v2701)进一步引入到细胞中。通过g418和杀稻瘟菌素s抗性筛选细胞。重组得到了pcr的证实。引物如下表所示。结果表明5-c3、1-h2、1-h9和1-g5为阳性克隆(图33)。
[0485]
表14
[0486][0487]
实施例8:获得可产生人源化抗体的小鼠
[0488]
将小鼠胚胎干细胞和实施例7中获得的细胞融合,并将经修饰的人2号染色体引入实施例6中获得的小鼠胚胎干细胞。选择只含有一条经修饰的人2号染色体的小鼠es细胞,
在cre介导的重组(cre介导的重组)后筛选细胞,如图34所示。对cre重组后的细胞进行测试,以确认人基因序列已整合到小鼠基因组中。将小鼠染色体全涂染探针(cytocell ltd,cambridge,uk;目录号amp06g)和人特异性igk断裂分离探针(cytocellltd,cambridge,uk;目录号lph 034)用于通过荧光原位杂交来验证pcr确认的阳性克隆。结果显示在图35中,证实人染色体片段存在于小鼠染色体中。细胞被注入囊胚。可以获得含有人源化轻链免疫球蛋白基因座的小鼠。
[0489]
选择嵌合小鼠与c57bl/6小鼠交配,产生灰色和黑色后代(f1代)。对从黑色小鼠尾部获得的dna进行pcr分析,并选择阳性f1代小鼠与flp工具小鼠交配。图36显示了flp介导的重组的示意图。通过该方法制备的小鼠含有c57bl/6背景。具有不同背景的小鼠可具有不同的有利方面,并且通过本文的方法制备的杂合或纯合小鼠可用于通过回交(例如,balb/c小鼠具有体液免疫有利方面)几代来产生具有一些其它背景的小鼠,以获得具有期望背景的小鼠。
[0490]
选择几只小鼠,与balb/c小鼠杂交数次,获得具有balb/c背景的杂合小鼠。然后将杂合小鼠相互杂交以获得纯合子小鼠。
[0491]
将具有人源化轻链免疫球蛋白基因座的小鼠和具有人源化重链免疫球蛋白基因座的小鼠相互杂交,以获得同时具有人源化重链免疫球蛋白基因座和人源化轻链免疫球蛋白基因座的小鼠。
[0492]
实施例9:转基因小鼠中的b细胞发育
[0493]
进行实验以比较人源化小鼠和野生型小鼠的免疫系统。选择3只9-10周龄野生型(wt)、3只具有杂合的人源化重链免疫球蛋白基因座的小鼠和3只具有纯合的人源化重链免疫球蛋白基因座的小鼠。其中,与野生型小鼠相比,杂合小鼠和纯合小鼠具有相似的体重、外观和活力。获得这些小鼠的外周血、脾脏、淋巴结和骨髓组织,未发现明显的解剖变化(例如,三组小鼠的脾脏尺寸、形态和重量没有可观察到的差异)。进行流式细胞术以分析小鼠外周血、脾脏和淋巴结中的淋巴细胞群和分布(图45-47)以及脾脏、淋巴结和骨髓中的b细胞群(图48-50)。在结果中,白细胞包括:b细胞(例如,特征为cd45+、cd19+、tcr-)、t细胞和天然杀伤(nk)细胞(例如,特征为cd45+、tcr-和nk1.1+)。t细胞的特征为cd45+、cd19-、tcr+。cd4+t细胞(cd4)的特征为cd45+、cd19-、tcr+、cd4+、cd8-。cd8+ t细胞(cd8)的特征为cd45+、cd19-、tcr+、cd4-、cd8+。流式细胞术分析中仅包括完整的单个活的白细胞。
[0494]
淋巴结和脾脏中b细胞的发育阶段分为t1(过渡1型b细胞,特征为b220
+
igm
+
igd-)、t2(过渡型2b细胞,特征为b220
+
igm
+
igd
+
)和成熟b细胞(特征为b220
+
igm
低
igd
+
)。图45-47显示了不同组织样品中白细胞的百分比。图48-49显示了不同发育阶段的b细胞百分比。
[0495]
另外,还在脾脏边缘区(边缘区b细胞,mz-b,特征为b220
+
cd21
+
cd23-)和滤泡区(滤泡b细胞,称为fo-b,特征为b220
+
cd21
低
cd23
+
)评估了b细胞的发育。图50显示脾边缘区(mz-b)和滤泡区(fo-b)的脾b细胞百分比。
[0496]
根据不同的发育阶段,骨髓中的b细胞可分为原b细胞(特征为b220
低
cd43
高
igm
低
)、前b细胞(特征为b220
低
cd43
中
igm
低
)和未成熟b细胞(特征为b220
高
cd43
低
igm
高
)。图51a-51c显示了骨髓中不同发育阶段的b细胞的百分比。
[0497]
与野生型小鼠相比,人源化小鼠中的免疫细胞和b细胞的百分比相似,不同组间无统计学差异。在重链人源化杂合小鼠(杂合子)或纯合子小鼠(纯合子)中未观察到显著的b
细胞分化缺陷。
[0498]
实施例10:血清免疫球蛋白同种型分析
[0499]
另外,分析了上述实施例中未免疫的小鼠血清中各种免疫球蛋白的水平。小鼠包括wt小鼠、具有杂合的人源化重链免疫球蛋白基因座的小鼠和具有纯合的人源化重链免疫球蛋白基因座的小鼠。
[0500]
实验使用克隆分型系统-b6/c57j-hrp(southern biotech,目录号5300-05b)试剂盒来进行。首先,捕获抗体山羊抗小鼠ig,人ads-unlb用pbs(solarbio,目录号p1020)稀释至10ug/ml。然后,将0.1ml稀释的抗体加入到酶联免疫吸附测定(elisa)板的每个孔中,并在37℃孵育2小时。接下来,洗涤该板,并在4℃下封闭12小时用1%bsa(cell signaling,目录号9998)梯度稀释。血清样品。向每个孔中加入0.1ml稀释的样品,然后在37℃孵育1小时。将1%bsa加入到孔中作为空白对照。
[0501]
接下来,用含有0.05%吐温-20的pbs(amresco,目录号m147)洗涤平板。加入缀合有辣根过氧化物酶的二抗(山羊抗小鼠iga、igg(1、2b、2c、3)、igm)(用1%bsa稀释300倍,每孔0.1ml),并在37℃下与样品一起孵育1小时。接下来,洗涤板,并通过向每个孔中加入0.1ml tmb色原溶液(beyotime biotechnology,目录号p0209)来对平板进行显影。在室温下于黑暗中孵育8分钟后,向每个孔中加入0.1ml反应溶液(beijing dingguo changsheng biotechnology co.ltd.,目录号eia-0032)。使用微孔板读数器(thermo multiskan go,thermo fisher scientific)测量450nm和570nm处的光吸收,并计算标准od值。
[0502]
结果显示,具有人源化重链免疫球蛋白基因座的小鼠能够产生iga、igg1、igg2b、igg2c、igg3和igm抗体同种型,并且与野生型小鼠相比,所述小鼠对于每种同种型具有相似的表达水平(图52-57)。这表明重链可变区基因区段的人源化对各种抗体同种型的抗体类别转换、表达或分泌没有显著的不利影响。
[0503]
实施例11:小鼠中人可变区基因区段的v(d)j重组
[0504]
通过下一代测序分析小鼠中重链可变区和轻链可变区的基因序列。
[0505]
选择一只未免疫(未暴露于特定抗原)的人源化重链纯合小鼠。从小鼠收集脾细胞用于提取rna。使用5’race试剂盒(smarter race 5
′
/3
′
kit,takara bio usa,inc.,目录号634858)进行逆转录以获得cdna。使用igm恒定区特异性引物和5’race试剂盒的upm引物对获得的cdna进行pcr扩增,以获得重链可变区序列片段,然后测序。igm恒定区特异性引物序列为5
′-
ccaagcttacgagggggaagacatttgggaa-3
′
(seq id no:50)。
[0506]
在另一个实验中,选择11只轻链人源化杂合小鼠,从眼眶采血提取rna。通过上述方法逆转录后,将引物vkf1和igkc-标签用于扩增vκi家族轻链基因,然后测序。使用了以下引物序列:
[0507]
vkf1序列:5
′-
cataagatctcgmcatccrgwtgacccagt-3’(seq id no:51);
[0508]
igkc-标签引物序列:5
′-
ctaacactcattcctgttgaagctcttgac-3
′
(seq id no:52)。
[0509]
将测序结果与人免疫球蛋白序列通过ncbi ig blast工具进行比较,以鉴定v(d)j重组后人v
h
、d
h
、j
h
和vκ、jκ基因的表达。在135个所分析的克隆中,初步结果检测到某些v
h
、d
h
基因区段和全部j
h
基因片段的表达(表15)。这些基因区段中的一些位于人源化片段上的修饰位点附近,一些远离修饰位点。这表明,在用人免疫球蛋白重链序列替换内源染色体片段后,整合到小鼠中的人染色体片段上的人v
h
、d
h
和j
h
基因可被重组以表达人重链。
[0510]
在源自人源化轻链杂合小鼠的441个克隆中检测到大多数vκi家族轻链基因(表16)。与重链检测结果相似,人源化片段上的这些基因中的一些位于非常接近修饰位点的位置,一些远离修饰位点。这表明在用人轻链免疫球蛋白可变区基因座替换内源轻链免疫球蛋白可变区基因座后,整合到小鼠基因组中的人vκ和jκ基因可被重组以表达具有人轻链可变区的轻链。对11只小鼠中的结果的进一步分析表明,检测到的igkv基因的分布在所测试的小鼠中没有显著差异(图58)。
[0511]
表15.检测到的vdj重组后表达的ighv基因、ighd基因和ighj的列表
[0512][0513]
表16.检测到的vj重组后表达的igkv基因的列表
[0514][0515][0516]
实施例12:人源化小鼠中的免疫和抗体产生
[0517]
随机选择5只野生型(wt)小鼠和5只人源化重链纯合小鼠(9-10周龄),并用外源抗原免疫。小鼠每两周重复免疫一次,共进行三次免疫。第二次免疫和第三次免疫后眼眶采血。收集血清,然后通过elisa或facs测定血清滴度,以确定和分析抗原特异性抗体反应。研究中使用了三种抗原,即hbtla、dpd1和ova(图59-63)。结果表明,第二次免疫后,大多数野生型(wt)和人源化重链纯合小鼠产生了抗原特异性抗体。第三次免疫后,抗体滴度增加至1x104至1x105。与野生型小鼠相比,人源化小鼠中的免疫效能测试结果基本相同,这表明小鼠中的人源化免疫球蛋白可变区基因座是有功能的,并且可产生抗原特异性抗体。
[0518]
实施例13:hvh/hvl小鼠中的b细胞发育
[0519]
将具有纯合人源化重链免疫球蛋白基因座的小鼠(人源化vh小鼠或hvh小鼠)和具有人源化轻链免疫球蛋白基因座的小鼠(人源化vl小鼠或hvl小鼠)相互杂交,以获得具有纯合人源化重链免疫球蛋白基因座和纯合人源化轻链免疫球蛋白基因座的小鼠(人源化vh/vl小鼠,或hvh/hvl小鼠)。hvh/hvl小鼠可用于体内产生人源化单克隆抗体。
[0520]
进行实验以比较未免疫的人源化vh/vl小鼠与未免疫的野生型小鼠的免疫系统。在野生型和hvh/hvl小鼠中测量体重和脾脏重量(图65a-65b)。在野生型小鼠与hvh/hvl小鼠之间未检测到平均体重和脾脏重量的显著差异。
[0521]
进行流式细胞术以分析小鼠脾脏中的淋巴细胞群和分布(图66)以及脾脏和骨髓中的b细胞群(图67a-67b、图68a-68c)。结果表明,在hvh/hvl小鼠中,脾脏中的b细胞、t细胞、nk细胞、cd4+ t细胞和cd8+ t细胞的百分比与野生型小鼠几乎完全相同。在结果中,白细胞包括:b细胞(例如,特征为cd45+、cd19+、tcr-)、t细胞和天然杀伤(nk)细胞(例如,特征为cd45+、tcr-和nk1.1+)。t细胞的特征为cd45+、cd19-、tcr+。cd4+ t细胞(cd4)的特征为cd45+、cd19-、tcr+、cd4+、cd8-。cd8+ t细胞(cd8)的特征为cd45+、cd19-、tcr+、cd4-、cd8+。流式细胞术分析中仅包括完整的单个活的白细胞。
[0522]
图67a显示了处于不同发育阶段的b细胞的百分比。脾脏中b细胞的发育阶段分为t1(过渡1型b细胞,特征为b220
+
igm
+
igd-)、t2(过渡2型b细胞,特征为b220
+
igm
+
igd
+
)和成熟
b细胞(特征为b220
+
igm
低
igd
+
)。另外,还在脾脏边缘区(边缘区b细胞,mz-b,特征为b220
+
cd21
+
cd23-)和滤泡区(滤泡b细胞,称为fo-b,特征为b220
+
cd21
低
cd23
+
)评估了b细胞的发育。图67b显示脾边缘区(mz-b)和滤泡区(fo-b)处的脾b细胞的百分比。野生型小鼠与hvh/hvl小鼠之间未观察到显著差异。
[0523]
图68a显示了骨髓中处于不同发育阶段的b细胞的百分比。通过流式细胞术分析了骨髓中的b细胞祖细胞。根据b220和cd43的表达水平,骨髓中的b细胞祖细胞可分为3个细胞群:原b细胞(特征为b220
低
cd43
高
igm
低
)、前b细胞(特征为b220
低
cd43
中
igm
低
)和未成熟b细胞(特征为b220
高
cd43
低
igm
高
)。在野生型小鼠与hvh/hvl小鼠之间未观察到显著差异。
[0524]
另外,还通过流式细胞术评估骨髓或脾脏中的b细胞发育,以对浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)进行选择性染色(图68b-68c)。在野生型小鼠与hvh/hvl小鼠之间没有观察到显著差异。
[0525]
通过elisa定量测量了hvh/hvl小鼠和野生型小鼠的血清中的不同免疫球蛋白(ig)亚型。每组共选择六只小鼠。未观察到iga、igg1、igg2b、igg2c、igg3和igm水平的显著差异(图69)。
[0526]
这些实验表明hvh/hvl小鼠的免疫系统是有功能的,并且hvh/hvl小鼠中的人源化免疫球蛋白基因座可与小鼠免疫球蛋白恒定区适当地相互作用。
[0527]
实施例14:hvh/hvl小鼠中的种系利用率(germline usage)的分析
[0528]
分析了未免疫的hvh/hvl小鼠(无抗原刺激)中的重链ighv、ighd和ighj利用率。结果如图70a-70d所示。另外,还分析了κ链igkv和igkj利用率。结果如图71a-71c所示。
[0529]
通过下一代测序(ngs)来确定未免疫的hvh/hvl小鼠中的种系利用率。例如,如图71c所示,igkj1、igkj2和igkj4在未免疫的hvh/hvl小鼠中频繁使用,而igkj3和igkj5较少被观察到。这种igkj种系使用模式与文献报道的人igkj种系使用情况一致。
[0530]
重链cdr3长度分布由来自未免疫的hvh/hvl小鼠(n=2)的脾细胞的免疫谱系的ngs测序确定。如图72所示,cdr3的中位长度为14个氨基酸。该结果与人免疫系统中人重链cdr3的中位长度一致。
[0531]
分析了重链cdr3(hcdr3)的每个位置处的氨基酸类型(图73)。观察到多种模式,包括酪氨酸使用频率的增加和dh2(ighd2)种系家族使用的增加。这些模式与人hcdr3中的氨基酸组成相似。
[0532]
半胱氨酸残基可形成二硫键。人hcdr3可含有一个半胱氨酸残基或两个半胱氨酸残基,而小鼠hcdr3通常不含半胱氨酸。图74中的结果显示了含有半胱氨酸残基的hvh/hvl小鼠的hcdr3频率,并且该频率随着hcdr3长度的增加而增加。这一结果与人外周血单核细胞(pbmc)中hcdr3的多样性一致。
[0533]
实施例15:淋巴器官组织学分析
[0534]
用苏木精-伊红对来自未免疫的野生型小鼠或未免疫的hvh/hvl小鼠的脾脏、腹股沟淋巴结和派伊尔斑进行染色。代表性切片如图75a-75f所示。野生型(c57bl/6)小鼠和hvh/hvl小鼠表现出具有明显边界的滤泡的正常结构,并且未观察到组织形态学的显著差异。
[0535]
实施例16:hvh/hvl小鼠中的抗体产生
[0536]
在用bcma、il4r、pd-1、siglec15和sirpα抗原进行第二次和第三次免疫后,收集血
液并通过elisa分析野生型(c57bl/6)小鼠和hvh/hvl小鼠的抗原特异性抗体滴度(图76a-76e)。结果表明,hvh/hvl小鼠可以产生与抗原特异性结合的抗体,并且野生型小鼠与hvh/hvl小鼠的免疫反应相似。
[0537]
实施例17:hvh/hvl小鼠中的b细胞发育
[0538]
进行实验以比较免疫后人源化vh/vl小鼠和野生型小鼠的免疫系统。在野生型小鼠和hvh/hvl小鼠中测量体重和脾脏重量(图77a-77b)。在野生型小鼠与hvh/hvl小鼠之间未检测到平均体重和脾脏重量的显著差异。
[0539]
进行流式细胞术以分析小鼠脾中的淋巴细胞群和分布(图78)以及脾脏和骨髓中的b细胞群(图79a-79b、图80a-80c)。
[0540]
图79a显示了脾脏中处于不同发育阶段的b细胞的百分比。另外,还在脾脏边缘区和滤泡区评估了b细胞的发育。图79b显示脾边缘区(mz-b)和滤泡区(fo-b)处的脾b细胞的百分比。在野生型小鼠与hvh/hvl小鼠之间未观察到显著差异。
[0541]
图80a显示了骨髓中处于不同发育阶段的b细胞的百分比。通过流式细胞术分析骨髓中的b细胞祖细胞。在野生型小鼠与hvh/hvl小鼠之间未观察到显著差异。
[0542]
此外,还通过流式细胞术评估了骨髓或脾脏中的b细胞发育,以对浆细胞(b220
低
igm-igd-cd138-)和记忆b细胞(b220
+
igm
+
igd-cd38
+
)进行选择性染色(图80b-80c)。在野生型小鼠与hvh/hvl小鼠之间未观察到显著差异。
[0543]
通过elisa定量测定了hvh/hvl小鼠和野生型小鼠的血清中的不同免疫球蛋白(ig)亚型。每组共选择六只小鼠。未观察到iga、igg1、igg2b、igg2c、igg3和igm水平的显著差异(图81)。另外,通过elisa定量测量了hvh/hvl小鼠和野生型小鼠的血清中的igg总量。未观察到显著差异(图82)。
[0544]
其它实施方案
[0545]
应当理解,虽然已经结合本发明的详细描述描述了本发明,但前述描述旨在说明而不是限制本发明的范围,本发明的范围由所附权利要求的范围限定。其它方面、有利方面和修改在以下权利要求的范围内。
[0546]
[0547]
[0548]
[0549]
[0550]
[0551]
[0552]
[0553]
[0554]
[0555]
[0556]
[0557]
[0558]
[0559]
[0560]
[0561]
[0562]
[0563]
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1