一种γ-谷酰胺甲胺合成酶的突变体,其编码基因、氨基酸序列及其应用的制作方法
本发明涉及一种酶突变体,特别涉及一种γ-谷酰胺甲胺合成酶的突变体、其编码基因、氨基酸及在生产茶氨酸中的应用。
背景技术:
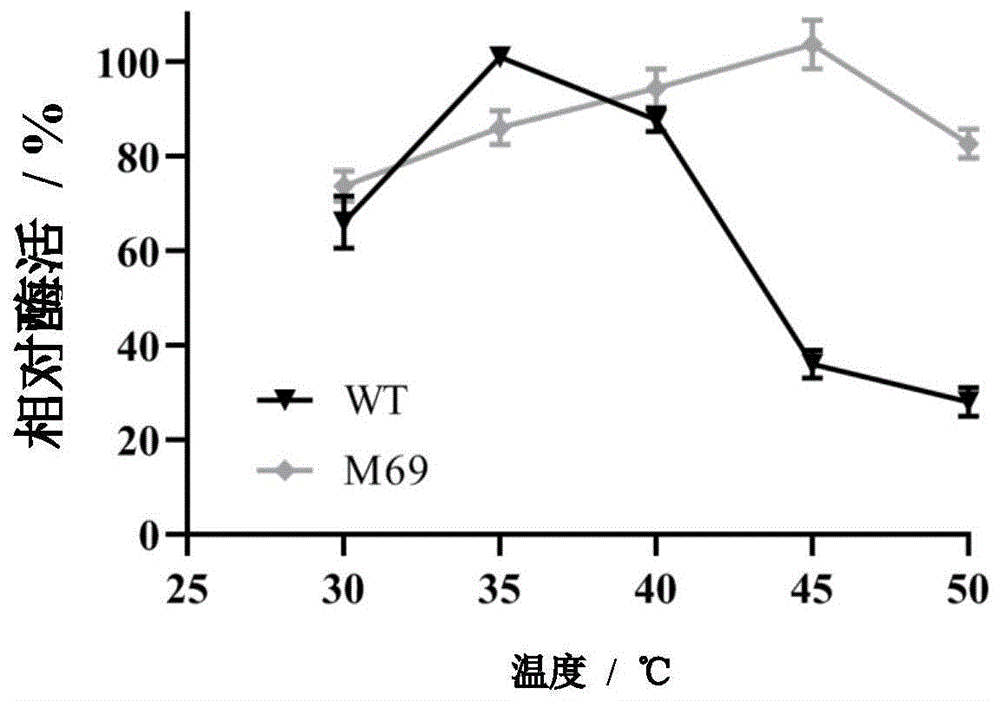
:l-茶氨酸(l-theanine)属于酰胺类化合物,是茶叶中含量最丰富的游离氨基酸,占总游离氨基酸的50%以上。茶氨酸为是茶叶香味和口感的主要成分,其含量对茶叶的风味和品质起到关键性的作用。茶氨酸的性质较稳定,将茶氨酸溶液煮沸5min,或将茶氨酸溶于ph3.0的溶液中并在25℃下储放12个月,茶氨酸含量不变。因此在通常的食品加工、杀菌过程中,茶氨酸的性质不会发生变化,且毒性试验中未发现其有毒性。l-茶氨酸作为一种安全、集多种生理功能于一体的功能性活性因子,被广泛应用于食品、保健品和医药行业。微生物酶法生产茶氨酸作为近年来较为经济和温和的方法,必将在未来产业应用中大有前景。γ-谷氨酰甲胺合成酶(γ-glutamylmethylamidesynthetase,gmas)是一种能够催化l-谷氨酸和甲胺合成n-甲基-l-谷氨酰胺的催化用酶,该反应需要消耗atp。专利cn201410316269利用大肠杆菌产γ-谷氨酰甲胺合成酶高效生产l-茶氨酸,在大肠杆菌中表达γ-谷酰胺甲胺合成酶基因,可以谷氨酸和乙胺为底物生产茶氨酸,50mm限制底物,8h谷氨酸转化率达到69.8%;专利cn201510755912在大肠杆菌中表达γ-谷酰胺甲胺合成酶基因,可以谷氨酸和乙胺为底物生产茶氨酸,150mm限制底物,转化率低于50%,生产周期长于30h,效率低,不适合工业生产。在cn201510973289中以γ-谷氨酰甲胺合成酶和磷酸激酶为催化剂,在咪唑溶液中以l-谷氨酸钠和乙胺盐酸盐为底物合成l-茶氨酸。100ml体系中限制底物最高200mm,转化率87%,反应体系较小,且咪唑为反应环境,对环境和成本均不友好。专利cn201880024038公开了一种使用以乙醛和丙氨酸作为底物生成乙胺的活性、以及γ-谷氨酰甲胺合成酶活性或谷氨酰胺酶活性增强的微生物,能够在不从外部添加乙胺且不会蓄积、残留乙胺作为副产物的情况下高效地制造茶氨酸,但乙醛并不安全,此举并无太多益处。γ-谷氨酰甲胺合成酶虽然是更为优良的催化剂,但据已有文献报道或专利发明文件来看,其生产茶氨酸的效率并不高。阻碍微生物酶法生产茶氨酸效率的一个原因是γ-谷氨酰甲胺合成酶容易失活。从相关文献的报道来看,茶氨酸的生产速率随着反应时间的延长而降低,符合酶的失活曲线,此为酶本身的性质决定。因此,如何提高γ-谷氨酰甲胺合成酶的热稳定性以及提高其生产茶氨酸的效率,为酶法工业化生产茶氨酸提供有效途径是当前亟需解决的技术问题。技术实现要素:针对现有技术中的不足之处,本发明通过基因工程方法获得了γ-谷氨酰甲胺合成酶突变体;并且,不仅在提高了γ-谷氨酰甲胺合成酶突变体的热稳定性同时,使酶的最适温度提高了10℃左右,并且该突变体不需要在缓冲盐溶液中就能发挥很好的催化效果,同时利用该突变体通过耦合多聚磷酸盐激酶的方法高效合成茶氨酸,该方法催化茶氨酸合成的同时可实现atp再生,为酶法工业化生产茶氨酸提供有效途径。本发明第一方面提供了一种γ-谷酰胺甲胺合成酶突变体,所述突变体的氨基酸序列如seqidno.3所示。进一步地,前文所述的γ-谷酰胺甲胺合成酶突变体,所述突变体的基因序列如seqidno.2所示;或编码所述突变体的基因序列为编码前文所述的如seqidno.3所示氨基酸序列的核苷酸序列。所述的γ-谷酰胺甲胺合成酶突变体即实施例中的m69突变体。本发明第二方面提供了一种γ-谷酰胺甲胺合成酶突变体的构建方法,包括如下步骤;按照随机突变试剂盒获得随机突变体,筛选其中具有优良性能的突变体。该方法中所述试剂盒为市面上可购买到的任意基因随机突变试剂盒。本发明第三方面提供了一种γ-谷酰胺甲胺合成酶突变体在生产茶氨酸中的应用。进一步地,所述应用包括将γ-谷酰胺甲胺合成酶突变体的全细胞用于茶氨酸生产的催化体系中进行反应。进一步地,所述茶氨酸生产的催化体系包括300~600mm谷氨酸钠和/或谷氨酸;350~600mm乙胺和/或乙胺盐酸盐;120~180mm含镁离子的盐,优选浓度为140~160mm;1~5mmatp和/或adp,优选浓度为2mm;以及100~200mm多聚磷酸盐;进一步地,所述茶氨酸生产的催化体系温度控制在40~50℃范围内,反应ph可在6~9范围内波动;进一步地,所述茶氨酸生产的催化体系中,添加40~70g/l的m69湿菌体和30~50g/l的能够以adp和/或amp为底物进行atp循环再生的菌体。优选的,所述能够以adp和/或amp为底物进行atp循环再生的菌体为专利cn201910642412-一种用于生产谷胱甘肽的重组大肠杆菌的混合培养工艺及其应用中所述的菌株bl21-pet30a-ppk2。对于上文所述的技术方案中,优选的所述含镁离子的盐包括氯化镁、硫酸镁等;对于上文所述的技术方案中,优选的所述多聚磷酸盐包括六偏磷酸钠、三聚磷酸钠等;对于上文所述的技术方案中,优选的谷氨酸钠和/或谷氨酸的浓度为350~450mm;对于上文所述的技术方案中,优选的乙胺和/或乙胺盐酸盐的浓度为420~450mm;对于上文所述的技术方案中,优选的含镁离子的盐浓度为140~160mm;对于上文所述的技术方案中,优选的atp和/或adp的优选浓度为2mm;对于上文所述的技术方案中,优选的多聚磷酸盐优选浓度为160~180mm;对于上文所述的技术方案中,优选的添加m69湿菌体的浓度为55~65g/l;对于上文所述的技术方案中,优选的添加能够以adp和/或amp为底物进行atp循环再生的菌体的浓度为35~45g/l;对于上文所述的技术方案中,优选的体系温度控制在43~46℃;对于上文所述的技术方案中,优选的反应ph可在6.5~7.5范围内。与现有技术相比,本发明具有如下有益效果:本发明通过基因工程方法获得的γ-谷氨酰甲胺合成酶突变体m69;其性质较野生株有显著提高,体现在以下几方面:(1)突变体的最适温度比原始菌提高了10℃;(2)最适温度下的生产强度是原始菌的2.88倍;(3)酶的热稳定性有显著提高,在35℃下,突变体的半衰期是原始菌的5.52倍,在45℃下半衰期提高到原来的8.19倍;(4)该突变体不需要在缓冲盐溶液中就能发挥很好的催化效果,提高了酶在水体系中反应的活性,从而降低了反应成本,同时有利于简化后续纯化工艺,缩减生产周期,对规模化生产有重要意义;(5)耦合多聚磷酸盐激酶,解除atp抑制,同时实现atp的再生,降低了工业成本。附图说明图1原始菌和突变体最适温度;图2原始菌和突变体在35℃和45℃下的热稳定性;图3原始菌和突变体最适ph;图4氯化镁最适添加量;图5m69加入量;图6ppk加入量。具体实施方式下面对本发明的具体实施方式进行详细地描述,但应当理解本发明的保护范围并不受具体实施方式的限制。本发明中,除非另有其他明确说明,否则百分比、百分含量均以质量计。如无特殊说明,所使用的实验方法均为常规方法,所用材料、试剂等均可从商业途径购买。本申请的实施例中ppk湿菌体为专利cn201910642412-一种用于生产谷胱甘肽的重组大肠杆菌的混合培养工艺及其应用中所述的菌株bl21-pet30a-ppk2。本申请的实施例中所涉及的酶学性质、热稳定性和半衰期检测方法如下:1.酶学性质测定酶活定义:1u酶活定义为在35℃下,每分钟内生成1μm的茶氨酸所需要的细胞量,反应体系包含50mm谷氨酸钠,55mm乙胺,40mm氯化镁和30mmatp以及15g/l的gmas细胞,于ph=7的pbs溶液中反应60min,加入等体积的20%三氯乙酸进行终止,hplc检测茶氨酸产量以确定酶活力。突变后gmas的酶学性质可能发生改变,探究最佳酶活条件,分别将未突变wt(原始菌)和突变体m69在相同条件下的酶学性质进行分析比较。2.热稳定性和半衰期检测温度稳定性的测定:用pbs将酶液的浓度稀释至30g/l,在35℃和45℃下孵育,测定孵育不同时间的残余酶活。t1/2值是指酶在特定温度下处理一段时间后残余酶活为50%时对应的时间。具体测定方法如下:以未进行热处理的gmas酶的活力作为100%,分别测定并计算出酶在35℃和45℃下处理不同时间后的残余酶活。以处理时间为横坐标,以ln(%残余酶活)为纵坐标,绘制时间—ln(%残余酶活)的曲线,据图计算t1/2=ln2/kd,kd为该图斜率。实施例1突变体的构建来源于亲甲基醇菌methylovorusmays的茶氨酸合成酶基因gmas序列,密码子优化后的序列为seqidno.1,连入质粒pet28(a),转入e.colibl21,得到原始菌e.colibl21-pet28(a)-gmas命名为wt。获得γ-谷酰胺甲胺合成酶突变体m69的方法,包括如下步骤:1.按照quickmutationtm基因随机突变试剂盒设计引物和随机突变,引物序列为:random-f:gatatacatatgaagagcctggaag;seqidno.4;random-r:cccaagcttgtagaattgaacatagcgg;seqidno.5;2.随机突变pcr反应随机突变pcr反应参考下表设置随机突变pcr反应体系:表1试剂最终浓度体积双蒸水或milliq水-12.2μlrandommutbuffer(10×)1×2μlmutationenhancer(10×)1×2uldntp(2.5mm)0.25mm2μl模板dna0.2pg-5ng/μl1μl引物混合物(各10μm)各0.2μm0.4μlrandommutdnapolymerase-0.4μl总体积-20μl按照如下参数设置pcr仪:表23.转化感受态细胞取10μl的pcr产物,1%的琼脂糖检测,观察到目的条带后加1μl的dpni酶于剩余的pcr产物中,混匀孵育1h。加入2-5μl消化产物与50μl感受态细胞中,混匀后冰浴30min,之后42℃水浴热击45s,立即放于冰上2-5min。之后加入250μllb培养基,200rpm,37℃培养1h,取100-200μl菌液于卡那抗性的平板上过夜培养。得到多个突变体,其中性质优良的突变体e.colibl21-pet28(a)-gmas-m69命名为突变体m69,突变体m69核苷酸序列如seqidno.2所示,其氨基酸序列如seqidno.3所示。野生酶和突变酶的表达:使用lb培养基培养原始菌和突变体m69,于37℃,200rpm的条件下过夜,以5%接种量接种到发酵培养基中,进行扩大培养,待细胞od600=0.4~1.0,添加0.1mm的iptg,低温诱导过夜,离心收获全细胞作为催化剂。所述的种子培养基和发酵培养基:酵母浸粉5g/l,蛋白胨10g/l,nacl10g/l,121℃灭菌20min,冷却后加入终浓度为10~100mg/ml的卡那霉素。实施例2以突变体m69湿菌体为生物催化剂,以谷氨酸钠和乙胺盐酸盐为底物,生物转化制备茶氨酸。催化体系为10l,催化体系包含400mm谷氨酸钠,440mm乙胺,160mm氯化镁,2mmatp,160mm六偏磷酸钠以及60g/l的m69湿菌体和40g/lppk湿菌体。反应体系于45℃,ph=7的水溶液中进行反应,反应结束后离心去除细胞,样品用0.22μm膜过滤后进行hplc检测茶氨酸产量。茶氨酸最高产量372mm,合65g/l,反应时长为12h,生产强度为5.42g/l/h,是原始菌的2.88倍。实施例3温度对酶性质的影响温度即能改变酶发生催化反应速度,也能导致酶蛋白活性降低或失活。同时测定未突变wt和突变体m69的最佳反应温度。分别置于30、35、40、45、50℃下反应60min。在ph=7的pbs溶液中,反应体系包含50mm谷氨酸钠,55mm乙胺,40mm氯化镁和30mmatp以及15g/l的gmas细胞。hplc定时测定反应液茶氨酸变化情况,将35℃下测定wt的酶活值作为100%,计算其他温度下的相对酶活。实验结果如图1所示,wt的最适温度为35℃,而m69的最适温度在45℃附近。将未突变wt和突变体m69置于35和45℃下孵育,以未孵育时酶活为100%,测定孵育不同时间的残余酶活。按照图2所示的测定结果计算半衰期,结果如表3,可知在35℃下,突变体的半衰期是原始菌的5.52倍,在45℃下半衰期提高到原来的8.19倍。表3半衰期/h35℃45℃wt12.482.78m6968.9622.78实施例4ph对酶性质的影响酶反应都有其最佳的ph值范围,ph值过高或者过低对酶发生催化反应的活性都会产生影响,本实验将同时测定未突变wt和突变体m69的最佳反应ph。准备若干组100ml反应液,反应体系包含50mm谷氨酸钠,55mm乙胺,40mm氯化镁和30mmatp以及15g/l的gmas细胞。分别调节ph值为5.0、6.0、7.0、8.0、9.0,置于35℃反应60min,hplc定时测定反应液茶氨酸变化情况,将ph=7下测得的未突变的酶活值作为100%,计算其他ph值下的相对酶活。结果如图3所示,实验结果表明未突变wt和突变体m69的最佳反应ph都是7.0。说明突变前后并未改变gmas的最适ph值。实施例5非缓冲体系中反应分别在pbs和纯水中溶解50mm谷氨酸钠,55mm乙胺,40mm氯化镁和30mmatp以及15g/l的原始菌和突变体m69的细胞,调ph=7,在原始菌和突变体m69各自的最适温度下进行反应,60min后hplc测定茶氨酸产量,结果如表4。可知突变体m69在非缓冲体系中可用且酶活更高。表4产量/mmpbs水wt35.431.2m6942.346.7实施例6加入能量循环atp作为底物之一在高浓度的工业化生产中有可能产物底物抑制效应,同时增加成本。利用ppk循环再生atp可有效避免上述问题。按照专利110387379a中的方法获得bl21-pet30a-ppk2细胞,以adp、amp和六偏磷酸钠为底物可进行atp再生。在100ml的反应体系中,以10g/l的bl21-pet30a-ppk2细胞和20mm六偏磷酸钠、2mm的atp替代30mm的atp,反应体系还包含50mm谷氨酸钠,55mm乙胺,40mm氯化镁以及15g/l的m69细胞;在45℃,ph=7的水溶液中进行反应,hplc定时测定反应液茶氨酸变化情况。反应60min,转化率达95.6%,是直接添加atp的1.06倍。实施例7提高底物浓度提高底物浓度在工业化生产中具有更高的经济性。然而,较高的底物浓度反应如果直接翻倍会面临底物抑制、激活剂过量或者酶的添加量不合适等问题。在100ml的反应体系中,添加400mm谷氨酸钠,440mm乙胺,2mmatp,160mm六偏磷酸钠以及120g/l的m69湿菌体和80g/l的ppk湿菌体。分别添加320mm、240mm、160mm、80mm的氯化镁,于45℃,ph=7下进行反应,hplc检测茶氨酸产量。根据图4中的曲线,选择160mm氯化镁作为反应中的激活剂。在100ml的反应体系中,添加400mm谷氨酸钠,440mm乙胺,160mm氯化镁,2mmatp,160mm六偏磷酸钠以及80g/l的ppk湿菌体。分别添加120g/l、90g/l、60g/l和30g/l的m69的湿菌体,于45℃,ph7下进行反应,hplc检测茶氨酸产量。根据图5的曲线,认为60g/l的湿菌体足够完成反应。在100ml的反应体系中,添加400mm谷氨酸钠,440mm乙胺,160mm氯化镁,2mmatp,160mm六偏磷酸钠以及60g/lm69的湿菌体,分别添加80g/l、60g/l、40g/l和20g/l的m69的湿菌体,于45℃,ph=7下进行反应,hplc检测茶氨酸产量。根据图6的曲线,反应中40g/l的ppk湿菌体足以提供足够的atp。对比例1原始菌转化效果以gmas野生菌湿菌体为生物催化剂,以谷氨酸钠和乙胺盐酸盐为底物,生物转化制备茶氨酸。催化体系为10l,催化体系包含400mm谷氨酸钠,440mm乙胺,160mm氯化镁,2mmatp,160mm六偏磷酸钠以及60g/l的gmas野生菌湿菌体和40g/l的ppk湿菌体。反应体系于35℃,ph=7的水溶液中进行反应,反应结束后离心去除细胞,样品用0.22μm膜过滤后进行hplc检测茶氨酸产量。茶氨酸最高产量282mm,合48.9g/l,反应时长为26h,生产强度为1.88g/l/h。其他突变体转化效果:以按照实施例1的方法获得的其他突变后氨基酸为半胱氨酸的突变体d103c、p337c湿菌体为生物催化剂,以谷氨酸钠和乙胺盐酸盐为底物,生物转化制备茶氨酸。催化体系为10l,催化体系包含400mm谷氨酸钠,440mm乙胺,160mm氯化镁,2mmatp,160mm六偏磷酸钠以及60g/l的d103c或p337c湿菌体和40g/l的ppk湿菌体。反应体系于35℃,ph=7的水溶液中进行反应,反应结束后离心去除细胞,样品用0.22μm膜过滤后进行hplc检测茶氨酸产量。茶氨酸最高产量分别为142mm和290mm,合24.5g/l和51g/l,反应时长为28h和23h,生产强度仅有0.88g/l/h和2.2g/l/h。与原始菌相比活性更低或相近。总之,通过本申请的对比例和实施例数据对比分析可知:所获得的突变体m69在应用于茶氨酸合成过程中可显著提高其生产强度,而突变体d103c、p337c虽然突变后的氨基酸也形成了半胱氨酸,却并没有达到同样正向效果。以上所述,仅为本发明较佳的具体实施方式,但本发明的保护范围并不局限于此,本领域的技术人员在本发明披露的技术范围内,可轻易想到的变化或替换,都应涵盖在本发明的保护范围之内。因此,本发明的保护范围应该以权利要求书的保护范围为准。序列表<110>大连医诺生物股份有限公司<120>一种γ-谷酰胺甲胺合成酶的突变体,其编码基因、氨基酸序列及其应用<160>5<170>siposequencelisting1.0<210>1<211>1332<212>dna<213>密码子优化后的茶氨酸合成酶gmas基因(无)<400>1atgaagagcctggaagaagcacaaaagtttctggaagaccaccatgtcaagtatgtcctg60gcacagttcgtcgatattcacggcgttgctaaagtgaagagcgttccggcgtctcatctg120aatgatattctgaccacgggtgcaggttttgcaggcggtgctatttggggtaccggtatc180gcaccgaacggtccggattatatggcaattggtgaactgagtacgctgtccctgatcccg240tggcagccgggttatgcacgtctggtctgcgacggccacgtgaatggtaaaccgtatgaa300tttgatacccgtgtggttctgaaacagcaaattgcacgtctggcagaaaagggttggacc360ctgtatacgggtctggaaccggaattttcactgctgaaaaaggatgaacatggcgcggtg420cacccgttcgatgactcggacacgctgcaaaaaccgtgttatgattacaagggtattacc480cgccatagcccgtttctggaaaaactgacggaatctctggttgaagtcggcctggacatt540tatcagatcgatcacgaagacgccaatggtcaatttgaaatcaactatacctacgccgat600tgcctgaaaagtgcagatgactacattatgttcaagatggcggcctccgaaatcgcgaac660gaactgggtattatctgtagttttatgccgaaaccgttctccaatcgtccgggcaacggt720atgcacatgcacatgtcaattggcgacggtaaaaagtcgctgtttcaggatgactcagat780ccgtcgggcctgggtctgagtaaactggcttatcatttcctgggcggtatcctggcacac840gcaccggcactggcagctgtttgcgcaccgaccgtcaattcttacaaacgtctggtcgtg900ggtcgcagcctgtctggtgctacctgggctccggcgtatattgcgtacggcaacaataac960cgtagcacgctggttcgcatcccgtatggccgtctggaactgcgcctgccggatggttct1020tgtaacccgtacctggcaaccgcagcagtgattgcagctggtctggacggtgttgcacgt1080gaactggatccgggcacgggtcgcgatgacaatctgtatgattacagcctggaacagctg1140gccgaatttggcattggtatcctgccgcaaaacctgggtgaagcactggatgctctggaa1200gcggaccaggtcatcatggatgcgatgggcccgggtctgtccaaagaatttgttgaactg1260aagcgcatggaatgggtggactatatgcgtcatgtgtcggactgggaaatcaaccgctat1320gttcaattctac1332<210>2<211>1332<212>dna<213>γ-谷酰胺甲胺合成酶突变体核苷酸序列(无)<400>2atgaagagcctggaagaagcacaaaagtttctggaagaccaccatgtcaagtatgtcctg60gcacagttcgtcgatattcacggcgttgctaaagtgaagagcgttccggcgtctcatctg120aatgatattctgaccacgggtgcaggttttgcaggcggtgctatttggggtaccggtatc180gcaccgaacggtccggattatatgtgcattggtgaactgagtacgctgtccctgatcccg240tggcagccgggttatgcacgtctggtctgcgacggccacgtgaatggtaaaccgtatgaa300tttgatacccgtgtggttctgaaacagcaaattgcacgtctggcagaaaagggttggacc360ctgtatacgggtctggaaccggaattttcactgctgaaaaaggatgaacatggcgcggtg420cacccgttcgatgactcggacacgctgcaaaaaccgtgttatgattacaagggtattacc480cgccatagcccgtttctggaaaaactgacggaatctctggttgaagtcggcctggacatt540tatcagatcgatcacgaagacgccaatggtcaatttgaaatcaactatacctacgccgat600tgcctgaaaagtgcagatgactacattatgttcaagatggcggcctccgaaatcgcgaac660gaactgggtattatctgtagttttatgccgaaaccgttctccaatcgtccgggcaacggt720atgcacatgcacatgtcaattggcgacggtaaaaagtcgctgtttcaggatgactcagat780ccgtcgggcctgggtctgagtaaactggcttatcatttcctgggcggtatcctggcacac840gcaccggcactggcagctgtttgcgcaccgaccgtcaattcttacaaacgtctggtcgtg900ggtcgcagcctgtctggtgctacctgggctccggcgtatattgcgtacggcaacaataac960cgtagcacgctggttcgcatcccgtatggccgtctggaactgcgcctgccggatggttct1020tgtaacccgtacctggcaaccgcagcagtgattgcagctggtctggacggtgttgcacgt1080gaactggatccgggcacgggtcgcgatgacaatctgtatgattacagcctggaacagctg1140gccgaatttggcattggtatcctgccgcaaaacctgggtgaagcactggatgctctggaa1200gcggaccaggtcatcatggatgcgatgggcccgggtctgtccaaagaatttgttgaactg1260aagcgcatggaatgggtggactatatgcgtcatgtgtcggactgggaaatcaaccgctat1320gttcaattctac1332<210>3<211>444<212>prt<213>γ-谷酰胺甲胺合成酶突变体氨基酸序列(无)<400>3metlysserleugluglualaglnlyspheleugluasphishisval151015lystyrvalleualaglnphevalaspilehisglyvalalalysval202530lysservalproalaserhisleuasnaspileleuthrthrglyala354045glyphealaglyglyalailetrpglythrglyilealaproasngly505560proasptyrmetcysileglygluleuserthrleuserleuilepro65707580trpglnproglytyralaargleuvalcysaspglyhisvalasngly859095lysprotyrglupheaspthrargvalvalleulysglnglnileala100105110argleualaglulysglytrpthrleutyrthrglyleugluproglu115120125pheserleuleulyslysaspgluhisglyalavalhispropheasp130135140aspseraspthrleuglnlysprocystyrasptyrlysglyilethr145150155160arghisserpropheleuglulysleuthrgluserleuvalgluval165170175glyleuaspiletyrglnileasphisgluaspalaasnglyglnphe180185190gluileasntyrthrtyralaaspcysleulysseralaaspasptyr195200205ilemetphelysmetalaalasergluilealaasngluleuglyile210215220ilecysserphemetprolyspropheserasnargproglyasngly225230235240methismethismetserileglyaspglylyslysserleuphegln245250255aspaspseraspproserglyleuglyleuserlysleualatyrhis260265270pheleuglyglyileleualahisalaproalaleualaalavalcys275280285alaprothrvalasnsertyrlysargleuvalvalglyargserleu290295300serglyalathrtrpalaproalatyrilealatyrglyasnasnasn305310315320argserthrleuvalargileprotyrglyargleugluleuargleu325330335proaspglysercysasnprotyrleualathralaalavalileala340345350alaglyleuaspglyvalalaarggluleuaspproglythrglyarg355360365aspaspasnleutyrasptyrserleugluglnleualagluphegly370375380ileglyileleuproglnasnleuglyglualaleuaspalaleuglu385390395400alaaspglnvalilemetaspalametglyproglyleuserlysglu405410415phevalgluleulysargmetglutrpvalasptyrmetarghisval420425430serasptrpgluileasnargtyrvalglnphetyr435440<210>4<211>25<212>dna<213>随机引物-f(无)<400>4gatatacatatgaagagcctggaag25<210>5<211>28<212>dna<213>随机引物-r(无)<400>5cccaagcttgtagaattgaacatagcgg28当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1