一种耐热性β-甘露聚糖酶突变体ManAK-2及其编码基因和应用
本发明属于基因工程和酶工程领域,特别涉及一种耐热性提高的β-甘露聚糖酶突变体manak-2及其编码基因和应用。
背景技术:
甘露聚糖是植物半纤维素的主要成分,广泛存在于自然界中,它是多种植物细胞壁的主要组成成分,作为植物性饲料原料被视为一种抗营养因子。由于甘露聚糖的多样性与结构的复杂性,它的完全水解需要多种酶的协同作用才能完成,包括内切-β-甘露聚糖酶、外切-β-甘露糖苷酶、β-葡萄糖苷酶、乙酞甘露聚糖酯酶和α-半乳糖苷酶等。其中,内切-β-甘露聚糖酶能够降解甘露聚糖主链的β-1,4-糖苷键,是甘露聚糖降解过程中发挥最重要作用的酶。
β-甘露聚糖酶(β-mannanase,ec3.2.1.78)主要来源于细菌、真菌和放线菌等微生物,目前,应用于工业生产β-甘露聚糖酶的菌种多为曲霉、芽抱杆菌、酵母等,在食品、医药、饲料等领域有广泛的应用。β-甘露聚糖酶在饲料工业中作为一种饲料添加剂,能有效地降解饲料中的甘露聚糖,提高动物对饲料的消化能力。目前饲料添加剂用酶都需要经过高温制粒,制备能够耐高温、耐酸、具有对消化酶耐受性提高的β-甘露聚糖酶,对于β-甘露聚糖酶在饲用酶和工业化应用具有重要价值。野生型的β-甘露聚糖酶基因经过异源表达和理性设计后已经能够获得较好的酶学性质,对于提高现有的β-甘露聚糖酶的耐高温性能发挥着至关重要的作用。
技术实现要素:
本发明的目的在于提供一种耐热性提高的β-甘露聚糖酶突变体,在饲料及其他领域具有更为良好的应用潜力。
具体而言,本发明针对前期开发的耐热酸性β-甘露聚糖酶manak进行必要的理性设计,改良酶学特性。manak最适反应ph为2,且当ph在2.0-6.0范围内具有较好的稳定性。最适作用温度为75℃,在60℃高温下30min可保持83%的酶活。然而,作为饲料添加剂应用,其耐热性和酶活力还有待进一步提高。
为了提高manak耐热性能,本发明对该酶的空间结构进行分子模拟对接分析,通过分析分子内二硫键,将该酶进行单位点和双位点突变,得到的突变体的最适温度(topt)和t1/2都有改变,耐热性能提高。
一种耐热性β-甘露聚糖酶突变体,是在野生β-甘露聚糖酶manak的氨基酸序列如seqidno.1所示基础上进行的突变,所述的突变位点选自g331c、s158y、q166w、s177r、s180i、k99w、n178r、n178i、d273c/v308c或n277c/t315c中的一种。
本发明的再一目的是提供编码上述突变体的基因。
本发明的再一目的是提供包含上述突变体基因的重组载体。
本发明的再一目的是提供包含上述突变体基因的重组菌株,所述重组菌株为毕赤酵母x33。
本发明还提供所述突变体在饲料或饲料添加剂中的应用。
本发明还提供含有所述突变体的饲料或饲料添加剂。
本发明还提供所述重组菌株在饲料或饲料添加剂中的应用。
本发明还提供所述重组载体在饲料或饲料添加剂中的应用。
根据本发明的技术方案,野生β-甘露聚糖酶manak的氨基酸序列如seqidno.1所示。
seqidno.1:
talpkaspapsssssssssastsfastsglqftidgetgyfagtnsywigfltddsdvdlvmshlkssglkilrvwgfndvttqpssgtvwyqlhqdgkstintgadglqrldyvvssaeqhdikliinfvnywtdyggmsayvsayggsdetdfytsdtmqsayqtyiktvverysnssavfawelaneprcpscdtsvlydwiektskfikgldadhmvcigdegfglntdsdgsypyqfaeglnftknlgidtidfgtlhlypdswgtsddwgngwisahgaackaagkpclleeygvtsnhcsvespwqktalnttgvsadlfwqygddlstgkspddgntiyygtsdyeclvtdhvaaigsa
野生β-甘露聚糖酶manak的核苷酸序列如seqidno.2所示。
seqidno.2:
actgctttgccaaaggcttctccagctccatcttcttcttcttcttcctcttcttctgcttctacttcttttgcttctacttctggtttgcaatttactattgatggtgaaactggttactttgctggtactaactcttactggattggttttcttactgatgattctgatgttgatttggttatgtctcatttgaagtcttctggtttgaagattttgagagtttggggttttaacgatgttactactcaaccatcttctggtactgtttggtaccaattgcatcaagatggtaagtctactattaacactggtgctgatggtttgcaaagattggattacgttgtttcttctgctgaacaacatgatattaagttgattattaactttgttaactactggactgattacggtggtatgtctgcttacgtttctgcttacggtggttctgatgaaactgatttttacacttctgatactatgcaatctgcttaccaaacttacattaagactgttgttgaaagatactctaactcttctgctgtttttgcttgggaattggctaacgaaccaagatgtccatcttgtgatacttctgttttgtacgattggattgaaaagacttctaagtttattaagggtttggatgctgatcatatggtttgtattggtgatgaaggttttggtttgaacactgattctgatggttcttacccataccaatttgctgaaggtttgaactttactaagaacttgggtattgatactattgattttggtactttgcatttgtacccagattcttggggtacttctgatgattggggtaacggttggatttctgctcatggtgctgcttgtaaggctgctggtaagccatgtttgttggaagaatacggtgttacttctaaccattgttctgttgaatctccatggcaaaagactgctttgaacactactggtgtttctgctgatttgttttggcaatacggtgatgatttgtctactggtaagtctccagatgatggtaacactatttactacggtacttctgattacgaatgtttggttactgatcatgttgctgctattggttctgcttaa
所述的点突变g331c为甘露聚糖酶突变体manak-1(g331c),由氨基酸序列为seqidno.1的甘露聚糖酶的第331位氨基酸甘氨酸(g)变为半胱氨酸(c)获得。
所述的点突变s158y为甘露聚糖酶突变体manak-2(s158y),由氨基酸序列为seqidno.1的甘露聚糖酶的第158位氨基酸丝氨酸(s)变为酪氨酸(y)获得。
所述的点突变q166w为甘露聚糖酶突变体manak-3(q166w),由氨基酸序列为seqidno.1的甘露聚糖酶的第166位氨基酸天冬酰胺(n)变为色氨酸(w)获得。
所述的点突变s177r为甘露聚糖酶突变体manak-4(s177r),由氨基酸序列为seqidno.1的甘露聚糖酶的第177位氨基酸丝氨酸(s)变为精氨酸(r)获得。
所述的点突变s180i为甘露聚糖酶突变体manak-5(s180i),由氨基酸序列为seqidno.1的甘露聚糖酶的第180位氨基酸丝氨酸(s)变为异亮氨酸(i)获得。
所述的点突变k99w为甘露聚糖酶突变体manak-6(k99w),由氨基酸序列为seqidno.1的甘露聚糖酶的第99位氨基酸赖氨酸(k)变为色氨酸(w)获得。
所述的点突变n178r为甘露聚糖酶突变体manak-7(n178r),由氨基酸序列为seqidno.1的甘露聚糖酶的第178位氨基酸天冬酰胺(n)变为精氨酸(r)获得。所述的点突变n178i为甘露聚糖酶突变体manak-8(n178i),由氨基酸序列为seqidno.1的甘露聚糖酶的第178位氨基酸天冬酰胺(n)变为异亮氨酸(i)获得。
所述的点突变d273c/v308c为甘露聚糖酶突变体manak-9(d273c/v308c),由氨基酸序列为seqidno.1的甘露聚糖酶的第273位氨基酸天冬氨酸(d)变为半胱氨酸(c),第308位氨基酸缬氨酸(v)变为半胱氨酸(c)获得。
所述的点突变n277c/t315c为甘露聚糖酶突变体manak-10(n277c/t315c),由氨基酸序列为seqidno.1的甘露聚糖酶的第277位氨基酸天冬酰胺(n)变为半胱氨酸(c),第315位氨基酸苏氨酸(t)变为半胱氨酸(c)获得。
本发明与现有技术相比的有益效果:
(1)本发明是以二硫键为基础进行理性设计,利用三维结构模拟结合酶学性质分析,筛选得到耐热性能提高的突变体,单点突变体对高温的耐受能力为s177r>n178r>n178i>g331c>k99w>s180i>q166w>s158y,表现出突变体耐热性的增强。可见分子内二硫键的引入显著提高了该酶在高温条件下稳定性。
(2)各突变体保持了野生型酶耐酸性及广范围ph的稳定性,
(3)最适温度除突变体n178r和d273-v308的最适温度为85℃外,其余所有单位点和双位点突变体的最适温度均在75℃,与甘露聚糖酶manak相比,本发明提供的突变体的最适作用温度没有发生较大变化。
附图说明
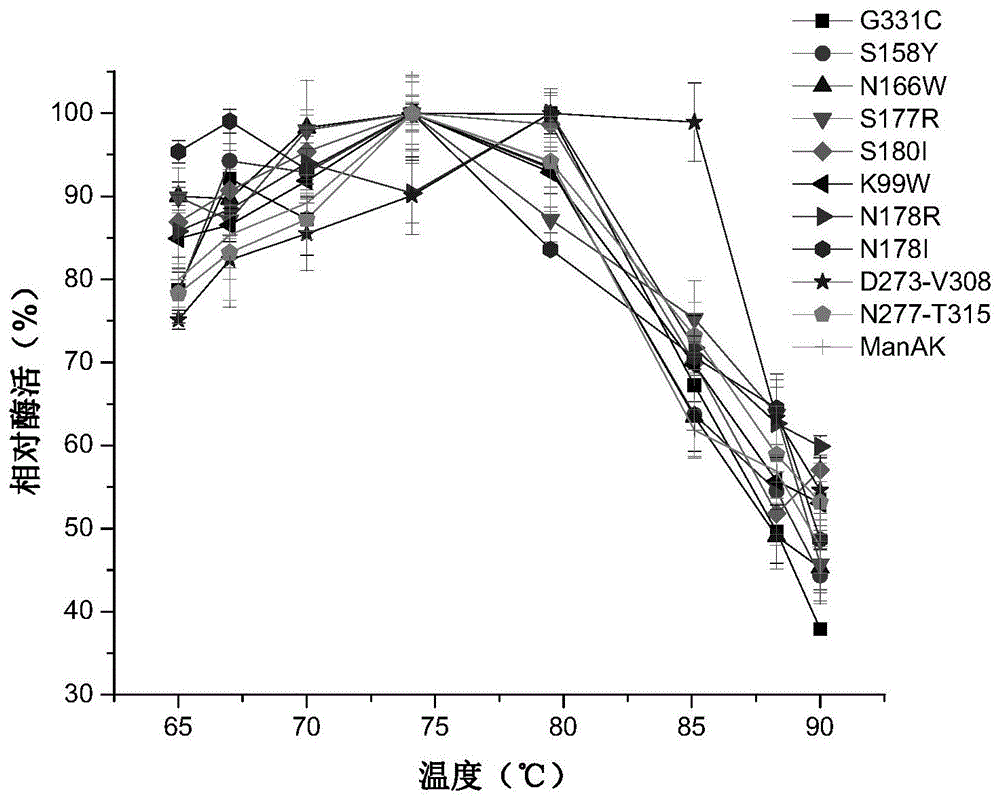
图1为β-甘露聚糖酶突变体与原始基因manak最适温度比较
图2为β-甘露聚糖酶突变体与原始基因manakt50值比较
图3为β-甘露聚糖酶突变体与原始基因manakt1/2值比较
具体实施方式
下面结合附图和具体实施例对本发明的技术方案做进一步详细的说明。以下结合实施实例对本发明做进一步说明,需要指出的是,本实施例仅用于解释本发明,而非对本发明范围的限制。
实验材料和试剂
1.菌株和载体:
表达宿主毕赤酵母x33、表达载体ppiczαa、amp、zeocine购自invitrogen公司。
2.酶与试剂盒:
pcr酶及连接酶购买自takara公司,质粒提取试剂盒及胶回收试剂盒购自omega公司。
3.培养基:
lb培养基:1%胰蛋白胨、0.5%酵母提取物、1%nacl;
ypd培养基:1%酵母提取物、2%胰蛋白胨,2%葡萄糖;
md培养基:1.34%ynb,0.4mg/l生物素,2%葡萄糖;
bmgy培养基:1%酵母提取物,2%蛋白胨,1.34%ynb,0.00004%biotin,1%甘油(v/v);
bmmy培养基:除以0.5%甲醇代替甘油,其余成份均与bmgy相同,ph4.0。
说明:以下实施例中未作具体说明的分子生物学实验方法,均参照《分子克隆实验指南》第三版j.萨姆布鲁克一书中所列的具体方法进行,或者按照试剂盒和产品说明书进行。
实验方法
实施例1
β-甘露聚糖酶突变体工程菌的构建
(1)表达载体的构建
以aspergilluskawachiiifo4308来源的β-甘露聚糖酶manak(核苷酸序列公开于genbankno.mn539024.1)为模板,通过常规方法在特定位点引入突变以制备人工基因。在基因的相应位点设计突变引物见表格1,利用pcr高保真酶进行扩增,与ppiczαa载体体外重组连接后热激转化大肠杆菌感受态细胞dh5α。涂布于lb+zeocin平板,37℃倒置培养,待转化子出现后挑取单克隆验证,挑选阳性转化子进行测序,测序表明序列正确的转化子用于大量制备重组质粒。
表1.突变体的引物序列
(2)在毕赤酵母中的表达
用限制性内切酶sac1进行线性化表达质粒载体dna,将100ul酵母感受态与10ul线性化载体混匀后转移到预冷的电转杯中电击转化,电转化的条件为1.5kv,5msec。电击后加入1ml山梨醇溶液,转移至1.5ml离心管中30℃温育1h。5000rpm离心5min,收集菌体涂布于ypd筛选平板上倒置30℃培养2-3天,挑取在平板上生长的转化子进行进一步的表达实验。
用灭过菌的牙签从长有转化子的板上挑取单菌落,按照编号先点到平板上,将平板置于30℃培养箱中培养2天,至菌落长出。按编号从平板上挑取转化子接种于3mlbmgy培养基中于30℃摇床培养48h,离心收集菌体,加入1ml含有1%甲醇的bmmy诱导培养基,继续30℃诱导培养,48h后取样检测各菌株上清液的酶活性,从中筛选出高甘露聚糖酶活性的转化子。
实施例2
重组甘露聚糖酶突变体在毕赤酵母中的发酵
(1)甘露聚糖酶突变体在摇瓶中大量表达
将筛选出的酶活较高的转化子接种于300mlbmgy液体培养基中,30℃200rpm振荡培养48h,进行菌体富集;4000×g离心5min,轻柔弃上清,将菌体转接到100ml含有1%甲醇的bmmy液体培养基中,30℃200rpm诱导培养72h。诱导培养期间,每间隔24h补加一次甲醇溶液,使甲醇终浓度保持在1%左右,10000×g离心10min收集上清。测定野生甘露聚糖酶及其突变体的活力。
(2)甘露聚糖酶突变体的纯化
收集摇瓶表达的甘露聚糖酶突变体上清液,首先利用10kda膜包进行脱盐和浓缩,再经阴离子交换柱层析纯化。以达到电泳纯的收集液作为表达酶学性质研究的样品。
实施例3
重组甘露聚糖酶酶的酶学性质分析
甘露聚糖酶活性检测根据中华人民共和国国家标准《gbt36861-2018》进行;甘露聚糖酶活性定义是指样品在ph为5.5,温度为37℃条件下,每分钟从浓度为3mg/ml的甘露聚糖(sigmag0753)溶液中降解释放1umol还原糖所需要的酶量为一个酶活力单位,以u表示。
采用dns法对本发明的甘露聚糖酶进行活性分析。
β-甘露聚糖酶活性的测定:吸取2ml经过适当稀释的酶液,加入到刻度试管中,再加入2ml浓度为0.6%(w/v)的甘露聚糖溶液,震荡,50℃下温育30min。加入5mldns试剂,震荡均匀,沸水浴加热10min,冷却至室温,加水定容到25ml,在540nm处测定吸光度。
(1)野生酶manak与突变体的最适温度
在柠檬酸-磷酸氢二钠缓冲体系及ph为5.5时,不同温度(65,70,75,80,85,90℃)下进行酶促反应,测定野生酶及突变体的最适温度。以最高酶活力为100%,计算相对酶活。
结果如图1所示,在ph相同的条件下,除突变体n178r和d273-v308的最适温度为80℃外,其余所有单位点和双位点突变体的最适温度均在75℃,与甘露聚糖酶manak相比,本发明提供的突变体的最适作用温度没有发生较大变化。
(2)野生酶manak与突变体的最适作用ph及ph稳定性
结果表明,除s180i突变体最适ph为3外,其余突变体的最适ph都保持与野生酶相同的ph=2。各突变酶残余酶活的变化趋势与manak保持一致,可见各突变体保持了野生型酶耐酸性及广范围ph的稳定性。
(3)野生酶manak与突变体的t50值和t1/2值
t50值和t1/2值是衡量耐热性能的两个重要指标。t50值是指按照甘露聚糖酶国家标准测定方法在测定时,酶活变为最高酶活的50%时的测定温度,温度越高耐热性能越好;t1/2值是指在同一温度处理不同时间,当酶活变为最高酶活一半时所需的时间,时间越长说明耐热性能越好。其中,后者是近年来判定耐热性能的主要指标。
结果显示(图2),野生manak酶的t50值为77.7℃,突变体中g331c和d273-v308的t50值超过80℃,分别比野生酶提高了2.8和5.4℃。而其余突变体的t50值在77.6-78.5℃之间,对高温的耐受能力为n277-t315>s158y>q166w=n178r>n178i>k99w>s177r=manak,而s180i低于野生manak酶0.1℃。由此可见g331c和d273-v308是促进甘露聚糖酶热稳定性提高的关键位点。
本文使用的“耐热性”是指即使在加热处理之后仍保持酶活性的性质,通过t1/2值的大小来体现。分别在75℃下处理5,10,15,20,25,30min,测定并计算相对酶活,结果如图3所示,根据拟合曲线计算相对酶活为50%时所需要的时间。通过拟合结果我们预测得到双位点突变体中n277-t315的耐热性能最好,在75℃条件下的t1/2值达到60.1min,其次是双位点突变体d273-v308,为57.2min。而其余突变体的t1/2值在51.5-55.5℃之间,均高于野生manak酶的32.9min。单点突变体对高温的耐受能力为s177r>n178r>n178i>g331c>k99w>s180i>q166w>s158y,表现出突变体耐热性的增强。可见分子内二硫键的引入显著提高了该酶在高温条件下稳定性。
综上,本发明以β-甘露聚糖酶manak为基础,提供了包含单点和双位点突变的甘露聚糖酶突变体。本发明确认了甘露聚糖酶中的关键耐热性位点,并证明了这些位点对于该酶高耐热性的重要性,对于研究甘露聚糖酶的热稳定机制提供了重要的线索,同时对于提高其它甘露聚糖酶的热稳定性提供了可靠的参考依据。
序列表
<110>中国海洋大学
<120>一种耐热性β-甘露聚糖酶突变体manak-2及其编码基因和应用
<160>26
<170>siposequencelisting1.0
<210>1
<211>367
<212>prt
<213>野生β-甘露聚糖酶(β-mannanase)
<400>1
thralaleuprolysalaserproalaproserserserserserser
151015
serserseralaserthrserphealaserthrserglyleuglnphe
202530
thrileaspglygluthrglytyrphealaglythrasnsertyrtrp
354045
ileglypheleuthraspaspseraspvalaspleuvalmetserhis
505560
leulysserserglyleulysileleuargvaltrpglypheasnasp
65707580
valthrthrglnproserserglythrvaltrptyrglnleuhisgln
859095
aspglylysserthrileasnthrglyalaaspglyleuglnargleu
100105110
asptyrvalvalserseralagluglnhisaspilelysleuileile
115120125
asnphevalasntyrtrpthrasptyrglyglymetseralatyrval
130135140
seralatyrglyglyseraspgluthraspphetyrthrseraspthr
145150155160
metglnseralatyrglnthrtyrilelysthrvalvalgluargtyr
165170175
serasnserseralavalphealatrpgluleualaasngluproarg
180185190
cysprosercysaspthrservalleutyrasptrpileglulysthr
195200205
serlyspheilelysglyleuaspalaasphismetvalcysilegly
210215220
aspgluglypheglyleuasnthraspseraspglysertyrprotyr
225230235240
glnphealagluglyleuasnphethrlysasnleuglyileaspthr
245250255
ileasppheglythrleuhisleutyrproaspsertrpglythrser
260265270
aspasptrpglyasnglytrpileseralahisglyalaalacyslys
275280285
alaalaglylysprocysleuleugluglutyrglyvalthrserasn
290295300
hiscysservalgluserprotrpglnlysthralaleuasnthrthr
305310315320
glyvalseralaaspleuphetrpglntyrglyaspaspleuserthr
325330335
glylysserproaspaspglyasnthriletyrtyrglythrserasp
340345350
tyrglucysleuvalthrasphisvalalaalaileglyserala
355360365
<210>2
<211>1104
<212>dna
<213>野生β-甘露聚糖酶(β-mannanase)
<400>2
actgctttgccaaaggcttctccagctccatcttcttcttcttcttcctcttcttctgct60
tctacttcttttgcttctacttctggtttgcaatttactattgatggtgaaactggttac120
tttgctggtactaactcttactggattggttttcttactgatgattctgatgttgatttg180
gttatgtctcatttgaagtcttctggtttgaagattttgagagtttggggttttaacgat240
gttactactcaaccatcttctggtactgtttggtaccaattgcatcaagatggtaagtct300
actattaacactggtgctgatggtttgcaaagattggattacgttgtttcttctgctgaa360
caacatgatattaagttgattattaactttgttaactactggactgattacggtggtatg420
tctgcttacgtttctgcttacggtggttctgatgaaactgatttttacacttctgatact480
atgcaatctgcttaccaaacttacattaagactgttgttgaaagatactctaactcttct540
gctgtttttgcttgggaattggctaacgaaccaagatgtccatcttgtgatacttctgtt600
ttgtacgattggattgaaaagacttctaagtttattaagggtttggatgctgatcatatg660
gtttgtattggtgatgaaggttttggtttgaacactgattctgatggttcttacccatac720
caatttgctgaaggtttgaactttactaagaacttgggtattgatactattgattttggt780
actttgcatttgtacccagattcttggggtacttctgatgattggggtaacggttggatt840
tctgctcatggtgctgcttgtaaggctgctggtaagccatgtttgttggaagaatacggt900
gttacttctaaccattgttctgttgaatctccatggcaaaagactgctttgaacactact960
ggtgtttctgctgatttgttttggcaatacggtgatgatttgtctactggtaagtctcca1020
gatgatggtaacactatttactacggtacttctgattacgaatgtttggttactgatcat1080
gttgctgctattggttctgcttaa1104
<210>3
<211>45
<212>dna
<213>人工序列(artificialsequence)
<400>3
ttggcaatactgtgatgatttgtctactggtaagtctccagatga45
<210>4
<211>42
<212>dna
<213>人工序列(artificialsequence)
<400>4
acaaatcatcacagtattgccaaaacaaatcagcagaaacac42
<210>5
<211>47
<212>dna
<213>人工序列(artificialsequence)
<400>5
tttttacacttacgatactatgcaatctgcttaccaaacttacatta47
<210>6
<211>44
<212>dna
<213>人工序列(artificialsequence)
<400>6
gcatagtatcgtaagtgtaaaaatcagtttcatcagaaccaccg44
<210>7
<211>54
<212>dna
<213>人工序列(artificialsequence)
<400>7
atctgcttactggacttacattaagactgttgttgaaagatactctaactcttc54
<210>8
<211>49
<212>dna
<213>人工序列(artificialsequence)
<400>8
taatgtaagtccagtaagcagattgcatagtatcagaagtgtaaaaatc49
<210>9
<211>39
<212>dna
<213>人工序列(artificialsequence)
<400>9
tgaaagatacagaaactcttctgctgtttttgcttggga39
<210>10
<211>48
<212>dna
<213>人工序列(artificialsequence)
<400>10
cagaagagtttctgtatctttcaacaacagtcttaatgtaagtttggt48
<210>11
<211>36
<212>dna
<213>人工序列(artificialsequence)
<400>11
ctctaactctattgctgtttttgcttgggaattggc36
<210>12
<211>52
<212>dna
<213>人工序列(artificialsequence)
<400>12
caaaaacagcaatagagttagagtatctttcaacaacagtcttaatgtaagt52
<210>13
<211>44
<212>dna
<213>人工序列(artificialsequence)
<400>13
tcaagatggttggtctactattaacactggtgctgatggtttgc44
<210>14
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>14
taatagtagaccaaccatcttgatgcaattggtaccaaac40
<210>15
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>15
aagatactctagatcttctgctgtttttgcttgggaat38
<210>16
<211>51
<212>dna
<213>人工序列(artificialsequence)
<400>16
cagcagaagatctagagtatctttcaacaacagtcttaatgtaagtttggt51
<210>17
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>17
aagatactctatttcttctgctgtttttgcttgggaat38
<210>18
<211>51
<212>dna
<213>人工序列(artificialsequence)
<400>18
cagcagaagaaatagagtatctttcaacaacagtcttaatgtaagtttggt51
<210>19
<211>38
<212>dna
<213>人工序列(artificialsequence)
<400>19
gggtacttcttgtgattggggtaacggttggatttctg38
<210>20
<211>43
<212>dna
<213>人工序列(artificialsequence)
<400>20
taccccaatcacaagaagtaccccaagaatctgggtacaaatg43
<210>21
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>21
ccattgttcttgtgaatctccatggcaaaagactgctttg40
<210>22
<211>46
<212>dna
<213>人工序列(artificialsequence)
<400>22
atggagattcacaagaacaatggttagaagtaacaccgtattcttc46
<210>23
<211>37
<212>dna
<213>人工序列(artificialsequence)
<400>23
tgattggggttgtggttggatttctgctcatggtgct37
<210>24
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>24
gaaatccaaccacaaccccaatcatcagaagtaccccaag40
<210>25
<211>41
<212>dna
<213>人工序列(artificialsequence)
<400>25
atggcaaaagtgtgctttgaacactactggtgtttctgctg41
<210>26
<211>40
<212>dna
<213>人工序列(artificialsequence)
<400>26
tgttcaaagcacacttttgccatggagattcaacagaaca40
- 还没有人留言评论。精彩留言会获得点赞!