通过临近使能反应疗法开发的共价蛋白质药物
序列表
本申请与电子形式的序列表一起提交。序列表的全部内容通过引用并入本文。
本申请总体上涉及用于生成共价蛋白质药物的临近使能反应疗法方法,以及包含一个潜在生物反应性非天然氨基酸(uaa)突变蛋白质,所述突变蛋白质可用于通过该临近使能反应疗法(proximity-enabledreactivetherapeutics)开发的共价蛋白质药物。更具体地,本申请涉及包含o-氟硫酸-l-酪氨酸(fluorosulfate-l-tyrosine,fsy)的突变的pd-1,其制备方法,和所述突变的pd-1的应用。
背景技术:
小分子药物与其靶标以共价方式结合,尽管在历史上被药学研究人员和工业有意地回避,但在近年内以巨大的成功再度崛起1,2,3。药物和靶标之间的共价键合相比常规药物的非共价相互作用提供了多种期望的特性,包括增加的生物化学效力和功效,使药物代谢动力学持久的延长的作用持续时间,具有减小的剂量和给药频次的提高的治疗指数,使靶标完全失活,抑制不易治愈的靶标的机会3,4。到目前为止,几乎30%的市场上销售的靶向酶的药物通过共价机制起作用,并且靶向共价抑制逐渐被应用于更宽范围的对非共价药物来说有挑战性的靶标5,4。
对于治疗挑战性的疾病来说,小分子共价药物相对于非共价药物提供多种期望的治疗特性。然而,由于蛋白质不能共价结合靶标,共价蛋白质药物的潜力仍然有大部分没有被开发出来。与小分子共价药物的顺利发展相反,共价蛋白质药物的治疗潜力仍然有大部分没有得到开发利用。蛋白质药物比小分子药物更适合于共价作用方式。小分子共价药物的一个主要问题是脱靶反应性1,脱靶反应性可以在共价蛋白质药物中被最小化,因为蛋白质具有较大的尺寸,并且通常具有比小分子更高的靶标特异性。开发共价小分子药物的关键是在反应性和选择性之间采取适宜的平衡4。开发共价蛋白质药物的挑战如下:天然蛋白质主要通过非共价相互作用进行相互作用并且对靶标上的天然氨基酸残基缺乏共价反应性。另外,氨基酸在蛋白质、细胞和体内的广泛存在对于实现针对靶标的反应选择性施加了巨大的障碍。
因此,在本领域中对于开发突变蛋白一直存在着需求,所述突变蛋白能够通过与在患病细胞诸如肿瘤细胞上表达的靶蛋白上的天然氨基酸的共价相互作用而进行相互作用。
技术实现要素:
本发明的目的是制备能够通过与肿瘤上表达的靶蛋白上的天然氨基酸共价相互作用而相互作用的突变蛋白质。所述突变蛋白质可以用来开发共价蛋白质药物。
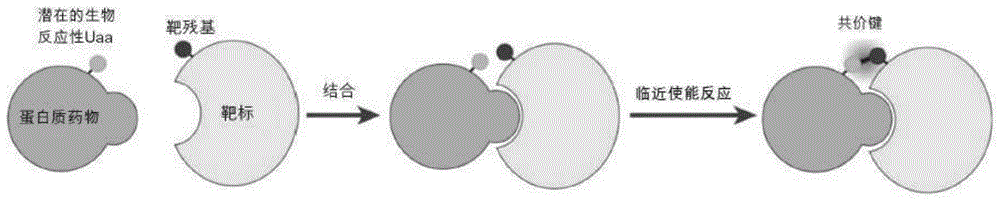
在此本申请公开了一种用于开发共价蛋白质药物的临近使能反应疗法策略(图1)。通过扩增遗传密码6,7,潜在的生物反应性非天然氨基酸(uaa)被引入至蛋白质药物中,所述uaa具有低的化学反应性,在蛋白质内部和细胞中保持惰性。在所述蛋白质药物与其靶标结合后,生物反应性的uaa将通过临近使能反应促进与靶标的天然残基选择性地反应8,在所述药物和所述靶标之间形成共价键。在一个实施方案中,发明人通过遗传方式将潜在的生物反应性uaa,即o-氟硫酸-l-酪氨酸(fsy)9,掺入到人程序性死亡蛋白1(pd-1)中,并且表明得到的pd-1(fsy)在体外与细胞表面上的和在体内与肿瘤组织中的其天然配体pd-l1共价结合。与pd-1(wt)相比,发现pd-1(fsy)增强人幼稚t细胞的功能活性,并在人源化小鼠模型中以显著更高的功效抑制肿瘤生长,显示了与治疗性抗-pd-l1单克隆抗体相当的抗肿瘤效应,突显了由共价键所促进的疗效。
在一个方面,本公开涉及用以生成共价蛋白质药物的临近使能反应疗法方法,所述方法包括:将潜在的生物反应性非天然氨基酸(uaa)掺入到蛋白质药物中以生成突变蛋白质药物,其中所述突变蛋白质药物在药物-靶标结合后通过临近使能反应与其靶蛋白的靶天然残基反应,由此使得所述蛋白质药物能够与其靶标共价结合。
在一个实施方案中,uaa经遗传方式掺入到所述蛋白质药物中,例如借助于对uaa特异的突变的氨酰-trna合成酶(aminoacyl-trnasynthetase,aars)通过在编码所述蛋白质药物的野生型蛋白的核苷酸序列中的期望位点处引入琥珀终止密码子(tag),所述对uaa特异的突变的氨酰-trna合成酶(aars)被进化为响应于tag密码子而掺入uaa。
在一个实施方案中,所述uaa是fsy(o-氟硫酸-l-酪氨酸),借助于用于正交trnapyl/fsyrs对的基因,通过在编码所述蛋白质药物的野生型蛋白的核苷酸序列中的期望位点处引入琥珀终止密码子(tag)经遗传方式将其引入到蛋白质药物中,所述用于正交trnapyl/fsyrs对的基因被进化为响应于tag密码子而掺入fsy。
在一个实施方案中,所述蛋白质药物可以是免疫检查点的一个成员,并且其靶标,即,免疫检查点的另一成员,可以是在肿瘤细胞表面上表达的蛋白质。
在一个实施方案中,所述免疫检查点可以选自,但不限于,pd-1/pd-l1、ctla-4/cd80(cd86)、4-1bb/配体、ox40/配体、gitr/配体、lag-3/配体、tim-3/配体,等等。
在一个实施方案中,所述突变蛋白质药物是在其氨基酸序列中含有一个fsy(o-氟硫酸-l-酪氨酸)的突变的pd-1(fsy),其中所述突变的pd-1(fsy)能够与其靶pd-l1中的接近的选自tyr、his或lys的一个靶天然氨基酸残基通过点击化学硫-氟化物交换(sufex)特异性地反应。
在一个方面,本公开涉及在其氨基酸序列中包含一个fsy(o-氟硫酸-l-酪氨酸)的突变的人pd-1(fsy)。
在一个实施方案中,所述突变的pd-1(fsy)中的所述fsy残基能够与其靶pd-l1中的接近的选自tyr、his或lys的一个靶天然氨基酸残基通过点击化学硫-氟化物交换(sufex)特异性地反应。fsy的使用将在pd-1(fsy)和pd-l1之间生成共价键,形成具有极大亲和力的不可逆拮抗剂,这种极大亲和力是利用通过天然氨基酸诱变进化的任意高亲和力突变体都无法达到的。
在一个实施方案中,所述fsy在下列位点中之一处引入至人pd-1中:75、77或129(根据野生型人pd-1(即,ncbi登录号:np_005009.2)的氨基酸序列编号),分别靶向pd-l1的lys124、lys124和his69。
在一个实施方案中,所述fsy在位点75或129处,优选在位点129处引入至人pd-1中。
在一个实施方案中,所述fsy残基通过在宿主细胞中表达包含在期望位点处引入的琥珀终止密码子(tag)的pd-1基因与用于正交trnapyl/fsyrs对的基因而掺入到人pd-1的胞外结构域中,所述用于正交trnapyl/fsyrs对的基因被进化为响应于tag密码子而掺入fsy。
在一个实施方案中,所述宿主细胞可以选自,但不限于,大肠杆菌(e.coli)细胞、酵母细胞、哺乳动物细胞(例如,cho细胞)或昆虫细胞。
在一个实施方案中,所述突变的人pd-1(fsy)可以在体外与人pd-l1特异性地共价结合,与在活的癌细胞的表面上生理性地表达的人pd-l1特异性地共价结合,以及在体内与肿瘤组织中的人pd-l1特异性地共价结合。然而,所述突变的人pd-1(fsy)表现出与小鼠pd-l1没有共价交联,尽管人pd-1已知与小鼠pd-l1交叉结合。因此,所述突变的人pd-1(fsy)具有高靶标特异性和低脱靶效应。
在一个实施方案中,所述突变的人pd-1(fsy)可以通过阻断pd-1/pd-l1相互作用来显著地增强t细胞激活。人pd-1(fsy)的t细胞激活效应与以相同摩尔浓度应用的阿特珠单抗的t细胞激活效应相当。
在一个实施方案中,所述突变的人pd-1(fsy)可以达到与以相同摩尔量施用的阿特珠单抗相同的抗肿瘤效应,但需要的pd-1(fsy)的质量量为阿特珠单抗的十分之一。
在一个方面,本公开涉及一种经遗传方式将fsy掺入至pd-1中的方法,所述方法包括:
在包含合成的fsy和用于诱导蛋白质表达的试剂的细胞培养基中,在宿主细胞中表达:
(i)包含在期望位点处引入的琥珀终止密码子(tag)的pd-1基因;和
(ii)用于正交trnapyl/fsyrs对的基因。
在一个实施方案中,所述宿主细胞是大肠杆菌,且用于诱导蛋白质表达的试剂是iptg。
在一个实施方案中,所述pd-1是人pd-1。
在一个实施方案中,所述琥珀终止密码子(tag)在选自人pd-1的位点75、77或129中的一个位点处引入。
在一个实施方案中,包含琥珀终止密码子(tag)的所述pd-1基因如seqidno:16-18中任一个所示。
在一个实施方案中,所述trnapyl如seqidno:19所示。
在一个实施方案中,所述fsyrs是指对fsy特异的突变的吡咯赖氨酰基-trna合成酶(pylrs)。fsyrs如seqidno:20所示并且是突变的马氏甲烷球菌(methanosarcinamazei)pylrs。
在一个方面,本公开涉及一种药物组合物,其包含有效量的包含潜在的生物反应性非天然氨基酸(uaa)的突变蛋白质和药学上可接受的载体和/或赋形剂。
在一个实施方案中,本公开涉及一种药物组合物,其包含有效量的突变的pd-1(fsy)和药学上可接受的载体和/或赋形剂。
在一个实施方案中,在突变的pd-1(fsy)中,所述fsy在下列位点中的一个处被引入至pd-1中:75、77或129(所述位点的编号依照野生型pd-1的氨基酸序列)。
在一个实施方案中,所述fsy在位点75或129处、优选在位点129处引入至pd-1中。
当将包含突变的pd-1(fsy)的药物组合物施用于有需要的受试者时,在pd-1(fsy)和pd-l1之间将生成共价键,形成具有极大亲和力的不可逆拮抗剂,这种极大亲和力是利用通过天然氨基酸诱变进化的任意高亲和力突变体都无法达到的。
在一个实施方案中,所述药物组合物用来预防和/或治疗pd-l1相关疾病,例如,黑色素瘤、肾细胞癌、头颈部癌、宫颈癌、成胶质细胞瘤、膀胱癌、食管癌、乳腺癌、肝细胞癌、霍奇金淋巴瘤和纵膈大b细胞淋巴瘤。
在一个方面,本公开涉及包含潜在的生物反应性非天然氨基酸(uaa)的突变蛋白质在制造用于预防和/或治疗疾病的药物组合物中的用途,所述疾病包括癌症。
在一个实施方案中,本公开涉及突变的pd-1(fsy)在制造用于预防和/或治疗pd-l1相关疾病的药物组合物中的用途,所述pd-l1相关疾病例如,黑色素瘤、肾细胞癌、头颈部癌、宫颈癌、成胶质细胞瘤、膀胱癌、食管癌、乳腺癌、肝细胞癌、霍奇金淋巴瘤和纵膈大b细胞淋巴瘤。
在一个实施方案中,所述药物组合物被包装成试剂盒。
在一个方面,本公开涉及试剂盒,所述试剂盒包含有效量的包含潜在的生物反应性非天然氨基酸(uaa)的突变蛋白质和药学上可接受的载体和/或赋形剂。
在一个实施方案中,所述试剂盒包含有效量的突变的pd-1(fsy)和药学上可接受的载体和/或赋形剂。
在一个方面,本公开涉及一种预防和/或治疗受试者中的疾病的方法,所述疾病包括癌症,所述方法包括:向所述受试者施用包含有效量的包含潜在的生物反应性非天然氨基酸(uaa)的突变蛋白质和药学上可接受的载体和/或赋形剂的药物组合物。
在一个实施方案中,本公开涉及一种预防和/或治疗受试者中的pd-l1-相关疾病的方法,所述方法包括:向所述受试者施用包含有效量的突变的pd-1(fsy)和药学上可接受的载体和/或赋形剂的药物组合物。
与现有技术相比,本申请具有下列优势:
(1)临近使能反应策略可以用于开发具有改善的功效和体内半衰期的共价蛋白质药物;
(2)临近使能反应可以用于从小蛋白质分子开发共价蛋白质药物;
(3)所述共价蛋白质药物因其改善的功效和体内半衰期而具有低剂量和低频次;以及
(4)所述临近使能反应策略用来修饰免疫检查点的一个成员,所述免疫检查点例如pd-1/pd-l1、ctla-4/cd80(cd86)、4-1bb/配体、ox40/配体、gitr/配体、lag-3/配体、tim-3/配体等,并且因此,修饰的蛋白质(例如,pd-1)可以特异性地结合在肿瘤细胞表面上表达的其靶配体(例如,pd-l1)。
前面是概述,因此必然包含对细节的简化、归纳和省略;因此,本领域技术人员要理解该概述仅是说明性的,并且不意在以任何方式限制。在本文所给出的教导中,本文所述的方法、组合物和/或装置和/或其他主题的其他方面、特征和优势将是很显然的。提供所述概述是为了以简化的形式引入各种概念,在下面的详述中进一步描述这些概念。此概述不意在识别所要求保护的主题的关键特征或必要特征,也不意在用作确定所要求保护的主题的范围的辅助。此外,在整个本申请中引用的所有参考文献、专利和公开的专利申请的内容通过引用全部并入本文。
附图说明
图1示出了利用临近使能反应开发共价蛋白质药物的原理。潜在的生物反应性uaa经遗传方式掺入到蛋白质药物中,在药物-靶标结合后,uaa经由临近使能反应与所述靶蛋白的靶天然残基反应,由此使得所述蛋白质药物与其靶标共价结合。
图2示出了将潜在的生物反应性uaafsy遗传掺入到pd-1中。
(a)潜在的生物反应性uaafsy的结构以及其与his经sufex的反应。
(b)人pd-1/pd-l1复合物的晶体结构。选择用于fsy掺入的pd-l的位点a129、q75和d77以及它们的靶天然残基pd-l1的his69和lys124以棒状物显示。pdb编码4zqk。
(c)大肠杆菌细胞的细胞裂解物在指示的不同表达条件下的蛋白质印迹分析。iptg用来诱导pd-1基因的表达,阿拉伯糖用来诱导fsyrs基因的表达。当在生长培养基中添加fsy时表达pd-1igv结构域,并在与pd-1(wt)igv结构域相同的位置处迁移。在生长培养基中不添加fsy,则翻译在129tag密码子处终止,因此不表达在期望的位置处的pd-1igv结构域。
(d)pd-1(wt)和pd-1(a129fsy)表达和纯化的sds-page分析。纯化和重折叠的pd-1(a129fsy)在与pd-1(wt)相同的位置处迁移。
(e)pd-1(a129fsy)的串联质谱。一系列b和y离子明确地指示fsy在pd-1的tag指定的129位置处掺入。u代表fsy。
图3示出了经由潜在的生物反应性uaafsy的临近使能反应在pd-1(fsy)与其配体pd-l1之间的共价键合。
(a)使用sds-page和蛋白质印迹在体外对纯化的pd-1突变体与pd-l1的结合的分析。pd-1/pd-l1的共价复合物以红色箭头指示。
(b)在pd-l1中his69突变为ala消除了其与pd-1(a129fsy)的共价结合。
(c)串联质谱明确地指示pd-1中的fsy129与pd-l1的his69共价交联。u代表fsy。
(d)pd-1(a129fsy)标记为pd-1(fsy),其以剂量依赖方式与癌细胞表面上的pd-l1共价结合。
(e)pd-1(a129fsy)标记为pd-1(fsy),其在体内与肿瘤上的pd-l1共价结合。
图4示出了pd-1(fsy)不与小鼠胸腺和脾细胞上表达的小鼠pd-l1共价交联。
(a)pd-1(wt)和pd-1(fsy)单独地与小鼠细胞孵育,将样品裂解并使用对小鼠pd-l1特异的一抗通过蛋白质印迹进行分析。
(b)人pd-l1和小鼠pd-l1的胞外igv结构域的序列比对。红色盒子强调了人pd-l1中的his69与小鼠pd-l1中的ala69对应。fsy通过临近使能反应与his反应但不与ala反应。因此,pd-1(fsy)与人pd-l1共价交联,但不与小鼠pd-l1共价交联。
图5示出了在同种异体刺激后在体外pd-1(fsy)增加了t细胞增殖和inf-γ产生。
(a)通过测量cfse染色强度获得的t细胞增殖的facs分析迹线。
(b)通过分裂的t细胞(分裂的t细胞的cfse强度低于亲代群体)百分比测量的t细胞增殖。n=4;*p<0.05;****p<0.0001;单因素anova,接着tukey多重比较检验。
(c)在mlr中由t细胞释放的inf-γ。n=8;ns,不显著;*p<0.05;****p<0.0001;单因素anova,接着tukey多重比较检验。
图6示出了在pbmc-肿瘤异种移植物小鼠模型中pd-1(fsy)比pd-1(wt)更有效地抑制肿瘤生长。
(a)说明实验设计的示意图。
(b-c)移植的肿瘤的生长曲线显示为单个肿瘤(b)或综合数据(c)。误差棒表示为s.e.m.(标准误),n=8/组。对于媒介物pbs对照,n=6。**p<0.01;****p<0.0001;双因素anova,接着tukey多重比较检验。
(d)从在第44天处死的小鼠切除的肿瘤的照片。
(e)切除的肿瘤的重量比较。误差棒表示为s.e.m.(标准误),n=8/组(例外是pbs组,n=6)。ns,不显著;**p<0.01;***p<0.001;单因素anova,接着tukey多重比较检验。
图7示出了来源于单核细胞的成熟人树突细胞的表型特征。在从用gm-csf和il-4诱导并用tnf-α和lps激活的单核细胞分化的成熟dc上cd83、cd86、hla-dc和pd-l1的表达增加。未被染色的cd83、cd86、hla-dr和pd-l1用作对照。
图8示出了在同种异体刺激后在体外pd-1(fsy)增加t细胞增殖。测量并比较在用指示的试剂处理的cd3+t细胞中cfse的平均荧光强度(mfi)。cfse荧光强度减小表示细胞分裂和增殖。pd-1(fsy)显示在50nm和250nm的浓度时比pd-1(wt)更显著地增强t细胞增殖,达到了与抗体阿特珠单抗相同的增强水平。n=4;ns,不显著;****p<0.0001;单因素anova,接着tukey多重比较检验。
具体实施方式
尽管本公开可以以多种不同的形式体现,但本文公开的是举例说明本公开的原理的其具体的说明性实施方案。应该强调本公开不限于所举例说明的具体实施方案。而且,本文使用的任何章节标题仅用于组织目的,并且不应被解释为限制所述的主题。
除非本文中另外限定,否则与本公开结合使用的科学和技术术语应当具有本领域普通技术人员普遍理解的含义。此外,除非上下文另外需要,否则单数术语应当包括复数并且复数术语应当包括单数。更具体地,如在本说明书和附带的权利要求书中所用,单数“一个/种”和“所述”包括复数指代物,除非上下文清晰地指示另外的含义。因此,例如,提到“一种蛋白质”包括多种蛋白质;提到“一种细胞”包括细胞的混合物,等等。在本申请中,“或”的使用意指“和/或”,除非另外指明。此外,术语“包括”以及其他形式诸如“包含和含有”的使用是非限制性的。另外,在说明书和附带的权利要求书中提供的范围包括两个端点以及端点之间的所有点。
通常,结合本文所述的细胞和组织培养、分子生物学、免疫学、微生物学、遗传学以及蛋白质和核酸化学和杂交使用的术语以及它们的技术是本领域众所周知和普遍使用的。除非另外指示,否则本公开的方法和技术一般是根据本领域中众所周知的常规方法执行的并且如在整个本说明书中引用和讨论的各篇一般的和更具体的参考文献中所述。参见,例如,abbas等人,cellularandmolecularimmunology,第6版,w.b.saunderscompany(2010);sambrookj.&russelld.molecularcloning:alaboratorymanual,第3版,coldspringharborlaboratorypress,coldspringharbor,n.y.(2000);ausubel等人,shortprotocolsinmolecularbiology:acompendiumofmethodsfromcurrentprotocolsinmolecularbiology,wiley,john&sons,inc.(2002);harlow和laneusingantibodies:alaboratorymanual,coldspringharborlaboratorypress,coldspringharbor,n.y.(1998);和coligan等人,shortprotocolsinproteinscience,wiley,john&sons,inc.(2003)。结合本文所述的分析化学、合成有机化学以及医学和药物化学使用的术语以及它们的实验室操作和技术是本领域众所周知和普遍使用的。而且,本文使用的任何章节标题仅用于组织目的,并且不应被解释为限制所述的主题。
定义
为了更好地理解本公开,相关术语的定义和解释提供如下。
术语“共价蛋白质药物”当在本文中使用时是指能够通过共价键合与其靶标相互作用的治疗性蛋白质。
术语“非天然氨基酸(uaa)”是指与20种天然氨基酸中的任一种均不同的氨基酸。uaa可以是修饰的天然氨基酸,例如,o-氟硫酸-l-酪氨酸(fsy)是一种修饰的酪氨酸。
术语“临近使能反应疗法”是指包含潜在的生物反应性非天然氨基酸(uaa)的突变蛋白质药物,其中所述突变蛋白质药物中的uaa可以与所述突变蛋白质药物的靶蛋白的一个靶天然残基特异性地反应,使得所述蛋白质药物与其靶标形成共价结合。
术语“程序性死亡1(pd-1)”,作为一种免疫检查点蛋白,是i型跨膜蛋白,它是ig基因超家族的一部分(agata等人(1996)bitimmunol8:765-72)且是与cd28具有同源性的免疫球蛋白超家族的抑制性成员。其在t细胞、激活的b细胞和髓样细胞上表达(agata等人,见前;okazaki等人(2002)curr.opin.immunol.14:391779-82;bennett等人(2003)jimmunol170:711-8),且在调节免疫系统的刺激性和抑制性信号中具有关键地位(okazaki,taku等人2007internationalimmunology19:813-824)。通过在凋亡细胞中筛选差异表达发现了pd-1(ishida等人(1992)emboj11:3887-95)。pd-1的备选名称或同义词包括pdcd1、pd1、cd279和sleb2等。人pd-1的代表性氨基酸序列由ncbi登录号:np_005009.2公开,且编码人pd-1的代表性核酸序列以ncbi登录号:nm_005018.2显示。
如本文所用,术语“pd-l1”是指程序性细胞死亡配体1(pd-l1,参见,例如,freeman等人(2000)j.exp.med.192:1027)。pd-l1的备选名称或同义词包括pdcd1l1、pdl1、b7h1、cd274和b7-h等。人pd-l1的代表性氨基酸序列以ncbi登录号:np_054862.1公开,且编码人pd-l1的代表性核酸序列以ncbi登录号:nm_014143.3显示。pd-l1在胎盘、脾、淋巴结、胸腺、心脏、胎肝中表达,也见于许多肿瘤或癌细胞中。pd-l1与其受体pd-1或b7-1结合,pd-1或b7-1在激活的t细胞、b细胞和髓样细胞上表达。pd-l1和其受体的结合诱导信号转导以抑制tcr-介导的对细胞因子产生和t细胞增殖的激活。因此,pd-l1在特定的事件(诸如妊娠、自身免疫病、组织同种异体移植)期间抑制免疫系统中具有主要地位,并且据信使肿瘤或癌细胞绕过免疫检查点并逃避免疫应答。
在激活的t细胞上表达的pd-1和在肿瘤细胞上表达的pd-l1之间的相互作用负性地调控免疫应答并限制抗肿瘤免疫性。pd-l1大量存在于多种人癌症中(dong等人(2002)nat.med8:787-9)。
如本文所用,术语“载体”是指其中可以插入多核苷酸的核酸运载体。当所述载体允许其中插入的多核苷酸所编码的蛋白质表达时,所述载体称为表达载体。载体可以通过转化、转导或转染到宿主细胞中而携带有在宿主细胞中表达的遗传材料元件。载体是本领域技术人员所熟知的,包括,但不限于,质粒,噬菌体,粘粒,人工染色体诸如酵母人工染色体(yac)、细菌人工染色体(bac)或p1-衍生的人工染色体(pac);噬菌体诸如λ噬菌体或m13噬菌体,和动物病毒。可以用作载体的动物病毒包括,但不限于,逆转录病毒(包括慢病毒)、腺病毒、腺相关病毒、疱疹病毒(诸如单纯疱疹病毒)、痘病毒、杆状病毒、乳头瘤病毒、乳多泡病毒(诸如sv40)。载体可以包括用于控制表达的多个元件,包括,但不限于,启动子序列、转录起始序列、增强子序列、选择元件和报告基因。另外,载体可以包括复制起始点。
如本文所用,术语“宿主细胞”是指可以被改造以生成感兴趣的蛋白质、蛋白质片段或肽的细胞系统。宿主细胞包括,但不限于,培养细胞,例如来源于啮齿动物(大鼠、小鼠、豚鼠或仓鼠)的哺乳动物培养细胞,诸如cho、bhk、nso、sp2/0、yb2/0;或人组织或杂交瘤细胞、酵母细胞和昆虫细胞,以及包含在转基因动物或培养组织内的细胞。该术语不仅涵盖特定的受试者细胞,还包括此类细胞的后代。因为因突变或环境影响在后续代中可以发生某些修饰,因此此类后代可以与亲代细胞不相同,但仍然包括在术语“宿主细胞”的范围内。
术语“受试者”包括任何人类或非人动物,优选人类。
如本文所用,术语“癌症”是指肿瘤或恶性细胞生长、增殖或转移介导的实体肿瘤或非实体肿瘤(诸如白血病)中的任一种,并且引起医学病况。
术语“治疗”如本文所用,在治疗病况的情形中,一般关于治疗和疗法,无论是人还是动物,其中达到一些期望的治疗效应,例如,抑制所述病况的进展,并且包括进展速度的减小,进展速度的停止,所述病况的消退,所述病况的改善和所述病况的治愈。还包括作为预防措施(即,预防法、预防)的治疗。对于癌症,“治疗”可以指遏制或减缓肿瘤或恶性细胞生长、增殖或转移,或其一些组合。对于肿瘤,“治疗”包括去除所有或部分肿瘤,抑制或减缓肿瘤生长和转移,预防或延缓肿瘤的发展,或其一些组合。
如本文所用,术语“有效量”关于活性化合物或包含活性化合物的材料、组合物或剂型的量,该量当根据期望的治疗方案施用时有效地用于产生与合理的收益/风险比相称的一些期望的治疗效应。
术语“预防”如本文所用,关于哺乳动物中的特定疾病,是指预防或减缓疾病的发作,或预防其临床表现或亚临床症状
如本文所用,术语“药学上可接受的”意指媒介物、稀释剂、赋形剂和/或其盐类,与制剂中的其他成分在化学上和/或物理上是相容的,并且与接受者生理上相容。
如本文所用,术语“药学上可接受的载体和/或赋形剂”是指与受试者和活性剂在药理学上和/或生理上相容的载体和/或赋形剂,它们在本领域中是众所周知的(参见,例如,remington'spharmaceuticalsciences.gennaroar编,第19版,pennsylvania:mackpublishingcompany,1995),并且包括,但不限于ph调节剂、表面活性剂、辅料和离子强度增强剂。例如,ph调节剂包括,但不限于,磷酸盐缓冲液;表面活性剂包括,但不限于,阳离子、阴离子或非离子表面活性剂,例如,tween-80;离子强度增强剂包括,但不限于,氯化钠。
序列表概述
本申请附带了序列表,包括一些氨基酸序列和核苷酸序列。下表a提供了所包括的序列的概述。
表a
实施例
参照下列的实施例将更容易理解由此一般描述的本公开,所述实施例通过举例说明的方式提供,不意在限制本公开。所述实施例不意在表示下面的实验是全部进行的实验或仅能进行的实验。
关于在实施例中所使用的市售材料的信息在表1中提供。除非另外指定,否则在实验中使用的试剂和其他材料均是市售的。
实施例1
人pd-1和pd-l1胞外igv结构域的克隆
通过将人pd-1(hpd-1)igv结构域编码基因(残基32-160,cys93替换为ser(c93s)以辅助蛋白质稳定性28)克隆进带有羧基端6x组氨酸标签的pet-26b载体中生成质粒pet-26b-pd1(wt)(seqidno:15和22)。简言之,利用下列的引物对pd-1wt-f和pd-1wt-r通过q5高保真dna聚合酶(neb,目录号m0491)从质粒puc57-kan-hpd-1igv结构域(32-160,c93s)扩增hpd-1igv结构域(32-160,c93s)的dna,所述质粒进行了密码子优化并由genewiz(china,suzhou)合成。pcr产物用ndei-hf和xhoi-hf消化,并使用t4dna连接酶(neb,目录号m0202l)连接到预切的pet-26b载体中。
pd-1wt-f:gttagactcatatgtggaatccgccgacctttagc(seqidno:1)
pd-1wt-r:actgctcgagcggactaggactcggatgtgcg(seqidno:2)
为了在位点d77或a129处引入tag密码子,利用q5高保真dna聚合酶并使用质粒pet-26b-pd1(wt)作为模板分别使用下列的引物对进行重叠pcr。扩增的pcr产物用ndei-hf和xhoi-hf消化,然后连接到预切的pet-26b载体中得到质粒pet-26b-pd-1(d77tag)或pet-26b-pd-1(a129tag)。
d77-f1:cctcatatgtggaatccgccgacctttagc(seqidno:3)
d77-r1:ggtctggttgctcgggctcatgc(seqidno:4)
d77-f2:cccgagcaaccagacctagaaactggccgcctttcc(seqidno:5)
d77-r2:ttagcagccggatctcagtggtgg(seqidno:6)a129-f1(与d77-f1相同):cctcatatgtggaatccgccgacctttagc(seqidno:3)
a129-r1:tgcaggctaatggcgccgcacagatag(seqidno:7)
a129-f2:ggcgccattagcctgtagccgaaagcccagattaag(seqidno:8)
a129-r2(与d77-r2相同):ttagcagccggatctcagtggtgg(seqidno:6)
为了在位点q75处引入tag密码子,使用快速位点定向诱变试剂盒(tiangen,km101)使用下列的引物对在质粒pet-26b-pd1(wt)上进行位点定向诱变。
q75f:tcgcatgagcccgagcaacagaccgataaactg(seqidno:9)
q75r:gttgctcgggctcatgcgataccaattcag(seqidno:10)
通过将人pd-l1igv结构域编码基因(残基19-134)克隆进质粒pet-26b中生成质粒pet-26b-pd-l1(wt)。简言之,用下列的引物对从质粒puc57-kan-hpd-l1igv结构域(19-134)扩增hpd-l1igv结构域(19-134),所述质粒进行了密码子优化并由genewiz(china,suzhou)合成。pcr产物用ndei-hf和xhoi-hf消化,然后连接到预切的pet-26b载体中。
pd-l1-f:ttccatatgtttaccgttaccgtg(seqidno:11)
pd-l1-r:gaactcgaggtacggggcat(seqidno:12)
使用下列的引物对和快速位点定向诱变试剂盒(tiangen,km101)在质粒pet-26b-pd-l1(wt)上进行位点定向诱变生成质粒pet-26b-pd-l1(h69a)。
h69a-f:cattcagtttgtggccggcgaagaagatctg(seqidno:13)
h69a-r:cagatcttcttcgccggccacaaactgaatg(seqidno:14)
实施例2
人pd-1igv结构域和pd-l1igv结构域的表达和纯化
质粒pet-26b-pd1(wt)、pet-26b-pd-l1(wt)和pet-26b-pd-l1(h69a)单独地转化到大肠杆菌bl21(de3)电感受态细胞中。质粒pet-26b-pd-1(q75tag)、pet-26b-pd-1(d77tag)或pet-26b-pd-1(a129tag)各自与质粒pevol-fsyrs29共转化到大肠杆菌bl21(de3)电感受态细胞中。
对于pd-1(wt)、pd-l1(wt)和pd-l1(h69a)的表达,将转化的细菌在37℃在含有50μg/ml卡那霉素的2xyt培养基中培养,并当od600达到0.8时用1mmiptg诱导。对于fsy掺入到蛋白质中,将转化的细菌在37℃在含有50μg/ml卡那霉素和34μg/ml氯霉素的2xyt培养基中培养,并当od600达到0.8时用1mmiptg、0.2%阿拉伯糖和1mmfsy诱导。
在诱导表达12小时后,通过在4℃以7000rpm离心5min收集细菌。将细胞沉淀物用10ml裂解缓冲液(20mmtris-hcl,ph8.0,200mmnacl,0.5%tritonx-100,溶菌酶1mg/ml,dnase0.1mg/ml和蛋白酶抑制剂)/克细菌湿重重悬。细胞悬浮液通过震摇在4℃裂解30min。随后利用超声波破碎仪(fisherscientific,30%输出,3min,1秒关,1秒开)在冰水浴中进行超声处理。通过将细胞裂解物在4℃以20,000g离心20min回收包涵体,然后将包涵体重悬于包涵体第一洗涤缓冲液(20mmtris-hcl,ph8.0,200mmnacl,10mmedta)中。将重悬溶液在4℃以20,000g再次离心20min,并重悬于包涵体第二洗涤缓冲液(20mmtris-hcl,ph8.0,200mmnacl)中。然后重悬溶液在4℃以20,000g离心20min,并重悬于包涵体溶解缓冲液(20mmtris-hcl,ph8.0,200mmnacl,8m尿素)中,接着在4℃搅拌过夜。溶解的级分通过在4℃以30,000g离心30min进行澄清。将适当体积的ni-nta琼脂糖珠子(qiagen,目录号30210)加入至上清液中并在4℃通过震摇温育30min。将混合物加样至柱中并用10个柱体积的洗涤缓冲液(20mmtris-hcl,ph8.0,200mmnacl,8m尿素,4mm咪唑)洗涤。将蛋白质用洗脱缓冲液(20mmtris-hcl,ph8.0,200mmnacl,8m尿素,300mm咪唑洗脱并用稀释缓冲液(20mmtris-hcl,ph8.0,200mmnacl,8m尿素)稀释到低于0.1mg/ml。将稀释的蛋白质溶液加样到透析袋(md44-1500-01,mwco=1.5kda)中进行透析,并通过尿素浓度的梯度减小(依次为6m,4m,2m,0m)在包含20mmtris-hcl,ph8.0,200mmnacl的缓冲液中将蛋白质重折叠。在每个尿素浓度下,通过在4℃搅拌6小时进行透析30。将在0m尿素缓冲液中的重折叠蛋白质溶液加样到超滤离心管(millipore,目录号ufc900324,mwco=3kda)中进行浓缩并将缓冲液交换到pbs(ph7.3)(用于细胞和动物实验)或20mmtris-hcl,ph8.0,200mmnacl(用于体外蛋白质交联实验)中三次。
通过nanodrop(thermofisher)基于蛋白质的分子量和消光系数测量蛋白质浓度。对于pd-1(wt),分子量为15.57kda,消光系数为14.11。对于pd-l1(wt),分子量为14.41kda,消光系数为17.55。重折叠蛋白质保存在-80℃。
结果
通过遗传密码扩展技术生成掺入fsy的pd-1
潜在的生物反应性uaa可以通过遗传密码的扩展经遗传方式掺入到蛋白质中,其与接近的靶天然氨基酸残基特异性地反应8,10。具体地,潜在的生物反应性uaafsy对细胞无毒,并且在掺入到蛋白质后仍然是惰性的9。fsy能够通过点击化学硫-氟化物交换(sufex)与接近的天然氨基酸残基tyr、his和lys反应(图2a)。本发明人推断将此类潜在的生物反应性uaa在结合界面处引入到蛋白质药物中将使得该蛋白质药物能够与其靶标共价结合。选择了pd-1/pd-l1免疫检查点来测试此想法。pd-1是调节t细胞活性的跨膜受体。pd-l1是pd-1的配体,通常在不同的肿瘤中过表达11。pd-1/pd-l1相互作用抑制t-淋巴细胞增殖、细胞因子的释放和细胞毒性,导致肿瘤特异性t细胞的耗竭和凋亡12,13。因此,阻断pd-1/pd-l1相互作用可以逆转受阻碍的抗肿瘤免疫应答,并且已经生成了几种单克隆抗体来靶向pd-1/pd-l1轴以治疗癌症14,15,16,17。然而,抗体具有固有的限制,包括不佳的组织/肿瘤穿透性和不利的fc-效应子功能(耗竭免疫细胞),并且对于测试的每种癌症,特定百分比的患者对现有的pd-1/pd-l1抗体治疗无应答18,19。因此,寻求具有较小分子量的替代的非抗体治疗剂20,21。例如,pd-1的胞外结构域已经被改造以具有对pd-l1的高亲和力并用作pd-l1的竞争拮抗剂19,22。这些高亲和力pd-1拮抗剂仅在同基因小鼠模型中显示了喜忧参半的结果,并且为了体内半衰期和功效需要额外附加ig结构域。fsy的使用将在pd-1和pd-l1之间形成共价键,形成具有极大亲和力的不可逆拮抗剂,这种极大亲和力是利用通过天然氨基酸诱变进化的任意高亲和力突变体都无法达到的。
基于人pd-1/pd-l1复合物的晶体结构23,本发明人决定单独地在pd-1的位点gln75、asp77或ala129处掺入fsy,分别靶向pd-l1的lys124、lys124和his69(图2b)。通过在大肠杆菌细胞中表达包含在期望位点处引入的琥珀终止密码子(tag)的pd-1基因与用于正交trnapyl/fsyrs对的基因,将fsy掺入到人pd-1的胞外结构域中,所述用于正交trnapyl/fsyrs对的基因被进化为响应于tag密码子而掺入fsy9。使用亲和色谱纯化并重折叠pd-1突变蛋白质(图2c,2d)。对突变蛋白质的质谱分析证实fsy以高保真度在tag-指定的位置处掺入到pd-1中(图2e)。
实施例3
pd-1(fsy)与pd-l1的交联
3.1pd-1(fsy)与pd-l1的体外交联
纯化和重折叠的pd-1(wt)、pd-1(q75fsy)、pd-1(d77fsy)或pd-1(a129fsy)与pd-l1(wt)或pd-l1(h69a)以1:1的摩尔比在pbs缓冲液中在37℃温育6小时。pd-l1的量为4μg。温育后,将5x还原上样缓冲液(cwbio,目录号cw0027)加入到温育液中并在100℃加热10min。然后这些样品通过15%sds-page凝胶进行分离,接着利用考马斯亮蓝染色。然后使用对his6x标签特异的一抗(mab抗-6xhis标签,abcam,目录号18184,1:1000稀释度)和二抗山羊pab抗小鼠igg(abcam,目录号97023,1:5000稀释度)进行蛋白质印迹。通过化学发光(bio-rad,目录号1705062)对蛋白质条带显影。
3.2质谱分析
将pd-1(fsy)和pd-1(fsy)/pd-l1交联蛋白质样品用胰蛋白酶消化。消化的肽利用内置easy-喷雾源和与elite质谱仪(thermofisher)连接的nano-lcultimate3000高效液相色谱系统(thermofisher)进行分析。easy-喷雾pepmapc18柱(50cm;粒径,2μm;孔径,
3.3pd-1(fsy)与细胞表面上表达的内源性pd-l1的交联
h460细胞(5x105个,购自cascellbank,目录号scsp-584)和u-87细胞(5x105个,购自cascellbank,目录号tchu138)分别接种在6孔板中并用rpmi-1640[+10%fbs(gibco,目录号10099-141)和1:100青霉素–链霉素(gibco,目录号15140-122]或dmem(+10%fbs和1:100青霉素-链霉素)培养。6小时后,将pd-1(wt)或pd-1(fsy)加入至培养基中,终体积为800μl。在37℃温育12小时后,细胞用0.25%胰蛋白酶-edta(gibco,目录号25200-056)解离,收集,并通过添加含有1x蛋白酶抑制剂混合物(cellsignalingtechnology,目录号5872)的100μlripa(beyotime,目录号p0013c)裂解,接着进行超声作用。蛋白质浓度使用bca蛋白质测定试剂盒(beyotime,目录号p0010)进行定量。然后在加入5x还原上样缓冲液(cwbio,目录号cw0027)后将样品在100℃加热10min。变性的样品通过在12%sds-page中电泳,接着使用一抗抗人pd-l1(abcam,目录号ab213524;1:1000)和内标准抗β-肌动蛋白(abcam,目录号ab179467;1:3000)和二抗抗兔igg(cellsignalingtechnology,目录号7074s;1:3000)进行蛋白质印迹来进行分析。蛋白质条带通过化学发光(bio-rad,目录号1705062)显影。
为了测试pd-1(fsy)是否可以与内源性小鼠pd-l1交联,从balb/c小鼠分离胸腺和脾脏,然后通过注射器研磨。将获得的细胞重悬于dpbs中,通过40μm滤器(corning,目录号352340)过滤并离心。然后将这些单细胞(1x106个)接种到6孔板中,并在含有10%fbs(gibco,目录号10099-141)和1:100青霉素-链霉素(gibco,目录号15140-122)的rpmi-1640中培养。将pbs、pd-1(wt)或pd-1(fsy)加入到孔中,终体积为800μl。在孵育12小时后,收获细胞用于蛋白质印迹分析,使用类似于上述用于h460和u-87细胞的程序进行。一抗为抗小鼠pd-l1(proteintech,目录号66248-1-ig;1:2000),二抗为抗小鼠igg(cst,目录号7076s;1:3000)。一抗抗-β-肌动蛋白抗体(abcam,目录号ab179467,1:3000)和二抗抗-兔igg(cellsignalingtechnology,目录号7074s,1:3000)用来检测用作内标准的β-肌动蛋白。
3.4pd-1(fsy)与体内肿瘤组织中的内源性pd-l1的交联
所有动物研究均得到机构动物看护和使用委员会的批准。h460或u-87细胞(2x106个)用200μldpbs重悬并分别注射到6-8周龄雄性m-nsg小鼠(nod-prkdc-/--il2rg-/y,shanghaimodelorganisms,china,目录号nm-nsg-001)的侧腹中。10天后,肿瘤大小为约4x4mm2。pd-1(wt)或pd-1(fsy)通过尾静脉注射或注射到肿瘤附近的皮肤部位中。3小时后,处死小鼠,并收集肿瘤。向肿瘤添加含有1x蛋白酶抑制剂混合物(cellsignalingtechnology,目录号5872)的200μlripa(beyotime,目录号p0013c),通过匀浆器(lukacexuyiqi,目录号lukym-i)破碎,并通过超声处理裂解。然后使用与章节3.3中所述相同的程序进行蛋白质印迹。
结果
通过fsy的临近使能反应pd-1与pd-l1共价结合
为了测试掺入fsy的pd-1是否可以与人pd-l1在体外共价结合,本发明人将含fsy的pd-1突变蛋白质与pd-l1蛋白质在缓冲液中在37℃温育,接着利用sds-page和蛋白质印迹进行分析(图3a)。纯化wt人pd-1(下文称为pd-1(wt))的胞外结构域并包括作为对照,其在变性分析条件下不与pd-l1形成共价复合物。对pd-1(q75fsy)和pd-1(a129fsy)检测pd-1/pd-l1的稳定共价复合物,其中pd-1(a129fsy)显示较高的交联效率(45±5%)。为简便起见pd-1(a129fsy)下文称为pd-1(fsy)。为了验证pd-1(fsy)是否共价地靶向人pd-l1的近端his69,本发明人生成了其中his69突变为ala的pd-l1(h69a)突变体,并且发现它不与pd-1(fsy)形成共价复合物(图3b)。另外,本发明人使用高分辨率质谱法分析了pd-1(fsy)与pd-l1的温育产物,清晰地表明pd-1的fsy与pd-l1的his69特异性地反应(图3c)。
本发明人下一步测试了pd-1(fsy)是否可以与在活的癌细胞的表面上生理性地表达的人pd-l1共价结合。pd-1(fsy)与h460细胞(人大细胞肺癌细胞系)或u87细胞(人原发性成胶质细胞瘤细胞系)孵育,这两种细胞均在细胞表面上表达pd-l1。孵育后,将细胞裂解并利用蛋白质印迹分析裂解产物(图3d)。如预期的那样,pd-1(wt)不与pd-l1交联,而pd-1(fsy)与两种癌细胞上的内源性全长pd-l1共价交联,并且交联效率随着所施加的pd-1(fsy)的浓度增加而增加。
本发明人进一步测试了pd-1(fsy)是否可以与肿瘤组织内的人pd-l1在体内共价交联。本发明人使用h460和u87细胞移植形成了患有这些肿瘤的免疫缺陷nodscidgamma(nsg)小鼠。当肿瘤尺寸达到4x4mm2时,通过尾静脉内注射或肿瘤旁注射递送pd-1(wt)或pd-1(fsy)。3小时后,收集肿瘤用于蛋白质印迹分析。如图3e中所示,pd-1(wt)也不与pd-l1产生任何交联,而pd-1(fsy)很显然在这两种肿瘤中和通过两种注射方法均与pd-l1发生共价交联,交联效率随着注射的pd-1(fsy)的剂量增加而增加。
已知人pd-1与小鼠pd-l1交叉结合22。因此本发明人测试了本发明的人pd-1(fsy)是否可以与小鼠细胞表面上的小鼠pd-l1共价交联。将来自小鼠胸腺和脾脏的免疫细胞解离并与pd-1(wt)和pd-1(fsy)孵育。在孵育和使用类似的程序进行蛋白质印迹分析后,没有检测到pd-1(fsy)与小鼠pd-l1的共价交联(图4a)。人和小鼠pd-l1氨基酸序列的同源性比对表明人pd-l1的靶标his69对应于小鼠pd-l1的ala69(图4b)24,其不能通过临近使能反应与fsy反应9。这种高选择性不仅表明可以通过临近使能反应策略实现高靶标特异性,而且提示通过临近使能反应允许的共价蛋白质药物的脱靶效应应该是低的。
总之,这些结果表明pd-1(fsy)与pd-l1通过pd-1的129fsy与pd-l1的his69反应而共价结合。所述共价结合在体外纯化的蛋白质上、在细胞培养中的癌细胞表面上以及在体内在小鼠中的肿瘤组织上有效地发生。
实施例4
混合淋巴细胞反应(mlr)
通过ficoll(gehealthcare,目录号17-5442-02)梯度离心从健康供体的外周血中分离新鲜的pbmc。使用cd14微珠(miltenyi,目录号130-050-201)从pbmc富集单核细胞。纯化的cd14+单核细胞在含有10%fbs(gibco,目录号10099-141)、1:100青霉素-链霉素(gibco,目录号15140-122)、100ng/mlgm-csf(peprotech,目录号300-03)和50ng/mlil-4(peprotech,目录号200-04)的rpmi-1640中培养以诱导dc分化。每3天添加新鲜培养基。在第6天,向培养基加入100ng/mltnf-α(peprotech,目录号300-01a)和3μg/mllps(sigma,目录号l4391)以诱导dc成熟。48小时后,通过流式细胞仪(bdlsrfortessatm)检测在dc上表达的cd11c(bdpharmingen,目录号555392)、pd-l1(bdpharmingen,目录号558065)、cd86(bdhorizon,目录号562999)、cd83(bdpharmingen,目录号561132)和hla-dr(bdpharmingen,目录号560652)。
对于同种异体dc-t细胞刺激,通过cd3微珠(miltenyi,目录号130-097-043)从健康供体的同种异体pbmc富集cd3+t细胞。cd3+t细胞(1x107个)用在1mlpbs中的2μmcfse(ebioscience,目录号65-0850-84)在37℃标记15min;通过添加3ml冷fbs终止标记,然后用冷pbs洗涤两次。
将成熟dc接种在96孔圆底培养板中。将pbs、pd-1或阿特珠单抗(crownbio,目录号e4543-t1901)加入到孔中。2小时后,将cfse标记的t细胞(1x105个)加入到孔中,并与同种异体dc共培养5天。在第3天时,向每个孔中补充30μl新鲜培养基。5天后,收集细胞,用抗人cd3(bdpharmingen,目录号561812)染色,使用流式细胞术检查t细胞增殖并对cd3+t细胞设门控。收获培养上清液,使用elisa试剂盒(invitrogen,目录号88-7316-76)测量inf-γ浓度。流式细胞术数据利用软件flowjo_v10分析。
结果
pd-1(fsy)体外增强t细胞激活
为了检查pd-1(fsy)是否可以通过阻断pd-1/pd-l1相互作用而增强t细胞激活,本发明人进行了体外同种异体混合淋巴细胞反应(mlr)。成熟的树突细胞(dc)来源于从健康供体的人外周血单核细胞(pbmc)分离的单核细胞,通过流式细胞术确认pd-l1表达和dc成熟(图s1)。成熟的dc单独地用pd-1(fsy)、pd-1(wt)或阿特珠单抗(fda-批准的一种用于治疗癌症的抗-pd-l1单克隆抗体)25处理2小时,然后与从健康供体的外周血中纯化并利用用于监测t细胞增殖的羧基荧光素琥珀酰亚胺酯(cfse)染料预先染色的同种异体cd3+t细胞共培养。在共培养5天后,使用流式细胞术分析cfse阳性cd3+t细胞(图5a)。如pbs对照中所示,与dc的共培养增加t细胞增殖,通过cfse强度低于亲代群体的cfse阳性细胞(cfselow)增加指示(图5b,图s2)。与pbs对照相比较,在50nm和250nm的浓度下pd-1(wt)不增加t细胞增殖。相反,pd-1(fsy)在这两个浓度下均引起t细胞增殖的显著剂量依赖的增强。pd-1(fsy)的t细胞激活效应与以相同摩尔浓度应用的阿特珠单抗相当。
作为t细胞激活的另一指标,利用elisa测量培养物中由t细胞释放的细胞因子干扰素γ(inf-γ)(图5c)。与pbs对照相比,在50nm的浓度下pd-1(wt)不增加inf-γ产生,在250nm有中度的增加(0.77倍)。相反,在50nm的浓度下pd-1(fsy)引起inf-γ产生的显著增加(3.4倍),并且在250nm进一步增加至4.1倍。pd-1(fsy)对inf-γ产生的效应与阿特珠单抗也相当。这些结果表明pd-1(fsy)可以通过阻断pd-1与pd-l1的相互作用显著增强由同种异体dc诱导的t细胞激活,而pd-1(wt)则不能。
实施例5
pbmc-肿瘤异种移植物模型研究
在第-10天,新鲜的pbmc用包被在12孔平板中的3μg/ml抗-cd3抗体(bdpharmingen,目录号555329)和1μg/ml抗-cd28抗体(bdpharmingen,目录号555725)激活并在包含10%fbs、1:100青霉素-链霉素(gibco,目录号15140-122)、100iu/mlil-2(peprotech,目录号af-200-02-500)和2mm谷氨酰胺(gibco,目录号25030-081)的rpmi-1640中培养。在第-7天,将5x105个h460细胞接种到6孔板中。6小时后,h460细胞用丝裂霉素c(10μg/ml)处理2小时,然后用pbs洗涤两次。然后向这些h460细胞中加入激活的pbmc(5x106个)进行共培养(pbmc:h460=10:1)。每2天收集pbmc,与用丝裂霉素c(10μg/ml)处理的h460细胞再培养。共培养3次后,收获pbmc并与新鲜的h460细胞以1:4的比率(h460=2x106个,pbmc=5x105个)混合。将细胞混合物注射到6-8周龄雄性m-nsg小鼠(nod-prkdc-/--il2rg-/y,shanghaimodelorganisms,china,目录号nm-nsg-001)的侧腹中。3小时后和每6天(在第6、12、18、24、30、36、42天)将pbs、22μgpd-1(wt)、22μgpd-1(fsy)或200μg阿特珠单抗通过尾静脉注射施用。3小时后和每3天(在第3、6、9、12、15、18、21、24、27、30、33、36、39、42天)将11μgpd-1(fsy)或100μg阿特珠单抗通过尾静脉注射施用。利用游标卡尺在两个维度上测量肿瘤生长,使用下面的公式计算肿瘤体积:肿瘤体积=长度x宽度2/2。在第44天,肿瘤长度接近于15mm,将小鼠处死用于分析。
使用prism6.0(graphpadsoftware)通过单因素anova,接着通过tukey多重比较检验比较对cfse增殖测定、cd69表达、肿瘤组织中的t细胞浸润(流式细胞术数据)、elisa数据和肿瘤重量数据的统计学分析。使用prism6.0(graphpadsoftware)通过双因素anova,接着通过tukey多重比较检验比较肿瘤生长数据。
结果
pd-1(fsy)在用人pbmc重建的肿瘤异种移植物模型中的抗肿瘤效应
鉴于pd-1(fsy)在体外不可逆地结合pd-l1和促进t细胞激活的能力,本发明人试图评价其体内抗肿瘤活性。由于pd-1(fsy)对人pd-l1有共价特异性,但对鼠pd-l1没有共价特异性,本发明人没有使用同基因有免疫能力的小鼠模型,而求助于用人免疫细胞重建的免疫缺陷小鼠模型,它是用于癌症免疫疗法的更有预测性的临床前模型26。相反,以前报道的人pd-1的两个高亲和力突变体均与小鼠pd-l1交叉结合,并且仅对在具有完全鼠源免疫系统的小鼠模型中抑制小鼠pd-l1来评价它们的抗肿瘤效应19,22。
本发明人利用人肿瘤h460细胞和人pbmc的混合物共同移植免疫缺陷nsg小鼠。人pbmc用抗-cd3和cd28抗体激活3天,然后与h460细胞共培养7天(图6a)。这些pbmc与h460(比率1:4)的混合物经皮下注射到免疫缺陷nsg小鼠的侧腹中。随后每3天(低剂量)或每6天(高剂量)通过尾静脉内注射施用pd-1蛋白质或阿特珠单抗。监测肿瘤生长持续44天。如期望的那样,媒介物处理的小鼠(pbs对照)的肿瘤生长迅速。用22μgpd-1(wt)治疗在一定程度上延缓了肿瘤生长,而用22μgpd-1(fsy)治疗显著地抑制了肿瘤生长(p<0.0001)(图6b,6c)持续整个44天。将pd-1(fsy)的剂量减小至11μg保持了利用22μg的相似的肿瘤抑制效应。事实上,pd-1(fsy)达到了与以相同摩尔量施用的阿特珠单抗相同的抗肿瘤效应,而所需的pd-1(fsy)的质量为阿特珠单抗的十分之一。在第44天时切除的肿瘤显示在图6d中,它们的重量在图6e中进行了比较。pd-1(fsy)-治疗的肿瘤在重量上显著小于用pd-1(wt)治疗的那些(p<0.001),并且与用阿特珠单抗治疗的那些相当。
本领域技术人员要进一步理解在不偏离本公开的精神或中心属性的前提下,本公开可以以其他具体的形式体现。由于前面对本公开的描述仅公开了其示例性的实施方案,因此要理解的是其他变化被认为在本公开的范围内。因此,本公开不限于已经在本文中详细描述的特定实施方案。而是,应当参照附带的权利要求作为本公开的范围和内容的指示。
参考文献
(1)singh,j.;petter,r.c.;baillie,t.a.;whitty,a.theresurgenceofcovalentdrugs.nat.rev.drugdiscov.2011,10(4),307–317.
(2)bauer,r.a.covalentinhibitorsindrugdiscovery:fromaccidentaldiscoveriestoavoidedliabilitiesanddesignedtherapies.drugdiscoverytoday2015,20(9),1061–1073.
(3)lonsdale,r.;ward,r.a.structure-baseddesignoftargetedcovalentinhibitors.chem.soc.rev.2018,47(11),3816–3830.
(4)johnson,d.s.;weerapana,e.;cravatt,b.f.strategiesfordiscoveringandderiskingcovalent,irreversibleenzymeinhibitors.futuremedchem2010,2(6),949–964.
(5)robertson,j.g.mechanisticbasisofenzyme-targeteddrugs.biochemistry2005,44(15),5561–5571.
(6)wang,l.;brock,a.;herberich,b.;schultz,p.g.expandingthegeneticcodeofescherichiacoli.science2001,292(5516),498–500.
(7)wang,l.;schultz,p.g.expandingthegeneticcode.angew.chem.int.ed.2004,44(1),34–66.
(8)xiang,z.;ren,h.;hu,y.s.;coin,i.;wei,j.;cang,h.;wang,l.addinganunnaturalcovalentbondtoproteinsthroughproximity-enhancedbioreactivity.nat.methods2013,10(9),885–888.
(9)wang,n.;yang,b.;fu,c.;zhu,h.;zheng,f.;kobayashi,t.;liu,j.;li,s.;ma,c.;wang,p.g.;wang,q.;wang,l.geneticallyencodingfluorosulfate-l-tyrosinetoreactwithlysine,histidine,andtyrosineviasufexinproteinsinvivo.j.am.chem.soc.2018,140(15),4995–4999.
(10)wang,l.geneticallyencodingnewbioreactivity.n.biotechnol.2017,38(pta),16–25.
(11)okazaki,t.;honjo,t.pd-1andpd-1ligands:fromdiscoverytoclinicalapplication.int.immunol.2007,19(7),813–824.
(12)francisco,l.m.;sage,p.t.;sharpe,a.h.thepd-1pathwayintoleranceandautoimmunity.immunol.rev.2010,236,219–242.
(13)sharpe,a.h.;pauken,k.e.thediversefunctionsofthepd1inhibitorypathway.nat.rev.immunol.2018,18(3),153–167.
(14)sharma,p.;allison,j.p.thefutureofimmunecheckpointtherapy.science2015,348(6230),56–61.
(15)rosenberg,j.e.;hoffman-censits,j.;powles,t.;vanderheijden,m.s.;balar,a.v.;necchi,a.;dawson,n.;o'donnell,p.h.;balmanoukian,a.;loriot,y.;srinivas,s.;retz,m.m.;grivas,p.;joseph,r.w.;galsky,m.d.;fleming,m.t.;petrylak,d.p.;perez-gracia,j.l.;burris,h.a.;castellano,d.;canil,c.;bellmunt,j.;bajorin,d.;nickles,d.;bourgon,r.;frampton,g.m.;cui,n.;mariathasan,s.;abidoye,o.;fine,g.d.;dreicer,r.atezolizumabinpatientswithlocallyadvancedandmetastaticurothelialcarcinomawhohaveprogressedfollowingtreatmentwithplatinum-basedchemotherapy:asingle-arm,multicentre,phase2trial.lancet2016,387(10031),1909–1920.
(16)syed,y.y.durvalumab:firstglobalapproval.drugs2017,77(12),1369–1376.
(17)kaufman,h.l.;russell,j.;hamid,o.;bhatia,s.;terheyden,p.;d'angelo,s.p.;shih,k.c.;lebbé,c.;linette,g.p.;milella,m.;brownell,i.;lewis,k.d.;lorch,j.h.;chin,k.;mahnke,l.;heydebreck,von,a.;cuillerot,j.-m.;nghiem,p.avelumabinpatientswithchemotherapy-refractorymetastaticmerkelcellcarcinoma:amulticentre,single-group,open-label,phase2trial.lancetoncol.2016,17(10),1374–1385.
(18)lee,c.m.;tannock,i.f.thedistributionofthetherapeuticmonoclonalantibodiescetuximabandtrastuzumabwithinsolidtumors.bmccancer2010,10(1),–11.
(19)maute,r.l.;gordon,s.r.;mayer,a.t.;mccracken,m.n.;natarajan,a.;ring,n.g.;kimura,r.;tsai,j.m.;manglik,a.;kruse,a.c.;gambhir,s.s.;weissman,i.l.;ring,a.m.engineeringhigh-affinitypd-1variantsforoptimizedimmunotherapyandimmuno-petimaging.proc.natl.acad.sci.u.s.a.2015,112(47),e6506–e6514.
(20)li,c.;zhang,n.;zhou,j.;ding,c.;jin,y.;cui,x.;pu,k.;zhu,y.peptideblockingofpd-1/pd-l1interactionforcancerimmunotherapy.cancerimmunologyresearch2018,6(2),178–188.
(21)zarganes-tzitzikas,t.;konstantinidou,m.;gao,y.;krzemien,d.;zak,k.;dubin,g.;holak,t.a.;
(22)lázár-molnár,e.;scandiuzzi,l.;basu,i.;quinn,t.;sylvestre,e.;palmieri,e.;ramagopal,u.a.;nathenson,s.g.;guha,c.;almo,s.c.structure-guideddevelopmentofahigh-affinityhumanprogrammedcelldeath-1:implicationsfortumorimmunotherapy.ebiomedicine2017,17,30–44.
(23)zak,k.m.;kitel,r.;przetocka,s.;golik,p.;guzik,k.;musielak,b.;
(24)tan,s.;chen,d.;liu,k.;he,m.;song,h.;shi,y.;liu,j.;zhang,c.w.h.;qi,j.;yan,j.;gao,s.;gao,g.f.crystalclear:visualizingtheinterventionmechanismofthepd-1/pd-l1interactionbytwocancertherapeuticmonoclonalantibodies.proteincell2016,7(12),866–877.
(25)akinleye,a.;rasool,z.immunecheckpointinhibitorsofpd-l1ascancertherapeutics.jhematoloncol2019,12(1),–13.
(26)larochere,de,p.;guil-luna,s.;decaudin,d.;azar,g.;sidhu,s.s.;piaggio,e.humanizedmiceforthestudyofimmuno-oncology.trendsinimmunology2018,39(9),748–763.
(27)xuan,w.;shao,s.;schultz,p.g.proteincrosslinkingbygeneticallyencodednoncanonicalaminoacidswithreactivearylcarbamatesidechains.angew.chem.int.ed.engl.2017,56(18),5096–5100.
(28)zak,k.m.;kitel,r.;przetocka,s.;golik,p.;guzik,k.;musielak,b.;
(29)wang,n.;yang,b.;fu,c.;zhu,h.;zheng,f.;kobayashi,t.;liu,j.;li,s.;ma,c.;wang,p.g.;wang,q.;wang,l.geneticallyencodingfluorosulfate-l-tyrosinetoreactwithlysine,histidine,andtyrosineviasufexinproteinsinvivo.j.am.chem.soc.2018,140(15),4995–4999.
(30)lee,j.y.;lee,h.t.;shin,w.;chae,j.;choi,j.;kim,s.h.;lim,h.;wonheo,t.;park,k.y.;lee,y.j.;ryu,s.e.;son,j.y.;lee,j.u.;heo,y.-s.structuralbasisofcheckpointblockadebymonoclonalantibodiesincancerimmunotherapy.nat.commun.2016,7,13354。
序列表
<110>中国科学院理化技术研究所杭州研究院
<120>通过临近使能反应疗法开发的共价蛋白质药物
<130>idc216008
<160>21
<170>patentinversion3.5
<210>1
<211>35
<212>dna
<213>人工序列
<220>
<223>引物
<400>1
gttagactcatatgtggaatccgccgacctttagc35
<210>2
<211>32
<212>dna
<213>人工序列
<220>
<223>引物
<400>2
actgctcgagcggactaggactcggatgtgcg32
<210>3
<211>30
<212>dna
<213>人工序列
<220>
<223>引物
<400>3
cctcatatgtggaatccgccgacctttagc30
<210>4
<211>23
<212>dna
<213>人工序列
<220>
<223>引物
<400>4
ggtctggttgctcgggctcatgc23
<210>5
<211>36
<212>dna
<213>人工序列
<220>
<223>引物
<400>5
cccgagcaaccagacctagaaactggccgcctttcc36
<210>6
<211>24
<212>dna
<213>人工序列
<220>
<223>引物
<400>6
ttagcagccggatctcagtggtgg24
<210>7
<211>27
<212>dna
<213>人工序列
<220>
<223>引物
<400>7
tgcaggctaatggcgccgcacagatag27
<210>8
<211>36
<212>dna
<213>人工序列
<220>
<223>引物
<400>8
ggcgccattagcctgtagccgaaagcccagattaag36
<210>9
<211>33
<212>dna
<213>人工序列
<220>
<223>引物
<400>9
tcgcatgagcccgagcaacagaccgataaactg33
<210>10
<211>30
<212>dna
<213>人工序列
<220>
<223>引物
<400>10
gttgctcgggctcatgcgataccaattcag30
<210>11
<211>24
<212>dna
<213>人工序列
<220>
<223>引物
<400>11
ttccatatgtttaccgttaccgtg24
<210>12
<211>20
<212>dna
<213>人工序列
<220>
<223>引物
<400>12
gaactcgaggtacggggcat20
<210>13
<211>31
<212>dna
<213>人工序列
<220>
<223>引物
<400>13
cattcagtttgtggccggcgaagaagatctg31
<210>14
<211>31
<212>dna
<213>人工序列
<220>
<223>引物
<400>14
cagatcttcttcgccggccacaaactgaatg31
<210>15
<211>138
<212>prt
<213>人工序列
<220>
<223>pd-1(wt)的氨基酸序列
<400>15
mettrpasnproprothrpheserproalaleuleuvalvalthrglu
151015
glyaspasnalathrphethrcysserpheserasnthrsergluser
202530
phevalleuasntrptyrargmetserproserasnglnthrasplys
354045
leualaalapheprogluaspargserglnproglyglnaspserarg
505560
pheargvalthrglnleuproasnglyargaspphehismetserval
65707580
valargalaargargasnaspserglythrtyrleucysglyalaile
859095
serleualaprolysalaglnilelysgluserleuargalagluleu
100105110
argvalthrgluargargalagluvalprothralahisproserpro
115120125
serproleugluhishishishishishis
130135
<210>16
<211>417
<212>dna
<213>人工序列
<220>
<223>包含琥珀终止密码子(75tag)的pd-1基因
<400>16
atgtggaatccgccgacctttagcccggccctgctggttgttaccgagggtgataatgcc60
accttcacctgcagcttcagcaacaccagcgagagcttcgtgctgaattggtatcgcatg120
agcccgagcaactagaccgataaactggccgcctttccggaagatcgtagccagccgggc180
caggatagccgctttcgtgttacccagctgccgaacggtcgcgattttcacatgagcgtg240
gtgcgcgcccgccgtaatgatagcggtacctatctgtgcggcgccattagcctggcaccg300
aaagcccagattaaggaaagcctgcgcgcagaactgcgcgtgaccgaacgtcgcgcagaa360
gtgccgaccgcacatccgagtcctagtccgctcgagcaccaccaccaccaccactga417
<210>17
<211>417
<212>dna
<213>人工序列
<220>
<223>包含琥珀终止密码子(77tag)的pd-1基因
<400>17
atgtggaatccgccgacctttagcccggccctgctggttgttaccgagggtgataatgcc60
accttcacctgcagcttcagcaacaccagcgagagcttcgtgctgaattggtatcgcatg120
agcccgagcaaccagacctagaaactggccgcctttccggaagatcgtagccagccgggc180
caggatagccgctttcgtgttacccagctgccgaacggtcgcgattttcacatgagcgtg240
gtgcgcgcccgccgtaatgatagcggtacctatctgtgcggcgccattagcctggcaccg300
aaagcccagattaaggaaagcctgcgcgcagaactgcgcgtgaccgaacgtcgcgcagaa360
gtgccgaccgcacatccgagtcctagtccgctcgagcaccaccaccaccaccactga417
<210>18
<211>417
<212>dna
<213>人工序列
<220>
<223>包含琥珀终止密码子(129tag)的pd-1基因
<400>18
atgtggaatccgccgacctttagcccggccctgctggttgttaccgagggtgataatgcc60
accttcacctgcagcttcagcaacaccagcgagagcttcgtgctgaattggtatcgcatg120
agcccgagcaaccagaccgataaactggccgcctttccggaagatcgtagccagccgggc180
caggatagccgctttcgtgttacccagctgccgaacggtcgcgattttcacatgagcgtg240
gtgcgcgcccgccgtaatgatagcggtacctatctgtgcggcgccattagcctgtagccg300
aaagcccagattaaggaaagcctgcgcgcagaactgcgcgtgaccgaacgtcgcgcagaa360
gtgccgaccgcacatccgagtcctagtccgctcgagcaccaccaccaccaccactga417
<210>19
<211>72
<212>dna
<213>人工序列
<220>
<223>trnapyl
<400>19
ggaaacctgatcatgtagatcgaacggactctaaatccgttcagccgggttagattcccg60
gggtttccgcca72
<210>20
<211>454
<212>prt
<213>人工序列
<220>
<223>fsyrs的氨基酸序列
<400>20
metasplyslysproleuasnthrleuileseralathrglyleutrp
151015
metserargthrglythrilehislysilelyshishisgluvalser
202530
argserlysiletyrileglumetalacysglyasphisleuvalval
354045
asnasnserargserserargthralaargalaleuarghishislys
505560
tyrarglysthrcyslysargcysargvalseraspgluaspleuasn
65707580
lyspheleuthrlysalaasngluaspglnthrservallysvallys
859095
valvalseralaprothrargthrlyslysalametprolysserval
100105110
alaargalaprolysproleugluasnthrglualaalaglnalagln
115120125
proserglyserlyspheserproalaileprovalserthrglnglu
130135140
servalservalproalaservalserthrserileserserileser
145150155160
thrglyalathralaseralaleuvallysglyasnthrasnproile
165170175
thrsermetseralaprovalglnalaseralaproalaleuthrlys
180185190
serglnthraspargleugluvalleuleuasnprolysaspgluile
195200205
serleuasnserglylysprophearggluleuglusergluleuleu
210215220
serargarglyslysaspleuglnglniletyralaglugluargglu
225230235240
asntyrleuglylysleugluarggluilethrargphephevalasp
245250255
argglypheleugluilelysserproileleuileproleuglutyr
260265270
ilegluargmetglyileaspasnaspthrgluleuserlysglnile
275280285
pheargvalasplysasnphecysleuargprometleuileproasn
290295300
leutyrasntyrleuarglysleuaspargalaleuproaspproile
305310315320
lysilephegluileglyprocystyrarglysgluseraspglylys
325330335
gluhisleuglugluphethrmetleuthrpheileglnmetglyser
340345350
glycysthrarggluasnleugluserileilethrasppheleuasn
355360365
hisleuglyileaspphelysilevalglyaspsercysmetvalleu
370375380
glyaspthrleuaspvalmethisglyaspleugluleuserserala
385390395400
valvalglyproileproleuaspargglutrpglyileasplyspro
405410415
lysileglyalaglypheglyleugluargleuleulysvallyshis
420425430
aspphelysasnilelysargalaalaargserglusertyrtyrasn
435440445
glyileserthrasnleu
450
<210>21
<211>1365
<212>dna
<213>人工序列
<220>
<223>fsyrs的核苷酸序列
<400>21
atggataaaaagcctttgaacactctgatttctgcgaccggtctgtggatgtcccgcacc60
ggcaccatccacaaaatcaaacaccatgaagttagccgttccaaaatctacattgaaatg120
gcttgcggcgatcacctggttgtcaacaactcccgttcttctcgtaccgctcgcgcactg180
cgccaccacaaatatcgcaaaacctgcaaacgttgccgtgttagcgatgaggacctgaac240
aaattcctgaccaaagctaacgaggatcagacctccgtaaaagtgaaggtagtaagcgct300
ccgacccgtactaaaaaggctatgccaaaaagcgtggcccgtgccccgaaacctctggaa360
aacaccgaggcggctcaggctcaaccatccggttctaaattttctccggcgatcccagtg420
tccacccaagaatctgtttccgtaccagcaagcgtgtctaccagcattagcagcatttct480
accggtgctaccgcttctgcgctggtaaaaggtaacactaacccgattactagcatgtct540
gcaccggtacaggcaagcgccccagctctgactaaatcccagacggaccgtctggaggtg600
ctgctgaacccaaaggatgaaatctctctgaacagcggcaagcctttccgtgagctggaa660
agcgagctgctgtctcgtcgtaaaaaggatctgcaacagatctacgctgaggaacgcgag720
aactatctgggtaagctggagcgcgaaattactcgcttcttcgtggatcgcggtttcctg780
gagatcaaatctccgattctgattccgctggaatacattgaacgtatgggcatcgataat840
gataccgaactgtctaaacagatcttccgtgtggataaaaacttctgtctgcgtccgatg900
ctgattccgaacttgtacaactatttacgtaaactggaccgtgccctgccggacccgatc960
aaaatattcgagatcggtccttgctaccgtaaagagtccgacggtaaagagcacctggaa1020
gaattcaccatgctgacattcattcagatgggtagcggttgcacgcgtgaaaacctggaa1080
tccattatcaccgacttcctgaatcacctgggtatcgatttcaaaattgttggtgacagc1140
tgtatggtgttaggcgatacgctggatgttatgcacggcgatctggagctgtcttccgca1200
gttgtgggcccaatcccgctggatcgtgagtggggtatcgacaaacctaaaatcggtgcg1260
ggttttggtctggagcgtctgctgaaagtaaaacacgacttcaagaacatcaaacgtgct1320
gcacgttccgagtcctattacaatggtatttctactaacctgtaa1365
<210>22
<211>417
<212>dna
<213>人工序列
<220>
<223>pd-1(wt)的核苷酸序列
<400>22
atgtggaatccgccgacctttagcccggccctgctggttgttaccgagggtgataatgcc60
accttcacctgcagcttcagcaacaccagcgagagcttcgtgctgaattggtatcgcatg120
agcccgagcaaccagaccgataaactggccgcctttccggaagatcgtagccagccgggc180
caggatagccgctttcgtgttacccagctgccgaacggtcgcgattttcacatgagcgtg240
gtgcgcgcccgccgtaatgatagcggtacctatctgtgcggcgccattagcctggcaccg300
aaagcccagattaaggaaagcctgcgcgcagaactgcgcgtgaccgaacgtcgcgcagaa360
gtgccgaccgcacatccgagtcctagtccgctcgagcaccaccaccaccaccactga417
<210>23
<211>138
<212>prt
<213>人工序列
<220>
<221>o-氟硫酸-l-酪氨酸(fsy)
<222>(45)
<223>pd-1(q75fsy)的氨基酸序列
<400>23
mettrpasnproprothrpheserproalaleuleuvalvalthrglu
151015
glyaspasnalathrphethrcysserpheserasnthrsergluser
202530
phevalleuasntrptyrargmetserproserasnxaathrasplys
354045
leualaalapheprogluaspargserglnproglyglnaspserarg
505560
pheargvalthrglnleuproasnglyargaspphehismetserval
65707580
valargalaargargasnaspserglythrtyrleucysglyalaile
859095
serleualaprolysalaglnilelysgluserleuargalagluleu
100105110
argvalthrgluargargalagluvalprothralahisproserpro
115120125
serproleugluhishishishishishis
130135
<210>24
<211>138
<212>prt
<213>人工序列
<220>
<221>o-氟硫酸-l-酪氨酸(fsy)
<222>(47)
<223>pd-1(d77fsy)的氨基酸序列
<400>24
mettrpasnproprothrpheserproalaleuleuvalvalthrglu
151015
glyaspasnalathrphethrcysserpheserasnthrsergluser
202530
phevalleuasntrptyrargmetserproserasnglnthrxaalys
354045
leualaalapheprogluaspargserglnproglyglnaspserarg
505560
pheargvalthrglnleuproasnglyargaspphehismetserval
65707580
valargalaargargasnaspserglythrtyrleucysglyalaile
859095
serleualaprolysalaglnilelysgluserleuargalagluleu
100105110
argvalthrgluargargalagluvalprothralahisproserpro
115120125
serproleugluhishishishishishis
130135
<210>25
<211>138
<212>prt
<213>人工序列
<220>
<221>o-氟硫酸-l-酪氨酸(fsy)
<222>(99)
<223>pd-1(a129fsy)的氨基酸序列
<400>25
mettrpasnproprothrpheserproalaleuleuvalvalthrglu
151015
glyaspasnalathrphethrcysserpheserasnthrsergluser
202530
phevalleuasntrptyrargmetserproserasnglnthrasplys
354045
leualaalapheprogluaspargserglnproglyglnaspserarg
505560
pheargvalthrglnleuproasnglyargaspphehismetserval
65707580
valargalaargargasnaspserglythrtyrleucysglyalaile
859095
serleuxaaprolysalaglnilelysgluserleuargalagluleu
100105110
argvalthrgluargargalagluvalprothralahisproserpro
115120125
serproleugluhishishishishishis
130135
2
- 还没有人留言评论。精彩留言会获得点赞!