重组人源嵌合GcFc基因片段及融合蛋白的制作方法
本发明涉及基因重组技术,特别涉及一种重组人源嵌合GcFc基因片段及融合蛋白。
背景技术:
近年来,在生物制药领域,人源化抗体和Fc融合蛋白两类技术发展迅速。根据重链恒定区的不同,人源抗体可以分为IgG、IgA、IgM、IgD和IgE 5类,其中IgG类抗体半衰期长达21天,人源化抗体药物多为IgG形式。Fc融合蛋白,又称抗体融合蛋白、Ig融合蛋白,是利用基因工程技术将具有活性功能的蛋白分子与抗体的Fc段通过铰链区的半胱氨酸残基连接,形成类似于IgG分子的同源二聚体蛋白质,但无CH1区域和轻链。由于结构上的同源,使Fc融合蛋白不仅具备功能蛋白分子的全部生物学活性,还表现出与同种型的人IgG相当的药物代谢动力学,具有长效的体内应用半衰期。
IgG的Fc段在消灭病原体或肿瘤细胞的免疫防御中起重要作用。IgG Fc的效应子功能包括两种主要机制:1、与细胞表面的FcγR结合,发挥调理作用或抗体依赖性细胞介导的细胞毒作用(ADCC);2、与补体成分C1q结合,引发补体依赖的细胞毒作用(CDC),从而裂解细胞。根据发现的先后顺序和在血清中浓度的不同,IgG又可分为IgGl、IgG2、IgG3和IgG4四种亚型。在四种人IgG同种型中,IgG1和IgG3能有效地结合FcγR。IgG4与FcγR的结合亲和力比IgG1和IgG3的低一个数量级,而IgG2与FcγR的结合低得难以测定。人IgG1和IgG3还能有效地结合C1q,激活补体级联反应,发挥CDC效应。人IgG2对补体的固定很弱,而IgG4表现出极端缺乏激活补体级联反应的能力。
在临床上,除了有目的的利用抗体的ADCC或CDC效应治疗疾病外,在有些时候,需要避免人源化抗体或Fc融合蛋白产生ADCC或CDC效应,亦即当人源化抗体或Fc融合蛋白结合于靶细胞表面时,抗体或融合蛋白的Fc区域要求必须不会介导不良效应子功能而裂解或除去这些细胞,因此,这类人源化抗体或Fc融合蛋白的Fc区域必须是非裂解性的,即在结合FcγR和C1q而触发效应子功能方面,Fc必须是无活性的。显然,没有一种天然的IgG同种型适合产生以上要求的人源化抗体或Fc融合蛋白。
制备非裂解性的人源化抗体或Fc融合蛋白,目前的策略是通过使天然Fc区域中的一些氨基酸突变而得到Fc变体,来消除其ADCC和CDC效应,但与此同时,氨基酸序列的局部改变不可避免的增加了Fc区域的免疫原性,不利于此类制剂的反复和长期应用。为了制备出稳定性好、半衰期长、非裂解性且免疫原性低的人源化抗体或Fc融合蛋白,有必要开发出新型的Fc区域结构,本发明由此而来。
技术实现要素:
本发明的目的是提供一种重组人源嵌合GcFc基因片段及融合蛋白,至少能够解决上述问题之一。
为实现上述目的,根据本发明的一个方面,提供了一种重组人源嵌合GcFc基因片段,包括三个依次连接的基因片段,三个基因片段的编码序列依次如SeqNo.1~3所示。
在一些实施方式中,重组人源嵌合GcFc基因片段基因编码序列如SeqNo.4所示。
相应地,本发明还提供了一种融合蛋白,包含免疫球蛋白的Fc段,Fc段包含GcFc多肽,GcFc多肽由上述的重组人源嵌合GcFc基因片段编码,GcFc多肽的氨基酸序列如SeqNo.5所示。
本发明的重组人源嵌合GcFc基因片段的设计思路来源如下。
通过比较人和鼠的IgG同种型的氨基酸序列,发现CH2区域氨基末端附近的Fc部分在IgG Fc与FcγR的结合中起作用,已用基因工程抗体证明234位到237位基序的重要性。在四种人IgG同种型中,IgG1和IgG3与FcγR的结合能力最强,且具有相同的序列Leu234-Leu-Gly-Gly237。在以低亲和力与FcγR结合的IgG4中,其序列含有单个氨基酸取代,Phe取代234位的Leu。在不结合FcγR的IgG2中,两个取代和一个缺失形成,Val234-Ala-Gly237。为了进一步减少IgG4Fc与FcγR的结合和ADCC活性,可用Ala替代IgG4中的Leu235。用IgG2序列中的Pro233-Val-Ala235替代IgG1序列中的Glu233-Leu-Leu235,可使IgG1变体在小鼠中失去由FcγR介导清除靶细胞的能力。
IgG的C1q结合位点在CH2结构域上,点突变实验证明IgG的Glu318-X-Lys320-X-Lys322对结合起重要作用。对Fc与C1q结合非常重要的另一部分序列位于人IgG的CH2区域羧基端附近。在四种人IgG同种型中,这部分中仅有一个位点显示取代:用IgG1、IgG2和IgG3中的Pro331替代IgG4中的Ser331,可以部分增强IgG4的补体固定活性。而用Ser替代IgG1的Pro331则可使IgG1失去与C1q的结合亲和力。
人IgG2不结合FcγR,但显示出极弱的补体结合活性。具有Pro331Ser突变的Fcγ2变体比天然Fcγ2的补体活性更低,而且依旧是FcγR非结合子。IgG4Fc在激活补体级联中有缺陷,且它与FcγR的结合亲和力比最高活性的同种型(IgG1)的低一个数量级。与天然Fcγ4相比,具有Leu235Ala突变的Fcγ4变体表现出最小的效应子功能。具有Leu234Val、Leu235Ala和Pro331Ser突变的Fcγ1也表现出比天然Fcγ1低的效应子功能。这些Fc变体比各种天然人IgG Fc更适于制备非裂解性人源化抗体或Fc融合蛋白。在非裂解Fc的制备中也可引入其它替换,而不影响循环半衰期或导致不良的构型变化。
IgG4与FcγR的结合亲和力比IgG1和IgG3的低一个数量级,且表现出极端缺乏激活补体级联反应的能力,因此有研究者选择应用IgG4的Fc制备非裂解性人源化抗体或Fc融合蛋白,但IgG4抗体容易发生半分子交换,或称半抗体交换反应,IgG4分子重链间的二硫键并不稳定,使得重链可以分开并随机重组,由此形成带有两个不同抗原结合位点的不对称抗体,一种天然双特异性抗体。IgG4抗体发生半分子交换需要满足三个条件:一是抗体铰链区特殊模体(motif);二是CH3结构域关键氨基酸残基;三要有还原条件的存在。IgG各亚型的铰链区都有一个CXXC蛋白模体(C:半胱氨酸;X:任意氨基酸),2个半胱氨酸形成重链间的二硫键。如IgGl的核心铰链区模体为226-CPPC-229(半胱氨酸-脯氨酸-脯氨酸-半胱氧酸),而IgG4的是226-CPSC-229(半胱氨酸-脯氨酸-丝氨酸-半胱氨酸),228位脯氨酸变为丝氧酸的这一改变使IgG4核心铰链区更容易被还原,链间二硫键极易变成链内二硫键,重链间共价键连接遭到破坏,最终导致IgG4重链解开,形成“半分子”。通过引入氨基酸突变,研宄人员发现抗体分子的CH3结构域而不是核心铰链区对半分子交换具有决定性作用。实验发现,将IgGl和IgG4的整个CH3结构域相互交换之后,IgG4失去了半分子交换能力,而IgGl则可以进行半分子交换,如果只交换铰链区,IgGl获得不了这种半分子交换能力。IgG4CH3结构域的第409位精氨酸残基(R)使2条重链之间CH3-CH3作用力减弱,有利于半分子交换进行。在有还原剂(如谷胱甘肽)存在,37℃的生理条件下IgG4半分子交换能有效的进行。由于半分子交换现象的存在,IgG4的Fc不是十分适合应用于人源化抗体或Fc融合蛋白的制备。此外,有文献报道,肥大细胞表面存在与IgE Fc受体相似的IgG4Fc受体,IgG4与其结合后可介导肥大细胞释放组胺等介质,可能在IgE介导的迟发性介质释放中发挥作用。
同IgG不同,IgM有两种形式,分泌型和膜结合型,分泌型IgM为五聚体,由J链和二硫键连接五个IgM单体形成,分子结构呈环形,是Ig中分子最大者。μ链Cys414(Cμ3)和Cys575(C端的尾部)对于IgM的多聚化极为重要。在J链存在下,通过两个邻近单体IgMμ链Cys之间以及J链与邻μ链Cys575之间形成二硫键组成五聚体。分泌型IgM的μ链在556位氨基酸后有20个氨基酸组成的尾片,C末端倒数第二个为Cys。膜表面型IgM的μ链在556位氨基酸后有41个氨基酸,其中富含带负电氨基酸胞膜外区12个氨基酸,26个疏水性氨基酸组成的穿膜区以及胞浆区3个氨基酸。μ链含有5个同源区,其CH3和CH4相当于IgG的CH2和CH3,无铰链区。IgG1、IgG2、IgG3的补体结合位点在CH2区内,而IgM补体结合位点在CH3区内,IgM无FcR结合能力。
本发明的有益效果如下。
本发明的重组人源嵌合GcFc基因片段,将SeqNo.1~3三段基因片段依次连接,如SeqNo.4所示。三个基因片段分别是IgG2的铰链区及其CH2结构域N端的部分基因、IgG4的CH2结构域C端的部分基因、IgG2的CH3结构域基因。
由于IgG2的Fc段基因和IgG4的Fc段基因具有高度同源性,使重组的嵌合GcFc片段中的分别来源于IgG2的Fc段基因和IgG4的Fc段基因序列无缝嵌合,过渡自然,局部无突变氨基酸,重组成了人源嵌合GcFc片段。
重组人源嵌合GcFc基因片段编码的GcFc多肽(228个氨基酸残基,含铰链区),其N端1~48个氨基酸序列与IgG2的铰链区和CH2结构域N端的部分序列相一致,其19~119位氨基酸序列与IgG4的CH2结构域C端的部分基因编码序列相一致,其C端113~228位氨基酸序列(不包括终止密码子)与IgG2的CH3结构域基因编码序列相一致。
该重组人嵌合GcFc基因片段,具有以下优点:继承了IgG4Fc无补体激活能力的特点;继承了IgG2Fc极弱的FcγR结合能力;无IgG4半抗体重组(半分子交换)的弊端;IgG2Fc和IgG4Fc两者无缝嵌合,过渡自然,局部无突变氨基酸,免疫原性最小化。
需要说明的是:本发明的重组人源嵌合GcFc基因片段并非是免疫球蛋白之不同结构域的杂合Fc区,而是根据免疫球蛋白的基因同源性,巧妙的将来源于不同免疫球蛋白Fc区的基因片段嵌合重组,表达人源免疫球蛋白嵌合Fc片段。该嵌合GcFc基因片的特点是:来源于不同免疫球蛋白的氨基酸残基序列无缝嵌合,过渡自然,序列局部无突变氨基酸。其中,有的氨基酸残基序列只来源于某一结构域的一部分,有的氨基酸残基序列则横跨两个相邻结构域的部分序列。此外,本发明的重组人源嵌合GcFc基因片段的概念中还包括铰链区,铰链区是本发明的重组人源嵌合GcFc基因片段不可分割的组成部分,特定的铰链区序列是确保本发明的重组人源嵌合GcFc基因片段功能发挥不可缺少的一部分。因此,本发明的重组人源嵌合GcFc基因片段与免疫球蛋白之不同结构域的杂合Fc区具有本质不同。
IgGl重链基因序列来源于GenBank:AF237583.1所公开的序列。IgG2重链基因序列来源于GenBank:AF237584.1所公开的序列。IgG3重链基因序列来源于GenBank:AF237585.1所公开的序列。IgG4重链基因序列来源于GenBank:JC036239.1所公开的序列。IgM重链基因序列来源于GenBank:AJ294734.1所公开的序列。IgGl、IgG2、IgG3、IgG4和IgM重链的其他等位基因也包括在本发明的权利要求中。
附图说明
图1为pEGFP-N1载体质粒结构示意图;
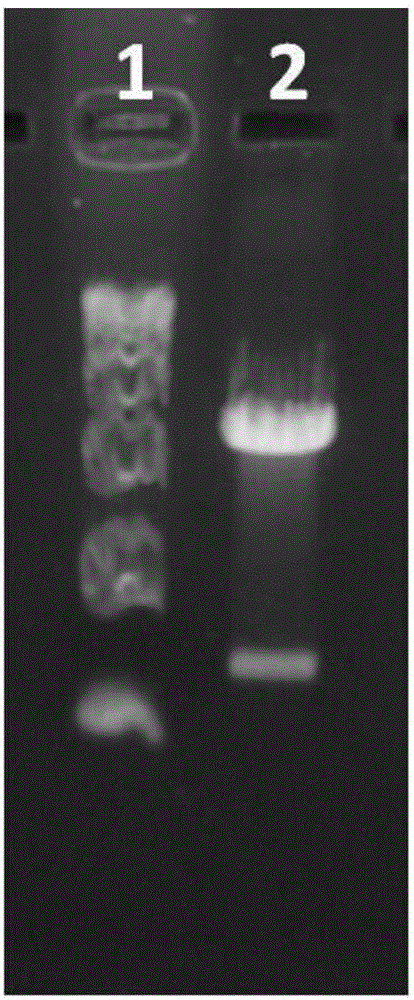
图2为pGcFc载体质粒酶切电泳检测示意图,其中,泳道1:DL15000Marker分子,泳道2为pGcFc载体酶切片段;
图3是构建成功的pGcFc-N1质粒结构示意图;
图4为实施例2中嵌合GcFc氨基酸残基序列。
具体实施方式
下面结合具体实施例对本发明作进一步详细的说明。
实施例1 pGcFc-N1质粒构建。
1.全基因合成GcFc基因序列,克隆到pUC57载体,构建成pUC57-GcFc载体。
全基因合成GcFc基因序列,即将IgG2的铰链区及其CH2结构域N端的部分基因SeqNo.1、IgG4的CH2结构域C端的部分基因SeqNo.3、IgG2的CH3结构域基因SeqNo.3合成为如SeqNo.4所示的基因片段。
其中,IgGl重链基因序列来源于GenBank:AF237583.1所公开的序列。IgG2重链基因序列来源于GenBank:AF237584.1所公开的序列。IgG3重链基因序列来源于GenBank:AF237585.1所公开的序列。IgG4重链基因序列来源于GenBank:JC036239.1所公开的序列。IgM重链基因序列来源于GenBank:AJ294734.1所公开的序列。
2.在两个无菌的0.2ml EP管,分别取pUC57-GcFc载体和pEGFP-N1载体各10μL,用BamHⅠ和NotⅠ限制性内切酶酶切目的基因片段和目的载体,反应体系和反应条件如下:
pEGFP-N1载体质粒结构图如图1所示。BamHI和NotI酶切pEGFP-N1,可将EGFP基因片段切掉。
3.琼脂糖凝胶电泳检测酶切效果,切割在琼脂糖凝胶电泳胶中的目的基因片段条带和线性化的载体,DNA回收试剂盒回收纯化。
4.目的片段与线性化的目的载体连接,构建成pGcFc-N1载体,具体反应体系如下:
5.转化感受态细胞:冰浴中将10μL连接产物分别加入至50μL Top10感受态细胞中,最后涂布于含Kan抗生素(100μg/mL)的LB平板过夜培养。
6.挑取数个菌落,进行PCR检测。
7.阳性克隆测序验证,将菌落PCR鉴定得到的阳性克隆送公司测序进行测序验证,测序完成后,软件比对测序结果,测序引物CMV-F:CGCAAATGGGCGGTAGGCGTG。Blast比对分析:基因GcFc已成功替换pEGFP-N1载体的EGFP序列,原始序列与已知序列进行BLAST 100%一致。构建成功的pGcFc-N1质粒结构图如图3所示。
8.用质粒提取试剂盒进行质粒小提后,取构建完成的pGcFc-N1载体10ul,用BamHⅠ和NotⅠ限制性内切酶双酶切,最后琼脂糖凝胶电泳检测酶切效果,结果如图2所示。
实施例2 重组人源嵌合GcFc基因片段的表达。
将实施例1得到的pGcFc-N1载体转染至表达细胞中,获得嵌合GcFc氨基酸残基,其氨基酸残基序列如图4所示。
图4中,单下划线标示的氨基酸残基来源于IgG2的序列,包括IgG2的铰链区序列、CH2结构域N端的部分序列、以及CH3结构域序列;灰色突出标示的氨基酸残基来源于IgG4CH2结构域C端的部分序列。两者无缝嵌合,过渡自然,局部无突变氨基酸,重组成了人源嵌合GcFc片段
在实施例1和实施例2中,实验仪器采用超净工作台,PCR仪,君意JY600E电泳仪。主要实验试剂:限制性内切酶(Thermo Scientific),DNA回收试剂盒(中美泰和),质粒提取试剂盒(中美泰和),PrimeSTAR Max Premix(2×)(Takara),T4DNA连接酶(Thermo Scientific),XL1-BLUE MRF’感受态细胞,Top10感受态细胞。测序部分为艾基生物公司完成。
以上所述的仅是本发明的一些实施方式。对于本领域的普通技术人员来说,在不脱离本发明创造构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。
- 还没有人留言评论。精彩留言会获得点赞!