一种α‑半乳糖苷酶基因及其应用的制作方法
本发明涉及生物技术领域,具体而言,涉及一种α-半乳糖苷酶基因及其应用。
背景技术:
α-半乳糖苷酶(α-galactosidase),也称蜜二糖酶,是一种外切糖苷水解酶,能够催化水解各类以α键结合的非还原性末端α-半乳糖苷化合物,如蜜二糖、棉子糖家族寡糖(棉子糖、水苏糖等)、乳糖(半乳糖)-甘露聚糖、半乳糖。
在饲料工业中,豆粕是应用最广泛的植物性蛋白质原料,但是豆粕中的α-半乳糖苷糖类含量比较高,如棉子糖、水苏糖和毛蕊花糖等,是饲料中的抗营养因子之一。在饲料中添加α-半乳糖苷酶可有效的降解饲料中的抗营养因子,减少胀气、消化不良等症状,同时提高饲料的营养利用率。
但是在工业生产加工中常常伴随着高温出现,如酶液的喷雾干燥,饲料的制粒、膨化,从而影响α-半乳糖苷酶的应用效果,因此在工业应用中就需要耐高温、高酶活的α-半乳糖苷酶
现在一般通过pcr技术从相关产α-半乳糖苷酶的微生物中扩增出α-半乳糖苷酶的基因,连接到表达载体中,再导入相关表达宿主,表达α-半乳糖苷酶。但是这样获得的α-半乳糖苷酶一般最适温度比较低,最适ph偏中性,蛋白的表达量较小,工业应用限制较大。还有研究人员通过各种手段构建外源基因(一般是小片段基因)的多拷贝表达载体来提高外源基因在宿主中的表达量;还有的将hac1、pdi等基因和外源基因分别导入宿主细胞内来提高外源基因的表达量。
传统上一般是从自然界产α-半乳糖苷酶的微生物中筛选菌株,然后对菌株进行改良,或者对发酵工艺进行优化。这样筛选的α-半乳糖苷酶的酶活和蛋白表达量一般不高,最适温度较低,在工业生产中易失活。
密士军克隆了来源于青霉的α-半乳糖苷酶基因,并在毕赤酵母gs115中表达,表达量最高的一株转化子在3l发酵罐水平α-半乳糖苷酶的酶活力达111u/ml,约为国外最高酶活力的2倍,是国内构建的α-半乳糖苷酶基因工程菌株的3倍。重组酶的最适ph5.0,最适温度为40℃。李荷等克隆了分枝犁头霉absidiaramosawl511的热稳定α-半乳糖苷酶cdna基因aga(genbankno.dq234280),将aga基因插入表达载体ppiczαa并电转化整合到毕赤酵母p.pastorisgs115上表达,酵母发酵上清的酶活达到32u/ml,最适ph为6.8,最适温度73℃。
毕赤酵母表达系统是目前广泛使用的真核生物表达体系。它具有生物安全性好、遗传元件稳定、高密度发酵工艺成熟以及目的蛋白分离纯化简便等优点,是异源蛋白质工业化生产的理想宿主。
然而,由于受到毕赤酵母对异源基因密码子使用偏好性、mrna二级结构的复杂性、基因中富含a/t或g/c区段、蛋白酶切割位点以及基因在宿主中的丰度等因素影响,异源基因在毕赤酵母中难以获得高效表达。
鉴于此,特提出本发明。
技术实现要素:
本发明的目的在于提供一种α-半乳糖苷酶基因,其适于编码α-半乳糖苷酶,该α-半乳糖苷酶基因的碱基序列经过密码子优化,采用了适用于毕赤酵母进行转录翻译表达出α-半乳糖苷酶的高频密码子,有利于提高该α-半乳糖苷酶基因在毕赤酵母细胞中α-半乳糖苷酶的表达量。
本发明的另一目的在于提供一种表达盒,该表达盒含有上述的α-半乳糖苷酶基因。
本发明的另一目的在于提供一种载体,其含有上述的α-半乳糖苷酶基因。
本发明的另一目的在于提供一种重组细胞,其含有上述的载体。
本发明的另一目的在于提供上述α-半乳糖苷酶基因在生产α-半乳糖苷酶中的应用。
本发明的另一目的在于提供上述α-半乳糖苷酶基因在水解棉籽糖或水苏糖中的应用。
本发明是这样实现的:
一种α-半乳糖苷酶基因,其适于编码α-半乳糖苷酶,其核苷酸序列如seqidno:1所示。
一种表达盒,其含有上述的α-半乳糖苷酶基因。
一种载体,其含有上述的α-半乳糖苷酶基因。
一种重组细胞,含有上述的载体。
上述α-半乳糖苷酶基因在生产α-半乳糖苷酶中的应用。
上述α-半乳糖苷酶基因在水解棉籽糖或水苏糖中的应用。
与现有技术相比,本发明的有益效果是:
本发明提供的α-半乳糖苷酶基因,其适于编码α-半乳糖苷酶,其核苷酸序列如seqidno:1所示。该人工设计的α-半乳糖苷酶基因的序列采用了毕赤酵母中的高频密码子替换原有的低频密码子,降低了mrna的自由能及其mrna二级结构的复杂性,并通过对次高频密码子的选用消除了基因中富含at或gc的区域以及蛋白酶作用位点,显著地提高了该人工合成的α-半乳糖苷酶基因在毕赤酵母细胞中的表达量;此外,本发明提供的上述α-半乳糖苷酶基因在生产α-半乳糖苷酶中的应用,通过培养含上述α-半乳糖苷酶基因的细胞,能够高效率、高产量地生产出α-半乳糖苷酶;还有,本发明提供上述α-半乳糖苷酶基因水解棉籽糖或水苏糖中的应用,利用该α-半乳糖苷酶基因编码出的α-半乳糖苷酶能够高效地降解棉籽糖和水苏糖。
附图说明
为了更清楚地说明本发明实施例的技术方案,下面将对实施例中所需要使用的附图作简单地介绍,应当理解,以下附图仅示出了本发明的某些实施例,因此不应被看作是对范围的限定,对于本领域普通技术人员来讲,在不付出创造性劳动的前提下,还可以根据这些附图获得其他相关的附图。
图1为本发明实施例1提供的pao815表达载体的图谱;
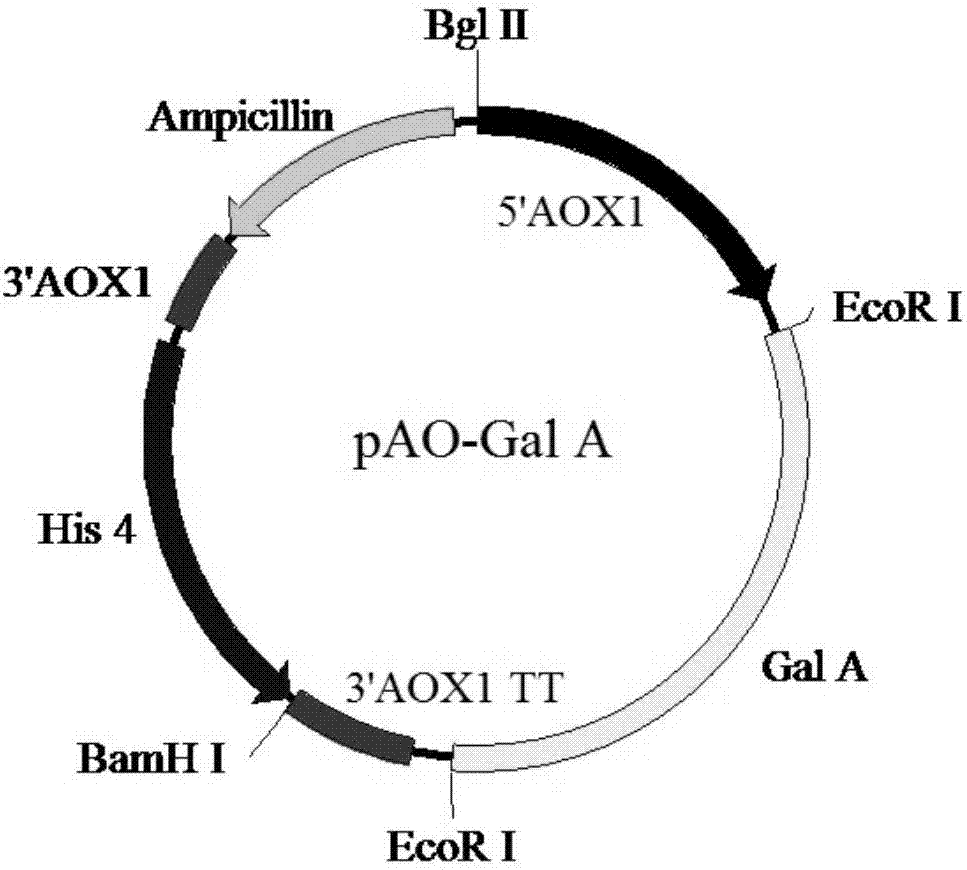
图2为本发明实施例1提供的pao-gala表达载体的图谱;
图3为本发明实施例1提供的单拷贝的pao-gala表达载体的双酶切检验结果图;
图4为本发明实施例1提供的两拷贝α-半乳糖苷酶基因的pao-gala-du表达载体的图谱;
图5为本发明实施例2提供的多拷贝重组表达载体的双酶切检验结果;
图6为本发明实施例2提供的多拷贝重组表达菌株发酵上清液的sds-pege结果;
图7为本发明实施例3提供的共表达重组载体的双酶切检验结果;
图8为本发明实施例3提供的共表达重组载体菌株的发酵上清液的sds-pege结果;
图9为本发明实施例4提供的两拷贝重组表达菌株在不同时间的发酵上清液的sds-pege结果;
图10为本发明实施例5提供的α-半乳糖苷酶水解棉子糖糖的薄层层析图;
图11为本发明实施例6提供的α-半乳糖苷酶水解水苏糖的薄层层析图。
具体实施方式
为使本发明实施例的目的、技术方案和优点更加清楚,下面将对本发明实施例中的技术方案进行清楚、完整地描述。实施例中未注明具体条件者,按照常规条件或制造商建议的条件进行。所用试剂或仪器未注明生产厂商者,均为可以通过市售购买获得的常规产品。
下面对本发明实施例的一种α-半乳糖苷酶基因及其应用进行具体说明。
一方面,本发明提供了一种α-半乳糖苷酶基因,其适于编码α-半乳糖苷酶,其核苷酸序列如seqidno:1所示。其所编码的α-半乳糖苷酶的氨基酸序列如seqidno:2所示。
该α-半乳糖苷酶基因采用了毕赤酵母中的高频密码子替换原有的低频密码子,降低了mrna的自由能及其mrna二级结构的复杂性,并通过对次高频密码子的选用消除了基因中富含at或gc的区域以及脂肪酶中的蛋白酶作用位点,大大地提高了该α-半乳糖苷酶基因在毕赤酵母细胞中的α-半乳糖苷酶的表达量。为后续的工业化高产量地生产出α-半乳糖苷酶奠定了基础。
另一方面,本发明提供了一种表达盒,其含有上述的α-半乳糖苷酶基因。
容易理解,该表达盒包括了用于驱动上述α-半乳糖苷酶基因表达的启动子,启动子位于该α-半乳糖苷酶基因的上游。该启动子的类型可以是强表达型启动子、组织特异性启动或诱导型启动子。在实际中,可根据实际情况选择相应的启动子来进行驱动表达。
优选地,在本发明的一些实施方案中,上述的表达盒包括了诱导型启动子,更优选地,该诱导型启动为甲醇诱导型启动子(aox1,碱基序列如seqidno:7所示),在甲醇的诱导下,该甲醇诱导型启动子可驱动上述的α-半乳糖苷酶基因转录表达出α-半乳糖苷酶。
优选地,在本发明的一些实施方案中,该表达盒还包括了信号肽序列,该信号肽序列位于启动子的下游与该α-半乳糖苷酶基因的上游之间,用于引导α-半乳糖苷酶的跨膜转移。
另一方面,本发明提供了一种载体,其含有上述的α-半乳糖苷酶基因。
优选地,在本发明的一些实施方案中,上述的载体上的α-半乳糖苷酶基因的拷贝数为多个。通过增加基因的拷贝数,可增加上述α-半乳糖苷酶基因的表达量,进一步提高α-半乳糖苷酶的表达量。当然,载体上的每个α-半乳糖苷酶基因具有独立的启动子驱动表达。
更优选地,在本发明的一些实施方案中,上述载体上的α-半乳糖苷酶基因的拷贝数为两个或三个。
优选地,在本发明的一些实施方案中,上述的载体为puc57载体。
上述的α-半乳糖苷酶基因通过酶切后连接至经相同酶切后的α-半乳糖苷酶基因载体上。
另一方面,本发明还提供了一种载体,其含有上述的表达盒。
优选地,在本发明的一些实施方案中,表达盒在载体上的拷贝数为多个,优选为两个或三个。通过表达盒数量的增加,进而增加α-半乳糖苷酶基因的数量,提高其α-半乳糖苷酶的表达量。
优选地,在本发明的一些实施方案中,上述的载体为pao815表达载体。
上述的表达盒通过酶切后连接至经相同酶切后的pao815表达载体上。
当外源基因毕赤酵母内大量表达时,外源蛋白由于无法及时折叠、加工而聚集在内质网中无法分泌到胞外,从而影响外源蛋白的分泌表达。此时细胞会激活内质网非折叠蛋白质反应(unfoldedproteinreponse,upr),提高内质网中蛋白二硫键异构酶(pdi)和分子伴侣等相关蛋白表达量,促进内质网中滞留蛋白的折叠,增加外源蛋白的分泌量。pdi能促进二硫键的形成和异构化,防止新生多肽多聚体的产生,使错误折叠的蛋白质正确折叠;内质网压力响应蛋白(hac1)是upr过程中重要的信号传递分子,hac1能与upre(unfoldedproteinresponseelement)结合,促进upre目的基因持续表达;hsp40家族的分子伴侣主要功能是结合错误折叠的蛋白并将这些蛋白转移到hsp70用于重折叠或降解;ero1在内质网中氧化还原态的pdi成氧化态,氧化态的pdi促进蛋白形成二硫键时被还原成还原态,从而间接帮助蛋白分泌。
基于此,优选地,在本发明的一些实施方案中,上述任一项载体含有细胞内质网分泌蛋白相关基因,该细胞内质网分泌蛋白相关基因选自hac1基因、pdi基因、ero1基因和hsp40基因中的任意一种或几种。
其中,hac1基因的核苷酸序列如seqidno:3所示,pdi基因的核苷酸序列如seqidno:4所示,ero1基因的核苷酸序列如seqidno:5所示,hsp40基因的核苷酸序列如seqidno:6所示。
将hac1基因、pdi基因、ero1基因和hsp40基因中的任意一种或几种基因的组合与上述的α-半乳糖苷酶基因构建至同一个表达载体上,形成共表达重组载体,通过提高内质网折叠加工新生多肽链、分解未折叠和错误折叠蛋白的能力,进一步提高α-半乳糖苷酶基因的表达量。
本发明通过构建α-半乳糖苷酶基因多拷贝载体并与细胞内质网分泌蛋白相关基因共表达的方法获得了高效表达的α-半乳糖苷酶基因工程载体,进一步显著提高了α-半乳糖苷酶基因的表达量。
另一方面,本发明提供含有上述载体的重组细胞。
需要说明的是,该重组细胞可以是大肠杆菌,也可以是酵母等细胞,或者是其他类型的细胞例如动物细胞等。其均属于本发明的保护范围。
优选地,在本发明的一些实施方案中,上述重组细胞由上述任一项载体导入毕赤酵母菌后得到。
再一方面,本发明提供了上述α-半乳糖苷酶基因在生产α-半乳糖苷酶中的用
进一步地,上述应用包括:培养含上述α-半乳糖苷酶基因的重组细胞。例如将含有上述载体(该载体含上述的α-半乳糖苷酶基因)的毕赤酵母进行发酵培养,发酵上清液经过提纯后即可得到α-半乳糖苷酶。
本发明提供的α-半乳糖苷酶基因经过密码子优化,采用了适用于毕赤酵母进行转录翻译表达出α-半乳糖苷酶的高频密码子,因此,以毕赤酵母作为宿主细胞,有利于提高该α-半乳糖苷酶基因在毕赤酵母细胞中α-半乳糖苷酶的表达量。相应地,采用上述α-半乳糖苷酶基因生产α-半乳糖苷酶的方法可高效率、高产量地生产出α-半乳糖苷酶。
另一方面,本发明还提供了上述α-半乳糖苷酶基因在水解棉籽糖或水苏糖中的应用。
进一步地,上述应用包括:用上述α-半乳糖苷酶基因表达得到的α-半乳糖苷酶接触棉籽糖或水苏糖。
采用本发明提供的α-半乳糖苷酶基因表达出的α-半乳糖苷酶具有最适ph为4.5、最适温度为50℃的特性,其具有正常的生物酶活性,能够高效地降解棉籽糖和水苏糖。
以下结合实施例对本发明的特征和性能作进一步的详细描述。
实施例1
构建含α-半乳糖苷酶单拷贝载体的菌株,方法如下。
1.1根据已有的α-半乳糖苷酶的基因(来自黑曲霉(aspergillusniger))序列进行人工设计,设计出一条全新的α-半乳糖苷酶基因序列,并通过人工合成的方法获得该α-半乳糖苷酶基因片段,其碱基序列如seqidno:1所示,命名为gala。该α-半乳糖苷酶基因编码出的α-半乳糖苷酶的氨基酸序列如seqidno:2所示。
为提高α-半乳糖苷酶基因在毕赤酵母中的表达量,本实施例在dna2.0等软件的辅助下首先insilico设计了该α-半乳糖苷酶基因序列。在设计过程中主要考虑了以下因素:
(1)采用p.pastoris中的高频密码子替换原有的低频密码子;
(2)降低了mrna的自由能;
(3)通过对次高频密码子的选用消除了基因中富含at或gc的区域;
(4)消除脂肪酶中的蛋白酶作用位点。
经过重新设计后,有效地提高了gala基因中含量较高氨基酸密码子的使用频率。同时,通过优化,有效地降低了gala基因转录出的mrna二级结构的复杂性,有利于α-半乳糖苷酶表达量的提高。
1.2将合成的gala基因片段连接到毕赤酵母表达载体pao815(结构如图1所示)中,得到具有单拷贝gala基因的pao-gala表达载体。
1.2.1在合成的gala基因两端加上酶切位点ecori,经ecori酶切后,连接至经ecori酶切后的中间载体puc57载体上,得到puc-gala载体。
1.2.2将puc-gala载体和pao815载体分别用ecori酶切,酶切体系:30μl的载体、2μl的ecori、10μl的10×buffer、ddh2o补齐至200μl,37℃条件下、酶切2h左右;
1.2.3电泳、胶回收gala基因片段和pao815片段,将两个片段用t4dna连接酶;
连接体系:1μl的t4buffer、1μl的t4dna连接酶、5.5μl的gala基因片段、2.5μl的pao815片段、ddh2o补齐至10μl;在16℃条件下连接8-10h后,转入大肠杆菌中,筛选阳性克隆,提取质粒,即得到具有单拷贝α-半乳糖苷酶基因的重组质粒pao-gala表达载体,其结构如图2所示,gala基因由aox1甲醇诱导型启动子驱动,由aox1(tt)终止子终止表达。
1.2.4利用电转仪将该pao-gala表达载体转化入巴斯德毕赤酵母(pichipastorisx-33,在其他的实施例中可选用其他的毕赤酵母菌株)中,然后在ypd平板上筛选得到阳性转化子,经pcr验证为正确的阳性转化子。提取质粒载体,进行双酶切(bglⅱ和bamhⅰ)验证,结果如图3所示(图中:m为dl5000dnamarker,1泳道为没有酶切的pao-gala重组表达载体;2泳道为双酶切后的pao-gala重组表达载体),在1300bp位置有条带,说明gala基因成功连接至pao815表达载体上。
实施例2
构建具有多拷贝α-半乳糖苷酶基因的重组表达载体
2.1通过同尾酶双酶切法获得含α-半乳糖苷酶基因的表达盒片段,将表达盒片段连接到α-半乳糖苷酶单拷贝表达载体上获得两拷贝表达载体。
2.1.1将实施例1得到的pao-gala重组表达载体用bglii和bamhi双酶切后,胶回收小片段(即aox-gala表达盒),酶切体系:30μl的pao-gala表达载体、1.5μl的bamhi、1.5μl的bglii、20μl的10×bufferk、20μl的bsa、20μl的tritonx-100、ddh2o补齐至200μl,37℃条件下,酶切4h左右。
2.1.2用bamhi单酶切新的pao-gala表达载体后胶回收,将其与步骤2.1.1得到的aox-gala片段用t4dna连接酶连接,得到具有两拷贝α-半乳糖苷酶基因的重组质粒即pao-gala-du表达载体,其结构如图4所示。
2.1.3根据上述方法,可获得含具有多拷贝α-半乳糖苷酶基因的重组表达载体。
2.1.4将得到的多拷贝重组表达载体分别转到毕赤酵母感受态细胞中,筛选得到阳性克隆子,可获得含多拷贝α-半乳糖苷酶基因的多拷贝重组表达菌株。
本实施例根据上述方法获得两拷贝重组表达菌株和三拷贝重组表达菌株,分别提取质粒,得到三拷贝α-半乳糖苷酶基因的重组质粒即pao-gala-th表达载体,进行双酶切验证,结果如图5所示(图中:第1泳道m为dnadl5000marker;第2/3/4泳道分别为α-半乳糖苷酶单拷贝、两拷贝、三拷贝重组表达质粒;第5/6/7泳道分别为单拷贝、双拷贝、三拷贝重组质粒经bglⅱ和bamhⅰ双酶切后的电泳结果)。图5结果说明了经双酶切验证后单拷贝、双拷贝、三拷贝重组表达质粒是正确的。
2.1.5培养多拷贝重组表达菌株,获取上清,进行sds-pege,结果如6所示。由图6可看出(图中:1-1、1-2、1-3为含单拷贝α-半乳糖苷酶基因的单拷贝重组表达菌株;2-1、2-2、2-3为含两拷贝α-半乳糖苷酶基因的两拷贝重组表达菌株;3-1、3-2、3-3为含三拷贝α-半乳糖苷酶基因的三拷贝重组表达菌株),两拷贝重组表达菌株(图中2-1、2-2、2-3)的上清液中的α-半乳糖苷酶含量较高,这也说明两拷贝的pao-gala-du表达载体的α-半乳糖苷酶的表达量较高。
2.2α-半乳糖苷酶的酶活测定,dns法测α-半乳糖苷酶酶活。
2.2.1取0.5ml适当稀释的上清液(步骤2.1.5得到的),加入到0.5ml1%棉子糖和1mlph4.5磷酸氢二钠-柠檬酸缓冲液混合液中。
2.2.250℃水浴反应10min,用2mldns(3,5-二硝基水杨酸)终止反应。
2.2.3置于沸水浴中5min,加水定容到25ml;对照组加入沸水浴灭活的酶,其它不变。
2.2.4测量od520吸光值,并对照葡萄糖标准曲线算出葡萄糖的浓度,根据后文的酶活公式计算出α-半乳糖苷酶酶活。
2.2.5每分钟水解棉子糖形成1μmol葡萄糖所需的酶量为1个酶活力单位(u)。酶活(u/ml)计算公式:u=(n×d×v1)/(v2×t),其中,n:酶液稀释倍数;d为根据od520吸光值得到的葡萄糖的摩尔浓度(μmol/ml);v1为反应定容体积(ml);v2为加入酶液的体积;t为反应时间(min)。
结果显示:单拷贝重组表达菌株的上清液酶活为:55.74u/ml;两拷贝重组表达菌株的上清液的酶活为:95.22u/ml,两拷贝重组表达菌株的酶活比单拷贝提高了70.83%。
实施例3
构建α-半乳糖苷酶基因与细胞内质网分泌蛋白相关基因共表达载体
3.1将与细胞内质网分泌蛋白相关基因例如hac1、pdi、ero1、hsp40分别与α-半乳糖苷酶基因进行共表达。
3.1.1将hac1、pdi、ero1、hsp40基因分别连接到pao815载体上,构建出细胞内质网分泌蛋白相关基因的重组载体(方法同步骤1.2)。
3.1.2再将含α-半乳糖苷酶基因的表达盒(步骤2.1.1得到的aox-gala表达盒)连接到上述得到的各细胞内质网分泌蛋白相关基因的重组载体中,得到共表达重组载体(方法可参考步骤2.1)。其中,hac1基因的核苷酸序列如seqidno:3所示,pdi基因的核苷酸序列如seqidno:4所示,ero1基因的核苷酸序列如seqidno:5所示,hsp40基因的核苷酸序列如seqidno:6所示。
3.1.3将构建的各共表达重组载体分别转到毕赤酵母感受态中,筛选得到阳性克隆子,可获得不同的细胞内质网分泌蛋白相关基因与gala基因共表达的共表达重组菌株。分别是:hac1-gala共表达重组菌株、pdi-gala共表达重组菌株、hsp40-gala共表达重组菌株、ero1-gala共表达重组菌株。
3.1.4然后,从上述共表达重组菌株中提取共表达重组载体,分别分别是pao-hac1-gala、pao-pdi-gala、pao-ero1-gala、pao-hsp40-gala,对其分别进行酶切验证和pcr验证,结果如图7所示。
由图7的结果显示(图中:第1、2、3、7泳道分别为pao-hac1-gala共表达重组载体、pao-pdi-gala共表达重组载体、pao-hsp40-gala共表达重组载体和pao-ero1-gala共表达重组载体;第4、5、6泳道分别为pao-hac1-gala共表达重组载体的hca1基因、pao-pdi-gala共表达重组载体的pdi基因、pao-hsp40-gala共表达重组载体的hsp40基因的pcr验证结果;第8泳道为pao-ero1-gala共表达重组载体上的ero1基因的pcr验证结果),hac1基因、pdi基因、hsp40基因以及ero1基因均独立与gal基因重组成功,形成了共表达重组载体。
3.2分别培养上述的共表达重组菌株,获取上清,检测各自的酶活(方法参考步骤2.2),结果显示:hac1-gala共表达重组菌株、pdi-gala共表达重组菌株、ero1-gala共表达重组菌株的培养上清液的酶活比单拷贝α-半乳糖苷酶重组表达菌株的酶活分别提高了15.25%、23.25%和26.86%。
3.3分别取共表达重组菌株发酵的上清进行sds-pege,结果如图8所示。由图8可看出(图中:1-1、1-2、1-3为hac1-gala共表达重组菌株,2-1、2-2、2-3为pdi-gala共表达重组菌株,3-1、3-2、3-3为hsp40-gala共表达重组菌株,4-1、4-2、4-3为ero1-gala共表达重组菌株),ero1-gala共表达重组菌株的培养上清液中的α-半乳糖苷酶含量较高。
3.4经鉴定,上述得到的各重组毕赤酵母菌种细胞呈白色球形,为好氧型酵母菌,生长过程中温度为28℃,产α-半乳糖苷酶需要以甲醇为碳源进行诱导且最适温度为25℃。
实施例4
本实施例提供了一种生产α-半乳糖苷酶的方法,其包括:培养上述的实施例2提供的含pao-gala-du表达载体的两拷贝重组表达菌。
4.1对含pao-gala-du表达载体的两拷贝重组表达菌进行发酵培养,参考如下步骤。
4.1.1将实施例2的两拷贝重组表达菌接种400ml至含7l发酵培养基的发酵罐中。
其中,发酵培养基参考表1中的成分进行配制。
表1
4.1.2生物量积累阶段:
控制参数如下:罐温28℃,ph6.0左右,转速200-500rpm,通气量2.0-5.5l/min,持续约20h。
4.1.3营养流加阶段:发酵开始20h后,流加甘油和葡萄糖的混合溶液(甘油和葡萄糖各称取70g,用水定容至280ml),补充营养,发酵参数不变,流速约为2.3ml/min,持续约2h。
4.1.4饥饿阶段:营养流加结束后开始计时,2h后开始流加甲醇,此时发酵罐溶氧会不断上升,逐渐降低通气量和转速。
4.1.5甲醇流加第一阶段:此阶段为酵母适应甲醇碳源阶段,在2h内使甲醇含量达到0.5%,温度降到25℃。
4.1.6甲醇流加第二阶段:酵母适应甲醇后,溶氧开始下降,调节转速和通气使溶氧维持在30%左右,温度25℃,ph5.5左右,甲醇流速为4.0ml/l/h。
4.1.7甲醇流加第三阶段:发酵到达后期时,溶氧波动较大,此时减少甲醇流速至2.5ml/l/h,使溶氧维持在20%左右。当发酵液ph迅速增加时表明酵母细胞开始破碎,此时结束发酵,离心,取上清,得到含α-半乳糖苷酶的发酵上清液
4.1.8将发酵上清液提纯后可得到α-半乳糖苷酶(可选步骤)。
4.1.9发酵过程中,于不同时间点(24h、48h、72h、96h,接种时间作为发酵起始时间开始计时即接种时间点记为发酵0h)取发酵液上清,进行sds-pege,结果图9所示。由图9的结果可知,在发酵120h时,α-半乳糖苷酶的产量最高。
此外,测得发酵液酶活在最适温度和ph,稀释倍数为40倍时酶活为791.23u/ml;加入终浓度20mmol/ml的fecl2到反应体系中时,测得酶活达到1400u/ml。金属离子对酶可以起到催化作用,活性中心的形成及结构稳定作用,调节作用等,反应体系中加入fecl2可以使α-半乳糖苷酶水解棉子糖的能力提高,即提高了α-半乳糖苷酶的酶活。
实施例5
本实施例提供了一种水解棉子糖的方法,其包括:用实施例4得到的α-半乳糖苷酶接触水苏糖。具体如下:
取实施例4得到中的含α-半乳糖苷酶的发酵上清液作为酶原液,作5倍、10倍、20倍、40倍稀释后,各取0.5ml分别加入到0.5ml1%棉子糖和1mlph4.5磷酸氢二钠-柠檬酸缓冲液混合液中,50℃水浴反应5min。
取反应液0.2μl进行薄层层析,展层剂配方:丙醇:乙酸:水(v:v:v)=1:1.5:0.1;显色剂配方:正丁醇:乙酸:水:磷酸(v:v:v:v)=80:8:5:7,再加入3%(m/v)的尿素。薄层层析方法参考如下:
(1)将含1%的蔗糖,1%的棉子糖,1%的水苏糖的溶液作为对照依次点2μl在同一点上,将不同稀释倍数的反应体系的产物依次点在不同点上。
(2)把薄层层析板置于含有展开剂的玻璃器皿中放置2h左右,取出薄层层析板用吹风机吹干
(3)往薄层层析板上均匀喷洒显色剂后用吹风机吹干
(4)将薄层层析板置于108℃烘箱中5min,结果如图10所示(图中:m为对照品,1为酶原液、2为5倍稀释液,3为10倍稀释液,4为20倍稀释液,5为40倍稀释液)。
从图10中可看出,随着酶原液稀释倍数的增加,水解能力下降,在20倍和40倍稀释倍数下,部分棉子糖未水解(如图中箭头所指),在酶原液和5倍、10倍稀释条件下,棉子糖完全水解,说明本发明实施例4得到的α-半乳糖苷酶可将棉子糖完全水解,其具有水解棉子糖的活性。
实施例6
本实施例提供了一种水解水苏糖的方法,其包括:用实施例4得到的α-半乳糖苷酶接触水苏糖。具体如下:
取实施例4得到中的含α-半乳糖苷酶的发酵上清液作为酶原液,作10倍、20倍稀释后,各取0.5ml分别加入到0.5ml1%水苏糖和1mlph4.5磷酸氢二钠-柠檬酸缓冲液混合液中,50℃水浴反应5min。
取反应液0.2ul进行薄层层析,方法同实施例5,结果图11所示(图中:m为对照品,1为酶原液、2为10倍稀释液,3为20倍稀释液)。
从图11中可看出,随着酶原液稀释倍数的增加,水苏糖的显色程度加深,说明本发明实施例4得到的α-半乳糖苷酶可将水苏糖完全水解,具有水解水苏糖的活性。
综上,本发明通过选择毕赤酵母偏好密码子等手段进行基因优化后,将合成的α-半乳糖苷酶基因(seqidno:1)与pao815表达载体连接后,转入毕赤酵母x-33中表达,重组表达菌株小摇瓶发酵上清的酶活达到55.74u/ml,α-半乳糖苷酶的最适ph为4.5,最适温度为50℃,加入终浓度20mmol的fecl2后酶活达到98.66u/ml。
此外,本发明将细胞内质网分泌蛋白相关基因hac1、pdi、ero1、和hsp40分别与α-半乳糖苷酶基因gala共表达后,hac1、pdi、ero1基因均能提高α-半乳糖苷酶的表达量。
还有,本发明还构建了多拷贝表达盒来增加α-半乳糖苷酶基因gala在毕赤酵母中的基因剂量。结果表明,两拷贝的重组表达菌株的酶活达到95.22u/ml,加入终浓度20mmol的fecl2后酶活达到168.54u/ml。在14l发酵罐中发酵120h的发酵上清液在ph4.6,50℃,稀释40倍,终浓度20mmol的fecl2条件下酶活达到1400u/ml。
总之,本发明提供的α-半乳糖苷酶基因(seqidno:1),采用了毕赤酵母中的高频密码子替换原有的低频密码子,降低了α-半乳糖苷酶基因转录出的mrna的自由能及其mrna二级结构的复杂性,并通过对次高频密码子的选用消除了基因中富含at或gc的区域以及脂肪酶中的蛋白酶作用位点,大大地提高了该α-半乳糖苷酶基因在毕赤酵母细胞中的表达量,且将含有该α-半乳糖苷酶基因的载体转入毕赤酵母菌株后得到的α-半乳糖苷酶具有生物酶活性,该α-半乳糖苷酶具有最适ph为4.5、最适温度为50℃的特性,其能够对棉子糖和水苏糖完全水解。本发明提供的α-半乳糖苷酶基因具有广阔的应用前景,可在发酵领域应用以高效率、高产量地生产出α-半乳糖苷酶。
以上所述仅为本发明的优选实施例而已,并不用于限制本发明,对于本领域的技术人员来说,本发明可以有各种更改和变化。凡在本发明的精神和原则之内,所作的任何修改、等同替换、改进等,均应包含在本发明的保护范围之内。
sequencelisting
<110>武汉轻工大学
<120>一种α-半乳糖苷酶基因及其应用
<160>7
<170>patentinversion3.5
<210>1
<211>1341
<212>dna
<213>人工序列
<400>1
atgagatggttgttgacttcttctgttttgttggttccagctactgctttggttagacca60
gatggtgttggtagaactccagctttgggttggaactcttggaacgcttactcttgtgat120
attgatgctgataagattgttactgctgctaacgaagttgttaacttgggtttgaaggat180
ttgggttacgaatacattaacattgatgattgttggtctgttaagtctggtagaaacact240
actactaagagaattattccagatccagataagtttccaaacggtatttctggtgttgct300
gatcaagttcatgctttgggtttgaagttgggtatttactcttctgctggtttgactact360
tgtgctggttacccagcttctttgggttacgaagaaattgatgctcaatcttttgctgaa420
tggggtgttgattacttgaagtacgataactgtggtgttccaactaacttgactgatcaa480
tacacttactgtgttccagatactactgatggttctaactttccaaacggtacttgtgtt540
gatttgactgatgctgctccaccaggttacgattgggctacttctactactgctaagaga600
taccaaagaatgagagatgctttgttgtctgttaacagaactattttgtactctttgtgt660
gattggggtcaagctgatgttaacgcttggggtaacgaaactggtaactcttggagaatg720
tctggtgatattactgctacttggtctagaattgctgaaattgctaacgaaaactctttt780
tacatgaactacgctaacttttggggttacccagatccagatatgttggaagttggtaac840
ggtaacttgactttggctgaaaacagagctcattttgctttgtgggctatgatgaaggct900
ccattgattattggtactccattggattctattgatacttctcatttgactattttgttg960
aacaagccattgttgacttttcatcaagatgctgttattggtagaccagcttacccatac1020
aagtggggttacaacccagattggacttttgatgctgaacatccagctgaatactggtct1080
ggtccaacttcttctggtgaagtttttgttttgatgttgaactctgaagatggtgttaag1140
actagaactgctgtttggggtgaagttccagaattgaagggtaagaacaagaactctaag1200
ggtaagaagggtaagaaggattctggttttaaggtttctgatgcttggactggtaaggat1260
ttgggttgtgttaaggataagtacgaagttaagttgcaagctcatgatgttgctgttttg1320
gttgttggtgaagaatgttaa1341
<210>2
<211>446
<212>prt
<213>人工序列
<400>2
metargtrpleuleuthrserservalleuleuvalproalathrala
151015
leuvalargproaspglyvalglyargthrproalaleuglytrpasn
202530
sertrpasnalatyrsercysaspileaspalaasplysilevalthr
354045
alaalaasngluvalvalasnleuglyleulysaspleuglytyrglu
505560
tyrileasnileaspaspcystrpservallysserglyargasnthr
65707580
thrthrlysargileileproaspproasplyspheproasnglyile
859095
serglyvalalaaspglnvalhisalaleuglyleulysleuglyile
100105110
tyrserseralaglyleuthrthrcysalaglytyrproalaserleu
115120125
glytyrglugluileaspalaglnserphealaglutrpglyvalasp
130135140
tyrleulystyraspasncysglyvalprothrasnleuthraspgln
145150155160
tyrthrtyrcysvalproaspthrthraspglyserasnpheproasn
165170175
glythrcysvalaspleuthraspalaalaproproglytyrasptrp
180185190
alathrserthrthralalysargtyrglnargmetargaspalaleu
195200205
leuservalasnargthrileleutyrserleucysasptrpglygln
210215220
alaaspvalasnalatrpglyasngluthrglyasnsertrpargmet
225230235240
serglyaspilethralathrtrpserargilealagluilealaasn
245250255
gluasnserphetyrmetasntyralaasnphetrpglytyrproasp
260265270
proaspmetleugluvalglyasnglyasnleuthrleualagluasn
275280285
argalahisphealaleutrpalametmetlysalaproleuileile
290295300
glythrproleuaspserileaspthrserhisleuthrileleuleu
305310315320
asnlysproleuleuthrphehisglnaspalavalileglyargpro
325330335
alatyrprotyrlystrpglytyrasnproasptrpthrpheaspala
340345350
gluhisproalaglutyrtrpserglyprothrserserglygluval
355360365
phevalleumetleuasnsergluaspglyvallysthrargthrala
370375380
valtrpglygluvalprogluleulysglylysasnlysasnserlys
385390395400
glylyslysglylyslysaspserglyphelysvalseraspalatrp
405410415
thrglylysaspleuglycysvallysasplystyrgluvallysleu
420425430
glnalahisaspvalalavalleuvalvalglygluglucys
435440445
<210>3
<211>717
<212>dna
<213>人工序列
<400>3
atggaaatgactgattttgaactaactagtaattcgcaatcgaacttggctatccctacc60
aacttcaagtcgactctgcctccaaggaaaagagccaagacaaaagaggaaaaggaacag120
cgaaggatcgagcgtattttgagaaacagaagagctgctcaccagagcagagagaaaaaa180
agactacatctgcagtatctcgagagaaaatgttctcttttggaaaatttactgaacagc240
gtcaaccttgaaaaactggctgaccacgaagacgcgttgacttgcagccacgacgctttt300
gttgcttctcttgacgagtacagggatttccagagcacgaggggcgcttcactggacacc360
agggccagttcgcactcgtcgtctgatacgttcacaccttcacctctgaactgtacaatg420
gagcctgcgactttgtcgcccaagagtatgcgcgattccgcgtcggaccaagagacttca480
tgggagctgcagatgtttaagacggaaaatgtaccagagtcgacgacgctacctgccgta540
gacaacaacaatttgtttgatgcggtggcctcgccgttggcagacccactctgcgacgat600
atagcgggaaacagtctaccctttgacaattcaattgatcttgacaattggcgtaatcca660
gaagcgcagtcaggtttgaattcatttgaattgaatgatttcttcatcacttcatga717
<210>4
<211>1399
<212>dna
<213>人工序列
<400>4
gaagactccgctgtcgttaagttggccaccgactccttcaatgagtacattcagtcgcac60
gacttggtgcttgcggagttttttgctccatggtgtggccactgtaagaacatggctcct120
gaatacgttaaagccgccgagactttagttgagaaaaacattaccttggcccagatcgac180
tgtactgaaaaccaggatctgtgtatggaacacaacattccagggttcccaagcttgaag240
attttcaaaaacagcgatgttaacaactcgatcgattacgagggacctagaactgccgag300
gccattgtccaattcatgatcaagcaaagccaaccggctgtcgccgttgttgctgatcta360
ccagcttaccttgctaacgagacttttgtcactccagttatcgtccaatccggtaagatt420
gacgccgacttcaacgccaccttttactccatggccaacaaacacttcaacgactacgac480
tttgtctccgctgaaaacgcagacgatgatttcaagctttctatttacttgccctccgcc540
atggacgagcctgtagtatacaacggtaagaaagccgatatcgctgacgctgatgttttt600
gaaaaatggttgcaagtggaagccttgccctactttggtgaaatcgacggttccgttttc660
gcccaatacgtcgaaagcggtttgcctttgggttacttattctacaatgacgaggaagaa720
ttggaagaatacaagcctctctttaccgagttggccaaaaagaacagaggtctaatgaac780
tttgttagcatcgatgccagaaaattcggcagacacgccggcaacttgaacatgaaggaa840
caattccctctatttgccatccacgacatgactgaagacttgaagtacggtttgcctcaa900
ctctctgaagaggcgtttgacgaattgagcgacaagatcgtgttggagtctaaggctatt960
gaatctttggttaaggacttcttgaaaggtgatgcctccccaatcgtgaagtcccaagag1020
atcttcgagaaccaagattcctctgtcttccaattggtcggtaagaaccatgacgaaatc1080
gtcaacgacccaaagaaggacgttcttgttttgtactatgccccatggtgtggtcactgt1140
aagagattggccccaacttaccaagaactagctgatacctacgccaacgccacatccgac1200
gttttgattgctaaactagaccacactgaaaacgatgtcagaggcgtcgtaattgaaggt1260
tacccaacaatcgtcttatacccaggtggtaagaagtccgaatctgttgtgtaccaaggt1320
tcaagatccttggactctttattcgacttcatcaaggaaaacggtcacttcgacgtcgac1380
ggtaaggccttgtacgaag1399
<210>5
<211>1692
<212>dna
<213>人工序列
<400>5
atgagattaagaaccgccattgccacactgtgcctcacggcttttacatctgcaacttca60
aacaatagctacatcgccaccgaccaaacacaaaatgcctttaatgacactcacttttgt120
aaggtcgacaggaatgatcacgttagtcccagttgtaacgtaacattcaatgaattaaat180
gccataaatgaaaacattagagatgatctttcggcgttattaaaatctgatttcttcaaa240
tactttcggctggatttatacaagcaatgttcattttgggacgccaacgatggtctgtgc300
ttaaaccgcgcttgctctgttgatgtcgtagaggactgggatacactgcctgagtactgg360
cagcctgagatcttgggtagtttcaataatgatacaatgaaggaagcggatgatagcgat420
gacgaatgtaagttcttagatcaactatgtcaaaccagtaaaaaacctgtagatatcgaa480
gacaccatcaactactgtgatgtaaatgactttaacggtaaaaacgccgttctgattgat540
ttaacagcaaatccggaacgatttacaggttatggtggtaagcaagctggtcaaatttgg600
tctactatctaccaagacaactgttttacaattggcgaaactggtgaatcattggccaaa660
gatgcattttatagacttgtatccggtttccatgcctctatcggtactcacttatcaaag720
gaatatttgaacacgaaaactggtaaatgggagcccaatctggatttgtttatggcaaga780
atcgggaactttcctgatagagtgacaaacatgtatttcaattatgctgttgtagctaag840
gctctctggaaaattcaaccatatttaccagaattttcattctgtgatctagtcaataaa900
gaaatcaaaaacaaaatggataacgttatttcccagctggacacaaaaatttttaacgaa960
gacttagtttttgccaacgacctaagtttgactttgaaggacgaattcagatctcgcttc1020
aagaatgtcacgaagattatggattgtgtgcaatgtgatagatgtagattgtggggcaaa1080
attcaaactaccggttacgcaactgccttgaaaattttgtttgaaatcaacgacgctgat1140
gaattcaccaaacaacatattgttggtaagttaaccaaatatgagttgattgcactatta1200
cagactttcggtagattatctgaatctattgaatctgttaacatgttcgaaaaaatgtac1260
gggaaaaggttaaacggttctgaaaacaggttaagctcattcttccaaaataacttcttc1320
aacattttgaaggaggcaggcaaatcgattcgttacaccatagagaacatcaattccact1380
aaagaaggaaagaaaaagactaacaattctcaatcacatgtatttgatgatttaaaaatg1440
cccaaagcagaaatagttccaaggccctctaacggtacagtaaataaatggaagaaagct1500
tggaatactgaagttaacaacgttttagaagcattcagatttatttatagaagctatttg1560
gatttacccaggaacatctgggaattatctttgatgaaggtatacaaattttggaataaa1620
ttcatcggtgttgctgattacgttagtgaggagacacgagagcctatttcctataagcta1680
gatatacaataa1692
<210>6
<211>1140
<212>dna
<213>人工序列
<400>6
atggttaaagaaactaagttttacgatattctaggtgtttcagtcactgccactgatgtc60
gaaattaagaaagcttatagaaaatgcgccttaaaataccatccagataagaatccaagt120
gaggaagctgcagaaaagttcaaagaagcttcagcagcctatgaaattttatcagattct180
gaaaagagagaggtatatgaccaatttggtgaagatggtctaagtggtgctggtggcgct240
ggcggattcccaggtggtggattcggttttggtgacgatatcttttcccaattctttggt300
gctggtggcgcacaaagaccaagaggtcctcaaagaggtaaagatatcaagcatgaaatt360
tctgcctcacttgaagaattatataagggtaggacagctaagttagcccttaacaaacag420
atcctatgtaaagaatgtgaaggtcgtggtggtaagaaaggcgccgtcaagaagtgtacc480
agctgtaatggtcaaggtattaaatttgtaacaagacaaatgggtccaatgatccaaaga540
ttccaaacagagtgtgatgtctgtcacggtactggtgatatcattgatcctaaggatcgt600
tgtaaatcttgtaacggtaagaaagttgaaaacgaaaggaagatcctagaagtccatgtc660
gaaccaggtatgaaagatggtcaaagaatcgttttcaaaggtgaagctgaccaagcccca720
gatgtcattccaggtgatgttgtcttcatagtttctgagagaccacacaagagtttcaag780
agagatggtgatgatttagtatatgaggctgaaattgatctattgactgctatcgctggt840
ggtgaatttgcattggaacatgtttctggtgattggttaaaggtcggtattgttccaggt900
gaagttattgccccaggtatgcgtaaggtcatcgaaggtaaaggtatgccaattccaaaa960
tacggtggctatggtaatttaatcatcaaatttactatcaagttcccagaaaaccatttc1020
acatcagaagaaaacttgaagaagttagaagaaattttgcctccaagaattgtcccagcc1080
attccaaagaaagctactgtggacgaatgtgtactcgcagactttgacccagccaaatac1140
<210>7
<211>939
<212>dna
<213>人工序列
<400>7
gatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacagg60
tccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgtt120
gcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaacca180
gcccagttattgggcttgattggagctcgctcattccaattccttctattaggctactaa240
caccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgtttat300
ttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctga360
gtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgctg420
tcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcgt480
tgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcggcataccgtttgtc540
ttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtctc600
tctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgctt660
tttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatactg720
ctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaata780
tataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagctta840
ctttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacgac900
aacttgagaagatcaaaaaacaactaattattcgaaacg939
- 还没有人留言评论。精彩留言会获得点赞!