大麦HvHOX9基因及其用途的制作方法
本发明涉及基因工程技术领域,具体涉及大麦hvhox9基因及其在调节大麦耐铝性能方面的用途。
背景技术:
铝是地壳中最丰富的金属元素,而在土壤中铝主要以难溶性的化合物存在。但是随着人类活动的加剧,酸雨沉降和生产肥料的大量使用导致土壤酸化严重。据统计,酸性土壤约占世界可耕地的50%(kochian等,2004)。当土壤ph低于5.0,铝就会从矿物态解离为离子态(其中以al3+离子毒性最强)释放到土壤中,并且ph越低,铝的溶解性越大,活性铝对植物有毒害作用,抑制根系生长以及水分和养分的吸收。铝毒害是酸性土壤上限制农作物产量的主要因素之一。
因此,开展作物耐酸铝胁迫的生理生化与分子机理研究,揭示作物酸铝耐性机制,具有重要的理论和实践价值。
铝毒害的作用部位主要是根尖,它能抑制植物根尖细胞伸长和细胞分裂,从而导致根发育不良、萎缩。植物耐铝机制包括外部排斥机制(质外体)和内部忍耐机制(共质体)。外部排斥机制主要包括:有机酸及磷酸盐的分泌、细胞壁对铝的固定、质膜对铝的选择透性、诱导产生的根际ph屏障、铝被主动输出细胞外等;内部忍耐机制包括共质体中有机酸与蛋白质对铝的螯合、液泡的区室化和抗氧化酶活性的提高等(maron等,2013)。
大麦是位于小麦、玉米和水稻之后的第四大谷类作物。但是大麦对铝胁迫极其敏感,这严重限制了大麦在全球许多酸性土壤农业区的种植,因此,迫切需要筛选耐铝种质,挖掘耐铝相关基因,以培育耐铝大麦品种。
青藏高原一年生野生大麦具有丰富的遗传多样性,与栽培大麦无生殖隔离,是一种无需特殊技术和设施即可直接加以利用的初级基因库,因此从野生大麦中发掘优异的抗(耐)性相关基因,进而应用于栽培大麦或其他作物,对作物品种改良具有重要意义。
技术实现要素:
本发明的目的在于提供一种从大麦中克隆得到的具有耐铝特性的基因,为培育耐铝大麦品种提供相关基因及理论基础。
为实现上述目的,本发明从青藏高原一年生野生大麦耐铝基因型xz16(hordeumvulgarel.ssp.spontaneum,六棱大麦)(dai等,2013)中鉴定克隆到的具有耐铝特性的基因hvhox9,来源于xz16。
hvhox9基因克隆与分析:基于根部mirna以及rna-seq结果挑选出一个铝胁迫条件下,在xz16中高表达的hd-zip家族基因,从xz16中克隆了该基因的全长cds区序列,命名为hvhox9。该基因的cds区核苷酸序列如seqidno.1所示。
hvhox9基因cds区全长2523bp,编码一个840aa的蛋白序列,该蛋白分子量为92.21kda,等电点pi=5.72。
本发明还提供了所述的大麦hvhox9基因编码的蛋白质,其氨基酸序列如seqidno.2所示。
hvhox9蛋白序列功能域预测分析,结果显示该蛋白含有3个功能域:hox功能域,start功能域以及mekhla功能域,因此,将该基因分类为hd-zipⅲ家族基因。
将hvhox9蛋白序列进行进化树分析,结果显示该蛋白与水稻oshox9位于同一进化枝上。
研究表明,hvhox9定位在细胞核和细胞膜上。对照情况下,hvhox9基因在植株的根、茎、叶中均有表达,但是铝胁迫下,只在根尖部分该基因的表达显著上调,hvhox9参与调控铝胁迫下xz16根系再生长,并降低细胞壁铝含量;另外,该基因仅受到铝诱导,单独酸、cr、la均不能诱导该基因的表达。
本发明还提供了一种重组质粒bsmv:hvhox9,包括rnaγ载体以及插入rnaγ载体的nheⅰ位点间的目的基因片段,所述目的基因片段的核苷酸序列如seqidno.3所示。本发明将一个281bp的hvhox9基因片段反向插入rnaγ载体的nheⅰ位点间,建立bsmv-vigs体系,成功沉默了xz16中的hvhox9基因。
本发明利用bsmv-vigs方法在野生大麦xz16上验证hvhox9基因功能,结果显示该基因受到铝胁迫的诱导,沉默后大麦的耐铝性显著降低,细胞壁的铝含量显著增加,表明该基因与xz16的耐铝性存在着重要的关系。
本发明提供了所述的大麦hvhox9基因在调控大麦耐铝性中的用途。研究表明,该基因受铝胁迫诱导,提高大麦的耐铝性。bsmv:hvhox9沉默大麦植株耐铝性降低。
本发明还提供一种重组表达载体,包括原始载体和插入所述原始载体的目标基因,所述目标基因的核苷酸序列如seqidno.1所示。
本发明具备的有益效果:
本发明通过对大麦hvhox9基因的克隆和分析,并结合bsmv-vigs技术在xz16上对该基因进行功能验证,结果显示,铝胁迫显著诱导hvhox9基因的上调表达,沉默hvhox9基因后的xz16植株耐铝性明显减弱。本发明为大麦耐铝育种与生产提供了理论依据和相关基因。
附图说明
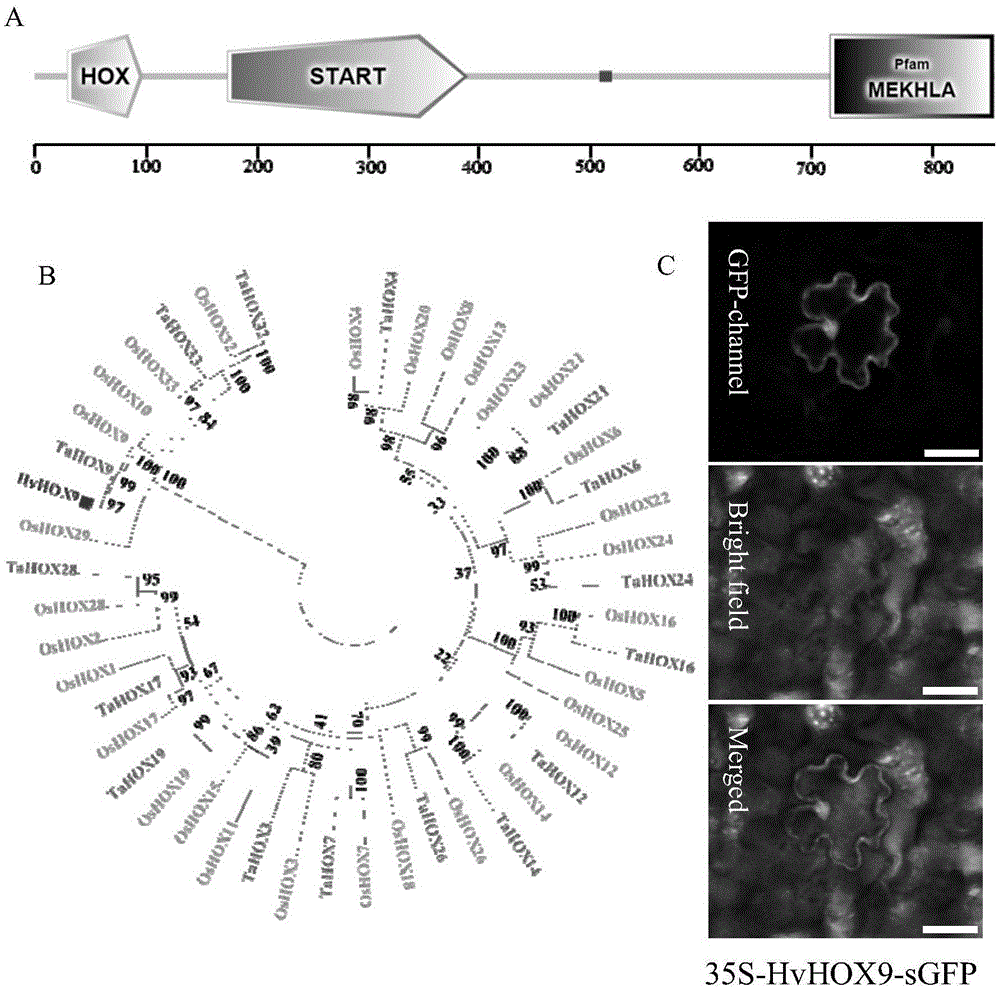
图1为hvhox9功能域预测图(a),hvhox9与水稻、小麦hd-zip家族基因的进化树分析(b),以及hvhox9的亚细胞定位分析(c)。
图2为hvhox9基因的表达模式。(a)铝处理和正常条件下的hvhox9基因的组织定位;(b)hvhox9基因在不同条件下的诱导表达;(c)hvhox9基因在基因型xz16中的时空表达;(d)在基因型xz16中接种bsmv:hvhox9后的基因表达。
图3为bsmv:hvhox9载体构建图、线性化图和体外转录图。(a)bsmv:hvhox9载体构建示意图;(b)rnaγ:hvhox9酶切产物琼脂糖凝胶电泳图;(c)rnaγ:hvhox9线性化琼脂糖凝胶电泳图;(d)rnaγ:hvhox9体外转录琼脂糖凝胶电泳图;其中1是rnaγ:hvhox9,2是rnaγ:hvhox9经mlui酶切,3是rnaγ:hvhox9经nhei酶切,箭头所指为从载体上酶切下来的hvhox9基因片段,4是rnaα经mlui酶切,5是rnaβ经spei酶切,6是rnaγ经mlui酶切,7是rnaγ:hvhox9经mlui酶切,8是体外转录的rnaα,9是体外转录的rnaβ,10是体外转录的rnaγ,11是体外转录的rnaγ:hvhox9。m1是15000bpdnamarker,m2是2000bpdnamarker。
图4为利用bsmv-vigs方法沉默hvhox9后铝胁迫对植株根系生长的影响。a图表示沉默hvhox9后影响根系生长的照片;b图表示沉默hvhox9后根系形态变化的照片;c图表示不同铝浓度处理下沉默hvhox9的根长变化;d图表示不同酸胁迫下沉默hvhox9植株的根长变化;e图表示在不同金属胁迫下沉默hvhox9后根系相对伸长的变化。
图5为利用bsmv-vigs方法沉默hvhox9后铝胁迫对植株干重及铝含量及的影响。a图表示沉默hvhox9后根系干重的变化;b图表示沉默hvhox9后根系铝含量的变化;c图表示沉默hvhox9后根系细胞汁液铝含量的变化;d图表示沉默hvhox9后根系细胞壁铝含量的变化。
图6为利用bsmv-vigs方法沉默hvhox9后铝胁迫对植株根部伸长区h+流以及根际ph的影响。a图表示沉默hvhox9后根部h+流的变化;b表示沉默hvhox9后根际ph的变化。
具体实施方式
下面通过具体实施例,对本发明的技术方案作进一步的具体说明。应当理解,本发明的实施并不局限于下面的实施例,对本发明所做的任何形式上的变通和/或改变都将落入本发明保护范围。
在本发明中,若非特指,所有的份、百分比均为重量单位,所采用的设备和原料等均可从市场购得或是本领域常用的。下述实施例中的方法,如无特别说明,均为本领域的常规方法。
本发明以课题组前期筛选到的青藏高原一年生野生大麦xz16(耐酸铝基因型)为主要材料(戴华鑫等,2013),克隆和分析调控大麦耐铝性的关键基因,对阐明大麦响应酸铝胁迫的分子机理和大麦耐酸铝的育种及生产工作具有重要的指导意义。
实施例1
hvhox9基因cds区克隆和分析
1、hvhox9基因cds区序列的克隆
基于课题组前期研究,从根部mirna以及rna-seq结果中挑选出一个铝胁迫条件下,在xz16中高表达的hd-zip家族基因,从xz16中克隆了该基因的全长cds区序列(如seqidno.1所示),将其命名为hvhox9。
使用总rna提取试剂盒(takara,日本)提取青藏高原一年生野生大麦xz16根部总rna,并用dnasei(takara,日本)去除总rna中基因组dna污染,用primescripttmii1ststrandcdnasynthesiskit反转录试剂盒(takara,日本)将提取的总rna反转录成单链的cdna。根据blast到的序列进行特异性引物设计,具体引物序列为:
hvhox9-cds-f:5'-atggcggcggcggtggcggcgat-3'(seqidno.4);
hvhox9-cds-r:5'-tcacacgaaagaccagttgacg-3'(seqidno.5)。
将扩增出来的产物,连接到pmd18-t(takara,日本)载体,转化大肠杆菌dh5α,挑取阳性克隆送公司测序,测序正确的分别进行提质粒和甘油保存,所得质粒命名为pmd18-t-hvhox9质粒。
pcr引物合成工作由生工生物工程(上海)股份有限公司完成,基因测序工作由上海铂尚生物技术有限公司完成。
2、hvhox9基因序列分析
将hvhox9蛋白序列通过smart(http://smart.embl-heidelberg.de/)网站进行功能域的预测和分析,结果显示该蛋白含有一个hox功能域,一个start功能域,一个mekhla功能域及一个低复杂性区(图1(a))。
通过clustalw和mega6软件将包含了hvhox9在内的不同物种中的hox蛋白序列进行分析,构建系统发育树。结果显示hvhox9在进化上与水稻oshox9位于同一进化枝(图1(b))。
3、hvhox9亚细胞定位分析
以保存的pmd18-t-hvhox9质粒为模板,设计一对引物hvhox9-35s-sgfp-f和hvhox9-35s-sgfp-r扩增hvhox9基因的orf片段(去除终止密码子tga),用于亚细胞定位分析。引物序列为(下划线标出的部分为酶切位点):
hvhox9-35s-sgfp-f:5'-atatgtcgacatggcggcggcggtggcggcgat-3'(seqidno.6),
hvhox9-35s-sgfp-r:5'-ctcagtcgaccacgaaagaccagttgacgaagc-3'(seqidno.7)。
胶回收纯化的orf片段经sali限制性内切酶酶切,并和经同样限制性内切酶酶切的pcambial300-35s-sgfp载体进行连接反应。将连接产物转化大肠杆菌dh5α,阳性克隆送公司测序,测序正确的单克隆(确保无移码,能翻译成完整的基因-sgfp融合表达蛋白)提质粒保存,所得质粒为35s-hvhox9-sgfp。
另取适量该质粒转化农杆菌eha105感受态细胞,然后挑单克隆接种到5ml含有相应抗生素的yeb液体培养基中,28℃过夜摇菌,并取500μl该菌液接种到50ml含有相应抗生素的yeb液体培养基中,摇菌至od600=1.0左右。4000g离心5min收集菌体,并用注射缓冲液(0.2mmas、10mmmes、10mmmgcl2,ph5.6)洗涤菌体两次,最后用该缓冲液重悬菌体至od600=1.0左右,室温下静置3h左右。用1ml注射器吸取该重悬菌液,从烟草叶片背面将重悬菌液注入烟草叶片中(培养两个月左右,5-6叶期烟草),继续置于培养箱中培养(24℃/26℃,16h/8h,白天/夜间),培养36h-72h之间进行荧光观察。观察时选取未受损伤的烟草叶片,撕取部分下表皮,制作样品玻片,于激光共聚焦显微镜下观察荧光分布情况(lsm780,carlzeiss,germany)。
结果显示hvhox9最终定位在细胞核和细胞膜上(图1(c))。
4、hvhox9基因的铝诱导表达情况
青藏高原一年生野生大麦xz16种子用2%的h2o2消毒30min,蒸馏水冲洗干净,之后将种子置于发芽盒内于生长室中暗培养(22℃/18℃)直至萌发,萌发之后补光(22℃/18℃)。6天后选取长势一致的幼苗转移至1l的黑色塑料桶中,每桶4个孔,每孔两株幼苗,用海绵固定,用0.5mmcacl2(ph4.3)溶液预处理1天,之后进行铝处理,设置两个处理,(1)对照(ph4.3的基本培养液);(2)铝处理(ph4.3cacl2+50μmalcl3),24小时后分别取根尖(0-1cm)、根基部(2-4cm)、茎、叶进行hvhox9组织定位及铝诱导表达分析,同样的处理条件分别于处理后0、1、3、6、12、24小时取根尖0-1cm进行hvhox9的时空表达分析。
rt-pcr检测hvhox9基因的表达量
用总rna提取试剂盒(takara,日本)分别提取不同处理样本的总rna,用revertraqpcrrtmastermixwithgdnaremover反转录试剂盒(toyobo,日本)将各样本总rna分别反转录成单链cdna。用sybr绿色荧光酶复合物(toyobo,日本)和lightcycler480pcr仪(roche,瑞士)对相应样本中hvhox9基因的表达进行荧光定量pcr分析(qrt-pcr),并用一个内参基因actin对表达值进行矫正处理。
pcr体系为:
pcr的具体程序为:95℃30s,(95℃5s,60℃10s,72℃15s)40个循环。溶解曲线程序为:60℃-95℃,每步反应5s,按0.5℃递增。利用2-δδcq相对定量方法计算基因表达值变化。每组实验重复三次。rt-pcr引物序列为:
hvhox9-qrt-pcr-f:5'-gatggtgggggttcaatcgt-3'(seqidno.8);
hvhox9-qrt-pcr-r:5'-tccccagcgcatatacaacc-3'(seqidno.9);
actin-f:5'-tggctgacggtgaggaca-3'(seqidno.10);
actin-r:5'-cgagggcgaccaactatg-3'(seqidno.11)。
如图2所示,通过qrt-pcr的结果显示,对照情况下,hvhox9基因在植株的根、茎、叶中均有表达,但是铝胁迫下,只在根尖部分该基因的表达显著上调,另外,该基因仅受到铝诱导,单独酸、cr、la均不能诱导该基因的表达。
该基因在耐铝基因型xz1624小时内表达量随着时间的延长而不断增加,沉默后,不论是对照还是铝胁迫情况下,该基因的表达均显著受到抑制。
实施例2
bsmv-vigs方法验证hvhox9基因功能
1、bsmv:hvhox9载体构建
用pmd18-t-hvhox9质粒为模板,设计引物,扩增得到一个281bp的hvhox9基因片段。
kod酶pcr反应体系:
扩增程序为:94℃5min,(98℃10s,58℃30s,68℃30s)35个循环,68℃10min。
引物序列为(下划线为酶切位点):
hvhox9-γ-f:5'-gtacgctagcctgctgttctgcggacattt-3'(seqidno.12);
hvhox9-γ-r:5'-gtacgctagcataatcagcccattcagacc-3'(seqidno.13)。
将hvhox9基因片段用nheⅰ限制性内切酶酶切,然后与经过同样限制酶内切酶酶切并且去磷酸化的rnaγ载体用t4连接酶连接。连接产物转化大肠杆菌dh5α,用hvhox9基因的正向引物hvhox9-γ-f和rnaγ载体上的引物γ-stain-f验证反向插入,将验证的反向插入的阳性克隆送公司测序,测序正确的单克隆进行摇菌提质粒并进行酶切再次验证(图3(b)),所得的质粒是rnaγ:hvhox9。
所用的rnaγ载体上的引物序列为:
γ-stain-f:5'-caactgccaatcgtgagtagg-3'(seqidno.14)。
pcr引物合成工作由生工生物工程(上海)股份有限公司完成,基因测序工作由上海铂尚生物技术有限公司完成。
2、bsmv载体线性化和体外转录
rnaα、rnaγ和rnaγ:hvhox9用mluⅰ限制性内切酶单酶切,同时rnaβ用speⅰ限制性内切酶单酶切,分别使其线性化,酶切产物跑胶,胶回收并纯化产物(图3(c))。将纯化后的rnaα、rnaβ、rnaγ和rnaγ:hvhox9线性化产物用ribomaxtmlargescalernaproductionsystem-t7kit和ribom7gcapanalogkit试剂盒进行体外转录(promega,美国),均在rnase-free环境下操作,并跑电泳验证结果(图3(d))。
将体外转录后的rnaα、rnaβ和rnaγ质粒按照1:1:1的体积比进行混合,将体外转录后的rnaα、rnaβ和rnaγ:hvhox9质粒按照1:1:1的体积比进行混合,分别加入两倍体积的rnase-free水进行稀释,随后在稀释产物中加入等体积的2×gkp缓冲液(1%膨润土、1%硅藻土、50mm甘氨酸和30mm磷酸氢二钾ph9.2),所得产物命名为bsmv:γ和bsmv:hvhox9,混匀之后用于接种。
3、bsmv接种前大麦幼苗培养
青藏高原一年生野生大麦xz16用2%的h2o2消毒30min,蒸馏水冲洗干净,之后将种子置于发芽盒内于生长室中暗培养(22℃/18℃)直至萌发,萌发之后补光(22℃/18℃)。6天后选取长势一致的幼苗转移至1l的黑色塑料桶中,每桶4个孔,每孔两株幼苗,用海绵固定,桶中盛有基本培养液且持续通气,基本培养液使用1/5hogland营养液配方。每3天更换一次培养液,营养液的ph用naoh或hcl调至5.8±0.1。
4、bsmv接种验证基因功能
当大麦幼苗生长至两叶一心期后,在rnase-free环境下对第二叶进行bsmv摩擦接种,每个植株使用10μl混合均匀的bsmv:γ或bsmv:hvhox9。接种后的植株立即喷施少量的depc水,并用透明的塑料罩罩住植株保湿3天,之后将玻璃罩取掉,在大麦生长培养室中继续培养(22℃/18℃),定时观察植株的表型,7天后进行以下处理。
表型,干重以及元素含量分析。实验共设4个处理:接种bsmv:γ+基本培养液(bns,ph4.3),接种bsmv:γ+bns+100μmalcl3(ph4.3),接种bsmv:hvhox9+bns(ph4.3),接种bsmv:hvhox9+bns+100μmalcl3(ph4.3)。处理5天后,结果显示,与模拟接种bsmv:γ的植株相比,接种bsmv:hvhox9的植株铝处理后主根明显变短(图4(a)),主根根尖部分受到的损伤更加严重(图4(b))。此外,接种bsmv:hvhox9的植株铝处理后根系的干重和铝含量显著提高(图5(a,b)),这些结果表明,hvhox9基因沉默后的xz16植株的耐铝性显著降低。
根系再伸长。分别对接种bsmv:γ和接种bsmv:hvhox9的植株设置以下5个处理,0.5mmcacl2+0/10/20/30/50μmalcl3(ph4.3);处理36小时后,结果显示,根系再伸长量随着铝浓度的增加而降低,且接种bsmv:hvhox9植株的根系伸长量明显低于模拟接种bsmv:γ的植株(图4(c))。这些结果表明,hvhox9参与调控铝胁迫下xz16根系再生长。
分别对接种bsmv:γ和接种bsmv:hvhox9的植株设置以下4个处理,0.5mmcacl2溶液,其ph分别为3.5,4.3,5.0,5.8。处理36h后测量其根长。结果显示,接种bsmv:hvhox9植株的根部抑制不受酸诱导(图4(d))。
分别对接种bsmv:γ和接种bsmv:hvhox9的植株设置以下4个处理,0.5mmcacl2溶液(ph4.3),0.5mmcacl2溶液+50μmalcl3(ph4.3),0.5mmcacl2溶液+20μmcrcl3(ph4.3),0.5mmcacl2溶液+5μmlacl3(ph4.3),处理36h后测量其根长。结果显示,只有在酸铝胁迫下,接种bsmv:hvhox9的植株的相对根系伸长量明显低于接种bsmv:γ植株,其他金属胁迫无显著差异(图4(e))。
根部细胞汁液和细胞壁铝含量测定。实验共设4个处理:(1)叶片接种bsmv:γ+0.5mmcacl2(ph4.3),(2)叶片接种bsmv:γ+0.5mmcacl2+50μmalcl3(ph4.3),(3)叶片接种bsmv:hvhox9+0.5mmcacl2(ph4.3),(4)叶片接种bsmv:hvhox9+0.5mmcacl2+50μmalcl3(ph4.3)。处理36小时后,结果显示,与模拟接种bsmv:γ的植株相比,接种bsmv:hvhox9的植株铝处理后根部细胞汁液的铝含量无变化(图5(c)),但是细胞壁的铝含量显著提高(图5(d))。这些结果表明,高表达hvhox9能降低根系细胞壁铝含量,从而提高植株耐铝性。
h+离子流和根际ph的变化。从接种bsmv:γ和接种bsmv:hvhox9的植株根尖切下大约5cm的根段,测定根部伸长区的离子流。根被固定后,浸泡在处理液中分别0、3、6、12或24h后,利用mife系统(universityoftasmania,hobart,australia)(shabala等,1997)测定离子流,记录时间为10min。电极制作时,硼硅酸盐毛细血管(gc150-10;clarkelectrochemicalinstruments)用电极拉出器拉出,拉好的电极在微波炉中220℃过夜后,加入100-200μl三丁基氯硅烷(catalogno.90796;fluka)使电极硅烷化,硅烷化15min后,关闭微波炉,待冷却后,电极填入back-filling溶液(h+:15mmnacl+40mmkh2po4)。电极的尖端填入相应的离子选择剂(h+95297,switzerlandsigma,buchs,switzerland)(shabala等,2006),制作好的电极保存在bsm中,当电极稳定时校准电极,只有相关系数大于0.999时电极才能被使用并用于后续的测定实验。测定时电极尖端离根大约40μm,由微距控制器(patchmannp2,eppendorf,hamburg,germany)进行位置调节,振幅为每6s移动80μm。保存数据后,利用mflux软件计算测定根部附近的净离子流(newman,2001)。结果显示,在铝胁迫下,沉默植株的h+内吸明显大于模拟接种bsmv:γ植株,且根际ph明显低于模拟接种bsmv:γ植株(图6)。这些结果表明,高表达hvhox9在铝胁迫下能使根际碱化,从而降低铝毒害。
5、rt-pcr验证hvhox9基因的表达量
参照实施例1第4部分的步骤进行rt-pcr检测。
如图2(d)所示,对于接种bsmv:hvhox9的植株来说,无论是在对照还是铝处理条件下,与模拟接种bsmv:γ植株相比,根系中hvhox9基因的表达量分别被抑制了62%和35%。
综上所述,通过对大麦hvhox9的克隆和分析,并结合bsmv-vigs技术在青藏高原一年生野生大麦xz16上对该基因进行功能验证发现,hvhox9参与调控铝胁迫下xz16根系再生长,并降低细胞壁铝含量,对xz16的耐铝性发挥重要作用。
序列表
<110>浙江大学
<120>大麦hvhox9基因及其用途
<160>14
<170>siposequencelisting1.0
<210>1
<211>2523
<212>dna
<213>青藏高原一年生野生大麦xz16(hordeumvulgarel.ssp.spontaneum)
<400>1
atggcggcggcggtggcggcgatgcgctgcgggagcgggagcggcagtgacggcggcggc60
ggcggctacgacaagggcgggatggactcgggcaagtacgtgcggtacacgccggagcag120
gtggaggcgctggagcgggtgtatgccgagtgccccaagccgacatccacgcgcaggcag180
cagctgctccgcgagtgccccattctgtccaacatcgagcccaggcagatcaaggtctgg240
ttccagaatcgaaggtgccgtgataagcagcggaaggagtcttccaggcttcaggccgtg300
aacagaaaactgagcgccatgaacaagcttctcatggaagagaatgaacgtctccagaag360
caggtttcccagttggttcatgagaatgcatacatgaagcagcagctgcagaatccttcg420
ttggccaatgatacaagttgtgagtcaaatgtgaccactcctccaaaccctctaagggat480
gcaagtaacccagctggactacttgcaattgcggaagagacgttgacagagttcctctcg540
aaggctacaggaactgctgttgattgggtcccgatgcctgggatgaagcctggtccggat600
tcgtttggtattgttgccatttcacatggttgccgaggtgtcgctgcccgtgcctgtggt660
ctggtgaatctagaaccaacaaagattgtggagatcttaaaagaccgcccatcttggttc720
cgtgattgtcggagtcttgaagtttttacaatgcttccagctggaaacggcgggactatt780
gaacttgtttacatgcagatgtatgctcctaccactttagttcccgcacgtgatttttgg840
acgctgagatacacaacgacaatggaagatggaagtctcgtggtttgtgagagatctttg900
agtggttcaggaggtggtccaagtaccgcctcagcacagcaatttgtaagggctgagatg960
cttcctagtggctatttagttcggccatgcgatggtgggggttcaatcgtgcatatagtg1020
gatcatctggaccttgaggcttggagtgtaccagaagtgcttcgcccgctctatgagtct1080
tctagggtagttgctcagaaaatgaccactgcggcattacgacacatcagacagattgct1140
caagaaactagtggggaggttgtatatgcgctggggaggcagcctgctgttctgcggaca1200
tttagtcagaggctgagtagaggatttaatgatgctataagtggcttcaatgatgatggt1260
tggtctgtaatggccggagatggcattgaagatgtgattatcgcttgcaactcaaagaag1320
attaggagcaataacactgctcccaatgcttttatagctcctggaggtgttatatgtgct1380
aaagcatcaatgttactgcagaatgttccaccagcagtactggttcgatttctaagggaa1440
catcggtctgaatgggctgattataactttgatgcatattcggcttcagcgctgaaatca1500
agttcatgctcacttcctgggttgcgccctatgagattttctgggagccagatcatcatg1560
ccacttgctcacacggtggagaatgaggagattctagaagttgttcgtcttgaagggcaa1620
gcgctcgatgagggtcttttatcaagagatatccacctgcttcagttttgcactggaata1680
gacgagaaatcaatggggtcctgcttccaacttgtctttgccccaattgatgagcttttc1740
cctgatgatgctccattaatatcttcaggcttccgtgttataccactggacatgaaaaca1800
gatggtgcacccaccggtagaacactagatttggcctctagccttgaagctggttcaact1860
acactgcaagcctcaggcaatgcggacgattgtaatctacgatcagtgctgacaattgcc1920
tttcaattcccttacgagatgcatctccaagatagtgttgcaaccatggcccggcagtat1980
gtccgcagcattgtctctgccgttcagagagtgtcgatggctatctctccctctcgatct2040
ggtttgaatgctgaacagaagataatttctggcttccctgaagctgcaacacttgctcgc2100
tggatatgccaaagctaccggttccatctgggggtcgagttatttagacaggcagatgaa2160
gctggggaatctttattgagaatgctctgggatcatgaagatgctattttgtgctgttct2220
ttcaaggaaaagcctgtatttacgtttgcaaacgagatggggattaacatgttggaaaca2280
tctttcgttgcccttcaagacctctcgttggacaagatatttgatgaagctggtagaaag2340
gcactatactctgagatcccaaagttgatggagcagggctttgtttacctgccaggtggt2400
gtgtgcttgtctgggatgggccgccatgtctccttcgagaatgctatagcatggaaagta2460
gtcggtgaggacaacaatgtgcactgccttgccttctgcttcgtcaactggtctttcgtg2520
tga2523
<210>2
<211>840
<212>prt
<213>青藏高原一年生野生大麦xz16(hordeumvulgarel.ssp.spontaneum)
<400>2
metalaalaalavalalaalametargcysglyserglyserglyser
151015
aspglyglyglyglyglytyrasplysglyglymetaspserglylys
202530
tyrvalargtyrthrprogluglnvalglualaleugluargvaltyr
354045
alaglucysprolysprothrserthrargargglnglnleuleuarg
505560
glucysproileleuserasnilegluproargglnilelysvaltrp
65707580
pheglnasnargargcysargasplysglnarglysgluserserarg
859095
leuglnalavalasnarglysleuseralametasnlysleuleumet
100105110
glugluasngluargleuglnlysglnvalserglnleuvalhisglu
115120125
asnalatyrmetlysglnglnleuglnasnproserleualaasnasp
130135140
thrsercysgluserasnvalthrthrproproasnproleuargasp
145150155160
alaserasnproalaglyleuleualailealaglugluthrleuthr
165170175
glupheleuserlysalathrglythralavalasptrpvalpromet
180185190
proglymetlysproglyproaspserpheglyilevalalaileser
195200205
hisglycysargglyvalalaalaargalacysglyleuvalasnleu
210215220
gluprothrlysilevalgluileleulysaspargprosertrpphe
225230235240
argaspcysargserleugluvalphethrmetleuproalaglyasn
245250255
glyglythrilegluleuvaltyrmetglnmettyralaprothrthr
260265270
leuvalproalaargaspphetrpthrleuargtyrthrthrthrmet
275280285
gluaspglyserleuvalvalcysgluargserleuserglysergly
290295300
glyglyproserthralaseralaglnglnphevalargalaglumet
305310315320
leuproserglytyrleuvalargprocysaspglyglyglyserile
325330335
valhisilevalasphisleuaspleuglualatrpservalproglu
340345350
valleuargproleutyrgluserserargvalvalalaglnlysmet
355360365
thrthralaalaleuarghisileargglnilealaglngluthrser
370375380
glygluvalvaltyralaleuglyargglnproalavalleuargthr
385390395400
pheserglnargleuserargglypheasnaspalaileserglyphe
405410415
asnaspaspglytrpservalmetalaglyaspglyilegluaspval
420425430
ileilealacysasnserlyslysileargserasnasnthralapro
435440445
asnalapheilealaproglyglyvalilecysalalysalasermet
450455460
leuleuglnasnvalproproalavalleuvalargpheleuargglu
465470475480
hisargserglutrpalaasptyrasnpheaspalatyrseralaser
485490495
alaleulyssersersercysserleuproglyleuargprometarg
500505510
pheserglyserglnileilemetproleualahisthrvalgluasn
515520525
glugluileleugluvalvalargleugluglyglnalaleuaspglu
530535540
glyleuleuserargaspilehisleuleuglnphecysthrglyile
545550555560
aspglulyssermetglysercyspheglnleuvalphealaproile
565570575
aspgluleupheproaspaspalaproleuileserserglyphearg
580585590
valileproleuaspmetlysthraspglyalaprothrglyargthr
595600605
leuaspleualaserserleuglualaglyserthrthrleuglnala
610615620
serglyasnalaaspaspcysasnleuargservalleuthrileala
625630635640
pheglnpheprotyrglumethisleuglnaspservalalathrmet
645650655
alaargglntyrvalargserilevalseralavalglnargvalser
660665670
metalaileserproserargserglyleuasnalagluglnlysile
675680685
ileserglypheproglualaalathrleualaargtrpilecysgln
690695700
sertyrargphehisleuglyvalgluleupheargglnalaaspglu
705710715720
alaglygluserleuleuargmetleutrpasphisgluaspalaile
725730735
leucyscysserphelysglulysprovalphethrphealaasnglu
740745750
metglyileasnmetleugluthrserphevalalaleuglnaspleu
755760765
serleuasplysilepheaspglualaglyarglysalaleutyrser
770775780
gluileprolysleumetgluglnglyphevaltyrleuproglygly
785790795800
valcysleuserglymetglyarghisvalserphegluasnalaile
805810815
alatrplysvalvalglygluaspasnasnvalhiscysleualaphe
820825830
cysphevalasntrpserpheval
835840
<210>3
<211>281
<212>dna
<213>青藏高原一年生野生大麦xz16(hordeumvulgarel.ssp.spontaneum)
<400>3
ctgctgttctgcggacatttagtcagaggctgagtagaggatttaatgatgctataagtg60
gcttcaatgatgatggttggtctgtaatggccggagatggcattgaagatgtgattatcg120
cttgcaactcaaagaagattaggagcaataacactgctcccaatgcttttatagctcctg180
gaggtgttatatgtgctaaagcatcaatgttactgcagaatgttccaccagcagtactgg240
ttcgatttctaagggaacatcggtctgaatgggctgattat281
<210>4
<211>23
<212>dna
<213>人工序列(artificialsequence)
<400>4
atggcggcggcggtggcggcgat23
<210>5
<211>22
<212>dna
<213>人工序列(artificialsequence)
<400>5
tcacacgaaagaccagttgacg22
<210>6
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>6
atatgtcgacatggcggcggcggtggcggcgat33
<210>7
<211>33
<212>dna
<213>人工序列(artificialsequence)
<400>7
ctcagtcgaccacgaaagaccagttgacgaagc33
<210>8
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>8
gatggtgggggttcaatcgt20
<210>9
<211>20
<212>dna
<213>人工序列(artificialsequence)
<400>9
tccccagcgcatatacaacc20
<210>10
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>10
tggctgacggtgaggaca18
<210>11
<211>18
<212>dna
<213>人工序列(artificialsequence)
<400>11
cgagggcgaccaactatg18
<210>12
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>12
gtacgctagcctgctgttctgcggacattt30
<210>13
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>13
gtacgctagcataatcagcccattcagacc30
<210>14
<211>21
<212>dna
<213>人工序列(artificialsequence)
<400>14
caactgccaatcgtgagtagg21
- 还没有人留言评论。精彩留言会获得点赞!