ILT4在肿瘤诊治中的应用的制作方法
本发明属于生物医药和分子生物学技术领域,具体涉及ilt4在肿瘤诊治中的应用。
背景技术:
本发明背景技术中公开的信息仅仅旨在增加对本发明的总体背景的理解,而不必然被视为承认或以任何形式暗示该信息构成已经成为本领域一般技术人员所公知的现有技术。
免疫球蛋白样转录子4(ilt4),也称为淋巴细胞免疫球蛋白样受体b2(lilrb2)、单核巨噬细胞免疫球蛋白样受体10(mir10)或cd85d,属于免疫球蛋白样转录子(ilts)家族。该家族包括6个活化型受体成员及5个抑制性受体成员,分别以跨膜区可交链免疫受体酪氨酸活化基序(itam)的精氨酸残基及胞内区免疫受体酪氨酸抑制基序(itim)为特征,向细胞内传递活化或抑制信号,调控免疫细胞的功能。ilt4是典型的ⅰ型跨膜蛋白,由587个氨基酸组成。其胞外段含有四个免疫球蛋白样结构域,胞内含有三个itims。生理条件下,ilt4主要表达于单核细胞、巨噬细胞、树突状细胞(dcs)和中性粒细胞等固有免疫细胞中。当ilt4与相应的受体结合时,细胞内的itim可以招募含sh-2结构域的shp-1或shp-2磷酸酶,抑制上述细胞中活化信号的传递及免疫功能。此外,ilt4也表达于造血干细胞(hscs)、破骨前体细胞、血小板及神经元等细胞中,能够促进hsc扩增、抑制破骨细胞发生、抑制血小板聚集及神经轴突再生[25]。ilt4在小鼠中的同源蛋白为配对的免疫球蛋白样受体-b(pir-b),其胞外段含有六个免疫球蛋白样结构域,胞内含有四个itims。pir-b主要表达于b细胞、dcs、粒细胞及巨噬细胞等免疫细胞中,抑制上述细胞的活化与免疫功能。
近年来,ilt4在肿瘤微环境中的表达越来越引起人们的重视。目前大量研究证实,ilt4在多种血液及实体肿瘤的肿瘤细胞中高表达,与患者的疾病进展及不良生存相关,是患者的阴性预后指标。此外,ilt4/pir-b在间质的肿瘤相关巨噬细胞(tams)及髓源性抑制细胞(mdscs)高表达,促使其向免疫抑制的m2表型极化,诱导了调节性t细胞(treg)的分化,进而促进肿瘤免疫逃逸。但是肿瘤源性ilt4对肿瘤浸润t细胞命运及功能的直接调控作用尚不清楚。
技术实现要素:
针对现有技术中的不足,本发明的目的在于提供免疫球蛋白样转录子4(ilt4)在肿瘤诊治中的应用。本申请采用real-timepcr及流式细胞术检测人肿瘤细胞株及上皮细胞株中ilt4的表达,采用免疫组化检测nsclc及乳腺癌患者(肿瘤)组织中ilt4的表达,并统计分析其与患者临床病理特征和总生存(os)的相关性;通过geo数据库分析人nsclc、乳腺癌、前列腺癌和恶性黑色素瘤肿瘤组织中ilt4的表达与患者预后的相关性;综合论证ilt4高表达是恶性肿瘤的普遍特征,预示患者的不良预后。同时利用基因转染及敲除技术,分别上/下调ilt4低/高表达肿瘤细胞中的ilt4,通过肿瘤细胞与t细胞的共培养体系明确ilt4对t细胞老化的诱导作用,从而完成本发明。
本发明的第一个方面,提供检测ilt4基因及其表达产物表达水平的物质在制备检测、诊断或预测肿瘤进展试剂中的应用。
所述肿瘤包括但不限于肺鳞癌、肺腺癌、乳腺癌、恶性黑色素瘤及前列腺癌。
根据试验证明,ilt4表达量增高与肿瘤严重程度及不良预后有关,具体表现为正相关。
本发明的第二个方面,提供一种用于检测、诊断或预测肿瘤进展的组合物,其包含基于高通量测序方法和/或基于定量pcr方法和/或基于探针杂交方法检测ilt4基因及其表达产物表达情况的物质;或者基于免疫检测方法检测ilt4蛋白表达情况的物质。
优选采用northern杂交方法、mirna表达谱芯片、核酶保护分析技术、rake法、原位杂交检测ilt4基因表达(转录)情况;
免疫检测方法优选包括westernblot、elisa、胶体金试纸条和蛋白质芯片等;
其中,所述肿瘤包括但不限于肺鳞癌、肺腺癌、乳腺癌、恶性黑色素瘤及前列腺癌。
本发明的第三个方面,提供抑制ilt4基因及其表达产物和/或活性降低的物质在如下a)-d)至少一种中的应用:
a)抑制肿瘤细胞的增殖,或制备用于抑制肿瘤细胞增殖的产品;
b)抑制肿瘤细胞的粘附,或制备用于抑制肿瘤细胞粘附的产品;
c)抑制肿瘤细胞的侵袭转移,或制备用于抑制肿瘤细胞侵袭转移的产品;
d)抑制t细胞的老化;或者制备用于抑制t细胞的老化的产品。
其中,所述t细胞为效应t细胞,具体包括cd4+t细胞和cd8+t细胞。本发明通过研究证实,肿瘤源性ilt4可以诱导效应t细胞的老化,抑制其杀伤功能,进而促进多种实体肿瘤的免疫逃逸及发生发展。
所述肿瘤包括但不限于肺鳞癌、肺腺癌、乳腺癌、恶性黑色素瘤及前列腺癌。
本发明的第四个方面,提供一种用于治疗肿瘤的药物组合物,其包含抑制ilt4基因及其表达产物和/或活性降低的物质。
所述肿瘤包括但不限于肺鳞癌、肺腺癌、乳腺癌、恶性黑色素瘤及前列腺癌。
所述抑制ilt4基因及其表达产物和/或活性降低的物质,包括ilt4蛋白特异的抗体、针对ilt4mrna的rna干扰分子或反义寡核苷酸、小分子抑制剂、sirna,以及实施慢病毒感染或基因敲除的物质;进一步的,所述抗体为人抗体或鼠抗体。
进一步的,所述药物组合物还可以包含通常使用的适量载体、赋形剂及稀释剂。而且,根据通常的方法,可以制作成粉剂、颗粒剂、片剂、胶囊剂、混悬剂、乳剂、糖浆剂、喷雾剂等的口服剂、外用剂、栓剂及无菌注射溶液形式的剂型使用。可以包含的载体、赋形剂及稀释剂有乳糖、葡萄糖、蔗糖、山梨糖醇、甘露醇、木糖醇、赤藓糖醇、麦芽糖醇、淀粉、阿拉伯胶、藻酸盐、明胶、磷酸钙、硅酸钙、纤维素、甲基纤维素、微晶纤维素、聚乙烯吡咯烷酮、水、羟基苯甲酸甲酯、羟基苯甲酸丙酯、滑石粉、硬脂酸镁和矿物油等。
同时,经试验验证,上述技术方案对在小鼠中的ilt4的同源分子——配对免疫球蛋白样受体b(pir-b)而言同样有效。同时,在本发明的技术方案中,ilt4与pir-b为肿瘤源性。
本发明的有益技术效果:
本发明首次证实了ilt4在多种肿瘤细胞中高表达,与患者不良预后相关;体内外研究证实肿瘤源性ilt4诱导了cd4+及cd8+t细胞的老化,抑制了其杀伤活性,进而促进肿瘤进展。
同时证明了,肿瘤源性ilt4通过活化mapkerk1/2信号通路调控了肿瘤细胞脂肪酸的合成及脂滴形成,进而诱导t细胞的老化,因此通过调控ilt4表达,可用于治疗多种肿瘤,具有良好的实际应用之价值。
附图说明
构成本发明的一部分的说明书附图用来提供对本发明的进一步理解,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。
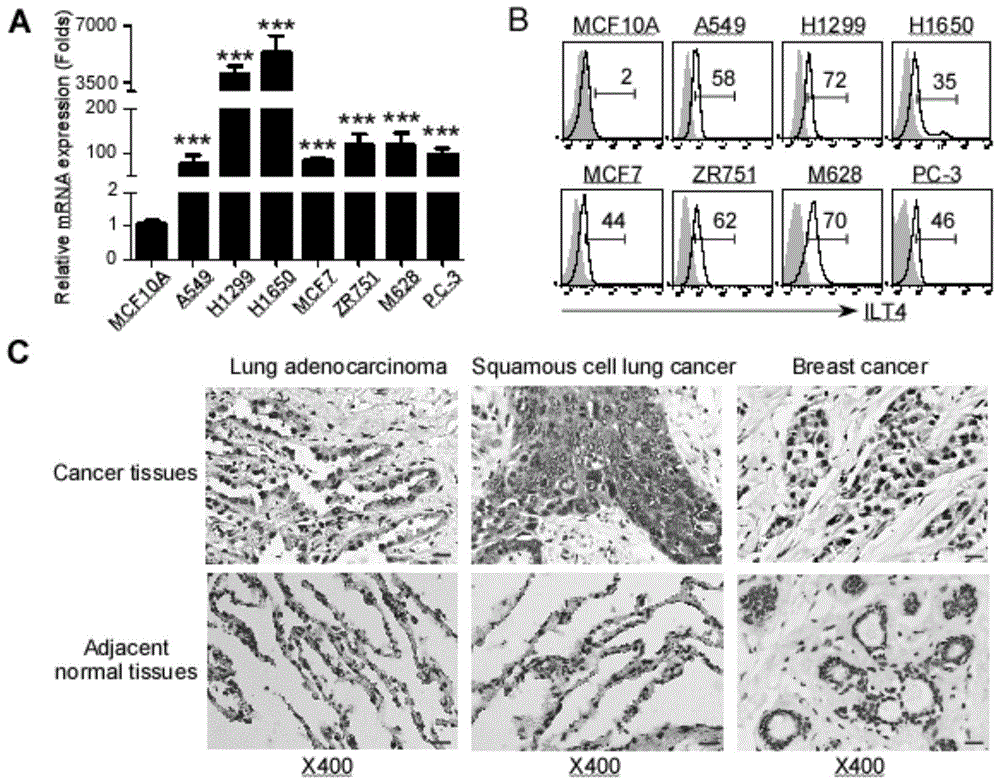
图1.ilt4在多种肿瘤的肿瘤细胞中高表达;a.real-timepcr结果显示,与正常乳腺上皮细胞mcf10a相比,ilt4在人肺癌(a549,h1299,h1650)、乳腺癌(mcf7,zr751)、恶性黑色素瘤(m628)、前列腺癌(pc-3)肿瘤细胞mrna水平高表达;b.流式细胞术分析结果显示,ilt4在上述肿瘤细胞中蛋白水平的表达显著高于正常乳腺上皮细胞;c.免疫组化结果显示,ilt4主要在肺鳞癌、肺腺癌及乳腺癌组织的肿瘤细胞中高表达,而在周围正常组织的上皮细胞中低表达。***,与上皮细胞株mcf10a中的表达水平相比较p<0.001。
图2.ilt4在肿瘤组织中高表达预示患者的不良预后;a.对临床nsclc及乳腺癌组织标本中ilt4的表达与患者总生存(os)数据的分析显示,在nsclc(n=116)及乳腺癌(n=84)中,ilt4低表达组患者的os显著优于ilt4高表达组,p值分别为0.0246及0.0493。b.geo数据库分析显示,ilt4高表达的nsclc(n=1906)、乳腺癌(n=159)及前列腺癌(n=281)患者中无进展生存(pfs)及os明显均差于ilt4低表达组。其中nsclc的pfs差异p=0.033,nsclc的os差异p=0.0077;乳腺癌的pfs差异p=0.007,乳腺癌的os差异p=0.012;前列腺癌os差异p=0.034。
图3.肿瘤细胞诱导了t细胞的老化;其中a.sa-β-gal染色显示,与正常上皮细胞mcf10a相比,nsclc(a549、h1299、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)肿瘤细胞明显诱导了t细胞的老化,表现为t细胞中蓝黑色颗粒的增加。人初始cd4+及cd8+t细胞由正常人pbmc中磁珠分选获得,并在2μg/ml的okt-3预包被的12孔板中活化。之后按1:1(肿瘤细胞:t细胞)的比例构建上述肿瘤细胞与cd4+及cd8+t细胞的共培养体系,共培养24小时后分离t细胞,单独培养72小时;采用sa-β-gal染色检测了老化t细胞的比例。b,c.与正常上皮细胞mcf10a相比,肿瘤细胞显著诱导了sa-β-gal阳性cd4+及cd8+t细胞数量的增加;d,e,f,g.流式细胞术结果显示,与正常上皮细胞mcf10a相比,nsclc(a549、h1299、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)细胞诱导了cd4+(d,e)及cd8+t(f,g)细胞中共刺激分子cd27/cd28的丢失及细胞周期抑制蛋白p53/p21表达上调。比例尺为10μm;***,与上皮细胞株mcf10a组相比较p<0.001。
图4.肿瘤源性ilt4介导了肿瘤细胞对t细胞老化的诱导;a.与同型对照组比较,肿瘤-t细胞共培养体系加入anti-ilt4显著逆转了t细胞中sa-β-gal阳性染色的细胞数;在肿瘤细胞-t细胞共培养体系中加入500ng/ml的ilt4中和抗体(anti-ilt4)或同型对照抗体(isotype),共培养24小时后分离cd4+及cd8+t细胞再单独培养72小时,sa-β-gal染色检测老化t细胞的比例;b,c.anti-ilt4逆转了nsclc(a549、h1299、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)诱导的cd4+及cd8+t细胞老化的统计图;d.肿瘤-t细胞共培养体系加入anti-ilt4(500ng/ml)部分逆转了nsclc(a549、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)诱导的t细胞中共刺激分子cd27/28的下调;e.分别用500ng/mlanti-ilt4预处理肿瘤细胞或t细胞2小时,然后洗去培养基中的anti-ilt4,再进行肿瘤细胞-t细胞共培养实验,结果显示anti-ilt4预处理肿瘤细胞而非t细胞显著逆转了cd4+t细胞的老化;f,g,h,i.选择ilt4相对低(h1650、mcf7、m628、pc-9)/高(a549、h1299、zr751、m628)表达的肿瘤细胞系,利用慢病毒转染分别过表达/敲除肿瘤细胞ilt4,转染48小时后再次进行了肿瘤-t细胞共培养,结果老化cd4+(f,g)及cd8+t(h,i)细胞的数量被相应上/下调。比例尺为10μm;*,p<0.05;**,p<0.01;***,p<0.001。
图5.pir-b在小鼠肿瘤细胞高表达;其中,pir-b(小鼠中ilt4同源分子)基因(a)及蛋白(b)在小鼠肺癌(mle12与ll/2)、乳腺癌(e0771)及恶性黑色素瘤(b16f0)细胞中的表达均显著高于正常上皮细胞株fsk4。***,与fsk4相比p<0.001。
图6.pir-b促进小鼠肿瘤细胞的恶性生物学行为;其中,a.细胞生长曲线显示,小鼠乳腺癌细胞e0771及恶性黑色素瘤细胞b16f0中pir-b过表达/敲除显著促进/抑制了肿瘤细胞的增殖;b.粘附实验结果显示,e0771及b16f0中pir-b过表达/敲除显著促进/抑制了肿瘤细胞的粘附能力;c.划痕实验证实e0771及b16f0中pir-b过表达/敲除显著促进/抑制了肿瘤细胞的侵袭移行能力。划痕图片分别在培养的0及12小时获得。与各自的对照组相比,*,p<0.05;**,p<0.01;***,p<0.001。
图7.免疫系统参与了pir-b调控的移植瘤进展;其中,a,b,c.c57bl/6j小鼠移植瘤模型中乳腺癌细胞过表达/敲除pir-b显著促进/抑制了小鼠体内肿瘤的生长;6-8周大小的雌性c57bl/6j小鼠皮下注射pir-b过表达/基因敲除的小鼠乳腺癌细胞e0771(2×105细胞/只)。每3天测量肿瘤长、短径评估大小。24天后当肿瘤最大径生长至2cm左右,co2处死小鼠,分离肿瘤组织并称重。d,e,f.nsg小鼠体内e0771移植瘤模型中pir-b的过表达/敲除同样促进/抑制了小鼠体内肿瘤的生长,但变化趋势明显小于全基因c57bl/6j小鼠,提示免疫系统参与了pir-b诱导的肿瘤发生发展。6-8周大小的雌性nsg小鼠中皮下注射pir-b过表达/基因敲除的乳腺癌细胞e0771(2×105细胞/只)。每3天测量肿瘤长、短径评估大小。24天后当肿瘤最大径生长至2cm左右,co2处死小鼠,分离肿瘤组织并称重。与各自的对照组相比,**,p<0.01;***,p<0.001。
图8.pir-b体内诱导了效应t细胞老化;其中,a.c57bl/6j小鼠移植瘤模型中pir-b过表达/敲除显著上/下调了外周血(bl)与肿瘤组织中浸润性t细胞(til)中sa-β-gal阳性淋巴细胞的数量;b,c.对肿瘤组织中tils数量及亚群分析显示:pir-b对tils浸润数量无显著影响(b),但是pir-b敲除增加了tils中cd8+t细胞的浸润比例(c)。d.分离不同组肿瘤组织中浸润的淋巴细胞进行流式分析显示pir-b过表达/敲除分别下/上调了浸润性cd4+/cd8+t细胞的免疫功能,表现为cd4+t细胞中ifn-γ减少/增加,cd8+t细胞中ifn-γ、granzymeb与perforin的减少/增加;d为典型流式分析结果;e.不同处理组中cd4+ifn-γ+t细胞、cd8+ifn-γ+t细胞、cd8+granzymeb+t细胞与cd8+perforin+t细胞占肿瘤浸润淋巴细胞的比例;e为d的统计结果。与lv-ctr-shrna相比,*,p<0.05;**,p<0.01;***,p<0.001。
具体实施方式
应该指出,以下详细说明都是例示性的,旨在对本申请提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本申请所属技术领域的普通技术人员通常理解的相同含义。
需要注意的是,这里所使用的术语仅是为了描述具体实施方式,而非意图限制根据本申请的示例性实施方式。如在这里所使用的,除非上下文另外明确指出,否则单数形式也意图包括复数形式,此外,还应当理解的是,当在本说明书中使用术语“包含”和/或“包括”时,其指明存在特征、步骤、操作、器件、组件和/或它们的组合。
现结合具体实例对本发明作进一步的说明,以下实例仅是为了解释本发明,并不对其内容进行限定。如果实施例中未注明的实验具体条件,通常按照常规条件,或按照试剂公司所推荐的条件;下述实施例中所用的试剂、耗材等,如无特殊说明,均可从商业途径得到。
以下通过实施例对本发明做进一步解释说明,但不构成对本发明的限制。应理解这些实施例仅用于说明本发明而不用于限制本发明的范围。下列实施例中为注明具体条件的试验方法,通常按照常规条件进行。
实施例1
1.细胞培养相关实验
1.1细胞培养
人细胞系a549、h1299、h1650、mcf7、zr751、m628、pc-3、293t及小鼠细胞系e0771、b16f0用含10%fbs及1%青链霉素双抗的rpmi-1640或dmem培养;小鼠肺癌细胞系mle12采用含2%胎牛血清、0.005mg/ml胰岛素、its(1:100)、10nm氢化可的松、10nmβ-雌二醇、2mml-谷氨酰胺的dmem培养;小鼠肺癌细胞系ll/2采用含10%fbs、2%l-谷氨酰胺及1%青链霉素双抗的高糖dmem培养;mcf10a采用含2%胎牛血清、1μg/ml氢化可的松、20ng/ml表皮生长因子及1.2μg/ml胰岛素的dmem培养基培养;fsk4采用含2%fbs、1%hepes溶液、10μg/ml胰岛素及5ng/ml表皮生长因子的dmem培养。
健康人pbmc采用ficoll-paque细胞分离液分离,人cd4+及cd8+t细胞采用相应磁珠细胞分选试剂盒从pbmc中分选,流式细胞术确认细胞分选纯度>97%。人t细胞用2μg/mlokt-3活化24小时,后采用含10%人ab血清、50iu/ml重组人il-2、2-me(1:1000)的rpmi-1640培养。
1.2细胞复苏
(1)将细胞从液氮罐取出,置于干冰上转运;
(2)将冻存管放入37℃水浴箱,快速晃动冻存管,尽量使细胞在1分钟内融化;
(3)将融化的细胞快速移入10ml培养液(如果是pbmc用25ml培养液)中,上下颠倒混匀细胞悬液;
(4)离心,1500rpm,5-10分钟;
(5)去上清,用新鲜培养基将细胞重悬于培养瓶中,晃动均匀,37℃恒温静置培养。每日观察细胞生长状态。
1.3细胞传代
(1)镜下观察细胞,当细胞融合度达到80-90%时给予传代;
(2)弃掉培养基,pbs洗一次,注意不要用力过大,以免冲起贴壁不牢的细胞;
(3)加入1-2ml的胰酶-edta消化液,轻晃使胰酶分布均匀,37℃消化2-10min左右,镜下观察,待细胞收缩变圆时立即加入培养基终止消化,轻轻吹打成单细胞悬液;
(4)离心,1500rpm×5-10分钟,弃掉上清;
(5)新鲜培养液重悬细胞,按1:1-1:4比例传代,放回37℃细胞培养箱内培养。
1.4细胞冻存
(1)首先将细胞冻存盒从-80℃冰箱中取出,恢复至室温;
(2)取生长状态良好的细胞,弃培养液;
(3)pbs洗一次;
(4)对于贴壁细胞,加入胰酶1-2ml消化,待细胞变圆后加入培养液终止消化,轻轻将细胞吹下来;悬浮细胞可省略此步骤;
(5)离心,1500rpm×5-10分钟,弃上清;
(6)1ml冻存液重悬细胞,肿瘤细胞加入含50%fbs的冻存液,t细胞加含90%fbs的冻存液;
(7)在冻存管上标明细胞的名称,冻存时间及操作者姓名;
(8)将细胞放入冻存盒,即刻置于-80℃冰箱,4小时后放入液氮保存。
1.5细胞计数
(1)清洗细胞计数板和盖玻片,并擦干备用;
(2)准备单细胞悬液,取30μl细胞液与30μl台盼蓝,在ep管内混合均匀,必要时pbs稀释细胞,使最终细胞计数量在100-200之间,以增加计数准确性;
(3)从计数板盖玻片一侧缓慢加入上述细胞悬液,使计数板和盖玻片间充满液体,注意盖片下不可产生气泡,也不要溢入两侧凹槽中,水平静置30s;
(4)镜下计数左上、右上、左下和右下4个大方格内的细胞总数,对于压线细胞,计数左侧及上方细胞数;
(5)最终细胞数=(4个大格细胞总数/4)×2×稀释倍数×104个/ml。
1.6肿瘤细胞与t细胞的共培养
(1)用500μl/孔2μg/ml的人okt3单抗包被24孔板4℃过夜;
(2)t细胞活化:去除包被液后将24孔板用pbs洗涤一次,用含10%fbs的1640将1-1.5×106的cd4+/cd8+t细胞重悬种入24孔板,37℃培养箱孵育24小时;
(3)将不同处理的肿瘤细胞接种于12或6孔板中,使细胞共培养之前的融合度达70%左右;
(4)轻轻洗涤肿瘤细胞,尽量去除悬浮的肿瘤细胞;
(5)计数t细胞数量,用含10%fbs的rpmi-1640将肿瘤细胞与t细胞按1:1的比例共培养18-24小时;
(6)轻轻晃动细胞,使t细胞悬浮于培养上清中,但尽量不要晃起肿瘤细胞,吸出细胞上清;
(7)1000rpm离心5-10分钟分离t细胞,后将t细胞继续置于含10%人ab血清、50iu/ml重组人il-2及2-me(1:1000)的rpmi-1640中培养72小时,用于后续实验。
2.质粒扩增与制备
2.1质粒构建
本实验所用ilt4及pir-b过表达与对照质粒购自美国genecopoeia公司;ilt4及pir-bshrna及对照质粒购自sigma-aldrich公司,慢病毒包装质粒pspax2及pmd2.g购自origene。
2.2质粒转化及扩增
(1)将感受态细菌从-80℃取出,并在冰上融化(大约20-30分钟);
(2)从4℃冷库取出提前制备好的琼脂糖凝胶板,恢复至室温或37℃;
(3)在ep管中将质粒原液1-5μl加入感受态细菌(20-50μl)中,轻弹混匀;
(4)将上述混合物冰上静置20-30min;
(5)将ep管下端1/2-2/3放入42℃水浴进行热休克处理30-60s,随后继续冰上静置2min;
(6)加入250-500μl的预冷的lb液体培养基,轻轻混匀,放在恒温摇床(200rpm),37℃摇晃45分钟;
(7)将部分或全部的转化物加入含有相应抗生素的10cm凝胶板中,置入37℃恒温孵箱培养,胶板向上培养1小时,后倒置培养过夜(16小时);
(8)待琼脂糖凝胶板中长出细菌菌落时,挑取单克隆菌落置于5ml含相应抗生素的lb液体培养基中,37℃恒温摇床,250rpm摇晃过夜。
2.3质粒提取
(1)2ml离心管中加入上述过夜的大肠杆菌培养基,离心≥12000g×1分钟;弃上清,颠倒ep管在纸上,去掉所有水痕;必要时重复上面步骤;
(2)重悬:加入250μl含有rna酶的重悬缓冲液,上下颠倒重悬细胞;
(3)溶解:加入250μl细胞裂解缓冲液,上下颠倒4-6次以彻底混匀(注意:这一步不能超过5分钟;不要旋转,以防剪短基因组dna);
(4)中和:加入350μl中和缓冲液,立即上下颠倒4-6次混匀,离心≥12000g×10分钟;
(5)纯化柱准备:将核酸纯化柱中加入500μl准备液,离心≥12000g×1分钟,弃去收集管中的液体;
(6)结合:将4中的上清转移到核酸纯化柱中,离心≥12000g×1分钟;去掉收集管中的液体,将核酸纯化柱放回收集管中;
(7)洗涤:核酸纯化柱中加入500μl洗涤缓冲液,离心≥12000g×1分钟,去掉收集管中的液体,将核酸纯化柱放回收集管中;
(8)晾干:再次离心≥12000g×1分钟,后将核酸纯化柱转移至干净的1.5ml的ep管中;
(9)洗提:核酸纯化柱中加入50μl的溶解缓冲液,室温放置1分钟。
(10)离心≥12000g×1分钟提洗dna(也可以将dna溶于30μl的溶解缓冲液中以增加质粒dna浓度)。
3.慢病毒包装与细胞感染
3.1慢病毒包装
(1)将1×106的293t细胞种在10cm平板中,使转染时细胞融合度达70%-80%;
(2)质粒转染前将细胞培养液更换为9ml含10%fbs的rpmi-1640;
(3)制备转染混合物:
a)ilt4/pir-bshrna慢病毒的转染混合物:1.2mlopti-mem+12μlx-tremegenetmhpdna转染试剂+6μg对照/ilt4/pir-bshrna+4μgpspax2+2μgpmd2.g质粒,混匀后室温放置25分钟;
b)ilt4/pir-b过表达慢病毒的转染混合物:ep管中加入200μlopti-mem稀释2.5μg对照/ilt4/pir-b过表达质粒+5μllenti-pachivmix(0.5g/l),另外准备一个ep管用200μlopti-mem稀释15μl的endofectinlenti,将后者逐滴加入dna溶液后轻轻混匀,室温放置10-25分钟;
(4)将转染混合物轻轻逐滴加入细胞培养液中,轻晃混匀,37℃培养箱恒温培养293t细胞;
(5)8小时后去掉上述细胞培养液,更换为含5%fbs的rpmi-1640培养液+200μltiterboost;
(6)分别于细胞培养的48小时及72小时收集293t上清液,1000g离心10分钟去掉细胞碎片,分装病毒上清并快速置入-80℃冰箱冷冻备用。
3.2细胞慢病毒感染
(1)用含10%fbs的rpmi-1640或dmem培养待转染的肿瘤细胞,使慢病毒感染前肿瘤细胞融合度为30%左右;
(2)用含5%fbs的rpmi-1640或dmem制备慢病毒混合物,根据不同待感染细胞株分别将慢病毒稀释10-100倍,并加入终浓度为5ng/mlpolybrene;
(3)24孔板中加入500μl/孔的慢病毒混合物;
(4)4℃离心,1000g×60分钟,后将细胞放回37℃细胞培养孵箱培养;
(5)8小时后更换培养液为含5%fbs的rpmi-1640或dmem;
(6)感染48h-72h后收集细胞准备下一步实验。
4.rt-pcr
4.1总rna提取
(1)匀浆处理:
a)组织:肿瘤组织称重并剪成50-100mg大小,干冰上快速冷却后置入-80℃冰冻备用;研磨前用rnasezap清洗研磨器1次,并用dh2o洗3-5次;组织中加入1mltrizol,用研磨棒磨碎组织至匀浆状态;
b)单层培养细胞:弃掉细胞培养基,pbs清洗细胞2遍,直接在培养板中加入trizol裂解细胞,按10cm2/ml加入trizol,使用移液器吹打使细胞充分裂解;
(2)匀浆样品静室温放置5min,使核酸蛋白复合物充分分离;
(3)每ml样品中加入0.2ml氯仿,振荡15s,室温静置2min;
(4)4℃离心,12000g×15min,取上清至新的ep管中;
(5)加入0.5ml异丙醇,将管中液体轻轻混匀,室温静置10min;
(6)4℃离心,12000g×10min,弃上清;
(7)每个ep管中加入1ml75%乙醇,轻轻洗涤沉淀。4℃离心,7500g×5min,弃上清(尽量将管内残留液体吸净);
(8)待rna晾干后,加入适量的去离子水溶解;
(9)使用nanodrop2000检测rna浓度及质量,-80℃保存。
4.2逆转录反应
(1)将高容量cdna逆转录试剂盒放置冰上提前融化;
(2)将2μg的mrna稀释至10μlnuclease-freeh2o中;
(3)参照如下比例配制2×rt混合物,总体系为20μl:
(4)准备cdna逆转录体系:每个反应管中加入上述准备好的10μlmrna及10μl2×rt混合物,轻轻混匀后快速离心避免损失液体,冰上转移至pcr仪;
(5)按照下列温度进行逆转录:
(6)取出cdna,-20℃保存备用。
4.3real-timepcr
(1)按照以下比例配置反应体系(10μl),每组设置3个重复孔:
(2)反应条件:
(3)基因表达量计算:2-δδct法计算基因表达差异。
5.流式细胞术
5.1肿瘤细胞表面分子ilt4、pir-b染色
(1)将待检测细胞系用胰酶消化后pbs洗2次,1500rpm离心5分钟,尽量去掉培养基;
(2)每个试验管中加入0.5-1×106个细胞,用含5%bsa的pbs100μl稀释一抗并重悬细胞,充分混匀(按1:100稀释);
(3)室温反应1小时;
(4)pbs洗涤2次,1500rpm离心5分钟;
(5)按推荐浓度将细胞重悬于荧光标记的二抗中,室温避光孵育30分钟;
(6)离心洗涤细胞2-3次;
(7)用0.5mlpbs重悬细胞,流式细胞仪检测。
5.2t细胞表面分子染色(荧光标记的一抗)
(1)每个试验管中加入0.5-1×106个待检测细胞,用含5%bsa的pbs100μl稀释荧光标记的一抗,充分混匀(抗体稀释浓度参照相应抗体说明书);
(2)冰上避光孵育30分钟;
(3)pbs洗涤2次,1500rpm离心5分钟;
(4)用0.5mlpbs重悬细胞,流式细胞仪检测。
5.3t细胞中胞浆分子p53、p21染色
(1)收集待检测t细胞,每个试验管中加入0.5-1×106个细胞,离心去上清,pbs洗涤两次;
(2)加入4%多聚甲醛37℃固定10分钟,冰上1分钟;
(3)加入90%的冰甲醇,冰上孵育30分钟破膜;
(4)pbs离心洗涤2-3次;
(5)加入含5%bsa的pbs稀释的一抗100μl(一抗按1:200稀释),重悬细胞并充分混匀,室温孵育1小时;
(6)pbs离心洗涤2-3次;
(7)加入100μlaf-488标记的抗兔二抗(1:1000稀释),室温避光孵育30分钟;
(8)pbs离心洗涤2-3次后用0.5mlpbs重悬,流式细胞仪检测荧光强度。
5.4t细胞中细胞因子染色
(1)将不同组织中的t细胞用pma、ionomycin及golgistop处理5小时后进行后续染色;
(2)参考6.2中表面分子染色方法,先进行t细胞表面cd3/cd4/cd8分子染色;
(3)用含0.5%bsa的pbs洗两遍,加入100μl/孔bdcytofix/cytoperm液,4℃20min;
(4)加入100μl/孔1×perm/washbuffer(用dh2o稀释10×perm/washbuffer),离心洗涤一次;
(5)加入200μl/孔1×perm/washbuffer再洗2次;
(6)用上述1×perm/washbuffer液稀释抗体(抗体稀释比例参照相应说明书),每孔50μl,4℃孵育30min;
(7)用200μl/孔1×perm/washbuffer洗涤2次;
(8)加入200μlpbs重悬,流式细胞仪检测。
5.5t细胞中转录因子foxp3染色
(1)参考6.2中表面分子染色方法,先进行t细胞表面cd3/cd4/cd8分子染色;
(2)用0.5%bsa的pbs洗两遍,加入1×fixsolution(用truenuclearfixdiluent稀释4×fixconcentration)每孔200μl,室温避光孵育1小时;
(3)离心去除fixsolution,加入200μl/孔的1×perm液(用dh2o稀释truenuclear10×perm),离心洗涤三次;
(4)用上述1×perm液稀释foxp3抗体(按1:100稀释),每孔50μl,室温避光孵育30min;
(5)用200μl/孔1×perm液离心液洗三次;
(6)加入pbs重悬,流式细胞仪检测。
6.老化相关β-半乳糖苷酶(sa-β-gal)染色
6.1染色步骤
(1)收获t细胞并pbs洗两次,1000rpm离心5分钟,去上清后pbs重悬,使细胞浓度为2-5×106/ml;
(2)取5μl细胞悬液,用枪头轻轻涂抹在在玻片上,注意动作轻柔,以免细胞破碎;
(3)等水分晾干后,4%多聚甲醛50μl固定3-5min;
(4)甩掉多聚甲醛,pbs轻轻洗涤2次;
(5)充分晾干载玻片后加入100μl显色液,37℃恒温箱避光孵育24小时;
(6)用水轻轻洗涤玻片,盖上盖玻片,400倍放大显微镜观察,细胞浆内蓝黑色颗粒为染色阳性细胞。
6.2sa-β-gal显色液的配制
7.肿瘤标本的免疫组化染色
(1)组织切片置入65℃恒温箱45min;
(2)二甲苯10min×2次进行脱蜡;
(3)采用100%、90%、80%及70%的梯度浓度酒精3min/浓度脱二甲苯并水化;
(4)edta缓冲液微波修复抗原20min,暴露抗原决定簇;
(5)3%过氧化氢室温孵育20min,灭活内源性过氧化物酶;
(6)山羊血清封闭非特异性抗原抗体反应;
(7)孵育一抗(affinityilt4抗体1:20稀释)4℃过夜;
(8)pbs洗涤3次,鼠兔通用型二抗(试剂a)37℃孵育30min;
(9)pbs洗涤3次,辣根酶标记链霉素卵白素(试剂b)37℃孵育30min;
(10)pbs洗涤3次,dab显色1-2min,流水冲洗终止显色;
(11)苏木素染核3min;
(12)盐酸酒精脱色并分化;
(13)70%、80%、90%、100%梯度浓度酒精脱水;
(14)二甲苯透明3min×2次;
(15)中性树胶封片,光学显微镜下观察并采集图像。
8.免疫组化评分标准
免疫组化染色结果由两个独立的病理学诊断医师在400倍镜下观察。ilt4染色的评估包括染色比例评估及染色强度分评估两部分。根据染色阳性的肿瘤细胞比例,比例得分可以分为0-3分:0分为肿瘤细胞无染色;1分为肿瘤细胞染色范围<25%;2分为肿瘤细胞染色范围处于25-75%;3分为肿瘤细胞染色范围>75%。根据肿瘤细胞中染色强度不同得分同样分为0-3分,其中0分为无染色,1分为肿瘤细胞染成淡黄色颗粒,2分为棕黄色颗粒,3分为褐色颗粒。总的免疫组化得分为比例得分与强度得分的乘积。根据最终得分将不同肿瘤组织分为ilt4高表达组(≥3分)及低表达组(<3分)。
9.geo数据库分析
我们利用km-plotter数据库的在线分析工具分析ilt4表达与nsclc患者生存的相关性。共有1926例及982例患者分别参与os及pfs的分析。截断值(cut-off)为系统自动选择的最佳值,其他参数为默认。分析结束系统在线生成hr及p值。我们从geo数据库选择了2个独立的包含患者临床病理参数及生存数据的mrna表达数据集(gse1456和gse16560)分别进行乳腺癌及前列腺癌患者的生存分析。gse1456依托gpl96及gpl97平台进行芯片检测,包含159例乳腺癌患者;gse16560依托gpl5474平台,包含281例前列腺癌患者。数据集矩阵文件数据从geo(https://www.ncbi.nlm.nih.gov/gds/)下载,采用r包“limma”进行数据标准化。我们首先提取并整合ilt4表达数据进行生存分析,然后利用kaplan-meier生存及log-rank(mantel-cox)检测验证ilt4表达与生存的相关性。不同组织中截断(cut-off)值由系统自行选择最佳值。
10.细胞生长试验
(1)将1-2×105的ilt4/pir-b过表达或基因敲除的肿瘤细胞接种于6孔板中,每组设置一个重复孔;
(2)每3天胰酶消化并计数肿瘤细胞数量,计数结束后根据细胞生长速度回种1/3-1/5的肿瘤细胞,计数细胞总数并绘出细胞生长曲线。
11.mtt增殖试验
(1)平底96孔板中每孔接种5000个不同处理的肿瘤细胞;
(2)接种24小时待细胞贴壁后加入15μl5mg/ml的mtt,37℃恒温培养4小时;
(3)吸出培养液,每孔加入15μldmso,37℃培养30分钟;
(4)采用酶标仪在570nm波长处检测每孔的吸光度,并计算细胞增殖速率。
12.粘附实验
(1)在平底96孔板中加入50μl/孔的纤维连接蛋白(10μg/ml),4℃包被过夜;
(2)去除纤维连接蛋白,用含2%bsa的pbs封闭孔底37℃两小时;
(3)去掉封闭液,将ilt4/pir-b过表达或基因敲除的肿瘤细胞按照1×105/100μlrpmi-1640接种于上述处理的96孔板中,37℃孵育45分钟;
(4)用pbs轻轻洗涤3次,去掉未粘附的细胞;
(5)4%多聚甲醛室温固定4分钟;
(6)0.03%结晶紫室温染色15分钟,pbs轻轻洗涤去掉染液,400×显微镜计数每个视野粘附肿瘤细胞的数量×10个视野,取平均值作为粘附细胞数量。
13.划痕实验
(1)将ilt4/pir-b过表达或基因敲除48小时后的肿瘤细胞按5×105/孔的数量接种于6孔板中;
(2)24小时后待细胞贴壁良好且融合度达90-100%,用枪头垂直划出宽约2mm的划痕;
(3)pbs洗去悬浮细胞,将细胞置于37℃孵箱培养,用不含血清的rpmi-1640培养肿瘤细胞,0小时、12小时、24小时镜下拍照观察划痕变化。
14.小鼠肿瘤生长模型
6-8周大小的c57bl/6j小鼠或nsg小鼠中皮下注射pir-b过表达/基因敲除的小鼠乳腺癌细胞e0771(2×105细胞/只)。每3天测量肿瘤长、短径评估大小。3-4周后当肿瘤最大径生长至2cm左右,co2处死小鼠,分离肿瘤组织并称重。
在全基因的c57bl/6j小鼠中,用ficoll-paque液及免疫磁珠分离小鼠血液、引流淋巴结、脾脏、肿瘤组织中的t细胞,sa-β-gal染色检测t细胞老化,流式细胞术检测t细胞亚群分布及功能。
15.统计分析
统计分析采用graphpadprism5软件完成。数据采用均数±sd表示。多组间比较采用单向anova方差分析,两组间比较采用student’st检验完成。如果样本量过小且不符合正态分布,采用非参t检验统计分析。p<0.05代表差异有统计学意义。
实验结果
1.ilt4在多种肿瘤细胞中高表达。
研究表明,ilt4在人非小细胞肺癌(nsclc)、乳腺癌及肝癌肿瘤细胞中高表达,且与患者较差的疾病分期及总生存期相关。为证实ilt4表达是恶性肿瘤的普遍标志,我们检测了ilt4在nsclc(a549、h1299、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)等细胞中的表达,结果显示与正常上皮细胞相比,ilt4基因及蛋白在所有上述肿瘤细胞中显著高表达(图1a,b)。此外,我们也采用免疫组化检测了ilt4在nsclc及乳腺癌患者肿瘤组织中的表达。结果发现,ilt4在上述肿瘤组织中的表达主要集中于肿瘤细胞,且其表达显著高于周围正常上皮组织(图1c)。
2.ilt4在肿瘤细胞的高表达预示患者的疾病进展及不良预后。
我们对临床nsclc及乳腺癌组织标本中ilt4的表达与患者的临床病理资料进行分析,结果发现,在nsclc中,ilt4的高表达与肿瘤低分化(p=0.026)、淋巴结转移(p=0.014)及较差的tnm分期(p=0.008)正相关(表1);在乳腺癌中,ilt4的高表达与肿瘤低分化(p=0.034)及淋巴结转移(p=0.024)正相关(表2)。对上述患者总生存(os)数据的分析显示,在nsclc及乳腺癌中,ilt4低表达组患者的os显著优于ilt4高表达组(图2a)。此外我们采用geo数据库收集并分析了1906例nsclc、159例乳腺癌及281例前列腺癌患者中ilt4表达的预后意义。与本课题组前期研究相一致,ilt4在上述肿瘤组织中的表达预示患者不良的pfs及os(图2b)。提示ilt4在肿瘤细胞中的表达代表了其恶性表型,驱动了肿瘤的进展,可作为恶性肿瘤患者不良的预后指标。
表1:nsclc组织中ilt4在肿瘤细胞的表达与患者临床病理参数的相关性(n=116)
表2:乳腺癌组织中ilt4在肿瘤细胞的表达与患者临床病理参数的相关性(n=84)
3.肿瘤细胞能够体外诱导cd4+及cd8+t细胞的老化。
前期研究证实,ilt4在nsclc细胞中的表达与肿瘤间质中减少的肿瘤浸润淋巴细胞(tils)相关,提示ilt4作为经典的免疫抑制性分子,也调控了t细胞的抗肿瘤免疫应答。t细胞老化是在微环境中介导肿瘤免疫逃逸的新机制。为探究ilt4是否参与了t细胞老化的诱导,我们首先确认了上述人肿瘤细胞对t细胞老化的诱导作用。人初始cd4+及cd8+t细胞由正常人pbmc中磁珠分选获得,并在2μg/ml的okt-3预包被的12孔板中活化。之后按1:1(肿瘤细胞:t细胞)的比例构建上述肿瘤细胞与cd4+或cd8+t细胞的共培养体系,共培养24小时后分离t细胞,单独培养72小时;采用sa-βgal染色检测了老化t细胞的比例,并采用流式细胞术检测老化t细胞的表型变化。结果发现:nsclc(a549、h1299、h1650)、乳腺癌(mcf7、zr751)、恶性黑色素瘤(m628)及前列腺癌(pc-3)肿瘤细胞明显诱导了cd4+及cd8+t细胞中sa-β-gal阳性细胞数的增加(图3a-c)。同时,流式细胞分析结果显示,肿瘤细胞诱导了上述cd4+及cd8+t细胞中共刺激分子cd27/cd28的丢失及细胞周期抑制蛋白p53/p21表达上调(图3d-g)。上述结果提示肿瘤细胞诱导了t细胞的老化。
4.肿瘤源性ilt4介导了肿瘤细胞诱导的t细胞老化。
为明确ilt4是否参与肿瘤细胞诱导的t细胞老化,我们阻断了共培养体系中ilt4,观察肿瘤细胞对t细胞老化的诱导作用。在肿瘤细胞-t细胞共培养体系中加入500ng/ml的ilt4中和抗体(anti-ilt4)或同型对照抗体,共培养24小时后分离cd4+及cd8+t细胞再单独培养72小时,sa-β-gal染色检测老化t细胞的比例。结果发现,anti-ilt4显著降低了sa-β-gal阳性的cd4+及cd8+t细胞的数量(图4a-c)。此外,anti-ilt4也部分逆转了cd4+t细胞cd27/cd28的下调(图4d),提示ilt4参与了肿瘤诱导的t细胞的老化。为进一步明确ilt4来源,我们分别用500ng/ml的anti-ilt4预处理肿瘤细胞或t细胞后2小时,然后洗去培养基中的抗体,再进行肿瘤细胞-t细胞共培养实验,结果发现,anti-ilt4预处理肿瘤细胞而非t细胞显著逆转了cd4+t细胞的老化(图4e),提示肿瘤源性ilt4而非t细胞来源的ilt4诱导了t细胞的老化。之后我们选择ilt4相对低(h1650、mcf7、m628、pc-9)/高(a549、h1299、zr751、m628)表达的肿瘤细胞系,利用慢病毒转染分别过表达/敲除肿瘤细胞ilt4,转染48小时后再次进行了肿瘤-t细胞共培养,结果发现老化cd4+(图4f-g)及cd8+t(图4h-i)细胞的数量被分别上/下调,再次验证了肿瘤源性ilt4诱导了cd4+及cd8+t细胞的老化。综合上述结果,ilt4在肿瘤细胞诱导的t细胞老化中发挥重要作用。
5.pir-b在小鼠肿瘤细胞中高表达。
ilt4在小鼠中的同源基因为pir-b。pir-b在小鼠肿瘤细胞中的表达及作用目前未见报道。我们首先采用real-timepcr及westernblot检测了pir-b在小鼠肺癌(mle12、ll/2)、乳腺癌(e0771)、恶性黑色素瘤(b16f0)细胞及正常乳腺上皮细胞株(fsk4)中mrna及蛋白水平的表达。结果显示,与正常小鼠乳腺上皮细胞fsk4相比,pir-b基因及蛋白在上述所有肿瘤细胞中均高表达(图5a,b)。
6.pir-b调控了小鼠肿瘤细胞的增殖、粘附、侵袭转移等生物学行为。
本课题组前期研究证实,ilt4在人nsclc细胞中的表达促进了其增殖、侵袭、迁移等恶性生物学行为。为明确小鼠肿瘤细胞pir-b表达是否同样调控其恶性生物学行为,我们选择小鼠乳腺癌(e0771)及恶性黑色素瘤(b16f0)细胞,通过慢病毒感染过表达/敲除了肿瘤细胞中pir-b表达,然后采用生长曲线、粘附实验及划痕实验分别检测肿瘤细胞的增殖、粘附及侵袭能力。结果发现:pir-b过表达/基因敲除显著促进/抑制了小鼠乳腺癌及恶性黑色素瘤细胞的增殖、粘附及侵袭迁移(图6a-c),提示pir-b的高表达同样可作为小鼠肿瘤的恶性表型。
7.免疫系统参与了pir-b调控的小鼠移植瘤的生长。
前面我们已证实,ilt4/pir-b不仅调控肿瘤的恶性生物学行为,同时调控t细胞的免疫功能。为验证ilt4对体内肿瘤微环境中t细胞免疫功能的影响,我们首先构建了野生型c57bl/6小鼠中pir-b过表达/敲除的e0771乳腺癌移植瘤模型评估pir-b对肿瘤生长与t细胞免疫的双重作用,同时利用免疫缺陷的nsg小鼠排除了pir-b对肿瘤恶性生物学行为的影响。6-8周大小的雌性c57bl/6j小鼠或nsg小鼠中皮下注射pir-b过表达/基因敲除的小鼠乳腺癌细胞e0771(2×105细胞/只)。每3天测量肿瘤长、短径评估大小。3-4周后当肿瘤最大径生长至2cm左右,co2处死小鼠,分离肿瘤组织并称重。结果发现,c57bl/6小鼠中pir-b过表达/敲除显著促进/抑制了体内肿瘤生长(图7a-c)。nsg小鼠中pir-b过表达/敲除同样促进/抑制了乳腺癌的生长,但组间差异明显小于全基因的c57bl/6小鼠模型(图7d-f),证明免疫系统确实参与了pir-b调控的肿瘤进展。
8.小鼠体内肿瘤源性pir-b通过诱导t细胞老化促进肿瘤的生长。
为体内证实肿瘤细胞pir-b对t细胞老化的诱导作用,我们从c57bl/6j荷瘤小鼠中分离血液(bl)、引流淋巴结(ln)、脾脏(sp)、肿瘤组织(til)中的t细胞,采用sa-β-gal染色检测t细胞老化,流式细胞术检测t细胞亚群分布及功能。结果发现,pir-b过表达/敲除分别促进/抑制了小鼠外周血(bl)及肿瘤组织(til)中淋巴细胞的免疫老化(图8a),但对脾脏(sp)及淋巴结(ln)中的淋巴细胞无影响。对于肿瘤组织中浸润t细胞数量及亚群构成的分析显示,pir-b未调控肿瘤组织中tils的浸润数量(图8b),但pir-b基因敲除似乎增加了cd8+t细胞亚群的构成(图8c)。功能研究显示,pir-b过表达/敲除分别下/上调了肿瘤浸润cd4+t细胞中ifn-γ的表达与cd8+t细胞中ifn-γ、granzymeb与perforin的表达(图8d-e),说明肿瘤源性pir-b体内调控了t细胞的杀伤能力。
结论:
1.ilt4在人的多种肿瘤细胞中高表达,与患者疾病进展及不良生存相关,可作为患者的不良预后指标;ilt4同源基因pir-b在小鼠肿瘤细胞高表达,促进其增殖、侵袭、迁移等恶性生物学行为。
2.肿瘤源性ilt4/pir-b在体内外诱导了cd4+及cd8+t细胞的免疫老化,抑制了其杀伤活性,促进了肿瘤进展。提示ilt4可以作为抗肿瘤免疫治疗靶点逆转肿瘤的免疫逃逸。
本发明未尽事宜为公知技术。
上述实施例只为说明本发明的技术构思及特点,其目的在于让熟悉此项技术的人士能够了解本发明的内容并据以实施,并不能以此限制本发明的保护范围。凡根据本发明精神实质所作的等效变化或修饰,都应涵盖在本发明的保护范围之内。
- 还没有人留言评论。精彩留言会获得点赞!