用于单细胞分选与增强IPSC重新编程的细胞培养平台的制作方法与工艺
用于单细胞分选与增强IPSC重新编程的细胞培养平台关于序列表的声明与本申请相关的序列表是以代替纸件形式的纯文本格式提供的,并通过引用并入本说明书。含有所述序列表的该文本文件的名称是FATE_094_00WO_ST25.txt。该文本文件的大小为4KB,其创建于2011年12月15日,并经由EFS-Web提交。引用的相关申请本申请根据35U.S.C.§119(e)要求2010年12月22日提交的美国临时专利申请61/429,369号,及2011年6月14日提交的美国临时专利申请61/496,991号的权益,在此通过引用将每一临时申请全文并入。发明
背景技术:
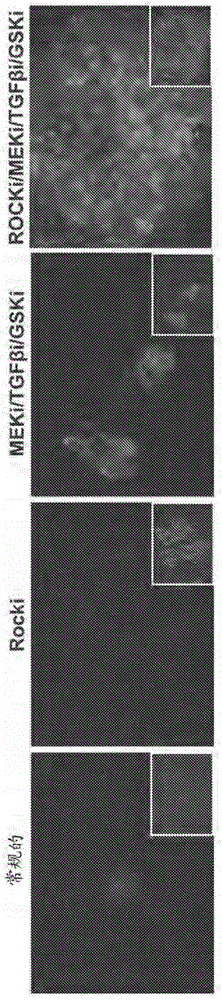
领域本发明通常涉及用于培养干细胞的细胞培养条件、培养基和培养平台,包括用于再生和培养人诱导性多能干细胞(iPSC)的无饲养层条件。相关领域描述多能干细胞生物学的应用为再生医学打开了新的大门。通过在生长因子的混合物中培养胚胎植入前囊胚,人胚胎干细胞(hESC)的衍生导致了许多有前途的细胞治疗方法,其中扩大的自我更新的细胞群可以在体外或在体内分化为与治疗有关的细胞系。在另一ESC生物学应用并且通过使用胚胎植入前基因分析,可能从几个遗传性疾病背景衍生ESC系从而在组织培养皿中模拟这些疾病。然而ESC技术存在一些局限性:自其中可以衍生ESC的遗传背景的范围受技术性与政治性限制,ESC的遗传背景并不总是已知的而且ESC衍生的细胞治疗本质上为同种异体移植物,冒着与传统的组织/器官移植相同的排斥风险。在一大进步中,多能细胞群产生于成年人的终末分化细胞:此类衍生的细胞被称作诱导性多能干细胞(iPSC)。iPSC技术允许来自任何供体的细胞重新编程为多能的自我更新的状态并且从而允许来自任何遗传背景的同种的细胞群扩增。iPSC克服了与ESC有关的伦理考量并且可以用于衍生任何遗传性人疾病的模型以用于高通量药物筛选或肝细胞与心肌细胞异型生物质的药物毒性筛选。另外,iPSC可最终导致自体移植中产生于患者自体细胞的防止移植排斥的细胞治疗。表达与差异分析显示iPSC与ESC在分子水平上非常接近,与多个ESC系相比时,所见到这些相似量级的克隆的iPSC培养物间差异。通常通过几个关键基因的异位表达产生iPSC,该关键基因为全部重新编程所需要的,即Oct4、Sox2、Klf4、c-Myc、Lin28以及Nanog的组合。最初使用整合病毒系统以表达关键转录因子产生iPSC。已成功地将包括多顺反子与诱导系统的逆转录病毒与慢病毒系统应用于iPSC产生。然而,由于插入突变而产生的永久性基因组变化与iPSC分化后外源基因再激活的潜在性可能为随后的药物筛选与这些方法所产生的细胞的治疗应用带来潜在的问题。确实已报道了使用相同的病毒系统所产生的iPSC克隆间显著的差异,在啮齿动物中作为分化神经球移植时有很大比例的形成肿瘤的克隆。研究表明使用相同的病毒方法所产生的iPSC一旦分化可有不同的表现。异位基因整合位点的差异可导致不同的插入突变与转基因表达的表观遗传调控。对包括整合系统的iPSC产生方法而言,需要衍生及筛选许多克隆以鉴定在多能的与分化的状态中都稳定的那些。因此,将来自给定的供体细胞源的克隆的iPSC的快速衍生的方法为有利的。尽管效率较低,已经证明了诸如腺病毒或附加型瞬时表达的用于iPSC产生的非整合系统的使用。这些系统可克服iPSC产生中的安全性与稳定性问题,然而当使用任何基于DNA的重新编程方法时存在基因组整合的潜在性并且这需要在它们用于研发iPSC衍生的细胞治疗之前进行评估。可切除的病毒系统与全基因组表达谱显示具有整合的表达组件的iPSC比相同的具有切除的病毒因子的克隆更不像ESC。而且,已证明了其中最常用的转录因子以具有细胞穿透肽的融合蛋白的形式表达于大肠杆菌(E.coli)中的只有蛋白重新编程。多剂量的纯化蛋白应用于小鼠成纤维细胞中导致iPSC产生。使用只有蛋白系统的重新编程的效率是很低的。这可归因于蛋白转导的效率、蛋白的特异活性和/或蛋白的稳定性。分化细胞通过多能性基因的异位表达重新编程的过程或它们经由蛋白转导或mRNA的引入需要几个月与熟练的干细胞生物学家的知识。最初通过眼睛进行重新编程的细胞的鉴定:ESC样集落形态的筛选。此类集落必须用手挑选,通常机械地传代及扩增。多能性因子的引入也产生转化细胞集落以及不完全重新编程的细胞。研究者能够从转化细胞的背景中鉴定出真正的iPSC集落,但这不是个高效的过程。然后需要进一步的特征与确认为真正的多能群体并且通常包括用于多能性标志物的免疫组织化学染色、基因表达与表观遗传分析以及多能群体分化为三个胚层(外胚层、中胚层与内胚层)的能力。一旦鉴定并且选择了多能细胞,通常此类细胞就会生长为集落并且需要在重新涂板之前通过挑选和机械地解离细胞来手工传代以长期保持细胞。来源于各种植入期之前与植入期之后的胚胎干细胞显示了不同的多能性状态。例如,衍生于囊胚的内细胞团的细胞被认为是更“原初的”并且具有完全不同于植入后衍生细胞的主要特性,该植入后衍生细胞被认为是更“成熟的”,具有较高的随机分化倾向性。原初细胞显得更“基态”并且不需要外在信号转导以保持它们的未分化状态。在另一方面,激活细胞需要包括TGFβ、激活素以及bFGF的关键细胞因子的外在信号转导并且非常依赖于ERK/MAPK细胞通路以保持它们的未分化状态。iPSC产生过程的改善可显著地降低技术壁垒,加速过程并且能够使随后的细胞按比例增加与分化以用于技术的工业应用诸如药物筛选与细胞治疗。需要用于更有效的不使用外源性物质iPSC生产以及更有效的重新编程细胞的鉴定与筛选的方法。促进人多能干细胞原初状态的产生iPSC的方法将极大地有利于将来在再生医学中的应用,诸如疾病校正、定向分化以及制造业规模扩张。而且,需要在使单细胞能够传代的特定的培养状态中更有效的产生iPSC的方法与可扩展性。发明概述本发明的一个实施方案提供了无饲养层环境中培养多能细胞的方法,其包括:在无饲养层环境中,在包含至少一种保持细胞多能性的制剂的培养基中,培养不是小鼠胚胎干细胞的多能细胞同时在培养期间保持细胞的多能性,其中所述制剂选自:i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂以及iv)Rock抑制剂,。在另一实施方案中,培养基包含足够量的制剂以允许至少一次细胞分裂同时保持细胞的多能性。在具体实施方案中,培养基包含至少三种或四种保持细胞多能性的制剂。仍在另一实施方案中,保持细胞多能性的制剂包含Rock抑制剂。在具体实施方案中,Rock抑制剂为thiazovivin或Y27632。在一实施方案中,保持细胞多能性的制剂包含TFGβ抑制剂,并且在具体实施方案中,TFGβ抑制剂为A-83-01或SB431542。在另一实施方案中,保持细胞多能性的制剂包含GSK3抑制剂,并且在具体实施方案中,GSK3抑制剂为CHIR99021或BIO。在一实施方案中,保持细胞多能性的制剂包含MEK抑制剂。在具体实施方案中,MEK抑制剂为PD98059或PD032901。在具体实施方案中,培养基包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂。在本发明更具体的实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在本发明的一些实施方案中,保持细胞的多能性至少五次细胞分裂。在其他的实施方案中,保持细胞的多能性至少十次细胞分裂。在一些实施方案中,在缺乏生长因子与细胞因子,任选地存在可溶性纤连蛋白的情况下培养细胞。在另一实施方案中,在缺乏MatrigelTM。的情况下培养细胞,并且仍在另一实施方案中,培养基基本上不含bFGF。在本发明的具体实施方案中,多能细胞为人胚胎干细胞或人诱导性多能干细胞。本发明的另一实施方案提供了包括在缺乏生长因子与细胞因子的情况下培养不是小鼠胚胎干细胞的多能细胞的培养多能细胞的方法。在某些实施方案中,方法包括在培养基中培养多能细胞,所述培养基包含至少一种保持细胞多能性的制剂以允许至少一次细胞分裂同时保持细胞的多能性,其中制剂选自:i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂以及iv)Rock抑制剂。在一实施方案中,多能细胞为人胚胎干细胞或人诱导性多能干细胞。在另外的实施方案中,培养基包含至少两种、至少三种或四种选自的制剂:i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂以及iv)Rock抑制剂;在具体的实施方案中,培养基包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在其他具体的实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。本发明的另一实施方案还提供了获得解离的人多能细胞的方法,包括解离人多能细胞以获得解离的细胞并且使解离的细胞接触培养基,所述培养基包含至少一种增强解离的细胞活力的制剂,其中制剂选自:i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂以及iv)Rock抑制剂,借此增强了解离的细胞的活力。在具体的实施方案中,解离的细胞的活力增强了至少10%、至少50%、至少100%、至少200%、或至少500%。在其他的实施方案中,本发明的方法还包括在培养基中培养解离的细胞至少一次、至少两次、至少五次、或至少十次传代,同时保持解离的细胞的多能性。在某些实施方案中,培养后解离的细胞的核型基本上与解离前细胞群的核型相似。在一些实施方案中,方法包括在制剂存在的情况下解离。仍在其他的实施方案中,方法包括在解离前将多能细胞与制剂接触。在具体的实施方案中,将解离的细胞与培养基接触包括将解离的细胞悬浮在培养基中。在其他的实施方案中,本发明提供了在无饲养层环境中增加细胞潜能的方法,包括在无饲养层环境中将细胞与包含至少一种小分子制剂的培养基接触以获得具有与接触培养基前的细胞相比潜能增加的细胞,其中小分子制剂选自:i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂以及iv)Rock抑制剂。在具体的实施方案中,培养基包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在一实施方案中,接触包括在足以增加细胞潜能的状态下培养细胞。在一些实施方案中,细胞选自:胚胎干细胞、多能细胞、专能细胞、非多能细胞以及体细胞。在具体的实施方案中,细胞不是小鼠胚胎干细胞。在其他的具体实施方案中,细胞为人细胞,并且在某些实施方案中,细胞为诱导性多能干细胞。在一些实施方案中,方法还包括将细胞与至少一种多能性因子接触。在一些实施方案中,多能性因子包含多核苷酸、多肽或小分子。在一些实施方案中,多能性因子为外源性转录因子。在具体的实施方案中,外源性转录因子包含Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸。在其他的实施方案中,多肽包含允许运输穿过细胞膜的氨基酸序列。在其他具体的实施方案中,外源性转录因子包含Oct4、Sox2以及Klf4多核苷酸。仍在其他的实施方案中,具有潜能增加的细胞通过下列的一种或多种进行表征:选自Oct4、Nanog、KLF4、SSEA4以及TRA1-81组成的至少一种多能干细胞标志物的表达;多能干细胞形态学;引起种系传递的能力;畸胎瘤的形成;分化或转分化为与起始谱系不同的谱系的能力,以及体外三系分化。在一些实施方案中,具有增加的潜能的细胞表达,与接触培养基前的细胞相比,至少高两倍的Oct4水平。仍在其他的实施方案中,具有增加的潜能的细胞具有与常规培养的iPSC相比至少低两倍的Xist活性。在另外的实施方案中,具有增加的潜能的细胞具有紧密的、半球形的集落形态。在一些实施方案中,在缺乏TFGβ、激活素以及MEK信号转导通路的外源性刺激,并且任选地缺乏bFGF通路的外源性刺激的情况下具有增加的潜能的细胞复制并且保持多能性。在其他的实施方案中,方法还包括在无饲养层环境中,在i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂或iv)Rock抑制剂中的至少一种存在的情况下,培养具有增加的潜能的细胞以允许至少一次细胞分裂同时保持细胞多能性。在具体的实施方案中,在TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂存在的情况下,培养细胞;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。本发明也提供了通过任何以上的实施方案所产生的具有增加的潜能的细胞。本发明还提供了在无饲养层环境中提高细胞群的重新编程的效率的方法,包括在足以诱导重新编程的条件下,将无饲养层环境中的细胞群与选自i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂,以及iv)Rock抑制剂组成的至少一种小分子制剂接触,借此,与没有接触小分子制剂的细胞群的重新编程的效率相比,重新编程的效率提高了至少10%、至少50%、至少100%、至少300%、或至少500%。在具体的实施方案中,细胞与TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂接触;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在一些实施方案中,重新编程之前细胞群包含非多能细胞。在其他的实施方案中,通过重新编程所需要的时间或重新编程的细胞数测量重新编程的效率。仍在其他的实施方案中,足以诱导重新编程的条件包括将细胞群与选自Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸的至少一种外源性转录因子接触。在具体的实施方案中,条件包括将细胞群与Oct4、Sox2及Klf4多肽或编码Oct4、Sox2及Klf4的多核苷酸接触。本发明的其他实施方案还提供了在无饲养层环境中分选细胞群以获得富含多能细胞的细胞群的方法,包括在无饲养层环境中获得包含混合细胞群的解离的细胞的悬浮液,并且分选悬浮液中的细胞以获得表达一种或多种多能性标志物的细胞,因此获得富含多能细胞的富集的细胞群。在一些实施方案中,悬浮液包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂或Rock抑制剂中的至少一种。在另外的实施方案中,分选为通过磁珠或流式细胞术。在具体的实施方案中,分选为通过磁珠。在其他的实施方案中,分选为通过流式细胞术。在一些实施方案中,方法还包括在包含i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂或iv)Rock抑制剂中的至少一种,任选地联合可溶性纤连蛋白的培养基中培养富集的细胞群。在具体的实施方案中,悬浮液中的混合细胞群在分选前与i)TFGβ抑制剂;ii)GSK3抑制剂;iii)MEK抑制剂或iv)Rock抑制剂中的至少一种接触。在某些实施方案中,混合的细胞群与TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂接触;在其他某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在具体的实施方案中,混合的细胞群包含表达一种或多种多能性标志物的细胞。在具体的实施方案中,一种或多种多能性标志物包含SSEA4、TRA160、TRA181、TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD30、CD50、CD133/prominin、CD140a、CD56、CD73、CD105、CD31、CD34、OCT4、Nanog或Sox2。在具体的实施方案中,多能性标志物选自SSEA4、TRA181、TRA160以及CD30。在具体的实施方案中,方法包括将混合的细胞群与一种或多种多能性因子接触以诱导重新编程。在一些实施方案中,接触包括将一种或多种多能性因子引入至混合的细胞群中的细胞。在某些实施方案中,多能性因子包含Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸。在其他具体的实施方案中,多能性因子包含Oct4、Sox2及Klf4多肽或编码Oct4、Sox2及Klf4多肽的多核苷酸。在某些具体的实施方案中,多能细胞为诱导性多能细胞。在一些实施方案中,就表达一种或多种多能性标志物而言,富集的细胞群丰富了至少20%、至少50%、至少100%、至少200%、或至少500%。在其他的实施方案中,本发明提供了获得诱导性多能干细胞的方法,包括处理细胞群以诱导重新编程;制备包含细胞群的解离的细胞的悬浮液;分选悬浮液中的细胞以获得表达一种或多种多能性标志物的分选细胞;培养表达一种或多种多能性标志物的分选细胞,其中获得iPSC。在具体的实施方案中,在缺乏细胞因子与生长因子,任选地在无饲养层环境中,并且任选地在可溶性纤连蛋白存在的情况下,培养分选细胞。在一些实施方案中,细胞群与i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂中的至少一种接触。在具体的实施方案中,细胞群与TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂接触;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在其他的实施方案中,处理细胞群以诱导重新编程,包括将细胞群与一种或多种多能性因子接触。在具体的实施方案中,多能性因子包含Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸。在某些实施方案中,多能性因子包含Oct4、Sox2及Klf4多肽或编码Oct4、Sox2及Klf4多肽的多核苷酸。在其他的实施方案中,处理细胞群以诱导重新编程还包括将细胞群与i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂中的至少一种接触。在具体的实施方案中,细胞群与TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂接触;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在其他的实施方案中,悬浮液包含i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂中的至少一种。在具体的实施方案中,悬浮液包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在一些实施方案中,分选通过流式细胞术或磁珠。在具体的实施方案中,分选细胞以获得表达至少一种、两种、三种、四种、或多种多能性标志物的细胞。在其他具体的实施方案中,一种或多种多能性标志物包含SSEA4、TRA160、TRA181、TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD30、CD50、CD133/prominin、CD140a、CD56、CD73、CD105、CD31、CD34、OCT4、Nanog或Sox2。在其他具体的实施方案中,一种或多种多能性标志物选自SSEA4、TRA160、TRA181以及CD30。在一些实施方案中,一种或多种多能性标志物为SSEA4、CD30以及TRA160或TRA181。在另外的实施方案中,使用特异的标志物分选细胞以从重新编程的群体中除去非重新编程的细胞。在一些实施方案中,培养包括在包含选自i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂的至少一种小分子制剂的培养基中培养细胞。在具体的实施方案中,培养基包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在具体的实施方案中,在无饲养层环境中培养细胞。在某些实施方案中,在无饲养层环境中处理、悬浮、分选以及培养细胞。在一些实施方案中,在约2天-22天中获得诱导性多能干细胞。在具体的实施方案中,在约4天-约18天中获得诱导性多能干细胞。在其他的实施方案中,在处理细胞群以诱导重新编程后约4天-约22天内获得诱导性多能干细胞。在某些实施方案中,在处理细胞群以诱导重新编程后约6天-约18天内获得诱导性多能干细胞,并且在其他某些实施方案中,在处理细胞群以诱导重新编程后约10天-约16天内获得诱导性多能干细胞。本发明也提供了在其他的实施方案中通过任何以上的方法获得诱导性多能干细胞。本发明还提供了从细胞群消耗多能细胞的方法,包括获得包含具有多能细胞的混合细胞群的解离的细胞的悬浮液,并且分选悬浮液中的细胞以去除表达一种或多种多能性标志物的细胞,因此从细胞群中消耗多能细胞。在一些实施方案中,混合的细胞群包含专能细胞或(成熟的)体细胞。在其他的实施方案中,获得悬浮液之前,在包含选自i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂的至少一种小分子制剂的培养基中培养悬浮液中的混合细胞群。在具体的实施方案中,培养基包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在本发明另外的实施方案中,悬浮液包含选自i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂的至少一种小分子制剂。在具体的实施方案中,悬浮液包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂;在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在一些实施方案中,分选通过流式细胞术。在其他的实施方案中,分选通过抗体包被的磁珠富集。本发明的一些实施方案提供了获得具有基因组稳定性的多能细胞的方法,包括在无饲养层环境中,在缺乏c-myc的情况下,在足以获得具有基因组稳定性的多能细胞的条件下,将细胞与选自i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂的至少一种小分子制剂接触。在一些实施方案中,细胞为胚胎干细胞、多能细胞、专能细胞、非多能细胞以及体细胞。在一些具体实施方案中,细胞包含非多能细胞。在某些实施方案中,条件包括细胞与至少一种多能性因子接触。在一些实施方案中,多能性因子为选自Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸的外源性转录因子。在具体的实施方案中,多能性因子包含Oct4、Sox2以及Klf4多核苷酸。在本发明的一些实施方案中,小分子制剂包含Rock抑制剂。在具体的实施方案中,Rock抑制剂为thiazovivin或Y27632,并且在更具体的实施方案中,Rock抑制剂为thiazovivin。在其他的实施方案中,小分子制剂包含TFGβ抑制剂。在一些实施方案中,TFGβ抑制剂为A-83-01或SB431542。在一些实施方案中,小分子制剂包含GSK3抑制剂,并且在具体的实施方案中,GSK3抑制剂为CHIR99021或BIO。在本发明其他的实施方案中,小分子制剂包含MEK抑制剂。在一些具体的实施方案中,MEK抑制剂为PD98059或PD032901。在本发明的一些实施方案中,小分子制剂包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂。在具体的实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。本发明额外地包含培养多能细胞以保持细胞基因组稳定性的方法,包括无饲养层环境中在包含至少一种保持多能细胞的基因组稳定性的制剂的培养基中培养多能细胞,同时保持培养期间多能细胞的基因组稳定性,其中制剂选自i)TFGβ抑制剂、ii)GSK3抑制剂、iii)MEK抑制剂以及iv)Rock抑制剂。在某些实施方案中,条件包括细胞与至少一种多能性因子接触。在一些实施方案中,多能性因子为选自Oct4、Sox、Klf、Myc、Lin28或Nanog多肽或编码Oct4、Sox、Klf、Myc、Lin28或Nanog的多核苷酸的外源性转录因子。在具体的实施方案中,多能性因子包含Oct4、Sox2以及Klf4多核苷酸。在一些实施方案中,培养基包含至少两种、至少三种、或四种制剂。在本发明的一些实施方案中,制剂包含Rock抑制剂。在具体的实施方案中,Rock抑制剂为thiazovivin或Y27632,并且在更具体的实施方案中,Rock抑制剂为thiazovivin。在本发明其他的实施方案中,制剂包含TFGβ抑制剂。在一些实施方案中,TFGβ抑制剂为A-83-01或SB431542。在一些实施方案中,制剂包含GSK3抑制剂,并且在具体的实施方案中,GSK3抑制剂为CHIR99021或BIO。在本发明其他的实施方案中,制剂包含MEK抑制剂。在一些具体的实施方案中,MEK抑制剂为PD98059或PD032901。在本发明的一些实施方案中,制剂包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂。在具体的实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在本发明的一些实施方案中,在培养基中培养多能细胞至少一次、至少两次、至少五次、至少十次、或至少15次传代,同时保持多能细胞的基因组稳定性。附图简要说明图1.各种小分子混合物对无饲养层基质上人多能干细胞的单细胞培养的影响。A.当在无饲养层环境中用小分子的各种组合处理单细胞解离的人多能干细胞时,不同的形态是明显的。包含ROCKi/MEKi/TGFβi/GSKi组合的培养基给予了人多能干细胞的强健的生长与活力。B.虽然一些小分子,即MEKi/TGFβi/GSKi支持多能干细胞的保持,并且其他的即Rocki促进多能干细胞的单细胞解离,这些小分子的独特组合,即ROCKi/MEKi/TGFβi/GSKi,却支持单细胞解离同时保持未分化的状态,如对多能性标志物Tra181反应阳性的高度增生的集落所示。Tra181,红色;DAPI,蓝色。C.当hiPSC以1×105个细胞接种于人工基底膜时,MEKi/TFGβi/GSKi/ROCKi的组合最佳地支持活力与增生并且4天后为活细胞数与台盼蓝所测量的活力百分比打分。D.当与MEKi/TFGβi/GSKi组合时,相对于MEKi/TFGβi/GSKi与Y27632所支持的培养,用Thiazovivin培养的集落在形态上显得更紧密。E.Y27632或Thiazovivin与MEKi/TFGβi/GSKi组合中所培养的hiPSC的SSEA4与Tra181的表面表达的流式细胞术分析显示,与具有Y27632的组合相比,Thiazovivin更好地支持细胞保持在未分化的状态。图2.人多能干细胞适应于无饲养层与单细胞培养。A.饲养层细胞上所产生的iPSC易于适应利用SMC4培养基的无饲养层与单细胞培养条件。然而,在常规培养条件下单细胞解离与培养可见扰乱细胞生存以及有限的细胞粘附与生长。B.一旦适应无饲养层环境,iPSC就作为单细胞在无饲养层表面进行常规地培养,同时持续保持在SMC4培养基中。图3.单细胞的长期保持与无饲养层人多能干细胞培养。A.单细胞解离的人多能干细胞保持在无饲养层SMC4培养基中培养10次传代,通过免疫荧光分析表达多能性标志物Nanog与Tra181。B.图3B显示了人多能干细胞被解离为单细胞并且置于常规地培养基或SMC4培养基中后4小时,7氨基放线菌素D(7AAD)染色细胞的百分比。当培养于SMC4培养基而不是常规的培养基中时,单细胞解离的人多能干细胞显示了显著增加的活力,如通过7AAD染色与流式细胞术分析所见到的。C.通过流式细胞术的定量群体分析揭示几乎所有的细胞解离为单细胞并且保持在SMC4培养基中表达多种未分化的状态的标志物。D.总体表达分析揭示作为单细胞传代并且保持在无饲养层细胞的SMC4培养基中的iPSC,与培养在饲养层细胞上的并且传代期间保持为细胞团块的人ESC,包括H1细胞,高度相关。E.使用SMC4培养基培养于无饲养层细胞系统中并且作为单细胞传代的iPSC,而不是它们的原始来源成纤维细胞,表达多能性标志物阵列,与人ESC相似,包括H1与HuES9。F.保持在无饲养层环境中的iPSC重新编程因子的外源性表达被有效地抑制,不同于外源性因子阳性对照(慢病毒感染4天后表达重新编程因子的成纤维细胞)。重新编程因子在无饲养层iPSC中的内源性表达与ESC相似,包括H1与HuES9。G.在无饲养层环境中作为单细胞11次传代后,人多能干细胞保持正常的核型。H.用常规分化的培养基替换SMC4培养基14天后,保持在无饲养层环境中的细胞显示了指示除了胚外的标志物之外,外胚层、内胚层以及中胚层胚层的基因表达,其通常只由原初的与全能干细胞表达。I.免疫荧光分析:使用SMC4培养基培养于无饲养层细胞系统中并且作为单细胞传代的iPSC,一旦转移到常规的分化培养基上,易于发育成为所有三种体细胞类型。中胚层,α平滑肌肌动蛋白(αSMA);外胚层,β微管蛋白III(βTUBIII);内胚层,Sox17。J.畸胎瘤形成分析证明SMC4培养基中培养的人多能干细胞产生代表三种胚层的细胞类型,并且因此保持它们的多能潜力。图4.增强的单细胞分选与人多能干细胞随后的无饲养层培养。基于多能性标志物表达的人多能干细胞的分选的演示以及随后在添加了优化SMC4培养基或SMC4+纤连蛋白培养基的无饲养层培养中保持分选细胞。A.分选前,人多能干细胞解离成为单细胞并且用抗体标志物标注至SSEA4与Tra-181。使用FACs技术,基于多能性标志物的表面标志物表达,SSEA4与Tra-181分选细胞。分选后最初将细胞接种于SMC4+纤连蛋白培养基,24-72小时后变换为SMC4培养基。分选后第1天,将分选的细胞分开并且到第5天,出现由数百个细胞组成的大的集落。B.分选的细胞保留它们的多能性标志物Tra160的表达。Tra160,红色;DAPI,蓝色。C.5次传代后,分选前,培养中大约1个月,基于SSEA4与Tra181染色,定量流式细胞术分析揭示大多数分选的细胞仍保留它们的多能状态。D.来源于SSEA4+与Tra181+人多能干细胞的各种密度在添加了SMC4+纤连蛋白培养基的无饲养层表面涂板,包括6孔平板每孔500个事件的集落密度,大约52个事件/cm2。分选后7天后,许多碱性磷酸酶阳性集落出现在每个接种孔中。E.计分所产生的碱性磷酸酶阳性集落。大约8-10%的接种事件产生每孔16000个事件的集落,产生了几乎融合的孔。由于融合阻止了精确的计数不计算N.C.。F.将SSEA4+与Tra181+人多能干细胞以各种密度直接分选至96孔平板,包括1个事件/孔,大约3个事件/cm2。分选后8天,用碱性磷酸酶将孔染色。G.在第8天计分每个孔碱性磷酸酶集落。图5.在SMC4培养基缺乏与存在的情况下的重新编程动力学。A.通过Oct4、Sox2、Klf4以及cMyc异位表达重新编程启动后7天,或将细胞保持在常规的培养基中或将细胞转移至SMC4培养基。重新编程20天后,常规的培养基培养中很少有任何类似iPSC集落形成,然而转移至SMC4培养基的细胞中出现了许多类似iPSC集落。B.SMC4培养基中许多类似iPSC集落表达碱性磷酸酶而常规培养中很少集落表达碱性磷酸酶。图6.在SMC4培养基中培养提高了原初特征。A.人多能干细胞适应SMC4培养基的示意图以及它与常规保持的人多能干细胞的比较。B.基于Affymetrix总体基因表达的分级群聚与各自培养基中都培养了11次传代,IMR90亲代细胞衍生的hiPSC(常规衍生的hiPSC(Conv)与SMC4培养基适应的hiPSC(SMC4培养基))间Pearson相关系数,都证明hiPSC彼此相似并且不同于它们的亲代IMR90细胞。C.虽然常规格式中培养的hiPSC与它们的亲代IMR90细胞系相比,Xist表达被中度抑制,而SMC4培养基中培养的hiPSC中Xist表达被显著地抑制,与雌性的hESC、HUES9水平相似。D.位于X染色体的基因在SMC4培养基中培养的hiPSC中表达程度更高,与常规培养的hiPSC中表达相比。E.当转移回到饲养层细胞时,保持在SMC4培养基中的hiPSC显示了更类似于小鼠ESC的形态,如通过与常规培养基中培养的hiPSC相比可见到更多的三维形状与更少的平面形态。图7.SMC4培养基中产生与保持改善了人多能干细胞的未分化状态。A.FTi91与93克隆平行产生的示意图,在SMC4培养基中产生并且保持在SMC4培养基FF培养中,以及FTi99克隆,在常规培养基中产生并且保持在饲养层细胞上。B.基于Affymetrix总体基因表达的分级群聚与Pearson相关系数描述了衍生的多能系间密切的关系以及与起始细胞系(FTC1)关系遥远。C.基于Hues9、H1、FTi99、FTi91以及FTi93间所有4倍差异表达的基因的分级群聚证明了常规系统中培养的多能系与SMC4培养基中培养的那些多能系之间的分离。D.SMC4培养基组(FTi91与93)与常规培养组(H1、Hues9以及FTi99)相比,与原初与谱系分化有关的选择基因的Affymetrix基因表达显示增加了许多与多能性有关的基因表达并且显著减少了SMC4培养基集中谱系特异的基因表达。E.各种获得原初状态的hiPSC方法的示意图。紫色的图层描述了原初状态超越成熟状态的优势。图8.重新编程过程期间产生的非iPSC集落。A.重新编程过程早期典型的细胞群:免疫染色显示大多数快速生长的细胞形成SSEA4阴性并且不可避免地非多能细胞的集落。这些细胞为转化的,高度增生的,生长迅速的并且在培养中占主导地位。集落1显示SSEA4阳性集落有变成iPSC集落的潜能而集落2显示了具有高增生率但是多能性标志物染色阴性的转化群。B.许多非iPSC集落显示了单一多能性标志物的表达,而真正的iPSC集落显示了多重多能性标志物的表达。重新编程的后一阶段,形成各种集落,一些集落SSEA4与Tra181双阳性,即集落1,而其他的集落只有标志物的其中之一为阳性或一点都没有,例如集落4与5。图9.使用细胞分选多能细胞的选择诸如iPSC。单细胞培养系统与用于从多能与非多能细胞的混合物中富集并且选择的细胞分选方法的示意图显示。图10.多能性标志物的鉴定。为研究对SSEA4+/Tra181+细胞特异的额外的标志物,评估各种表面标志物。Tra160表达的检测用作阳性对照,因为已经证实它与Tra181表达很好地关联。CD200与CD90似乎没有针对多能细胞特异,因为它们相似地染色SSEA-/Tra181-与SSEA+/Tra181+群,而没有见到CD9针对SSEA+/Tra181+群特异性、然而,CD50与CD30似乎越过SSEA-/Tra181-群优选地鉴定SSEA4+/Tra181+群。图11.CD30对Nanog表达基因具有特异性。A.各种hiPSC系的SSEA4/Tra181表达的代表性流型证明所有衍生的克隆代表大多数表达SSEA4与Tra181的群。B.基于(A)中所见的门控,评估每一细胞系的CD30与CD9流型。只有克隆FTC8克隆1与FTi31似乎表达CD30而所有的克隆都表达CD9。C.也评估各谱系的相对Nanog表达。D.汇总表证明只有CD30表达与Nanog表达相关。图12.重新编程期间去除非重新编程的细胞使重新编程的细胞富集。重新编程过程期间,存在重新编程与非重新编程的细胞的混合群,只有较少的细胞群代表重新编程为完整多能性的细胞,如通过SSEA4与Tra181表达所示(通过黑色虚线箭头所示)。然而,当将非重新编程的细胞从混合群中移除或去除时,显示了SSEA4与Tra181阳性群的显著富集。如灰色实线箭头所示,CD13阴性细胞群代表SSEA4与Tra181阳性细胞超过CD13低与高群体细胞的显著富集。图13.通过单细胞分选增强细胞重新编程并且从包括IMR90与ASC的各种群中富集多能干细胞。A.通过SSEA4+细胞的单细胞分选富集重新编程过程早期的细胞群。细胞的起始群,包含重新编程的与非重新编程的细胞,被酶促解离成单细胞,独特的多能性标志物染色,以及基于表面标志物表达通过免疫连接的磁珠分选以获得对SSEA4阳性的细胞群。这一SSEA4+细胞富集后,将10000个SSEA4+细胞接种于MatrigelTM包被的皿并且在FF环境常规的培养基或SMC4培养基中培养。分选后8天进行AP染色。这些右边角落中较小的插入图层为代表性细胞形态的图像。没有在常规培养中检测到碱性磷酸酶阳性的集落而许多碱性磷酸酶阳性的集落衍生于SMC4培养基。B.计分衍生于SMC4培养基存在或缺乏的情况下MatrigelTM上细胞的重新编程的SSEA4+/Tra181+集落的量。C.无饲养层条件下衍生于IMR90成纤维细胞或脂肪干细胞的iPSC克隆与使用重新编程与SMC4培养基的单细胞培养表达多能性标志物。D.使用单细胞分选方法衍生iPSC克隆的全群体流式细胞术分析揭示大多数细胞对关键的多能性标志物为阳性的。E.使用单细胞分选SSEA4与无饲养层培养而不是它们原始源成纤维细胞或脂肪干细胞所产生的iPSC表达多能性标志物的阵列,与人ESC相似,包括H1与HuES9。F.重新编程因子的外源性表达在使用单细胞分选与无饲养层培养所产生的iPSC中有效地沉默,不同于被慢病毒感染3天后表达重新编程因子的对照的成纤维细胞。G.使用基于SSEA4单细胞分选所产生的iPSC一旦分化易于发育为所有三种体细胞类型。内胚层,FoxA2;中胚层,α平滑肌肌动蛋白(αSMA);外胚层,β微管蛋白III(βTUBIII)。图14.用于独特的多能干细胞群的选择的单细胞分选。A.使用FACS分选,选择重新编程过程早期为SSEA4+/Tra181+的独特稀少的细胞群并且转移到添加了SMC4+纤连蛋白培养基,24-72小时后变换为SMC4培养基的无饲养层培养。培养另外6天后,SSEA4+/Tra181+衍生的iPSC集落似乎在无饲养层培养中生长。然而,当SSEA4-/Tra181-集落被转移并且保持在无饲养层培养中时,培养14天后检测不到表达碱性磷酸酶的集落。B.各种SSEA4+/Tra181+分选的细胞发育为集落同时保持它们的SSEA4与Tra181表达。图15.高通量产生方法与无饲养层条件下iPSC的主要特征。使用SMC4培养基(当分选时使用SMC4+纤连蛋白培养基),以两步骤的方式分选三种因子(Oct、Sox、Klf)诱导的成纤维细胞以送递有效的96孔平板平台。标记包含个体集落的孔并且扩展到4×96孔平板。当一组被设计为主平板并且扩增时,处理其他三个平板以表征,包括用于包括SSEA4与Tra181的表面标志物表达的流式细胞术分析,用于包括Nanog的关键标志物表达的qRTPCR,转基因沉默以及用于包括Oct4与Nanog多能性标志物的免疫荧光。基于特征资料解析,扩增堆积选择的hiPSC克隆以用于进一步的分析。数据图层代表FTC5的hiPSC产生高通量平台期间所研究的克隆的快照(通过96孔平板的31-40孔所标记)。qPCR图层中,将表达标化为Gapdh。Nanog表达与H1hESC有关而转基因表达与FTC5感染后第4天有关(第4天感染)。基于特征资料解析,扩增堆积选择的hiPSC克隆以用于进一步的分析。在突出的实例中,基于它的多参数多能性分布图鉴定37孔作为扩增候选并且称为FTC5克隆1。以5×放大率取得免疫荧光图像。图16.SMC4存在的情况下,3-因子(多顺反子性的-OKS)hiPSC的高通量平台,克隆的以及FF衍生。A.染色衍生于以上策略的FTi91与FTi93用于多能性标志物的表达。B.针对FTC1(包皮成纤维细胞),H1与Hues9(hESC系),FTi91与93(FTCI衍生的hiPSC克隆)以及第4天P.I.(感染后第4天)的多能性标志物的外源性表达的qRT-PCR。表达标化为Gapdh以及每一基因组内相关性。C.Oct4启动子甲基化状态。开环代表未甲基化的CpG岛而黑圈代表甲基化的CpG岛。D.分化28天后FTi91与93克隆的EB形成与分化。内胚层,FoxA2;中胚层,α平滑肌肌动蛋白(αSMA);外胚层,Tuj1。E.衍生于FF培养与SMC4中所产生并且保持的hiPSC克隆的畸胎瘤的组织切片。黑箭头指向感兴趣的区域:中胚层,白色脂肪组织;外胚层,神经元;内胚层,腺体。图17.重新编程中多能干细胞的产生与保持与SMC4培养基保持基因组完整性并且能够富集多能细胞。A.如通过阵列比较基因组杂交所评估的拷贝数变异。底表为数据的解释汇总,展示了SMC4培养基培养的人多能干细胞与它们亲代的细胞系间最小的拷贝数变异。B.G带中期细胞的细胞遗传学分析。底表为数据的解释汇总,描述了SMC4培养基中长期无饲养层培养后基因组的稳定性。图18.FF与SMC4培养下FTC5与FTC7衍生的克隆的hiPSC的特征。为检测高通量平台的重复性,测试额外的患者同意的FTC5与FTC7以用于hiPSC产生。A.多能性标志物Oct4、Tra181、Nanog以及Tra160代表的免疫荧光染色,其表达于OKS诱导的并且使用多重平台所产生的个体的FTC5与FTC7衍生的hiPSC克隆。B.诱导分化后28天hiPSC衍生的FTC5与FTC7的代表的谱系特异的染色。内胚层,FoxA2;中胚层,α平滑肌肌动蛋白(αSMA);外胚层,Tuj1。C.来源于各种传代中长期FF与单细胞培养后FTC5与FTC7衍生的hiPSC克隆的G带中期细胞的细胞遗传学分析。图19.使用细胞表面标志物保持多能培养的方法。A.如通过无饲养层与SMC4平台中Tra181富集所示的,培养期间进一步富集多能干细胞群。B.多能细胞群培养期间,群内一定比例的细胞可开始分化并且失去它们的多能性。使用用于多能性的选择方法,即,Tra181富集,获得未分化细胞的同种培养。图20.从非、部分以及完全多能的干细胞的异源群体分离分化的细胞。重新编程的人成纤维细胞群的FACS分选:当分选对多能性标志物SSEA4-/Tra181-双阴性的细胞(蓝色框;左下部框)时,没有检测到碱性磷酸酶(AP)阳性的集落,表明培养一周后失去了多能性与潜在的致癌性,反之SSEA4+/Tra181+双阳性群体的分选(桔色框,右上部框)导致了多能细胞的选择与富集,如通过AP+集落形成所证明的。图21.无饲养层培养上人多能干细胞的无细胞因子培养。A.当在缺乏包括bFGF的任何细胞因子的情况下,培养于明胶包被的表面并且添加了SMC4培养基时,人多能干细胞保持它们的生长与形态。B.放大的图像描述了人多能干细胞集落内个体的细胞。C.在传代第3次SSEA4与Tra181的共表达进一步地证明了无细胞因子培养的人多能干细胞保持它们的未分化状态。发明详述本发明提供了强力的用于培养干细胞的培养系统,包括用于产生及培养诱导性多能干细胞(iPSC)的无饲养层条件。具体地,本发明提供了培养平台,其允许多能细胞在无饲养层环境中长期培养;细胞在无饲养层环境中重新编程;多能细胞的单细胞解离;多能细胞的细胞分选;提高重新编程效率;原初多能细胞的产生;以及用于多能细胞鉴定与选择的标志物鉴定。本发明的培养基与培养方法支持单细胞解离的人干细胞的活力与存活,并且保持干细胞的未分化状态以允许没有分化的解离的单细胞培养与传代。定义如本文所用的,术语“重新编程”或“去分化”或“增加细胞潜能”或“提高发育能力”指增加细胞潜能或使细胞去分化为分化程度较低状态的方法。例如,具有增加的细胞潜能的细胞,与非重新编程的状态中相同的细胞相比,具有更大的发育可塑性(即能够分化为更多的细胞类型)。换句话说,重新编程的细胞为比非重新编程状态中相同的细胞分化程度更低的细胞。如本文所用的,术语“潜能”指所有可得到细胞的发育选择(即发育能力)的总数。本领域中技术人员应理解细胞潜能为连续体,范围自具有最大的发育能力的最可塑的细胞,全能干细胞至具有最小的发育能力的最不可塑的细胞,终末分化细胞。细胞潜能的连续体包括,但不限于,全能细胞、多能细胞、专能细胞、寡能细胞、单能细胞以及终末分化细胞。在严格意义上,干细胞为多能的;因此能够产生任何成熟的细胞类型。然而,专能细胞、寡能细胞或单能的祖细胞有时候被称为谱系限制性干细胞(例如间充质干细胞、脂肪组织衍生的干细胞等)和/或祖细胞。如本文所用的,术语“多能的”指细胞形成所有机体或体细胞(即胚体)的谱系的能力。例如,胚胎干细胞为一种类型的多能干细胞,其能够形成来源于三种胚层,外胚层、中胚层以及内胚层的每一种的细胞。多能性为发育能力的连续体,范围自不能产生完整的有机体的不完全地或部分地多能细胞(例如外胚层干细胞或EpiSC)至能够产生完整的有机体的更原始的、更多能的细胞(例如胚胎干细胞)。通过评估细胞的多能性特征可以确定细胞多能性的水平。多能性特征包括,但不限于i)多能干细胞形态;ii)多能干细胞标志物的表达,其包括但不限于SSEA1(只有小鼠),SSEA3/4;TRA1-60/81;TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD105、CD31、CD34、OCT4、Nanog和/或Sox2以及如本发明所述的CD30与CD50;iii)小鼠多能干细胞促进小鼠嵌合体中种系传递的能力;iv)使用四倍体补偿测定多能干细胞对胚体贡献的能力;v)多能干细胞的畸胎瘤形成;vi)胚体的形成;以及vii)失活的X染色体重新激活。如本文所用的,术语“多能干细胞形态”指胚胎干细胞经典的形态特征。通过形状圆而小,高的核质比,核仁的显著存在,以及典型的细胞间间距表征正常的胚胎干细胞形态。如本文所用的,术语“专能的”指成体干细胞形成一个谱系多重细胞类型的能力。例如,造血干细胞能够形成血细胞谱系的所有细胞,例如,淋巴细胞与骨髓细胞。“粘附”指细胞依附于器皿,例如,在适当的培养基存在的情况下,细胞依附于无菌塑料的(包被塑料的)细胞培养皿或烧瓶。某些细胞种类在培养中不能持续或不能生长,除非它们粘附在细胞培养皿上。某些细胞种类(“非粘附细胞”)不需要粘附而在培养中保持和/或增殖。“培养”或“细胞培养”指在体外环境中细胞的保持、生长和/或分化。“细胞培养基”、“培养基”(在每种情况下单数的“培养基”)、“添加”以及“培养基添加”指培养细胞培养物的营养组分。“培养”指组织以外或体外,例如在无菌塑料的(或包被塑料的)细胞培养皿或烧瓶中,细胞的保持、增殖(生长)和/或分化。“培养法”可利用培养基作为营养素、激素和/或其他有助于增殖和/或保持细胞的因子的来源。如本文所用的,“解离的”细胞指从其他的细胞或从表面(例如培养皿表面)基本上分离或纯化掉的细胞。例如,细胞能够通过机械的或酶促的方法从动物或组织分离。可选地,在体外聚集的细胞能够彼此分离,诸如通过酶促地或机械地解离成为群集、单细胞或单细胞与群集混合的悬浮液。仍然在其他可选的实施方案中,粘附细胞从培养皿或其他的表面分离。因此解离可以包括破坏细胞与细胞外基质以及基质(例如培养表面)的相互作用或破坏细胞间ECM。如本文所用的,术语“富集”与“使……富集”指增加组合物中特异组分的量,诸如细胞组分,并且当用于描述细胞组合物诸如细胞群时,“富集的”指具有与富集前细胞群中此类组分的比例,特异组分的量成比例地增加的细胞群。例如,对靶细胞类型(即具有特异特征的细胞)而言,富集诸如细胞群的组合物,因此与富集前存在于细胞群中的靶细胞的比例相比,具有靶细胞类型的比例或百分比增加。可通过本领域中已知的细胞选择与分选方法针对靶细胞类型进行细胞群富集。在本发明的一些实施方案中,通过本文实施例中所述的分选或选择过程富集细胞群。在本发明具体的实施方案中,针对靶细胞群富集的方法,对靶细胞群而言细胞群富集了至少大约20%,意指富集的细胞群包含比群体富集前群体中的靶细胞类型成比例地多大约20%的靶细胞类型。在一实施方案中,针对靶细胞群富集的方法,对靶细胞群而言,细胞群富集了至少大约30+%、40+%、50+%、60+%、70+%、80%、85%、90%、95%、97%、98%或99%、或至少大约98%或在具体的实施方案中,大约99%。在本发明的某些方面,对多能细胞的量或细胞展示的多能性特征而言,富集细胞群。在本发明具体的实施方案中,针对具有诸如多能性标志物表达的多能性特征的靶细胞,富集进行重新编程的细胞群,多能性标志物包括但不限于SSEA4、TRA1-60、TRA-1-81、CD30或CD50。在本发明其他具体的实施方案中,使用对分化或非多能细胞特异的表面标志物消耗细胞群,诸如进行重新编程的细胞群中的非多能细胞,例如对分化或非多能细胞特异的表面标志物其包括CD13、CD26、CD34、CD45、CD31、CD46或CD7。因此,所得到的细胞群被描述为针对多能细胞富集的细胞群。在本发明的某些方面,针对靶细胞富集的富集细胞群中的细胞具有独特的基因或蛋白表达谱,例如,至少两种诸如SSEA4、TRA1-60、TRA-1-81、CD30以及CD50的多能性标志物的细胞表面表达。在一些实施方案中,针对表达两种或多种多能性标志物的靶细胞富集细胞群。在具体的实施方案中,针对表达SSEA4与Tra181或Tra160组合的靶细胞富集细胞群。在更具体的实施方案中,针对表达SSEA4、Tra181以及CD30的靶细胞富集细胞群。在一实施方案中,富集细胞群中至少大约5%、10%、15%、20%、25%、30%、40%、50%、70%、75%、80%、90%、95%、97%、98%或99%的细胞为靶细胞类型,诸如多能细胞。因此,在一些实施方案中,本发明提供了针对多能细胞富集细胞群的方法,通过基于多能性标志物的细胞表面表达,诸如SSEA4、TRA1-60、TRA-1-81、CD30以及CD50,分选细胞群并且收集表达此类标志物的细胞以获得针对多能细胞富集的细胞群。在本发明其他的实施方案中,针对多能细胞富集细胞群,通过基于分化或分化细胞标志物的细胞表面表达,诸如CD13、CD26、CD34、CD45、CD31、CD46以及CD7,分选细胞群并且消耗此类细胞的细胞群以获得针对多能细胞富集的细胞群。在具体的实施方案中,基于CD13的表达分选细胞群,并且将CD13+细胞从细胞群移除以获得针对多能细胞富集的细胞群。如本文所用的,“饲养层细胞”或“饲养层”为用于描述一种类型的细胞与第二种类型的细胞共培养以提供第二种类型的细胞可以在其中生长的环境的术语,因为饲养层细胞为第二种细胞类型的支持提供了生长因子与营养素。随着它们支持的细胞饲养层细胞任选地来源于不同的种类。例如,某些人细胞类型,包括干细胞,可以由小鼠胚胎成纤维细胞与永生化小鼠胚胎成纤维细胞的原代培养支持。与其他的细胞共培养时,饲养层细胞通常为灭活的,通过辐射或用诸如丝裂霉素C的抗有丝解离剂处理以防止它们过大而不适于它们支持的细胞。不限制前述事项,特异的饲养层细胞类型可为人饲养层细胞,诸如人皮肤成纤维细胞或人胚胎干细胞。另一种饲养层细胞类型可为小鼠胚胎成纤维细胞(mEF)。如本文所用的,“无饲养层”(FF)环境指诸如细胞培养或培养基基本上无饲养层细胞的环境并且不受饲养层细胞培养的预调节。“预调节”培养基指饲养层细胞已在培养基内培养一段时间,诸如至少一天后收获的培养基。预调节培养基包含许多中介物质,诸如培养基中培养的饲养层细胞所分泌的生长因子与细胞因子。在本发明的一些实施方案中,无饲养层环境基本上没有人饲养层细胞并且在具体的实施方案中,额外地不受饲养层细胞预调节,人饲养层细胞包括但不限于人成纤维细胞、角质细胞以及胚胎干细胞。在本发明另外的实施方案中,无饲养层环境基本上没有动物饲养层细胞,并且进一步地,在具体的实施方案中,不受饲养层细胞预调节。在本发明的某些实施方案中,无饲养层环境基本上没有人与动物的饲养层细胞并且不受饲养层细胞预调节。基因组稳定性指细胞忠实地复制DNA并且保持DNA复制过程完整性的能力。如本文所用于描述本发明细胞的,“基因组稳定的细胞”与“具有基因组稳定性的细胞”指显示突变与染色体畸变(诸如易位、非整倍性、拷贝数变异以及重复)频率的细胞,其频率与正常人体细胞的突变与染色体畸变频率基本上相似。“成分”指可用于细胞培养基以保持和/或促进生长和/或细胞分化的任何化合物或其他的材料,无论是化学起源的还是生物起源的。术语“组分”“营养素”以及“成分”可互换地使用。细胞培养基所用的常规的成分可包括但不限于氨基酸、盐、金属、糖、脂质、核酸、激素、维生素、脂肪酸、蛋白等。如所期望的效果所需,通过本领域中技术人员选择在体外促进和/或保持细胞培养的其他成分。“分离”或“使分离”指从自然环境分离并且收集组合物或材料,诸如从组织或机体分离个体的细胞或细胞培养物。在一方面,细胞群或细胞组合物基本上没有与它的本质有关的细胞与材料。“分离的”或“纯化的”或“基本上纯的”,对靶细胞群而言,指至少大约50%,至少大约75%,至少大约85%,至少大约90%,并且在具体的实施方案中,至少大约95%纯的细胞群,对靶细胞组成全部细胞群而言。通过本领域中熟知的适当的方法可以评估细胞群或组合物的纯度。例如,基本上纯的多能细胞群指至少大约50%,至少大约75%,至少大约85%,至少大约90%,并且在具体的实施方案中,至少大约95%,以及在某些实施方案中,大约98%纯的细胞群,对靶细胞组成全部细胞群而言。在本文术语“本质上纯的”与“基本上纯的”可互换地使用。“传代”或“使……传代”指当细胞增生到所希望的程度时,细分并且将细胞平板接种到多个细胞培养表面或器皿的行为。在一些实施方案中,“传代”或“使……传代”指细分、稀释并且平板接种细胞。因为细胞从原代培养表面或器皿传代到随后的一套表面或器皿,因此随后的培养物可称为“传代培养”或“首次传代”等。每次细分并且平板接种到新的培养皿的行为被认为是一次传代。“平板接种”指将细胞或细胞们置于培养皿从而使得细胞粘附并且涂抹在细胞培养皿上。“多能性因子”指单独或与其他的制剂组合,能够增加细胞的发育能力的制剂。多能性因子包括但不限于多核苷酸、多肽以及能够增加细胞的发育能力的小分子。例如,示例性多能性因子包括转录因子与小分子重新编程剂。转录因子可指蛋白(即多肽)以及编码蛋白的多核苷酸,除非本文用法另有指示。例如,示例性转录因子包括Oct、Klf、Myc与Sox多肽,以及编码这些多肽的多核苷酸。本文提供了额外的转录因子的实例。如本文所用的,术语“多肽”与“蛋白”可互换地使用,除非有相反的规定,并且根据常规意义,即氨基酸序列。多肽不限于特定长度,例如它们可包含全长蛋白序列或全长蛋白的片段,并且可包括多肽的翻译后修饰,例如糖基化作用、乙酰化作用、磷酸化作用等,以及本领域中已知的其他的修饰,天然存在的与非天然存在的。使用任何各种熟知的重组和/或合成技术制备本发明的方法中所用的多肽。本发明的方法,在某些实施方案中,使用本文所述的多肽的活性片段(例如Sox-2、c-Myc、Oct3/4、Klf4、Lin28、Nanog等,或基质、辅因子和/或其下游效应物),例如包含至少大约10个、15个、20个、25个、50个、75个、100个、200个、300个、400个、500个、1000个等连续氨基酸或更多本文所述的多肽,包括所有中间的长度。在一具体的实施方案中,当用于本文所述的方法中时,使用的片段或片段组合保留了调节、诱导和/或保持多能性的能力。在其他的方面,本发明使用本文所述的多肽组合物(例如Sox-2、c-Myc、Oct3/4、Klf4、Lin28、Nanog等,或基质、辅因子和/或其下游效应物)的变体。本发明通常所包含的多肽变体,沿其长度方向,通常展示了与本文所示的多肽序列的至少约70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高同一性(例如,如下文所述所测定的)。在具体的实施方案中,使用的变体或其变体的组合保留了如本文所述的诱导多能性的能力。在其他方面,本发明使用多肽变体,沿其长度方向,展示了与本公开内容所用的野生型哺乳动物多肽的相应区域的至少约70%、75%、80%、85%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%或更高同一性(例如,如下文所述所测定的)。多肽变体在一种或多种取代、缺失、添加和/或插入方面不同于天然存在的多肽。此类变体可为天然存在的或可为合成产生的,例如,通过修饰一种或多种本发明方法中所用的以上的多肽序列并且使用本领域中熟知的许多技术的任一种评估它们的效果。“增生的”指一个细胞分成两个本质上相同的细胞或细胞群在数量上增加(例如以再生)的性能。“增殖”指在组织或机体外,例如在诸如塑料的(包被塑料的)细胞培养皿或烧瓶的无菌容器中使细胞生长(例如经由细胞增生再生)。“原代培养”指细胞、组织和/或培养物,其中分离的细胞与培养基置于初次培养皿中。细胞、组织和/或培养物可为保持的和/或可增生的,然而,只要细胞、组织和/或培养物依然在首次培养皿中,细胞、组织和/或培养物就称为原代培养。术语“小分子重新编程剂”或“小分子重新编程化合物”在本文中可互换地使用并且指能够单独或与其他的多能性因子组合增加细胞发育能力的小分子。“小分子”指具有分子量小于约5KD,小于约4KD,小于约3KD,小于约2KD,小于约1KD或小于约5KD的制剂。小分子可以为核酸、模拟肽、类肽、碳水化合物、脂质或其他有机的或无机的分子。化学的和/或生物的混合物的文库,诸如真菌、细菌、或海藻提取物,为本领域中已知的并且可用任何本发明的测定筛选。在具体的实施方案中,本文所用的小分子重新编程剂具有小于10000道尔顿,例如,小于8000道尔顿,6000道尔顿,4000道尔顿,2000道尔顿,例如50-1500道尔顿、500-1500道尔顿、200-2000道尔顿、500-5000道尔顿的分子量。如本文所用的,术语“基本上无”与“本质上无”可互换地使用,并且当用于描述组合物,诸如细胞群或培养基时,指无指定物质的组合物,诸如95%无、96%无、97%无、98%无、99%无指定物质或通过常规方法测定检测不到。相似的意义可以应用于术语“缺乏”,其中提及特定物质或组合物组分的缺乏。本发明中使用的细胞本发明中使用的起始细胞群可来自于基本上任何合适的来源,并且对细胞类型或多能性状态而言可为异源的或同源的。在一实施方案中,细胞为哺乳动物细胞,并且在具体的实施方案中,细胞从选自啮齿动物、猫、狗、猪、山羊、绵羊、马、牛或灵长类动物的哺乳动物分离出来。在某些实施方案中,哺乳动物为人。在其他的实施方案中,本发明待被使用或处理的细胞为成年细胞,包括基本上任何可得到的成年细胞类型。细胞可为体细胞的、非多能的、不完全地或部分多能的干细胞、专能细胞、寡聚多能细胞、单能细胞、终末分化细胞、或包含任何前述组合的混合细胞群。本发明的方法中所用的多能细胞可为天然存在的干细胞,包括胚胎干细胞、或可以为诱导性多能干细胞。“混合的”细胞群为发育能力不同程度的细胞群。例如,混合的细胞群可包含进行重新编程的细胞,从而使得混合群包含多能细胞、部分多能的细胞以及非多能细胞,诸如完全分化细胞。在一实施方案中,起始的细胞群选自成年的或新生的干/祖细胞。在具体的实施方案中,起始的干/祖细胞群选自中胚层干/祖细胞、内胚层干/祖细胞以及外胚层干/祖细胞。在其他的实施方案中,起始的干/祖细胞群为中胚层干/祖细胞。中胚层干/祖细胞的示例性实例包括但不限于中胚层干/祖细胞、内皮干/祖细胞、骨髓干/祖细胞、脐带干/祖细胞、脂肪组织来源的干/祖细胞、造血干/祖细胞(HSCs)、间充质干/祖细胞、肌肉干/祖细胞、肾脏干/祖细胞、成骨细胞干/祖细胞、软骨细胞干/祖细胞等。在其他有关的实施方案中,起始的干/祖细胞群为外胚层干/祖细胞。外胚层干/组细胞的示例性实例包括,但不限于神经干/祖细胞、视网膜干/祖细胞、皮肤干/祖细胞等。在其他有关的实施方案中,起始的干/组细胞群为内胚层干/祖细胞。内胚层干/祖细胞的示例性实例包括,但不限于肝脏干/祖细胞、胰腺干/祖细胞、上皮干/祖细胞等。在某些实施方案中,起始的细胞群可为异源的或同源的细胞群,选自胰岛细胞、CNS细胞、PNS细胞、心肌细胞、骨骼肌细胞、平滑肌细胞、造血细胞、骨细胞、肝细胞、脂肪细胞肾细胞、肺脏细胞、软骨细胞、皮肤细胞、卵泡细胞、血管细胞、上皮细胞、免疫细胞、内皮细胞等。诱导细胞的重新编程与增加细胞潜能继续在细胞中进行各种策略以诱导多能性或增加潜能(Takahashi,K.与Yamanaka,S.,Cell126,663-676(2006);Takahashi等,Cell131,861-872(2007);Yu等,Science318,1917-1920(2007);Zhou等,CellStemCell4,381-384(2009);Kim等,CellStemCell4,472-476(2009);Yamanaka等,2009;Saha,K.,Jaenisch,R.,CellStemCell5,584-595(2009)),并且提高重新编程的效率(Shi等,CellStemCell2,525-528(2008a);Shi等,CellStemCell3,568-574(2008b);Huangfu等,NatBiotechnol26,795-797(2008a);Huangfu等,NatBiotechnol26,1269-1275(2008b);Silva等,PlosBio6,e253.doi:10.1371/journal.pbio.0060253(2008);Lyssiotis等,PNAS106,8912-8917(2009);Ichida等,CellStemCell5,491-503(2009);Maherali,N.,Hochedlinger,K.,CurrBiol19,1718-1723(2009b);Esteban等,CellStemCell6,71-79(2010);Feng等,CellStemCell4,301-312(2009))。通常,用于重新编程的技术包括直接或间接地调节特异性细胞通路,使用多核苷酸-,多肽-和/或基于小分子的方法。例如,通过将细胞与一种或多种多能性因子接触增加细胞潜能。如本文所用的,“接触”可以包括在多能性因子(诸如,例如小分子、蛋白、肽等)存在的情况下培养细胞或将多能性因子引入到细胞。通过在包括诸如蛋白的转录因子的因子存在的情况下,在允许将转录因子引入到细胞的条件下,可以将多能性因子引入到细胞。例如,见ZhouH等,CellStemCell.2009May8;4(5):381-4;WO/2009/117439。例如,使用瞬态法,例如蛋白转导、显微注射、非整合基因传递、mRNA转导等或任何其他合适的技术可促进引入到细胞中。在一些实施方案中,通过已引入到细胞中的重组载体表达或通过在外源性转录因子多肽存在的情况下孵育细胞从而使得多肽进入细胞,将转录因子引入到细胞中。在具体的实施方案中,多能性因子为转录因子。与增加、确立、或保持细胞潜能有关的示例性转录因子包括但不限于Oct-3/4、Cdx-2、Gbx2、Gsh1、HesX1、HoxA10、HoxA11、HoxB1、Irx2、Isl1、Meis1、Meox2、Nanog、Nkx2.2、Onecut、Otx1、Oxt2、Pax5、Pax6、Pdx1、Tcf1、Tcf2、Zfhx1b、Klf-4、Atbf1、Esrrb、Gcnf、Jarid2、Jmjd1a、Jmjd2c、Klf-3、Klf-5、Mel-18、Myst3、Nac1、REST、Rex-1、Rybp、Sall4、Sall1、Tif1、YY1、Zeb2、Zfp281、Zfp57、Zic3、Coup-Tf1、Coup-Tf2、Bmi1、Rnf2、Mta1、Pias1、Pias2、Pias3、Piasy、Sox2、Lef1、Sox15、Sox6、Tcf-7、Tcf7l1、c-Myc、L-Myc、N-Myc、Hand1、Mad1、Mad3、Mad4、Mxi1、Myf5、Neurog2、ngn3、Olig2、Tcf3、Tcf4、Foxc1、Foxd3、BAF155、C/EBPβ、mafa、Eomes、Tbx-3;Rfx4、Stat3、Stella以及UTF-1。示例性转录因子包括Oct4、Sox2、Klf4、c-Myc以及Nanog。小分子重新编程剂也为多能性因子并且也可用于本发明的方法中以诱导重新编程并且保持或增加细胞潜能。在本发明的一些实施方案中,一种或多种小分子重新编程剂用于诱导体细胞的多能性,增加或保持细胞潜能或提高重新编程的效率。在一些实施方案中,小分子重新编程剂用于本发明的方法中以提高重新编程的效率。通过(1)重新编程与多能细胞产生所需要的时间的减少(例如,通过与没有小分子相似的或相同的过程相比,缩短了产生多能细胞的时间至少一天)或可选地或组合,(2)通过特定过程所产生的多能细胞数量的增加(例如,通过与没有小分子相似的或相同的过程相比,某个时间段内重新编程的细胞数量增加了至少10%、30%、50%、100%、200%、500%等)可以测量重新编程效率的提高。在一些实施方案中,观察到重新编程效率2倍或20倍的提高。在一些实施方案中,重新编程效率提高了超过20倍。在一些实施方案中,观察到超过没有小分子重新编程剂的方法多于100倍的效率(例如,产生的多能细胞的数量有超过100倍的增加)。几类小分子重新编程剂对增加、确立和/或保持细胞潜能是重要的。示例性小分子重新编程剂包括,但不限于抑制H3K9甲基化作用或促进H3K9脱甲基作用的制剂;抑制H3K4脱甲基作用或促进H3K4甲基化作用的制剂;抑制组蛋白脱乙酰作用或促进组蛋白乙酰化作用的制剂;L型钙通道激动剂;cAMP通路的激活剂;DNA甲基转移酶(DNMT)抑制剂;核受体配体;GSK3抑制剂;MEK抑制剂;TGFβ受体/ALK5抑制剂;HDAC抑制剂;Erk抑制剂;ROCK抑制剂;FGFR抑制剂;以及PARP抑制剂。示例性小分子重新编程剂包括GSK3抑制剂;MEK抑制剂;TGFβ受体/ALK5抑制剂;HDAC抑制剂;Erk抑制剂;以及ROCK抑制剂。这些类别的小分子剂每一个在下文有详细的描述。在本发明的一些实施方案中,小分子重新编程剂用于取代本发明的方法中一种或多种转录因子以诱导多能性,提高重新编程的效率,和/或增加或保持细胞潜能。例如,在一些实施方案中,细胞与一种或多种小分子重新编程剂接触,其中,包括足以提高重新编程的效率的剂量。在其他的实施方案中,除了转录因子以外,还在本发明的方法中使用一种或多种小分子重新编程剂。在一实施方案中,在条件下,细胞与至少一种多能性转录因子及至少一种小分子重新编程剂接触以增加、确立和/或保持细胞潜能或提高重新编程过程的效率。在其他的实施方案中,在条件下,细胞与至少一种多能性转录因子及至少两种、至少三种、至少四种、至少五种、至少六种、至少七种、至少八种、至少九种或至少十种小分子重新编程剂接触一段时期足以增加、确立和/或保持细胞潜能或提高重新编程过程的效率。通过监测本文别处所述的多能性特征可以评估多能性状态或细胞分化。在一实施方案中,细胞与包含一种或多种多能性因子的组合物和/或小分子重新编程剂的组合接触,其中多能性因子与小分子增加或诱导细胞的多能性。可以预期的是在体外、离体或在体内可接触本发明的细胞。表征多能细胞继诱导重新编程后,基于有关的与可检测的形态、与多能性有关的分子和/或生物化学变化可以选择重新编程的细胞。细胞多能性的特异特征,其在评估细胞潜能中可单独地或组合地监测,包括但不限于基因表达、甲基化作用以及体内与体外特征诸如:i)圆且扁的多能干细胞形态;ii)多能干细胞标志物的表达,其包括SSEA1(小鼠多能干细胞)、SSEA3/4(人多能干细胞);TRA1-60/81;TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD105、CD31、CD34、OCT4、Nanog和/或Sox2,以及如本发明提供的,CD30与CD50,以及前述事物的组合;iii)多能干细胞促进小鼠嵌合体中种系传递的能力;iv)使用四倍体补偿测定多能干细胞对胚体贡献的能力;v)多能干细胞的畸胎瘤形成;vi)胚体的形成与体外三系分化;以及vii)失活的X染色体重新激活。在某些实施方案中,任何以上的特征的亚型用于监测细胞多能性。在一实施方案中,多能细胞通过具有扁的、圆形集落形态,SSEA4与Oct4的表达,以及形成嵌合体与畸胎瘤的能力表征多能细胞。如本文所讨论的,多能性作为连续体存在并且诱导性多能干细胞似乎以“成熟的”状态与“原初的”状态存在,原初状态中的细胞可能具有更大的分化潜能。常规的培养基中产生的诱导性多能干细胞以成熟的状态存在并且更接近地类似来源于植入后囊胚的细胞,而原初iPSC显示了更接近地类似小鼠胚胎干细胞或来源于植入前囊胚的细胞的多能性特征。可以通过各种差异定义成熟的与原初的细胞状态,包括集落形态的差异、对抑制或激活关键信号转导的细胞反应、基因表达信号以及再激活与胚胎外细胞有关的基因的能力。例如,常规的iPSC,代表成熟的多能状态,展示了扁的集落形态,而原初iPSC展示了与小鼠胚胎干细胞相似的紧密半球形的集落形态。而且,常规的iPSC需要诸如TGFβ、激活素以及bFGF的关键细胞因子的外在信号转导,并且依赖于ERK/MEK细胞信号转导以保持未分化的状态,并且当例如通过细胞与TGFβ或MEK抑制剂接触来抑制这些通路时分化。相比之下,原初细胞不需要外在的信号转导,甚至当用TGFβ与MEK信号转导通路的抑制剂处理时,还保持多能性。额外地,基因表达分析揭示了原初的与成熟的多能细胞间的显著差异。例如,原初iPSC显著地抑制了Xist表达而常规的iPSC只显示了对Xist表达的适度抑制;原初细胞显示了显著的X染色体再激活并且位于X染色体上增加的基因表达,超过了常规的iPSC中所见的表达;原初细胞表达胚胎外干细胞标志物,包括但不限于Gata6、CDX2以及CGB。相比之下,早期的分化标志物,诸如谱系特异性基因,诸如Foxa2、Sox17以及Brachyury,超过了原初iPSC,在常规的iPSC中更高表达。额外的用于鉴定多能性的原初状态中的细胞的标志物包括Klf4、Tbx3、Gbx2、Lin28、Soc3中的增加或Otx2、Sox17、Cer1、FoxA2、Zic1、Lhx2、Xist中的减少。在本发明具体的实施方案中,原初细胞显示与常规的iPSC相比,Xist表达减少了至少两倍、至少五倍或至少十倍。在本发明的一些实施方案中,多能性的原初状态中细胞具有低于常规的iPSC两倍的Xist表达并且位于X染色体上至少五个基因的表达水平比常规的iPSC高三倍。X染色体再激活可以显示通过位于X染色体上的至少5个基因、至少10个基因、或在具体的实施方案中,至少100个基因的表达水平超过常规的iPSC中此类基因水平至少两倍、三倍、五倍或更多倍。在本发明的具体实施方案中,对多次细胞传代而言,例如对诸如至少1次、3次、5次、7次、10次、15次、20次或更多次的传代而言,本发明的多能细胞保留了多能性的特征。本发明的方法中使用的培养基平台本发明的培养基(即培养平台)包含化学定义的常备基础培养基与各种小分子的组合,包括小分子抑制剂,其允许:多能细胞在无饲养层环境中长期培养;细胞在无饲养层环境中重新编程;多能细胞的单细胞解离;多能细胞的细胞分选;未分化状态的保持;重新编程效率的提高;和原初多能细胞的产生。本发明的培养基中所用的化学定义的常备基础培养基可为任何定义的适用于支持干细胞的保持、生长和/或分化的基础培养基,诸如常规的人胚胎干细胞培养基。本发明所用的定义的基础培养基的实例包括,但不限于贝科改良Eagle培养基(“DMEM”)、Eagle基础培养基(BME)、DMEM/F-12(1:1DMEM与F-12体积:体积)、培养基199、F-12(Ham)营养混合物、F-10(Ham)营养混合物、最小必需培养基(MEM)、Williams′培养基E以及RPMI1640,除了别的以外,所有这些都可以从Gibco-BRL/Life技术公司,盖瑟堡,马里兰州购得。可使用许多这些培养基的几个版本,并且特别用于构建本发明的培养基的那些包括,但不限于DMEM11966、DMEM10314、MEM11095、Williams′培养基E12251、HamF1211059、αMEM12561以及培养基-19911151(都可从Gibco-BRL/Life技术公司购得(1995-1996目录))。例如培养基可包括下列的一种或多种:氨基酸、维生素、有机盐、无机盐、微量成分、缓冲盐、糖、ATP等(合适的基础培养基成分从圣路易斯,密苏里州的Sigma-Aldrich购得)。本发明的细胞培养基中所用的小分子及其种类在下文进行更加详细地描述。在具体的实施方案中,本发明的培养基包含一种或多种,两个或多个或三个或多个的TGFβ、GSK3抑制剂、MEK抑制剂以及ROCK抑制剂。在某些实施方案中,本发明的培养基包含TGFβ、GSK3抑制剂、MEK抑制剂以及ROCK抑制剂。本发明的细胞培养基与方法中所用的示例性TGFβ、GSK3抑制剂、MEK抑制剂以及ROCK抑制剂在下文进行了描述。培养基可额外地包含PARP抑制剂,诸如奥拉帕尼(AZD-2281)。GSK-3β抑制剂GSK-3β抑制剂包括,但不限于结合GSK-3β的抗体、显性阴性的GSK-3β变体与siRNA以及将GSK-3β作为靶标的反义核酸。其他示例性GSK-3β抑制剂包括但不限于Kenpaullone,1-Azakenpaullone,CHIR99021,CHIR98014,AR-A014418,CT99021,CT20026,SB216763,AR-A014418,锂,SB415286,TDZD-8,BIO,BIO-丙酮肟,(5-甲基-lH-吡唑-3-基)-(2-苯基喹唑啉-4-基)胺,吡啶咔唑-环青霉素二烯钌复合体,TDZD-84-苄基-2-甲基-l,2,4-t噻二唑嗪-3,5-二酮,2-硫代(3-碘苄基)-5-(l-吡啶基)-[l,3,4]-恶二唑,OTDZT,α-4-对溴溴代苯乙酮,AR-AO144-18,3-(l-(3-羟丙基)-lH-吡咯并[2,3-b]吡啶-3-基]-4-吡嗪-2-基-吡咯-2,5-二酮;TWSl19吡咯并嘧啶化合物,L803H-KEAPPAPPQSpP-NH2或它的十八烷基化形式;2-氯-l-(4,5-二溴-thiophen-2-基)-乙酮,SB216763,以及SB415286。本发明的细胞培养基中所用的示例性GSK3抑制剂包括CHIR99021,BIO以及Kenpaullone,而且CHIR99021在具体的实施方案中为优选的。ERK/MEK抑制剂示例性ERK/MEK通路抑制剂包括但不限于MEK或ERK抗体、显性阴性MEK或ERK变体、与siRNA以及抑制MEK和/或ERK表达的反义核酸。其他示例性ERK/MEK抑制剂包括但不限于PD0325901、PD98059、UO126、SL327、ARRY-162、PD184161、PD184352,舒尼替尼,索拉非尼,凡德他尼,帕唑帕尼,阿西替尼,GSKl120212、ARRY-438162、RO5126766、XL518、AZD8330、RDEAl19、AZD6244、FR180204以及PTK787。额外的ERK/MEK抑制剂包括国际公开的专利申请WO99/01426,WO02/06213,WO03/077914,WO05/051301以及WO2007/044084中公开的那些化合物。更多的示例性ERK/MEK抑制剂包括下列的化合物:--6-(4-溴-2-氯-苯胺)-7-氟-3-甲基-3H-苯并咪唑-e-5-羧酸(2,3-二羟基-丙氧基)-胺;6-(4-溴-2-氯-苯胺)-7-氟-3-(四氢-吡喃-2-ylm-乙基)-3H-苯并咪唑-5-羧酸(2-羟基-乙氧基)-胺,1-[6-(4-溴-2-氯-苯胺)-7-氟-3-甲基-3H-苯并咪唑-唑-5-基]-2-羟基-乙酮,6-(4-溴-2-氯-苯胺)-7-氟-3-甲基-3H-苯并咪唑-e-5-羧酸(2-羟基-1,1-二甲基-乙氧基)-胺,6-(4-溴-2-氯-苯胺)-7-氟-3-(四氢呋喃-2-ylm-乙基)-3H-苯并咪唑-5-羧酸(2-羟基-乙氧基)-胺,6-(4-溴-2-氟-苯胺)-7-氟-3-甲基-3H-苯并咪唑-e-5-羧酸(2-羟基-乙氧基)-胺,6-(2,4-二氯-苯胺)-7-氟-3-甲基-3H-苯并咪唑-5--羧酸(2-羟基-乙氧基)-胺,6-(4-溴-2-氯-苯胺)-7-氟-3-甲基-3H-苯并咪唑-e-5-羧酸(2-羟基-乙氧基)-胺,在下文中称为MEK抑制剂1;2-[(2-氟-4-碘苯基)胺]-N-(2-羟乙氧基)-1,5-二甲基-6--氧-1,6-二氢吡啶-3-羧胺;在下文中称为MEK抑制剂2;以及4-(4-溴-2-氟苯基胺)-N-(2-羟乙氧基)-1,5-二甲基-6--氧-1,6-二氢吡啶-3-羧胺或其药学上可接受的盐。在某些实施方案中,本发明的细胞培养基中所用的MEK/ERK抑制剂为PD98059。TGFβ受体/ALK5抑制剂示例性ALK5抑制剂包括ALK5抗体,ALK5的显性阴性变体,以及抑制ALK5表达的反义核酸。其他示例性ALK5抑制剂包括但不限于SB431542、A-83-01、2-(3-(6-甲基吡啶-2-基)-lH-吡唑-4-基)-l,5-萘啶、Wnt3a/BIO、BMP4、GW788388、SM16、IN-1130、GW6604、SB-505124以及嘧啶衍生物,例如参见,WO2008/006583,通过引用并入本文。另外,虽然“ALK5抑制剂”不打算包含非特异性激酶抑制剂,应理解“ALK5抑制剂”包含除了ALK5以外抑制ALK4和/或ALK7的抑制剂,例如,诸如SB-431542(例如参见Inman等,JMoI.Phamacol.62(1):65-74(2002)。鉴于本文显示抑制ALK5的效应的数据,应相信的是抑制TGFβ/激活剂通路具有相似的效应。因此,任何抑制剂,例如TGFβ/激活剂通路的上游或下游可以与ALK5组合或代替本文每段中所述的ALK5抑制剂使用。示例性TGFβ/激活剂通路包括但不限于:TGFβ受体抑制剂,SMAD2/3磷酸化作用的抑制剂,SMAD2/3与SMAD4相互作用的抑制剂,以及SMAD6与SMAD7激活剂/激动剂。而且,下文所述的分类仅仅出于组织的目的,并且本领域中技术人员应理解化合物可以影响一种或多种通路中的点,因此化合物可以一个以上所定义的分类发挥功能。TGFβ受体抑制剂可以包括以TGFβ受体为靶标的抗体、显性阴性变体与siRNA或反义核酸。抑制剂的特定实例包括但不限于SU5416;2-(5-苯并[l,3]二氧-5-基-2-叔丁基-3H-咪唑-4-基)-6-甲基吡啶氢氯化(SB-505124);lerdelimμMb(CAT-152);metelimμMab(CAT-192);GC-1008;IDl1;AP-12009;AP-11014;LY550410;LY580276;LY364947;LY2109761;SB-505124;SB-431542;SD-208;SM16;NPC-30345;Ki26894;SB-203580;SD-093;格列卫;3,5,7,2′,4′-戊羟基黄酮(莫林);激活素-M108A;P144;可溶的TBR2-Fc;以及以TGFβ受体为靶标的反义转染的肿瘤细胞。(例如参见Wrzesinski等,ClinicalCancerResearch13(18):5262-5270(2007);Kaminska等,ActaBiochimicaPolonica52(2):329-337(2005);以及Chang等,FrontiersinBioscience12:4393-4401(2007)。本发明的细胞培养基中所用的示例性TGFβ受体抑制剂包括SB431542,A-83-01以及RepSox。在具体的实施方案中,TGFβ抑制剂为SB431542。ROCK抑制剂ROCK为充当靶蛋白Rho(其中有三种亚型存在--RhoA,RhoB以及RhoC)的丝氨酸/苏氨酸激酶。示例性ROCK抑制剂包括但不限于ROCK抗体,显性阴性ROCK变体,与siRNA以及抑制ROCK表达的反义核酸。其他示例性ROCK抑制剂包括但不限于thiazovivin,Y27632、法舒地尔、AR122-86、Y27632H-1152、Y-30141、Wf-536、HA-1077、羟基-HA-1077、GSK269962A、SB-772077-B、N-(4-吡啶基)-N′-(2,4,6-三氯苯基)尿素、3-(4-吡啶基)-1H-吲哚以及(R)-(+)-反式-N-(4-吡啶基)-4-(1-氨乙基)-环己酰胺。本发明的细胞培养基中所用的示例性ROCK抑制剂包括thiazovivin,Y27632,pyrintegrin以及Blebbistatin。在某些实施方案中,ROCK抑制剂为thiazovivin。FGFR抑制剂示例性FGFR抑制剂包括但不限于FGFR抗体、显性阴性FGFR变体与siRNA以及抑制FGFR表达的反义核酸。其他示例性FGFR抑制剂包括但不限于RO-4396686、CHIR-258、PD173074、PD166866、ENK-834、ENK-835、SU5402、XL-999、SU6668、CHIR-258、R04383596以及BIBF-1120。PARP抑制剂PARP抑制剂抑制多聚二磷酸腺苷核糖聚合酶(“PARP”)。PARP蛋白为DNA修复酶,其发挥在细胞中调节DNA修复通路的功能。PARP参与剪辑切除修复(BER)通路,并且PARP抑制可促进多能细胞重新编程或保持期间细胞基因组的稳定性。本发明的细胞培养基中所用的示例性PARP抑制剂包括但不限于iniparib、veliparib以及奥拉帕尼(AZD-2281)。本发明的细胞培养基中小分子的量可以变化并且根据特定的培养条件优化,包括所用的特异性分子与组合,培养基中培养的细胞类型以及本发明的培养基所用的特异性应用。在一些实施方案中,小分子以足以诱导多能性、提高重新编程的效率或增加或保持细胞潜能的浓度存在于培养基中。在具体的实施方案中,本发明细胞培养基中小分子的优选浓度与组合显示于表1中。在本发明细胞培养基的具体实施方案中,细胞培养基为“SMC4”培养基,如表1中所示。SMC4培养基包含如表1中所示的常规的人ESC培养基与特异性通路调节剂以及添加剂。培养基组分以表1中所示此类组分的最佳范围的量存在于培养基中,并且以表1中所示的最佳浓度存在。SMC4培养基的实施方案可任选地包含任何一种或多种可选择的培养基以及表2中所示的通路调节剂与激活剂,以表2中所示的最佳范围内浓度,并且在某些实施方案中,以表2中所示的最佳浓度内浓度。在培养基的具体实施方案中,SMC4培养基包含可溶性纤连蛋白,并在本文被称为“SMC4+纤连蛋白”。在一些实施方案中,本发明的培养基还包含一种或多种Oct多肽、Klf多肽、Myc多肽以及Sox多肽。在一些实施方案中,培养基不包含细胞。在一些实施方案中,培养基还包含细胞,例如,非多能细胞、部分多能细胞、多能细胞或包含各种潜能状态的细胞的混合的细胞群。表1.细胞培养组分与添加剂。下列的表格列出了分子的实例,以及它们影响的可以用于增强重新编程过程以及保持未分化的状态中的多能干细胞信号转导通路。SMC4+纤连蛋白是指以约5μg/mL的浓度添加可溶性纤连蛋白的上文所述的SMC4培养基。纤连蛋白可以以约0.05-500μg/mL的浓度存在于SMC4培养基中。表2.具体可选择的细胞培养组分与添加剂以增强在单细胞传代5上、细胞分选以及无饲养层系统上培养细胞的多能性与活力细胞因子与生长因子在本发明的一些实施方案中,本发明的细胞培养基基本上无细胞因子和/或生长因子,并且任选地为无饲养层环境。在其他的实施方案中,细胞培养基包含添加物,诸如血清、提取物、生长因子、激素、细胞因子等。各种生长因子与它们在培养基中的使用为本领域中熟知的并且例如,包括ECM蛋白、层粘连蛋白1、纤连蛋白、IV型胶原蛋白同种型、蛋白酶抑制剂、细胞表面粘附蛋白、细胞信号转导蛋白、钙粘素、胞内氯离子通道蛋白1、跨膜受体PTK7、胰岛素样生长因子、抑制素βA、TGFβ/激活素/nodal信号转导通路的诱导物以及激活素A。例如,培养基中所用的细胞因子可包括下列的一种或多种:生长因子诸如表皮生长因子(EGF)、酸性成纤维细胞生长因子(aFGF)、碱性成纤维细胞生长因子(bFGF)、肝细胞生长因子(HGF)、胰岛素样生长因子1(IGF-1)、胰岛素样生长因子2(IGF-2)、角质细胞生长因子(KGF)、神经生长因子(NGF)、血小板源性生长因子(PDGF)、转化生长因子β(TGF-β)、血管内皮细胞生长因子(VEGF)、转铁蛋白、各种白细胞介素(诸如IL-1至IL-18)、各种集落刺激因子(诸如粒细胞/巨噬细胞集落刺激因子(GM-CSF))、各种干扰素(诸如γ干扰素)以及其他具有对干细胞有影响的细胞因子,诸如干细胞因子(SCF)与红细胞生成素(Epo)。这些细胞因子可商购获得,例如从R&D系统、明尼阿波利斯、明尼苏达州商购,并且可为天然的或重组的。在一些实施方案中,对各种各样的哺乳动物细胞的培养而言,基础培养基将包含浓度约0.01-100ng/ml、约0.2-20ng/ml,并且在具体的实施方案中约0.5-10ng/ml的FGF。其他的细胞因子,如果使用的话,将以凭经验确定的或如通过已确立的细胞因子技术指导的浓度添加。本发明的培养基中可包括的额外的组分为胰岛素(特别是胰岛素Zn++)与转铁蛋白。这些额外的成分,商购获得(例如从Sigma-Aldrich,圣路易斯,密苏里州商购),可以约0.1-约100μg/ml或约1μg/ml-约10μg/ml的浓度范围配制到本发明的培养基中。额外地,重组胰岛素或胰岛素基本锌的盐可被替换为动物源性或人源性的胰岛素。可添加其他的成分或替代品至本领域中普通技术人员已知的那些添加组分。细胞因子与类似的添加物组分可替换(或另外)被包括在基础培养基中。此类组分通常与添加组分包括在一起,因为添加组分常规储存在约-20℃下而不是常规用于储存基础培养基的约4℃温度。细胞因子与类似的组分在更接近-20℃的温度下一般更好。本发明的方法中所用的基质任何合适的器皿或细胞培养皿都可用作基础培养基中的细胞培养物和/或细胞培养添加物的支持物。支持物上无基质包被为必需的。然而,用粘附促进基质(例如胶原蛋白、纤连蛋白、包含RGD的多肽、明胶等)包被培养皿的表面促进细胞附着并且因此增强细胞培养基与本文公开的添加物的影响。用于培养并且传代细胞的合适的基质为本领域中已知的并且包括但不限于明胶、层粘连蛋白、纤连蛋白、胶原蛋白、弹性蛋白、骨桥蛋白、诸如MatrigelTM的天然存在的细胞系产生的矩阵混合物以及合成的或人造的表面,诸如聚胺单层与羧基末端的单层。无饲养层环境虽然细胞通常培养在饲养层细胞上或在用饲养层细胞预调节并且包含胎牛血清的培养环境中,但此类环境不适合于产生用于临床与治疗用途的细胞。例如,通常认为在此类外源污染的环境中培养的细胞不适合于人的细胞移植,因为动物组分的暴露可存在免疫排斥与将未鉴定的病原体传递至被治疗患者的严重风险,并且可潜在地再激活动物的逆转录病毒。使用无动物培养基的培养系统,诸如本发明的无饲养层环境,促进临床级细胞系,特别是hESC与iPSC细胞系的产生。在本发明的一些实施方案中,本发明的无饲养层环境基本上无人饲养层细胞,包括但不限于人成纤维细胞、角质细胞以及胚胎干细胞,并且不受饲养层细胞预调节。在本发明另外的实施方案中,无饲养层环境基本上无动物饲养层细胞,另外,在一些实施方案中,无饲养层环境没有用饲养层细胞预调节。在本发明具体的实施方案中,无饲养层环境基本上无人的与动物的饲养层细胞,并且在更具体的实施方案中,无饲养层环境基本上无人的与动物的饲养层细胞并且没有用饲养层细胞预调节。本发明的无饲养层细胞培养基用于本发明的方法中,包括多能细胞的培养,细胞的重新编程,多能细胞的单细胞解离,多能细胞的细胞分选,原初多能细胞的产生以及细胞未分化状态的保持。在本发明的具体方法中,无饲养层环境用于诱导多能性、提高重新编程的效率,和/或增加或保持细胞潜能的方法中。在某些实施方案中,无饲养层环境基本上没有细胞因子与生长因子,包括bFGF。解离通过酶促的或机械的方法可以实现细胞解离为单细胞,诸如解离为单细胞悬浮液。本领域中已知的允许细胞解离为单细胞的任何酶促剂可用于本发明的方法中。在本发明的一实施方案中,解离剂选自胰蛋白酶/EDTA、TrypLE选择剂、IV型胶原蛋白酶以及中性蛋白酶。螯合剂,诸如EDTA、Accutase或AccμMax,也可单独地或与酶促剂组合,用于本发明的方法中解离细胞。解离剂可溶于无钙与镁的PBS以促进解离为单细胞。为增强解离期间与解离后细胞的存活,可以添加生存促进物质(例如生长因子、参与细胞死亡与凋亡的细胞通路的抑制剂、或条件培养基)。在一些实施方案中,解离常规培养基中培养的细胞并且将单细胞置于具有一种或多种小分子抑制剂的本发明的细胞培养中,诸如SMC4培养基或SMC4+纤连蛋白。可任选地将解离的单细胞置于无饲养层环境中。在其他的实施方案中,解离前在无饲养层环境中培养细胞并且置于具有一种或多种小分子抑制剂的本发明的细胞培养中,诸如SMC4培养基或SMC4+纤连蛋白,其可任选地为无饲养层环境。通过机械力支持酶促解离为单细胞。可选地,解离剂可仅为机械力,诸如通过使用机械工具,诸如移液管或尖头的微毛细管以分离细胞。细胞培养与培养基收集中的通用技术在以下中概述:大规模的哺乳动物细胞培养(Hu等,Curr.Opin.Biotechnol.8:148,1997);无血清培养基(K.Kitano,Biotechnology17:73,1991);大规模的哺乳动物细胞培养(Curr.Opin.Biotechnol.2:375,1991);以及哺乳动物细胞的悬浮培养(Birch等,BioprocessTechnol.19:251,1990)。其他感兴趣的读物包括理解培养基(M.McLuhan,MentorN.Y.,1964)与培养基即信息(M.McLuhan&Q.Fiore,BantamN.Y.,1967)。为详细阐述本发明实践中所用的通用技术,实践者可以参考细胞生物学、组织培养以及胚胎学方面标准的教科书与综述。包括:“畸胎癌与胚胎干细胞:实践的方法”(E.J.Robertson,ed.,IRLPressLtd.1987);“小鼠发育中的技术指南”(P.M.Wasserman等.eds.,AcademicPress1993);“胚胎干细胞体外分化”(M.V.Wiles,Meth.Enzymol.225:900,1993);“胚胎干细胞的特性与用途”(P.D.Rathjen等,al.,1993)。Robertson,Meth.CellBiol.75:173,1997与Pedersen,Reprod.Fertil.Dev.10:31,1998中回顾了干细胞的分化。富集与消耗策略本发明也提供了用于富集多能细胞的细胞群的策略,作为增加产生iPSC效率的方法。富集策略提供了在相对短的时间内衍生克隆的iPSC集落的方法,提高了iPSC产生效率。本发明的富集方法包括分选细胞群,已诱导其重新编程以鉴定及获得表达多能性标志物的细胞,因此获得富集的多能细胞的细胞群。待被分选的细胞已被诱导重新编程并且可包含进行重新编程的混合的细胞群,从而使得群体包含多能细胞、部分多能细胞以及非多能细胞,诸如终末分化细胞。在一实施方案中,待被分选的细胞群已被诱导重新编程并且表达多能性标志物。在一些实施方案中,诱导重新编程约4天-30天、约4天-24天、约6天-22天、或约8天-约12天后培养细胞。额外的富集方法涉及表达分化标志物或非多能性细胞的消耗以获得富集的多能细胞群。本发明的富集策略包括获得待被分选的细胞群的单细胞悬浮液。在本发明的一实施方案中,通过解离群体中的细胞并且重悬浮细胞获得单细胞悬浮液。解离的细胞可重悬浮在任何合适的用于保持细胞或进行细胞分选的溶液或培养基。在具体的实施方案中,单细胞悬浮液包含一种或多种TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂。在某些实施方案中,单细胞悬浮液包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂,以及在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在本发明的富集过程中,分选细胞以获得多能细胞或消耗非重新编程的或非多能细胞,因此获得富集的多能细胞群。在一实施方案中,制备单细胞悬浮液,然后制备用于分选的单细胞,诸如通过多能性标志物染色,例如使用适当的抗体。可通过任何合适的分选细胞的方法,诸如通过磁珠或流式细胞术(FACS)分选,分选细胞。基于各种多能性标志物分选细胞,包括Oct、Sox、Nanog、SSEA3/4、TRA1-60/81、TRA1-85、TRA2-54、GCTM-2、TG343、TG30、CD9、CD29、CD133/prominin、CD140a、CD56、CD73、CD105、CD31、CD34、OCT4、KLF4、SSEA1(小鼠)以及如本发明中所示的CD30与CD50。在各种实施方案中,基于至少两种、至少三种、或至少四个多能性标志物分选细胞。在某些实施方案中,基于SSEA4的表达,并且在某些具体的实施方案中,基于SSEA4与TRA181或TRA160组合的表达分选细胞。在某些实施方案中,基于SSEA4、Tra181或TRA160以及CD30分选细胞。在某些实施方案中,最初,使用分化细胞的表面标志物,其可包括但不限于CD13、CD26、CD34、CD45、CD31、CD46或CD7,针对非重新编程的细胞消耗细胞,然后针对多能性标志物诸如SSEA4、Tra181以及CD30富集细胞。分选以获得针对多能性标志物阳性的细胞后,理想的细胞分数为富含多能细胞的细胞群。可将富含多能细胞的群体置于细胞培养系统中,诸如常规的hESC培养基或本发明的细胞培养基。细胞培养系统可添加饲养层细胞或任选地为无饲养层环境。在一些实施方案中,将表达多能性标志物的分选的细胞置于添加了饲养层细胞的培养系统且然后转移至无饲养层环境。细胞培养系统可添加一种或多种表1与2中所示的特异性通路调节剂与添加剂。在一实施方案中,细胞培养基为无饲养层环境并且包含TFGβ抑制剂、GSK3抑制剂、MEK抑制剂以及Rock抑制剂的其中之一;并且在某些实施方案中,TFGβ抑制剂为SB431542,GSK3抑制剂为CHIR99021,MEK抑制剂为PD0325901,以及Rock抑制剂为thiazovivin。在本发明其他具体的实施方案中,细胞培养系统为包含MatrigelTM包被的组织平板、常规的hESC培养基以及表1中所示的特异性通路调节剂。在一实施方案中,细胞培养系统包含表1中所述的SMC4培养基,任选地与任何可选的表2中所示的培养基与通路调节剂组合。富集的细胞群可培养于本文所述的细胞培养系统中以获得iPSC集落,iPSC集落通常出现在分选后约3天-约25天;分选后约5-9天或分选后约5-7天。可以选择或分选iPSC集落用于克隆扩增。使用本发明的富集策略,富集3倍多能细胞的细胞群。本发明也提供了消耗不合需要的细胞的细胞群的方法。在一些实施方案中,细胞群,诸如进行重新编程的细胞群或多能细胞群,消耗分化细胞。在本发明的方法中,被诱导重新编程的多能细胞群或细胞可以消耗具有分化细胞的细胞表面标志物的细胞。基于分化细胞的表面标志物,诸如CD13、CD26、CD34、CD45、CD31、CD46、或CD7可以分选细胞群,并且可以从细胞群移除表达分化细胞的标志物的细胞以获得富含多能细胞的细胞群。在本发明的具体实施方案中CD13用作分化细胞的表面标志物。在其他的实施方案中,诱导以分化的细胞群,诸如诱导以分化为所需谱系的细胞群,消耗多能细胞以获得分化群或分化细胞。在一些实施方案中,分化细胞群包含已诱导其分化为特定谱系的细胞群,诸如ESC或iPSC。使用上文所述的分选技术,诸如根据磁珠或基于多能性标志物的FACs分选群体中的细胞,细胞群可消耗多能细胞。在一些实施方案中,通过FACs使用多能性标志物,分选包含分化细胞的细胞群并且获得消耗了表达多能性标志物的细胞的一小部分。在其他的实施方案中,通过FACs基于分化标志物,诸如谱系特异性标志物,像CD13、CD26、CD34、CD45、CD31、CD46、或CD7分选细胞群以获得消耗了多能性标志物的一小部分。在本发明的具体实施方案中CD13用作分化细胞的表面标志物。本发明实践将使用,除非明确指示意思相反,化学、生物化学、有机化学、分子生物学、微生物学的常规方法,重组DNA技术、遗传学、免疫学、细胞生物学、干细胞技术,细胞培养与转基因生物学,它们都在技术人员掌握范围内。出于示例目的下文描述了其中许多。在文献中充分地解释此类技术,例如参见Sambrook等,分子克隆:实验室手册(第3版,2001年);Sambrook等,分子克隆:实验室手册(第2版,1989年);Maniatis等,分子克隆:实验室手册(1982年);Ausubel等,当前的分子生物学技术(JohnWiley与Sons,2008年7月升级);分子生物学中简短技术:来源于当前分子生物学技术的方法概略,GreenePub.AssociatesandWiley-interscience;Glover,DNA克隆:实践方法,I&II卷(IRLPress,Oxford,1985年);Anand,用于复合基因组分析的技术(学术出版社,纽约,1992年);Guthrie与Fink,酵母遗传学与分子生物学指南(学术出版社,纽约,1991年);寡核苷酸合成(N.Gait,Ed.,1984年);核酸杂交(B.Hames&S.Higgins,Eds.,1985年);转录与转译(B.Hames&S.Higgins,Eds.,1984年);动物细胞培养(R.Freshney,Ed.,1986年);Perbal,分子克隆的实践指南(1984年);Fire等,RNA干扰技术:从基础科学到药物研发(剑桥大学出版社,剑桥,2005年);Schepers,实践中RNA干扰(Wiley-VCH,2005年);Engelke,RNA干扰(RNAi):siRNA技术的螺母与螺栓(DNA出版社,2003年);Gott,RNA干扰,编辑与修正:方法与技术(分子生物学中的方法;人出版社,托托瓦,新泽西州,2004年);Sohail,受RNA干扰的基因沉默:技术与应用(CRC,2004年);Clarke与Sanseau,微RNA:生物学,功能&表达(螺母&螺栓系列;DNA出版社,2006年);固定化细胞与酶(IRL出版社,1986年);论述,酶学中的方法(学术出版社,有限公司,纽约);用于哺乳动物细胞的基因转移载体(J.H.Miller与M.P.Caloseds.,1987年,冷泉港实验室);Harlow与Lane,抗体(冷泉港实验室出版社,冷泉港,纽约,1998年);细胞与分子生物学中的免疫化学方法(Mayer与Walker,eds.,学术出版社,伦敦,1987年);实验免疫学手册,I-IV卷(D.M.Weir与CCBlackwell,eds.,1986年);Riott,基础免疫学,第6版,(Blackwell科学出版社,牛津,1988年);胚胎干细胞:方法与技术(分子生物学中的方法)(KurstadTurksen,Ed.,2002年);胚胎干细胞技术:I卷:分离与表征(分子生物学中的方法)(KurstadTurksen,Ed.,2006年);胚胎干细胞技术:II卷:分化模型(分子生物学中的方法)(KurstadTurksen,Ed.,2006年);人胚胎干细胞技术(分子生物学中的方法)(KursadTurksenEd.,2006);间充质干细胞:方法与技术(分子生物学中的方法)(DarwinJ.Prockop,DonaldG.Phinney,以及BruceA.BunnellEds.,2008年);造血干细胞技术(分子医学中的方法)(ChristopherA.Klug以及CraigT.JordanEds.,2001年);造血干细胞技术(分子医学中的方法)(KevinD.BuntingEd.,2008年)神经干细胞:方法与技术(分子生物学中的方法)(LeslieP.WeinerEd.,2008年);Hogan等,操作小鼠胚胎的方法(第2版,1994年);Nagy等,操作小鼠胚胎的方法(第3版,2002年),以及斑马鱼书,斑马鱼使用的实验室指南(斑马鱼),第4版,(俄勒冈大学出版社,尤金,俄勒冈州,2000年)。如本说明书与所附的权利要求中所用的,单数形式“一(a)”,“一(an)”以及“所述(the)”包括复数参考,除非内容另有明确地规定。贯穿本说明书,除非上下文另有要求,应理解词语“包含(comprise)”,“包含(comprises)”以及“包含(comprising)”暗示规定的步骤或成分或步骤或成分的组的包含,但是不排除任何其他的步骤或成分或步骤或成分的组。所谓“由…组成(consistingof)”意指包括并且限于跟随在短语“consistingof”无论什么。因此,短语“由…组成(consistingof)”指示所列成分是必须的或强制的,并且没有其他的成分存在。所谓“基本上由…组成(consistingessentiallyof)”意指包括短语后所列的任何成分,并且限于不干扰或有助于活性或所列成分公开内容中指定的活动其他的成分。因此,短语“基本上由…组成(consistingessentiallyof)”指示所列成分是必须的或强制的,但是其他的成分为任选的以及可能或不可能存在,取决于它们是否影响所列成分的活性或活动。可以组合上文所述的各种实施方案,以便提供其他的实施方案。通过引用,全文并入本说明书所涉及和/或申请资料页所列举的所有美国专利、美国专利申请公布、美国专利申请、国外的专利、国外的专利申请和非专利出版物。如有必要,可更改实施方案的各种方面,以便应用各种专利、申请和出版物的概念再提供其他的实施方案。根据上文的详述,可对实施方案做出这些或其他改变。总之,在所附的权利要求中,所用的术语不应当解释为,限制说明书和权利要求中所公开的具体实施方案,而应当解释为包括所有可能的实施方案,以及授予权利的此类权利要求的等同物的全部范围。因此,权利要求并不限于公开的内容。实施例实施例1:实验方法A.多能干细胞的细胞培养适应无饲养层前,将人诱导性多能干细胞(hiPSC)保持在饲养层细胞(丝裂霉素C处理的小鼠胚胎成纤维细胞(MEF)细胞(Millipore))上,并且与常规的培养基一起培养。如本申请中所用的,“常规的培养基”指包含DMEM/F12(Mediatech),10ng/mLbFGF(Invitrogen),20%v/v基因敲除的血清置换(Invitrogen),1%v/v非必需氨基酸(Mediatech),2mM左旋谷氨酰胺(Mediatech)以及100μMβ-巯基乙醇(Invitrogen)的基础的人胚胎干细胞(hESC)培养基。表1的第一部分中也描述了常规的培养基。使用精尖的玻璃吸管,通过机械地切开将集落解体为小块每5-7天传代(团块传代)hiPSC,收集并以1:3-1:6稀释传代至新近接种的丝裂霉素C处理的MEF细胞上,每日添加hESC培养基。将细胞培养保持在设置成37℃、5%CO2湿润的孵育箱内。在具有饲养层细胞的常规培养基中培养细胞并且使用团块传代在本文被称为“常规的培养”。对单细胞解离而言,用磷酸盐缓冲盐水(PBS)(Mediatech)洗涤hiPSC一次并且在37℃下用Accutase(Millipore)或TrypL(Invitrogen)处理3-5分钟随后通过移液管打破成单细胞。然后以等体积将单细胞悬浮液与上文所述的常规培养基混合,以300g旋转沉降5分钟并且重悬浮在SCM4培养基或SMC4+纤连蛋白培养基。在大多数情况下,将单细胞解离的细胞保持在SMC4培养基,SMC4培养基由添加了包括0.4μMPD0325901、1μMCHIR99021、5μMThiazovivin以及2μMSB431542(所有Biovision)的各种小分子与添加剂的常规hESC培养基组成。20℃下,在添加至培养基前将小分子以储备浓度5-25μM保持DMSO中。将所有的工作培养基保持在4℃下长达4周。ROCK抑制剂的培养,与Thiazovivin一起优于Y27632用于保持细胞在未分化的状态。重悬浮在适当的培养基中后,将细胞转移到预先在37℃下用MatrigelTM(1:25稀释;BDBiosciences)包被1-2小时的无饲养层组织培养平板(BDFalcon)。在这种格式中,每隔一天细胞常规地接收新鲜的培养基并且当融合达到66-75%时传代,这通常发生在传代后4-5天。随着每一传代,将细胞再解离为单细胞并且以1:5-1:10的稀释传代转移至新的MatrigelTM(BDBiosciences)包被的组织培养平板。对于界定,无生长因子培养,将细胞悬浮液添加至预先用1%明胶(Mediatech)包被的组织培养平板。如上文所述的保持并且传代细胞,除了SMC4培养基基本上无所有的细胞因子与生长因子,包括bFGF之外。出于冷冻的目的,将细胞解离为单细胞,重悬浮于添加了10%v/vDMSO(Mediatech)的SMC4+纤连蛋白中并且置于冷冻管(Nalgene)内。一旦盖上盖子,就将冷冻管置于Mr.Frosty(Nalgene)内并且在-80℃下保持过夜。第二天将冷冻管转移至液氮以长期储存。为解冻,将冷冻管置于37℃水浴大约1-2分钟,直到大多数的冰融化。然后将解冻的细胞溶液与新鲜的常规hESC培养基轻微地混合并且以300g旋转沉降5分钟。将细胞溶液重悬浮在SMC4+纤连蛋白培养基中并且转移至MatrigelTM(BDBiosciences)包被的组织培养平板。随着与所有其他的细胞培养孵育,细胞保持在设置成37℃与5%CO2的湿润孵育箱中。B.重新编程的诱导为启动重新编程过程,使用慢病毒转导或其他的方法诸如只有蛋白处理获得重新编程因子(人Oct4、Sox2、Klf4、c-Myc、Lin28以及Nanog的可变组合)的异位表达。在大多数的情况下,将起始细胞以10%融合(即1x105个细胞/6孔平板的孔)接种至明胶(Mediatech)包被的表面上。对病毒感染的方法而言,以1:2的稀释将新近收集的慢病毒添加至起始细胞,添加4μg/mL聚凝胺(Millipore),在32℃下以650g旋转感染1.5小时。将培养物转移至37℃与5%CO2下再保持7小时。孵育完成后,用PBS洗涤细胞三次并且用新鲜的培养基供给。难以感染细胞,诸如无饲养层培养系统中的IMR90成纤维细胞,初始感染后48小时再重复一次这个过程。对诱导重新编程的非遗传方法诸如直接的蛋白应用至细胞的使用而言,将蛋白混合物或由重新编程因子组成的混合物以8μg/mL添加至细胞溶液并且在培养基变化前保持24小时。重复这一步骤额外的2-4次。所有起始细胞都培养在它们自己各自的体细胞培养基直到第4天后添加起始的蛋白,在这一点上,培养基转换为一部分体细胞培养基与一部分常规的hESC培养基。根据融合(通常在4-6天)使细胞胰蛋白酶化,与相等部分的培养基混合,以300g旋转沉降5分钟,重悬浮在1:1体细胞的/常规的hESC培养基中并且以1:4-1:6扩增至更大的培养平板。例如,6孔平板的两个孔中的细胞通常扩增到10cm皿上。扩增后第二天,培养基完全转换为常规的hESC培养基。一旦扩增的细胞达到融合(通常在8-12天),它们将进行富集(参见独特的群富集)。在所有的情况下,每隔一天常规地改变培养基。C.独特的群富集在用各种策略,包括个体的慢病毒构建体或包含Oct4和/或Klf4和/或Sox2和/或Myc的多顺反子性载体诱导起始细胞重新编程并且培养大约8-12天(参见上文)后,细胞解离为单细胞(参见多能干细胞的细胞培养)并且用各种多能性的表面标志物、体细胞的标志物和/或不完全的重新编程的标志物将其染色。简而言之,解离的细胞重悬浮在包含汉克斯平衡盐溶液(Invitrogen)、4%胎牛血清(Invitrogen)以及10mMHepes(Invitrogen)的染色溶液中并且保持在冰上。按照制造商推荐的稀释,将结合的第一抗体添加至细胞溶液并且在冰上孵育溶液15分钟。洗涤细胞溶液,重悬浮在染色缓冲液中并且保持在冰上。在这一点上,采用各种富集/消耗策略,包括荧光激活的细胞分选(BDBiosciences,参见下文)与磁珠细胞分选(MiltenyiBiotec,参见下文)。在FACSAria(BD,Biosciences)上进行流式细胞术分选。按照说明,所用的第一抗体包括SSEA4(BDBiosciences)、Tra-181(Biosciences)、Tra-161(BDBiosciences)、CD30(BDBiosciences)以及CD50(BDBiosciences)。然后旋转沉降分选的细胞并且重悬浮在SMC4+纤连蛋白培养基中,转移至MatrigelTM包被的组织培养平板。当分选至微孔,即96孔平板时,以300g旋转沉降平板2分钟。每隔一天替换SMC4+纤连蛋白培养基持续3-4天。3-4天后,在培养剩余的时间里SMC4+纤连蛋白培养基通常被SMC4培养基取代。通常分选后7-9天可见到集落形成。在GuavaEasyCyte8HT(Millipore)上进行流式细胞术分析。根据实验方案进行MACS微珠(MiltenyiBiotec)分离。简单地说,细胞解离为单细胞(参见多能干细胞的细胞培养)并且用适当的FITC-结合的第一抗体,按照说明,包括SSEA4(BDBiosciences)、Tra-181(Biosciences)、Tra-161(BDBiosciences)、CD30(BDBiosciences)以及CD50(BDBiosciences)。然后细胞被抗FITC微珠(MiltenyiBiotec)磁性标记。然后将标记的细胞悬浮液装载到LSMACS柱(MiltenyiBiotec)上。以300g旋转沉降从阳性或阴性选择的部分收集的细胞5分钟并且重悬浮在SMC4+纤连蛋白中,转移至MatrigelTM(BDBiosciences)包被的组织培养平板。翌日,将新鲜的培养基添加至培养物并且随后每隔一天替换。3-4天后,在培养剩余的时间里SMC4+纤连蛋白培养基通常被SMC4培养基取代。通常分选后5-7天出现集落。D.碱性磷酸酶染色在4%v/v多聚甲醛(AlfaAesar)中固定细胞30秒,用PBS洗涤三次并且用碱性磷酸酶染色试剂盒染色(Sigma-Aldrich)。简单地说,将1mL亚硝酸钠溶液添加至1mLFRV-碱性溶液,混合并且在25℃下孵育2分钟。然后将溶液与45mLH2O混合,随后添加1mL萘酚AS-BI碱性溶液。将碱性染色混合物添加至固定的细胞并且在25℃下孵育15分钟随后用PBS洗涤。然后在碱性磷酸酶存在的情况下给细胞计分。E.免疫荧光染色在4%v/v多聚甲醛(AlfaAesar)中固定细胞15分钟,用包含0.2%v/v吐温(PBST)(FisherScientific)洗涤三次并且在25℃下使用0.15%v/vTritonX-100(Sigma-Aldrich)在PBS中渗透1小时。透化作用后,在25℃下用1%v/v牛血清白蛋白(BSA)(Invitrogen)的PBST(PBSTB)(FisherScientific)封闭细胞30分钟。PBSTB温和去除后,在4℃下将细胞与第一抗体在PBSTB中孵育过夜。本研究中所用的第一抗体包括Nanog(Abcam)、Tra-1-60(BDBiosciences)、Tra-181(BDBiosciences)、SSEA4(BDBiosciences)、β-III微管蛋白(R&DSystems)、α-平滑肌肌动蛋白(Sigma)以及Sox17(R&DSystems)。过夜孵育后,用PBST洗涤细胞三次并且在25℃下,以1:200稀释在PBSTB中用第二抗体(Alexa488or555;Invitrogen)染色1小时。在PBST中洗涤细胞三次并且用Hoechst染料(Invitrogen)染色。使用荧光显微镜或CCD照相机捕获染色细胞的图像。F.分化诱导与畸胎瘤形成无饲养层iPSC分化为单层与拟胚体。对单层分化而言,转变分化培养基之前允许iPSC达到接近融合,因为一旦分化细胞通常减少它们的增生。简单地说,一旦融合,SMC4培养基转变为包含DMEM/F12(Mediatech)、20%胎牛血清(Invitrogen)、1%非必需氨基酸(Mediatech)、2mM左旋谷氨酰(Mediatech)以及100μMβ-巯基乙醇的分化培养基。一旦培养基转变,就允许iPSC分化14天。每2-3天改变培养基。对胚体(“EB”)形成与分化而言,hiPSC为用Accutase(Millipore)解离的单细胞并且以最终的浓度75,000个细胞/mL重悬浮在分化培养基中,添加5μMThiazovivan。细胞以100μL/孔接种于V底96孔非组织培养平板(Nunc)并且以950g用离心机分离5分钟。第二天,使用P1000以大约30-40EBs/孔将紧密的“球样团块”转移至超低结合的6孔平板(Corning)。7天后,以1:1转移转移EBs至Matrigel包被的6孔平板。培养3周后,将细胞固定并且染色。通过应用的干细胞(MenloPark,CA)进行畸胎瘤抑制与分析。简单地说,将1000,000-2000,000个单细胞解离的hiPSC混合在添加了培养基与100μLMatrigel的100μLSMC4培养基中并且引入至肾被膜与浅褐色SCID小鼠的睾丸。收获发育的畸胎瘤,将其切片并且分析各种分化的细胞类型与结构。G.RT-qPCR与qPCR分析使用PicoPureRNA分离试剂盒(MDSAnalyticalTechnologies)分离RNA,使用iScriptcDNA合成试剂盒(Bio-Rad)0.5μgRNA用于产生cDNA的第一链。使用TaqManFastUniversalPCRMasterMix(AppliedBiosystems)与以下表3中所列的FAM标记的TaqMan探针测定相对基因表达水平。表3.ABI引物与探针H.基因表达分析使用PicoPureRNA分离试剂盒(MolecularDevices,Sunnyvale,CA)从细胞分离总RNA。简而言之,使用MessageAmpIIαRNA放大试剂盒(AppliedBiosystems/Ambion,Austin,TX)的标准实验方案,利用任选的第二轮放大,然后使用MessageAmpII生物素增强试剂盒(AppliedBiosystems/Ambion,Austin,TX)标准实验方案转录到生物素标记的αRNA,制备生物素化的αRNA。根据Affymetrix推荐纯化生物素标记的αRNA并使其成碎片。45℃下,20μg片段的αRNA用于与人基因组U133Plus2.0芯片(AffymetrixInc.SantaClara,CA)杂交16小时。洗涤阵列并且在Affymetrix孵育装置450中染色,使用Affymetrix基因芯片扫描仪30007G扫描。使用Affymetrix表达控制台软件,应用缺省分析设置分析图像数据。通过对数分度强大的多阵列分析(RMA)标准化阵列,并且在针对基因组3.1的Spotfire(TibcoSpotfire,PaloAlto,CA)中使其形象化。I.核型分析与拷贝数变异分析在20G带解离中期相细胞上,通过位于Madison,WI细胞系遗传学进行细胞遗传学分析。通过WiCell(Madison,WI)进行高分辨率比较基因组杂交(NimbleGen12x135k)以及随后的拷贝数变异分析。实施例2:细胞培养条件与方法以使无饲养层培养环境与酶促的单细胞能够解离以及多能干细胞的传代本实施例涉及多能细胞群的培养与解离。此类细胞群包括但不限于,胚胎干细胞与诱导性多能细胞,诸如通过体细胞核移植(SCNT)或经由多能性因子-诱导性多能干细胞(iPSC)的引入所产生的那些。多能干细胞培养条件传统上包括饲养层细胞的使用,饲养层细胞经由辐照或丝裂霉素C处理使有丝解离失活,但是提供了支持干细胞培养所需的生长因子与营养素。不使用饲养层细胞的干细胞群的培养对于研究与工业应用是有利的,其中同源的干细胞群是必需的或其中换算,工业行为需要无异种的,特定的干细胞产生培养条件。在本实施例中,测试几个特异的细胞信号转导通路的小分子调节剂以建立,如果个体的小分子或小分子的组合(“混合物”)可用于增强多能细胞在无饲养层系统中的培养。当培养多能细胞群不使用饲养层反而使用更多确定的细胞外基质诸如常规的hESC细胞培养基中的MatrigelTM(诸如表1与2的第一部分中所述的常规的/基础的培养基制)不支持细胞活力与多能性(图1)。然而,Rock激酶的小分子抑制剂的使用提高了在这些条件下的细胞活力。而且,MAP激酶,TGFβ以及Wnt/β-连环蛋白通路的小分子抑制剂的使用在缺乏饲养层的情况下保持细胞的多能性质,尽管可能会注意到细胞活力中的变化。Rock激酶抑制剂的组合,与MEK、TGFβ以及GSK3抑制剂组合都保持多能干细胞的活力与多能性,当培养在无饲养层环境中时。多能细胞,诸如ESC或iPSC通常生长为团块。传统上,通过本领域中熟练的研究人员所识别的形态人工选择集落这些细胞扩增并且传代。本文的实施例1描述了此类程序(描述为团块传代)。然后将选择的集落打碎并且将解离的细胞再接种于平板。多能细胞群的快速扩增得益于酶促的、单细胞传代的使用。诸如胰蛋白酶与accutase的酶通常用于传代期间细胞的单细胞解离。在特定的演示中,iPSC细胞显示了显著的活力下降当酶促传代并且作为单细胞平板接种在无饲养层环境中时,如通过图3B中的7AAD合并可以见到。通过使用包含培养基组分诸如表1与2中所列的那些的本发明的培养平台,将多能细胞群培养在无饲养层培养基中并且酶促地解离单细胞传代,极大地提高了细胞的活力以及多能性的保持。更具体地说,通过使用常规的/基础的hESC培养基制剂与细胞信号转导通路诸如MAP激酶通路、TGFβ通路、Wnt/β-连环蛋白通路以及Rho/Rock通路的调节剂的组合,在需要单细胞解离与无饲养层培养的应用期间保持了多能干细胞的活力与多能性。如图2中所示,无饲养层细胞培养基上所产生的并且使用团块传代的iPSC为使用诸如accutase的酶解离的单细胞并且作为单细胞平板接种于无饲养层系统中,诸如在MatrigelTM与表1中所述的SMC4或SMC4+纤连蛋白培养基组合物的使用。只有当将解离的细胞置于包含使用这些培养基中所述的小分子的组合的培养基中时,保持细胞活力。常规培养基的使用,诸如表1中的基础培养基制剂,导致了大量的细胞损失当多能细胞酶促地解离为单细胞并且平板接种在无饲养层环境中时。一旦细胞适应了无饲养层培养基与单细胞传代,它们就以这种与SMC4培养基使用的方式不断地保持。适应无饲养层、但细胞传代条件的hiPSC细胞的全部特征显示于图3中。多重传代后保持这些细胞的多能状态:通过免疫荧光与基因表达鉴定多能性标志物。而且,约99.8%的细胞保持SSEA4与Tra1-81阳性染色,如通过流式细胞术测量的。这些细胞的基因表达谱与饲养层上培养的人ESC相似。保持hiPSC中的转基因沉默,核型为正常的并且与在饲养层细胞培养中生长的相同的克隆一样。而且,以这种方式传代及保持的hiPSC分化为三个胚层,如通过体外分化与畸胎瘤形成(图3I与3J)所示。实施例3:细胞培养条件与方法以使多能细胞能够单细胞分选同时保持多能性与细胞活力如实施例2中所述的,ESC或iPSC的干细胞培养物常规培养在饲养层细胞上并且通过细胞集落的人工选择然后在平板再接种前细胞集落机械地解离传代。熟练的研究人员基于集落形态能够识别具有多能的、非分化的特征的干细胞集落并且使用这种作为选择多能细胞的一种方法。因此可以挑选并且人工地从其中一些细胞具有不太理想的特征的细胞群,诸如在培养或死细胞的团块中显示分化迹象的细胞,富集多能群或具有理想特征的群。这种过程是辛苦的并且取决于熟练的研究人员挑选理想的细胞群。其中根据理想的特征单独地选择细胞细胞富集或分选技术的使用对本领域大有裨益。使用目前可获得的技术,诸如磁性激活细胞分选(MACS)或荧光激活的细胞分选(FACS),此类富集步骤将需要在富集前多能细胞群的酶促传代至单细胞格式并且接种回到培养基。而且,饲养层支持的培养物的使用对这些技术不太理想,因此有必要使用无饲养层培养系统。在具体的实施方案中,使用如表1中所述的本发明的培养基组分以及实施例1与2中所述的方法,多能细胞群为解离的单细胞,使用细胞分选过程,诸如磁性激活细胞分选(MACS)或荧光激活的细胞分选(FACS)富集并且接种在无饲养层培养上而没有损失多能性或细胞活力。在具体的实例中,如图4中所示,通过基于对细胞表面标志物SSEA4+/Tra181+阳性的细胞选择及分选解离的多能细胞保持在SMC4培养基(表1)中的单细胞群。这些表面抗原通常用作多能性标志物。然后将分选的双阳性的群转移至无饲养层培养(MatrigelTMECM包被)并且在转回到SMC4培养基(表1)之前允许生长在SMC4培养基(表1)或SMC4+纤连蛋白培养基2-4天。分选24小时内见到细胞分裂并且分选后第5天细胞接近融合。分选的细胞为培养另外5次传代的单细胞并且显示不但表达未分化状态的标志物而且代表基本上纯净的多能细胞群,根据SSEA4+/Tra181+细胞双染色流式细胞术(图4B与4C)判断。在培养期间多能干细胞的富集与分选的实例中,对Tra181阳性的细胞而言,富集包含多能的与分化细胞的培养,然后Tra181+细胞的传代继续保持纯净的多能干细胞群(图17C)。研究多能细胞的单细胞分选的效率。如图4D与E中所见到的各种分选后事件数目,如通过FACS仪器所测量的,使用SMC4培养基或SMC4+纤连蛋白培养基平板接种于无饲养层培养系统,并且对碱性磷酸酶阳性的细胞数量计分。碱性磷酸酶为多能集落形成的早期诱导剂。量化的分选后接种效率,如通过碱性阳性的集落数/接种的细胞数所示,在这个系统中见到大约8-10%(图4E)。在其他的多能细胞群的保持中单细胞分选的潜能的演示,iPSC培养为解离的、对多能性标志物SSEA4与Tra181特异的结合荧光的抗体标记的单细胞,应用于FACS并且基于这些标记选择。以1-9个事件/孔从FACS仪器直接平板接种选择的SSEA4+/Tra181+细胞至96孔平板。尽可能少的1事件/孔产生克隆的碱性磷酸酶-阳性的集落(图4F与4G)。因此,此类单细胞分选系统用于多能细胞的克隆选择,基于优选与某些细胞表面标志物有关的特征。实施例4:方法与条件以使有效的细胞重新编程能够无饲养层培养系统中多能状态iPSC用于工业的和/或临床的应用的用途使细胞在完全确定的培养条件,特别是无异种的条件中细胞的产生、选择以及保持成为必要。因此,无饲养层培养条件中的细胞重新编程是高度理想的。然而,虽然由于经由皮肤活检或毛囊的通路,成纤维细胞与角质细胞为最常用的用于重新编程的细胞类型,这些细胞类型重新编程的效率非常低,并且还要证明无饲养层培养中这些细胞重新编程的有效方法。如实施例2中所述,在细胞信号转导通路-特异地MAP激酶通路、TGFβ通路、Wnt/β-连环蛋白通路以及Rho/Rock通路的抑制剂存在的情况下,常规的hESC干细胞培养基使多能培养物在缺乏饲养层细胞的情况下能够生长并且保持以及使用酶促的单细胞传代扩增这些培养物。在本实施例中,iPSC细胞产生于缺乏饲养层细胞的培养系统中。具体地,人成纤维细胞被表达多能性因子Oct4、KLF4、Sox2以及C-myc的病毒感染。如实施例1中所述的与平板接种在MatrigelTM的细胞而不是饲养层细胞一起进行重新编程实验方案。与由无饲养层细胞重新编程相比,使用常规的hESC干细胞培养基或添加了SMC4培养基(表1)中所列的特异性通路调节剂的常规的hESC培养基。如图5中可以见到的,使用个体的慢病毒表达Oct4、Klf4、Sox2以及Myc,诱导重新编程后,在添加了SMC4培养基的无饲养层培养中见到许多iPSC样集落而在单独的常规的hESC培养基中保持的培养物中很少见到或见不到集落。SMC4培养基中产生的集落的随后的特征显示它们许多为真正的iPSC集落。通过多能性标志物Oct4、Nanog、Sox2、KLF4、SSEA4以及TRA1-81的表达测定细胞的多能性,使用免疫荧光、流式细胞术以及基因表达谱(图13C-E)。而且,细胞显示有效地分化为所有三个胚层当培养在分化培养基中时。因此,通过本发明证明了无饲养层培养系统中iPSC的有效产生。这种方法的效率,与其他的重新编程方法的比较,描述于实施例8与表4中。实施例5.用于产生的细胞培养组合物与原初状态的多能细胞的保持最近的研究已证明通过表观遗传重新编程,终末分化细胞具有求助于祖细胞状状态(Xie,H.,Ye,M.,Feng,R.以及Graf,T.2004),不同的分化细胞状态(Szabo,Bhatia2010)或甚至回到胚胎状状态(Takahashi等,2006),诸如iPSC的能力。尽管iPSC的产生已变得更加常规,给定实验中只有很小百分比的体细胞重新编程为iPSC。归因于这种低效率的几个参数包括体细胞的增生状态,导致基因激活或抑制的额外的突变形成,基因送递的格式以及环境线索。也已报道了不是所有鉴定为iPSC的细胞与ESC表现相似。例如,基因表达谱已证明了许多iPSC显示了在它们的ESC相对物表达谱方面显著的差异。另外,Xist活性的研究与X染色体再激活分析显示虽然一些ESC处于原初状态(即多能性的基态),大多数,如果不是所有衍生的iPSC处于成熟的状态(即成熟为分化)。组合的,这些差异可有助于减少的多能性与对于特定的细胞类型iPSC分化中低效率,减小了再生医学中iPSC的值。通过把涉及原初的与成熟的状态后面的机制的关键的细胞通路作为目标,本发明人已证实了转变以成熟的状态存在的包括常规的iPSC的多能干细胞为原初状态的能力。使用表1与2中所列的培养基组分,可能的是此类成熟的多能细胞与常规的iPSC进一步重新编程为原初状态。更具体地,通过在包含细胞信号转导通路的调节剂,诸如MAP激酶通路、TGFβ通路和/或Wnt/β-连环蛋白通路的培养基中重新编程体细胞或培养iPSC,产生的或培养的iPSC的基因表达信号变得比常规的iPSC(常规的培养基中并且未与细胞信号转导通路的调节剂接触所产生和/或培养的iPSC)更像ESC。如分级群聚中所示,常规的培养基与衍生hiPSC的SMC4培养基彼此相似并且不同于它们的亲代系IMR90(图6B)。然而,当SMC4培养基中培养的iPSC平板接种回到饲养层细胞时,它们比常规培养的iPSC更紧密地类似小鼠ESC,原初状态的另一个论证(图6E)。而且,SMC4培养基中培养的iPSC显著地减小了Xist活性并且增强X染色体的基因表达(图6C与6D)。最终,SMC4培养基中培养的iPSC,不但分化为所有三个胚层,而且证明了再激活与胚胎外细胞有关的基因的能力,只保留原初状态中细胞的能力(图3H)。因此,SMC4培养基中培养多能干细胞,包括以成熟的状态存在的iPSC,促进了原初状态并且增强分化潜能。为测定使用添加SMC4的培养基与细胞分选平台所产生的hiPSC克隆的多能状态(图7),hiPSC衍生于相同的起始成纤维细胞系与表达Oct4、Klf4以及Myc的3-因子多顺反子性载体但是保持在常规培养下,包括团块传代与饲养层细胞(FTi99)。进行各种克隆以及来源于hESC的mRNA的Affymetrix全球基因表达分析(图7B)。可见到所有多能细胞系具有与对照的成纤维细胞系(FTC1,Pearson得分0.886)不同的谱系并且彼此基本上相似的表达谱系(Pearson得分0.969)。产生的热图签名代表hESC/FTi99(饲养层细胞与常规培养基上产生并且保持的hESC或hiPSC)与FTi91/FTi93(无饲养层与SMC4培养基上产生与保持的hiPSC)组(图7C)间4倍差异性表达的1739个探针。1739个差异性表达转录内基因的深度分析中鉴定了一个感兴趣的趋势:虽然通常与原初状态有关的几个多能性基因在FTi91/FTi93组中被上调,更显著地,与成熟状态有关的许多分化基因在这个组内被抑制(图7D)。SMC4培养基所产生的hiPSC的全分析提示培养条件在决定未分化状态方面发挥了更有影响力的作用,相对于起始细胞系或衍生策略。数据也提示来自或适应SMC4培养条件的克隆展示了与优选的质量有关的原初特征,诸如与早期谱系标志物的减小的表达一起高度未分化状态(图7E)。实施例6:鉴定真正的hiPSC的抗体混合物的鉴定研究多能干细胞表面标志物。除了SSEA4与Tra181表达以外,也鉴定了CD30与CD50的表达并且认为代表额外的多能性表面标志物(图10)。用具有真正的多能性标志物,诸如Nanog表达的很少数细胞鉴定使用代表各种潜能状态的多顺反子性慢病毒表达Oc4、Klf4以及Sox2重新编程的细胞。迄今为止,没有基于表面标志物表达鉴定真正多能的hiPSC的可靠的方法。例如,如图11中所见,一些鉴定为针对SSEA4、Tra181以及CD9阳性的克隆群不表达Nanog,并且被错误地鉴定为hiPSC。目前的发明提供了鉴定表达Nanog,一个真正多能的细胞标志物的细胞群的细胞表面标志物的组合。具体地,针对CD30、SSEA4以及Tra181表面标志物阳性的细胞鉴定表达Nanog的细胞(图11)。本研究证明了CD30/SSEA4/Tra181表达的组合代表与Nanog表达与真正多能的hiPSC相关的表面标志物的独特组合。在其他增加的富集的实例中,分选进行重新编程的细胞群以鉴定针对CD13表面标志物表达阳性的细胞并且将这些CD13+细胞从重新编程的细胞群移除。关联体细胞的CD13+群与非重新编程的细胞,以及消耗CD13+细胞的重新编程的细胞群增强了SSEA4/Tra181阳性细胞的富集(图12)。如图12中所示,当使用表达Oct4、Klf4以及Sox2的多顺反子性载体重新编程体细胞21天时,建立各种不同的表面标志物表达模式的细胞,只有一个小的细胞子集代表了SSEA4与Tra181阳性细胞(虚线箭头,图12)。然而,当基于CD13表达(实线箭头,图12)首次评估相同的群时,针对CD13阴性的群子集(即CD13细胞的耗尽)代表了已显著地富集了SSEA4与Tra181表达细胞的群(11.04%,图12)。与此相反,表达CD13高水平的细胞子集代表表达SSEA4与Tra181的少数细胞(1.14%,图12)。实施例7:来源于分化细胞的诱导性多能干细胞的产生中单细胞分选的使用如图8A中所见,重新编程过程中早期的成纤维细胞培养,其中使用个体的表达Oct4、Klf4、Sox2以及myc的慢病毒诱导重新编程,包含形态不同的细胞的集落。这些细胞的一些针对多能性标志物染色阳性而其他的仅为转化的、快速生长的细胞。在重新编程的这个阶段,尚不清楚由单独的细胞形态其细胞集落将继续形成iPSC。更快的生长转化但不是多能细胞迅速占据培养基。因此有利的是具有在重新编程过程早期选择iPSC的富集步骤。而且,如图8中所示,重新编程过程期间一些集落表达几个多能性标志物,并且为真正的iPSC,而一些集落未完全地重新编程并且只表达一些多能性标志物。图8B中集落1针对SSEA4与Tra181阳性的,而集落4与5只针对一个或另一个标志物阳性。因此,同时地使用多能性的细胞表面标志物或几个标志物而不是通过集落形态选择多能细胞,这是更有效率的且更少技术上挑战的。本文也提供了使用本文所述的与细胞富集和/或分选方法组合的细胞培养组分,可能的是通过重新编程过程期间选择显示多能性的表面标志物的个体的细胞,以更多数量并且在更短的时间段内衍生的iPSC。更具体地,如图9A与B中概要所示,使用本发明的方法进行重新编程期间使用单细胞分选与基于多能性标志物的富集的两条iPSC的产生途径。途径A中,启动重新编程后,产生各种潜能状态的混合细胞群。混合的细胞群包含分化细胞、部分重新编程的细胞、重新编程的细胞以及进行重新编程的细胞。使用诸如磁珠分选或流式细胞术分选表达多能性标志物诸如SSEA4的细胞(参见实施例1方法)富集细胞群。一旦富集,细胞就保持在SMC4培养基中(表1)或在具体的实施方案中,SMC4+纤连蛋白培养基大约3天随后SMC4培养基,并且大约6-10天的培养期后,基于诸如SSEA4与TRA181的活的培养物标志物染色鉴定iPSC集落,挑选并且分选用于克隆扩增(图9)。在途径B示意图(图9)下,重新编程启动后早期,分选混合的细胞群以获得稀少的针对两种或多种多能性标志物阳性的细胞群。此类标志物包括但不限于SSEA4与TRA181。将表达多能性标志物的组合的选择的细胞转移至添加了饲养层细胞的培养系统或无饲养层培养系统,特别是添加了表1与2中所述的细胞培养基组分的那些。用这种方法,与早先描述的方法相比,显著减少的时间线与技术壁垒产生了iPSC集落。在这种技术的具体示范中,IMR90成纤维细胞被表达Oct4、Sox2、Klf4以及c-Myc的慢病毒感染(OSKM)。无饲养层培养几天后,重新编程细胞从它们的体细胞培养基转换到添加了SMC4培养基的无饲养层培养(表1)。重新编程启动后第8天,通过流式细胞术分析见到感染的细胞群以包含表达多能性标志物SSEA4的适度的亚细胞群(图13A)。使用如实施例1中所述的磁性激活的细胞分选,富集表达SSEA4细胞3倍的多能细胞群(图13A)。富集针对SSEA4表达细胞的细胞后,将分选的细胞转移至包含MatrigelTM并且添加了常规的hESC培养基或SMC4培养基或在具体的实施方案中,SMC4+纤连蛋白培养基的无饲养层培养。培养物的碱性磷酸酶染色显示MEK、GSK3、Rock激酶以及TGFβ的小分子抑制剂的使用支持使用单细胞分选多能细胞的富集而常规的基础培养基不能(图13A)。三个独立的实验的定量明确证明了只有在小分子抑制剂存在的情况下通过单细胞分选的多能细胞选择(图13B)。进一步在SMC4培养基中培养用这种方法富集的细胞集落并且表征为真正的iPSC:免疫荧光与流式细胞术中,针对多能性标志物SSEA4与Tra181,染色阳性的真正的iPSC(图13C与D);显示与人ESC相似的基因表达谱(图13E);显示显著的外源性转基因的沉默(图13F)并且能够分化为所有三个胚层(图13G)。当使用相同的实验方案但用常规的培养条件(由包含饲养层细胞与缺乏的SMC4的常规培养基组成)时很少观察到多能细胞集落。因此细胞重新编程期间,基于多能性的细胞表面标志物,使用单细胞分选强力地富集多能细胞群的能力取决于表1与2中所列的细胞信号转导通路抑制剂与添加剂的使用。另外,当添加的纤连蛋白用于培养系统中3天时,观察到提高的分选后接种。也针对其他起始的细胞类型包括人脂肪源性干细胞完成这种重新编程过程。在技术的其他实例中,FACS用作从非重新编程的、部分重新编程的以及完全重新编程的细胞的混合物富集多能细胞群。与前面的实例一样,IMR90成纤维细胞被表达Oct4、Sox2、Klf4/c以及Myc的慢病毒(OSKM)感染。成纤维细胞感染后几天将细胞培养转换至SMC4培养基中无饲养层培养。感染的细胞群包含对多能性的SSEA4与Tra181标志物阳性的适度的细胞群(图14A)。选择对两种多能性标志物阳性的细胞并且从针对两个标志物阴性的或只针对一个标志物阳性的细胞分选出来。随后的无饲养层培养与添加如表1或2中所述的培养基的比较来自于针对两种标志物阳性的细胞的培养物形成随后表征为多能iPSC的集落而通过FACS门控针对多能性标志物阴性的细胞不产生碱性磷酸酶阳性的集落(图14A,B)。更具体地,信号转导分子MEK、GSK、Rock以及TGFβ的抑制剂作为基础培养基制剂的添加剂促进了FACS在重新编程过程早期分选选择与多能细胞富集中的使用。另外,当添加的纤连蛋白用于培养系统3天时观察到提高的分选后接种。SMC4培养基与培养系统的优势其次用于发展高通量产生无饲养层与克隆衍生的hiPSC的方法。设计方案以处理诱导的细胞与SMC4培养基重新编程并且选择如通过多能性标志物的组合所示忠实地重新编程稀少的个体细胞。而且,我们将重新编程过程与多重平台连接以基于双标记流式细胞术的选择测定、qRTPCR以及免疫荧光(图15)有效地选择顶级克隆。在优化的多重实验方案中,使用3因子(OKS)多顺反子性病毒启动重新编程,感染后第20天随后在第30天通过FACS分选SSEA4+/Tra181+细胞至96孔平板(图15与B)完成SSEA4+/Tra181+群的最初的大部分FACS分选。这种策略考虑到许多克隆的衍生并且导致表达多能性标志物,展示外源基因活性衰减以及Oct4启动子的去甲基化作用(图15A-C)的克隆FTC1克隆1与2的产生。通过体外与体内分化为三个体细胞谱系(图15D,E),FTC1克隆1与2也显示了多能的。也评估选择的克隆染色体完整性并且显示与它们亲代细胞系与正常的核型甚至在FF培养中20次连续的单细胞传代后具有最小的拷贝数变异,超过其他重新编程的策略显著的改善(图17)。为测定平台的再生性,使用表达Oct4、Klf4及Sox2的多顺反子性载体诱导额外的成纤维细胞系、FTC5以及FTC7重新编程并且应用于如图15中所述的高通量平台。如图18中所述,衍生于FTC5与FTC7成纤维细胞系的集落显示保持它们的未分化状态,保留它们的多能性以及基因组稳定性。因此,3种多能性因子、SMC4培养基以及多重特征化平台的组合显著增强了无饲养层重新编程的动力学而同时以高通量的方式能够鉴定、选择以及扩增克隆衍生的与基因组稳定的hiPSC。实施例8:用于多重iPSC克隆的快速产生的方法与培养组分通过多能性基因诸如Oct4、Sox2、Klf4、c-myc、Lin28以及Nanog的异位表达人iPSC的产生为效率低的且技术要求高的过程。涉及多能性因子转基因的慢病毒或逆转录病毒整合至宿主细胞基因组,与包括饲养层细胞支持组合的策略传统上为用于iPSC产生的最有效的方法。使用用于人iPSC产生的病毒与饲养层细胞方法的历史研究的文献综述显示0.001%-0.01%的受感染的细胞变成iPS细胞的效率,其中在感染后第21-30天期间见到潜在的多能细胞并且这些为感染后第30-45天间通过人工“团块”传代克隆衍生的。其他用于引入多能性基因的方法包括附加型载体系统与修饰蛋白的转导。此类细胞被认为是关于iPSC技术最终的临床应用的重要发展。然而,这些方法甚至效率比使用病毒系统重新编程的效率更低。而且,也减小了重新编程的效率或在某些情况下为不可能的,当无饲养层细胞系统与常规的干细胞培养基制剂组合使用,妨碍iPSC用于工业与治疗用途的发展。体细胞重新编程已表征为随机过程;大多数细胞随着时间流逝将最终重新编程。然而还必须描述一种强力的、技术简单的、有效率的并且可扩展的用于单细胞重新编程中产生多重iPSC克隆的方法。本发明提供了细胞培养条件与方法以在相对短的时间内并且比目前的方法技术壁垒更低衍生克隆的iPSC集落。具体地,并且如图13B中所见,特异的信号转导通路的小分子抑制剂用作SMC4培养基中添加剂允许在无饲养层培养环境中比不使用SMC4培养基时iPSC集落的产生效率大得多。MEK、GSK、Rock及TGFβ信号转导通路的抑制剂用于允许无饲养层环境中有效的重新编程。如图13与表4中所见,在三个独立的重新编程实验,当在无饲养层环境重新编程过程中使用常规hESC培养基时看不到或只有一个集落,而小分子培养添加剂(即SMC4培养基)增强无饲养层重新编程事件达到0.035%的效率,并且导致感染后14-21天集落以及28-35天克隆的iPSC的衍生。因此,小分子抑制剂的使用增加了重新编程的效率,就重新编程时间与重新编程的细胞的百分比而言。使用这种方法产生的集落表征为使用标准的程序,诸如免疫荧光与基因表达谱的多能的。在这种技术的另外示范中,分化细胞被表达个体的多能性基因Oct4、Klf4、Sox2以及Myc的病毒感染,并且被培养在SMC4培养基与无饲养层环境中(表1)8-12天。在该时间点上,并且如实施例6及图13与14中所述,使用FACS或MACS单细胞分选富集细胞以获得针对多能性标志物诸如SSEA4与Tra181阳性染色所确定的小的多能细胞群。使用这种方法,大大减少了iPSC产生时间轴:证明了在重新编程的细胞的百分比方面效率0.22%,分选后即刻出现集落(第10-16天)。重新编程的效率允许到感染后21-28天克隆的iPSC的衍生。在提高重新编程的效率的其他示范中,其中3种(Oct4、Klf4及Sox2)或4种(Oct4、Klf4、Sox2及myc)多能性因子表达于相同的启动子元件的多顺反子性载体系统与优化的SMC4培养基、无饲养层培养以及使用SSEA4与TRA181的单细胞分选系统组合使用,导致重新编程效率0.756%,在感染后第6-8天首先见到集落。这种方法如此有效率的从而使得仅在感染后第4天iPSC集落就出现。这些技术代表了优于传统的iPSC产生方法显著的改善。表4:各种策略中重新编程的动力学。N.I.,未鉴定;*iPSC样集落的形态表现;**计算为SSEA4+/Tra181+数量/接种的细胞;***iPSC集落扩增并且保持为克隆细胞系所需的时间(基于Tra181/SSEA4染色)。灰色框代表文献检索来源的数据。单独的因子病毒;通过组合每个表达一个关键转录因子的单独的病毒进行感染,3代表Oct4/Klf4/Sox2组合并且4代表Oct4/Klf4/Sox2/cMyc组合。实施例9:使用单细胞分选与富集从分化细胞群消耗多能细胞群的方法药物筛选与干细胞生物学的一些临床应用需要从多能细胞诸如ESC或iPSC分化为特定谱系的同源性细胞群的产生。分化细胞群与多能细胞的接触导致误导的筛选结果或甚至体内肿瘤/畸胎瘤形成。富集分化细胞的细胞群或从细胞群消耗多能细胞的方法可包括本文实施例3与6中所述的分选技术。细胞培养基中使用小分子添加剂以特异地阻止单细胞分选过程期间多能细胞分化或部分分化,如本发明所提供的,允许多能细胞从完全分化的细胞群阴性地选择。相反地,在多能细胞针对多能性的表面标志物仍保持完全阳性的培养条件下,通过细胞分选从细胞群阳性选择分化细胞可以更加有效的。如图20中所见,当细胞在表1中所述的培养环境中预培养时,使用FACS,有效地分离混合的完全分化的成纤维细胞与多能细胞群。分选程序期间也可以使用小分子培养添加剂以稳定单细胞悬浮液。可以从图20见到,在随后的培养与碱性磷酸酶染色上,多能性标志物阴性选择的细胞完全见不到多能细胞。实施例10:无饲养层培养上多能干细胞的无细胞因子与生长因子培养如实施例2中所讨论的,常规人多能培养系统包括饲养层细胞与细胞因子,诸如bFGF,其用作外源性刺激保持未分化状态中多能干细胞。饲养层细胞与产生重组的细胞因子的过程用作异种污染的来源。另外,尚未鉴定饲养层细胞分泌的关键因子与细胞因子刺激的精确的细胞通路。因此,人多能干细胞的常规培养代表定义不清的系统并且阻碍转化至临床级大规模应用。为处理这种情况,本发明包括另外的实施方案,其中通过从SMC4培养基制剂移除bFGF与其他的细胞因子及生长因子进一步地修饰实施例2中所讨论的无饲养层与单细胞传代系统。而且,在本发明的一实施方案中,用明胶取代MatrigelTM,因为MatrigelTM代表动物源性并且没有充分表征的细胞外基质。本发明的这些实施方案提供了完全定义的无细胞因子的培养系统,其考虑到内源性自我更新并且保持多能干细胞,包括iPSC。如图21A与21B中所示,人多能干细胞,诸如用包含Oct4、Klf4及Sox2的多顺反子性载体系统产生并且保持在无饲养层环境中的iPSC,易于单细胞传代到明胶包被的缺乏额外的细胞因子或生长因子诸如bFGF的SMC4培养基培养表面。在该完全定义的系统中几次传代后,iPSC保持如通过Tra181与SSEA4共表达所示的它们的多能状态(图21C)。而且,我们已证明了在该完全定义的无细胞因子的系统中iPSC的产生。因此,在一实施方案中,本发明提供了缺乏生长因子与细胞因子,包含SMC4培养基与明胶组合的完全定义的培养系统。实施例11.多能干细胞的产生与保持中基因组稳定性研究提示重新编程过程与随后多能干细胞培养可导致基因组异常倾向较高。另外,无饲养层培养已显示引起核型异常细胞的克隆生长。如图17A、图17B及18C中所示,本发明提供了重新编程的方法以获得具有基因组稳定性的细胞以及保持具有基因组稳定性重新编程的细胞的方法。在本发明的方法中,重新编程过程期间以及使用包含Oct4、Klf4及Sox2的3因子多顺反子性构建体重新编程细胞并且培养于SMC4培养基中时长期无饲养层培养期间,保持基因组稳定性。如图17A中所见,高分辨率比较基因组杂交证明了当使用SMC4培养基产生hiPSC并且保持在长期无饲养层培养中时,检测到最小的拷贝数变异。另外,在如图17B与18C中所示的常规的长期无饲养层与单细胞培养期间,保持基因组稳定性。实施例12:在培养期间使用细胞表面标志物保持多能细胞的细胞群通常需要或有必要从多能细胞培养移除分化细胞以保持多能干细胞诸如hESC与hiPSC的多能性。迄今为止,这个过程需要从细胞培养人工挑选出分化细胞或从极大分化的群体收集未分化细胞。这两个过程都是劳动密集型,需要技能训练,并且依赖于基于形态的细胞选择,其不一定总是真正的培养中细胞状态的标示(图8)。在改善的方法中,本发明提供了在常规培养期间有效地并且精确地选择未分化细胞的能力。如图19A中所示,保持在SMC4培养基与FF培养中的hiPSC群易于富集或分选以保持未分化的多能细胞的细胞培养。在其他的实例中,如图19B所示,通常被丢弃的大多数分化的细胞群,可以从细胞培养移除以获得大多数未分化的多能细胞,如通过Tra181表达所述的。
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1