基于乙型肝炎核心抗原的疫苗的制作方法
本发明涉及包含乙型肝炎核心抗原(hbcag)和流感病毒a表面多肽m2或其免疫原性片段的蛋白。本发明还涉及由所述蛋白形成的颗粒、编码所述蛋白的核酸分子、制备所述蛋白的方法、包含所述蛋白的药物组合物以及所述蛋白用于在对象中诱导免疫反应的用途。
背景技术:
乙型肝炎(hepatitisb)病毒核心(hbc)蛋白具有某种程度上的独特结构,它由两个反平行的α螺旋组成,其形成特有的“穗(spike)”结构。然后两个hbc分子自发地二聚化以形成双穗束。该双穗束是病毒样颗粒(vlp)的构成部件。vlp是具有吸引力的疫苗系统,因为它们的高度重复的性质递送抗原的多个拷贝。进一步地,病毒核酸的缺乏使得它们成为安全的载体。hbc作为疫苗载体是特别有趣的,因为它具有几个可能让抗原序列插入的位点。然后hbc的极端免疫原性也赋予被插入的序列,因此使得其也具有过度的免疫原性。最佳的插入位点是主要插入区域(mir)。然而,先前表明,当较大的或疏水性的序列被插入mir时,则单体的hbc不能二聚化且vlp没有形成(pumpen&grens2001),因此使疫苗无效。
流感病毒是正粘病毒科的成员。存在命名为a、b和c的三种亚型的流感病毒会感染人类。流感病毒粒子包含分段的负链rna基因组。包膜的流感a病毒粒子具有:三个膜蛋白,血凝素(ha)、神经氨酸酶(na)和质子离子通道蛋白(m2);基质蛋白(m1),刚好在脂双层之下;核糖核蛋白核心,由8个病毒rna片段和三个蛋白(聚合酶酸性蛋白(pa)、聚合酶碱性蛋白1(pb1)和聚合酶碱性蛋白2(pb2))组成;以及非结构蛋白2(ns2)。流感b病毒粒子具有在包膜中的四个蛋白:ha、na、nb和bm2。与流感a病毒的m2蛋白相似,bm2蛋白是对于脱壳过程至关重要的质子通道。nb蛋白被相信是离子通道,但是它对于细胞培养中的病毒复制是不需要的。
流感c病毒有些不同。与流感a和b病毒相似,流感c病毒的核心由核糖核蛋白组成,它由病毒rna和四个蛋白组成。如同在流感a和b病毒粒子中一样,m1蛋白刚好位于膜之下。小病毒包膜蛋白是cm2,作为离子通道。主要流感c病毒包膜糖蛋白被称为hef(血凝素-酯酶-融合物),因为它同时具有ha和na的功能。
ha和na蛋白是包膜糖蛋白,负责病毒附着和病毒颗粒渗透进细胞,并且是病毒中和以及保护性免疫力的免疫显性表位。但是,这些蛋白能够并且常常在菌株之间发生变化。由于这两个蛋白的可变性,广谱的、长效的流感疫苗迄今尚未被开发。常用的流感疫苗几乎每年都必须进行调整,以遵循病毒的抗原漂移。当在病毒中发生更多的急剧变化(被称为抗原转移)时,疫苗就不再有保护性。
发明概述
本发明涉及一种基于乙型肝炎(hbv)核心蛋白的疫苗递送系统。当前的对于流感病毒的疫苗要求它们每年都被重新设计,这是因为病毒的快速突变率造成了不被包含在先前年份的疫苗中的逃脱变种的出现。它们依赖于预测主要的流行性流感菌株,但是它们在疫苗和流行性菌株之间存在不匹配的时候变得不理想。此外,它们不能防范新型的、先前未见过的病毒菌株。一个解决方案是设计基于流感的保守的蛋白结构域的疫苗,这些结构域每年都保持大体不变并且在新出现的变种中是保守的。靶定保守结构域的许多先前的尝试都失败了,因为这些区域的自身的免疫原性低或者不能在野生型病毒之外以它们的天然构象呈现。发明人已经设法将来自流感病毒a表面多肽m2(流感基质蛋白2)的保守区域插入进串联构建体。所述保守区域是流感病毒a表面多肽m2胞外域(m2e)。得到的vlp能够产生对相关的m2肽的血清转化,并且提供对致死性h1n1流感感染的保护(见实施例1)。发明人还已经设法将来自流感血凝素(ha)的保守区域插入进串联构建体。得到的vlp能够产生对ha蛋白的血清转化,并且提供对致死性同源h1n1流感感染的保护(见实施例2)。进一步地,发明人还已经设法将来自流感血凝素(ha)和基质2蛋白胞外域(m2e)的保守区域同时插入进串联构建体。得到的vlp能够产生对ha蛋白和m2e肽的血清转化,并且提供对致死性同源h1n1流感感染的保护(见实施例3)。发明人还制造了包含两种不同的vlp的疫苗:包括来自流感ha和m2e的保守区域的第一种vlp的串联构建体;以及包括来自源于流感的不同亚型的流感ha的保守区域的第二种vlp的串联构建体。这两种vlp一共包括5个来自流感的保守抗原,并且可以作为单一的疫苗而被同时递送。该组合疫苗能够产生对组-1和组-2ha蛋白、以及m2e肽的血清转化,并且提供对致死性h1n1和h3n2流感感染的保护(见实施例4)。
发明人发现,将原生的m2e插入进el环会使得vlp不能正确地形成,这是因为存在蛋白错误折叠。发明人意外地发现,可以通过加入在m2e插入体侧翼的接头序列、和/或通过使用替代的氨基酸取代位于m2e的第17和19位的两个半胱氨酸残基中的一个或两个,解决上述问题。所述接头允许vlp的形成,它能够诱导血清转化并且提供对致病性挑战的保护。例如,实施例1中的表2的第1和第4列证明了,将接头引入m2e和el环之间形成了能够诱导血清转化的vlp。因此,所述接头提供了将m2e插入进串联构建体的方法,而如果没有接头,会导致错误折叠并且因此会不能产生在诱导免疫反应上有效的vlp。
下文进一步讨论的是可以被结合进串联构建体中的免疫原性多肽的例子。特别地,当前的发明人已经发现,包含流感病毒a表面多肽m2胞外域和/或血凝素(ha)的茎(stalk)区域的串联构建体产生vlp,该vlp在诱导血清转化和提供对流感挑战的保护上特别有效(见实施例)。
在m2e的第17和/或19位使用丝氨酸取代半胱氨酸允许vlp的形成,该vlp能够诱导血清转化并提供对致病性挑战的保护。如果没有取代一个或两个半胱氨酸,则vlp不会正确地形成,除非如上所述地将接头插入m2e插入体和el环之间。例如,实施例1中的表2的第1至第3列证明了,在第17和/或19位使用丝氨酸取代半胱氨酸能够形成vlp,该vlp能够诱导血清转化,而在使用具有在第17和19位的原生半胱氨酸的vlp时不存在血清转化。据认为,半胱氨酸残基的取代的存在防止了二硫键的形成,它可能干扰vlp的形成。
因此,本发明提供了一种蛋白,其包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的拷贝的一个或两个包括在el环中的流感病毒a表面多肽m2或其免疫原性片段。所述流感病毒a表面多肽m2或其免疫原性片段在一侧或两侧的侧翼有接头,所述接头将所述多肽或片段连接至hbcag序列。
本发明还提供了一种蛋白,其包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的拷贝的一个或两个包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在所述流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代。
本发明的蛋白还可以包括在hbcag的另一个拷贝中的另一种免疫原性多肽。该免疫原性多肽可以衍生自流感。但是,它也可以衍生自不同的病原体或过敏原。因此,本发明的蛋白可以用于诱导对流感病毒的免疫反应(取决于存在何种其他的免疫原性多肽),它还可以用于同时诱导对不同的病原体或过敏原的免疫反应。
发明人还发现,可以通过将抗原肽呈现在hbcag的拷贝中的一个的el环中,并且使得在hbcag的第二拷贝的el环中具有“空(null)”插入体,从而解决被插入的抗原肽的错误折叠的可能问题。特别地,发明人通过将来自流感h3n2病毒血凝素ha2蛋白结构域(lah3)的保守区域插入进串联构建体中的hbcag的一个拷贝,开发了另一个类型的vlp(vlp2)。他们发现,仅将小于20个氨基酸的短序列插入进hbcag的第二拷贝,使得第一插入体(lah3)正确地配置并且赋予整个vlp更大的溶解度(实施例4)。特别地,他们插入了单一的赖氨酸(k)残基,其在侧翼有由甘氨酸和丝氨酸残基形成的短的柔性的接头区域,它有效地是“空”插入体。
因此,本发明还提供了一种蛋白,其包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地为ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列。ha的所述片段任选地来自流感病毒血凝素ha2蛋白结构域,并且进一步任选地来自流感病毒亚型h3n2。hbcag的第二拷贝可以包括在el环中的赖氨酸(k)残基,其在每侧的侧翼有包括甘氨酸和丝氨酸残基的接头序列。
本发明进一步提供了一种蛋白,其包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的拷贝的一个或两个包括根据本发明的在el环中的流感病毒a表面多肽m2或其免疫原性片段,其中hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列。hbcag的第二拷贝可以包括在el环中的赖氨酸(k)残基,其在每侧的侧翼有包括甘氨酸和丝氨酸残基的接头序列。
本发明还提供:
-一种颗粒,其包括一个或多个本发明的蛋白的多个拷贝;
-一种核酸分子,其编码本发明的蛋白;
-一种宿主细胞,其包括一个或多个本发明的核酸分子;
-一种用于制备本发明的蛋白的方法,所述方法包括:在表达所述蛋白的条件下培养包含编码所述蛋白的核酸分子的宿主细胞,以及回收所述蛋白;
-一种药物组合物,其包括本发明的蛋白、本发明的颗粒或本发明的核酸分子,以及药学上可接受的载体或稀释剂;
-一种疫苗,其包括本发明的蛋白、本发明的颗粒或本发明的核酸分子,以及药学上可接受的载体或稀释剂;
-一种在对象中诱导针对流感的免疫反应的方法,所述方法包括向所述对象施用本发明的蛋白、本发明的颗粒或本发明的核酸分子;
-用于在针对流感的人类或动物体的免疫接种的方法中使用的本发明的蛋白、本发明的颗粒或本发明的核酸分子;
-本发明的蛋白、本发明的颗粒或本发明的核酸分子用于制备用于针对流感的人类或动物体的疫苗接种的药物的应用。
本发明的蛋白、颗粒、核酸、药物组合物和疫苗可以被单独使用来针对流感进行保护,或者它们可以被联合使用。包含不同流感抗原的不同vlp的联合使用允许更广水平的保护,例如同时针对流感的不同亚型。考虑到这一点,发明人开发了联合疫苗,其包括两个vlp的混合物,它们一起包括来自流感ha和m2e的5个保守抗原,它们能够作为单一的疫苗而被同时递送(实施例4)。
因此,本发明还提供了:
-一种药物组合物,其包括:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂;
-一种药物组合物,其包括:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂;
-一种疫苗,其包括:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂;
-一种疫苗,其包括:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂;
-一种在对象中诱导针对流感的免疫反应的方法,所述方法包括向所述对象施用:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;
-一种在对象中诱导针对流感的免疫反应的方法,所述方法包括向所述对象施用:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;
-用于在针对流感的人类或动物体的免疫接种的方法中使用的:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列;联合(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;
-用于在针对流感的人类或动物体的免疫接种的方法中使用的:(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代;联合(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;
-(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列;联合(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;用于制备用于针对流感的人类或动物体的疫苗接种的药物的应用;以及
-(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中该蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代;联合(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中该蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;用于制备用于针对流感的人类或动物体的疫苗接种的药物的应用。
附图说明
图1:使用含有流感3xm2e序列的串联核心进行的免疫产生抗体,该抗体识别重组m2e肽。在初级免疫之后3周从使用串联核心+流感3xm2evlp(圆圈)、仅有佐剂(三角形)或对m2e肽的14c2单克隆抗体(方形)进行免疫接种的5只小鼠收集血清。在3种不同的血清稀释度,对m2e使用elisa一式两份地测试来自5只小鼠的血清池。
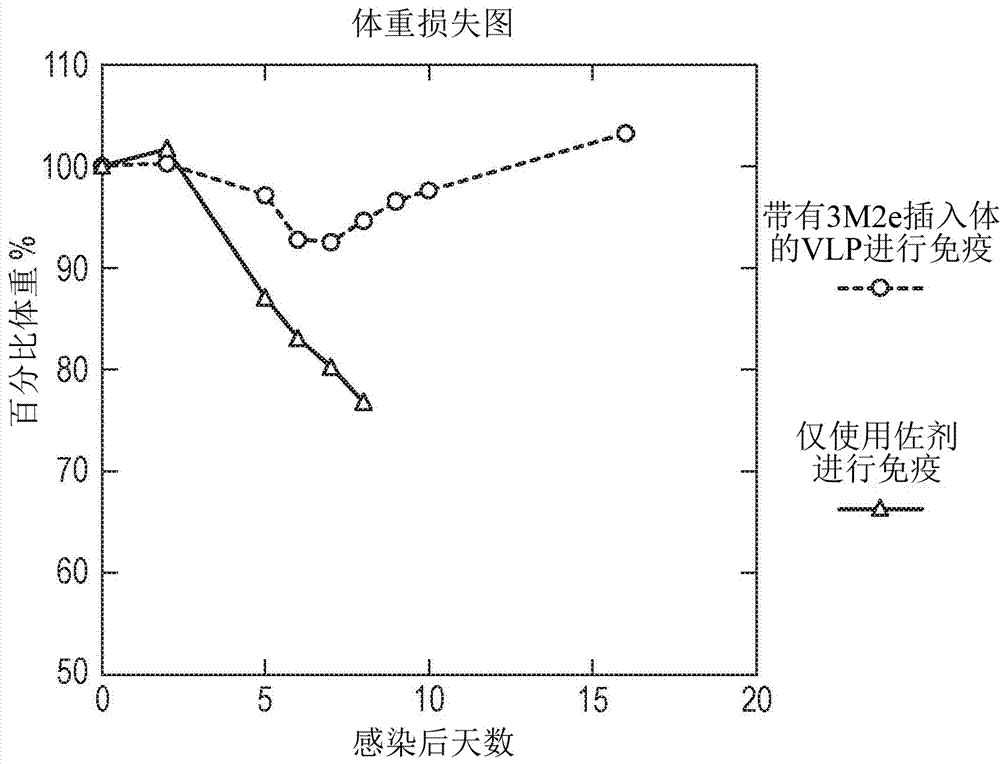
图2:使用含有3xm2e插入体的串联核心vlp进行的免疫在致命流感挑战期间减轻了体重损失。在使用含有3xm2e插入体的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比体重被计算为:%体重=100-(100x(第0天体重–第n天体重/第0天体重)。每组5只小鼠,代表2个单独的实验。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图3:使用含有3xm2e插入体的串联核心vlp进行的免疫在致命流感挑战期间减少了临床疾病。在使用含有ha3xm2e的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。群组临床分数被计算为群组中每只小鼠(每组5只小鼠)的单独的临床分数的加和。通过表3所示的程度确定单独的临床分数。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图4:使用含有3xm2e的串联核心vlp进行的免疫在致命的pr8流感挑战之后消除了死亡。在使用含有3xm2e插入体的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比存活率被计算为:%存活率=100–(100*(第0天小鼠数量-第n天存活小鼠数量/第0天小鼠数量)。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图5:描绘流感病毒的表面界面的示意图。修改自parketal.,j.virol.march1998vol.72no.32449-2455。
图6:使用含有流感茎序列的串联核心进行的免疫生成抗体,该抗体识别重组的血凝素蛋白。在初级免疫之后3周从使用串联核心+流感茎vlp(圆圈)、仅有佐剂(三角形)进行免疫接种的5只小鼠或pr8感染的小鼠(方形)收集血清。在3种不同的血清稀释度,对来自a/pr8h1n1的rha使用elisa一式两份地测试血清池。
图7:使用含有ha茎插入体的串联核心vlp进行的免疫在致命流感挑战期间减轻了体重损失。在使用含有ha茎插入体的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比体重被计算为:%体重=100-(100x(第0天体重–第n天体重/第0天体重)。每组5只小鼠,代表2个单独的实验。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图8:使用含有ha茎插入体的串联核心vlp进行的免疫在致命流感挑战期间减少了临床疾病。在使用含有ha茎插入体的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。群组临床分数被计算为群组中每只小鼠(每组5只小鼠)的单独的临床分数的加和。通过表3所示的程度确定单独的临床分数。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图9:使用含有ha茎插入体的串联核心vlp进行的免疫在致命的pr8流感挑战之后消除了死亡。在使用含有ha茎插入体的串联核心(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比存活率被计算为:%存活率=100–(100*(第0天小鼠数量-第n天存活小鼠数量/第0天小鼠数量)。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图10:示出ha蛋白茎区域的示意图。修改自kaminskiandlee,frontimmunol.2011;2:76.publishedonlinedec16,2011.prepublishedonlinesep12,2011.doi:10.3389/fimmu.2011.00076。
图11:使用含有流感ha茎和3xm2e序列的串联核心进行的免疫生成抗体,该抗体识别重组的血凝素蛋白和m2e肽。在初级免疫之后3周从使用串联核心vlp(圆圈)、仅有佐剂(三角形)进行免疫接种的5只小鼠或pr8感染的小鼠(方形)收集血清。在m2eelisa中将对m2e的单克隆抗体(c14)用作阳性对照(交叉)。在3种不同的血清稀释度,对来自a/pr8h1n1的rha或m2e肽使用elisa一式两份地测试血清池。
图12:使用含有流感茎和3xm2e序列的串联核心vlp进行的免疫在致命流感挑战期间减轻了体重损失。在使用串联核心vlp(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比体重被计算为:%体重=100-(100x(第0天体重–第n天体重/第0天体重)。每组5只小鼠,代表2个单独的实验。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图13:使用含有流感茎和3xm2e序列的串联核心vlp进行的免疫在致命流感挑战期间减少了临床疾病。在使用串联核心vlp(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。群组临床分数被计算为群组中每只小鼠(每组5只小鼠)的单独的临床分数的加和。通过表3所示的程度确定单独的临床分数。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图14:使用含有流感茎和3xm2e序列的串联核心vlp进行的免疫在致命的pr8流感挑战之后消除了死亡。在使用串联核心vlp(圆形)或仅有佐剂(三角形)进行免疫之后4周,使用5xmld50的pr8病毒感染小鼠。百分比存活率被计算为:%存活率=100–(100*(第0天小鼠数量-第n天存活小鼠数量/第0天小鼠数量)。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图15:在两个mir中都含有流感衍生的插入体的串联核心vlp的示意图。核心1以交叉格子示出,其在mir含有ha茎插入体。核心2以黑色示出,其含有三重m2e插入体。所示的二聚体(a)组装成病毒样颗粒(vlp),其在外侧(b)与它们的插入体一起同时显示核心1和核心2。
图16:在一个或两个mir中含有流感衍生的插入体的串联核心vlp1和vlp2的示意图。核心1以交叉格子示出,其在mir含有ha茎插入体。核心2以黑色示出,其含有三重m2e插入体或含有赖氨酸残基的“空”插入体。所示的二聚体(a)组装成病毒样颗粒(vlp1),相似地示出了用于vlp2的构建块。组装的vlp在外侧(c)与它们的插入体一起同时显示核心1和核心2。
图17:描绘vlp1(ha2.3)和vlp2(lah3)里面的流感病毒ha茎插入体的二级结构的模型。
图18:使用含有流感茎和m2e序列的串联核心vlp进行的免疫生成抗体,该抗体识别重组的血凝素蛋白和m2e肽。图上的点代表来自每组5只小鼠的汇集的血清的2个elisa吸光度值的平均值。从血凝素至h3n2的血清转换以三角形示出,至h1n1的以方形示出,至m2胞外域的以菱形示出。阴性对照代表对来自仅有佐剂的汇集的血清的全部3种蛋白的反应性(圆形)。该实验独立地重复3次。
图19:使用含有流感衍生的插入体的串联核心vlp进行的免疫在流感挑战期间减轻了体重损失。在使用串联核心vlp(方形)或仅有佐剂(圆形)进行免疫之后4周,使用5xmld50的pr8病毒(2a)或x31h3n2(2b)感染小鼠。百分比体重被计算为:%体重=100-(100x(第0天体重–第n天体重/第0天体重)。每组5只小鼠,代表2个单独的实验。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
图20:使用含有流感衍生的插入体的串联核心vlp进行的免疫在流感挑战期间减少了临床疾病。在使用串联核心vlp(方形)或仅有佐剂(圆形)进行免疫之后4周,使用5xmld50的pr8病毒(3a)或x31h3n2(3b)感染小鼠。群组临床分数被计算为群组中每只小鼠(每组5只小鼠)的单独的临床分数的加和。通过表3所示的程度确定单独的临床分数。达到20%体重损失、或个体分数15的的小鼠死亡或者根据英国内政部准则而被淘汰。
图21:使用含有流感衍生的插入体的串联核心vlp进行的免疫在致命流感挑战之后消除了死亡。在使用串联核心vlp(方形)或仅有佐剂(圆形)进行免疫之后4周,使用5xmld50的pr8h1n1(4a)或3xmld50x31h3n2(4b)病毒感染小鼠。百分比存活率被计算为:%存活率=100–(100*(第0天小鼠数量-第n天存活小鼠数量/第0天小鼠数量)。达到20%体重损失的小鼠死亡或者根据英国内政部准则而被淘汰。
序列简述
seqidno:1是ayw亚型的183个氨基酸的蛋白加上hbcag的29个氨基酸的前序列以及相应的核苷酸序列。
seqidno:2是ayw亚型的183个氨基酸的蛋白加上hbcag的29个氨基酸的前序列。
seqidno:3是来自流感病毒a菌株a/34/pr8的m2氨基酸序列。
seqidno:4是用于连接相邻的hbcag单元的可能的接头序列,其用于处于el环中的免疫原性多肽插入体的侧翼或者用于将el环中的多个免疫原性多肽结合在一起。
seqidno:5是用于连接相邻的hbcag单元的可能的接头序列,其用于处于el环中的免疫原性多肽插入体的侧翼或者用于将el环中的多个免疫原性多肽结合在一起。seqidno:5是seqidno:4的序列的三个重复。
seqidno:6是野生型m2e序列。
seqidno:7是和seqidno:6的序列一样的,除了在第17位的半胱氨酸已经被丝氨酸取代之外。
seqidno:8是和seqidno:6的序列一样的,除了在第17和19位的半胱氨酸都已经被丝氨酸取代之外。
seqidno:9是通用的m2e共识序列,除了在第17和19位的每个半胱氨酸都已经被丝氨酸取代之外。
seqidno:10是变异的m2e序列。
seqidno:11是串联核心的氨基酸序列,其中三个m2e序列被插入在hbcag的一个拷贝中。被插入的三个m2e序列在两侧有接头序列。一个接头跨越seqidno:11的氨基酸80至94,而另一个跨越seqidno:11的氨基酸167至181(在实施例1中以斜体显示的氨基酸)。所述接头序列与seqidno:5的序列是相同的。所述三个m2e序列跨越seqidno:11的氨基酸95至166(在实施例1中下划线显示的氨基酸)。跨越seqidno:11的氨基酸95至118的m2e序列与seqidno:9的序列是相同的。跨越seqidno:11的氨基酸119至142的m2e序列与seqidno:8的序列是相同的。跨越seqidno:11的氨基酸142至166的m2e序列与seqidno:10的序列是相同的。
seqidno:12是串联核心的氨基酸序列,其中在hbcag的一个拷贝中具有ha茎插入体。所述ha茎插入体跨越seqidno:12的氨基酸80至151(在实施例2中下划线显示的氨基酸)。所述ha茎插入体跨越分离自流感a病毒h1n1/lux/09的氨基酸403-474。
seqidno:13是串联核心的氨基酸序列,其中在hbcag的一个拷贝中具有ha茎插入体,而三个m2e序列被插入在hbcag的另一个拷贝中。所述ha茎插入体跨越seqidno:13的氨基酸80至151(在实施例3中双下划线显示的氨基酸)。所述ha茎插入体跨越分离自流感a病毒h1n1/lux/09的氨基酸403-474。被插入的三个m2e序列在两侧有接头序列。一个接头跨越seqidno:13的氨基酸328至342,而另一个跨越seqidno:13的氨基酸415至429(在实施例3中以斜体显示的氨基酸)。所述接头序列与seqidno:5的序列是相同的。所述三个m2e序列跨越seqidno:13的氨基酸343至414(在实施例3中下划线显示的氨基酸)。跨越seqidno:13的氨基酸343至366的m2e序列与seqidno:9的序列是相同的。跨越seqidno:13的氨基酸367至390的m2e序列与seqidno:8的序列是相同的。跨越seqidno:13的氨基酸391至414的m2e序列与seqidno:10的序列是相同的。
seqidno:14是与seqidno:6相同的序列,其中在c末端加入了–oh基团。
seqidno:15是hbcag可以包含从而平衡α-螺旋的序列。
seqidno:16是用于连接相邻的hbcag单元的可能的接头序列,其用于处于el环中的免疫原性多肽插入体的侧翼或者用于将el环中的多个免疫原性多肽结合在一起。它被用作靠近实施例4中的vlp1中的m2e插入体的接头。
seqidno:17是串联核心的dna序列,其中在hbcag的一个拷贝中具有ha茎插入体,而三个m2e序列被插入在hbcag的另一个拷贝中(实施例4中的vlp1)。
seqidno:18是串联核心的氨基酸序列,其中在hbcag的一个拷贝中具有ha茎插入体,而三个m2e序列被插入在hbcag的另一个拷贝中(实施例4中的vlp1)。所述ha茎插入体跨越seqidno:18的氨基酸80至151(在实施例4中下划线显示的氨基酸)。所述ha茎插入体跨越分离自流感a病毒h1n1/lux/09的氨基酸403-474。被插入的三个m2e序列在两侧有接头序列。一个接头跨越seqidno:18的氨基酸328至342,而另一个跨越seqidno:18的氨基酸415至428(在实施例4中以斜体显示的氨基酸)。所述接头序列分别是seqidno:5和seqidno:16的序列。所述三个m2e序列跨越seqidno:18的氨基酸343至414(在实施例4中下划线显示的氨基酸)。跨越seqidno:18的氨基酸343至366的m2e序列与seqidno:9的序列是相同的。跨越seqidno:18的氨基酸367至390的m2e序列与seqidno:8的序列是相同的。跨越seqidno:18的氨基酸391至414的m2e序列与seqidno:10的序列是相同的。
seqidno:19是串联核心的dna序列,其中在hbcag的一个拷贝中具有ha茎插入体,而在hbcag的另一个拷贝中具有“空”插入体(实施例4中的vlp2)。
seqidno:20是串联核心的氨基酸序列,其中在hbcag的一个拷贝中具有ha茎插入体,而在hbcag的另一个拷贝中具有“空”插入体(实施例4中的vlp2)。所述ha茎插入体跨越seqidno:20的氨基酸80至134(在实施例4中下划线显示的氨基酸)。所述ha茎插入体跨越分离自流感a病毒h3n2/hk/68的氨基酸421-475。所述“空”插入体跨越seqidno:20的氨基酸311至324并且对应于seqidno:21。
seqidno:21是被插入进实施例4中的vlp2中的hbcag的第二拷贝中的“空”插入体的氨基酸序列。
seqidno:22是跨越分离自流感a病毒h1n1/lux/09的ha蛋白的氨基酸403-474的ha茎的氨基酸序列。这对应于seqidno:12、seqidno:13和seqidno:18的氨基酸80至151。
seqidno:23是跨越分离自流感a病毒h1n1/lux/09的ha蛋白的氨基酸403-474的ha长α螺旋的氨基酸序列。这对应于seqidno:12、seqidno:13和seqidno:18的氨基酸97至151。
seqidno:24是跨越分离自流感a病毒h3n2/hk/68的ha蛋白的氨基酸421-475的ha茎区域的氨基酸序列。这对应于seqidno:20的氨基酸80至134。
发明详述
此外,如在本说明书和在附属的权利要求书中使用的,除非另有明确说明,否则单数形式“一”、“一个”和“该”包括复数引用。因此,例如,对“一个免疫原性多肽”的引用包括两个或更多个这样的免疫原性多肽。
本文所引用的所有出版物、专利和专利申请,无论是在上文或下文中引用的,在此通过引用以其整体合并入本文中。
本发明的发明人已经研发了串联核心构建体,其已经克服了与将免疫原性多肽(包括流感病毒a表面多肽m2)引入hbcag核心相关的困难。
所述串联构建体是两个hbcag基因的基因融合,使得得到的重组蛋白形成两个平行的“穗”,它与自然地二聚化的野生型核心蛋白是不能区分的。所述串联核心蛋白以与单体核心蛋白相似的方式形成vlp。“串联核心”、“串联构建体”和“串联核心构建体”在本文中被可互换地使用。该术语可以被用来描述串联hbcag核心,其在hbcag的一个或两个拷贝的el环中具有或不具有免疫原性多肽。
本发明提供了串联核心构建体,其包括在被插入hbcag的一个或两个拷贝的el环中的流感病毒a表面多肽m2或其免疫原性片段的一侧或两侧上的接头。本发明还提供了串联核心构建体,其包括在hbcag的一个或两个拷贝的el环中的流感病毒a表面多肽m2或其免疫原性片段,并且其中在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代。本发明还提供了串联核心构建体,其包括被插入hbcag的一个拷贝的el环中的血凝素(ha)或其免疫原性片段以及小于20个氨基酸的序列,其任选地包括被插入进hbcag的另一个拷贝的侧翼有甘氨酸和丝氨酸残基的赖氨酸(k)残基。
本文描述的特征可以应用至上述的串联构建体的任何一个。例如,包括在流感病毒a表面多肽m2或其免疫原性片段的一侧或两侧上的接头的上述串联核心构建体中的流感病毒a表面多肽m2或其免疫原性片段可以具有在第17和/或19位的被取代的半胱氨酸。
乙型肝炎核心抗原(hbcag)
串联核心是基于hb核心蛋白的病毒样颗粒,其已知是高度免疫原性的并且具有将免疫原性赋予在其结构之内的蛋白插入体的能力。进一步地,串联核心的病毒样特性能够显示外来抗原,而同时维持在多聚体显示平台上的结构表位。此外,由于其双插入位点,串联核心能够携带多个插入体。
根据hbv的亚型,hbcag具有183或185个氨基酸(aa)。ayw亚型的183个氨基酸的蛋白的序列加上29个氨基酸的前序列显示于seqidno:2中。成熟的hbcag是第30位的met残基至最c末端的cys残基,其中第1至29位的序列是前序列。
所述蛋白包括形成二聚体的hbcag的两个拷贝(“第一拷贝”和“第二拷贝”)。hbcag的“拷贝”和“单元”在本文中被可互换地使用。hbcag的二聚体形成vlp的结构性构成部件。该hbcag单元通常以头至尾的形式连接在一起,即一个单元的c末端连接至相邻单元的n末端。“第一拷贝”可以是n末端拷贝或c末端拷贝。这些单元可以通过共价键(例如肽键)直接连接,但优选地它们通过接头连接,所述接头将相邻的单元隔开,从而避免破坏相邻单元的包装的任何问题。接头的性质在下文讨论。被连接的二聚体形成“串联构建体”、“串联核心”或“串联核心构建体”,其可以具有被插入进el环的一个或两个中的免疫原性多肽。
所述蛋白中的hbcag可以是天然的全长hbcag。但是,根据本发明的一个方面,所述单元中的至少一个是具有在el环中的流感病毒a表面多肽m2或其免疫原性片段的hbcag的修改形式。hbcag的另一个拷贝可以是天然的hbcag或者可以是本文描述的hbcag的修改版本。所述hbcag的修改版本可以具有在el环中的另一种免疫原性多肽。所述串联构建体可以具有被插入两个hbcag拷贝的el环中的流感病毒a表面多肽m2或其免疫原性片段。在一个hbcag拷贝中可以存在一种以上的类型的免疫原性多肽。可能的免疫原性多肽的例子在本文中讨论。
根据本发明的另一个方面,所述单元中的一个是hbcag的修改形式,其具有在el环中的流感病毒血凝素(ha)多肽或其免疫原性片段。hbcag的另一个拷贝可以是天然的hbcag或者可以是本文描述的hbcag的修改版本。
所述免疫原性多肽可以在一侧或两侧的侧翼具有接头。因此,串联构建体可以包括一个或多个接头,所述接头位于在hbcag的一个拷贝中的或在hbcag的两个拷贝中的免疫原性多肽的侧翼。如果在el环的一个或每个中存在一个以上的免疫原性多肽,或者如果在el环的一个或每个中存在相同的免疫原性多肽的一个以上的拷贝,则可以存在接头,所述接头在免疫原性多肽的一侧或两侧上位于免疫原性多肽的一个或每个的侧翼。免疫原性多肽的每一个都可以在两侧的一个上具有或不具有接头。可以存在连接相邻的免疫原性多肽的侧翼接头和/或将免疫原性多肽连接至el环的侧翼接头。串联构建体也可以具有连接hbcag单元的接头。接头的性质在下文讨论。
作为总的原则,任何修饰都被选择得不干扰hbcag的构象及其组装成颗粒的能力。这些修饰在蛋白中对于维持其构象不重要的位点进行,例如在e1环、c末端和/或n末端中。hbcag的e1环可以耐受例如1至600个氨基酸的插入而不会破坏蛋白的颗粒形成能力。
hbcag序列可以通过取代、插入、缺失或延伸而被修饰。插入、缺失或延伸的尺寸可以是例如1至600个aa、1至500个aa、1至400个aa、1至300个aa、1至200个aa、3至100个aa或6至100个aa。取代可以涉及hbcag序列的长度上的多个氨基酸,多达例如1、2、5、10、20或50个氨基酸。延伸可以在hbcag的n末端或c末端。缺失可以在蛋白的n末端、c末端或内部位点。取代可以在蛋白序列的任何位置进行。插入也可以在蛋白序列的任何位点进行,但通常在蛋白的表面暴露区域(例如e1环)进行。被插入的序列可以携带免疫原性多肽。可以对每个hbcag单元进行多于一个的修饰。因此,可以进行末端延伸或缺失并进行内部插入。例如,可以在c末端进行截平并在e1环中进行插入。
本发明的蛋白中的hbcag序列的每个部分优选具有与天然的hbcag蛋白的相应序列的至少70%序列一致性,例如具有如seqidno:2所示的序列的蛋白。更优选地,所述一致性为至少80%、至少90%、至少97%、至少98%或至少99%。测定蛋白同源性的方法在本领域中是熟知的,本领域技术人员将会理解的是,在本说明书中,同源性是基于氨基酸一致性计算的(有时称为“硬同源性(hardhomology)”)。
例如uwgcg软件包(devereuxetal(1984)nucleicacidsresearch12:387-395)提供bestfit程序,它可以被用来计算同源性(例如使用其默认设置)。pileup和blast算法可以被用来计算同源性或对齐序列(通常使用它们的默认设置),例如在altschuls.f.(1993)jmolevol36:290-300;altschul,s,fetal(1990)jmolbiol215:403-10中描述的。
进行blast分析的软件可以通过美国国家生物技术信息中心(http://www.ncbi.nlm.nih.gov/)得到。该算法涉及首先识别出高分值序列对(hsp),其通过识别查询序列中的长度为w的短字段而进行,所述短字段在与数据库序列中相同长度的字段对齐时匹配或满足某些正值的阈值分值t。t是指相邻字段分值阈值(altschuletal,同上)。这些初始的相邻字段匹配用作启动查找包含它们的hsp的检索的种子。字段匹配沿各个序列在两个方向上延伸,只要累积的对齐分值可以被升高。当出现以下情况时字段匹配在每个方向上的延伸停止:累积的对齐分值从其最大达到值下降数量x;由于累积一个或多个负分值的残基对齐,累积分值达到零或以下;或达到了任一序列的末端。blast算法参数w、t和x决定了对齐的灵敏度和速度。blast程序的默认值如下:字段长度(w)为11;blosum62分值矩阵(参见heikoffandhenikoff(1992)proc.natl.acad.sci.usa89:10915-10919)对齐(b)为50;期望值(e)为10;m=5;n=4;并且比较双链。
blast算法进行两个序列之间的相似性的统计分析;参见例如,karlinandaltschul(1993)proc.natl.acad.sci.usa90:5873-5787。blast算法提供的相似性的一个量度为最小和概率(p(n)),它表示两个核苷酸序列或氨基酸序列偶然匹配的概率。例如,当第一序列与第二序列比较时,如果最小和概率小于约1、优选小于约0.1、更优选小于约0.01、最优选小于约0.001,则认为一个序列与另一个序列相似。
hbcag的e1环位于成熟序列的第68至90位,免疫原性多肽可以被插入这些位置之间的任何位置。优选地,免疫原性多肽被插入进第69至90、71至90或75至85的区域中。最优选的是将用于免疫原性多肽插入到第79和第80个氨基酸残基之间或第80和第81个残基之间。当插入免疫原性多肽时,hbcag的整个序列可以被维持,或者替代地e1环序列的整个或其一部分可以被缺失并被蛋白序列取代。因此,第69至90个、第71至90个或第75至85个氨基酸残基可以被免疫原性多肽取代。当免疫原性多肽取代e1环序列时,取代序列通常不短于被其取代的序列。
hbcag的c末端截平通常不超出aa144,因为如果进行任何进一步的截平,就可能不形成颗粒。因此,被删除的氨基酸可以包括例如aa144至c末端aa(aa183或185)、aa150至c末端aa、aa164至c末端aa或aa172至c末端aa。hbcag的c末端结合dna,因此c末端的截平降低或完全从hbcag和hbcag杂合蛋白的制剂中移除dna。
本发明的蛋白形成颗粒,其优选类似于由天然hbcag形成的颗粒。本发明的颗粒包含本发明的一个或多个蛋白的多个拷贝。所述颗粒可以是vlp的形式。本发明的颗粒通常直径为至少10nm,例如直径为10至50nm或20至40nm,但优选地,它们的直径为约27nm(这是天然hbcag颗粒的尺寸)。它们包括多个hbcag单元,例如150至300个单元,但通常它们被固定至约180或约240个单元(这是天然hbcag颗粒中单元的数量)。因为本发明的蛋白可以是二聚体,这意味着颗粒中的蛋白单体的数量可以是75至150个,但通常是约90或约120个。
组成hbc穗区域的两个α螺旋不是对称的,因此所得到的mir不会从vlp完全竖直地指向,而是略微偏移。因此分子模型表明已插入的任何抗原可以平行于vlp,而不是成直角。这有可能导致空间位阻和免疫原性的降低。hbcag可以包含这样的插入序列,它通过将一个或多个额外的转向增加至第一螺旋(其位于成熟序列的第50至73位)从而“平衡”α螺旋。这导致出现垂直定向至vlp的被插入的蛋白表位。这可以通过将3至12个氨基酸(例如3、5或7个氨基酸)插入至hbcag来实现。这些氨基酸优选是不带电荷的氨基酸,例如丙氨酸、亮氨酸、丝氨酸和苏氨酸。被插入的序列优选是aaalaaa(seqidno:15)。插入可以在成熟序列的第50和75个氨基酸之间的位点,例如在第60和75个残基或第70和73个残基之间的位点。
本发明的颗粒可以包括一个以上本发明的蛋白,即混合的颗粒。发明人发现,将长度小于20个氨基酸的“空”插入体以及如本文进一步描述的插入进串联构建体中的hbcag的一个拷贝,使得在hbcag的另一个拷贝中的被插入的抗原能够折叠和/或被正确地呈现。包括在hbcag的一个拷贝中的一个类型的被插入的抗原以及在hbcag的另一个拷贝中的“空”插入体的蛋白,与包括在hbcag的一个拷贝中的不同类型的被插入的抗原以及在hbcag的另一个拷贝中的“空”插入体的蛋白,可以被组合进相同的颗粒。这使得所述颗粒能够包括两个类型的被插入的抗原的多个拷贝,这些抗原被很好地隔开并且处于稳定的形式。被插入的抗原本质上被放置得相互之间存在“分隔物”,由此提供空间来正确地折叠。单体的hbcag不会能够实现该效果。
接头
在相邻的hbcag单元之间、在el环中的插入体侧翼和/或连接el环中的相邻的插入体的接头,通常是长度至少为1.5nm(15å)的氨基酸的链,例如1.5至10nm、1.5至5nm或1.5至3nm。例如,它可以包括4至40aa或10至30aa,优选15至21aa。所述接头通常是柔性的。在接头中的氨基酸可以例如包括甘氨酸、丝氨酸和/或脯氨酸,或者完全由它们组成。例如,接头可以包括序列glynser(gns)的一个或多个重复,其中n是2、3、4、5、6、7或8。优选的接头包括序列glyglyglyglyser(ggggs)(seqidno:4)的一个、两个或更多个重复。例如,ggggsggggsggggs(seqidno:5)。替代地,接头可以包括一个或多个glypro(gp)二肽重复。重复的数量可以例如是1至18,优选3至12。在g2s重复的情况下,5、6或7个重复的使用已经被发现允许颗粒的形成。在相邻的hbcag单元之间的优选的接头是g2s的7个重复。接头可以对应于抗体的铰链区;该铰链区被认为提供抗体的抗原结合和尾部结构域之间的柔性连接。
包含接头的串联构建体的结构的例子包括以下:
[hbcag的“第一拷贝”的对于el环是n末端的部分]-[第一接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第二接头]-[hbcag的“第一拷贝”的对于el环是c末端的部分]-[第三接头]-[hbcag的“第二拷贝”的对于el环是n末端的部分]-[任选的免疫原性多肽(例如ha茎)]-[hbcag的“第二拷贝”的对于el环是c末端的部分]。
如果在串联构建体中存在一个以上的接头,则它们可以是相同的或相互不同的。例如,它们可以是全部相同的,它们可以是全部相互不同的,两个或以上接头可以是相同的但是与一个或以上其他接头不同,等等。如果在“第二拷贝”中存在免疫原性多肽,则在每个el环中被插入的序列可以在一侧或两侧上具有接头。如果在两个插入体的两侧都存在接头,则串联构建体的结构的例子包括以下:
[hbcag的“第一拷贝”的对于el环是n末端的部分]-[第一接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第二接头]-[hbcag的“第一拷贝”的对于el环是c末端的部分]-[第三接头]-[hbcag的“第二拷贝”的对于el环是n末端的部分]-[第四接头]-[任选的免疫原性多肽(例如ha茎)]-[第五接头]-[hbcag的“第二拷贝”的对于el环是c末端的部分]。
本发明的一个方面的串联核心构建体包括在hbcag的一个拷贝的el环中的流感病毒a表面多肽m2或其免疫原性片段,以及任选的在hbcag的另一个拷贝的el环中的另一个免疫原性多肽。在hbcag的另一个拷贝中的所述免疫原性多肽(在以上设置中被描述为“任选的免疫原性多肽”)可以是本文描述的任何免疫原性多肽。因此,所述免疫原性多肽可以是另一种流感多肽或其免疫原性片段。它可以是流感病毒a表面多肽m2或其免疫原性片段。因此,可以在hbcag的两个拷贝的el环中都存在流感病毒a表面多肽m2或其免疫原性片段。在每个el环中的流感病毒a表面多肽m2或其免疫原性片段可以是相同的或不同的。所述免疫原性多肽可以衍生自ha。例如,所述免疫原性多肽可以是ha茎或其免疫原性片段。因此,可以在hbcag的一个拷贝的el环中存在流感病毒a表面多肽m2或其免疫原性片段,并在hbcag的另一个拷贝的el环中存在ha茎或其免疫原性片段。
本发明的另一个方面的串联核心构建体包括在hbcag的一个拷贝的el环中的流感病毒血凝素(ha)多肽或其免疫原性片段。hbcag的另一个拷贝的el环包括小于20个氨基酸的序列。
如本文所述,在hbcag的一个拷贝的el环中可以存在免疫原性多肽的一个或多个拷贝。例如,在一个el环中可以存在流感病毒a表面多肽m2或其免疫原性片段的一个或多个拷贝。在一个el环中可以存在ha茎或其免疫原性片段的一个或多个拷贝。在一个el环中可以存在多达免疫原性多肽的2、3、4、6或8个拷贝。在每个el环中可以存在免疫原性多肽的多个拷贝。例如,串联构建体可以包括在一个hbcag的el环中的流感病毒a表面多肽m2或其免疫原性片段的一个、两个、三个、四个或五个拷贝,以及在另一个hbcag的el环中的ha茎或其免疫原性片段的一个、两个或三个拷贝。因此,串联构建体可以包括在一个hbcag的el环中的流感病毒a表面多肽m2或其免疫原性片段的三个拷贝,以及在另一个hbcag的el环中的ha茎或其免疫原性片段的一个拷贝。进一步地,串联构建体可以包括在一个hbcag的el环中的流感病毒a表面多肽m2或其免疫原性片段的三个拷贝,以及在另一个hbcag的el环中的ha茎或其免疫原性片段的两个或三个拷贝。在一个el环中的每个拷贝可以是相同的或不同的。例如,在一个el环中可以存在流感病毒a表面多肽m2或其免疫原性片段的两个、三个、四个或五个(优选三个)不同的序列。在被插入进一个el环的免疫原性多肽的拷贝之间可以存在接头。因此,可以存在将一个或多个免疫原性多肽连接至el环的接头,以及将被插入进一个el环中的多个免疫原性多肽连接在一起的接头。例如,串联构建体的设置可以如下:
[hbcag的“第一拷贝”的对于el环是n末端的部分]-[第一接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第二接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第三接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第四接头]-[hbcag的“第一拷贝”的对于el环是c末端的部分]-[第五接头]-[hbcag的“第二拷贝”的对于el环是n末端的部分]-[任选的免疫原性多肽(例如ha茎)]-[hbcag的“第二拷贝”的对于el环是c末端的部分]。
流感病毒a表面多肽m2(m2)
本发明的蛋白的目的是它能够被用来诱导对流感(特别是流感病毒a)的免疫反应,并且因此能够被用作流感疫苗。本发明的一个方面的蛋白具有被插入进hbcag的一个或两个拷贝的el环中的流感病毒a表面多肽m2或其免疫原性片段。本发明的蛋白可以具有被插入进hbcag的两个拷贝的el环中的m2或其免疫原性片段。流感病毒a表面多肽m2是质子选择性的离子通道蛋白,其被整合在流感病毒的病毒包膜中。通道本身是同源四聚体(由四个相同的m2单元组成),其中所述单元由两个二硫键键稳定的螺旋。流感病毒a表面多肽m2单元由三个蛋白结构域组成:在n末端的24个氨基酸,暴露至外侧环境;在跨膜区上的19个疏水氨基酸;以及在c末端的54个氨基酸,被朝着病毒颗粒的内侧定向。来自流感a病毒菌株a/34/pr8的m2蛋白的全长序列被示出于seqidno:3中。
将被插入进hbcag的el环的流感病毒a表面多肽m2衍生自流感病毒a。它可以衍生自seqidno:3中的序列。全长的流感病毒a表面多肽m2,即全长97个氨基酸序列,可以被插入进hbcag的el环。例如,seqidno:3的全长序列可以被插入。流感病毒a表面多肽m2可以是天然存在的m2蛋白或者可以是天然存在流感病毒a表面多肽m2的变种。
流感病毒a表面多肽m2或其免疫原性片段的一个以上拷贝可以被插入hbcag的一个或两个拷贝的el环。例如,流感病毒a表面多肽m2或其免疫原性片段的2、3、4、5、6、7或8个拷贝可以被插入hbcag的一个或两个拷贝的el环。例如,1、2或3个拷贝可以被插入hbcag的一个或两个拷贝的el环。因此,流感病毒a表面多肽m2或其免疫原性片段的1、2或3个拷贝可以被插入两个el环或el环中的一个。在el环中存在流感病毒a表面多肽m2或其免疫原性片段的一个以上拷贝的时候,用于每个拷贝的序列可以是相同的或可以是不同的。如果序列是不同的,则它们可以以任意顺序被插入。例如,“第一拷贝”、“第二拷贝”、“第三拷贝”等可以是从el环中的n末端至c末端的任意顺序。例如,“第三拷贝”可以是对于“第一拷贝”的n末端。
流感病毒a表面多肽m2序列被记载于seqidno:3中。流感病毒a表面多肽m2的序列可以与seqidno:3或任何天然存在的流感病毒a表面多肽m2具有同源性,例如在整个序列上或者在至少20个、例如至少30个、至少40个、至少50个、至少60个、至少80个或更多个连续的氨基酸的区域上,例如至少60%、至少70%、至少80%、至少90%、至少95%、至少97%、至少98%或至少99%的一致性。测定蛋白同源性的方法在本领域中是公知的,并且在上文中关于hbv核心蛋白进行了讨论。
同源蛋白通常通过取代、插入或缺失而与天然存在的流感病毒a表面多肽m2序列不同,例如1、2、3、4、5或8个或更多个的取代、插入或缺失。取代优选是“保守的”,并且可以例如根据表1进行。在第二列的同一区块的氨基酸,优选在第三列的同一行的氨基酸可以相互取代。
表1
流感病毒a表面多肽m2或其免疫原性片段插入体的序列可以衍生自流感类型a的任何亚型(参见例如sharp2002cellvol.108,305-312,“originsofhumanvirusdiversity”和shietal2010,plosone,5(12)“acompleteanalysisofhaandnagenesofinfluenzaaviruses”)。例如,来自ha亚型的任何一个,例如h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15或h16,和/或来自na亚型的任何一个,例如n1、n2、n3、n4、n5、n6、n7、n8或n9。优选地,流感病毒a表面多肽m2或其免疫原性片段插入体可以衍生自h1n1、h5n1、h3n2、h7n7、h1n2、h2n2、h7n3、h5n2、h1n7、h9n2、h7n2或h10n7。更优选地,流感病毒a表面多肽m2或其免疫原性片段插入体可以衍生自h3n2、h5n1、h1n1或h7n7。
将被用作插入体的流感病毒a表面多肽m2的免疫原性片段是全长流感病毒a表面多肽m2的缩短版本,其保留了诱导免疫反应的能力。在一些情况下,片段可以是天然存在的流感病毒a表面多肽m2序列或seqidno:3的序列的长度的至少10,例如至少20%、至少30%、至少40%或至少50%、优选至少60%、更优选至少70%、更优选至少80%、甚至更优选至少90%和更优选至少95%。例如,片段的长度可以是6至96个aa、6至50个aa或6至25个aa。
优选地,所述免疫原性片段来自流感病毒a表面多肽m2的暴露在病毒粒子的表面上的区域。优选地,所述免疫原性片段是流感病毒a表面多肽m2胞外域(m2e),它是流感病毒a表面多肽m2蛋白的外部结构域。m2e的序列可以是通用m2e共识序列。seqidno:9示出了通用序列,其在第17和19位的半胱氨酸已经被丝氨酸取代。m2e的序列可以是通用m2e共识序列的变种。m2e的序列可以与通用m2e共识序列或任何天然存在的m2e具有同源性,例如在整个序列上或者在至少8个、例如至少10个、至少15个或至少20个或更多个连续的氨基酸的区域上,例如至少60%、至少70%、至少80%、至少90%、至少95%、至少97%、至少98%或至少99%的一致性。测定蛋白同源性的方法在本领域中是公知知的,并且在上文中关于hbv核心蛋白进行了讨论。所述序列可以通过一个或多个氨基酸的取代、添加和/或缺失而改变。例如,可以存在多达18个、多达15个、多达12个、多达10个或多达5个取代、缺失或添加。所述序列可以仅通过缺失、仅通过添加或仅通过取代而改变。所述序列可以通过缺失和添加、缺失和取代、添加和取代、或者缺失、添加和取代的组合而改变。优选地,存在一个、两个、三个、四个、五个或六个缺失、添加和取代。取代优选是“保守的”,并且可以例如根据上述的表1进行。在实施例1和3中使用了m2e的取代形式的例子。m2e的序列可以衍生自流感病毒a的任何亚型,例如在上文关于流感病毒a表面多肽m2而列出的。优选地,使用m2e最常见的变种。例如,参见在实施例1和3中使用的m2e的序列。流感病毒a表面多肽m2的免疫原性片段可以是这些序列的任何一个。这些序列中的一些在第17和/或19位的半胱氨酸已经被丝氨酸取代。可以通过这种方法如下文进一步讨论地修改流感病毒a表面多肽m2或其免疫原性片段(例如m2e)的任何一个。衍生自流感a病毒菌株a/34/pr8的m2e序列是seqidno:3的前24个氨基酸。流感病毒a表面多肽m2的片段可以包括或者可以是seqidno:3的第1至24个氨基酸。
在流感病毒a表面多肽m2中的半胱氨酸的取代或缺失
在流感病毒a表面多肽m2或其免疫原性片段的第17位的半胱氨酸氨基酸、第19位的半胱氨酸氨基酸或者第17和19位的半胱氨酸氨基酸,可以被删除或者被替代的氨基酸取代。可以存在缺失和取代的组合。例如,在第17位的半胱氨酸被删除而在第19位的半胱氨酸被取代,或者在第17位的半胱氨酸被取代而在第19位的半胱氨酸被删除。流感病毒a表面多肽m2或其免疫原性片段的第17和19位是来自成熟的流感病毒a表面多肽m2的n末端的第17和19位。例如,seqidno:3的第17和19位。如上描述的,流感病毒a表面多肽m2的片段可以是m2e。因此,在m2e的的第17位的半胱氨酸、第19位的半胱氨酸或者第17和19位的半胱氨酸,可以被删除或者取代为替代的氨基酸。
“替代的氨基酸”可以是不为半胱氨酸并且使得vlp能够形成从而反过来能够诱导对象中的免疫反应的任何氨基酸。取代优选是“保守的”,并且可以例如根据上述表1进行。半胱氨酸优选被丝氨酸、苏氨酸或甲硫氨酸取代。丝氨酸是最优选的。在实施例1的表2中示出了一个或两个半胱氨酸被取代的示例性m2e序列。
免疫原性多肽
串联核心系统的灵活性是指,本发明的蛋白除了包括m2或其免疫原性片段之外,可以包括一个或多个进一步的免疫原性多肽。例如,本发明的蛋白可以包括一个或多个进一步的流感病毒衍生的免疫原性多肽,从而诱导对于流感病毒的优秀的免疫反应。替代地,本发明的蛋白可以包括一个或多个衍生自不同的来源(例如不同的病原体或过敏原)的免疫原性多肽,从而同时诱导对于流感病毒和对于不同的病原体或过敏原的免疫反应。因此,虽然本发明的蛋白必须具有被插入至hbcag的至少一个拷贝的el环的流感病毒a表面多肽m2或其免疫原性片段,但是在该蛋白中的hbcag的另一个拷贝的el环可以包括任何其他类型的免疫原性多肽。
所述免疫原性多肽包括能够诱导免疫反应的氨基酸的序列。所述免疫原性多肽可以是构象性的或线性的。它可以是例如6至600个aa、6至300个aa、6至200个aa、50至200个aa、100至200个aa、6至120个aa、20至90个aa、40至90个aa或60至90个aa的序列。
较大的和/或疏水性的插入可以被容纳而不破坏vlp。被用作插入物的免疫原性多肽可以是不破坏vlp形成的任何适当的尺寸。其优选小于100kda,例如小于80kda、小于60kda、小于40kda、小于20kda、小于10kda或小于5kda。其可以多于5kda、10kda、20kda或30kda。
本发明的蛋白可以含有多于一个免疫原性多肽,例如多达2、3、4、6或8个免疫原性多肽。免疫原性多肽的多于一个的拷贝可以被插入到hbcag的一个或两个拷贝中;例如,可以插入2至8个拷贝,例如可以插入2、3、4、5、6、7或8个拷贝。当本发明的蛋白中有两个或更多个免疫原性多肽时,它们可以来自于相同或不同的生物体以及来自相同或不同的蛋白。
所述免疫原性多肽可以包括一个或多个t细胞表位或b细胞表位。如果它包括t细胞表位,则它可以是细胞毒性t-淋巴细胞(ctl)表位或t辅助细胞(th)表位(例如th1或th2表位)。在本发明的一个优选实施方式中,免疫原性多肽包括t辅助细胞表位和b细胞或ctl表位。t辅助细胞表位的存在增强了对b细胞或ctl表位的免疫反应。
免疫原性多肽的选择取决于疫苗接种期望对抗的疾病。例如,所述免疫原性多肽可以来自病原性生物、癌症相关的抗原或过敏原。所述病原性生物可以例如是病毒、细菌或原生动物。
所述免疫原性多肽可以衍生自任何病原体,所述病原体例如但不限于病毒,包括正粘病毒科(orthomyxoviridae)(包括例如流行性感冒a、b和c病毒)、腺病毒科(adenoviridae)(包括例如人腺病毒)、杯状病毒科(caliciviridae)(例如诺沃克病毒群(norwalkvirusgroup))、疱疹病毒科(herpesviridae)(包括例如hsv-1、hsv-2、ebv、cmv和vzv)、乳多空病毒科(papovaviridae)(包括例如人类乳头状瘤病毒(hpv))、痘病毒科(poxviridae)(包括例如天花病毒和牛痘病毒)、小dna病毒科(parvoviridae)(包括例如细小病毒b19)、呼肠孤病毒科(reoviridae)(包括例如轮状病毒)、冠状病毒科(coronaviridae)(包括例如sars)、黄病毒科(flaviviridae)(包括例如黄热病毒、西尼罗河病毒、登革热病毒、丙型肝炎病毒和森林脑炎病毒)、小rna病毒科(picornaviridae)(包括肠道病毒、脊髓灰质炎病毒、鼻病毒和甲型肝炎病毒)、披膜病毒科(togaviridae)(包括例如风疹病毒)、丝状病毒科(filoviridae)(包括例如马尔堡病毒和埃博拉病毒)、副粘液病毒科(paramyxoviridae)(包括副流感病毒、呼吸道合胞病毒(rsv)、流行性腮腺炎病毒和麻疹病毒)、弹状病毒科(rhabdoviridae)(包括例如狂犬病病毒)、尼亚病毒科(bunyaviridae)(包括例如汉坦病毒)、逆转录病毒(retroviridae)(包括例如hiv和人类t细胞淋巴瘤病毒(htlv))和肝病毒科(hepadnaviridae)(包括例如乙型肝炎病毒)的成员。
所述免疫原性多肽可以衍生自细菌,所述细菌包括伯克霍尔德氏菌(burkholderia)、结核分枝杆菌(m.tuberculosis)、衣原体(chlamydia)、淋病奈瑟菌(n.gonorrhoeae)、志贺氏杆菌(shigella)、沙门氏菌(salmonella)、霍乱弧菌(vibriocholera)、梅毒螺旋体(treponemapallidua)、假单胞菌属(pseudomonas)、百日咳杆菌(bordetellapertussis)、布鲁氏菌(brucella)、土拉弗朗西斯菌(franciscellatulorensis)、幽门螺杆菌(helicobacterpylori)、钩端螺旋体菌(leptospriainterrogaus)、嗜肺军团菌(legionellapnumophila)、鼠疫杆菌(yersiniapestis)、链球菌(streptococcus,a型和b型)、肺炎球菌(pneumococcus)、脑膜炎球菌(meningococcus)、流感嗜血杆菌(hemophilusinfluenza,b型)、弯曲杆菌病(complybacteriosis)、卡他莫拉菌(moraxellacatarrhalis)、杜诺凡病(donovanosis)和放线菌(actinomycosis)、真菌病原体(包括念珠菌(candidiasis)和曲霉病(aspergillosis))、以及寄生虫病原体(包括弓形虫(toxoplasmagondii)、绦虫(taenia)、吸虫(flukes)、蛔虫(roundworms)、扁虫(flatworms)、阿米巴病(amebiasis)、贾第虫病(giardiasis)、隐孢子虫(cryptosporidium)、血吸虫(schitosoma)、卡氏肺孢子虫(pneumocystiscarinii)、滴虫病(trichomoniasis)和旋毛虫病(trichinosis))。
所述免疫原性多肽可以衍生自通过a)呼吸道、b)泌尿生殖系统或c)胃肠道进行感染的病原体。此类病原体的例子包括a)腺病毒(adenoviridae)、副粘液病毒科(paramyxoviridae)和痘病毒科(poxviridae)、鼻病毒、流行性感冒、和汉坦病毒的成员;b)解脲支原体(ureaplasmaurealyticum)、淋球菌(neisseriagonorrhoeae)、阴道加德菌(gardnerellavaginalis)、阴道毛滴虫(trichomonasvaginalis)、梅毒螺旋体(treponemapallidum)、沙眼衣原体(chlamydiatrachomatis)、杜克雷嗜血杆菌(haemophilusducreyi)、单纯疱疹病毒(herpessimplexvirus)、hpv、hiv、白色念珠菌(candidaalbicans)、梅毒螺旋体(treponemapallidum)、以及肉芽肿荚膜杆菌(calmatobacterium);以及c)志贺氏菌(shigella)、沙门氏菌(salmonella)、霍乱弧菌(vibriocholera)、大肠杆菌(e.coli)、溶组织内阿米巴(entamoebahistolytica)、弯曲杆菌属(campylobacter)、梭菌属(clostridium)、耶尔森氏菌属(yersinia)、轮状病毒、诺如病毒、腺病毒、星状病毒、蛔虫、扁虫、贾第鞭毛虫病(giardiasis)、以及隐孢子虫(cryptosporidium)。
将在本发明中使用的免疫原性多肽可以衍生自癌症,所述癌症例如但不限于肺癌、胰腺癌、肠癌、结肠癌、乳房癌、子宫癌、宫颈癌、卵巢癌、睾丸癌、前列腺癌、黑色素瘤、卡波济氏肉瘤、淋巴瘤(例如ebv诱导的b细胞淋巴瘤)和白血病。肿瘤相关的抗原的具体例子包括但不限于:癌症睾丸抗原(例如mage家族(mage1、2、3等)的成员、ny-eso-1和ssx-2)、分化抗原(例如酪氨酸酶、gp100、psa、her-2和cea)、突变的自身抗原和病毒性肿瘤抗原(例如来自致癌性hpv类型的e6和/或e7)。特定的肿瘤抗原的进一步的例子包括mart-1、melan-a、p97、β-hcg、gainac、mage-1、mage-2、mage-4、mage-12、muc1、muc2、muc3、muc4、muc18、cea、ddc、p1a、epcam、黑色素瘤抗原gp75、hker8、高分子量的黑色素瘤抗原、k19、tyrl、tyr2、pmel17基因家族的成员、c-met、psm(前列腺粘蛋白抗原)、psma(前列腺特异膜抗原)、前列腺分泌蛋白、甲胎蛋白、ca125、ca19.9、tag-72、brca-1和brca-2抗原。
本发明使用的其它候选的免疫原性多肽的例子包括以下抗原:流感抗原ha(血凝素)、na(神经氨酸酶)、np(核蛋白/核壳蛋白)、m1、m2、pb1、pb2、pa、ns1和ns2;hiv抗原gp120、gp160、gag、pol、nef、tat和ref;疟疾抗原cs蛋白和子孢子表面蛋白2;疱疹病毒抗原ebvgp340、ebvgp85、hsvgb、hsvgd、hsvgh、hsv早期蛋白产物、巨细胞病毒gb、巨细胞病毒gh和ie蛋白gp72;人类乳头状瘤病毒抗原e4、e6和e7;呼吸道合胞病毒抗原f蛋白、g蛋白和n蛋白;百日咳杆菌(b.pertussis)的百日咳杆菌粘附素抗原、肿瘤抗原癌cea、癌相关的粘蛋白、癌p53、黑色素瘤mpg、黑色素瘤p97、mage抗原、癌neu的癌基因产物、前列腺特异性抗原(psa)、前列腺相关的抗原、ras蛋白和myc;以及屋尘螨过敏原。
优选地,所述免疫原性多肽衍生自流感病毒。所述免疫原性多肽可以衍生自流感病毒a、b或c。优选地,它衍生自流感病毒a。所述免疫原性多肽可以衍生自任何流感抗原,例如ha(血凝素)、na(神经氨酸酶)、np(核蛋白/核衣壳蛋白)、m1、m2、pb1、pb2、pa、ns1和ns2抗原,特别是m2、ha和na抗原。所述免疫原性多肽优选是上述的流感病毒a表面多肽m2或其免疫原性片段。
所述免疫原性多肽优选是血凝素(ha)或其免疫原性片段。关于流感病毒a表面多肽m2或其免疫原性片段的上述讨论也涉及ha。ha的序列可以与任何天然存在的ha具有同源性,例如在整个序列上或者在至少20个、例如至少30个、至少50个、至少70个、至少100个、至少150个、至少200个或更多个连续的氨基酸的区域上,例如至少60%、至少70%、至少80%、至少90%、至少95%、至少97%、至少98%或至少99%的一致性。测定蛋白同源性的方法在本领域中是公知的,并且在上文中关于hbv核心蛋白进行了讨论。
所述免疫原性多肽优选是ha或其免疫原性片段。ha的片段可以衍生自ha1或ha2。片段的长度可以是6至565个aa、6至300个aa、6至200个、或6至100个aa。ha的片段可以是ha2,它被称为ha的茎区域。图10示出了ha茎区域的结构。所述片段可以包括ha2单体的已知的结构域环b、螺旋c、螺旋cd和螺旋d(参见图10)。在表4中示出了用于插入在el环中的ha的片段的例子。被插入的ha茎序列可以是seqidno:12的第80至151个氨基酸。ha茎的序列可以与任何天然存在的ha茎具有同源性,例如在整个序列上或者在至少8个、例如至少10个、至少20个、至少30个、至少50个、至少60个、至少70个或更多个连续的氨基酸的区域上,例如至少60%、至少70%、至少80%、至少90%、至少95%、至少97%、至少98%或至少99%的一致性。测定蛋白同源性的方法在本领域中是公知的,并且在上文中关于hbv核心蛋白进行了讨论。所述序列可以通过一个或多个氨基酸的取代、添加和/或缺失而改变。例如,可以存在多达18个、多达15个、多达12个、多达10个或多达5个取代、缺失或添加。所述序列可以仅通过缺失、仅通过添加或仅通过取代而改变。所述序列可以通过缺失和添加、缺失和取代、添加和取代、或者缺失、添加和取代的组合而改变。优选地,存在一个、两个、三个、四个、五个或六个缺失、添加和取代。取代优选是“保守的”,并且可以例如根据上述的表1进行。
ha或其免疫原性片段的一个以上的拷贝可以被插入进el环。例如,ha或其免疫原性片段的2、3、4、5、6、7或8个拷贝可以被插入el环。例如,1、2或3个拷贝可以被插入el环。因此,ha或其免疫原性片段的1、2或3个拷贝可以被插入el环。在el环中存在ha或其免疫原性片段的一个以上拷贝的时候,用于每个拷贝的序列可以是相同的或可以是不同的。如果序列是不同的,则它们可以以任意顺序被插入。例如,“第一拷贝”、“第二拷贝”、“第三拷贝”等可以是从el环中的n末端至c末端的任意顺序。例如,“第三拷贝”可以是对于“第一拷贝”的n末端。
根据本发明的一个方面,串联构建体包括在hbcag的“第一拷贝”中的流感病毒a表面多肽m2或其免疫原性片段,以及在hbcag的“第二拷贝”中的ha或其免疫原性片段。如上所述,“第一拷贝”可以是n末端或c末端拷贝。
在hbcag的一个拷贝中的“空”插入体
根据本发明的另一个方面,串联构建体包括在hbcag的一个拷贝中的“空”插入体。发明人发现,这使得在hbcag的另一个拷贝中的抗原能够折叠和/或被正确地呈现。“空”插入体是短的序列,通常长度小于20个氨基酸,它使得在hbcag的另一个拷贝中的抗原能够折叠和/或被正确地呈现。
更详细地,发明人将来自流感h3n2病毒ha2蛋白结构域(lah3)的保守区域插入串联构建体中的hbcag的一个拷贝。他们发现,与在单体核心中表达lah3相比,将小于20个氨基酸的短序列插入进hbcag的第二拷贝,使得第一插入体(lah3)正确地配置并且赋予整个vlp更大的溶解度(实施例4)。特别地,他们将以下序列插入进hbcag的第二拷贝,该序列单一的赖氨酸(k)残基,其在侧翼有由甘氨酸和丝氨酸残基形成的短的柔性的接头区域(“空”插入体)。这样的插入体可以被用在串联构建体中的hbcag的一个拷贝中,该串联构建体在hbcag的第二拷贝中具有任意抗原,从而帮助抗原折叠和/或正确地呈现。
因此,本发明提供了一种蛋白,其包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个拷贝包括在el环中的在一侧或两侧具有接头的流感病毒a表面多肽m2或其免疫原性片段,所述接头将所述多肽或片段连接至hbcag序列,并且hbcag的另一个拷贝包括在el环中的少于20个氨基酸的序列。
因此,本发明提供了一种蛋白,其包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代,并且hbcag的另一个拷贝包括在el环中的少于20个氨基酸的序列。
根据本发明的这些方面的任何一个,所述m2或其片段可以是上述的m2或其片段。
本发明还提供了一种蛋白,其包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个拷贝包括在el环中的ha或其免疫原性片段,其中ha的片段任选地是ha茎区域,并且hbcag的另一个拷贝包括在el环中的少于20个氨基酸的序列。
根据本发明的该方面,所述ha或其片段可以是上述的ha或其片段。所述ha的片段可以例如来自ha2蛋白结构域,例如来自流感h3n2病毒,使得该蛋白能够被用来免疫流感h3n2病毒感染。例如,所述片段可以包括分离自流感a病毒h3n2的ha蛋白的第421至475个氨基酸。所述片段可以包括seqidno:24或与该序列具有同源性的序列。与天然存在的ha序列具有同源性的序列是如上所述的。同样如上所述地,可以存在ha或其片段的一个以上的拷贝。
hbcag的第二拷贝包括el环中的短序列,即“空”插入体。与在单体hbcag核心中的hbca的第一拷贝中表达插入体相比,该序列使得hbca的第一拷贝中的插入体正确地配置和/或赋予整个vlp更大的溶解度。所述序列的长度小于20个氨基酸,例如长度小于或等于18个、小于或等于15个、小于或等于12个、小于或等于10个、小于或等于5个、小于或等于3个氨基酸。例如,序列的长度可以是18、16、14、12、10、8、6、4、2、1或0个氨基酸。序列的长度可以是14个氨基酸,如seqidno:21。所述序列可以包括赖氨酸(k)残基,其在每侧具有接头序列(在以下结构中的“第二”和“第三”接头)。所述接头通常是柔性的。在接头中的氨基酸可以例如包括甘氨酸和丝氨酸残基或者完全由它们组成。赖氨酸残基可以例如在侧翼有接头序列,该接头序列包括1至10个甘氨酸或丝氨酸残基,例如2至9个、或3至7个甘氨酸或丝氨酸残基。
包括m2或其免疫原性片段的串联构建体的结构的例子包括以下:
[hbcag的第一拷贝的对于el环是n末端的部分]-[第一接头]-[流感病毒a表面多肽m2或其免疫原性片段]-[第二接头]-[hbcag的第一拷贝的对于el环是c末端的部分]-[第三接头]-[hbcag的第二拷贝的对于el环是n末端的部分]-[第四接头]-[赖氨酸(k)残基]-[第五接头]-[hbcag的第二拷贝的对于el环是c末端的部分]。
包括ha或其免疫原性片段的串联构建体的结构的例子包括以下:
[hbcag的第一拷贝的对于el环是n末端的部分]-[血凝素(ha)或其免疫原性片段]-[hbcag的第一拷贝的对于el环是c末端的部分]-[第一接头]-[hbcag的第二拷贝的对于el环是n末端的部分]-[第二接头]-[赖氨酸(k)残基]-[第三接头]-[hbcag的第二拷贝的对于el环是c末端的部分]。
hbcag的第二拷贝可以包括在el环中的seqidno:21的序列、或同源性序列。所述序列可以与seqidno:21具有同源性,例如在整个序列上或者在至少6个、例如至少8个、至少10个、至少12个或更多个连续的氨基酸的区域上,例如至少60%、至少70%、至少80%、至少90%、至少95%、至少97%、至少98%或至少99%的一致性。测定蛋白同源性的方法在本领域中是公知的,并且在上文中关于hbv核心蛋白进行了讨论。所述序列可以通过一个或多个氨基酸的取代、添加和/或缺失而改变。例如,可以存在多达8个、多达6个、多达4个或多达2个取代、缺失或添加。所述序列可以仅通过缺失、仅通过添加或仅通过取代而改变。所述序列可以通过缺失和添加、缺失和取代、添加和取代、或者缺失、添加和取代的组合而改变。优选地,存在一个、两个、三个、四个、五个或六个缺失、添加和取代。取代优选是“保守的”,并且可以例如根据上述的表1进行。
可以在包括蛋白的多个拷贝的颗粒中表达包括在hbcag的一个拷贝中的抗原和在hbcag的第二拷贝中的空插入体的蛋白。替代地,包括在hbcag的一个拷贝中的抗原(例如m2或其免疫原性片段)和在hbcag的第二拷贝中的空插入体的蛋白,与包括在hbcag的一个拷贝中的第二抗原(例如ha或其免疫原性片段)和在hbcag的第二拷贝中的空插入体的蛋白,可以被联合使用来制造混合颗粒。所述空插入体使得两个抗原能够被良好地隔开地呈现,得到稳定的颗粒。
制备本发明的蛋白
本发明的蛋白通常通过重组dna技术制备蛋白。本发明包括编码本发明的蛋白(例如表达载体)的核酸分子(例如dna或rna)。可以利用已知的操纵核酸的技术制备核酸分子。通常,制备编码两个hbcag单元的两个独立的dna构建体,然后通过重叠pcr连接在一起。
可以通过在表达蛋白的条件下,培养包含编码该蛋白的核酸分子的宿主细胞,以及回收该蛋白来生产本发明的蛋白。合适的宿主细胞包括细菌(例如e.coli)、酵母菌、哺乳动物细胞和其他的真核细胞(例如昆虫sf9细胞)。
可以通过利用编码本发明的蛋白的一个以上的核酸分子转化宿主细胞,同时制备一个以上的本发明的蛋白。例如,可以利用以下核酸分子转化宿主细胞:编码以下蛋白的核酸分子:包括在hbcag的一个拷贝中的抗原(例如m2或其免疫原性片段)和在hbcag的第二拷贝中的空插入体的蛋白;以及编码以下蛋白的核酸分子:包括在hbcag的一个拷贝中的第二抗原(例如ha或其免疫原性片段)和在hbcag的第二拷贝中的空插入体的蛋白。可以在相同的或者分开的核酸分子上编码这两种蛋白。
构建根据本发明的核酸分子的载体可以是例如质粒载体或病毒载体。它们可含有复制的起点、用于表达编码蛋白的序列的启动子、启动子的调节子(例如增强子)、转录终止信号、翻译起始信号和/或翻译终止信号。所述载体也可以包含一个或多个选择性的标记物基因,例如在细菌质粒的情况中的氨苄青霉素抗性基因或在哺乳动物载体的情况中的新霉素抗性基因。载体可以在体外用于例如生产rna或用来转化或转染宿主细胞。所述载体也可以适于在体内被用在例如基因治疗或dna疫苗接种的方法中。
启动子、增强子和其他表达调节信号可以被选择来与表达载体所设计的宿主细胞兼容。例如,可以使用原核启动子,特别是适用于e.coli菌株(例如e.colihb101)中的启动子。可以使用其活性响应于周围环境(例如厌氧条件)中的变化而被诱导的启动子。优选地,可以使用htra或nirb启动子。这些启动子可以特别是被用来在减毒细菌中表达蛋白,例如被用作疫苗。当所述蛋白的表达在哺乳动物细胞中进行时,无论是在体外还是在体内进行,都可以使用哺乳动物启动子。也可以使用组织特异性启动子,例如肝细胞特异性启动子。也可以使用病毒启动子,例如莫洛尼鼠白血病病毒长末端重复序列(mmlvltr)、劳斯氏肉瘤病毒(rsv)ltr启动子、sv40启动子、人巨细胞病毒(cmv)ie启动子、单纯疱疹病毒启动子和腺病毒启动子。所有这些启动子都可从现有技术中容易得到。
本发明的蛋白可以使用用于纯化蛋白的常规技术来纯化。所述蛋白例如可以以纯化的、纯的或分离的形式提供。对于在疫苗中使用,所述蛋白通常必须以高水平的纯度提供,例如以在制剂中包含多于80%、多于90%、多于95%或多于98%的蛋白的水平。但是,在最终疫苗制剂中将所述蛋白与其他蛋白混合也是可取的。
诱导免疫反应
本发明的蛋白、颗粒或核酸可以被用来诱导免疫反应,特别是针对流感(例如流感病毒a)。所述蛋白、颗粒或核酸可以被用作疫苗。本发明提供了在对象中诱导免疫反应的方法,所述方法包括向所述对象施用本发明的蛋白、颗粒或核酸。佐剂可以与所述蛋白、颗粒或核酸联合施用。所述蛋白、颗粒或核酸可以被用来引起对所有组分(hbcag、流感病毒a表面多肽m2、或ha以及可能的一种或多种其他免疫原性多肽)的多个同时的免疫反应。如果所述免疫原性多肽也衍生自流感,则这可以诱导针对流感的增强的免疫反应。如果所述免疫原性多肽不衍生自流感,则这可以诱导针对所述免疫原性多肽的来源以及流感的同时的免疫反应。如果存在一种以上的免疫原性多肽以及流感病毒a表面多肽m2,则所述一种以上的免疫原性多肽可以来自一个来源(例如病原体或过敏原),或者来自不同的来源(例如一种以上的病原体或过敏原)。如果所述免疫原性多肽衍生自一种以上的来源,则这可以诱导针对不同的来源(例如一种以上的病原体或过敏原)的同时的免疫反应。本发明的优点之一是,它允许对在疫苗中被递送的不同的免疫原性多肽的比率的精确控制。例如,在hbcag的第一拷贝中的流感病毒a表面多肽m2与在hbcag的第二拷贝中的免疫原性多肽的比率可以精确地是1:1。
所述蛋白、颗粒或核酸可以单独使用或用作组合物的一部分,所述组合物包括但不限于药物组合物、疫苗组合物或免疫治疗组合物。因此本发明提供了一种药物组合物(例如疫苗组合物),其包括本发明的蛋白、包含本发明蛋白的多个拷贝的颗粒或编码本发明的蛋白的核酸分子、以及药学上可接受的载体或稀释剂。并且,如上解释的,本发明提供“混合的”颗粒,所述颗粒包括一种以上的本发明的蛋白。因此,本发明还提供包含这样的混合的颗粒的药物组合物、疫苗组合物或免疫治疗组合物。所述组合物可以进一步包括佐剂。所述组合物可以被用于人体或动物体的治疗。所述组合物可以被用于人体或动物体的疫苗接种。本发明提供了用于人类或动物个体的治疗的方法,所述方法包括向所述个体施用所述组合物。所述组合物可以被用来针对本文描述的任何病原体进行疫苗接种。特别地,所述组合物可以被用来针对流感(例如流感病毒a)进行疫苗接种。
本发明的蛋白、本发明的颗粒或本发明的核酸可以被用在人体或动物体的治疗的方法中。本发明提供了用于人类或动物个体的治疗的方法,所述方法包括向所述个体施用本发明的蛋白、本发明的颗粒或本发明的核酸。佐剂可以与所述蛋白、颗粒或核酸联合施用。本发明还提供了本发明的蛋白、本发明的颗粒或本发明的核酸用于制备用于人类或动物体的治疗的药物的应用。所述蛋白、颗粒或核酸可以被用来治疗本文描述的任何病原体。特别地,所述蛋白、颗粒或核酸可以被用于流感(例如流感病毒a)的治疗。
本发明的蛋白、本发明的颗粒或本发明的核酸可以被用在人体或动物体的治疗的疫苗接种的方法中。本发明提供了用于人类或动物个体的疫苗接种的方法,所述方法包括向所述个体施用本发明的蛋白、本发明的颗粒或本发明的核酸。本发明提供了本发明的蛋白、本发明的颗粒或本发明的核酸用于制备用于人类或动物体的疫苗接种的药物的应用。所述蛋白、颗粒或核酸可以被用来针对本文描述的病原体的任一种进行疫苗接种。特别地,所述蛋白、颗粒或核酸可以被用于流感(例如流感病毒a)的疫苗接种。
疫苗接种背后的原理是在宿主中诱导免疫应答,从而在宿主中产生免疫记忆。这意味着,当宿主被暴露至有毒病原体时,它引发有效的(保护性的)免疫反应,即使病原体失活和/或杀死病原体的免疫反应。本发明形成了针对流感病毒的疫苗的基础,并且根据何种其他的免疫原性多肽被包括在蛋白中,能够针对范围广泛的其他疾病和病症对个体同时进行疫苗接种,所述疾病和病症例如hbv、hav、hcv、口蹄病、脊髓灰质炎、疱疹、狂犬病、aids、登革热、黄热病、疟疾、肺结核、百日咳、伤寒、食物中毒、腹泻、脑膜炎和淋病。对在本发明的蛋白中的免疫原性多肽进行选择,以使其适于该疫苗旨在提供的保护所针对的疾病。
本发明的蛋白、颗粒或核酸能够诱导针对流感病毒a的任何或所有亚型的免疫反应。因此,本发明的蛋白、颗粒或核酸能够针对流感病毒a的任何或所有亚型进行免疫接种(参见例如sharp2002cellvol.108,305-312,“originsofhumanvirusdiversity”和shietal2010,plosone,5(12)“acompleteanalysisofhaandnagenesofinfluenzaaviruses”)。例如,ha亚型的任何一个,例如h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15或h16,和/或来自na亚型的任何一个,例如n1、n2、n3、n4、n5、n6、n7、n8或n9。优选是亚型h1n1、h5n1、h3n2、h7n7、h1n2、h2n2、h7n3、h5n2、h1n7、h9n2、h7n2和/或h10n7。更优选的是亚型h3n2、h5n1、h1n1或h7n7。例如,串联构建体可以包括来自流感病毒a的多个亚型的流感病毒a表面多肽m2或其免疫原性片段,并且因此可以被用作诱导免疫反应的通用疫苗,并且因此提供针对流感病毒a的子集或所有亚型的保护。例如,包括通用m2e序列和通用m2e序列的一个、两个或多个变种(如本文所述)的串联构建体可以诱导针对流感病毒a的子集或所有亚型的免疫反应。例如,参见在实施例1和3中的包含串联构建体的三重me2。所述通用m2e序列的一个、两个或多个变种可以是在ha亚型中发现的最常见的变种中的一个或多个(例如一个、两个或三个),例如h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15或h16,和/或来自na亚型的任何一个,例如n1、n2、n3、n4、n5、n6、n7、n8或n9。优选是亚型h1n1、h5n1、h3n2、h7n7、h1n2、h2n2、h7n3、h5n2、h1n7、h9n2、h7n2和/或h10n7。更优选的是亚型h3n2、h5n1、h1n1或h7n7。
并且,作为例子,串联构建体可以包括流感病毒a表面多肽m2或其免疫原性片段以及衍生自流感病毒a的一个或多个亚型的一个或多个进一步的免疫原性多肽,并且因此可以被用作诱导免疫反应的通用疫苗,并且因此提供针对流感病毒a的子集或所有亚型的保护。例如,包括通用m2e序列和通用m2e序列的一个、两个或多个变种(如本文所述)以及ha或其免疫原性片段(例如ha茎)的一个或多个(例如一个、两个或三个)拷贝的串联构建体可以诱导针对流感病毒a的子集或所有亚型的免疫反应。例如,参见在实施例3中的串联构建体。所述通用m2e序列的一个、两个或多个变种可以是在ha亚型中发现的最常见的变种中的一个或多个(例如一个、两个或三个),例如h1、h2、h3、h4、h5、h6、h7、h8、h9、h10、h11、h12、h13、h14、h15或h16,和/或来自na亚型的任何一个,例如n1、n2、n3、n4、n5、n6、n7、n8或n9。优选是亚型h1n1、h5n1、h3n2、h7n7、h1n2、h2n2、h7n3、h5n2、h1n7、h9n2、h7n2和/或h10n7。更优选的是亚型h3n2、h5n1、h1n1或h7n7。所述免疫原性多肽(例如ha或其免疫原性片段,例如ha茎)还可以衍生自任何流感病毒a亚型,例如在上文中列出的任何一种。
并且,作为例子,串联构建体可以包括流感病毒ha或其免疫原性片段(例如ha茎)区域,其也可以衍生自任何流感病毒a亚型,例如在上文中列出的任何一种。
可以制备多个不同的串联构建体,其每个都包括来自流感的相同或不同的亚型的不同的流感病毒抗原,以同时使用于针对流感的人类或动物体的疫苗接种的方法中。例如,包含本发明的流感病毒a表面多肽m2或其免疫原性片段的串联构建体,与包含本发明的ha或其免疫原性片段的串联构建体,可以被联合使用。特别地,提供对h1n1、h1n7和/或h5n1流感感染的保护的包含本发明的流感病毒a表面多肽m2或其免疫原性片段的串联构建体,与提供对h3n2流感感染的保护的包含本发明的ha或其免疫原性片段的串联构建体,可以被联合使用。例如,参见在实施例4中的串联构建体。
因此,本发明还提供了包含一种以上本发明的不同的蛋白、颗粒或核酸的药物组合物和疫苗。例如,本发明提供了药物组合物和疫苗,其包括(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中所述第一蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,所述多肽或片段在一侧或两侧的侧翼具有接头,所述接头将所述多肽或片段连接至hbcag序列,并且任选地hbcag的第二拷贝包括在el环中的另一种免疫原性多肽或小于20个氨基酸的序列;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中所述第二蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂。
本发明还提供了药物组合物和疫苗,其包括(i)第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸,其中所述第一蛋白包括串联的hbcag的第一拷贝和第二拷贝,其中hbcag的一个或两个拷贝包括在el环中的流感病毒a表面多肽m2或其免疫原性片段,并且在流感病毒a表面多肽m2或其免疫原性片段的第17和/或19位的半胱氨酸氨基酸被删除或者被替代的氨基酸取代,并且任选地hbcag的第二拷贝包括在el环中的另一种免疫原性多肽或小于20个氨基酸的序列;以及(ii)第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,其中所述第二蛋白包括串联的乙型肝炎核心抗原(hbcag)的第一拷贝和第二拷贝,其中hbcag的第一拷贝包括在el环中的血凝素(ha)或其免疫原性片段,其中ha的所述片段任选地是ha茎区域,并且hbcag的第二拷贝包括在el环中的小于20个氨基酸的序列;以及药学上可接受的载体或稀释剂。
在所述药物组合物和疫苗中的第一蛋白和第二蛋白可以提供针对上述流感的亚型的任一种的保护,但优选提供针对不同的亚型的保护。例如,第一蛋白可以提供针对h1n1、h1n7和/或h5n1的保护,并且第二蛋白可以提供针对h3n2流感感染的保护(例如实施例4中的串联构建体)。
因此,本发明的第一蛋白和第二蛋白可以被一起使用在在对象中诱导针对流感的免疫反应的方法中。所述方法可以包括向所述对象施用:(i)本发明的第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸;以及(ii)本发明的第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸。这两种实体可以被施用在相同或不同的组合物中,优选相同的组合物。
本发明还提供了用于在针对流感的人类或动物体的疫苗接种的方法中使用的(i)本发明的第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸;以及(ii)本发明的第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸。本发明还提供了(i)本发明的第一蛋白、包含所述第一蛋白的多个拷贝的颗粒、或编码所述第一蛋白的核酸;以及(ii)本发明的第二蛋白、包含所述第二蛋白的多个拷贝的颗粒、或编码所述第二蛋白的核酸,用于制备用于针对流感的人类或动物体的疫苗的应用。术语“个体”和“对象”在本文中可互换使用,是指脊索动物亚门的任何成员,包括但不限于:人和其他灵长类,包括非人灵长类,例如黑猩猩和其他猿类和猴类物种;农场动物,例如牛、绵羊、猪、山羊和马;家养哺乳动物,例如狗和猫;实验动物,包括啮齿动物,例如小鼠、大鼠和豚鼠以及猪;鸟类,包括家养的、野生的和竞技的鸟类,例如鸡、火鸡和其他鸡形鸟类、鸭、鹅等。所述术语不指示特定的年龄。因此,成体和新生个体都旨在被包括在内。本发明所述的方法旨在用于以上脊椎动物物种的任何一种,因为所有这些脊椎动物的免疫系统都以类似方式运作。
在一些情况下,本发明可以被施用至任何合适的对象,特别是给定物种的任何合适对象,优选是合适的人类对象。因此,尽可能多的对象可以例如接受施用,而不强调对象的任何特定的群体。例如,作为整体的或尽可能多的对象的群体可以接受施用。
本发明的蛋白、颗粒或核酸用于施用至对象。它可以与佐剂一起同时施用或按顺序施用。因此,包含蛋白、颗粒或核酸的本发明的组合物还可以包含佐剂。本发明的组合物可以通过注射(例如皮内、皮下、肌内、静脉内、骨内和腹膜内)、经皮颗粒递送、吸入、局部地、口服地或经黏膜(例如鼻腔、舌下、阴道或直肠)的方式而递送。
所述组合物可以被配制为常规的药物制剂。这可以使用标准的药物制剂化学和方法学来实现,这对于本领域技术人员都是可获得的。例如,包含蛋白、颗粒或核酸的组合物(含有或不含有佐剂)可以与一种或多种药学上可接受的赋形剂或载体组合以提供液体制剂。因此,本发明还提供了一种药物组合物,其包含所述蛋白、颗粒或核酸以及药学上可接受的载体或稀释剂。所述组合物任选地包含佐剂。
可以存在辅助性物质,例如润湿剂或乳化剂、ph缓冲物质等。这些载体、稀释剂和辅助性物质通常是这样的药物试剂,它可以被施用而不引起不适当的毒性,并且在抗原性组合物的情况下在其本身不会诱导在接收该组合物的个体中的免疫反应。药学上可接受的载体包括但不限于液体,例如水、盐水、聚乙二醇、透明质酸、甘油和乙醇。药学上可接受的盐也可以被包括在其中,例如无机酸盐,例如盐酸盐、氢溴酸盐、磷酸盐、硫酸盐等;以及有机酸的盐,例如乙酸盐、丙酸盐、丙二酸盐、苯甲酸盐等。虽然不是必需的,但优选地,制剂将含有用作稳定剂的药学上可接受的载体,特别是用于肽、蛋白或其他类似的分子(如果它们被包括在组合物中的话)。也用作肽的稳定剂的合适的载体的例子包括但不限于药品级的葡萄糖、蔗糖、乳糖、海藻糖、甘露醇、山梨糖醇、肌糖、葡聚糖等。其他合适的载体包括但不限于淀粉、纤维素、磷酸钠或磷酸钙、柠檬酸、酒石酸、甘氨酸、高分子量的聚乙二醇(peg)及其组合。药学上可接受的赋形剂、载体和辅助性物质的深入讨论在remington'spharmaceuticalsciences(mackpub.co.,n.j.1991)中可获得,其通过引用合并至本文中。
某些核酸摄取和/或表达的促进剂(“转染促进剂”)也可以被包括在组合物中,例如:促进剂,例如布比卡因、心脏毒素和蔗糖,以及转染促进载体,例如常规地被用来递送核酸分子的脂质体或脂质制剂。阴离子和中性脂质体广泛可用并且熟知用于递送核酸分子(参见例如liposomes:apracticalapproach,(1990)rpcnewed.,irlpress)。阳离子脂质制剂也是熟知的用于递送核酸分子的载体。合适的脂质制剂包括:dotma(n-[1-(2,3-二油烯基氧基)丙基]-n,n,n-三甲基氯化铵),商品名为lipofectin™,以及dotap(1,2-双(油烯基氧基)-3-(三甲基铵基)丙烷),参见例如felgneretal.(1987)proc.natl.acad.sci.usa84:7413-7416;maloneetal.(1989)proc.natl.acad.sci.usa86:6077-6081;美国专利号5,283,185和5,527,928,以及国际公开号wo90/11092、wo91/15501和wo95/26356。这些阳离子脂质可以优选地与中性脂质配合使用,例如dope(二油烯基磷脂酰乙醇胺)。可以被加入至上述脂质或脂质体制剂的进一步的转染促进组合物包括精胺衍生物(参见例如国际公开号wo93/18759)以及膜透化化合物,例如gala、gramicidines和阳离子胆汁盐(参见例如国际公开号wo93/19768))。
替代地,所述蛋白、颗粒或核酸和/或佐剂可以被包封、吸附至颗粒载体,或与颗粒载体连接。合适的颗粒载体包括来源于聚甲基丙烯酸甲酯聚合物的颗粒载体、以及来源于聚乳酸和聚乳酸共乙交酯的plg微颗粒。参见例如jefferyetal.(1993)pharm.res.10:362-368。也可以使用其他颗粒系统和聚合物,例如聚合物,例如聚赖氨酸、聚精氨酸、聚鸟氨酸、精胺、亚精胺以及这些分子的轭合物。例如,可以在存在多核苷酸缩合剂和金属离子螯合剂的情况下,将多核苷酸沉淀在载体上。优选的缩合剂包括阳离子聚合物,特别是聚胺,特别是聚精氨酸或聚赖氨酸。在优选的情况中,所述聚胺是(arg)4或(arg)6。可以参考在wo2004/208560中讨论的可以使用的技术。
一旦配制完成,所述组合物可以使用多种已知的途径和技术在体内递送至对象。例如,液体制剂可以作为注射溶液、悬浮液或乳剂而提供,并使用常规的针头和注射器或使用液体喷射注射系统,经过肠胃外、皮下、皮内、肌内、静脉内、骨内和腹膜内注射而施用。液体制剂也可以被局部地施用至皮肤或粘膜组织(例如鼻腔、舌下、阴道或直肠),或作为适于吸入施用或肺部施用的细分散喷雾而提供。其他施用方式包括口服施用、栓剂和主动或被动的经皮递送技术。
本发明的蛋白、颗粒或核酸以将有效地调节免疫反应的量而被施用至对象。合适的有效量将会落入相对较大的范围,但可以由本领域技术人员通过常规的试验而容易地确定。“医生桌面参考(physiciansdeskreference)”和“古德曼和吉尔曼的治疗药理基础(goodmanandgilman’sthepharmacologicalbasisoftherapeutics)”可以用于确定所需的量。通常,所述蛋白或颗粒以0.1至200mg、优选1至100mg、更优选10至50mg体重的剂量施用。可以使用本领域已知的技术或使用本领域已知的载体,作为裸核酸构建体直接施用本发明的核酸。施用的核酸的量通常是1μg至10mg、优选100mg至1mg。疫苗可以以单剂量方案或多剂量方案(例如2至32个剂量或4至16个剂量)给出。上述施用的途径和剂量仅旨在作为指导,途径和剂量最终由医生决定。
在某些情况下,在初始施用之后,可以进行本发明的组合物的后续施用。特别地,在初始施用之后,对象可被给予“加强剂”。所述加强剂可以是例如选自任何以上所述剂量的剂量。所述加强剂施用可以在初始施用之后例如至少一周、两周、四周、六周、一个月、两个月或六个月进行。
本发明的蛋白、颗粒或核酸及佐剂可以按顺序或同时施用,优选同时施用。两种实体可以在相同或不同的组合物中施用,优选相同的组合物。递送所述佐剂,使得观察到佐剂效应,即,观察到的免疫反应不同于佐剂没有与抗原一起施用时的响应。两种实体可以被施用在相同或不同的位点,优选相同的位点。优选地,两种实体在相同的时间在相同的位点被施用在相同的组合物中,并且优选经注射施用。
可以使用任何合适的佐剂。当前使用的疫苗佐剂包括:
-无机化合物,例如铵盐(如氢氧化铵和磷酸铝)或磷酸钙。铝盐也被称为矾。
-基于油乳剂和表面活性剂的制剂,例如mf59(微流化的洗涤剂稳定化的水包油乳剂)、qs21(纯化的皂素)、as02[sbas2](水包油乳剂+mpl+qs-21)、montanideisa-51和isa-720(稳定化的油包水乳剂)。
-颗粒佐剂,例如病毒颗粒(结合了例如流感血凝素的单层脂质体载体)、as04([sbas4]具有mpl的al盐)、iscoms(皂素和脂质的结构化复合物)、以及聚乳酸乙交酯共聚物(plg)。
-微生物衍生物(天然的和合成的),例如单磷酰脂a(mpl)、detox(mpl+m.phlei细胞壁骨架)、agp[rc-529](合成的酰化单糖)、dc_chol(能够自身组织成脂质体的类脂免疫刺激剂)、om-174(脂质a衍生物)、cpg模体(含有免疫刺激cpg模体的合成寡核苷酸)、以及经修饰的lt和ct(经遗传修饰的细菌毒素以提供无毒佐剂效应)。
-内源性人免疫调节剂,例如hgm-csf或hil-12(能够以蛋白或编码的质粒的形式施用的细胞因子)、以及immudaptin(c3d串联阵列)。
-惰性载体,例如金颗粒。
优选的佐剂是矾。最优选地,所述佐剂是氢氧化铝和氢氧化镁的混合物,例如注射用矾(piercelaboratories),它不适于用在人类中。
本发明通过以下实施例来阐释:
实施例1
材料与方法:
vlp序列:
在bl21e.coli中制备包含在mir中的来自流感病毒保守蛋白的插入体的串联核心。对应于所述插入体的序列属于流感病毒基质蛋白2胞外域(m2e)的区域。它跨越编码已知的m2蛋白的n末端外部序列的24个氨基酸(图5);mslltevetpirnewgcrcngssd(seqidno:6)。该野生型序列被修改来使用丝氨酸残基取代在第17和19位(以上下划线的)的半胱氨酸残基(其影响vlp形成)。最终的插入体包含该序列的三个变种。第一个版本是通用m2e共识序列,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外(seqidno:9)。第二个(seqidno:8)和第三个(seqidno:10)是通用序列的突变版本,其对应于在h3n2和h5n1流感病毒中发现的最常见的变种,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外。m2e序列在侧翼有柔性的分隔物,其由15个氨基酸形成;ggggsggggsggggs(seqidno:5)。还制备了包含m2e序列一致性、拷贝数目以及侧翼序列的变化的其他构建体;这些构建体被描述于以下的表2中。
表2.通过在插入体序列上的变化而生成的替代的vlp的表。
*侧翼序列是m2e序列的上游和下游
$该插入体在m2e序列之间具有所示的侧翼序列。
具有3xm2e插入体的串联核心的氨基酸序列如下所示(单个字母aa编码)。来自串联核心的氨基酸为加粗,而来自流感m2的氨基酸为下划线,柔性的连接区域为斜体:
mdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlegs
ggggsggggsggggs
mslltevetptrngwgsksngssd
mslltevetpirnewgsrsngssd
mslltevetptrnewesrssgssd
ggggsggggsggggs
grdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvggssggsggsggsggsggsggstmdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlefagasdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvrrrdrgrsprrrtpsprrrrsqsprrrrsqsresqclehhhhhh-(seqidno:11)
理论上的pi/mw:6.29/51438.34
动物:
从harlan(wyton,uk)购得6-8周龄的balb/c雌性小鼠并关在ivc2类遏制设施中。所有的动物护理和程序都是根据英国内政部对实验目的的动物使用的规定而进行的。
免疫:
对于初级免疫,个体小鼠被给予腹腔内(i.p.)注射,其含有15μgvlp材料、20μgsas(sigma佐剂系统-sigmaaldrich)、20μgpierceimject明矾(thermoscientific)、以及无菌盐水溶液,至总体积为100μl。对于加强免疫,在初级免疫之后7和14日施用,个体小鼠接受皮下(s.c.)注射,其含有5ugvlp材料、20μgmlpa(sigma)、20μg胞壁酰二肽(sigma)、20μgpierceimject明矾(thermoscientific),并且用无菌盐水溶液形成100μl的最终体积。在最终的加强免疫之后一日,通过面部静脉提取对小鼠取血,以确认血清转化。对照组仅通过不包含vlp材料的佐剂进行免疫。
血清转化:
通过elisa进行抗-m2e抗体的检测。使用在1mnacl缓冲液中的6.25μg/mlm2e肽序列mslltevetpirnewgcrcngssd-oh(seqidno:14)(activotec)涂覆96孔nuncmaxisorp板。使用pbs-tween200.05%洗涤3次被涂覆的板,并在37°c使用10%牛奶溶液封闭1小时。加入在2.5%牛奶中的稀释的血清,并在37°c孵育1小时,然后像之前一样洗涤3次。20分钟加入tmb基质(sigma),使用1mh2so4停止反应。使用tecansunrise平板读取器读出在450nm的具有630nm修正的吸光度,并使用magellan™软件进行分析。所有孔都做两份,至少3次稀释重复。
挑战:
在初级免疫之后4周,使用5xmld50剂量的a/pr8h1n1流感病毒感染小鼠。通过在盐水中的氯胺酮/甲苯噻嗪2:1混合物的腹膜内(i.p.)施用对小鼠进行麻醉,然后鼻内(i.n.)施用病毒。在感染的时候(第0天)对小鼠进行称重,此后每半日称重直至进行完全恢复。还使用在表3中发现的疾病程度对小鼠进行评分。
结果:
免疫的小鼠从流感病毒血清转化为m2e肽。
在初级免疫之后3周,对来自小鼠的抗血清进行收集、汇集并通过elisa测试针对来自流感病毒的合成的m2e肽的反应性。使用含有3xm2e插入体的串联核心vlp进行免疫的小鼠产生抗体,所述抗体能够结合来自流感病毒的m2e肽,而仅使用佐剂进行免疫的小鼠则不会(图1)。
免疫的小鼠被保护免受致命的a/pr8h1n1流感病毒感染。
在初级免疫之后4周,使用5xmld50的pr8流感病毒对小鼠进行挑战。在仅有佐剂的群组中的小鼠快速地体重减轻并且临床分数高(图2和图3),在感染后第8日达到100%死亡(图4)。反过来,使用具有流感3xm2e插入体的串联核心进行免疫的小鼠达到8%的峰值体重损失,这与更温和的临床评分相关,并且在第7天开始恢复,在感染后第12日实现完全恢复(图2和图3)。在免疫组中的存活率是100%(图4)。这些结果合在一起显示,在pr8流感病毒的致命挑战之后,接受含有流感3xm2e插入体的串联核心的小鼠表现出较少的体重损失、较低的发病率并且没有死亡。串联核心所赋予的保护并不无菌,这是因为抗-m2e抗体滴度不足够高来进行完全保护,或者因为抗-m2e通过非中和机制进行保护。替代地,保护可以通过非抗体依赖机制介导,但是它与抗-m2e保护水平相关性良好。不管具体的机制如何,保护是通过具有串联核心vlp的疫苗接种赋予的,因为无免疫的小鼠表现出更为严重的症状和高的死亡率。
表3.对流感症状的严重程度。个体的临床分数是使用以下程度来确定的。
实施例2
材料与方法:
vlp序列:
在bl21e.coli中制备包含在mir中的来自流感病毒保守蛋白的插入体的串联核心。对应于所述插入体的序列属于流感病毒血凝素ha2蛋白结构域的茎区域。它跨越编码已知的结构域的核酸;ha单体的环b、螺旋c、螺旋cd和螺旋d(图10)。该序列跨越分离自流感a病毒h1n1/lux/09的ha蛋白的氨基酸(aa)403-474。存在包含ha茎插入体序列的其他构建体,这些被描述于以下表4中。
表4.通过在插入体序列上的变化而生成的替代的vlp的表。
具有ha茎流感插入体的串联核心的氨基酸序列(单个字母aa编码)如下所示。来自串联核心的氨基酸为加粗,而来自流感ha的氨基酸为下划线:
mdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlegs
mntqftavgkefnhlekrienlnkkvddgfldiwtynaellvllenertldyhdsnvknlyekvrsqlknna
sgrdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvggssggsggsggsggsggsggstmdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlefagasdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvrrrdrgrsprrrtpsprrrrsqsprrrrsqsresqclehhhhhh-(seqidno:12)
理论上的pi/mw:6.98/50293.85
动物:
从harlan(wyton,uk)购得6-8周龄的balb/c雌性小鼠并关在ivc2类遏制设施中。所有的动物护理和程序都是根据英国内政部对实验目的的动物使用的规定而进行的。
免疫:
对于初级免疫,个体小鼠被给予腹腔内(i.p.)注射,其含有15μgvlp材料、20μgsas(sigma佐剂系统-sigmaaldrich)、20μgpierceimject明矾(thermoscientific)、以及无菌盐水溶液,至总体积为100μl。对于加强免疫,在初级免疫之后7和14日施用,个体小鼠接受皮下(s.c.)注射,其含有5ugvlp材料、20μgmlpa(sigma)、20μg胞壁酰二肽(sigma)、20μgpierceimject明矾(thermoscientific),并且用无菌盐水溶液形成100μl的最终体积。在最终的加强免疫之后一日,通过面部静脉提取对小鼠取血,以确认血清转化。对照组仅通过不包含vlp材料的佐剂进行免疫。
血清转化:
通过elisa进行抗-ha抗体的检测。使用在碳酸二碳酸酯缓冲液中的来自a/pr8流感病毒(lifetechnologies)的1μg/mlrha涂覆96孔nuncmaxisorp板。使用pbs-tween200.05%洗涤3次被涂覆的板,并在37°c使用10%牛奶溶液封闭1小时。加入在2.5%牛奶中的稀释的血清,并在37°c孵育1小时,然后像之前一样洗涤3次。20分钟加入tmb基质(sigma),使用1mh2so4停止反应。使用tecansunrise平板读取器读出在450nm的具有630nm修正的吸光度,并使用magellan™软件进行分析。所有孔都做两份,至少3次稀释重复。
挑战:
在初级免疫之后4周,使用5xmld50剂量的a/pr8h1n1流感病毒感染小鼠。通过在盐水中的氯胺酮/甲苯噻嗪2:1混合物的腹膜内(i.p.)施用对小鼠进行麻醉,然后鼻内(i.n.)施用病毒。在感染的时候(第0天)对小鼠进行称重,此后每半日称重直至进行完全恢复。还使用在表3中发现的疾病程度对小鼠进行评分。
结果:
免疫的小鼠从a/pr8h1n1流感病毒血清转化为rha蛋白。
在初级免疫之后3周,对来自小鼠的抗血清进行收集、汇集并通过elisa测试针对来自pr8流感病毒的重组血凝素蛋白的反应性。使用含有ha茎插入体的串联核心vlp进行免疫的小鼠产生抗体,所述抗体能够结合来自流感病毒的rha蛋白,而仅使用佐剂进行免疫的小鼠则不会(图6)。
免疫的小鼠被保护免受致命的a/pr8h1n1流感病毒感染。
在初级免疫之后4周,使用5xmld50的pr8流感病毒对小鼠进行挑战。在仅有佐剂的群组中的小鼠快速地体重减轻并且临床分数高(图7和图8),在感染后第8日达到100%死亡(图9)。反过来,使用具有流感茎插入体的串联核心进行免疫的小鼠达到15%的峰值体重损失,这与更温和的临床评分相关,并且在第8天开始恢复,在感染后第16日实现完全恢复(图7和图8)。在免疫组中的存活率是100%(图9)。这些结果合在一起显示,在pr8流感病毒的致命挑战之后,接受含有流感茎插入体的串联核心的小鼠表现出较少的体重损失、较低的发病率并且没有死亡。串联核心所赋予的保护并不无菌,这是因为抗-ha抗体滴度不足够高来进行完全保护,或者因为抗-ha通过非中和机制进行保护。替代地,保护可以通过非抗体依赖机制介导,但是它与抗-ha保护水平相关性良好。不管具体的机制如何,保护是通过具有串联核心vlp的疫苗接种赋予的,因为无免疫的小鼠表现出更为严重的症状和高的死亡率。
实施例3
材料与方法:
vlp序列:
在bl21e.coli中制备包含在两个主要插入区域(mir)之内的来自流感病毒保守蛋白的插入体的串联核心(图15)。对应于第一插入体的序列属于流感病毒血凝素ha2蛋白结构域的茎区域。它跨越编码已知的结构域的核酸;ha2单体的环b、螺旋c、螺旋cd和螺旋d(图10)。该序列跨越分离自流感a病毒h1n1/lux/09的ha蛋白的氨基酸(aa)403-474。对应于第二插入体的序列属于流感病毒基质蛋白2胞外域(m2e)的区域。它跨越编码已知的m2蛋白的n末端外部序列的24个氨基酸;mslltevetpirnewgcrcngssd(seqidno:6)(图5)。该野生型序列被修改来使用丝氨酸残基取代在第17和19位(以上下划线的)的半胱氨酸残基(其影响vlp形成)。最终的插入体包含该序列的三个变种。第一个版本是通用m2e共识序列,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外(seqidno:9)。第二个(seqidno:8)和第三个(seqidno:10)是通用序列的突变版本,其对应于在h7n7和h5n1流感病毒中发现的最常见的变种,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外。具有ha茎流感插入体和m2ex3的串联核心的氨基酸序列如下所示(单个字母aa编码)。来自串联核心的氨基酸为加粗,而来自流感的氨基酸为下划线:
mdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlegs
mntqftavgkefnhlekrienlnkkvddgfldiwtynaellvllenertldyhdsnvknlyekvrsqlknna
sgrdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvggssggsggsggsggsggsggstmdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlef
ggggsggggsggggs
mslltevetptrngwgsksngssd
mslltevetpirnewgsrsngssd
mslltevetptrnewesrssgssd
ggggsggggsggggs
asdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvrrrdrgrsprrrtpsprrrrsqsprrrrsqsresqclehhhhhh-(seqidno:13)
理论上的pi/mw:6.17/59834.78
动物:
从harlan(wyton,uk)购得6-8周龄的balb/c雌性小鼠并关在ivc2类遏制设施中。所有的动物护理和程序都是根据英国内政部对实验目的的动物使用的规定而进行的。
免疫:
对于初级免疫,个体小鼠被给予腹腔内(i.p.)注射,其含有15μgvlp材料、20μgsas(sigma佐剂系统-sigmaaldrich)、20μgpierceimject明矾(thermoscientific)、以及无菌盐水溶液,至总体积为100μl。对于加强免疫,在初级免疫之后7和14日施用,个体小鼠接受皮下(s.c.)注射,其含有5ugvlp材料、20μgmlpa(sigma)、20μg胞壁酰二肽(sigma)、20μgpierceimject明矾(thermoscientific),并且用无菌盐水溶液形成100μl的最终体积。在最终的加强免疫之后一日,通过面部静脉提取对小鼠取血,以确认血清转化。对照组仅通过不包含vlp材料的佐剂进行免疫。
血清转化:
通过elisa进行抗-ha或抗-m2e抗体的检测。使用在碳酸二碳酸酯缓冲液中的来自a/pr8流感病毒(lifetechnologies)的1μg/mlrha、或者在1mnacl缓冲液中的6.25μg/mlm2e肽序列mslltevetpirnewgcrcngssd-oh(seqidno:14)(activotec),涂覆96孔nuncmaxisorp板。使用pbs-tween200.05%洗涤3次被涂覆的板,并在37°c使用10%牛奶溶液封闭1小时。加入在2.5%牛奶中的稀释的血清,并在37°c孵育1小时,然后像之前一样洗涤3次。以1/2500稀释(sigma)加入山羊抗小鼠igg-过氧化物酶二次抗体,并在37°c孵育1小时,然后像之前一样洗涤3次。20分钟加入tmb基质(sigma),使用1mh2so4停止反应。使用tecansunrise平板读取器读出在450nm的具有630nm修正的吸光度,并使用magellan™软件进行分析。所有孔都做两份,至少3次稀释重复。
挑战:
在初级免疫之后4周,使用5xmld50剂量的a/pr8h1n1流感病毒感染小鼠。通过在盐水中的氯胺酮/甲苯噻嗪2:1混合物的腹膜内(i.p.)施用对小鼠进行麻醉,然后鼻内(i.n.)施用病毒。在感染的时候(第0天)对小鼠进行称重,此后每半日称重直至进行完全恢复。还使用在表3中发现的疾病程度对小鼠进行评分。
结果:
免疫的小鼠从a/pr8h1n1流感病毒和m2e肽血清转化为rha蛋白。
在初级免疫之后3周,对来自小鼠的抗血清进行收集、汇集并通过elisa测试针对来自pr8流感病毒的重组血凝素蛋白和m2e肽的反应性。使用含有ha茎和3xm2e插入体的串联核心vlp进行免疫的小鼠产生抗体,所述抗体能够结合来自流感病毒的rha蛋白和m2e肽,而仅使用佐剂进行免疫的小鼠则不会(图11)。
免疫的小鼠被保护免受致命的a/pr8h1n1流感病毒感染。
在初级免疫之后4周,使用5xmld50的pr8流感病毒对小鼠进行挑战。在仅有佐剂的群组中的小鼠快速地体重减轻并且临床分数高(图12和图13),在感染后第10日达到100%死亡(图14)。反过来,使用串联核心vlp进行免疫的小鼠达到5%的峰值体重损失,这与更温和的临床评分相关,并且在第6天开始恢复,在感染后第10日实现完全恢复(图12和图13)。在免疫组中的存活率是100%(图14)。这些结果合在一起显示,在pr8流感病毒的致命挑战之后,接受含有流感衍生的插入体的串联核心的小鼠表现出较少的体重损失、较低的发病率并且没有死亡。串联核心所赋予的保护并不无菌,这是因为抗流感抗体滴度不足够高来进行完全保护,或者因为vlp疫苗通过非中和机制进行保护。不管具体的机制如何,保护是通过具有串联核心vlp的疫苗接种赋予的,因为无免疫的小鼠表现出更为严重的症状和高的死亡率。
实施例4
材料与方法:
vlp序列:
在pichiapastoris酵母中制备包含在两个主要插入区域(mir)之内的来自流感病毒保守蛋白的插入体的串联核心vlp(图16)。第一vlp(vlp1)包含两个流感插入体,在每个mir中有一个。第一插入体是对应于流感h1n1病毒血凝素ha2蛋白结构域的茎区域的序列。它跨越编码已知的结构域的核酸;ha2单体的环b、螺旋c、螺旋cd和螺旋d(图10)。该序列跨越分离自流感a病毒h1n1/lux/09的ha蛋白的氨基酸(aa)403-474。vlp1的该插入体此后被称为ha2.3(图17)。
对应于在vlp1上的第二插入体的序列属于流感病毒基质蛋白2胞外域(m2e)的区域。它跨越编码已知的m2蛋白的n末端外部序列的24个氨基酸(图5);mslltevetpirnewgcrcngssd(seqidno:6)。该野生型序列被修改来使用丝氨酸残基取代在第17和19位(以上下划线的)的半胱氨酸残基(其影响vlp形成)。最终的插入体包含该序列的三个变种。第一个版本是通用m2e共识序列,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外(seqidno:9)。第二个(seqidno:8)和第三个(seqidno:10)是通用序列的突变版本,其对应于在h7n7和h5n1流感病毒中发现的最常见的变种,除了在第17和19位的半胱氨酸残基已经被丝氨酸取代之外。vlp1的该三重插入体此后被称为m2ex3。所述m2e序列侧翼有柔性的分隔物:ggggsggggsggggs(seqidno:5)和ggggsggggsgggg(seqidno:16)。
vlp1的完整dna序列是seqidno:17,并且vlp1的氨基酸序列如下所示(单个字母aa编码)。来自串联核心的氨基酸为加粗,而来自流感的氨基酸为下划线,柔性的连接区域为斜体:
mdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlegs
mntqftavgkefnhlekrienlnkkvddgfldiwtynaellvllenertldyhdsnvknlyekvrsqlknna
sgrdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvggssggsggsggsggsggsggstmdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlef
ggggsggggsggggs
mslltevetptrngwgsksngssd
mslltevetpirnewgsrsngssd
mslltevetptrnewesrssgssd
ggggsggggsgggg
asdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvrrrdrgrsprrrtpsprrrrsqsprrrrsqsresqc
(seqidno:18)
理论上的pi/mw:6.01/58769.66
第二vlp(vlp2)在其第一个mir中包含对应于流感h3n2病毒血凝素ha2蛋白结构域的序列。它跨越编码已知的结构域螺旋c、螺旋cd和螺旋d的核酸,这些结构域有时被统称为“长α-螺旋”或“lah”(图10)。该序列包括分离自流感a病毒h3n2/hk/68的ha蛋白的氨基酸(aa)421-475。该插入体此后将被称为lah3(图17)。
vlp2的第二插入体是侧翼有柔性的接头区域的单个赖氨酸(k)残基,所述接头区域由甘氨酸和丝氨酸残基形成。所施用的插入体的序列是gsgsgggkgggsgs(seqidno:21)。这有效地是空插入体,其使得第一插入体(lah3)正确地配置并且赋予整个vlp2更大的溶解度。
vlp2的完整dna序列是seqidno:19,并且vlp2的氨基酸序列如下所示(单个字母aa编码)。来自串联核心的氨基酸为加粗,而来自流感的氨基酸为下划线,接头区域为斜体:
mdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlegs
riqdlekyvedtkidlwsynaellvalenqhtidltdsemnklfektrrqlrena
sgrdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvggssggsggsggsggsggsggstmdidpykefgatvellsflpsdffpsvrdlldtasalyrealespehcsphhtalrqailcwgelmtlatwvgnnlef
gsgsgggkgggsgs
asdpasrdlvvnyvntnmglkirqllwfhiscltfgretvleylvsfgvwirtppayrppnapilstlpettvvrrrdrgrsprrrtpsprrrrsqsprrrrsqsresqc
(seqidno:20)
理论上的pi/mw:6.77/48186.40
动物:
从harlan(wyton,uk)购得6-8周龄的balb/c雌性小鼠并关在ivc2类遏制设施中。所有的动物护理和程序都是根据英国内政部对实验目的的动物使用的规定而进行的。
免疫:
对于初级免疫,个体小鼠被给予i.p.注射,其含有30μgvlp材料、20μgsas(sigma佐剂系统-sigmaaldrich)、20μgpierceimject明矾(thermoscientific)、以及无菌盐水溶液,至总体积为100μl。对于加强免疫,在初级免疫之后7和14日施用,个体小鼠接受s.c.注射,其含有15ugvlp材料、20μgmlpa(sigma)、20μg胞壁酰二肽(sigma)、20μgpierceimject明矾(thermoscientific),并且用无菌盐水溶液形成100μl的最终体积。在最终的加强免疫之后一日,对小鼠取血,以确认血清转化。对照组仅通过不包含vlp材料的佐剂进行免疫。
血清转化:
通过elisa进行抗-ha或抗-m2e抗体的检测。使用在碳酸二碳酸酯缓冲液中的来自a/pr8或x31流感病毒的1μg/mlrha(lifetechnologies)、或者在1mnacl缓冲液中的6.25μg/mlm2e肽序列mslltevetpirnewgcrcngssd-oh(seqidno:14)(activotec),涂覆96孔nuncmaxisorp板。使用pbs-tween200.05%洗涤3次被涂覆的板,并在37°c使用10%牛奶溶液封闭1小时。加入在2.5%牛奶中的稀释的血清,并在37°c孵育1小时,然后像之前一样洗涤3次。以1/2500稀释(sigma)加入山羊抗小鼠igg-过氧化物酶二次抗体,并在37°c孵育1小时,然后像之前一样洗涤3次。20分钟加入tmb基质(sigma),使用1mh2so4停止反应。使用tecansunrise平板读取器读出在450nm的具有630nm修正的吸光度,并使用magellan™软件进行分析。所有孔都做两份,至少3次稀释重复。
挑战:
在初级免疫之后4周,使用5xmld50剂量的a/pr8h1n1或3xmld50x31h3n2流感病毒感染小鼠。施用经过氧气扩散室递送的异氟醚对小鼠进行麻醉,然后鼻内(i.n.)施用病毒。在感染的时候(第0天)对小鼠进行称重,此后每半日称重直至进行完全恢复。还使用在表3中发现的疾病程度对小鼠进行评分。
结果:
免疫的小鼠从h1n1(a/pr8)、h3n2(x31)流感病毒和m2e肽血清转化为rha蛋白。
在初级免疫之后3周,对来自5只小鼠的抗血清进行收集、汇集并通过elisa测试针对来自pr8(h1)或x31(h3)流感病毒的重组血凝素蛋白和m2e肽的反应性。使用含有ha茎和m2e插入体的串联核心vlp1和vlp2进行免疫的小鼠产生抗体,所述抗体能够结合来自流感病毒的rha蛋白和m2e肽。仅使用佐剂(阴性-)进行免疫的小鼠则不会检测到对任何流感抗原的阳性(图18)。
使用串联核心vlp进行免疫的小鼠被保护免受h1n1和h3n2流感病毒感染。
在初级免疫之后4周,使用5xmld50的pr8h1n1(a)或3xmld50x31h3n2(b)流感病毒对小鼠进行挑战。在仅有佐剂的群组中的小鼠快速地体重减轻(图19a和19b)并且临床分数高(图20a和图20b),在感染后第8日达到高度的死亡率(图21a和图21b)。反过来,使用串联核心vlp进行免疫的小鼠达到10%的峰值体重损失,这与更温和的临床评分相关,并且在第5-7天开始恢复,在感染后第15日实现完全恢复(图19和图20)。在免疫组中的存活率是100%(图21)。这些结果合在一起显示,在pr8h1n1或x31h3n2流感病毒的致命挑战之后,接受含有流感衍生的插入体的串联核心vlp1和vlp2的小鼠表现出较少的体重损失、较低的发病率并且没有死亡。串联核心所赋予的保护容许感染,这是因为抗流感抗体滴度不足够高来进行无菌免疫,或者因为vlp疫苗通过非中和机制进行保护。不管具体的机制如何,明确的益处是通过具有串联核心vlp的疫苗接种赋予的,因为阴性对照组中的小鼠表现出更为严重的症状和高的死亡率。
序列表
<110>iqur有限公司
<120>基于乙型肝炎核心抗原的疫苗
<130>n403290wo
<150>gb1421692.3
<151>2014-12-05
<160>24
<170>patentinversion3.5
<210>1
<211>639
<212>dna
<213>乙型肝炎病毒
<220>
<223>hbcagayw亚型核苷酸和氨基酸序列
<220>
<221>cds
<222>(1)..(639)
<400>1
atgcaactttttcacctctgcctaatcatctcttgttcatgtcctact48
metglnleuphehisleucysleuileilesercyssercysprothr
151015
gttcaagcctccaagctgtgccttgggtggctttggggcatggacatc96
valglnalaserlysleucysleuglytrpleutrpglymetaspile
202530
gacccttataaagaatttggagctactgtggagttactctcgtttttg144
aspprotyrlysglupheglyalathrvalgluleuleuserpheleu
354045
ccttctgacttctttccttcagtacgagatcttctagataccgcctca192
proseraspphepheproservalargaspleuleuaspthralaser
505560
gctctgtatcgggaagccttagagtctcctgagcattgttcacctcac240
alaleutyrargglualaleugluserprogluhiscysserprohis
65707580
catactgcactcaggcaagcaattctttgctggggggaactaatgact288
histhralaleuargglnalaileleucystrpglygluleumetthr
859095
ctagctacctgggtgggtgttaatttggaagatccagcgtctagagac336
leualathrtrpvalglyvalasnleugluaspproalaserargasp
100105110
ctagtagtcagttatgtcaacactaatatgggcctaaagttcaggcaa384
leuvalvalsertyrvalasnthrasnmetglyleulysphearggln
115120125
ctcttgtggtttcacatttcttgtctcacttttggaagagaaacagtt432
leuleutrpphehisilesercysleuthrpheglyarggluthrval
130135140
atagagtatttggtgtctttcggagtgtggattcgcactcctccagct480
ileglutyrleuvalserpheglyvaltrpileargthrproproala
145150155160
tatagaccaccaaatgcccctatcctatcaacacttccggagactact528
tyrargproproasnalaproileleuserthrleuprogluthrthr
165170175
gttgttagacgacgaggcaggtcccctagaagaagaactccctcgcct576
valvalargargargglyargserproargargargthrproserpro
180185190
cgcagacgaaggtctcaatcgccgcgtcgcagaagatctcaatctcgg624
argargargargserglnserproargargargargserglnserarg
195200205
gaatctcaatgttag639
gluserglncys
210
<210>2
<211>212
<212>prt
<213>乙型肝炎病毒
<220>
<223>hbcagayw亚型蛋白序列
<400>2
metglnleuphehisleucysleuileilesercyssercysprothr
151015
valglnalaserlysleucysleuglytrpleutrpglymetaspile
202530
aspprotyrlysglupheglyalathrvalgluleuleuserpheleu
354045
proseraspphepheproservalargaspleuleuaspthralaser
505560
alaleutyrargglualaleugluserprogluhiscysserprohis
65707580
histhralaleuargglnalaileleucystrpglygluleumetthr
859095
leualathrtrpvalglyvalasnleugluaspproalaserargasp
100105110
leuvalvalsertyrvalasnthrasnmetglyleulysphearggln
115120125
leuleutrpphehisilesercysleuthrpheglyarggluthrval
130135140
ileglutyrleuvalserpheglyvaltrpileargthrproproala
145150155160
tyrargproproasnalaproileleuserthrleuprogluthrthr
165170175
valvalargargargglyargserproargargargthrproserpro
180185190
argargargargserglnserproargargargargserglnserarg
195200205
gluserglncys
210
<210>3
<211>97
<212>prt
<213>流感a病毒
<220>
<223>a/34/pr8菌株m2蛋白
<400>3
metserleuleuthrgluvalgluthrproileargasnglutrpgly
151015
cysargcysasnglyserseraspproleuthrilealaalaasnile
202530
ileglyileleuhisleuthrleutrpileleuaspargleuphephe
354045
lyscysiletyrargargphelystyrglyleulysglyglyproser
505560
thrgluglyvalprolyssermetarggluglutyrarglysglugln
65707580
glnseralavalaspalaaspaspglyhisphevalserilegluleu
859095
glu
<210>4
<211>5
<212>prt
<213>人工序列
<220>
<223>可能的接头序列
<400>4
glyglyglyglyser
15
<210>5
<211>15
<212>prt
<213>人工序列
<220>
<223>可能的接头序列
<400>5
glyglyglyglyserglyglyglyglyserglyglyglyglyser
151015
<210>6
<211>24
<212>prt
<213>人工序列
<220>
<223>野生型m2e序列
<400>6
metserleuleuthrgluvalgluthrproileargasnglutrpgly
151015
cysargcysasnglyserserasp
20
<210>7
<211>24
<212>prt
<213>人工序列
<220>
<223>m2e序列,第17位的cys被取代为ser
<400>7
metserleuleuthrgluvalgluthrproileargasnglutrpgly
151015
serargcysasnglyserserasp
20
<210>8
<211>24
<212>prt
<213>人工序列
<220>
<223>m2e序列,第17和19位的cys被取代为ser
<400>8
metserleuleuthrgluvalgluthrproileargasnglutrpgly
151015
serargserasnglyserserasp
20
<210>9
<211>24
<212>prt
<213>人工序列
<220>
<223>通用m2e共识序列(cys被取代为ser)
<400>9
metserleuleuthrgluvalgluthrprothrargasnglytrpgly
151015
serlysserasnglyserserasp
20
<210>10
<211>24
<212>prt
<213>人工序列
<220>
<223>变异的m2e序列
<400>10
metserleuleuthrgluvalgluthrprothrargasnglutrpglu
151015
serargserserglyserserasp
20
<210>11
<211>476
<212>prt
<213>人工序列
<220>
<223>具有三个m2e序列的串联核心
<400>11
metaspileaspprotyrlysglupheglyalathrvalgluleuleu
151015
serpheleuproseraspphepheproservalargaspleuleuasp
202530
thralaseralaleutyrargglualaleugluserprogluhiscys
354045
serprohishisthralaleuargglnalaileleucystrpglyglu
505560
leumetthrleualathrtrpvalglyasnasnleugluglysergly
65707580
glyglyglyserglyglyglyglyserglyglyglyglysermetser
859095
leuleuthrgluvalgluthrprothrargasnglytrpglyserlys
100105110
serasnglyserseraspmetserleuleuthrgluvalgluthrpro
115120125
ileargasnglutrpglyserargserasnglyserseraspmetser
130135140
leuleuthrgluvalgluthrprothrargasnglutrpgluserarg
145150155160
serserglyserseraspglyglyglyglyserglyglyglyglyser
165170175
glyglyglyglyserglyargaspproalaserargaspleuvalval
180185190
asntyrvalasnthrasnmetglyleulysileargglnleuleutrp
195200205
phehisilesercysleuthrpheglyarggluthrvalleuglutyr
210215220
leuvalserpheglyvaltrpileargthrproproalatyrargpro
225230235240
proasnalaproileleuserthrleuprogluthrthrvalvalgly
245250255
glyserserglyglyserglyglyserglyglyserglyglysergly
260265270
glyserglyglyserthrmetaspileaspprotyrlysgluphegly
275280285
alathrvalgluleuleuserpheleuproseraspphepheproser
290295300
valargaspleuleuaspthralaseralaleutyrargglualaleu
305310315320
gluserprogluhiscysserprohishisthralaleuargglnala
325330335
ileleucystrpglygluleumetthrleualathrtrpvalglyasn
340345350
asnleugluphealaglyalaseraspproalaserargaspleuval
355360365
valasntyrvalasnthrasnmetglyleulysileargglnleuleu
370375380
trpphehisilesercysleuthrpheglyarggluthrvalleuglu
385390395400
tyrleuvalserpheglyvaltrpileargthrproproalatyrarg
405410415
proproasnalaproileleuserthrleuprogluthrthrvalval
420425430
argargargaspargglyargserproargargargthrproserpro
435440445
argargargargserglnserproargargargargserglnserarg
450455460
gluserglncysleugluhishishishishishis
465470475
<210>12
<211>447
<212>prt
<213>人工序列
<220>
<223>在hbcag的一个拷贝中具有ha茎的串联核心
<400>12
metaspileaspprotyrlysglupheglyalathrvalgluleuleu
151015
serpheleuproseraspphepheproservalargaspleuleuasp
202530
thralaseralaleutyrargglualaleugluserprogluhiscys
354045
serprohishisthralaleuargglnalaileleucystrpglyglu
505560
leumetthrleualathrtrpvalglyasnasnleugluglysermet
65707580
asnthrglnphethralavalglylysglupheasnhisleuglulys
859095
argilegluasnleuasnlyslysvalaspaspglypheleuaspile
100105110
trpthrtyrasnalagluleuleuvalleuleugluasngluargthr
115120125
leuasptyrhisaspserasnvallysasnleutyrglulysvalarg
130135140
serglnleulysasnasnalaserglyargaspproalaserargasp
145150155160
leuvalvalasntyrvalasnthrasnmetglyleulysilearggln
165170175
leuleutrpphehisilesercysleuthrpheglyarggluthrval
180185190
leuglutyrleuvalserpheglyvaltrpileargthrproproala
195200205
tyrargproproasnalaproileleuserthrleuprogluthrthr
210215220
valvalglyglyserserglyglyserglyglyserglyglysergly
225230235240
glyserglyglyserglyglyserthrmetaspileaspprotyrlys
245250255
glupheglyalathrvalgluleuleuserpheleuproseraspphe
260265270
pheproservalargaspleuleuaspthralaseralaleutyrarg
275280285
glualaleugluserprogluhiscysserprohishisthralaleu
290295300
argglnalaileleucystrpglygluleumetthrleualathrtrp
305310315320
valglyasnasnleugluphealaglyalaseraspproalaserarg
325330335
aspleuvalvalasntyrvalasnthrasnmetglyleulysilearg
340345350
glnleuleutrpphehisilesercysleuthrpheglyarggluthr
355360365
valleuglutyrleuvalserpheglyvaltrpileargthrpropro
370375380
alatyrargproproasnalaproileleuserthrleuprogluthr
385390395400
thrvalvalargargargaspargglyargserproargargargthr
405410415
proserproargargargargserglnserproargargargargser
420425430
glnserarggluserglncysleugluhishishishishishis
435440445
<210>13
<211>547
<212>prt
<213>人工序列
<220>
<223>具有ha茎和三个m2e序列的串联核心
<400>13
metaspileaspprotyrlysglupheglyalathrvalgluleuleu
151015
serpheleuproseraspphepheproservalargaspleuleuasp
202530
thralaseralaleutyrargglualaleugluserprogluhiscys
354045
serprohishisthralaleuargglnalaileleucystrpglyglu
505560
leumetthrleualathrtrpvalglyasnasnleugluglysermet
65707580
asnthrglnphethralavalglylysglupheasnhisleuglulys
859095
argilegluasnleuasnlyslysvalaspaspglypheleuaspile
100105110
trpthrtyrasnalagluleuleuvalleuleugluasngluargthr
115120125
leuasptyrhisaspserasnvallysasnleutyrglulysvalarg
130135140
serglnleulysasnasnalaserglyargaspproalaserargasp
145150155160
leuvalvalasntyrvalasnthrasnmetglyleulysilearggln
165170175
leuleutrpphehisilesercysleuthrpheglyarggluthrval
180185190
leuglutyrleuvalserpheglyvaltrpileargthrproproala
195200205
tyrargproproasnalaproileleuserthrleuprogluthrthr
210215220
valvalglyglyserserglyglyserglyglyserglyglysergly
225230235240
glyserglyglyserglyglyserthrmetaspileaspprotyrlys
245250255
glupheglyalathrvalgluleuleuserpheleuproseraspphe
260265270
pheproservalargaspleuleuaspthralaseralaleutyrarg
275280285
glualaleugluserprogluhiscysserprohishisthralaleu
290295300
argglnalaileleucystrpglygluleumetthrleualathrtrp
305310315320
valglyasnasnleuglupheglyglyglyglyserglyglyglygly
325330335
serglyglyglyglysermetserleuleuthrgluvalgluthrpro
340345350
thrargasnglytrpglyserlysserasnglyserseraspmetser
355360365
leuleuthrgluvalgluthrproileargasnglutrpglyserarg
370375380
serasnglyserseraspmetserleuleuthrgluvalgluthrpro
385390395400
thrargasnglutrpgluserargserserglyserseraspglygly
405410415
glyglyserglyglyglyglyserglyglyglyglyseralaserasp
420425430
proalaserargaspleuvalvalasntyrvalasnthrasnmetgly
435440445
leulysileargglnleuleutrpphehisilesercysleuthrphe
450455460
glyarggluthrvalleuglutyrleuvalserpheglyvaltrpile
465470475480
argthrproproalatyrargproproasnalaproileleuserthr
485490495
leuprogluthrthrvalvalargargargaspargglyargserpro
500505510
argargargthrproserproargargargargserglnserproarg
515520525
argargargserglnserarggluserglncysleugluhishishis
530535540
hishishis
545
<210>14
<211>24
<212>prt
<213>人工序列
<220>
<223>具有添加在c末端的oh基团的m2e序列
<220>
<221>mod_res
<222>(24)..(24)
<223>在c末端的oh基团的添加
<400>14
metserleuleuthrgluvalgluthrproileargasnglutrpgly
151015
cysargcysasnglyserserasp
20
<210>15
<211>7
<212>prt
<213>人工序列
<220>
<223>平衡hbcag中的α螺旋
<400>15
alaalaalaleualaalaala
15
<210>16
<211>14
<212>prt
<213>人工序列
<220>
<223>可能的接头
<400>16
glyglyglyglyserglyglyglyglyserglyglyglygly
1510
<210>17
<211>4807
<212>dna
<213>人工序列
<220>
<223>用于vlp1的dna序列
<400>17
agatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacag60
gtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgt120
tgcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaacc180
agcccagttattgggcttgattggagctcgctcattccaattccttctattaggctacta240
acaccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgttta300
tttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctg360
agtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgct420
gtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcg480
ttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcggcataccgtttgt540
cttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtct600
ctctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgct660
ttttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatact720
gctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaat780
atataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagctt840
actttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacga900
caacttgagaagatcaaaaaacaactaattattcgaaacgaggaattgccgtgatcactg960
cagaaaaaaatggacatcgacccatacaaagagttcggagctactgttgagttgttgtcc1020
ttcttgccttccgacttcttcccatccgttagagacttgttggacactgcttccgctttg1080
tacagagaagctttggaatctccagagcactgttctccacatcacactgctttgagacag1140
gctatcttgtgttggggtgagttgatgactttggctacttgggttggtaacaatctagaa1200
ggatccatgaatacacagttcacagcagtaggtaaagagttcaaccacctggaaaaaaga1260
atagagaatttaaataaaaaagttgatgatggtttcctggacatttggacttacaatgcc1320
gaactgttggttctattggaaaatgaaagaactttggactaccacgattcaaatgtgaag1380
aacttatatgaaaaggtaagaagccagttaaaaaacaatgccagcggccgcgatcctgct1440
tcaagagatttggttgttaattacgttaacactaacatgggtttgaagatcagacagttg1500
ttgtggttccacatctcctgtttgactttcggtagagagactgttttggagtacttggtt1560
tccttcggtgtttggatcagaactccaccagcttacagaccaccaaacgctccaattttg1620
tccactttgccagagactactgttgttggtgggagctctggtggttcaggtggatcaggt1680
ggttccggtggatctggtggatcaggtgggtcgactatggatatcgatccttacaaagaa1740
tttggtgctacagttgagttgttgtcttttttgccatctgatttttttccatctgttaga1800
gatttgttggatacagcttctgctttgtatagagaggctttggagtcccctgaacactgt1860
tcacctcaccatacagctttgagacaagctattttgtgttggggagaattgatgacattg1920
gctacatgggttggaaacaacttggaattcggtggtggtggtagcggaggtggtggttct1980
ggtggtggtggttcaatgagtcttctaaccgaggtcgaaacgcctactagaaacggttgg2040
gggtccaaatccaacggttcaagtgatatgagtcttctaaccgaggtcgaaacgcctatc2100
agaaacgaatgggggtccagatccaacggttcaagtgatatgagtcttctaaccgaggtc2160
gaaacgcctactagaaacgaatgggaatccagatcctccggttcaagtgatggtggtggt2220
ggaagcggaggtggtggttctggtggtggtggtgctagcgatccagcttccagagacttg2280
gttgttaattatgttaacacaaatatgggattgaaaattagacagttgttgtggtttcat2340
atttcttgtttgacatttggaagagaaacagttttggaatatttggtttcttttggagtt2400
tggattagaacacctcctgcttatagacctcctaatgctcctatcttgtctacattgcct2460
gaaacaacagttgttagaagaagagacagaggtagatccccaagaagaagaactccatct2520
cctagaagaagaagatcccagtcaccacgtagaagaagatcacaatccagagaatcccag2580
tgttaaggcgcctgatgactcgagggcccaccggtcttgctagattctaatcaagaggat2640
gtcagaatgccatttgcctgagagatgcaggcttcatttttgatacttttttatttgtaa2700
cctatatagtataggattttttttgtcattttgtttcttctcgtacgagcttgctcctga2760
tcagcctatctcgcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagt2820
ttgatgtttttcttggtatttcccactcctcttcagagtacagaagattaagtgagacct2880
tcgtttgtgcggatcccccacacaccatagcttcaaaatgtttctactccttttttactc2940
ttccagattttctcggactccgcgcatcgccgtaccacttcaaaacacccaagcacagca3000
tactaaattttccctctttcttcctctagggtgtcgttaattacccgtactaaaggtttg3060
gaaaagaaaaaagagaccgcctcgtttctttttcttcgtcgaaaaaggcaataaaaattt3120
ttatcacgtttctttttcttgaaatttttttttttagtttttttctctttcagtgacctc3180
cattgatatttaagttaataaacggtcttcaatttctcaagtttcagtttcatttttctt3240
gttctattacaactttttttacttcttgttcattagaaagaaagcatagcaatctaatct3300
aaggggcggtgttgacaattaatcatcggcatagtatatcggcatagtataatacgacaa3360
ggtgaggaactaaaccatggccaagttgaccagtgccgttccggtgctcaccgcgcgcga3420
cgtcgccggagcggtcgagttctggaccgaccggctcgggttctcccgggacttcgtgga3480
ggacgacttcgccggtgtggtccgggacgacgtgaccctgttcatcagcgcggtccagga3540
ccaggtggtgccggacaacaccctggcctgggtgtgggtgcgcggcctggacgagctgta3600
cgccgagtggtcggaggtcgtgtccacgaacttccgggacgcctccgggccggccatgac3660
cgagatcggcgagcagccgtgggggcgggagttcgccctgcgcgacccggccggcaactg3720
cgtgcacttcgtggccgaggagcaggactgacacgtccgacggcggcccacgggtcccag3780
gcctcggagatccgtcccccttttcctttgtcgatatcatgtaattagttatgtcacgct3840
tacattcacgccctccccccacatccgctctaaccgaaaaggaaggagttagacaacctg3900
aagtctaggtccctatttatttttttatagttatgttagtattaagaacgttatttatat3960
ttcaaatttttcttttttttctgtacagacgcgtgtacgcatgtaacattatactgaaaa4020
ccttgcttgagaaggttttgggacgctcgaaggctttaatttgcaagctggagaccaaca4080
tgtgagcaaaaggccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttt4140
tccataggctccgcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggc4200
gaaacccgacaggactataaagataccaggcgtttccccctggaagctccctcgtgcgct4260
ctcctgttccgaccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcg4320
tggcgctttctcaatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctcca4380
agctgggctgtgtgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaact4440
atcgtcttgagtccaacccggtaagacacgacttatcgccactggcagcagccactggta4500
acaggattagcagagcgaggtatgtaggcggtgctacagagttcttgaagtggtggccta4560
actacggctacactagaaggacagtatttggtatctgcgctctgctgaagccagttacct4620
tcggaaaaagagttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtt4680
tttttgtttgcaagcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttga4740
tcttttctacggggtctgacgctcagtggaacgaaaactcacgttaagggattttggtca4800
tgagatc4807
<210>18
<211>538
<212>prt
<213>人工序列
<220>
<223>串联核心,ha茎,3个m2e拷贝,接头
<400>18
metaspileaspprotyrlysglupheglyalathrvalgluleuleu
151015
serpheleuproseraspphepheproservalargaspleuleuasp
202530
thralaseralaleutyrargglualaleugluserprogluhiscys
354045
serprohishisthralaleuargglnalaileleucystrpglyglu
505560
leumetthrleualathrtrpvalglyasnasnleugluglysermet
65707580
asnthrglnphethralavalglylysglupheasnhisleuglulys
859095
argilegluasnleuasnlyslysvalaspaspglypheleuaspile
100105110
trpthrtyrasnalagluleuleuvalleuleugluasngluargthr
115120125
leuasptyrhisaspserasnvallysasnleutyrglulysvalarg
130135140
serglnleulysasnasnalaserglyargaspproalaserargasp
145150155160
leuvalvalasntyrvalasnthrasnmetglyleulysilearggln
165170175
leuleutrpphehisilesercysleuthrpheglyarggluthrval
180185190
leuglutyrleuvalserpheglyvaltrpileargthrproproala
195200205
tyrargproproasnalaproileleuserthrleuprogluthrthr
210215220
valvalglyglyserserglyglyserglyglyserglyglysergly
225230235240
glyserglyglyserglyglyserthrmetaspileaspprotyrlys
245250255
glupheglyalathrvalgluleuleuserpheleuproseraspphe
260265270
pheproservalargaspleuleuaspthralaseralaleutyrarg
275280285
glualaleugluserprogluhiscysserprohishisthralaleu
290295300
argglnalaileleucystrpglygluleumetthrleualathrtrp
305310315320
valglyasnasnleuglupheglyglyglyglyserglyglyglygly
325330335
serglyglyglyglysermetserleuleuthrgluvalgluthrpro
340345350
thrargasnglytrpglyserlysserasnglyserseraspmetser
355360365
leuleuthrgluvalgluthrproileargasnglutrpglyserarg
370375380
serasnglyserseraspmetserleuleuthrgluvalgluthrpro
385390395400
thrargasnglutrpgluserargserserglyserseraspglygly
405410415
glyglyserglyglyglyglyserglyglyglyglyalaserasppro
420425430
alaserargaspleuvalvalasntyrvalasnthrasnmetglyleu
435440445
lysileargglnleuleutrpphehisilesercysleuthrphegly
450455460
arggluthrvalleuglutyrleuvalserpheglyvaltrpilearg
465470475480
thrproproalatyrargproproasnalaproileleuserthrleu
485490495
progluthrthrvalvalargargargaspargglyargserproarg
500505510
argargthrproserproargargargargserglnserproargarg
515520525
argargserglnserarggluserglncys
530535
<210>19
<211>4495
<212>dna
<213>人工序列
<220>
<223>用于vlp2的dna序列
<400>19
agatctaacatccaaagacgaaaggttgaatgaaacctttttgccatccgacatccacag60
gtccattctcacacataagtgccaaacgcaacaggaggggatacactagcagcagaccgt120
tgcaaacgcaggacctccactcctcttctcctcaacacccacttttgccatcgaaaaacc180
agcccagttattgggcttgattggagctcgctcattccaattccttctattaggctacta240
acaccatgactttattagcctgtctatcctggcccccctggcgaggttcatgtttgttta300
tttccgaatgcaacaagctccgcattacacccgaacatcactccagatgagggctttctg360
agtgtggggtcaaatagtttcatgttccccaaatggcccaaaactgacagtttaaacgct420
gtcttggaacctaatatgacaaaagcgtgatctcatccaagatgaactaagtttggttcg480
ttgaaatgctaacggccagttggtcaaaaagaaacttccaaaagtcggcataccgtttgt540
cttgtttggtattgattgacgaatgctcaaaaataatctcattaatgcttagcgcagtct600
ctctatcgcttctgaaccccggtgcacctgtgccgaaacgcaaatggggaaacacccgct660
ttttggatgattatgcattgtctccacattgtatgcttccaagattctggtgggaatact720
gctgatagcctaacgttcatgatcaaaatttaactgttctaacccctacttgacagcaat780
atataaacagaaggaagctgccctgtcttaaacctttttttttatcatcattattagctt840
actttcataattgcgactggttccaattgacaagcttttgattttaacgacttttaacga900
caacttgagaagatcaaaaaacaactaattattcgaaacgaggaattgccgtgatcactg960
cagaaaaaaatggacatcgacccatacaaagagttcggagctactgttgagttgttgtcc1020
ttcttgccttccgacttcttcccatccgttagagacttgttggacactgcttccgctttg1080
tacagagaagctttggaatctccagagcactgttctccacatcacactgctttgagacag1140
gctatcttgtgttggggtgagttgatgactttggctacttgggttggtaacaatctagaa1200
ggatccagaatccaagatttggaaaagtacgttgaagatactaagattgatttgtggtct1260
tacaacgctgaattgttggttgctttggaaaaccaacatactattgatttgactgattct1320
gaaatgaacaagttgtttgaaaagactagaagacaattgagagaaaacgctagcggccgc1380
gatcctgcttcaagagatttggttgttaattacgttaacactaacatgggtttgaagatc1440
agacagttgttgtggttccacatctcctgtttgactttcggtagagagactgttttggag1500
tacttggtttccttcggtgtttggatcagaactccaccagcttacagaccaccaaacgct1560
ccaattttgtccactttgccagagactactgttgttggtgggagctctggtggttcaggt1620
ggatcaggtggttccggtggatctggtggatcaggtgggtcgactatggatatcgatcct1680
tacaaagaatttggtgctacagttgagttgttgtcttttttgccatctgatttttttcca1740
tctgttagagatttgttggatacagcttctgctttgtatagagaggctttggagtcccct1800
gaacactgttcacctcaccatacagctttgagacaagctattttgtgttggggagaattg1860
atgacattggctacatgggttggaaacaacttggaattcggttccggatcaggtggtgga1920
aagggtggtggatcaggttctgctagcgatccagcttccagagacttggttgttaattat1980
gttaacacaaatatgggattgaaaattagacagttgttgtggtttcatatttcttgtttg2040
acatttggaagagaaacagttttggaatatttggtttcttttggagtttggattagaaca2100
cctcctgcttatagacctcctaatgctcctatcttgtctacattgcctgaaacaacagtt2160
gttagaagaagagacagaggtagatccccaagaagaagaactccatctcctagaagaaga2220
agatcccagtcaccacgtagaagaagatcacaatccagagaatcccagtgttaaggcgcc2280
tgatgactcgagggcccaccggtcttgctagattctaatcaagaggatgtcagaatgcca2340
tttgcctgagagatgcaggcttcatttttgatacttttttatttgtaacctatatagtat2400
aggattttttttgtcattttgtttcttctcgtacgagcttgctcctgatcagcctatctc2460
gcagctgatgaatatcttgtggtaggggtttgggaaaatcattcgagtttgatgtttttc2520
ttggtatttcccactcctcttcagagtacagaagattaagtgagaccttcgtttgtgcgg2580
atcccccacacaccatagcttcaaaatgtttctactccttttttactcttccagattttc2640
tcggactccgcgcatcgccgtaccacttcaaaacacccaagcacagcatactaaattttc2700
cctctttcttcctctagggtgtcgttaattacccgtactaaaggtttggaaaagaaaaaa2760
gagaccgcctcgtttctttttcttcgtcgaaaaaggcaataaaaatttttatcacgtttc2820
tttttcttgaaatttttttttttagtttttttctctttcagtgacctccattgatattta2880
agttaataaacggtcttcaatttctcaagtttcagtttcatttttcttgttctattacaa2940
ctttttttacttcttgttcattagaaagaaagcatagcaatctaatctaaggggcggtgt3000
tgacaattaatcatcggcatagtatatcggcatagtataatacgacaaggtgaggaacta3060
aaccatggccaagttgaccagtgccgttccggtgctcaccgcgcgcgacgtcgccggagc3120
ggtcgagttctggaccgaccggctcgggttctcccgggacttcgtggaggacgacttcgc3180
cggtgtggtccgggacgacgtgaccctgttcatcagcgcggtccaggaccaggtggtgcc3240
ggacaacaccctggcctgggtgtgggtgcgcggcctggacgagctgtacgccgagtggtc3300
ggaggtcgtgtccacgaacttccgggacgcctccgggccggccatgaccgagatcggcga3360
gcagccgtgggggcgggagttcgccctgcgcgacccggccggcaactgcgtgcacttcgt3420
ggccgaggagcaggactgacacgtccgacggcggcccacgggtcccaggcctcggagatc3480
cgtcccccttttcctttgtcgatatcatgtaattagttatgtcacgcttacattcacgcc3540
ctccccccacatccgctctaaccgaaaaggaaggagttagacaacctgaagtctaggtcc3600
ctatttatttttttatagttatgttagtattaagaacgttatttatatttcaaatttttc3660
ttttttttctgtacagacgcgtgtacgcatgtaacattatactgaaaaccttgcttgaga3720
aggttttgggacgctcgaaggctttaatttgcaagctggagaccaacatgtgagcaaaag3780
gccagcaaaaggccaggaaccgtaaaaaggccgcgttgctggcgtttttccataggctcc3840
gcccccctgacgagcatcacaaaaatcgacgctcaagtcagaggtggcgaaacccgacag3900
gactataaagataccaggcgtttccccctggaagctccctcgtgcgctctcctgttccga3960
ccctgccgcttaccggatacctgtccgcctttctcccttcgggaagcgtggcgctttctc4020
aatgctcacgctgtaggtatctcagttcggtgtaggtcgttcgctccaagctgggctgtg4080
tgcacgaaccccccgttcagcccgaccgctgcgccttatccggtaactatcgtcttgagt4140
ccaacccggtaagacacgacttatcgccactggcagcagccactggtaacaggattagca4200
gagcgaggtatgtaggcggtgctacagagttcttgaagtggtggcctaactacggctaca4260
ctagaaggacagtatttggtatctgcgctctgctgaagccagttaccttcggaaaaagag4320
ttggtagctcttgatccggcaaacaaaccaccgctggtagcggtggtttttttgtttgca4380
agcagcagattacgcgcagaaaaaaaggatctcaagaagatcctttgatcttttctacgg4440
ggtctgacgctcagtggaacgaaaactcacgttaagggattttggtcatgagatc4495
<210>20
<211>434
<212>prt
<213>人工序列
<220>
<223>串联核心,ha茎,空插入体
<400>20
metaspileaspprotyrlysglupheglyalathrvalgluleuleu
151015
serpheleuproseraspphepheproservalargaspleuleuasp
202530
thralaseralaleutyrargglualaleugluserprogluhiscys
354045
serprohishisthralaleuargglnalaileleucystrpglyglu
505560
leumetthrleualathrtrpvalglyasnasnleugluglyserarg
65707580
ileglnaspleuglulystyrvalgluaspthrlysileaspleutrp
859095
sertyrasnalagluleuleuvalalaleugluasnglnhisthrile
100105110
aspleuthraspserglumetasnlysleupheglulysthrargarg
115120125
glnleuarggluasnalaserglyargaspproalaserargaspleu
130135140
valvalasntyrvalasnthrasnmetglyleulysileargglnleu
145150155160
leutrpphehisilesercysleuthrpheglyarggluthrvalleu
165170175
glutyrleuvalserpheglyvaltrpileargthrproproalatyr
180185190
argproproasnalaproileleuserthrleuprogluthrthrval
195200205
valglyglyserserglyglyserglyglyserglyglyserglygly
210215220
serglyglyserglyglyserthrmetaspileaspprotyrlysglu
225230235240
pheglyalathrvalgluleuleuserpheleuproseraspphephe
245250255
proservalargaspleuleuaspthralaseralaleutyrargglu
260265270
alaleugluserprogluhiscysserprohishisthralaleuarg
275280285
glnalaileleucystrpglygluleumetthrleualathrtrpval
290295300
glyasnasnleuglupheglyserglyserglyglyglylysglygly
305310315320
glyserglyseralaseraspproalaserargaspleuvalvalasn
325330335
tyrvalasnthrasnmetglyleulysileargglnleuleutrpphe
340345350
hisilesercysleuthrpheglyarggluthrvalleuglutyrleu
355360365
valserpheglyvaltrpileargthrproproalatyrargpropro
370375380
asnalaproileleuserthrleuprogluthrthrvalvalargarg
385390395400
argaspargglyargserproargargargthrproserproargarg
405410415
argargserglnserproargargargargserglnserarggluser
420425430
glncys
<210>21
<211>14
<212>prt
<213>人工序列
<220>
<223>空插入体
<400>21
glyserglyserglyglyglylysglyglyglyserglyser
1510
<210>22
<211>72
<212>prt
<213>人工序列
<220>
<223>ha茎403-474(来自h1n1)
<400>22
metasnthrglnphethralavalglylysglupheasnhisleuglu
151015
lysargilegluasnleuasnlyslysvalaspaspglypheleuasp
202530
iletrpthrtyrasnalagluleuleuvalleuleugluasngluarg
354045
thrleuasptyrhisaspserasnvallysasnleutyrglulysval
505560
argserglnleulysasnasnala
6570
<210>23
<211>55
<212>prt
<213>人工序列
<220>
<223>ha长α螺旋(lah)420-474(来自h1n1)
<400>23
argilegluasnleuasnlyslysvalaspaspglypheleuaspile
151015
trpthrtyrasnalagluleuleuvalleuleugluasngluargthr
202530
leuasptyrhisaspserasnvallysasnleutyrglulysvalarg
354045
serglnleulysasnasnala
5055
<210>24
<211>55
<212>prt
<213>人工序列
<220>
<223>ha长α螺旋(lah)421-475(来自h3n2)
<400>24
argileglnaspleuglulystyrvalgluaspthrlysileaspleu
151015
trpsertyrasnalagluleuleuvalalaleugluasnglnhisthr
202530
ileaspleuthraspserglumetasnlysleupheglulysthrarg
354045
argglnleuarggluasnala
5055
- 还没有人留言评论。精彩留言会获得点赞!