在多肽的重组生产期间防止二硫键还原的制作方法
本申请是申请日为2008年07月08日、中国申请号为200880105932.5、发明名称为“在多肽的重组生产期间防止二硫键还原”的发明申请的分案申请。
发明领域
本发明关注用于在含双硫的多肽(disulfide-containingpolypeptide)的重组生产期间防止二硫键还原的方法和手段。具体而言,本发明关注在自重组宿主细胞培养物收获含双硫的多肽(包括抗体)期间防止二硫键还原。
发明背景
在生物技术产业中,制药应用需要使用重组dna技术生产多种蛋白质。一般而言,由细胞培养物生成重组蛋白,使用经改造而生成感兴趣蛋白质的真核细胞(诸如哺乳动物细胞)或原核细胞(诸如细菌细胞),这通过插入含有编码期望蛋白质的核酸的重组质粒来进行。为了蛋白质保留生物学活性,必须在其纯化和分离期间维持蛋白质的构象(包括其三级结构),而且必须保护蛋白质的多个官能团免于降解。
哺乳动物细胞已经成为生产用于临床应用的哺乳动物蛋白质的主要系统,主要是由于它们生成正确折叠和装配的异源蛋白质的能力及它们翻译后修饰的能力。中国仓鼠卵巢(cho)细胞和自各种其它哺乳动物来源(诸如例如小鼠骨髓瘤(ns0)、幼仓鼠肾(bhk)、人胚肾(hek-293)和人肾细胞诸如自人肾细胞分离、提供人糖基化特征、且能够天然生成与人生理学匹配的抗体的
通常,为了开始生产周期,容许少量的经转化重组宿主细胞在培养物中生长几天(见例如图23)。一旦细胞进行了几轮复制,就将它们转移至更大的容器,在那里准备用它们进行发酵。用于培养细胞的培养基及生产周期期间存在的氧、氮和二氧化碳的水平可能对生产过程有显著影响。为每种细胞系具体确定培养参数,而且频繁地测量这些参数以确保最佳的生长和生产条件。
当细胞生长至足够数目时,将它们转移至大规模生产罐并培养更长的一段时间。在该过程中的此点,可以收获重组蛋白。典型的是,将细胞改造成分泌多肽入细胞培养液中,使得纯化过程的第一步是将细胞与培养液分开。典型的是,收获包括离心和过滤以生成已收获的细胞培养液(harvestedcellculturefluid,hccf)。然后将培养液进行几个另外的纯化步骤,除去任何细胞碎屑、不想要的蛋白质、盐、矿物或其它不想要的成分。在纯化过程结束时,重组蛋白是高度纯的,而且是适合于人治疗性用途的。
虽然在过去的几十年里对此过程进行了许多研究和改进,但是重组蛋白的生产仍然是困难重重。如此,例如在包含二硫键的多肽(尤其是包含链间二硫键的多链多肽,诸如抗体)的重组生产期间,重要的是要在整个生产、回收和纯化过程中保护和保持二硫键,以生成具有必要生物学活性的正确折叠的多肽。
发明概述
本发明一般涉及一种用于防止重组宿主细胞中表达的多肽中二硫键还原的方法,包括给所述重组宿主细胞的收获前的培养液(pre-harvestculturefluid)或已收获的培养液(harvestedculturefluid)补充硫氧还蛋白(thioredoxin)的抑制剂或硫氧还蛋白样蛋白(thioredoxin-likeprotein)。
在一个实施方案中,向所述收获前的培养液添加所述硫氧还蛋白抑制剂。
在另一个实施方案中,向所述已收获的培养液添加所述硫氧还蛋白抑制剂。
在又一个实施方案中,所述硫氧还蛋白抑制剂是硫氧还蛋白的直接抑制剂。
在所有实施方案中,所述硫氧还蛋白抑制剂可以是例如烃基-2-咪唑基二硫化物(alkyl-2-imidazolyldisulfide)或萘醌螺酮缩醇衍生物(naphthoquinonespiroketalderivative)。
在又一个实施方案中,所述硫氧还蛋白抑制剂是硫氧还蛋白还原酶的特异性抑制剂。
在还有一个实施方案中,所述硫氧还蛋白抑制剂是金复合物,其中所述金复合物可以是例如金硫葡糖(aurothioglucose,atg)或金硫苹果酸(aurothiomalate,atm)。虽然有效抑制浓度可以有所变化,但是它通常介于约0.1mm和1mm之间。类似地,最小有效抑制浓度随多肽性质和总体环境而变化,而且通常在atg或atm浓度是收获前的或已收获的培养液中硫氧还蛋白浓度的至少约四倍时达到。
在本发明此方面的另一个实施方案中,所述硫氧还蛋白抑制剂是金属离子,其中所述金属离子可以(不限于)选自下组:hg2+、cu2+、zn2+、co2+、和mn2+。当所述金属离子以硫酸铜的形式添加时,有效抑制浓度一般介于约5μm和约100μm之间,或介于约10μm和约80μm之间,或介于约15μm和约50μm之间。硫酸铜的最小抑制浓度也有所变化,但是通常在硫酸铜以收获前或已收获的培养液中硫氧还蛋白浓度至少约两倍的浓度添加时达到。
在不同的实施方案中,所述硫氧还蛋白抑制剂是氧化剂,例如g6pd的抑制剂,诸如例如吡哆醛-5’-磷酸、1-氟-2,4-二硝基苯、脱氢表雄酮(dhea)或表雄酮(ea);胱氨酸或半胱氨酸。dhea的典型有效抑制浓度介于约0.05mm和约5mm之间,或介于约0.1mm和约2.5mm之间。
在又一个实施方案中,所述硫氧还蛋白抑制剂是己糖激酶活性的抑制剂,包括但不限于金属离子的螯合剂,诸如例如乙二胺四乙酸(edta)。edta通常以如下浓度添加且在如下浓度是有效的:介于约5mm和约60mm之间,或介于约10mm和约50mm之间,或介于约20mm和约40mm之间。
在其它优选实施方案中,所述己糖激酶活性的抑制剂选自下组:山梨糖-1-磷酸、多磷酸、6-脱氧-6-氟葡萄糖、2-c-羟基-甲基葡萄糖、木糖、和来苏糖。
其它抑制剂包括胱氨酸、半胱氨酸、和氧化型谷胱甘肽,它们通常以收获前的或已收获的培养液中所讨论多肽浓度至少约40倍的浓度添加。
在还有一个实施方案中,所述硫氧还蛋白抑制剂是与硫氧还蛋白还原酶特异性结合的抗体、反义核苷酸、或sirna。
在另一个实施方案中,所述硫氧还蛋白抑制剂是间接导致对硫氧还蛋白活性的抑制的措施。此实施方案包括例如给重组宿主细胞的已收获的培养液鼓入空气和/或降低重组宿主细胞的已收获的培养液的ph。
在各种实施方案中,用于抑制硫氧还蛋白活性的间接手段(诸如鼓入空气和/或降低ph)可以与直接硫氧还蛋白抑制剂(诸如上文所列的那些)的使用组合。
在所有实施方案中,所述多肽可以是例如抗体或有生物学功能的抗体片段。代表性的抗体片段包括fab、fab′、f(ab′)2、scfv、(scfv)2、dab、互补决定区(cdr)片段、线性抗体、单链抗体分子、微型抗体、双抗体、和自抗体片段形成的多特异性抗体。
治疗性抗体包括但不限于抗her2抗体;抗cd20抗体;抗il-8抗体;抗vegf抗体;抗cd40抗体、抗cd11a抗体;抗cd18抗体;抗ige抗体;抗apo-2受体抗体;抗组织因子(tf)抗体;抗人α4β7整联蛋白抗体;抗egfr抗体;抗cd3抗体;抗cd25抗体;抗cd4抗体;抗cd52抗体;抗fc受体抗体;抗癌胚抗原(cea)抗体;针对乳房上皮细胞的抗体;与结肠癌细胞结合的抗体;抗cd38抗体;抗cd33抗体;抗cd22抗体;抗epcam抗体;抗gpiib/iiia抗体;抗rsv抗体;抗cmv抗体;抗hiv抗体;抗肝炎抗体;抗ca125抗体;抗αvβ3抗体;抗人肾细胞癌抗体;抗人17-1a抗体;抗人结肠直肠肿瘤抗体;抗人黑素瘤抗体;针对gd3神经节苷酯的r24;抗人鳞状细胞癌抗体;和抗人白细胞抗原(hla)抗体,和抗hladr抗体。
在其它实施方案中,所述治疗性抗体是与her受体、vegf、ige、cd20、cd11a、cd40、或dr5结合的抗体。
在又一个实施方案中,所述her受体是her1和/或her2,优选her2。所述her2抗体可以例如包含选自下组的重链和/或轻链可变域序列:seqidno:16、17、18、和19。
在另一个实施方案中,所述治疗性抗体是与cd20结合的抗体。所述抗cd20抗体可以例如包含选自下组的重链和/或轻链可变域序列:seqidno:1-15。
在还有一个实施方案中,所述治疗性抗体是与vegf结合的抗体。所述抗vegf可以例如包含选自下组的重链和/或轻链可变域序列:seqidno:20-25。
在另一个实施方案中,所述治疗性抗体是与cd11a结合的抗体。所述抗cd11a抗体可以例如包含选自下组的重链和/或轻链可变域序列:seqidno:26-29。
在又一个实施方案中,所述治疗性抗体与dr5受体结合。所述抗dr5抗体可以是例如选自下组:apomab1.1、2.1、3.1、4.1、5.1、5.2、5.3、6.1、6.2、6.3、7.1、7.2、7.3、8.1、8.3、9.1、1.2、2.2、3.2、4.2、5.2、6.2、7.2、8.2、9.2、1.3、2.2、3.3、4.3、5.3、6.3、7.3、8.3、9.3、和25.3,而且优选apomab8.3或apomab7.3,而且最优选apomab7.3。
在本发明方法的其它实施方案中,所述在重组宿主细胞中表达的多肽是治疗性多肽。例如,所述治疗性多肽可以选自下组:生长激素,包括人生长激素和牛生长激素;生长激素释放因子;甲状旁腺素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;促卵泡激素;降钙素;黄体生成素;高血糖素;凝血因子,诸如因子viiic、因子ix、组织因子、和vonwillebrands因子;抗凝血因子,诸如蛋白c;心房钠尿因子;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或人尿或组织型纤溶酶原激活剂(t-pa);铃蟾肽;凝血酶;造血生长因子;肿瘤坏死因子-α和-β;脑啡肽酶;rantes(在激活后受到调节,正常情况下由t细胞表达和分泌);人巨噬细胞炎症蛋白(mip-1-α);血清清蛋白,诸如人血清清蛋白;muellerian抑制性物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺素相关肽;微生物蛋白质,诸如β-内酰胺酶;dna酶;ige;细胞毒性t-淋巴细胞相关抗原(ctla),诸如ctla-4;抑制素;激活素;血管内皮生长因子(vegf);激素的或生长因子的受体;蛋白a或d;类风湿因子;神经营养性因子,诸如骨衍生神经营养性因子(bdnf)、神经营养因子-3、-4、-5、或-6(nt-3、nt-4、nt-5、或nt-6)、或神经生长因子,诸如ngf-β;血小板衍生生长因子(pdgf);成纤维细胞生长因子,诸如afgf和bfgf;表皮生长因子(egf);转化生长因子(tgf),诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4、或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-i)、胰岛素样生长因子结合蛋白;cd蛋白,诸如cd3、cd4、cd8、cd19、cd20、cd34、和cd40;红细胞生成素;骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);干扰素,诸如干扰素-α、-β,和-γ;集落刺激因子(csf),例如m-csf、gm-csf、和g-csf;白介素(il),例如il-1至il-10;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子;病毒抗原,诸如例如aids包膜的一部分;运输蛋白;归巢受体;地址素;调节蛋白;整联蛋白,诸如cd11a、cd11b、cd11c、cd18、icam、vla-4和vcam;肿瘤相关抗原,诸如her2、her3或her4受体;及以上所述多肽的片段。
在所有实施方案中,所述重组宿主细胞可以是真核细胞,诸如哺乳动物细胞,包括例如中国仓鼠卵巢(cho)细胞。
在所有实施方案中,所述重组宿主细胞也可以是原核细胞,诸如细菌细胞,包括但不限于大肠杆菌细胞。
本申请涉及下述实施方案。
1.一种用于防止重组宿主细胞中表达的多肽中二硫键还原的方法,包括给所述宿主细胞的收获前的或已收获的培养液补充硫氧还蛋白抑制剂。
2.实施方案1的方法,其中向所述收获前的培养液添加所述硫氧还蛋白抑制剂。
3.实施方案1的方法,其中向已收获的培养液添加所述硫氧还蛋白抑制剂。
4.实施方案1的方法,其中所述硫氧还蛋白抑制剂是硫氧还蛋白的直接抑制剂。
5.实施方案4的方法,其中所述硫氧还蛋白抑制剂是烃基-2-咪唑基二硫化物或萘醌螺酮缩醇衍生物。
6.实施方案1的方法,其中所述硫氧还蛋白抑制剂是硫氧还蛋白还原酶的特异性抑制剂。
7.实施方案6的方法,其中所述硫氧还蛋白抑制剂是金复合物。
8.实施方案7的方法,其中所述金复合物是金硫葡糖(atg)或金硫苹果酸(atm)。
9.实施方案8的方法,其中所述atg或所述atm以介于约0.1mm和约1mm之间的浓度添加。
10.实施方案8的方法,其中所述atg或所述atm以所述收获前的或已收获的培养液中硫氧还蛋白还原酶浓度至少约四倍的浓度添加。
11.实施方案1的方法,其中所述硫氧还蛋白抑制剂是金属离子。
12.实施方案11的方法,其中所述金属离子选自下组:hg2+、cu2+、zn2+、co2+和mn2+。
13.实施方案12的方法,其中所述金属离子是以硫酸铜的形式存在的cu2+。
14.实施方案13的方法,其中所述硫酸铜是五水合物的形式或无水的形式。
15.实施方案13的方法,其中所述硫酸铜以介于约5μm和约100μm之间的浓度添加。
16.实施方案13的方法,其中所述硫酸铜以介于约10μm至约80μm之间的浓度添加。
17.实施方案13的方法,其中所述硫酸铜以介于约15μm和约50μm之间的浓度添加。
18.实施方案13的方法,其中所述硫酸铜以所述收获前的或已收获的培养液中硫氧还蛋白浓度至少约两倍的浓度添加。
19.实施方案1的方法,其中所述硫氧还蛋白抑制剂是g6pd的抑制剂。
20.实施方案19的方法,其中所述硫氧还蛋白抑制剂选自下组:吡哆醛-5’-磷酸、1-氟-2,4-二硝基苯、脱氢表雄酮(dhea)和表雄酮(ea)。
21.实施方案20的方法,其中所述dhea以介于约0.05mm和约5mm之间的浓度添加。
22.实施方案20的方法,其中所述dhea以介于约0.1mm和约2.5mm之间的浓度添加。
23.实施方案1的方法,其中所述硫氧还蛋白抑制剂是己糖激酶活性的抑制剂。
24.实施方案23的方法,其中所述硫氧还蛋白抑制剂是金属离子的螯合剂。
25.实施方案24的方法,其中所述金属离子的螯合剂是乙二胺四乙酸(edta)。
26.实施方案25的方法,其中所述edta以介于约5mm和约60mm之间的浓度添加。
27.实施方案25的方法,其中所述edta以介于约10mm和约50mm之间的浓度添加。
28.实施方案25的方法,其中所述edta以介于约20mm和约40mm之间的浓度添加。
29.实施方案23的方法,其中所述硫氧还蛋白抑制剂选自下组:山梨糖-1-磷酸、多磷酸、6-脱氧-6-氟葡萄糖、2-c-羟基-甲基葡萄糖、木糖和来苏糖。
30.实施方案1的方法,其中所述硫氧还蛋白抑制剂是胱氨酸、半胱氨酸、或氧化型谷胱甘肽。
31.实施方案30的方法,其中所述胱氨酸、半胱氨酸或氧化型谷胱甘肽以所述收获前的或已收获的培养液中所述多肽浓度至少约40倍的浓度添加。
32.实施方案1的方法,其中所述硫氧还蛋白抑制剂是与硫氧还蛋白还原酶特异性结合的抗体、反义核苷酸、或sirna。
33.实施方案1的方法,其中所述硫氧还蛋白抑制剂是间接导致对硫氧还蛋白活性的抑制的措施。
34.实施方案33的方法,其中所述措施是给所述重组宿主细胞的已收获的培养液鼓入空气。
35.实施方案33的方法,其中所述措施是降低所述重组宿主细胞的已收获的培养液的ph。
36.实施方案1-33任一项的方法,进一步包括给所述重组宿主细胞的已收获的培养液鼓入空气的步骤。
37.实施方案1-33任一项的方法,进一步包括降低所述重组宿主细胞的已收获的培养液的ph的步骤。
38.实施方案1的方法,其中所述多肽是抗体或有生物学功能的抗体片段。
39.实施方案38的方法,其中所述抗体片段选自下组:fab、fab′、f(ab′)2、scfv、(scfv)2、dab、互补决定区(cdr)片段、线性抗体、单链抗体分子、微型抗体、双抗体和自抗体片段形成的多特异性抗体。
40.实施方案38的方法,其中所述抗体或抗体片段是治疗性抗体或其生物学功能片段。
41.实施方案40的方法,其中所述治疗性抗体选自下组:抗her2抗体;抗cd20抗体;抗il-8抗体;抗vegf抗体;抗cd40抗体,抗cd11a抗体;抗cd18抗体;抗ige抗体;抗apo-2受体抗体;抗组织因子(tf)抗体;抗人α4β7整联蛋白抗体;抗egfr抗体;抗cd3抗体;抗cd25抗体;抗cd4抗体;抗cd52抗体;抗fc受体抗体;抗癌胚抗原(cea)抗体;针对乳房上皮细胞的抗体;与结肠癌细胞结合的抗体;抗cd38抗体;抗cd33抗体;抗cd22抗体;抗epcam抗体;抗gpiib/iiia抗体;抗rsv抗体;抗cmv抗体;抗hiv抗体;抗肝炎抗体;抗ca125抗体;抗αvβ3抗体;抗人肾细胞癌抗体;抗人17-1a抗体;抗人结肠直肠肿瘤抗体;抗人黑素瘤抗体;针对gd3神经节苷酯的r24;抗人鳞状细胞癌抗体;和抗人白细胞抗原(hla)抗体和抗hladr抗体。
42.实施方案40的方法,其中所述治疗性抗体是与her受体、vegf、ige、cd20、cd11a、cd40或dr5结合的抗体。
43.实施方案42的方法,其中所述her受体是her1和/或her2。
44.实施方案43的方法,其中所述her受体是her2。
45.实施方案44的方法,其中所述治疗性抗体包含选自下组的重链和/或轻链可变域序列:seqidno:16、17、18和19。
46.实施方案42的方法,其中所述治疗性抗体是与cd20结合的抗体。
47.实施方案46的方法,其中所述治疗性抗体包含选自下组的重链和/或轻链可变域序列:seqidno:1-15。
48.实施方案42的方法,其中所述治疗性抗体是与vegf结合的抗体。
49.实施方案48的方法,其中所述治疗性抗体包含选自下组的重链和/或轻链可变域序列:seqidno:20-25。
50.实施方案42的方法,其中所述治疗性抗体是与cd11a结合的抗体。
51.实施方案50的方法,其中所述治疗性抗体包含选自下组的重链和/或轻链可变域序列:seqidno:26-29。
52.实施方案42的方法,其中所述治疗性抗体与dr5受体结合。
53.实施方案52的方法,其中所述治疗性抗体选自下组:apomab1.1、2.1、3.1、4.1、5.1、5.2、5.3、6.1、6.2、6.3、7.1、7.2、7.3、8.1、8.3、9.1、1.2、2.2、3.2、4.2、5.2、6.2、7.2、8.2、9.2、1.3、2.2、3.3、4.3、5.3、6.3、7.3、8.3、9.3和25.3。
54.实施方案52的方法,其中所述治疗性抗体是apomab8.3或apomab7.3。
55.实施方案54的方法,其中所述治疗性抗体是apomab7.3。
56.实施方案1的方法,其中所述多肽是治疗性多肽。
57.实施方案56的方法,其中所述治疗性多肽选自下组:生长激素,包括人生长激素和牛生长激素;生长激素释放因子;甲状旁腺素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;促卵泡激素;降钙素;黄体生成素;高血糖素;凝血因子,诸如因子viiic、因子ix、组织因子、和vonwillebrands因子;抗凝血因子,诸如蛋白c;心房钠尿因子;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或人尿或组织型纤溶酶原激活剂(t-pa);铃蟾肽;凝血酶;造血生长因子;肿瘤坏死因子-α和-β;脑啡肽酶;rantes(在激活后受到调节,正常情况下由t细胞表达和分泌);人巨噬细胞炎症蛋白(mip-1-α);血清清蛋白,诸如人血清清蛋白;muellerian抑制性物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺素相关肽;微生物蛋白质,诸如β-内酰胺酶;dna酶;ige;细胞毒性t-淋巴细胞相关抗原(ctla),诸如ctla-4;抑制素;激活素;血管内皮生长因子(vegf);激素的或生长因子的受体;蛋白a或d;类风湿因子;神经营养性因子,诸如骨衍生神经营养性因子(bdnf)、神经营养因子-3、-4、-5或-6(nt-3、nt-4、nt-5或nt-6)、或神经生长因子,诸如ngf-β;血小板衍生生长因子(pdgf);成纤维细胞生长因子,诸如afgf和bfgf;表皮生长因子(egf);转化生长因子(tgf),诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4、或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-i)、胰岛素样生长因子结合蛋白;cd蛋白,诸如cd3、cd4、cd8、cd19、cd20、cd34和cd40;红细胞生成素;骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);干扰素,诸如干扰素-α、-β和-γ;集落刺激因子(csf),例如m-csf、gm-csf和g-csf;白介素(il),例如il-1至il-10;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子;病毒抗原,诸如例如aids包膜的一部分;运输蛋白;归巢受体;地址素;调节蛋白;整联蛋白,诸如cd11a、cd11b、cd11c、cd18、icam、vla-4和vcam;肿瘤相关抗原,诸如her2、her3或her4受体;及以上所述多肽的片段。
58.实施方案1的方法,其中所述重组宿主细胞是真核细胞。
59.实施方案58的方法,其中所述真核细胞是哺乳动物细胞。
60.实施方案59的方法,其中所述哺乳动物细胞是中国仓鼠卵巢(cho)细胞。
61.实施方案1的方法,其中所述重组宿主细胞是原核细胞。
62.实施方案61的方法,其中所述原核细胞是细菌细胞。
63.实施方案62的方法,其中所述细菌细胞是大肠杆菌细胞。
附图简述
图1。透析实验:自bioanalyzer分析获得的数字凝胶样成像(每条带代表一个时间点),证明了透析袋内的ocrelizumab(rhumab2h7-变体a)在温育期间保持完整。
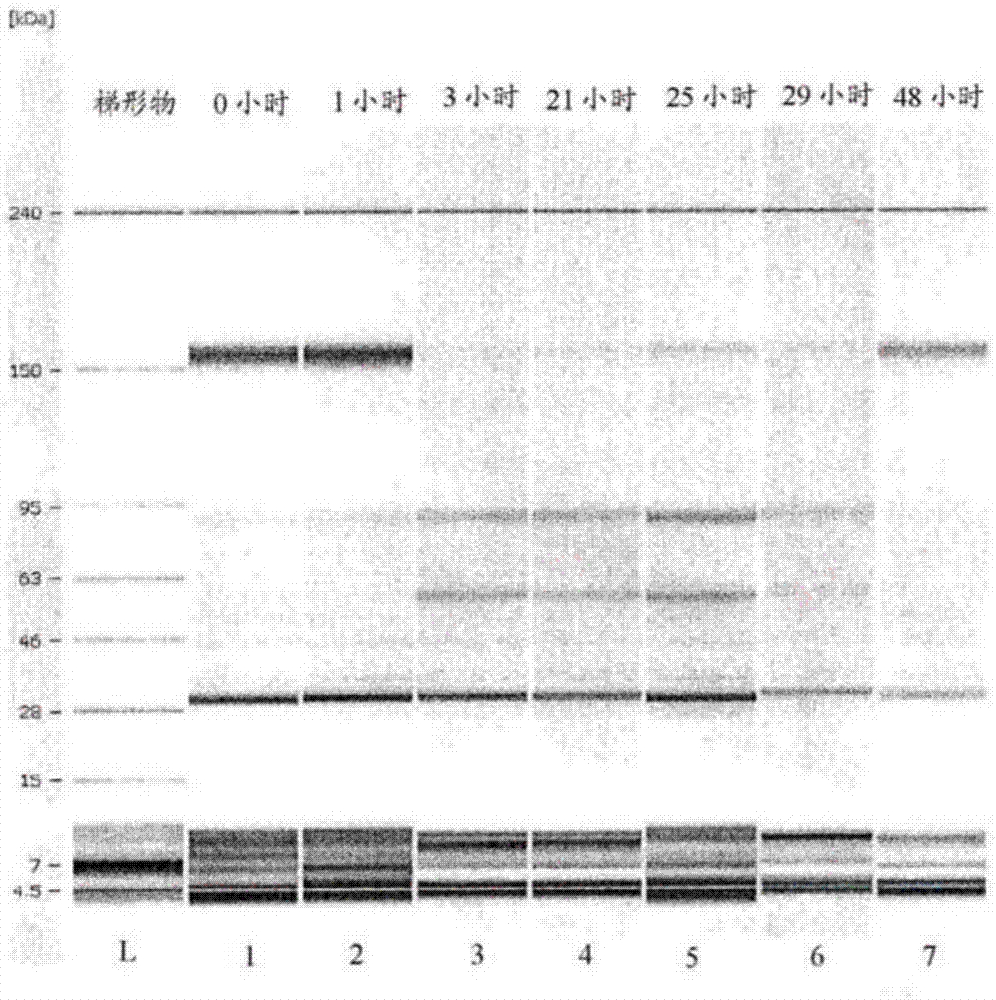
图2。透析实验:自bioanalyzer分析获得的数字凝胶样成像(每条带代表一个时间点),显示了透析袋外的ocrelizumab在温育期间被还原。这是通过图中所绘完整抗体(约150kda)的丢失和抗体片段的形成而证明的。在48小时时间点(第7道),被还原的抗体表现出被重新氧化,大概是已收获的细胞培养液(hccf)中还原活性丢失的结果。刚好在28kda标志物上方出现的条带源自抗体的轻链。在温育开始前hccf中就有显著量的游离轻链存在。hccf中过量的游离轻链和轻链二聚体的存在对于生成ocrelizumab的细胞系是典型的。
图3。来自透析实验的游离硫醇水平:磷酸盐缓冲盐水(pbs)中纯化的ocrelizumab在透析袋内,而含有ocrelizumab的hccf在袋外。透析袋内(方框)和外(菱形)游离硫醇在数小时内达到可比水平,指明hccf中的小分子成分在透析袋内与外之间有较好的交换。
图4。硫氧还蛋白系统和其它涉及抗体还原的反应:硫氧还蛋白系统(包含硫氧还蛋白(trx)、硫氧还蛋白还原酶(trxr)和nadph)为蛋白质中二硫键的还原发挥氢供体系统的功能。trx是具有经由硫醇-二硫化物交换催化许多氧化还原反应的cxxc活性位点基序的小单体蛋白质。被氧化的trx能被nadph经trxr还原。然后被还原的trx能够催化蛋白质中二硫化物的还原。硫氧还蛋白系统所需要的nadph是经由磷酸戊糖途径和糖酵解中的反应提供的。
图5。硫氧还蛋白系统的体外活性:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),证明了将完整ocrelizumab(1mg/ml)与pbs中的0.1mmtrxr(大鼠肝)、5mmtrx(人)、和1mmnadph一起温育导致ocrelizumab的完全还原;ocrelizumab在短于21小时里被完全还原。
图6。硫氧还蛋白系统的体外活性受到金硫葡糖抑制:以1mm的浓度向上文图5说明中所述相同反应混合物添加金硫葡糖(atg)有效抑制ocrelizumab还原。这是通过来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点)看到的。
图7。硫氧还蛋白系统的体外活性受到金硫苹果酸抑制:以1mm的浓度向上文图5说明中所述相同反应混合物添加金硫苹果酸(atm)有效抑制ocrelizumab还原。这是通过来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点)看到的。
图8。硫氧还蛋白系统的体外活性:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了将完整ocrelizumab(1mg/ml)与10mm硫酸组氨酸缓冲液中的0.1mmtrxr(大鼠肝)、5mmtrx(人)、和1mmnadph一起温育导致ocrelizumab在短于1小时里被还原。
图9。硫氧还蛋白系统的体外活性受到cuso4抑制:以50μm的浓度向图8说明中所述相同反应混合物添加cuso4有效抑制ocrelizumab还原,如来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点)中所示。
图10。ocrelizumab还原:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了ocrelizumab在使用来自匀浆化ccf(其是自3l发酵罐生成的)的hccf进行的温育实验中被还原。
图11。hccf中的ocrelizumab还原受到金硫葡糖抑制:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了向图10说明中所述温育实验所使用的相同hccf添加1mm金硫葡糖有效抑制ocrelizumab还原。
图12。hccf中的ocrelizumab还原受到金硫苹果酸抑制:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),指明了向图10说明中所述温育实验所使用的相同hccf添加1mm金硫苹果酸有效抑制ocrelizumab还原。
图13。hccf中还原活性丢失:进行过数轮冻融循环的、来自ocrelizumab大规模制造运行之一(“β”运行)(12kl运行8)的hccf在温育实验中使用时证明了没有ocrelizumab还原。这是通过bioanalyzer分析(每条带代表一个时间点)显示的,而且能与先前在新鲜融化的、来自同一发酵批次的hccf中看到的抗体还原形成对比。
图14。hccf中丢失的还原活性通过添加nadph而恢复:在以5mm的浓度向hccf(那里的还原活性在上文图13中所述条件下已经被消除)中添加nadph后在bioanalyzer分析(每条带代表一个时间点)中再次观察到ocrelizumab还原。
图15。hccf中丢失的还原活性通过添加葡萄糖-6-磷酸而恢复:在以10mm的浓度向hccf(那里的还原活性由于上文图13中所述处理已经被消除)中添加g6p后在bioanalyzer分析(每条带代表一个时间点)中再次观察到ocrelizumab还原。
图16。ocrelizumab还原:来自bioanalyzer分析的数字凝胶样图像,显示了ocrelizumab在使用来自大规模制造运行(“α”运行)(12kl运行9)的hccf进行的温育实验中被还原。
图17。edta抑制ocrelizumab还原:来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了ocrelizumab还原在使用来自α运行的hccf进行的温育实验中受到抑制,其中以20mm的浓度向hccf(其还原活性展示于图16中)添加edta。
图18。“β”运行hccf中丢失的还原活性通过添加葡萄糖-6-磷酸而恢复,但是edta不抑制还原:在以5mm的浓度向hccf(其还原活性已经丢失,见图13)中添加g6p和20mmedta后在bioanalyzer分析(每条带代表一个时间点)中观察到ocrelizumab还原。与图17中所示结果形成对比,edta的存在不阻断ocreliumab还原。
图19。通过(i)添加edta、(ii)添加cuso4、或(iii)调节ph至5.5来抑制ocrelizumab还原:独立使用的所有三种不同方法,即(1)添加edta、(2)添加cuso4、和(3)调节ph至5.5,都有效抑制ocrelizumab还原。这是通过所描绘的定量bioanalyzer结果证明的,该结果显示了几乎100%的完整(150kda)抗体保留在蛋白a洗脱合并液中。与此对比,在对照hccf中在20小时hccf保持时间后ocrelizumab被完全还原。
图20。鼓入空气抑制ocrelizumab还原:给hccf鼓入空气有效抑制ocrelizumab二硫键还原。这是通过定量bioanalyzer结果证明的,该结果显示了几乎100%的完整(150kda)保留在蛋白a洗脱合并液中。与此对比,在对照hccf中在鼓入氮气5小时后ocrelizumab被几乎完全还原。
图21显示了抗her2抗体(trastuzumab)的vl(seqidno.24)氨基酸序列。
图22显示了抗her2抗体(trastuzumab)的vh(seqidno.25)氨基酸序列。
图23是示意图,显示了典型的大规模制造过程的一些步骤。
图24是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸中。
图25是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在1mm硫酸组氨酸+1mmatg中。
图26是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+0.6μmatg中(6:1atg:trxr)。
图27是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+0.4μmatg中(4:1atg:trxr)。
图28是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+0.2μmatg中(2:1atg:trxr)。
图29是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+0.1mm金硫苹果酸(atm)中。
图30是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+0.01mm金硫苹果酸(atm)中。
图31是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+20μmcuso4中(4:1cu2+:trx)。
图32是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+10μmcuso4中(2:1cu2+:trx)。
图33是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+5μmcuso4中(1:1cu2+:trx)。
图34是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+532μm胱胺中(20:1胱胺:2h7二硫化物)。
图35是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+266μm胱胺中(10:1胱胺:2h7二硫化物)。
图36是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+133μm胱胺中(5:1胱胺:2h7二硫化物)。
图37是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+26.6μm胱胺中(1:1胱胺:2h7二硫化物)。
图38是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸(ph=7.6)+2.6mm胱氨酸中。
图39是来自bioanalyzer分析的数字凝胶样图像:2h7(变体a)+1mmnadph+5μm硫氧还蛋白+0.1μm硫氧还蛋白还原酶(重组体),在10mm硫酸组氨酸+2.6mmgssg(氧化型谷胱甘肽)中。
图40。重建的酶促还原系统。1mg/ml2h7(变体a)+10μg/ml己糖激酶、50μg/ml葡萄糖-6-磷酸脱氢酶、5μm硫氧还蛋白、0.1μm硫氧还蛋白还原酶、2mm葡萄糖、0.6mmatp、2mmmg2+、和2mmnadp,在ph=7.38的50mm硫酸组氨酸缓冲液中。
图41。硫氧还蛋白系统需要nadph。1mg/ml2h7(变体a)+5μm硫氧还蛋白、0.1μm硫氧还蛋白还原酶、和2mmnadp,在ph=7.38的50mm硫酸组氨酸缓冲液中。
发明详述
i.定义
在本发明中,在蛋白质(包括抗体)的语境中,一般而言或就任何特定蛋白质或抗体而言,术语“还原”用于指蛋白质或抗体的一个或多个二硫键的还原。如此,例如,术语“ocrelizumab还原”与术语“ocrelizumab二硫键还原”可互换使用,而术语“抗体(ab)还原”与术语“抗体(ab)二硫键还原”可互换使用。
术语“还原”或“二硫键还原”以最广义使用,而且包括完全的和部分的还原及蛋白质(诸如抗体)中存在的一些或所有(链间的或链内的)二硫键的还原。
“蛋白质”意指链长度足以生成更高水平的三级和/或四级结构的氨基酸序列。这与没有此类结构的“肽”或其它小分子量药物有区别。典型的是,本文中的蛋白质会具有至少约15-20kd、优选至少约20kd的分子量。本文中定义内所涵盖的蛋白质的例子包括所有哺乳动物蛋白质,特别是治疗性和诊断性蛋白质,诸如治疗性和诊断性抗体,及一般而言含有一个或多个二硫键的蛋白质,包括包含一个或多个链间和/或链内二硫键的多链多肽。
术语“治疗性蛋白质”或“治疗性多肽”指在疾病治疗中使用的蛋白质,不管其适应症或作用机制。为了使治疗性蛋白质在临床中有用,必须大量制造它。治疗性蛋白质或其它蛋白质的“制造规模”生产利用范围为约400l至约80,000l的细胞培养物,取决于所生产的蛋白质和需求。典型的是,此类制造规模生产利用量为约400l至约25,000l的细胞培养物。在此范围内,利用特定的细胞培养物量,诸如4,000l、约6,000l、约8,000、约10,000、约12,000l、约14,000l、或约16,000l。
术语“治疗性抗体”指在疾病治疗中使用的抗体。治疗性抗体可具有各种作用机制。治疗性抗体可结合并中和与抗原有关的靶物的正常功能。例如,阻断癌细胞存活所需要的蛋白质的活性的单克隆抗体引起细胞死亡。另一种治疗性单克隆抗体可结合并活化与抗原有关的靶物的正常功能。例如,单克隆抗体能结合细胞上的蛋白质并触发凋亡信号。又一种单克隆抗体可结合只在患病组织上表达的靶抗原;给单克隆抗体偶联毒性净荷(有效药剂)(诸如化疗剂或放射性药剂)能创建向患病组织特异性投递毒性净荷的药剂,降低对健康组织的损害。治疗性抗体的“生物学功能性片段”会展现至少一项(如果不是一些或所有的话)归于完整抗体的生物学功能,该功能至少包含特异性结合靶抗原。
术语“诊断性蛋白质”指在疾病诊断中使用的蛋白质。
术语“诊断性抗体”指作为疾病诊断试剂使用的抗体。诊断性抗体可结合与特定疾病特异性相关的或在特定疾病中显示表达升高的靶抗原。诊断性抗体可用于例如检测来自患者的生物学样品中的靶物,或用在患者中疾病部位(诸如肿瘤)的诊断成像中。诊断性抗体的“生物学功能性片段”会展现至少一项(如果不是一些或所有的话)归于完整抗体的生物学功能,该功能至少包含特异性结合靶抗原。
“纯化的”意味着分子以至少80-90%(以包含它的样品的重量计)的浓度存在于样品中。
纯化的蛋白质(包括抗体)优选是基本上纯的且期望是基本上同质的(即不含污染性蛋白质等)。
“基本上纯的”蛋白质意味着基于组合物的总重量,蛋白质组合物包含至少约90%(以重量计)、优选至少约95%(以重量计)的蛋白质。
“基本上同质的”蛋白质意味着基于组合物的总重量,蛋白质组合物包含至少约99%(以重量计)的蛋白质。
如上所述,在某些实施方案中,所述蛋白质是抗体。“抗体”(ab)和“免疫球蛋白”(ig)是具有相同结构特征的糖蛋白。虽然抗体展现出对特定抗原的结合特异性,但是免疫球蛋白包括抗体和一般缺乏抗原特异性的其它抗体样分子二者。后一类多肽例如由淋巴系统以低水平生成而由骨髓瘤以升高的水平生成。
术语“抗体”以最广义使用,明确覆盖单克隆抗体(包括具有免疫球蛋白fc区的全长抗体)、多克隆抗体、具有多表位特异性的抗体组合物、双特异性抗体、双抗体、和单链分子诸如scfv分子、以及抗体片段(例如fab、f(ab')2、和fv)。
术语“单克隆抗体”在用于本文时指从一群基本上同质的抗体获得的抗体,即构成群体的各个抗体相同,除了可能以极小量存在的可能的突变,例如天然存在的突变。如此,修饰语“单克隆”表明抗体不是分立的抗体的混合物的特征。在某些实施方案中,此类单克隆抗体典型地包括包含结合靶物的多肽序列的抗体,其中靶物结合多肽序列是通过包括从众多多肽序列中选择单一靶物结合多肽序列在内的过程得到的。例如,选择过程可以是从众多克隆诸如杂交瘤克隆、噬菌体克隆或重组dna克隆的集合中选择独特克隆。应当理解,所选择的靶物结合序列可进一步改变,例如为了提高对靶物的亲和力、将靶物结合序列人源化、提高其在细胞培养物中的产量、降低其在体内的免疫原性、创建多特异性抗体等,而且包含改变后的靶物结合序列的抗体也是本发明的单克隆抗体。与典型的包含针对不同决定簇(表位)的不同抗体的多克隆抗体制备物不同,单克隆抗体制备物的每个单克隆抗体针对抗原上的单一决定簇。除它们的特异性外,单克隆抗体制备物的优势还在于它们通常未受到其它免疫球蛋白的污染。
修饰语“单克隆”表明抗体从基本上同质的抗体群获得的特征,不应解释为要求通过任何特定方法来生产抗体。例如,将依照本发明使用的单克隆抗体可通过多种技术来生成,包括例如杂交瘤法(例如kohleretal.,nature256:495(1975);harlowetal.,antibodies:alaboratorymanual,coldspringharborlaboratorypress,2nded.1988;hammerlingetal.,于:monoclonalantibodiesandt-cellhybridomas,563-681,elsevier,n.y.,1981)、重组dna法(参见例如美国专利no.4,816,567)、噬菌体展示技术(参见例如clacksonetal.,nature352:624-628(1991);marksetal.,j.mol.biol.222:581-597(1992);sidhuetal.,j.mol.biol.338(2):299-310(2004);leeetal.,j.mol.biol.340(5):1073-1093(2004);fellouse,proc.nat.acad.sci.usa101(34):12467-12472(2004);leeetal.,j.immunol.methods284(1-2):119-132(2004))、及用于在具有部分或整个人免疫球蛋白基因座或编码人免疫球蛋白序列的基因的动物中生成人或人样抗体的技术(参见例如wo98/24893;wo96/34096;wo96/33735;wo91/10741;jakobovitsetal.,proc.natl.acad.sci.usa90:2551(1993);jakobovitsetal.,nature362:255-258(1993);bruggemannetal.,yearinimmuno.7:33(1993);美国专利no.5,545,807;5,545,806;5,569,825;5,625,126;5,633,425;5,661,016;marksetal.,bio/technology10:779-783(1992);lonbergetal.,nature368:856-859(1994);morrison,nature368:812-813(1994);fishwildetal.,naturebiotechnol.14:845-851(1996);neuberger,naturebiotechnol.14:826(1996);lonbergandhuszar,intern.rev.immunol.13:65-93(1995))。
单克隆抗体在本文中明确包括“嵌合”抗体,以及此类抗体的片段,只要它们展现出期望的生物学活性,其中所述抗体的重链和/或轻链的一部分与衍生自特定物种或属于特定抗体类别或亚类的抗体中的相应序列相同或同源,而链的剩余部分与衍生自另一物种或属于另一抗体类别或亚类的抗体中的相应序列相同或同源,(美国专利no.4,816,567和morrisonetal.,proc.natl.acad.sci.usa81:6851-6855(1984))。
非人(例如鼠)抗体的“人源化”形式指最低限度包含衍生自非人免疫球蛋白的序列的嵌合抗体。在一个实施方案中,人源化抗体指人免疫球蛋白(受体抗体)中的高变区残基用具有期望特异性、亲和力和/或能力的非人物种(供体抗体)(诸如小鼠、大鼠、家兔、或非人灵长类动物)的高变区残基替换的免疫球蛋白。在有些情况中,将人免疫球蛋白的框架区(fr)残基用相应的非人残基替换。此外,人源化抗体可包含在受体抗体中或在供体抗体中没有找到的残基。进行这些修饰可以是为了进一步改进抗体的性能。一般而言,人源化抗体将包含至少一个、通常两个基本上整个如下的可变域,其中所有或基本上所有高变环对应于非人免疫球蛋白的高变环,且所有或基本上所有fr是人免疫球蛋白序列的fr区。人源化抗体任选还将包含至少部分免疫球蛋白恒定区(fc),通常是人免疫球蛋白的恒定区。更多细节参见jonesetal.,nature321:522-525(1986);riechmannetal.,nature332:323-329(1988);及presta,curr.op.struct.biol.2:593-596(1992)。还可参见以下综述及其引用的参考文献:vaswaniandhamilton,ann.allergy,asthma&immunol.1:105-115(1998);harris,biochem.soc.transactions23:1035-1038(1995);hurleandgross,curr.op.biotech.5:428-433(1994)。人源化抗体包括primatizedtm(灵长类化)抗体,其中抗体的抗原结合区衍生自通过用感兴趣抗原免疫猕猴而生成的抗体。
“人抗体”指拥有与由人生成的抗体的氨基酸序列对应的氨基酸序列和/或使用本文所公开的用于生成人抗体的任何技术生成的抗体。人抗体的这种定义明确排除包含非人抗原结合残基的人源化抗体。
“亲和力成熟的”抗体指在抗体的一个或多个cdr/hvr中具有一处或多处改变、导致该抗体对抗原的亲和力与没有这些改变的亲本抗体相比有所改进的抗体。优选的亲和力成熟的抗体具有纳摩尔或甚至皮摩尔量级的对靶抗原的亲和力。亲和力成熟的抗体可通过本领域已知规程来生成。marksetal.,bio/technology10:779-783(1992)记载了通过vh和vl结构域改组进行的亲和力成熟。以下文献记载了cdr/hvr和/或框架残基的随机诱变:barbasetal.,proc.nat.acad.sci.usa91:3809-3813(1994);schieretal.,gene169:147-155(1995);yeltonetal.,j.immunol.155:1994-2004(1995);jacksonetal.,j.immunol.154(7):3310-9(1995);hawkinsetal.,j.mol.biol.226:889-896(1992)。
抗体的“可变区”或“可变域”指抗体重链或轻链的氨基末端结构域。重链的可变域可以称为“vh”。轻链的可变域可以称为“vl”。这些结构域一般是抗体的最易变部分且包含抗原结合位点。
术语“可变的”指可变域中的某些部分在抗体序列间差异广泛且用于每种特定抗体对其特定抗原的结合和特异性的实情。然而,变异性并非均匀分布于抗体的整个可变域。它集中于轻链和重链可变域中称作互补决定区(cdr)或高变区(hvr)的三个区段。可变域中相对较高度保守的部分称作框架区(fr)。天然重链和轻链的可变域各自包含四个fr,它们大多采取β-折叠片构象,通过形成环状连接且在有些情况中形成β-折叠片结构一部分的三个cdr连接。每条链中的cdr通过fr非常接近地保持在一起,并与另一条链的cdr一起促成抗体的抗原结合位点的形成(参见kabatetal.,sequencesofproteinsofimmunologicalinterest,第5版,nationalinstituteofhealth,bethesda,md.(1991))。恒定域不直接参与抗体与抗原的结合,但展现出多种效应器功能,诸如抗体依赖性细胞的细胞毒性中抗体的参与。
根据其恒定域的氨基酸序列,来自任何脊椎动物物种的抗体(免疫球蛋白)的“轻链”可归入两种截然不同的型中的一种,称作卡帕(κ)和拉姆达(λ)。
根据其重链恒定域的氨基酸序列,抗体(免疫球蛋白)可归入不同的类。免疫球蛋白有五大类:iga、igd、ige、igg和igm,其中有些可进一步分为亚类(同种型),例如igg1、igg2、igg3、igg4、iga1和iga2。将与不同类的免疫球蛋白对应的重链恒定域分别称作α、δ、ε、γ和μ。不同类别免疫球蛋白的亚基结构和三维构造是众所周知的,一般性描述于例如abbasetal.,cellularandmol.immunology,4thed.(2000)。抗体可以是抗体与一种或多种其它蛋白质或肽共价或非共价连接而形成的更大融合分子的一部分。
术语“全长抗体”和“完整抗体”在本文中可互换使用,指基本上完整形式的抗体而非如下文所定义的抗体片段。该术语具体指重链包含fc区的抗体。
“抗体片段”只包含完整抗体的一部分,其中所述部分保留该部分存在于完整抗体中时通常与之有关的至少一项、多至大多数或所有功能。在一个实施方案中,抗体片段包含完整抗体的抗原结合位点,如此保留结合抗原的能力。在另一个实施方案中,抗体片段,例如包含fc区的抗体片段,保留通常与fc区存在于完整抗体中时通常与之有关的至少一项生物学功能,诸如fcrn结合、抗体半衰期调控、adcc功能和补体结合。在一个实施方案中,抗体片段是体内半衰期与完整抗体基本上相似的单价抗体。例如,这样的抗体片段可包含一个抗原结合臂且其与能够赋予该片段以体内稳定性的fc序列相连。
用木瓜蛋白酶消化抗体产生两个相同的抗原结合片段,称为“fab”片段,各自具有一个抗原结合位点,及剩余的“fc”片段,其名称反映了它易于结晶的能力。胃蛋白酶处理产生f(ab')2片段,它具有两个抗原结合位点且仍能够交联抗原。
fab片段包含重链可变域和轻链可变域,而且还包含轻链的恒定域和重链的第一恒定域(ch1)。fab'片段与fab片段的不同之处在于重链ch1结构域的羧基末端增加了少数残基,包括来自抗体铰链区的一个或多个半胱氨酸。fab'-sh是本文中对其中恒定域半胱氨酸残基携带游离硫醇基的fab'的称谓。f(ab')2抗体片段最初是作为在成对fab'片段之间有铰链半胱氨酸的成对fab'片段生成的。还知道抗体片段的其它化学偶联形式。
“fv”是包含完整抗原结合位点的最小抗体片段。在一个实施方案中,双链fv种类由紧密、非共价结合的一个重链可变域和一个轻链可变域的二聚体组成。在单链fv(scfv)种类中,一个重链可变域和一个轻链可变域可以通过柔性肽接头共价相连接,使得轻链和重链在与双链fv种类类似的“二聚体”结构中相结合。正是在这种构造中,每个可变域的三个cdr相互作用而在vh-vl二聚体表面上限定了抗原结合位点。六个cdr一起赋予抗体以抗原结合特异性。然而,即使是单个可变域(或是只包含对抗原特异性的三个cdr的半个fv)也具有识别和结合抗原的能力,只是亲和力低于完整结合位点。
“单链fv”或“scfv”抗体片段包含抗体的vh和vl结构域,其中这些结构域存在于一条多肽链上。一般而言,scfv多肽在vh与vl结构域之间进一步包含多肽接头,其使得scfv能够形成结合抗原的期望结构。关于scfv的综述参见pluckthun,于《thepharmacologyofmonoclonalantibodies》,第113卷,rosenburg和moore编,springer-verlag,newyork,第269-315页,1994。
术语“双抗体”指具有两个抗原结合位点的小型抗体片段,该片段在同一条多肽链(vh-vl)中包含相连的重链可变域(vh)和轻链可变域(vl)。通过使用过短的接头使得同一条链上的两个结构域之间不能配对,迫使这些结构域与另一条链的互补结构域配对,从而产生两个抗原结合位点。双抗体可以是二价的或双特异性的。双抗体更完整的记载于例如ep404,097;wo93/1161;hudsonetal.(2003)nat.med.9:129-134;及hollingeretal.,proc.natl.acad.sci.usa90:6444-6448(1993)。三抗体(triabody)和四抗体(tetrabody)也记载于hudsonetal.(2003)nat.med.9:129-134。
抗体可结合任何蛋白质,包括但不限于her受体家族的成员,诸如her1(egfr)、her2、her3和her4;cd蛋白,诸如cd3、cd4、cd8、cd19、cd20、cd21、cd22、和cd34;细胞粘附分子,诸如lfa-1、mol、p150、95、vla-4、icam-1、vcam和αvβ3整联蛋白,包括其α或β亚基(例如抗cd11a、抗cd18或抗cd11b抗体);生长因子,诸如血管内皮生长因子(vegf);ige;血型抗原;flk2/flt3受体;肥胖(ob)受体;和蛋白c。其它例示性蛋白质包括生长激素(gh),包括人生长激素(hgh)和牛生长激素(bgh);生长激素释放因子;甲状旁腺素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;促卵泡激素;降钙素;黄体生成素;高血糖素;凝血因子,诸如因子viiic、因子ix、组织因子、和vonwillebrands因子;抗凝血因子,诸如蛋白c;心房钠尿因子;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或组织型纤溶酶原激活剂(t-pa);铃蟾肽;凝血酶;肿瘤坏死因子-α和-β;脑啡肽酶;rantes(在激活后受到调节,正常情况下由t细胞表达和分泌);人巨噬细胞炎症蛋白(mip-1-α);血清清蛋白,诸如人血清清蛋白(hsa);muellerian抑制性物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺素相关肽;dna酶;抑制素;激活素;激素的或生长因子的受体;整联蛋白;蛋白a或d;类风湿因子;神经营养性因子,诸如骨衍生神经营养性因子(bdnf)、神经营养因子-3、-4、-5、或-6(nt-3、nt-4、nt-5、或nt-6)、或神经生长因子,诸如ngf-β;血小板衍生生长因子(pdgf);成纤维细胞生长因子,诸如afgf和bfgf;表皮生长因子(egf);转化生长因子(tgf),诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4、或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-i);胰岛素样生长因子结合蛋白(igfbp);红细胞生成素(epo);血小板生成素(tpo);骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);干扰素,诸如干扰素-α、-β、和-γ;集落刺激因子(csf),例如m-csf、gm-csf、和g-csf;白介素(il),例如il-1至il-10;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子(daf);病毒抗原,诸如例如aids包膜的一部分;运输蛋白;归巢受体;地址素;调节蛋白;免疫粘附素;抗体;及任何上文所列多肽的生物学活性片段或变体。许多其它抗体和/或其它蛋白质可依照本发明使用,而且上文列表并非意欲限制本发明。
抗体的“生物学功能性片段”只包含完整抗体的一部分,其中所述部分保留该部分存在于完整抗体中时通常与之有关的至少一项、多至大多数或所有功能。在一个实施方案中,抗体的生物学功能性片段包含完整抗体的抗原结合位点,如此保留结合抗原的能力。在另一个实施方案中,抗体的生物学功能性片段(例如包含fc区的)保留通常与fc区存在于完整抗体中时通常与之有关的至少一项生物学功能,诸如fcrn结合、抗体半衰期调控、adcc功能和补体结合。在一个实施方案中,抗体的生物学功能性片段是体内半衰期与完整抗体基本上相似的单价抗体。例如,这样的抗体的生物学功能性片段可包含一个抗原结合臂且其与能够赋予该片段以体内稳定性的fc序列相连。
术语“硫氧还蛋白抑制剂”和“trx抑制剂”可互换使用,包括有效抑制硫氧还蛋白活性的所有试剂和措施。如此硫氧还蛋白(trx)抑制剂包括阻断trx、g6pd和/或己糖激酶系统的任何成分的所有试剂和措施。在此语境中,“抑制”包括完全消除(阻断)和降低硫氧还蛋白活性,及因此完全或部分消除蛋白质(诸如抗体)中的二硫键还原。
“分离的”抗体指已经鉴定且自其天然环境的成分分开和/或回收的抗体。其天然环境的污染性成分指将会干扰该抗体的研究、诊断或治疗用途的物质,可包括酶、激素、和其它蛋白质性质或非蛋白质性质的溶质。在有些实施方案中,将抗体纯化至(1)根据例如lowry法的测定,抗体重量超过95%,而在有些实施方案中,重量超过99%,(2)足以通过使用例如转杯式测序仪获得至少15个残基的n-末端或内部氨基酸序列的程度,或(3)根据还原性或非还原性条件下的sds-page及使用例如考马斯蓝或银染色,达到同质。既然抗体天然环境的至少一种成分不会存在,那么分离的抗体包括重组细胞内的原位抗体。然而,分离的抗体通常将通过至少一个纯化步骤来制备。
术语“蛋白a”和“proa”在本文中可互换使用,涵盖自其天然来源回收的蛋白a、合成生成的蛋白a(例如通过肽合成或通过重组技术)、及其保留结合具有ch2/ch3区(诸如fc区)的蛋白质的能力的变体。蛋白a可购自repligen、gehealthcare和fermatech。蛋白a一般固定化在固相支持材料上。术语“蛋白a”还指含有对其共价附着蛋白a的层析固体支持基质的亲和层析树脂或柱。
术语“层析”指根据混合物中的各种溶质在流动相的影响下或在结合和洗脱过程中穿过静止的介质迁移的速率的差异,将混合物中的感兴趣溶质与混合物中的其它溶质分开的过程。
术语“亲和层析”和“蛋白质层析”在本文中可互换使用,指其中感兴趣蛋白质或感兴趣抗体可逆地且特异性地结合至生物特异性配体的蛋白质分离技术。优选的是,所述生物特异性配体共价附着至层析固相材料且在溶液接触层析固相材料时是溶液中的感兴趣蛋白质可及的。在层析步骤期间,所述感兴趣蛋白质(例如抗体、酶、或受体蛋白质)保留其对生物特异性配体(例如抗原、底物、辅因子、或激素)的特异性结合亲和力,而混合物中的其它溶质和/或蛋白质没有可观地或特异性地结合配体。感兴趣蛋白质对固定化配体的结合容许污染性蛋白质或蛋白质杂质流过层析介质而感兴趣蛋白质保持特异性结合至固相材料上的固定化配体。然后,特异性结合的感兴趣蛋白质用低ph、高ph、高盐、竞争性配体、等等以活性形式自固定化配体消除,并随洗脱缓冲液流过层析柱,不含早先容许流过柱的污染性蛋白质或蛋白质杂质。任何成分都能用作用于纯化其相应特异性结合蛋白质(例如抗体)的配体。
术语“非亲和层析”和“非亲和纯化”指其中不利用亲和层析的纯化过程。非亲和层析包括依赖感兴趣分子(诸如蛋白质,例如抗体)与固相基质之间的非特异性相互作用的层析技术。
“阳离子交换树脂”指如下的固相,其带负电荷且因此有游离阳离子供与流过该固相的水溶液中的阳离子交换。附着至固相以形成阳离子交换树脂的、带负电荷的配体可以是例如羧酸根或磺酸根。商品化的阳离子交换树脂包括羧甲基纤维素、在琼脂糖上固定化的磺丙基(sp)(例如gehealthcare的sp-sepharosefastflowtm或sp-sepharosehighperformancetm)和固定化在琼脂糖上的磺酰基(例如gehealthcare的s-sepharosefastflowtm)。“混合模式离子交换树脂”指经阳离子、阴离子、和疏性模块共价修饰的固相。商品化的混合模式离子交换树脂是bakerbondabxtm(j.t.baker,phillipsburg,nj),其含有附着至硅胶固相支持介质的弱阳离子交换基团、低浓度阴离子交换基团、和疏水性配体。
术语“阴离子交换树脂”在本文中用于指带正电荷的固相,例如具有附着于它的一个或多个带正电荷的配体,诸如季氨基团。商品化的阴离子交换树脂包括deae纤维素、qaesephadextm和fastqsepharosetm(gehealthcare)。
“缓冲液”指通过其酸-碱成对成分的作用来抵抗ph变化的溶液。buffers.aguideforthepreparationanduseofbuffersinbiologicalsystems,gueffroy,d.,ed.calbiochemcorporation(1975)中记载了取决于例如期望的缓冲液ph而可采用的多种缓冲液。在一个实施方案中,所述缓冲液具有约2至约9、或者约3至约8、或者约4至约7、或者约5至约7范围内的ph。会将ph控制在此范围内的缓冲液的例示性例子包括mes、mops、mopso、tris、hepes、磷酸盐、乙酸盐、柠檬酸盐、琥珀酸盐、和铵缓冲液,以及这些缓冲液的组合。
“加载缓冲液”指用于将包含感兴趣多肽分子和一种或多种杂质的组合物加载到离子交换树脂上的缓冲液。加载缓冲液具有如下的电导率和/或ph,其使得感兴趣多肽分子(和一般一种或多种杂质)结合至离子交换树脂或使得感兴趣蛋白质流过柱而杂质结合至树脂。
“中间缓冲液”指用于在洗脱感兴趣多肽分子之前自离子交换树脂洗脱一种或多种杂质的缓冲液。中间缓冲液的电导率和/或ph使得一种或多种杂质自离子交换树脂洗脱,但不洗脱显著量的感兴趣多肽。
术语“清洗缓冲液”在本文中使用时指用于在洗脱感兴趣多肽分子之前清洗或再平衡离子交换树脂的缓冲液。方便的是,清洗缓冲液和加载缓冲液可以是相同的,但这不是必须的。
“洗脱缓冲液”用于自固相洗脱感兴趣多肽。洗脱缓冲液的电导率和/或ph使得感兴趣多肽自离子交换树脂洗脱。
“再生缓冲液”可用于再生离子交换树脂,使得它能再使用。再生缓冲液具有自离子交换树脂清除基本上所有杂质和感兴趣多肽所要求的电导率和/或ph。
短语“基本上相似”或“基本上相同”在用于本文时表示两个数值(例如,一个涉及本发明的抗体而另一个涉及参照/比较抗体)之间足够高的相似程度,以致本领域技术人员将认为在用所述数值(例如kd值)所测量的生物学特性背景内两个数值之间的差异具有很小的或没有生物学和/或统计学显著性。作为参照/比较值的函数,所述两个数值之间的差异例如小于约50%,小于约40%,小于约30%,小于约20%,和/或小于约10%。
短语“实质性降低”或“实质性不同”在用于本文时(关于量或数值,而非提到还原的化学过程)表示两个数值(通常一个与某分子有关而另一个与参照/比较分子有关)之间足够高的差异程度,以致本领域技术人员将认为在用所述数值(例如kd值)所测量的生物学特性背景内两个数值之间的差异具有统计学显著性。作为参照/比较分子该数值的函数,所述两个数值之间的差异例如大于约10%,大于约20%,大于约30%,大于约40%,和/或大于约50%。
术语“载体”在用于本文时意指能够运输与其连接的其它核酸的核酸分子。一类载体是“质粒”,指其中可连接另外的dna区段的环状双链dna。另一类载体是噬菌体载体。另一类载体是病毒载体,其中可将另外的dna区段连接到病毒基因组中。某些载体能够在其所导入的宿主细胞中自主复制(例如具有细菌复制起点的细菌载体和附加型哺乳动物载体)。其它载体(例如非附加型哺乳动物载体)可在导入宿主细胞后整合到宿主细胞的基因组中,由此随着宿主基因组一起复制。此外,某些载体能够指导与其可操作连接的基因表达。此类载体在本文中称为“重组表达载体”(或简称为“表达载体”)。通常,在重组dna技术中有用的表达载体常常是质粒形式。在本说明书中,“质粒”和“载体”可互换使用,因为质粒是载体的最常用形式。
关于参考多肽序列的“百分比(%)氨基酸序列同一性”定义为对比序列并在必要时引入缺口以获取最大百分比序列同一性后,且不将任何保守替代视为序列同一性的一部分时,候选序列中与参考多肽序列中的氨基酸残基相同的氨基酸残基的百分率。为测定百分比氨基酸序列同一性目的的对比可以本领域技术范围内的多种方式进行,例如使用公众可得到的计算机软件,诸如blast、blast-2、align或megalign(dnastar)软件。本领域技术人员可决定用于对比序列的适宜参数,包括对所比较序列全长获得最大对比所需的任何算法。然而,为了本发明的目的,%氨基酸序列同一性值是使用序列比较计算机程序align-2获得的。align-2序列比较计算机程序由genentech公司编写,源代码已经连同用户文档一起提交给美国版权局(uscopyrightoffice,washingtond.c.,20559),并以美国版权注册号txu510087注册。公众可自genentech公司(southsanfrancisco,california)得到align-2程序,或者可以自源代码编译。align2程序应当编译成在unix操作系统,优选数码unixv4.0d上使用。所有序列比较参数由align-2程序设定且不变。
在采用align-2来比较氨基酸序列的情况中,给定氨基酸序列a相对于(to)、与(with)、或针对(against)给定氨基酸序列b的%氨基酸序列同一性(或者可表述为具有或包含相对于、与、或针对给定氨基酸序列b的某一%氨基酸序列同一性的给定氨基酸序列a)如下计算:
分数x/y乘100
其中x是由序列对比程序align-2在该程序的a和b对比中评分为相同匹配的氨基酸残基数,且其中y是b中的氨基酸残基总数。可以领会,若氨基酸序列a的长度与氨基酸序列b的长度不相等,则a相对于b的%氨基酸序列同一性将不等于b相对于a的%氨基酸序列同一性。
除非另有具体说明,本文中所使用的所有%氨基酸序列同一性值都是依照上一段所述,使用align-2计算机程序获得的。
“百分比(%)核酸序列同一性”定义为对比序列并在必要时引入缺口以获取最大百分比序列同一性后,候选序列中与参考因子d编码序列中的核苷酸相同的核苷酸的百分率。本领域技术人员可决定用于测量对比的适宜参数,包括对所比较序列全长获得最大对比所需的任何算法。然后相对于较长的序列计算序列同一性,即,即使较短的序列显示与较长的序列一部分的100%序列同一性,整体序列同一性也会小于100%。
“治疗”或“处理”指治疗性处理和预防性或防范性措施二者。需要治疗的受试者包括早就患有病症的受试者以及要预防病症的受试者。“治疗”或“处理”在本文中涵盖缓解疾病和缓解特定疾病的征候和症状。
“病症”指任何会受益于蛋白质治疗的疾患。这包括慢性和急性病症,或包括那些使哺乳动物倾向于所讨论病症的病理状况的疾病。本文中要治疗的病症的非限制性例子包括癌瘤和变态反应。
为了治疗目的,“哺乳动物”指归入哺乳类的任何动物,包括人、非人高等灵长类、其它脊椎动物,家畜和牲畜,及动物园、运动或宠物动物,诸如犬、马、猫、牛等。优选的是,哺乳动物指人。
“干扰rna”或“小干扰rna(sirna)”指长度小于约30个核苷酸且降低靶基因表达的双链rna分子。干扰rna可使用已知方法来鉴定和合成(shiy.,trendsingenetics19(1):9-12(2003);wo2003/056012;wo2003/064621),而且sirna文库可通过商业途径获得,例如dharmacon,lafayette,colorado。通常,sirna能成功设计成靶向基因的5'端。
ii.本发明的组合物和方法
除非另有说明,本发明的实施将采用分子生物学等的常规技术,这些都在本领域的技术范围内。这些技术在文献中有充分解释。参见例如molecularcloning:alaboratorymanual,(j.sambrooketal.,coldspringharborlaboratory,coldspringharbor,n.y.,1989);currentprotocolsinmolecularbiology(f.ausubeletal.,eds.,1987updated);essentialmolecularbiology(t.browned.,irlpress1991);geneexpressiontechnology(goeddeled.,academicpress1991);methodsforcloningandanalysisofeukaryoticgenes(a.bothwelletal.,eds.,bartlettpubl.1990);genetransferandexpression(m.kriegler,stocktonpress1990);recombinantdnamethodologyii(r.wuetal.,eds.,academicpress1995);pcr:apracticalapproach(m.mcphersonetal.,irlpressatoxforduniversitypress1991);oligonucleotidesynthesis(m.gaited.,1984);cellcultureforbiochemists(r.adamsed.,elseviersciencepublishers1990);genetransfervectorsformammaliancells(j.miller&m.caloseds.,1987);mammaliancellbiotechnology(m.butlered.,1991);animalcellculture(j.pollardetal.,eds.,humanapress1990);cultureofanimalcells,2nded.(r.freshneyetal.,eds.,alanr.liss1987);flowcytometryandsorting(m.melamedetal.,eds.,wiley-liss1990);theseriesmethodsinenzymology(academicpress,inc.);wirthm.andhauserh.(1993);immunochemistryinpractice,3rdedition,a.johnstone&r.thorpe,blackwellscience,cambridge,ma,1996;techniquesinimmunocytochemistry,(g.bullock&p.petruszeds.,academicpress1982,1983,1985,1989);handbookofexperimentalimmunology,(d.weir&c.blackwell,eds.);currentprotocolsinimmunology(j.coliganetal.,eds.1991);immunoassay(e.p.diamandis&t.k.christopoulos,eds.,academicpress,inc.,1996);goding(1986)monoclonalantibodies:principlesandpractice(2ded)academicpress,newyork;edharlowanddavidlane,antibodiesalaboratorymanual,coldspringharborlaboratory,coldspringharbor,newyork,1988;antibodyengineering,2ndedition(c.borrebaeck,ed.,oxforduniversitypress,1995);andtheseriesannualreviewofimmunology;theseriesadvancesinimmunology。
1.防止二硫键还原
本发明关注用于防止重组生产期间蛋白质二硫键还原的方法。具体而言,本发明关注用于防止发酵后加工期间重组蛋白二硫键还原的方法。本发明的方法对于含有二硫键的蛋白质的大规模生产(诸如制造规模的)特别有价值。在一个实施方案中,本发明的对于规模超过5,000l的大规模蛋白质生产是有用的。
通过实验发现,在加工含有二硫键的重组蛋白制造期间生成的已收获的细胞培养液(hccf)期间发生二硫键还原。典型的是,在细胞裂解后观察到此还原,尤其是收获操作期间的机械细胞裂解,当它达到某阈值时,诸如例如约30%至约70%、或约40%至约60%、或约50%至约60%的总细胞裂解时。此阈值会有变化,取决于所生成的蛋白质(例如抗体)的本质、重组宿主、生产系统、所使用的生产参数、等等,而且能通过实验容易地确定。
理论上,此类还原可源自制造过程期间的多种因素和条件,而且可源自多种还原剂。本发明(至少部分)基于如下的认识,即此还原的根本原因是hccf中的活性硫氧还蛋白(trx)或硫氧还蛋白样系统。
trx酶系统(由trx、硫氧还蛋白还原酶(trxr)和nadph构成)是蛋白质中二硫键还原的氢供体系统。trx是具有经由硫醇-二硫化物交换催化许多氧化还原反应的cxxc活性位点基序的小单体蛋白质。氧化型trx能被nadph经trxr还原。然后还原型trx能够催化蛋白质中二硫化物的还原。硫氧还蛋白系统所需要的nadph是经由磷酸戊糖途径和糖酵解中的反应提供的。本文中所呈现的结果证明了nadph(其是trx系统的活性所需要的)是由葡萄糖-6-磷酸脱氢酶(g6pd)活性(其通过己糖激酶自葡萄糖和atp生成nadph)提供的(见图4)。在细胞裂解时,这些细胞酶(trx系统、g6pd、和己糖激酶)与它们的底物一起被释放入ccf中,从而容许还原发生。因而,可以通过trx酶系统的或为活性trx系统提供成分的上游酶系统(诸如g6pd和己糖激酶活性)的抑制剂来防止二硫化物还原。
关于这些酶系统的进一步详情或关于蛋白质生产的其它详情参见例如:babson,a.l.andbabson,s.r.(1973)kineticcolorimetricmeasurementofserumlactatedehydrogenaseactivity.clin.chem.19:766-769;michaelw.lairdetal.,“optimizationofblysproductionandpurificationfromeschericiacoli,”proteinexpressionandpurification39:237-246(2005);johnc.jolyetal.,“overexpressionofeschericiacolioxidoreductasesincreasesrecombinantinsulin-likegrowthfactor-iaccumulation,”proc.natl.acad.sci.usa95:2773-2777(march1998);danac.andersenetal.,“productiontechnologiesformonoclonalantibodiesandtheirfragments,”currentopinioninbiotechnology15:456-462(2004);yarivmazoretal.,“isolationofengineered,full-lengthantibodiesfromlibrariesexpressedinescherichiacoli,”naturebiotech.25,563-565(01jun2007);laurac.simmonsetal.,“expressionoffull-lengthimmunoglobulinsinescherichiacoli:rapidandefficientproductionofaglycosylatedantibodies,”journalofimmunologicalmethods263:133-147(2002);paulh.bessetteetal.,“efficientfoldingofproteinswithmultipledisulfidebondsintheescherichiacolicytoplasm,”proc.natl.acad.sci.96(24):13703-08(1999);chaderjian,w.b.,chin,e.t.,harris,r.j.,andetcheverry,t.m.,(2005)“effectofcoppersulfateonperformanceofaserum-freechocellcultureprocessandtheleveloffreethiolintherecombinantantibodyexpressed,”biotechnol.prog.21:550-553;gordong.,mackowm.c.,andlevyh.r.,(1995)“onthemechanismofinteractionofsteroidswithhumanglucose6-phosphatedehydrogenase,”arch.biochem.biophys.318:25-29;gromers.,urigs.,andbeckerk.,(2004)“thetrxsystem-fromsciencetoclinic,”medicinalresearchreviews,24:40-89;hammesg.g.andkochavid.,(1962a)“studiesoftheenzymehexokinase.i.steadystatekineticsatph8,”j.am.chem.soc.84:2069-2073;hammesg.g.andkochavid.,(1962b)“studiesoftheenzymehexokinase.iii.theroleofthemetalion,”j.am.chem.soc.84:2076-2079;johanssonc.,lilligc.h.,andholmgrena.,(2004)“humanmitochondrialglutaredoxinreducess-glutathionylatedproteinswithhighaffinityacceptingelectronsfromeitherglutathioneorthioredoxinreductase,”j.biol.chem.279:7537-7543;legrand,c.,bour,j.m.,jacob,c.,capiaumontj.,martial,a.,marc,a.,wudtke,m.,kretzmer,g.,demangel,c.,duval,d.,andhachej.,(1992)“lactatedehydrogenase(ldh)activityofthenumberofdeadcellsinthemediumofculturedeukaryoticcellsasmarker,”j.biotechnol.,25:231-243;mcdonald,m.r.,(1955)“yeasthexokinase:atp+hexose-->hexose-6-phosphate+adp,”methodsinenzymology,1:269-276,academicpress,ny;sols,a.,delafuente,g.,villar-palasi,c.,andasensio,c.,(1958)“substratespecificityandsomeotherpropertiesofbakers'yeasthexokinase,”biochimbiophysacta30:92-101;kirkpatrickd.l.,kuperusm.,dowdeswellm.,potiern.,donaldl.j.,kunkelm.,berggrenm.,angulom.,andpowisg.,(1998)“mechanismsofinhibitionofthetrxgrowthfactorsystembyantitumor2-imidazolyldisulfides,”biochem.pharmacol.55:987-994;kirkpatrickd.l.,watsons.,kunkelm.,fletchers.,ulhaqs.,andpowisg.,(1999)“parallelsynthesesofdisulfideinhibitorsofthetrxredoxsystemaspotentialantitumoragents,”anticancerdrugdes.14:421-432;milhausen,m.,andlevy,h.r.,(1975)“evidenceforanessentiallysineing6pdfromleuconostocmesenteroides,”eur.j.biochem.50:453-461;pleasants,j.c.,guo,w.,andrabenstein,d.l.,(1989)“acomparativestudyofthekineticsofselenol/diselenideandthiol/disulfideexchangereactions,”j.am.chem.soc.111:6553-6558;whitesides,g.m.,lilburn,j.e.,andszajewski,r.p.,(1977)“ratesofthioldisulfideinterchangereactionsbetweenmono-anddithiolsandellman’sreagent,”j.org.chem.42:332-338;及wipfp.,hopkinst.d.,jungj.k.,rodriguezs.,birminghama.,southwicke.c.,lazoj.s.,andpowisg,(2001)“newinhibitorsofthetrx–trxrsystembasedonanaphthoquinonespiroketalnaturalproductlead,”bioorg.med.chem.lett.11:2637-2641。
依照本发明的一个方面,可通过阻断trx、g6pd和己糖激酶酶系统的任何成分来防止二硫键还原。这些酶系统的抑制剂在本文中统称为“硫氧还蛋白抑制剂”或“trx抑制剂”。典型的是,将trx抑制剂添加至含有重组宿主细胞和培养液的细胞培养液(ccf)和/或通过离心、过滤、或类似分离方法收获后获得的已收获的细胞培养液(hccf)。hccf缺少完整细胞,但通常含有宿主细胞蛋白质和其它污染物,包括dna,它们在后续纯化步骤中清除。如此,可以在收获之前和/或在收获期间、优选在收获之前添加trx抑制剂。
或者/另外,也可以使用其它非特异性方法来防止重组蛋白重组生产期间发酵后的二硫键还原,诸如鼓入空气或调节ph。下文表1中列举了本文中所涵盖的某些还原抑制方法。
表1:还原抑制方法
1在收获之前应用至ccf或在收获之后立即应用于hccf。
2当前尚未商品化。
供本发明方法中使用的“trx抑制剂”包括但不限于(1)trx的直接抑制剂,诸如烃基-2-咪唑基二硫化物和相关化合物(例如1-甲基丙基-2-咪唑基二硫化物)(kirkpatricketal.,1998and1999,supra)和萘醌螺酮缩醇衍生物(例如palmarumycincp1)(wipfetal.,2001,supra);(2)trxr的特异性抑制剂,包括金复合物,诸如金硫葡糖(atg)和金硫苹果酸(atm)(参见例如综述gromeretal.,2004),它们是trxr的不可逆抑制剂的例子;(3)金属离子,诸如hg2+、cu2+、zn2+、co2+、和mn2+,它们能与硫醇和硒醇容易地形成复合物,如此可在本发明的实施方案中用作trxr或trx的抑制剂;(4)g6pd的抑制剂,诸如例如吡哆醛-5’-磷酸和1-氟-2,4-二硝基苯(milhausenandlevy1975,supra),某些类固醇,诸如脱氢表雄酮(dhea)和表雄酮(ea)(gordonetal.,1995,supra);及(4)己糖激酶活性(由此为g6pd生成g6p)的抑制剂,包括金属离子(例如mg2+)的螯合剂(诸如edta)和与sh基团起反应的化合物,山梨糖-1-磷酸、多磷酸、6-脱氧-6-氟葡萄糖、2-c-羟基-甲基葡萄糖、木糖和来苏糖(solsetal.,1958,supra;mcdonald,1955,supra);其它己糖激酶抑制剂披露于美国专利no.5,854,067,题为“hexokinaseinhibitors”。应当理解,这些抑制剂是仅作为例示而列举的。存在其它trx抑制剂,而且能单独地或以各种组合在本发明的方法中使用。
供本发明方法中使用的“trx抑制剂”还包括如下的试剂,其通过在生产活动期间的各个点降低trx系统、磷酸戊糖途径或己糖激酶的酶的水平,从而降低或防止重组生成的抗体或蛋白质的还原。在一些实施方案中,所述酶水平的降低可以通过使用靶向sirna、反义核苷酸、或抗体来实现。为了设计针对cho细胞中找到的基因的靶向sirna或反义核苷酸,这些基因序列能自公开数据库得到,以选择用于靶向不同生物体中的酶的序列。关于大肠杆菌和小鼠trx系统的基因的例子,见下文实施例9。
在使用上文所述抑制剂之外,在本发明的某些实施方案中还可能通过给hccf鼓入空气以维持hccf中的氧化性氧化还原势来防止要纯化的重组蛋白还原。这是一种非直接措施,能通过连续氧化还原型trx和trxr来消减葡萄糖、g6p和nadph。给hccf罐鼓入空气可以例如用约100升每分钟至约200升每分钟、诸如例如150升每分钟的气流来实施。可以实施鼓入空气以达到终点饱和百分比;例如,可以继续鼓入空气直至hccf约100%的空气饱和,或者可以继续鼓入空气直至hccr约30%的空气饱和,或直至介于约100%空气饱和至约30%空气饱和之间。期望的抑制性效果所需要的最小溶氧量(do2)还取决于所生成的抗体或其它重组蛋白。如此,例如,约10%的do2(或连续流约10sccm)会在抗体2h7(变体a)的生产期间具有期望的效果,而apomab可要求更高(约30%)的do2。
在本发明的其它实施方案中,阻断重组蛋白还原有用的另一种非直接方法是降低hccf的ph。此实施方案利用较低ph值时特别慢的硫醇-二硫化物交换(whitesidesetal.,1977,supra;pleasantsetal.,1989,supra)。因此,trx系统在6以下的ph值时的活性显著较低,如此能抑制重组蛋白(诸如ocrelizumab)的还原。
非直接办法还能彼此和/或与一种或多种trx抑制剂的使用组合。
在细胞培养过程完成后,优选对于ccf在收获之前进行或对于hccf在收获后立即进行,通过使用一种或多种trx抑制剂和/或应用非直接办法,能抑制(即部分或完全阻断)二硫键还原。最佳应用时间和模式及有效量取决于要纯化的蛋白质的本质、重组宿主细胞、和所使用的具体生产方法。最佳参数的确定完全在本领域普通技术人员的技术范围内。
例如,在哺乳动物细胞培养过程中,诸如本文实施例中所描述的cho抗体生产过程,如果使用硫酸铜(cuso4,五水合物的形式或无水形式)作为trx抑制剂,那么以约5μm至约100μm、诸如约10μm至约80μm、优选约15μm至约50μm的浓度范围添加它以补充ccf或hccf。由于有些细胞培养物早就含有铜(例如本文实施例中所使用的cho细胞培养物中的约0.04μmcuso4),因此此量是细胞培养物中早就存在的铜(如果有的话)之外的。可使用任何铜(ii)盐代替cuso4,只要溶解度不成问题。例如,乙酸铜和氯化铜(二者都是在水中可溶性的)可用于代替cuso4。最小有效浓度还可能取决于所生成的抗体和使用抑制剂的阶段。如此,例如,当在裂解前添加硫酸铜时,用于抗体2h7(变体a)的最小有效浓度是约30μm,用于apomab的是约75μm,而用于抗体变体c(见表2)的是约50μm。当在cc培养基中添加硫酸铜时,用于抗体2h7(变体a)的最小有效浓度是约15μm,用于apomab的是约25μm,而用于抗体变体c的是约20μm。一种典型的最小cuso4抑制剂浓度是2倍trx浓度(或trx当量)。
edta能在宽浓度范围中使用,取决于细胞裂解程度、所使用的重组宿主细胞、和生产过程的其它参数。例如,当使用cho或其它哺乳动物细胞时,典型的是,能以介于约5mm至约60mm、诸如约10mm至约50mm、或约20mm至约40mm之间的浓度添加edta,取决于细胞裂解程度。对于较低的细胞裂解程度,较低的edta浓度会是足够的,而对于约75%-100%的细胞裂解,所需要的edta浓度较高,诸如例如约20mm至约40mm。最小有效浓度还可能取决于所生成的抗体。如此,例如对于抗体2h7(变体a),最小有效edta浓度是约10mm。
典型的是,dhea在较低的浓度作为trx抑制剂是有效的,诸如例如在约0.05mm至约5mm、优选约0.1mm至约2.5mm的浓度范围内。
其它trx抑制剂,诸如金硫葡糖(atg)和金硫苹果酸(atm),在μm浓度范围内抑制二硫键还原。如此,例如,atg或atm可以以介于约0.1mm至约1mm之间的浓度添加。虽然最小抑制性浓度随实际条件而变化,但是对于atg和atm,典型的是4倍trxr浓度左右。
注意到,所有抑制剂都能以过量使用,因此,并非总是必须知道所述系统中trx或trxr的量。
在一个优选的实施方案中,制造过程中所使用的哺乳动物细胞是中国仓鼠卵巢(cho)细胞(urlaubetal.,proc.natl.acad.sci.usa77:4216(1980))。其它哺乳动物细胞包括但不限于经sv40转化的猴肾cv1系(cos-7,atcccrl1651);人胚肾系(293或为在悬浮培养中生长而亚克隆的293细胞,grahametal.,j.genvirol.36:59(1977));幼仓鼠肾细胞(bhk,atccccl10);小鼠塞托利细胞(tm4,mather,biol.reprod.23:243-251(1980));猴肾细胞(cv1,atccccl70);非洲绿猴肾细胞(vero-76,atcccrl-1587);人宫颈癌细胞(hela,atccccl2);犬肾细胞(mdck,atccccl34);牛鼠(buffalorat)肝细胞(brl3a,atcccrl1442);人肺细胞(w138,atccccl75);人肝细胞(hepg2,hb8065);小鼠乳腺肿瘤(mmt060562,atccccl51);tri细胞(matheretal.,annalsn.y.acad.sci.383:44-68(1982));mrc5细胞;fs4细胞;人肝瘤系(hepg2);和骨髓瘤或淋巴瘤细胞(例如y0、j558l、p3和ns0细胞)(见美国专利no.5,807,715)。
用于在本文中生产多肽的一种优选细胞是cho细胞系dp12(chok1dhfr-)。这是了解最多的cho细胞系之一,广泛用于实验室实践(参见例如1989年3月15日公布的ep0,307,247)。另外,知道其它cho-k1(dhfr-)细胞系,而且能在本发明的方法中使用。
可以在多种培养基中培养用于生成肽、多肽和蛋白质的哺乳动物细胞。商品化的培养基,诸如ham氏f10(sigma)、极限必需培养基((mem),sigma)、rpmi-1640(sigma)、和dulbecco氏改良eagle氏培养基((dmem,sigma),适合于培养细胞。另外,可以使用下列文献中记载的任何培养基作为细胞的培养基:hamandwallace(1979),meth.inenz.58:44;barnesandsato(1980),anal.biochem.102:255;美国专利no.4,767,704;4,657,866;4,927,762;或4,560,655;wo90/03430;wo87/00195;美国专利no.re.30,985;或美国专利no.5,122,469,通过述及将它们所有的公开内容收入本文。任何这些培养基可根据需要补充激素和/或其它生长因子(诸如胰岛素、运铁蛋白、或表皮生长因子)、盐(诸如氯化钠、钙、镁、和磷酸盐)、缓冲剂(诸如hepes)、核苷酸(诸如腺苷和胸苷)、抗生素(诸如gentamycintm药物)、痕量元素(定义为通常以微摩尔范围的终浓度存在的无机化合物)、和葡萄糖或等效能源。任何其它必需补充物也可以以本领域技术人员会知道的适宜浓度包括在内。培养条件(诸如温度、ph等)就是那些先前为表达而选择用于细胞的,这对于普通技术人员而言是显而易见的。
用于在哺乳动物(诸如cho)细胞中生成、回收和纯化重组抗体的方案可包括下列步骤:可以在搅拌罐生物反应器系统中培养细胞,而且采用补料分批培养规程。在优选的补料分批培养中,首先向培养容器提供哺乳动物细胞和培养基,并在培养期间连续地或以离散的增量给培养物补充另外的培养营养物,在培养终止之前有或没有定期的细胞和/或产物收获。补料分批培养可以包括例如半连续补料分批培养,其中定期清除全部培养物(包括细胞和培养基)并用新鲜培养基替换。补料分批培养与单批培养的区别在于在培养过程开始时向培养容器提供细胞培养的所有成分(包括细胞和所有培养营养物)。补料分批培养与灌注培养的区别在于在该过程期间没有自培养容器清除上清液(在灌注培养中,通过例如过滤、封装、锚定至微载体等将细胞限制在培养物中,而且连续地或间歇地向培养容器中引入培养基并自培养容器清除培养基)。
另外,可以依照对所涵盖的特定细胞和特定生产计划合适的任何方案或惯例繁殖培养物的细胞。因此,可采用单步骤或多步骤培养规程。在单步骤培养中,将细胞接种入培养环境中,并在细胞培养的单一生产阶段采用该过程。或者,可使用多阶段培养。在多阶段培养中,可以在许多步骤或阶段培养细胞。例如,可以在第一步或生长阶段培养细胞,其中将细胞(可能是取自贮藏的)接种入对于促进生长和高存活力合适的培养基。在生长阶段通过向细胞培养物添加新鲜培养基而将细胞维持合适的一段时间。
在某些实施方案中,可设计补料分批或连续细胞培养条件来增强哺乳动物细胞在细胞培养生长阶段的生长。在生长阶段,培养细胞的条件和时间使生长最大化。培养条件(诸如温度、ph、溶氧(do2)等等)就是那些用于特定宿主的,而且对于普通技术人员是显而易见的。一般而言,使用酸(例如co2)或碱(例如na2co3或naoh)将ph调节至介于约6.5和7.5之间的水平。用于培养哺乳动物细胞(诸如cho细胞)的合适温度范围介于约30℃至38℃之间,而合适的do2介于5-90%空气饱和之间。
在特定阶段,可以使用细胞来接种细胞培养的生产阶段或步骤。或者,如上所述,生产阶段或步骤可以是与接种或生长阶段或步骤连续的。
细胞培养的生产阶段期间的细胞培养环境通常是受到控制的。如此,如果生产糖蛋白,那么可以操纵影响哺乳动物细胞的细胞比生产力(cellspecificproductivity)的因素,使得在所得糖蛋白中实现期望的唾液酸含量。在一个优选的方面,细胞培养过程的生产阶段前面是细胞培养的过渡阶段,其中涉及用于细胞培养的生产阶段的参数。关于此过程的进一步详情参见美国专利no.5,721,121及chaderjianetal.,biotechnol.prog.21(2):550-3(2005),通过述及明确将其完整公开内容收入本文。
发酵后,纯化蛋白质。用于自细胞碎屑纯化蛋白质的规程首先取决于蛋白质表达的部位。一些蛋白质能自细胞直接分泌入周围培养基中,其它蛋白质是胞内制备的。对于后一类蛋白质,纯化过程的第一步涉及裂解细胞,这可以通过多种方法来进行,包括机械剪切、渗压震扰、或酶处理。此类破坏将细胞的整个内含物释放入匀浆中,而且另外生成由于尺寸小而难以清除的亚细胞碎片。这些一般通过差速离心或通过过滤来清除。由于蛋白质生成运行的过程中细胞的天然死亡和胞内细胞蛋白质和成分的释放,直接分泌的蛋白质存在同样的问题,只是程度较小。
一旦获得含有感兴趣蛋白质的澄清溶液,通常使用不同层析步骤的组合来试图将它与细胞生成的其它蛋白质分开。这些技术基于蛋白质的电荷、疏水性程度、或大小将它们的混合物分开。这些技术每一种可利用数种不同层析树脂,从而容许为所涉及的具体蛋白质精确剪裁纯化方案。这些分离方法每一种的本质是能使蛋白质以不同速率沿长柱向下移动,从而实现随它们沿柱进一步向下移动时增大的物理分离,或者是能使蛋白质选择性粘附至分离介质,然后用不同溶剂差异洗脱。在一些情况中,当杂质特异性粘附至柱而感兴趣蛋白质不粘附至柱时,将期望蛋白质与杂质分开,也就是说,感兴趣的蛋白质存在于“流出液”中。如此,自哺乳动物细胞的细胞培养物纯化重组蛋白可包括一个或多个亲和(例如蛋白a)和/或离子交换层析步骤。
离子交换层析是常用于纯化蛋白质的一种层析技术。在离子交换层析中,溶质表面上带电荷的部分(patch)受到附着于层析基质的相反电荷的吸引,前提是周围缓冲液的离子强度低。一般通过提高缓冲液的离子强度(即电导率)以与溶质竞争离子交换基质带电荷的位点来实现洗脱。改变ph,由此改变溶质的电荷是实现溶质洗脱的另一种方式。电导率或ph的变化可以是逐渐的(梯度洗脱)或逐步的(分步洗脱)。在过去,这些变化是渐进的;即,ph或电导率以单一方向升高或降低。
关于治疗性抗体的工业性纯化的进一步详情参见例如fahrneretal.,biotechnol.genet.eng.rev.18:301-27(2001),通过述及明确将其完整公开内容收入本文。
在哺乳动物细胞之外,可以使用其它真核生物体作为用于表达重组蛋白的细胞。为了在酵母细胞(诸如常用的面包酵母或啤酒糖酵母/酿酒酵母(saccharomycescerevisiae))中表达,合适的载体包括基于2微米质粒的附加体复制载体、整合载体、和酵母人工染色体(yac)载体。其它适合于重组生产异源蛋白质的酵母包括粟酒裂殖糖酵母(schizosaccharomycespombe)(beachandnurse,nature,290:140(1981);ep139,383published2may1985);克鲁维氏酵母属(kluyveromyces)宿主(美国专利no.4,943,529;fleeretal.,bio/technology,2:968975(1991))诸如例如乳酸克鲁维氏酵母(k.lactis)(mw98-8c,cbs683,cbs4574;louvencourtetal.,j.bacteriol.,737(1983))、脆壁克鲁维氏酵母(k.fragilis)(atcc12,424)、保加利亚克鲁维氏酵母(k.bulgaricus)(atcc16,045)、威克克鲁维氏酵母(k.wickeramii)(atcc24,178)、k.waltii(atcc56,500)、果蝇克鲁维氏酵母(k.drosophilarum)(atcc36,906;vandenbergetal.,bio/technology,8:135(1990))、耐热克鲁维氏酵母(k.thermotolerans)、和马克思克鲁维氏酵母(k.marxianus);亚罗酵母属(yarrowia)(ep402,226);巴斯德毕赤氏酵母(pichiapastoris)(ep183,070;sreekrishnaetal.,j.basicmicrobiol.,28:265278(1988));假丝酵母属(candida);瑞氏木霉(trichodermareesia)(ep244,234);粗糙脉孢霉(neurosporacrassa)(caseetal.,proc.natl.acad.sci.usa,76:52595263(1979));许旺酵母属(schwanniomyces),诸如西方许旺酵母(schwanniomycesoccidentalis)(ep394,538published31oct.1990);和丝状真菌,诸如例如脉孢菌属(neurospora)、青霉属(penicillium)、弯颈霉属(tolypocladium)(wo91/00357published10jan.1991)、和曲霉属(aspergillus)宿主诸如构巢曲霉(a.nidulans)(ballanceetal.,biochem.biophys.res.commun.,112:284289(1983);tilburnetal.,gene,26:205221(1983);yeltonetal.,proc.natl.acad.sci.usa,81:14701474(1984))和黑曲霉(a.niger)(kellyandhynes,emboj.,4:475479(1985))。甲基营业性酵母在本文中是合适的,包括但不限于能够在甲醇上生长的酵母,选自下列属:汉逊氏酵母属(hansenula)、假丝酵母属(candida)、克勒克氏酵母属(kloeckera)、毕赤氏酵母属(pichia)、糖酵母属(saccharomyces)、球拟酵母属(torulopsis)、和红酵母属(rhodotorula)。此类酵母的例示性的具体物种的列表可见于c.anthony,thebiochemistryofmethylotrophs,269(1982)。用于所列和其它酵母的表达系统是本领域公知的和/或是商品化的。
为了在昆虫细胞(诸如sf9细胞)中表达,合适的载体包括杆状病毒载体。为了在植物宿主细胞(特别是双子叶植物宿主,诸如烟草)中表达,合适的表达载体包括自根癌土壤杆菌(agrobacteriumtumefaciens)的ti质粒衍生的载体。
本发明的方法还延伸至原核细胞的培养。适合于表达要通过本发明的手段来保护的抗体和其它蛋白质的原核细胞包括古细菌和真细菌,诸如革兰氏阴性或革兰氏阳性生物体。有用的细菌的例子包括埃希氏菌属(escherichia)(例如大肠埃希氏菌/大肠杆菌(e.coli))、芽孢杆菌属(bacilli)(例如枯草芽孢杆菌(b.subtilis))、肠杆菌属(enterobacteria)、假单胞菌属(pseudomonas)物种(例如铜绿假单胞菌(p.aeruginosa))、鼠伤寒沙门氏菌(salmonellatyphimurium)、粘质沙雷氏菌(serratiamarcescans)、克雷伯氏菌属(klebsiella)、变形菌属(proteus)、志贺氏菌属(shigella)、根瘤菌属(rhizobia)、透明颤菌属(vitreoscilla)、或副球菌属(paracoccus)。在一个实施方案中,使用革兰氏阴性细胞。大肠杆菌菌株的实例包括菌株w3110(bachmann,cellularandmolecularbiology,vol.2(washington,d.c.:americansocietyformicrobiology,1987),pp.1190-1219;atcc保藏物no.27,325)及其衍生物,包括具有基因型w3110δfhua(δtona)ptr3laciqlacl8δomptδ(nmpc-fepe)degp41kanr的菌株33d3(美国专利no.5,639,635)。其它菌株及其衍生物,诸如大肠杆菌294(atcc31,446)、大肠杆菌b、大肠杆菌1776(atcc31,537)和大肠杆菌rv308(atcc31,608)也是合适的。这些实例只是例示而非限制。本领域知道用于构建具有指定基因型的任何上述细菌衍生物的方法,参见例如bassetal.,proteins,8:309-314(1990)。通常必需考虑复制子在细菌细胞中的可复制性来选择适宜的细菌。例如,在使用众所周知的质粒诸如pbr322、pbr325、pacyc177、或pkn410来提供复制子时,大肠杆菌、沙雷氏菌属、或沙门氏菌属物种可能适合于用作宿主。通常,宿主细胞应当分泌最小量的蛋白水解酶,而且可能希望在细胞培养中掺入额外的蛋白酶抑制剂。
用于自非哺乳动物细胞生成、回收和纯化重组蛋白的方法也是本领域公知的。如果在非哺乳动物细胞中生成多肽,例如微生物,诸如真菌或大肠杆菌,那么会在细胞内部或在周质空间中回收多肽(kipriyanovandlittle,molecularbiotechnology,12:173201(1999);skerraandpluckthun,science,240:10381040(1988))。因此,必须通过提取,诸如细胞裂解,使蛋白质自细胞释放至胞外培养基。此类破坏将细胞的整个内含物释放入匀浆中,而且另外生成由于尺寸小而难以清除的亚细胞碎片。这些一般通过差速离心或通过过滤来清除。
细胞裂解通常使用机械破碎技术来实现,诸如匀浆化或头研磨(headmilling)。虽然一般有效解放感兴趣蛋白质,但是此类技术具有数项缺点(engler,proteinpurificationprocessengineering,harrisoneds.,3755(1994))。加工期间常常发生的温度升高可导致蛋白质灭活。此外,所得悬浮液含有广谱地污染性蛋白质、核酸、和多糖。核酸和多肽提高溶液粘度,可能伴有通过离心、交叉流过滤、或层析进行的后续加工。这些污染物与感兴趣蛋白质的复杂关联能使纯化过程变复杂,导致不可接受的低产率。自微生物发酵液或匀浆纯化异源蛋白质的改良方法记载于例如美国专利no.7,169,908,通过述及明确将其完整公开内容收入本文。
要强调,本文所述发酵、回收和纯化方法只用于例示目的。本发明的方法可以与为重组蛋白的生成、回收和纯化而开发的任何制造过程组合。
2.抗体
在一个优选的实施方案中,使用本发明的方法来防止抗体(包括治疗性和诊断性抗体)的链间和/或链内二硫键还原。本发明范围内的抗体包括但不限于:抗her2抗体,包括曲妥单抗(trastuzumab)
这些抗体许多在临床实践中广泛用于治疗各种疾病,包括癌症。
在某些具体的实施方案中,本发明的方法用于生产下列抗体和重组蛋白。
抗cd20抗体
利妥昔单抗(rituximab)
关注cd20抗体的专利和专利公开文本包括美国专利no.5,776,456,5,736,137,6,399,061,和5,843,439,以及美国专利申请no.us2002/0197255a1,us2003/0021781a1,us2003/0082172a1,us2003/0095963a1,us2003/0147885a1(andersonetal.);美国专利no.6,455,043b1和wo00/09160(grillo-lopez,a.);wo00/27428(grillo-lopez和white);wo00/27433(grillo-lopez和leonard);wo00/44788(braslawskyetal.);wo01/10462(rastetter,w.);wo01/10461(rastetter和white);wo01/10460(white和grillo-lopez);美国申请no.us2002/0006404和wo02/04021(hanna和hariharan);美国申请no.us2002/0012665a1和wo01/74388(hanna,n.);美国申请no.us2002/0058029a1(hanna,n.);美国申请no.us2003/0103971a1(hariharan和hanna);美国申请no.us2002/0009444a1,和wo01/80884(grillo-lopez,a.);wo01/97858(white,c.);美国申请no.us2002/0128488a1和wo02/34790(reff,m.);wo02/060955(braslawskyetal.);wo2/096948(braslawskyetal.);wo02/079255(reff和davies);美国专利no.6,171,586b1,和wo98/56418(lametal.);wo98/58964(raju,s.);wo99/22764(raju,s.);wo99/51642,美国专利no.6,194,551b1,美国专利no.6,242,195b1,美国专利no.6,528,624b1和美国专利no.6,538,124(idusogieetal.);wo00/42072(presta,l.);wo00/67796(curdetal.);wo01/03734(grillo-lopezetal.);美国申请no.us2002/0004587a1和wo01/77342(miller和presta);美国申请no.us2002/0197256(grewal,i.);美国申请no.us2003/0157108a1(presta,l.);美国专利no.6,090,365b1,6,287,537b1,6,015,542,5,843,398,和5,595,721,(kaminskietal.);美国专利no.5,500,362,5,677,180,5,721,108,和6,120,767(robinsonetal.);美国专利no.6,410,391b1(raubitscheketal.);美国专利no.6,224,866b1和wo00/20864(barbera-guillem,e.);wo01/13945(barbera-guillem,e.);wo00/67795(goldenberg);美国申请no.us2003/01339301a1和wo00/74718(goldenberg和hansen);wo00/76542(golayetal.);wo01/72333(wolin和rosenblatt);美国专利no.6,368,596b1(ghetieetal.);美国申请no.us2002/0041847a1,(goldenberg,d.);美国申请no.us2003/0026801a1(weiner和hartmann);wo02/102312(engleman,e.);美国专利申请no.2003/0068664(albitaretal.);wo03/002607(leung,s.);wo03/049694和us2003/0185796a1(wolinetal.);wo03/061694(sing和siegall);us2003/0219818a1(bohenetal.);us2003/0219433a1和wo03/068821(hansenetal.),通过述及明确将每一篇收入本文。还可参见美国专利no.5,849,898和欧洲专利申请no.330,191(seedetal.);美国专利no.4,861,579和ep332,865a2(meyer和weiss);美国专利no.4,861,579(meyeretal.)和wo95/03770(bhatetal.)。
关注用利妥昔单抗进行的疗法的出版物包括:perottaandabuel"responseofchronicrelapsingitpof10yearsdurationtorituximab"abstract#3360blood10(1)(part1-2):p.88b(1998);stashietal.,"rituximabchimericanti-cd20monoclonalantibodytreatmentforadultswithchronicidopathicthrombocytopenicpurpura"blood98(4):952-957(2001);matthews,r."medicalheretics"newscientist(7apr.,2001);leandroetal.,"clinicaloutcomein22patientswithrheumatoidarthritistreatedwithblymphocytedepletion"annrheumdis61:833-888(2002);leandroetal.,"lymphocytedepletioninrheumatoidarthritis:earlyevidenceforsafety,efficacyanddoseresponse.arthritis&rheumatism44(9):s370(2001);leandroetal.,"anopenstudyofblymphocytedepletioninsystemiclupuserythematosus",arthritis&rheumatism46(1):2673-2677(2002);edwardsandcambridge"sustainedimprovementinrheumatoidarthritisfollowingaprotocoldesignedtodepleteblymphocytes"rheumatology40:205-211(2001);edwardsetal.,"b-lymphocytedepletiontherapyinrheumatoidarthritisandotherautoimmunedisorders"biochem.soc.trans.30(4):824-828(2002);edwardsetal.,"efficacyandsafetyofrituximab,ab-celltargetedchimericmonoclonalantibody:arandomized,placebocontrolledtrialinpatientswithrheumatoidarthritis.arthritis&rheumatism46(9):s197(2002);levineandpestronk"igmantibody-relatedpolyneuropathies:b-celldepletionchemotherapyusingrituximab"neurology52:1701-1704(1999);devitaetal.,"efficacyofselectivebcellblockadeinthetreatmentofrheumatoidarthritis"arthritis&rheumatism46:2029-2033(2002);hidashidaetal.,"treatmentofdmard-refractoryrheumatoidarthritiswithrituximab."presentedattheannualscientificmeetingoftheamericancollegeofrheumatology;october24-29;neworleans,la.2002;tuscano,j."successfultreatmentofinfliximab-refractoryrheumatoidarthritiswithrituximab"presentedattheannualscientificmeetingoftheamericancollegeofrheumatology;october24-29;neworleans,la.2002。sarwaletal.,n.eng.j.med.349(2):125-138(july10,2003)报告了通过dna微阵列序型分析鉴定的急性肾同种异基因移植物排斥中的分子异质性。
在各种实施方案中,本发明提供了包含人源化2h7抗cd20抗体的药物组合物。在具体的实施方案中,所述人源化2h7抗体是表2中所列举的抗体。
表2:人源化抗cd20抗体及其变体
表2的抗体变体a、b和i每一个都包含轻链可变序列(vl):
diqmtqspsslsasvgdrvtitcrasssvsymhwyqqkpgkapkpliyapsnlasgvpsrfsgsgsgtdftltisslqpedfatyycqqwsfnpptfgqgtkveikr(seqidno:1);和
重链可变序列(vh):
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngdtsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysnsywyfdvwgqgtlvtvss(seqidno:2)。
表2的抗体变体c、d、f和g每一个都包含轻链可变序列(vl):
diqmtqspsslsasvgdrvtitcrasssvsylhwyqqkpgkapkpliyapsnlasgvpsrfsgsgsgtdftltisslqpedfatyycqqwafnpptfgqgtkveikr(seqidno:3),和
重链可变序列(vh):
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysasywyfdvwgqgtlvtvss(seqidno:4)。
表2的抗体变体2包含seqidno:3(见上文)的轻链可变序列(vl)和重链可变序列(vh):
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysyrywyfdvwgqgtlvtvss(seqidno:5)。
表2的抗体变体a、b和i每一个都包含全长轻链序列:
diqmtqspsslsasvgdrvtitcrasssvsymhwyqqkpgkapkpliyapsnlasgvpsrfsgsgsgtdftltisslqpedfatyycqqwsfnpptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec(seqidno:6)。
表2的变体a包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngdtsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysnsywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:7)。
表2的变体b包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngdtsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysnsywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnkalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:8)。
表2的变体i包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngdtsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysnsywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnaalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:15)。
表2的抗体变体c、d、f、g和h每一个都包含全长轻链序列:
diqmtqspsslsasvgdrvtitcrasssvsylhwyqqkpgkapkpliyapsnlasgvpsrfsgsgsgtdftltisslqpedfatyycqqwafnpptfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec(seqidno:9)。
表2的变体c包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysasywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnkalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:10)。
表2的变体d包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysasywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykcavsnkalpapieatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:11)。
表2的变体f包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysasywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnaalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:12)。
表2的变体g包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysasywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnaalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhwhytqkslslspgk(seqidno:13)。
表2的变体h包含全长重链序列:
evqlvesggglvqpggslrlscaasgytftsynmhwvrqapgkglewvgaiypgngatsynqkfkgrftisvdkskntlylqmnslraedtavyycarvvyysyrywyfdvwgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynatyrvvsvltvlhqdwlngkeykckvsnaalpapiaatiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:14)。
在某些实施方案中,本发明的人源化2h7抗体进一步包含iggfc中的氨基酸改变,且展现出比具有野生型iggfc的抗体升高至少60倍、至少70倍、至少80倍、更优选至少100倍、优选至少125倍、甚至更优选至少150倍至约170倍的对人fcrn的结合亲和力。
igg中的n-糖基化位点位于ch2域中的asn297。本发明的人源化2h7抗体组合物包括任何前述具有fc区的人源化2h7抗体的组合物,其中组合物中约80-100%(优选约90-99%)的抗体包含缺少附着至糖蛋白fc区的岩藻糖的成熟核心碳水化合物结构。此类组合物在本文中证明展示出乎意料的对fcγriiia(f158)的结合的改善,在与人igg相互作用方面fcγriiia(f158)不如fcγriiia(v158)有效。在正常的、健康的非洲裔美国人和高加索人种中,fcγriiia(f158)比fcγriiia(v158)更常见。参见lehrnbecheretal.,blood94:4220(1999)。历史上,在中国仓鼠卵巢细胞(cho)(最常用的工业宿主之一)中生成的抗体中,群体中约2-6%是未岩藻糖基化的。然而,yb2/0和lec13能生成78-98%未岩藻糖基化的抗体。shinkawaetal.,jbio.chem.278(5),3466-347(2003)报告了在具有较低fut8活性的yb2/0和lec13细胞中生成的抗体在体外显示显著升高的adcc活性。具有降低的岩藻糖含量的抗体的生成也记载于例如lietal.,(glycofi)“optimizationofhumanizediggsinglycoengineeredpichiapastoris”innaturebiologyonlinepublication22jan.2006;niwar.etal.,cancerres.64(6):2127-2133(2004);us2003/0157108(presta);us6,602,684和us2003/0175884(glycartbiotechnology);us2004/0093621,us2004/0110704,us2004/0132140(都是kyowahakkokogyo的)。
双特异性人源化2h7抗体涵盖如下的抗体,其中抗体的一条臂至少具有本发明人源化2h7抗体h和/或l链的抗原结合区,且另一条臂具有针对第二抗原的v区结合特异性。在具体的实施方案中,所述第二抗原选自下组:cd3、cd64、cd32a、cd16、nkg2d或其它nk活化性配体。
抗her2抗体
鼠her2抗体4d5的一种重组人源化型式(humab4d5-8,rhumabher2,曲妥单抗(trastuzumab)或
在一个实施方案中,所述抗her2抗体包含下列vl和vh域序列:
人源化2c4型式574抗体vl(seqidno:16)
diqmtqspsslsasvgdrvtitckasqdvsigvawyqqkpgkapklliysasyrytgvpsrfsgsgsgtdftltisslqpedfatyycqqyyiypytfgqgtkveik,
和人源化2c4型式574抗体vh(seqidno:17)
evqlvesggglvqpggslrlscaasgftftdytmdwvrqapgkglewvadvnpnsggsiynqrfkgrftlsvdrskntlylqmnslraedtavyycarnlgpsfyfdywgqgtlvtvss。
在另一个实施方案中,所述抗her2抗体包含图21和图22分别所示曲妥单抗的vl(seqidno:18)和vh(seqidno:19)域序列。
具有各种特性的其它her2抗体记载于tagliabueetal.,int.j.cancer47:933-937(1991);mckenzieetal.,oncogene4:543-548(1989);maieretal.,cancerres.51:5361-5369(1991);bacusetal.,molecularcarcinogenesis3:350-362(1990);stancovskietal.,pnas(usa)88:8691-8695(1991);bacusetal.,cancerresearch52:2580-2589(1992);xuetal.,int.j.cancer53:401-408(1993);wo94/00136;kasprzyketal.,cancerresearch52:2771-2776(1992);hancocketal.,cancerres.51:4575-4580(1991);shawveretal.,cancerres.54:1367-1373(1994);arteagaetal.,cancerres.54:3758-3765(1994);harwerthetal.,j.biol.chem.267:15160-15167(1992);美国专利no.5,783,186;及klapperetal.,oncogene14:2099-2109(1997)。
抗vegf抗体
抗vegf抗体可以包含例如下列序列:
在一个实施方案中,所述抗vegf抗体包含下列vl序列(seqidno:20):diqmtqttsslsaslgdrviiscsasqdisnylnwyqqkpdgtvkvliyftsslhsgvpsrfsgsgsgtdysltisnlepediatyycqqystvpwtfgggtkleikr;和
下列vh序列(seqidno:21):
eiqlvqsgpelkqpgetvrisckasgytftnygmnwvkqapgkglkwmgwintytgeptyaadfkrrftfsletsastaylqisnlknddtatyfcakyphyygsshwyfdvwgagttvtvss。
在另一个实施方案中,所述抗vegf抗体包含下列vl序列(seqidno:22):
diqmtqspsslsasvgdrvtitcsasqdisnylnwyqqkpgkapkvliyftsslhsgvpsrfsgsgsgtdftltisslqpedfatyycqqystvpwtfgqgtkveikr;和
下列vh序列(seqidno:23):
evqlvesggglvqpggslrlscaasgytftnygmnwvrqapgkglewvgwintytgeptyaadfkrrftfsldtskstaylqmnslraedtavyycakyphyygsshwyfdvwgqgtlvtvss。
在第三个实施方案中,所述抗vegf抗体包含下列vl序列(seqidno:24):
diqltqspsslsasvgdrvtitcsasqdisnylnwyqqkpgkapkvliyftsslhsgvpsrfsgsgsgtdftltisslqpedfatyycqqystvpwtfgqgtkveikr;和
下列vh序列(seqidno:25):
evqlvesggglvqpggslrlscaasgydfthygmnwvrqapgkglewvgwintytgeptyaadfkrrftfsldtskstaylqmnslraedtavyycakypyyygtshwyfdvwgqgtlvtvss。
抗cd11a抗体
人源化抗cd11a抗体efalizumab或
vl(seqidno:26):
diqmtqspsslsasvgdrvtitcrasktiskylawyqqkpgkapklliysgstlqsgvpsrfsgsgsgtdftltisslqpedfatyycqqhneypltfgqgtkveikr;和
vh(seqidno:27):
evqlvesggglvqpggslrlscaasgysftghwmnwvrqapgkglewvgmihpsdsetrynqkfkdrftisvdkskntlylqmnslraedtavyycargiyfygttyfdywgqgtlvtvss。
抗人cd11a抗体可包含seqidno:27的vh和humhm24的全长l链,后者具有序列:
diqmtqspsslsasvgdrvtitcrasktiskylawyqqkpgkapklliysgstlqsgvpsrfsgsgsgtdftltisslqpedfatyycqqhneypltfgqgtkveikrtvaapsvfifppsdeqlksgtasvvcllnnfypreakvqwkvdnalqsgnsqesvteqdskdstyslsstltlskadyekhkvyacevthqglsspvtksfnrgec(seqidno:28),或
上文l链及具有下列序列的h链:
evqlvesggglvqpggslrlscaasgysftghwmnwvrqapgkglewvgmihpsdsetrynqkfkdrftisvdkskntlylqmnslraedtavyycargiyfygttyfdywgqgtlvtvssastkgpsvfplapsskstsggtaalgclvkdyfpepvtvswnsgaltsgvhtfpavlqssglyslssvvtvpssslgtqtyicnvnhkpsntkvdkkvepkscdkthtcppcpapellggpsvflfppkpkdtlmisrtpevtcvvvdvshedpevkfnwyvdgvevhnaktkpreeqynstyrvvsvltvlhqdwlngkeykckvsnkalpapiektiskakgqprepqvytlppsreemtknqvsltclvkgfypsdiavewesngqpennykttppvldsdgsfflyskltvdksrwqqgnvfscsvmhealhnhytqkslslspgk(seqidno:29)。
针对dr5受体的抗体(抗dr5抗体)也可以依照本发明来生成。此类抗dr5抗体具体包括pct公开文本no.wo2006/083971中披露的所有抗体变体,诸如称作apomab1.1、2.1、3.1、4.1、5.1、5.2、5.3、6.1、6.2、6.3、7.1、7.2、7.3、8.1、8.3、9.1、1.2、2.2、3.2、4.2、5.2、6.2、7.2、8.2、9.2、1.3、2.2、3.3、4.3、5.3、6.3、7.3、8.3、9.3和25.3的抗dr5抗体,尤其是apomab8.3和apomab7.3,优选apomab7.3。通过述及明确收录wo2006/083971的完整内容。
3.其它含有二硫化物的蛋白质
在抗体之外,本发明的方法可用于制造包括二硫键的其它多肽。此类多肽的代表性例子包括但不限于下列治疗性蛋白质:组织纤溶酶原激活剂(t-pa),诸如人组织纤溶酶原激活剂(htpa,阿替普酶(alteplase),
含有二硫化物的生物学重要蛋白质的例子包括生长激素,包括人生长激素和牛生长激素;生长激素释放因子;甲状旁腺素;促甲状腺激素;脂蛋白;α-1-抗胰蛋白酶;胰岛素a链;胰岛素b链;胰岛素原;促卵泡激素;降钙素;黄体生成素;高血糖素;凝血因子,诸如因子viiic、因子ix、组织因子、和vonwillebrands因子;抗凝血因子,诸如蛋白c;心房钠尿因子;肺表面活性剂;纤溶酶原激活剂,诸如尿激酶或人尿或组织型纤溶酶原激活剂(t-pa);铃蟾肽;凝血酶;造血生长因子;肿瘤坏死因子-α和-β;脑啡肽酶;rantes(在激活后受到调节,正常情况下由t细胞表达和分泌);人巨噬细胞炎症蛋白(mip-1-α);血清清蛋白,诸如人血清清蛋白;muellerian抑制性物质;松弛素a链;松弛素b链;松弛素原;小鼠促性腺素相关肽;微生物蛋白质,诸如β-内酰胺酶;dna酶;ige;细胞毒性t-淋巴细胞相关抗原(ctla),诸如ctla-4;抑制素;激活素;血管内皮生长因子(vegf);激素的或生长因子的受体;蛋白a或d;类风湿因子;神经营养性因子,诸如骨衍生神经营养性因子(bdnf)、神经营养因子-3、-4、-5、或-6(nt-3、nt-4、nt-5、或nt-6)、或神经生长因子,诸如ngf-β;血小板衍生生长因子(pdgf);成纤维细胞生长因子,诸如afgf和bfgf;表皮生长因子(egf);转化生长因子(tgf),诸如tgf-α和tgf-β,包括tgf-β1、tgf-β2、tgf-β3、tgf-β4、或tgf-β5;胰岛素样生长因子-i和-ii(igf-i和igf-ii);des(1-3)-igf-i(脑igf-i)、胰岛素样生长因子结合蛋白;cd蛋白,诸如cd3、cd4、cd8、cd19、cd20、cd34、和cd40;红细胞生成素;骨诱导因子;免疫毒素;骨形态发生蛋白(bmp);干扰素,诸如干扰素-α、-β、和-γ;集落刺激因子(csf),例如m-csf、gm-csf、和g-csf;白介素(il),例如il-1至il-10;超氧化物歧化酶;t细胞受体;表面膜蛋白;衰变加速因子;病毒抗原,诸如例如aids包膜的一部分;运输蛋白;归巢受体;地址素;调节蛋白;整联蛋白,诸如cd11a、cd11b、cd11c、cd18、icam、vla-4和vcam;肿瘤相关抗原,诸如her2、her3或her4受体;及任何上文所列多肽的片段。
4.用于重组生产抗体的一般方法
本文中的抗体和其它重组蛋白可通过重组dna技术的公知技术来生产。如此,在上文具体鉴定的抗体之外,熟练从业人员能生成针对感兴趣抗原的抗体,例如使用下文所述技术。
抗原选择和制备
本文中的抗体针对感兴趣抗原。优选的是,所述抗原是生物学重要多肽,而且给患有疾病或病症的哺乳动物施用所述抗体能在该哺乳动物中产生治疗性好处。然而,也涵盖针对非多肽抗原(诸如肿瘤相关糖脂抗原;参见美国专利5,091,178)的抗体。若抗原是多肽,则它可以是跨膜分子(例如受体)或配体(诸如生长因子)。例示性的抗原包括下文章节(3)中描述的那些蛋白质。本发明所涵盖的抗体的例示性分子靶物包括cd蛋白,诸如cd3、cd4、cd8、cd19、cd20、cd22、cd34、cd40;erbb受体家族的成员,诸如egf受体、her2、her3或her4受体;细胞粘附分子,诸如lfa-1、mac1、p150、95、vla-4、icam-1、vcam和αv/β3整联蛋白,包括其α或β亚基(例如抗cd11a、抗cd18或抗cd11b抗体);生长因子,诸如vegf;ige;血型抗原;flk2/flt3受体;肥胖(ob)受体;mpl受体;ctla-4;蛋白c,或本文所述任何其它抗原。上文所列抗体所针对的抗原明确包括在本发明范围内。
可溶性抗原或其片段(任选偶联有其它分子的)可作为免疫原用于生成抗体。对于跨膜分子,诸如受体,它们的片段(例如受体的胞外结构域)可用作免疫原。或者,表达跨膜分子的细胞可用作免疫原。此类细胞可衍生自天然来源(例如癌细胞系),或者可以是经重组技术转化而表达跨膜分子的细胞。
对于制备抗体有用的其它抗原及其形式对于本领域技术人员会是显而易见的。
多克隆抗体
多克隆抗体优选在动物中生成,通过多次皮下(sc)或腹膜内(ip)注射相关抗原和佐剂。使用双功能或衍生化试剂,例如马来酰亚氨基苯甲酰基磺基琥珀酰亚胺酯(通过半胱氨酸残基偶联)、n-羟基琥珀酰亚胺(通过赖氨酸残基)、戊二醛、琥珀酸酐、socl2或r1n=c=nr(其中r和r1是不同的烃基),将抗原与在待免疫物种中有免疫原性的蛋白质偶联可能是有用的,例如匙孔虫戚血蓝蛋白、血清白蛋白、牛甲状腺球蛋白或大豆胰蛋白酶抑制剂。
通过将例如100μg或5μg蛋白质或偶联物(分别用于兔或小鼠)与3倍体积的弗氏完全佐剂混和并将该溶液皮内注射于多个部位,将动物针对抗原、免疫原性偶联物或衍生物进行免疫。1个月后,通过多个部位的皮下注射,用弗氏完全佐剂中初始量1/5-1/10的抗原或偶联物对动物进行加强免疫。7-14天后,采集动物的血液并测定血清的抗体滴度。对动物进行加强免疫直到滴度达到平台(plateau)。优选的是,将动物用相同抗原但与不同蛋白质和/或通过不同交联剂偶联得到的偶联物进行加强免疫。偶联物还可在重组细胞培养中作为蛋白质融合物来制备。同样,适当使用凝聚剂诸如明矾来增强免疫应答。
单克隆抗体
单克隆抗体可以使用最初由kohler等,nature,256:495(1975)记载的杂交瘤方法来生成,或者可以通过重组dna方法(美国专利no.4,816,567)来生成。
在杂交瘤方法中,如上所述免疫小鼠或其它适宜的宿主动物,诸如仓鼠或猕猴,以引发生成或能够生成如下抗体的淋巴细胞,所述抗体将特异性结合用于免疫的蛋白质。或者,可以在体外免疫淋巴细胞。然后使用合适的融合剂诸如聚乙二醇将淋巴细胞与骨髓瘤细胞融合,以形成杂交瘤细胞(goding,monoclonalantibodies:principlesandpractice,pp.59-103,academicpress,1986)。
将如此制备的杂交瘤细胞在合适的培养基中接种和培养,所述培养基优选含有抑制未融合的亲本骨髓瘤细胞生长或存活的一种或多种物质。例如,若亲本骨髓瘤细胞缺少次黄嘌呤鸟嘌呤磷酸核糖基转移酶(hgprt或hprt),则用于杂交瘤的培养基典型的将含有次黄嘌呤、氨基喋呤和胸苷(hat培养基),这些物质阻止hgprt缺陷细胞生长。
优选的骨髓瘤细胞是那些高效融合、支持所选择的抗体生产细胞稳定且高水平的生成抗体、且对培养基诸如hat培养基敏感的骨髓瘤细胞。其中,优选的骨髓瘤细胞系是鼠骨髓瘤系,诸如那些自可从索尔克研究所细胞分配中心(salkinstitutecelldistributioncenter,sandiego,californiausa)获得的mopc-21和mpc-11小鼠肿瘤和可从美国典型培养物保藏中心(americantypeculturecollection,rockville,marylandusa)获得的sp-2或x63-ag8-653衍生的。人骨髓瘤和小鼠-人异源骨髓瘤细胞系也有用于生成人单克隆抗体的记载(kozbor,j.immunol.,133:3001(1984);brodeur等,monoclonalantibodyproductiontechniquesandapplications,pp.51-63,marceldekker,inc.,newyork,1987)。
可对杂交瘤细胞正在其中生长的培养基测定针对抗原的单克隆抗体的生成。优选的是,通过免疫沉淀或通过体外结合测定法,诸如放射性免疫测定法(ria)或酶联免疫吸附测定法(elisa),测定由杂交瘤细胞生成的单克隆抗体的结合特异性。
在鉴定出生成具有期望特异性、亲和力和/或活性的抗体的杂交瘤细胞后,所述克隆就可通过有限稀释规程进行亚克隆,并通过标准方法进行培养(goding,monoclonalantibodies:principlesandpractice,pp.59-103,academicpress,1986)。适于这一目的的培养基包括例如d-mem或rpmi-1640培养基。另外,杂交瘤细胞可在动物中作为腹水瘤进行体内培养。
可通过常规免疫球蛋白纯化规程,诸如例如蛋白a-sepharose、羟磷灰石层析、凝胶电泳、透析、或亲和层析,将由亚克隆分泌的单克隆抗体与培养基、腹水或血清适当分开。优选的是,使用本文所述蛋白a层析规程。
编码单克隆抗体的dna易于使用常规规程分离和测序(例如通过使用能够特异性结合编码单克隆抗体重链和轻链的基因的寡核苷酸探针)。杂交瘤细胞是此类dna的优选来源。一旦分离,就可将dna置于表达载体中,然后将该表达载体转染到不另外生成免疫球蛋白蛋白质的宿主细胞中,诸如大肠杆菌细胞、猿猴cos细胞、中国仓鼠卵巢(cho)细胞或骨髓瘤细胞,以在重组宿主细胞中获得单克隆抗体的合成。
也可以修饰dna,例如通过用人重链和轻链恒定域的编码序列替代同源鼠序列(美国专利no.4,816,567;morrison等,proc.natl.acad.sci.usa,81:6851(1984)),或通过将免疫球蛋白编码序列与非免疫球蛋白多肽的整个或部分编码序列共价连接。
典型地,用此类非免疫球蛋白多肽替代抗体的恒定域,或者用它们替代抗体的一个抗原结合位点的可变域,以产生嵌合二价抗体,它包含对一种抗原具有特异性的一个抗原结合位点和对不同抗原具有特异性的另一个抗原结合位点。
在又一个实施方案中,可从使用mccaffertyetal.,nature,348:552-554(1990)所记载的技术构建的抗体噬菌体文库中分离单克隆抗体。clacksonetal.,nature,352:624-628(1991)和marksetal.,j.mol.biol.,222:581-597(1991)分别记载了使用噬菌体文库分离鼠和人抗体。后续出版物记载了通过链改组(marksetal.,bio/technology,10:779-783(1992)),以及组合感染和体内重组作为构建非常大的噬菌体文库的策略(waterhouseetal.,nuc.acids.res.,21:2265-2266(1993)),生成高亲和力(nm范围)的人抗体。如此,这些技术是用于分离单克隆抗体的传统杂交瘤技术的可行替代方法。
人源化和人抗体
人源化抗体具有一个或多个导入其中的来自非人来源的氨基酸残基。这些非人氨基酸残基常常称作“输入”残基,它们通常取自“输入”可变域。人源化可基本上遵循winter及其同事的方法进行(jones等,nature,321:522-525(1986);riechmann等,nature,332:323-327(1988);verhoeyen等,science,239:1534-1536(1988)),即用啮齿类cdr或cdr序列替代人抗体的相应序列。因此,此类“人源化”抗体是嵌合抗体(美国专利4,816,567),其中本质上少于整个人可变域用来自非人物种的相应序列替代。在实践中,人源化抗体通常是其中一些cdr残基和可能的一些fr残基用来自啮齿类抗体中类似位点的残基替代的人抗体。
用于制备人源化抗体的人可变域的选择,包括轻链和重链,对于降低抗原性非常重要。依照所谓的“最适”(best-fit)法,用啮齿类抗体的可变域序列对已知的人可变域序列的整个文库进行筛选。然后选择与啮齿类序列最接近的人序列作为人源化抗体的人fr(sims等,j.immunol.,151:2296(1993))。另一种方法使用由特定轻链或重链亚组的所有人抗体的共有序列衍生的特定框架。同一框架可用于数种不同的人源化抗体(carter等,proc.natl.acad.sci.usa,89:4285(1992);presta等,j.immunol.,151:2623(1993))。
更为重要的是,抗体在人源化后保持对抗原的高亲和力及其它有利的生物学特性。为了实现这一目的,依照一种优选的方法,通过使用亲本和人源化序列的三维模型分析亲本序列和各种概念性人源化产物的方法来制备人源化抗体。三维免疫球蛋白模型通常是可获得的,且为本领域技术人员所熟悉。还可获得图解和显示所选候选免疫球蛋白序列的可能三维构象结构的计算机程序。检查这些显示图像能够分析残基在候选免疫球蛋白序列行使功能中的可能作用,即分析影响候选免疫球蛋白结合其抗原的能力的残基。这样,可从受体和输入序列中选出fr残基并进行组合,从而获得期望抗体特征,诸如对靶抗原的亲和力提高。通常,cdr残基直接且最实质性地涉及对抗原结合的影响。
或者,现在有可能生成在缺乏内源免疫球蛋白生成的情况下能够在免疫时生成人抗体完整全集的转基因动物(例如小鼠)。例如,已经描述了嵌合和种系突变小鼠中抗体重链连接区(jh)基因的纯合删除导致内源抗体生成的完全抑制。在此类种系突变小鼠中转移大量人种系免疫球蛋白基因将导致在抗原攻击时生成人抗体。参见例如jakobovits等,proc.natl.acad.sci.usa,90:2551(1993);jakobovits等,nature,362:255-258(1993);bruggermann等,yearinimmunol.,7:33(1993);及duchosal等,nature,355:258(1992)。还可从噬菌体展示库衍生人抗体(hoogenboom等,j.mol.biol.,227:381(1991);marks等,j.mol.biol.,222:581-597(1991);vaughan等,naturebiotech.,14:309(1996))。
抗体片段
已经开发了用于生成抗体片段的多种技术。传统上,通过蛋白水解消化完整抗体来衍生这些片段(参见例如morimoto等,journalofbiochemicalandbiophysicalmethods24:107-117(1992);及brennan等,science,229:81(1985))。然而,现在可直接由重组宿主细胞生成这些片段。例如,可以从上文讨论的噬菌体抗体库分离抗体片段。或者,可直接从大肠杆菌回收fab'-sh片段并化学偶联以形成f(ab')2片段(carter等,bio/technology10:163-167(1992))。依照另一种方法,可直接从重组宿主细胞培养物分离f(ab')2片段。用于生成抗体片段的其它技术对熟练从业人员将是显而易见的。在其它实施方案中,所选抗体是单链fv片段(scfv)(参见wo93/16185)。
多特异性抗体
多特异性抗体具有对至少两种不同抗原的结合特异性。尽管此类分子通常只结合两种抗原(即双特异性抗体,bsab),但是此表述在用于本文时涵盖具有额外特异性的抗体,诸如三特异性抗体。
用于构建双特异性抗体的方法是本领域已知的。全长双特异性抗体的传统生产基于两对免疫球蛋白重链-轻链的共表达,其中两种链具有不同的特异性(millstein等,nature,305:537-539(1983))。由于免疫球蛋白重链和轻链的随机分配,这些杂交瘤(四源杂交瘤(quadroma))生成10种不同抗体分子的潜在混合物,其中只有一种具有正确的双特异性结构。通常通过亲和层析步骤进行的正确分子的纯化相当麻烦且产物产量低。类似的规程披露于wo93/08829及traunecker等,emboj.,10:3655-3659(1991)。
依照wo96/27011中记载的另一种方法,可改造一对抗体分子间的界面,以将从重组细胞培养物回收的异二聚体的百分比最大化。优选的界面包含抗体恒定域的至少部分ch3结构域。在该方法中,将第一抗体分子界面的一个或多个小氨基酸侧链用较大侧链(例如酪氨酸或色氨酸)替换。通过将大氨基酸侧链用较小氨基酸侧链(例如丙氨酸或苏氨酸)替换,在第二抗体分子的界面上产生与大侧链相同或相似大小的补偿性“空腔”。这提供了较之其它不想要的终产物诸如同二聚体提高异二聚体产量的机制。
双特异性抗体包括交联的或“异源偶联的”抗体。例如,可以将异源偶联物中的一种抗体与亲合素偶联,而将另一种抗体与生物素偶联。此类抗体已经建议用于例如将免疫系统细胞靶向不想要的细胞(美国专利no.4,676,980)和用于治疗hiv感染(wo91/00360,wo92/200373,及ep03089)。异源偶联抗体可以使用任何方便的交联方法来制备。合适的交联剂连同许多交联技术是本领域众所周知的,而且披露于美国专利no.4,676,980。
文献中还记载了由抗体片段生成双特异性抗体的技术。例如,可使用化学连接来制备双特异性抗体。brennan等,science,229:81(1985)记载了通过蛋白水解切割完整抗体以生成f(ab')2片段的规程。将这些片段在存在二硫醇络合剂亚砷酸钠的情况下还原,以稳定邻近的二硫醇并防止分子间二硫键的形成。然后将产生的fab'片段转变为硫代硝基苯甲酸酯(tnb)衍生物。然后将fab'-tnb衍生物之一通过巯基乙胺的还原重新恢复成fab'-硫醇,并与等摩尔量的另一种fab'-tnb衍生物混合,以形成双特异性抗体。产生的双特异性抗体可用作酶的选择性固定化试剂。
最近的进展推动从大肠杆菌直接回收fab'-sh片段,该片段可以化学偶联以形成双特异性抗体。shalaby等,j.exp.med.,175:217-225(1992)记载了完全人源化的双特异性抗体f(ab')2分子的生成。由大肠杆菌分开分泌每种fab'片段,并在体外进行定向化学偶联以形成双特异性抗体。如此形成的双特异性抗体能够结合过表达erbb2受体的细胞和正常人t细胞,以及触发人细胞毒性淋巴细胞针对人乳腺肿瘤靶物的溶解活性。
还记载了从重组细胞培养物直接生成和分离双特异性抗体片段的多种技术。例如,已使用亮氨酸拉链生成双特异性抗体。kostelny等,j.immunol.,148(5):1547-1553(1992)。将来自fos和jun蛋白的亮氨酸拉链肽通过基因融合与两种不同抗体的fab'部分连接。抗体同二聚体在铰链区还原以形成单体,然后重新氧化以形成抗体异二聚体。这种方法也可用于生成抗体同二聚体。hollinger等,proc.natl.acad.sci.usa,90:6444-6448(1993)记载的“双抗体”技术提供了构建双特异性抗体片段的替代机制。该片段包含通过接头相连的重链可变域(vh)和轻链可变域(vl),所述接头太短使得同一条链上的两个结构域之间不能配对。因此,迫使一个片段上的vh和vl结构域与另一个片段上的互补vl和vh结构域配对,由此形成两个抗原结合位点。还报道了通过使用单链fv(sfv)二聚体构建双特异性抗体片段的另一种策略。参见gruber等,j.immunol.,152:5368(1994)。或者,所述抗体可以是zapataetal.,proteineng.8(10):1057-1062(1995)中所记载的“线性抗体”。简言之,这些抗体包含一对串联的fd区段(vh-ch1-vh-ch1),其形成一对抗原结合区。线性抗体可以是双特异性的,或者是单特异性的。
设想了具有超过两个效价的抗体。例如,可制备三特异性抗体。tutt等,j.immunol.,147:60(1991)。
免疫粘附素
最简单且最直接的免疫粘附素设计将粘附素的结合结构域(例如受体的胞外结构域(ecd))与免疫球蛋白重链的铰链和fc区组合在一起。通常,在制备本发明的免疫粘附素时,将编码粘附素结合结构域的核酸融合在编码免疫球蛋白恒定域序列n-末端的核酸的c-末端,然而n-末端融化也是可能的。
通常,在此类融合物中,所编码的嵌合多肽将至少保留有功能活性的免疫球蛋白重链恒定区铰链、ch2和ch3结构域。还可以对恒定域fc部分的c-末端进行融合,或者紧挨着重链ch1或轻链对应区的n-末端进行。进行融合的精确位点不是至关重要的;具体位点是众所周知的,而且可以进行选择以优化免疫粘附素的生物学活性、分泌或结合特征。
在一个优选的实施方案中,将粘附素序列融合在免疫球蛋白g1(igg1)fc域的n-末端。有可能将整个重链恒定区与粘附素序列融合。然而,更优选的是,在融合中使用在铰链区中紧挨着在化学上定义iggfc的木瓜蛋白酶切割位点(即残基216,以重链恒定区的第一个残基为114)或其它免疫球蛋白的类似位点上游开始的序列。在一个特别优选的实施方案中,将粘附素氨基酸序列与igg重链的(a)铰链区、ch2和ch3或(b)ch1、铰链、ch2和ch3结构域融合。
对于双特异性免疫粘附素,将免疫粘附素装配成多聚体,特别是异二聚体或异四聚体。一般而言,这些装配好的免疫球蛋白将具有已知的单元结构。基本的四链结构单元就是igg、igd和ige存在的形式。四链单元在更高分子量的免疫球蛋白中重复;igm一般作为通过二硫键保持在一起的四链基本单元的五聚体存在。iga球蛋白及偶尔的igg球蛋白也可以以多聚体的形式存在于血清中。在多聚体的情况中,各个四链单元可以是相同的或不同的。
下面示意性的列举了在本文范围内的各种例示性的装配好的免疫粘附素:
acl-acl;
ach-(ach,acl-ach,acl-vhch,或vlcl-ach);
acl-ach-(acl-ach,acl-vhch,vlcl-ach,或vlcl-vhch);
acl-vhch-(ach,acl-vhch,或vlcl-ach);
vlcl-ach-(acl-vhch,或vlcl-ach);和
(a-y)n-(vlcl-vhch)2,
其中各个a代表相同或不同的粘附素氨基酸序列;
vl是免疫球蛋白轻链可变域;
vh是免疫球蛋白重链可变域;
cl是免疫球蛋白轻链恒定域;
ch是免疫球蛋白重链恒定域;
n是大于1的整数;
y代表共价交联剂的残基。
为简短起见,上述结构只显示了关键特征;它们没有标示免疫球蛋白的连接(j)或其它结构域,也没有显示二硫键。然而,若结合活性需要此类结构域,则它们应当构建成存在于它们在免疫球蛋白分子中所占据的通常位置。
或者,可以将粘附素序列插入免疫球蛋白重链与轻链序列之间,从而得到包含嵌合重链的免疫球蛋白。在该实施方案中,将粘附素序列融合在免疫球蛋白每个臂中的免疫球蛋白重链的3’端,或是在铰链与ch2结构域之间,或是在ch2与ch3结构域之间。类似的构建物已报道于hoogenboometal.,mol.immunol.28:1027-1037(1991)。
尽管本发明的免疫粘附素中不需要存在免疫球蛋白轻链,然而可以存在免疫球蛋白轻链,或是与粘附素-免疫球蛋白重链融合多肽共价结合,或是直接与粘附素融合。在前一种情况中,通常将编码免疫球蛋白轻链的dna与编码粘附素-免疫球蛋白重链融合蛋白的dna共表达。在分泌后,杂合重链与轻链将共价结合以提供包含两对二硫键相连的免疫球蛋白重链-轻链的免疫球蛋白样结构。适于制备此类结构的方法披露于例如1989年3月28日公告的美国专利no.4,816,567。
最方便的是通过将编码粘附素部分的cdna序列与免疫球蛋白cdna序列以符合读码框的形式融合来构建免疫粘附素。然而,也可以使用与基因组免疫球蛋白片段的融合(参见例如aruffoetal.,cell61:1303-1313(1990);stamenkovicetal.,cell66:1133-1144(1991))。后一种类型的融合要求存在ig调控序列以供表达。编码igg重链恒定区的cdna可以根据已发表的序列自衍生自脾或外周血淋巴细胞的cdna文库分离,通过杂交或者通过聚合酶链式反应(pcr)技术。将编码免疫粘附素的“粘附素”和免疫球蛋白部分的cdna串联插入在所选宿主细胞中指导高效表达的质粒载体。
提供下文实施例仅仅为了例示目的,而非意图以任何方式限制本发明的范围。
在此完整收入本说明书中引用的所有专利和文献作为参考。
实施例
除非另有说明,实施例中提及的商品化试剂依照制造商的说明书使用。下文实施例和整篇说明书中以atcc编号所鉴别的那些细胞的来源是美国典型培养物保藏中心(americantypeculturecollection,manassas,virginia)。实施例1:对材料和方法的描述
实施例2-8中使用了下列材料和方法。
材料
实验实施例中所描述的实验中所使用的材料和装置包括:不锈钢管形瓶(微型罐,flowcomponents,dublin,ca;矮的(50cc)和高的(55cc));透析管(spectra/por,6-8000mwco,产品目录#132645);0.22μm滤器(milliporemillipakgammagold,产品目录#mpgl04gh2);磷酸盐缓冲盐水(pbs,emd,产品目录#6506);乙二胺四乙酸(edta,sigma,产品目录#e4884);α-烟酰胺腺嘌呤二核苷酸磷酸(nadph,calbiochem,产品目录#481973);脱氢表雄酮(dhea,tci,产品目录#d0044);硫酸铜(sigma,产品目录#c8027);葡萄糖-6-磷酸(g6p,calbiochem,产品目录#346764);金硫葡糖(atg,usp,产品目录#1045508);金硫苹果酸(atm,alfaaesar,产品目录#39740);还原型谷胱甘肽(gsh,j.t.baker,产品目录#m770-01);monobromobimane(mbb,fluka,产品目录#69898);组氨酸(j.t.baker,产品目录#2080-05);硫酸钠(j.t.baker,产品目录#3897-05);trx(sigma,产品目录#t8690);trxr(sigma,产品目录#t9698)。所有化学药品和试剂以收到的形式使用,没有进一步纯化。配制edta(250mm,ph7.5)、cuso4(10mm)、atg(30mm)、atm(30mm)、nadph(75mm)、g6p(300mm)的储备溶液,供微型罐时程研究中使用。
细胞培养液(ccf)的生成
为了生成供各种抗原研究用的ocrelizumabccf,利用与先前所述方法(chaderjianetal.,2005)类似的代表性小规模发酵过程。简言之,使用配有斜片叶轮的3升玻璃搅拌罐
已收获的细胞培养液(hccf)制备
使用microfluidicshc-8000匀浆器通过高压匀浆化来实现ccf的完全裂解。将该仪器的压力调节器设置成4,000-8,000psi,并使ccf穿过匀浆器以在单次通过后获得完全细胞裂解(膜破裂)。一旦穿过系统清除了水,就收集ccf匀浆。将匀浆转移至离心瓶,并在sorvalrc-3b转子离心机中于20℃以4,500rpm离心30分钟。倒出离心液,然后深部过滤,接着使用蠕动泵及硅管0.22μm无菌过滤,用以自匀浆化的ccf(100%细胞裂解)生成最终的hccf。或者,将来自发酵罐的ccf直接离心,没有任何匀浆化,然后用无菌0.22μm滤器过滤离心液以生成hccf。
微型罐操作
在操作所有微型罐时使用层流通风橱,而且hccf温育实验中使用的所有材料或是高压灭菌或是用70%异丙醇漂洗以使细菌污染最小化。
乳酸脱氢酶测定法
关于乳酸脱氢酶测定法,参见babson&babson(1973)及legrandetal.,(1992),通过述及将其收入本文。
透析实验
实施透析实验以确定引起ocrelizumab还原的成分是小分子还是大分子(即酶)。将纯化和配制好的ocrelizumab(30.2mg/ml)的3ml样品针对1l磷酸盐缓冲盐水(pbs,10mm,ph7.2)透析24小时,并在8小时后更换pbs。然后使用280nm吸光度将ocrelizumab样品的浓度调节至1mg/ml。使用前将等分试样保存于-70℃。将透析管在0.05%叠氮化物溶液中水合过夜,并在使用前用无菌水漂洗。融化将来自3l发酵罐的ccf匀浆化获得的hccf,并使用蠕动泵经由0.22μmmillipak滤器过滤。给六个矮微型罐各装30mlhccf。给每个微型罐添加密封透析管中的500μlocrelizumab样品。密封微型罐,并加载入以35rpm于环境温度运转的台式混合仪(barnsteadlab-linemaxq4000)中。对于每个时间点,自混合仪中取出一个微型罐,并采集hccf(在微型罐中)和ocrelizumab样品(在透析袋中)的等分试样,保存于-70℃,直至用游离硫醇测定法和bioanalyzer测定法(下文所述)分析。
对抑制剂测试小规模体外系统中的还原
给高微型罐装27mlhccf。根据实验设计,添加各种试剂(nadph、g6p、g6pd抑制剂或trxr)至期望浓度,并用pbs(10mm,ph7.2)使微型罐中的终体积达到30ml。密封微型罐,并加载入以35rpm于环境温度运转的台式混合仪。在采样的每个时间点,将微型罐的外部用70%ipa灭菌,并在层流通风橱中打开以取出等分试样。然后将微型罐重新密封并再次加载入台式混合仪中。将所有等分试样保存于-70℃,直至用游离硫醇测定法和bioanalyzer测定法(下文所述)分析。
体外trx/trx还原酶研究
将商品化trxr(大鼠肝)溶液(4μm)用水稀释以产生2.86μm溶液。将冻干的trx(人)用pbs(10mm,ph7.2)重建,产生500μm溶液。在水中配制20mmnadph溶液及10mmatg和atm溶液。
在黑色聚丙烯1.5ml微量离心管中,温和混合437μlpbs、25μlnadph、16μl配制好的ocrelizumab溶液(30.2mg/ml)和5μltrx。通过添加17.5μltrxr来启动反应。将反应于室温温育24小时。在每个采样时间点取出20μl等分试样,并保存于-70℃,直至通过bioanalyzer测定法(见下文)分析。实施对照以确定当通过用等体积的pbs替代反应混合物中的trx和/或trxr来省略酶时酶途径是否有活性。
使用上文所述相同反应条件及添加5μlatg或atm证明了对trx系统的抑制。为了证明cu2+对trx系统的抑制,向反应混合物添加2.5μlcuso4(10mm),使用相同的酶但不同的缓冲液(10mm组氨酸、10mmna2so4、137mmnacl、2.5mmkcl,ph7.0)以防止不溶性cu3(po4)2形成。
游离硫醇测定法
在pbs(10mm,ph6.0±0.05)中生成了gsh的标准曲线。自110mmgsh溶液,经由连续稀释配制了0、5.5、11、22、44、55、110和550μm浓度的标准品。自mbb的乙腈储备溶液(10mm,保存于-20℃),在pbs(10mm,ph10.0±0.05)中配制了mbb的100μm溶液,并避光保存。
在黑色平底96孔板中,向每个孔中分配100μlmbb。对于标准曲线,添加10μl标准gsh溶液,产生8.0±0.2的工作ph。对于样品,向孔中添加10μl样品。所有孔都以一式三份准备。将板在黑暗中于室温温育1小时,然后使用荧光读板仪(moleculardevices
bioanalyzer测定法
使用agilent2100bioanalyzer获得了毛细管电泳测量。样品制备如agilentprotein230测定法方案(手册部分号g2938-90052)中所述进行,有轻微变化。制备前,用水将hccf样品1∶4稀释并将蛋白a样品稀释至1.0g/l。对于变性步骤的hccf样品,在所提供的2μl变性溶液之外还添加24μl50mm碘乙酰胺(iam)、0.5%sds溶液。对于蛋白a样品,使用不含iam的0.5%sds和2μl变性溶液。使用agilent2100expert软件生成了数字凝胶样图像。
用于hccf保持时间研究的储备溶液
在实验室规模hccf保持时间研究中使用了三种不同的储备溶液:(1)使用edta二钠二水合物(mallinckrodt,产品目录#7727-06或sigma,产品目录#e-5134)和edta四钠二水合物(sigma,产品目录#e-6511)配制的250mmedta储备溶液(ph7.4);(2)硫酸铜五水合物(cuso4,sigma,产品目录#c-8027)的50mm储备溶液;和(3)1m乙酸溶液(mallinckrodt,产品目录#v193)。
抑制剂添加和细胞培养液(ccf)掺合
匀浆化前将250mmedta或50mmcuso4的储备溶液添加至ccf以评估防止抗体二硫化物还原的终浓度范围。一旦自匀浆化的ccf生成最终的hccf,就将这些溶液与自未匀浆化的ccf生成的hccf(也含有edta或cuso4)混合以稀释和降低细胞裂解总水平至100%最大值以下。或者,将1m乙酸的储备溶液添加至最终的掺合hccf溶液(匀浆化的ccf和未匀浆化的ccf)以降低溶液的ph以防止抗体二硫化物还原。
在50ml316l不锈钢管形瓶中装大约30-50ml每种hccf溶液(含有edta、cuso4、乙酸、或不添加而作为对照)。用夹子将管形瓶密封,不给溶液充气或不搅动。将管形瓶保存于室温(18-22℃)。在预定的时间点,取出溶液并在实验室规模蛋白a亲和树脂上纯化。
用其它氧化剂(诸如例如胱氨酸和氧化型谷胱甘肽)能获得相似的结果。
鼓入空气
为了评估给自匀浆化ccf生成的hccf鼓入空气以防止抗体二硫化物还原,利用3l玻璃或15l不锈钢容器。将大约1-5lhccf通过0.22μm无菌过滤入每个经过灭菌的容器中。实验条件维持于18-22℃和50(15l发酵罐)或275rpm(3l发酵罐)搅动,有或无通过添加二氧化碳进行的ph控制。或是给溶液鼓入空气以提高溶氧水平至空气饱和,或是鼓入氮气(对照)以清除溶液中的任何溶氧。给每个容器的气流是可变的,取决于是使用恒定充气速率还是维持最小溶氧水平。在预定的时间点,自两种容器取出25-50ml样品,并在实验室规模蛋白a亲和树脂上纯化,之后分析。
蛋白a加工
可以使用特异性亲和层析树脂来捕捉和纯化已收获的细胞培养液样品中的抗体。选择蛋白a树脂(millipore,prosep-vahighcapacity)作为用于抗体纯化的亲和树脂。将树脂填充到床高14cm、内径0.66cm(最终柱体积4.8ml)的玻璃柱
将树脂暴露于介于350-560cm/hr之间的线性流速的缓冲液和hccf。用25mmtris、25mmnacl、5mmedta,ph7.1平衡树脂。对于每次纯化,给树脂加载5-15mg抗体每ml树脂。使用固定化蛋白ahplc柱(appliedbiosystems,porosa)测定hccf中的抗体浓度。加载后,用25mmtris、25mmnacl、5mmedta、0.5mtmac,ph7.1清洗树脂,然后使用0.1m乙酸,ph2.9洗脱抗体。洗脱合并基于柱后联机测量的280nmuv吸光度。使用1mhepes钠将纯化后的洗脱合并液调节ph至ph5.0-5.5。用0.1m磷酸再生树脂后,使用相同或相似填充的树脂进行后续对其它hccf溶液的纯化。
使用280nmuv光谱测定术测量纯化后的蛋白a合并液的抗体浓度。通过bioanalyzer测定法分析纯化后的蛋白a洗脱合并液,用以对150kda分子量的完整抗体的百分比定量。
实施例2:透析实验
设计并实施透析实验来确定ocrelizumab的还原是由还原性小分子还是大分子(例如酶)引起的。在此透析实验中,将纯化后的完整ocrelizumab放置在分子量截留(mwco)7000的透析袋中,并将透析袋在不锈钢微型罐中含ocrelizumab的hccf中温育。如图1和2所示,袋内的ocrelizumab在温育期后没有被还原(图1),而hccf中袋外的ocrelizumab在温育开始后不久就被显著还原了。这是通过完整ocrelizumab(约150kda)的损失和ocrelizumab片段(各种重链和轻链组合)的形成证明的(图2)。对来自被还原制造运行的蛋白a洗脱合并液中ocrelizumab的质谱术分析指明那些观察到的片段是通过只还原链间二硫键而形成的。
游离硫醇测量显示在温育开始时透析袋内不存在游离硫醇;然而,温育开始后不到5小时,透析袋内外的游离硫醇水平变得可比,指明hccf中的小分子成分在透析袋内外是完全平衡的(图3)。由于只在mwco7000da的透析袋外观察到还原,但在透析袋内没有观察到还原,因此还原性分子的分子量必然大于7000da。如此,酶促反应导致ocrelizumab的还原。
实施例3:在体外trx/trxr对ocrelizumab(rhumab2h7,变体a)的还原
通过与trx、trxr、和nadph一起温育完整ocrelizumab,对trx系统测试它在体外还原ocrelizumab的能力。bioanalyzer结果指明在体外ocrelizumab被trx系统还原(图5)。此体外系统中的还原速率表现为比hccf慢(例如与图2所示还原比较时)。这可能是由于体外反应中所使用的酶(trx和trx-r)的较低浓度和/或缓冲系统,因为trx系统的反应速率取决于酶浓度和缓冲系统。
实施例4:trx系统的抑制剂
(i)硫酸铜对重组抗体还原的抑制
已知硫酸铜有能力提供氧化还原势,而且已经在细胞培养过程中用于使重组抗体分子中的游离硫醇水平最小化(即使未配对半胱氨酸最小化)(chaderjianetal.,2005,supra)。对硫酸铜测试在体外抑制trx系统和后续ocrelizumab还原的功效。在此体外还原实验中,将缓冲系统由pbs变成硫酸组氨酸以避免不溶性cu3(po4)2形成。图8显示了ocrelizumab易于被硫酸组氨酸缓冲液中的trx系统还原(甚至比pbs缓冲液中的快)。向此反应添加cuso4明显抑制ocrelizumab还原(图9)。
(ii)atg和atm对hccf中重组抗体还原的抑制
对trxr的两种商品化特异性抑制剂,即金硫葡糖(atg)和金硫苹果酸(atm),测试它们在体外抑制trx系统和ocrelizumab还原的能力。atg和atm都能在上文所述测定法中有效抑制ocrelizumab还原(见图6和7)。以1mm浓度向与图5说明所述相同的反应混合物添加金硫葡糖或金硫苹果酸有效抑制ocrelizumab还原,如来自bioanalyzer分析的数字凝胶样图像所示。
如果trx系统在hccf中有活性并还原ocrelizumab,如导致被还原的抗体分子的制造运行中或实验室规模实验中观察到的,那么这两种金化合物(atg和atm)都应能够抑制hccf中的ocrelizumab还原。图10显示了ocrelizumab在来自匀浆化cct(其是自3l发酵罐生成的)的hccf中在温育一段时间后易于还原。然而,当向hccf添加1mmatg或atm时,ocrelizumab还原事件受到完全抑制(图11和12)。这些结果证明了trx系统在hccf中有活性且直接造成ocrelizumab还原。
实施例5:用于trx系统活性的nadph的来源及g6p和葡萄糖在还原机制中的作用
trx系统对二硫化物的还原需要来自nadph的还原性当量(图4)。为所有还原性生物合成反应提供nadph的主要细胞代谢途径是磷酸戊糖途径。为了使抗体还原事件发生,此途径中的酶必须在hccf中仍有活性以保持trx系统有活性。最低限度,磷酸戊糖途径中的第一步(由g6pd催化)必须有活性以还原nadp+成nadph同时转变g6p成6-磷酸葡萄糖酸内酯。另外,g6p最有可能是hccf中由己糖激酶活性自葡萄糖和腺苷-5’-三磷酸(atp)生成的。图4总结了ocrelizumab还原的整个机制。
hccf中的还原活性表现出在一些情况中是瞬时的,而且可以在某些贮藏条件下或在多轮冻融后随时间受到抑制。完全丧失还原活性的hccf提供了机会来探索nadph和g6p在trx系统还原ocrelizumab中的作用。
将来自大规模制造运行(“β”运行)的hccf进行数轮冻融并在设计用于测量还原的实验中使用;没有观察到ocrelizumab还原(图13),尽管它有能力引起抗体还原,像先前在新鲜融化的来自此同一发酵的hccf中看到的。以5mm的浓度向此非还原性hccf添加nadph,则还原事件恢复了(图14)。此外,trx系统在不再发生还原且在提供辅因子的情况中能够还原蛋白质和/或抗体的hccf中仍是完整的且有活性的。另外,还原活性随时间丢失,因为nadph源被消减(大概是由于所有竞争nadph的还原性反应对nadph的氧化),而不是因为trx系统被降解或灭活。
这得到了另一项实验的证实。向经过反复冻融的来自β运行的hccf添加10mmg6p。此g6p添加再活化trx系统,其随后在hccf温育实验中还原ocrelizumab(图15)。这证明了hccf中ocrelizumab的还原是由trx系统和g6pd二者的活性引起的。另外,g6pd在经过反复冻融的β运行hccf中仍有活性;此经过反复冻融的hccfβ运行中还原活性的损失表现为是由于g6p的消减,如此消除nadp+向nadph的转变。
在我们的研究中,我们观察到edta在hccf温育实验中能有效抑制ocrelizumab还原。如图16所示,将来自12,000l规模ocrelizumab制造运行(没有反复冻融且没有丢失还原活性)的hccf于环境温度温育超过19小时后,ocrelizumab被还原。然而,当向12klhccf添加20mmedta并在分开的不锈钢微型罐中保持时,还原被完全抑制(图17)。在糖酵解的第一步中,己糖激酶催化磷酸基团从mg2+-atp转移至葡萄糖,即需要mg2+与atp络合的反应(hammes&kochavi,1962a&1962b,supra)。由于edta是金属离子螯合剂,尤其对于mg2+而言,因此它会是己糖激酶的有效抑制剂。过量的edta能有效阻断还原的观察结果指明ocrelizumab还原的机制中涉及己糖激酶(即提供g6p)。不限于此或任何其它理论,edta通过消除己糖激酶活性并由此降低g6pd可利用的g6p水平及随后trx系统可利用的nadph水平来阻断ocrelizumab还原。
虽然edta每一次都有效阻断新鲜hccf中的ocrelizumab还原,但是它没有能力防止丧失trx系统活性、然后通过添加g6p而再活化的β运行hccf中的ocerlizumab还原。例如,在向完全丧失还原活性的β运行hccf添加5mmg6p和20mmedta(终浓度)的hccf温育实验中观察到ocrelizumab还原(图18)。然而,在不添加g6p和edta的对照温育实验中没有看到还原。不限于此或任何其它理论,以此方式使用的edta因此可能不抑制trx系统或g6pd,而且可能发挥为g6pd生成g6p的己糖激酶的抑制剂的功能。若没有g6p,trx系统不会得到必要的nadph供应来实现活性。
实施例6:dhea对重组抗体还原的抑制
脱氢表雄酮(dhea)像其它类似的g6pd抑制剂一样有效阻断g6pd活性(gordonetal.,1995,supra)。g6pd抑制剂还通过阻断nadph生成来防止hccf中抗体(例如ocrelizumab)的还原。在hccf温育实验中证明了dhea抑制orcelizumab还原的能力。向hccf添加dhea防止抗体还原。
dhea通常以约0.05mm至约5mm的浓度范围使用。dhea通常也以约0.1mm至约2.5mm的浓度范围使用。
实施例7:添加(i)edta、(ii)硫酸铜、和(iii)乙酸对重组抗体还原的抑制
在不锈钢管形瓶中保存和保持四种不同hccf。各溶液中细胞裂解的量是相似的,它们是通过用来自未匀浆化ccf的hccf稀释来自匀浆化ccf的hccf而生成的。例如,将150ml第一裂解溶液与50ml第二溶液混合。在此研究中评估的四种hccf混合物含有以下任一项:(1)20mmedta、(2)30μmcuso4、(3)15mm乙酸(ph5.5)、和(4)不添加化学抑制剂而作为对照溶液。使用蛋白a层析立即(t=0hr)自所有四种混合物纯化ocrelizumab抗体,然后再次在不锈钢管形瓶中保存20hr和40hr。通过bioanalyzer测定法分析纯化后的蛋白a洗脱合并液,用以对完整抗体(150kda)的百分比定量。结果显示了在初始时间点所有四种混合物中存在超过90%的完整抗体(图19)。然而,在20hr时间点,在对照混合物(不进行任何添加)中没有检测到完整抗体,指明抗体二硫键的还原。在其它三种混合物中,在20hr和40hr时间点都仍检测到超过90%的完整抗体,证明了测试的所有三种抑制剂都防止二硫键还原。
实施例8:给hccf鼓入空气对重组抗体还原的抑制
在两个分开的10l不锈钢发酵罐中储存和保持自匀浆化ccf生成的一份hccf混合物。给一个容器鼓入空气,而给另一个容器鼓入氮气。使用蛋白a层析立即(t=0hr)自初始混合物纯化ocrelizumab抗体。在选定的时间点,自每个容器取出50ml样品,并使用蛋白a层析纯化抗体。然后通过bioanalyzer测定法分析纯化后的蛋白a洗脱合并液,用以对150kda完整抗体的百分比定量。结果显示了初始溶液中存在大约85%的完整抗体(图20),指明暴露于氧(即发酵罐中鼓入的空气)之前抗体二硫键的一些早期还原。一旦给混合物鼓入空气2小时,对36小时研究的剩余物测量到超过90%的完整抗体。相反,当给混合物鼓入氮气时,继续发生抗体还原事件,像2小时(28%150kda峰)和6小时(5%150kda峰)测量到的。这些结果证明了当自匀浆化ccf生成的hccf混合物暴露于氧时防止抗体中的二硫键还原。
实施例9:靶向sirna或反义核苷酸trx抑制剂的设计
针对cho细胞中找到的基因的靶向sirnas或反义核苷酸的设计可使用公众可得到的序列进行,诸如大肠杆菌硫氧还蛋白trxa(seqidno:30)、大肠杆菌硫氧还蛋白还原酶trxb(seqidno:31);小鼠硫氧还蛋白1(seqidno:32)、小鼠硫氧还蛋白2(seqidno:33)、小鼠硫氧还蛋白还原酶1(seqidno:34)、和小鼠硫氧还蛋白还原酶2(seqidno:35)的序列。本领域普通技术人员能使用这些序列来选择用于设计靶向不同生物体和/或细胞(诸如cho细胞)中的酶的trx抑制剂的序列。
大肠杆菌硫氧还蛋白trxa的序列是:
atgttacaccaacaacgaaaccaacacgccaggcttattcctgtggagttatatatgagcgataaaattattcacctgactgacgacagttttgacacggatgtactcaaagcggacggggcgatcctcgtcgatttctgggcagagtggtgcggtccgtgcaaaatgatcgccccgattctggatgaaatcgctgacgaatatcagggcaaactgaccgttgcaaaactgaacatcgatcaaaaccctggcactgcgccgaaatatggcatccgtggtatcccgactctgctgctgttcaaaaacggtgaagtggcggcaaccaaagtgggtgcactgtctaaaggtcagttgaaagagttcctcgacgctaacctggcgtaa(seqidno:30)。
大肠杆菌硫氧还蛋白trxb的序列是:
atgggcacgaccaaacacagtaaactgcttatcctgggttcaggcccggcgggatacaccgctgctgtctacgcggcgcgcgccaacctgcaacctgtgctgattaccggcatggaaaaaggcggccaactgaccaccaccacggaagtggaaaactggcctggcgatccaaacgatctgaccggtccgttattaatggagcgcatgcacgaacatgccaccaagtttgaaactgagatcatttttgatcatatcaacaaggtggatctgcaaaaccgtccgttccgtctgaatggcgataacggcgaatacacttgcgacgcgctgattattgccaccggagcttctgcacgctatctcggcctgccctctgaagaagcctttaaaggccgtggggtttctgcttgtgcaacctgcgacggtttcttctatcgcaaccagaaagttgcggtcatcggcggcggcaataccgcggttgaagaggcgttgtatctgtctaacatcgcttcggaagtgcatctgattcaccgccgtgacggtttccgcgcggaaaaaatcctcattaagcgcctgatggataaagtggagaacggcaacatcattctgcacaccaaccgtacgctggaagaagtgaccggcgatcaaatgggtgtcactggcgttcgtctgcgcgatacgcaaaacagcgataacatcgagtcactcgacgttgccggtctgtttgttgctatcggtcacagcccgaatactgcgattttcgaagggcagctggaactggaaaacggctacatcaaagtacagtcgggtattcatggtaatgccacccagaccagcattcctggcgtctttgccgcaggcgacgtgatggatcacatttatcgccaggccattacttcggccggtacaggctgcatggcagcacttgatgcggaacgctacctcgatggtttagctgacgcaaaataa(seqidno:31)。
小鼠硫氧还蛋白1的序列是:
atggtgaagctgatcgagagcaaggaagcttttcaggaggccctggccgccgcgggagacaagcttgtcgtggtggacttctctgctacgtggtgtggaccttgcaaaatgatcaagcccttcttccattccctctgtgacaagtattccaatgtggtgttccttgaagtggatgtggatgactgccaggatgttgctgcagactgtgaagtcaaatgcatgccgaccttccagttttataaaaagggtcaaaaggtgggggagttctccggtgctaacaaggaaaagcttgaagcctctattactgaatatgcctaa(seqidno:32)。
小鼠硫氧还蛋白2的序列是:
atggctcagcggctcctcctggggaggttcctgacctcagtcatctccaggaagcctcctcagggtgtgtgggcttccctcacctctaagaccctgcagacccctcagtacaatgctggtggtctaacagtaatgcccagcccagcccggacagtacacaccaccagagtctgtttgacgacctttaacgtccaggatggacctgactttcaagacagagttgtcaacagtgagacaccagttgttgtggactttcatgcacagtggtgtggcccctgcaagatcctaggaccgcggctagagaagatggtcgccaagcagcacgggaaggtggtcatggccaaagtggacattgacgatcacacagaccttgccattgaatatgaggtgtcagctgtgcctaccgtgctagccatcaagaacggggacgtggtggacaagtttgtggggatcaaggacgaggaccagctagaagccttcctgaagaagctgattggctga(seqidno:33)。
小鼠硫氧还蛋白还原酶1的序列是:
atgaatggctccaaagatccccctgggtcctatgacttcgacctgatcatcattggaggaggctcaggaggactggcagcagctaaggaggcagccaaatttgacaagaaagtgctggtcttggattttgtcacaccgactcctcttgggaccagatggggtctcggaggaacgtgtgtgaatgtgggttgcatacctaagaagctgatgcaccaggcagctttgctcggacaagctctgaaagactcgcgcaactatggctggaaagtcgaagacacagtgaagcatgactgggagaaaatgacggaatctgtgcagagtcacatcggctcgctgaactggggctaccgcgtagctctccgggagaaaaaggtcgtctatgagaatgcttacgggaggttcattggtcctcacaggattgtggcgacaaataacaaaggtaaagaaaaaatctattcagcagagcggttcctcatcgccacaggtgagaggccccgctacctgggcatccctggagacaaagagtactgcatcagcagtgatgatcttttctccttgccttactgcccggggaagaccctagtagttggtgcatcctatgtcgccttggaatgtgcaggatttctggctggtatcggcttagacgtcactgtaatggtgcggtccattctccttagaggatttgaccaagacatggccaacaaaatcggtgaacacatggaagaacatggtatcaagtttataaggcagttcgtcccaacgaaaattgaacagatcgaagcaggaacaccaggccgactcagggtgactgctcaatccacaaacagcgaggagaccatagagggcgaatttaacacagtgttgctggcggtaggaagagattcttgtacgagaactattggcttagagaccgtgggcgtgaagataaacgaaaaaaccggaaagatacccgtcacggatgaagagcagaccaatgtgccttacatctacgccatcggtgacatcctggaggggaagctagagctgactcccgtagccatccaggcggggagattgctggctcagaggctgtatggaggctccaatgtcaaatgtgactatgacaatgtcccaacgactgtatttactcctttggaatatggctgttgtggcctctctgaagaaaaagccgtagagaaatttggggaagaaaatattgaagtttaccatagtttcttttggccattggaatggacagtcccatcccgggataacaacaaatgttatgcaaaaataatctgcaaccttaaagacgatgaacgtgtcgtgggcttccacgtgctgggtccaaacgctggagaggtgacgcagggctttgcggctgcgctcaagtgtgggctgactaagcagcagctggacagcaccatcggcatccacccggtctgtgcagagatattcacaacgttgtcagtgacgaagcgctctgggggagacatcctccagtctggctgctga(seqidno:34)。
小鼠硫氧还蛋白还原酶2的序列是:
atggcggcgatggtggcggcgatggtggcggcgctgcgtggacccagcaggcgcttccggccgcggacacgggctctgacacgcgggacaaggggcgcggcgagtgcagcgggagggcagcagagctttgatctcttggtgatcggtgggggatccggtggcctagcttgtgccaaggaagctgctcagctgggaaagaaggtggctgtggctgactatgtggaaccttctccccgaggcaccaagtggggccttggtggcacctgtgtcaacgtgggttgcatacccaagaagctgatgcatcaggctgcactgctggggggcatgatcagagatgctcaccactatggctgggaggtggcccagcctgtccaacacaactggaagacaatggcagaagccgtgcaaaaccatgtgaaatccttgaactggggtcatcgcgtccaactgcaggacaggaaagtcaagtactttaacatcaaagccagctttgtggatgagcacacagttcgcggtgtggacaaaggcgggaaggcgactctgctttcagctgagcacattgtcattgctacaggaggacggccaaggtaccccacacaagtcaaaggagccctggaatatggaatcacaagtgacgacatcttctggctgaaggagtcccctgggaaaacgttggtggttggagccagctatgtggccctagagtgtgctggcttcctcactggaattggactggataccactgtcatgatgcgcagcatccctctccgaggctttgaccagcaaatgtcatctttggtcacagagcacatggagtctcatggcacccagttcctgaaaggctgtgtcccctcccacatcaaaaaactcccaactaaccagctgcaggtcacttgggaggatcatgcttctggcaaggaagacacaggcacctttgacactgtcctgtgggccatagggcgagttccagaaaccaggactttgaatctggagaaggctggcatcagtaccaaccctaagaatcagaagattattgtggatgcccaggaggctacctctgttccccacatctatgccattggagatgttgctgaggggcggcctgagctgacgcccacagctatcaaggcaggaaagcttctggctcagcggctctttgggaaatcctcaaccttaatggattacagcaatgttcccacaactgtctttacaccactggagtatggctgtgtggggctgtctgaggaggaggctgtggctctccatggccaggagcatgtagaggtttaccatgcatattataagcccctagagttcacggtggcggatagggatgcatcacagtgctacataaagatggtatgcatgagggagcccccacaactggtgctgggcctgcacttccttggccccaacgctggagaagtcacccaaggatttgctcttgggatcaagtgtggggcttcatatgcacaggtgatgcagacagtagggatccatcccacctgctctgaggaggtggtcaagctgcacatctccaagcgctccggcctggagcctactgtgactggttgctga(seqidno:35)。
实施例10:体外trx/trx还原酶研究
测量和方法
将商品化trxr(大鼠肝)溶液(4μm)用水稀释以产生2.86μm溶液。将冻干的trx(人)用pbs(10mm,ph7.2)重建,产生500μm溶液。在水中配制20mmnadph溶液及10mmatg和atm溶液。
在黑色聚丙烯1.5ml微量离心管中,温和混合437μl反应缓冲液(10mm组氨酸、10mmna2so4、137mmnacl、2.5mmkcl,ph7.0)、25μlnadph、16μl配制好的ocrelizumab溶液(30.2mg/ml)和5μltrx。通过添加17.5μltrxr来启动反应。将反应于室温温育24小时。在每个采样时间点取出20μl等分试样,并保存于-70℃,直至通过bioanalyzer测定法分析。
使用上文所述相同反应条件及添加各种抑制剂证明了对trx系统的抑制。
1.硫氧还蛋白系统的体外活性
图24显示了来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了将完整ocrelizumab(“2h7”,一种人源化抗cd20抗体,上文称作“变体a”)(1mg/ml)与10mm硫酸组氨酸缓冲液中的0.1μmtrxr(大鼠肝)、5μmtrx(人)和1mmnadph一起温育导致不到1小时里ocrelizumab的还原。
2.受到金硫葡糖抑制的硫氧还蛋白系统的体外活性
以下列浓度向上文所述ocrelizumab混合物添加金硫葡糖(atg):1mm;0.6μm(6∶1atg∶trxr);0.4μm(4∶1atg∶trxr);和0.2μm(2∶1atg∶trxr)。
如图25-27所示来自bioanalyzer分析的数字凝胶样图像所证明的,以1mm、0.6μm、和0.4μm的浓度添加的金硫葡糖有效抑制硫氧还蛋白系统对ocrelizumab的还原。然而,如图28所示,在这些实验条件下,以0.2μm的浓度添加的金硫葡糖在24小时后不能抑制ocrelizumab还原。
3.受到金硫苹果酸抑制的硫氧还蛋白系统的体外活性
以0.1mm和0.01mm的浓度向上文所述ocrelizumab混合物添加金硫苹果酸(atm)。如图29和30所示来自bioanalyzer分析的数字凝胶样图像所证明的,atm以所测试的两个浓度都有效抑制硫氧还蛋白系统对ocrelizumab的还原。
4.受到cuso4抑制的硫氧还蛋白系统的体外活性
以20μm(4∶1cu2+∶trx);10μm(2∶1cu2+∶trx);和5μm(1∶1cu2+∶trx)的浓度向上文所述ocrelizumab混合物添加cuso4。如图31-33所示,cuso4在20μm和10μm的浓度有效抑制硫氧还蛋白诱导的ocrelizumab还原(图31和32),但是5μm浓度不足以导致对还原的完全抑制(图33)。
5.受到胱胺抑制的硫氧还蛋白系统的体外活性
以下列浓度向上文所述ocrelizumab混合物添加胱胺:532μm(20∶1胱胺∶2h7(变体a)二硫化物);266μm(10:1胱胺:2h7(变体a)二硫化物);133μm(5:1胱胺:2h7二硫化物);和26.6μm(1:1胱胺:2h7(变体a)二硫化物)。如图34-37所示,胱胺在532μm(20:1胱胺:2h7(变体a)二硫化物)和266μm(10:1胱胺:2h7(变体a))的浓度有效抑制硫氧还蛋白诱导的ocrelizumab还原(图34和35),但是133μm(5∶1胱胺:2h7(变体a)二硫化物)和26.6μm(1:1胱胺:2h7(变体a)二硫化物)浓度不足以在24小时后抑制ocrelizumab还原(图36和37)。
6.受到胱氨酸抑制的硫氧还蛋白系统的体外活性
以2.6mm的浓度向上文所述ocrelizumab混合物添加胱氨酸。如图38所示,在此浓度,胱氨酸有效抑制硫氧还蛋白系统对ocrelizumab的还原。应当注意,胱氨酸的最小有效浓度(正如其它抑制剂的有效最小浓度)取决于实际情况,而且对于不同蛋白质(诸如抗体)可以是不同的,而且可以随添加的时间而变化。如此,例如,如果在裂解前添加胱氨酸,那么对于抗体2h7(变体a)的最小有效浓度为约1.3mm,对于apomab为约1mm而对于抗体变体c为约4.5mm。当在细胞培养基中添加胱氨酸时,最小有效浓度通常要高一些,而且对于2h7(变体a)是约5.2mm,对于apomab是6mm而对于抗体变体c是9mm。通常,对于胱氨酸、胱胺和氧化型谷胱甘肽(见下文),最小有效抑制浓度是约40倍抗体浓度(以μm计)。
7.受到氧化型谷胱甘肽(gssg)抑制的硫氧还蛋白系统的体外活性
以2.6mm的浓度向上文所述ocrelizumab混合物添加gssg。如图39所示,在此浓度,gssg有效抑制硫氧还蛋白系统对ocrelizumab的还原。然而,应当注意,氧化型谷胱甘肽的最小有效浓度(正如其它抑制剂的)取决于实际情况,诸如例如所生成的蛋白质(例如抗体)的本质和添加的时间。例如,对于抗体2h7(变体a),裂解前添加的最小有效浓度为约1.3mm。
8.酶促还原系统的体外活性
图40显示了来自bioanalyzer分析的数字凝胶样图像(每条带代表一个时间点),显示了将完整ocrelizumab(“2h7”,一种人源化抗cd20抗体,变体a)(1mg/ml)与50mm硫酸组氨酸缓冲液ph7.38中的10μg/ml己糖激酶、50μg/ml葡萄糖-6-磷酸脱氢酶、5μm硫氧还蛋白、0.1μm硫氧还蛋白还原酶、2mm葡萄糖、0.6mmatp、2mmmg2+、和2mmnadp一起温育在约1小时里导致ocrelizumab还原。添加0.1mmhdea(一种已知的葡萄糖-6-磷酸脱氢酶抑制剂)不抑制还原。
9.酶促还原系统的体外活性需要nadph
如图41来自bioanalyzer分析的数字凝胶样图像所示,将完整ocrelizumab(1mg/ml)与50mm硫酸组氨酸缓冲液ph7.38中的5μm硫氧还蛋白、0.1μm硫氧还蛋白还原酶、和2mmnadp一起温育不导致ocrelizumab抗体的还原。ocrelizumab还原不能在没有己糖激酶和葡萄糖-6-磷酸脱氢酶和它们的底物来生成nadph的情况中发生。
本文中合适地例示性描述的发明可以在缺乏未在本文中明确公开的任何一种或多种成分、一项或多项限制的情况中实施。如此,例如在本文中的每种情况中,术语“包含”、“包括”、“含有”等应当读成开放式的且没有限制。另外,本文中所采用的术语和表述作为描述的术语使用而并非限制,而且使用此类术语和表述并不意图排除所显示的发明的任何等同方案或其部分,而是认识到在要求保护的发明的范围内的各种变化是有可能的。如此,应当理解的是,尽管已经通过优选的实施方案和任选的特征明确地公开了本发明,但是本领域技术人员可以容易地做出本文中公开的发明的修改和变化,并且此类修饰和变化被认为是在本文所公开的发明的范围内的。本文中已经广泛地和一般性地描述了本发明。落在一般公开内的每一种更窄的种类和亚类的分组也构成这些发明的一部分。这在每一项发明的一般描述内包括一项附带条件或负面限制,其会容许自该类别中消除任何主题,不管要消除的材料是否是具体叙述的。另外,在按马库什组描述本发明的特征或方面时,本领域技术人员会认可本发明也由此是以马库什组的任何成员个体或成员亚组的方式描述的。另外,当提到本发明的一个方面列出成员个体的一个范围时,像例如“seqidno:1至seqidno:100(含两端点)”,等同于个别地列出该列表的每一个成员,而且应当理解,权利要求中可以个别地排除或包括每一个成员个体。
本文方法中所描绘的和/或所使用的步骤可以以与所描绘的和/或所叙述的不同的次序实施。这些步骤仅仅是这些步骤可发生的次序的例示。这些步骤可以以期望的任何次序发生,使得它仍执行要求保护的发明的目的。
根据本文中对本发明的描述,显然可以使用各种等效方案来执行本发明的概念而不背离其范围。此外,虽然已经具体参照某些实施方案描述了本发明,但是本领域普通技术人员会认识到可以在形式和细节方面做出变化而不背离本发明的精神和范围。所描述的实施方案在所有方面视为例示性的而非限制性的。还应当理解,本发明不限于本文所述特定实施方案,但是能够进行许多等效方案、重新安排、改良、和替换,而不背离本发明的范围。如此,别的实施方案也在本发明的范围内且在所附权利要求内。
通过述及将本文中提到的所有美国专利和申请;外国专利和申请;科学论文;书籍;和出版物完整收入本文,就像通过述及明确且个别地指明收录每一篇单独的专利或出版物一样,包括任何图表,就像完整列出一样。
- 还没有人留言评论。精彩留言会获得点赞!