一种重组免疫细胞因子及其应用的制作方法
本发明属于生物技术领域,更具体地,本发明涉及一种靶向pd-l1与il-2受体的重组免疫细胞因子及其应用。
背景技术:
pd-1是ⅰ型跨膜糖蛋白,由与cd28家族有21%-22%的同源基因的igv型胞外域、跨膜区和胞内尾区组成。其中胞内尾区由免疫受体酪氨酸抑制基序上两个磷酸化位点和免疫受体络氨酸开关模块构成。这一结构,在磷酸化后可以通过磷酸化的src同源性磷酸酶1(shp-1)和shp-2来负性调节tcr信号。因其缺乏形成二聚体所必需的临近细胞膜的半胱氨酸残基,故pd-1以单体的形式存在。编码pd-1的基因pdcd1在人2号染色体上,包含5个外显子(keir,etal.,pd-1anditsligandsintoleranceandimmunity.annurevimmunol,2008.26:p.677-704)。pd-1有两个配体,pd-l1和pd-l2,它们同属于b7跨膜蛋白家族(freeman,etal.,engagementofthepd-1immunoinhibitoryreceptorbyanovelb7familymemberleadstonegativeregulationoflymphocyteactivation.jexpmed,2000.192(7):p.1027-34)。
pd-1配体(pd-l1和pd-l2)为组成型表达,或在多种细胞类型中可被诱导,这些细胞类型包括非造血组织以及各种肿瘤类型。pd-l1在b细胞、t细胞、髓样细胞和树突细胞(dc)上表达,但也在外周细胞(例如微血管内皮细胞)和非淋巴器官(如心脏、肺等)中表达。与此相反,pd-l2仅在巨噬细胞和dc中存在,与pd-l2只表达于apc上不同,pd-l1表达在多种细胞表面,并且会因ifn-γ和il-4等前炎性因子通过信号转导子和转录激活子1(stat1)和干扰素调节因子1(irf1)而上调表达(sharpe,etal.,thefunctionofprogrammedcelldeath1anditsligandsinregulatingautoimmunityandinfection.natimmunol,2007.8(3):p.239-45.)。
pd-l1由一个跨膜区和两个igc和igv胞外域构成(dong,etal.,b7-h1,athirdmemberoftheb7family,co-stimulatest-cellproliferationandinterleukin-10secretion.natmed,1999.5(12):p.1365-9),由人9号染色体上cd274基因编码,包含7个外显子(keir,etal.,pd-1anditsligandsintoleranceandimmunity.annurevimmunol,2008.26:p.677-704),每个外显子都编码pd-l1的不同部分。与b7家族其他共刺激蛋白相似,pd-l1也在人体循环中以可溶性形式存在,并且主要存在于骨髓来源单核细胞,巨噬细胞和树突细胞表面。虽然激活状态的t细胞不会产生大量spd-l1,但是spd-l1仍然保留它的igv配体结合区可以和t细胞表面的pd-1结合来抑制t细胞的激活。spd-l1的生理重要性还有待研究,但是目前已知在年长的成人体内spd-l1含量普遍较高。在产生spd-l1的时候膜表面的pd-l1(mpd-l1)会被金属蛋白酶(mmps)清除。spd-l1在多种人类肿瘤细胞中存在,包括非小细胞肺癌(h1299)细胞,淋巴瘤细胞株,卵巢癌细胞株(ho8910),肺腺癌细胞株(spca-1)和恶性胶质瘤(u251)(chen,etal.,developmentofasandwichelisaforevaluatingsolublepd-l1(cd274)inhumanseraofdifferentagesaswellassupernatantsofpd-l1+celllines.cytokine,2011.56(2):p.231-8.)。研究表明血液中升高的spd-l1水平与乳腺癌和b细胞淋巴瘤的进展和不良愈后有关(rossille,etal.,highlevelofsolubleprogrammedcelldeathligand1inbloodimpactsoverallsurvivalinaggressivediffuselargeb-celllymphoma:resultsfromafrenchmulticenterclinicaltrial.leukemia,2014.28(12):p.2367-75.)。
随着对免疫检查点抑制t细胞功能的深入研究,开发免疫检查点抑制剂用以治疗癌症和肿瘤的进程也在加快。目前,靶向ctla-4的封闭性抗体ipilimumab(伊匹单抗)和靶向pd-1抗体nivolumab(百时美施贵宝)和pembrolizumab(默沙东)都已通过fda批准用于治疗进展性黑色素瘤和非小细胞肺癌(nsclc)。而靶向pd-l1单抗mpdl3280a(罗氏)也已被fda批准用于治疗pd-l1阳性的非小细胞肺癌(nsclc)。虽然免疫检查点抗体能够使患者肿瘤维持持续的消退,提高患者的无进展生存期(pfs)和整体生存期(os),但较低的客观反应率仍然是大部分患者治疗无效的原因,这一问题阻碍了这类抗体的广泛应用和发展pardoll,d.m.,theblockadeofimmunecheckpointsincancerimmunotherapy.natrevcancer,2012.12(4):p.252-64.)。从免疫调节的效果上说,靶向ctla-4和pd-1抗体的联合应用显示出了非常好的治疗效果,例如nivolumab和ipilimumab联合应用能显著提高进展性黑色素瘤患者2年生存率(wolchok,j.d.,etal.,nivolumabplusipilimumabinadvancedmelanoma.nengljmed,2013.369(2):p.122-33.)。作为一个重要的调节机制,在病理状态下,pd-l1在大多数肿瘤细胞表面的表达量增高后,会对t细胞激活形成负性调节,抑制自身免疫反应。作为一种免疫抑制蛋白,pd-l1在许多类型的肿瘤中表达量都会上升,这类患者也常常伴有对pd-1/pd-l1抗体治疗产生抵抗和不良愈后。但是,那些细胞表面未检测到pd-l1表达的肿瘤类型有时也同样可以对抗pd-l1抗体治疗产生应答,这表明肿瘤表面pd-l1检测的预测作用可能并不适用于所有的肿瘤类型。而且,多种肿瘤内和外部都有广泛的不同程度的pd-l1表达,这种现象同时也会影响pd-l1的检测结果(blackburn,s.d.,etal.,tissue-specificdifferencesinpd-1andpd-l1expressionduringchronicviralinfection:implicationsforcd8t-cellexhaustion.jvirol,2010.84(4):p.2078-89)。
利用单克隆抗体抑制pd-l1功能在临床上显著提高了癌症患者的整体生存率。因为pd-l1表达同时受到mapk和pi3k/akt调节,反之抑制这些信号通路也可减少pd-l1的表达。但是受体络氨酸激酶抑制剂却能导致高表达pd-l1的肺癌患者有更好的愈后(velcheti,etal.,programmeddeathligand-1expressioninnon-smallcelllungcancer.labinvest,2014.94(1):p.107-16.)。
肿瘤发生和发展都伴随着对免疫系统的入侵,免疫功能不全的个体常常有较高的癌症发生率和较差的愈后。因此研究者都投入大量的时间精力提高癌症患者的免疫功能,包括给予免疫刺激性的细胞因子治疗,如ifn、il-2等;利用树突细胞(dc)为基础的疫苗来激活免疫系统以提高内源性免疫反应;利用自体癌症特异性细胞毒性t细胞(ctls)激活和扩增的过继细胞转移治疗;或者改造t细胞使其表达嵌合抗原受体用以识别肿瘤细胞特异性抗原等。
通过信号抑制减少pd-l1表达可以与抗pd-l1治疗联合,将pd-1/pd-l1通路控制在低水平可以显著增加治疗效果,因此免疫治疗和靶向治疗可以联合应用。肿瘤免疫包含了固有免疫和适应性免疫两种。目前已在多种小鼠肿瘤模型中验证了nk细胞和t细胞,尤其是细胞毒性t细胞在肿瘤免疫过程中的作用。多个临床试验正在评估某些细胞因子单用,或者与多种化疗药物和肿瘤靶向的单克隆抗体或和其他细胞因子联用的抗癌疗效。例如,主要针对于固态肿瘤如黑色素瘤,肾癌,乳腺癌的多种重组细胞因子il-7,il-12,il-15,il-21等,和多用于多种血液和淋巴系统疾病的il-2和ifn-α等。
对于细胞因子的选择应基于细胞因子本身的免疫治疗能力,应具有激活t细胞,nk细胞,或肿瘤巨噬细胞或调解肿瘤基质或肿瘤内的信号通路的能力。大多数被选为肿瘤治疗的细胞因子均可以刺激t细胞免疫,或是通过推动树突细胞成熟分泌细胞因子(ifns,gm-csf,tnf-α)帮助杀伤肿瘤细胞。
il-2单独应用可通过激活t细胞,nk细胞和单核细胞刺激免疫系统,曾用于治疗进展性肾细胞肾癌和进展性黑色素瘤。在il-2在治疗的283名转移性黑色素瘤和转移性肾癌患者中,其整体反应率分别为17%和20%(rosenberg,etal.,treatmentof283consecutivepatientswithmetastaticmelanomaorrenalcellcancerusinghigh-dosebolusinterleukin2.jama,1994.271(12):p.907-13.)。然而il-2广泛应用最大的限制是以高血压和发热为特点的血管渗漏综合征这一严重不良反应和只有5-7分钟的半衰期,随后的二次清除时间为30-120分钟。il-2的另外一个缺点就是它广泛与表达在免疫细胞上的受体结合,使调节性t细胞(treg)扩增,而阻碍机体的抗癌免疫(boyman,theroleofinterleukin-2duringhomeostasisandactivationoftheimmunesystem.natrevimmunol,2012.12(3):p.180-90.)。
虽然多种免疫制剂均可引起有效的免疫反应来抵抗肿瘤细胞的入侵,然而通过激活免疫系统来治疗癌症仍存在一定的缺点:严重的不良反应、严重的脱靶效应和癌症细胞表面上调的免疫抑制因子导致的低反应率。t细胞的激活必须依靠从抗原提呈细胞(apc)接受两种信号:第一种为t细胞受体识别mhc分子与apc表面抗原形成的复合物而介导的抗原特异性信号。第二种为t细胞表面的cd28和apc表面的cd80或cd86结合引起的共刺激信号。刺激或抑制受体与配体之间的相互作用可以调控t细胞的激活。t细胞,apc细胞和其他类型的细胞上表达的成对的受体与配体都统称为“免疫检查点”。ctla-4和pd-1就是其中的代表。
为了避免il-2的毒性,并充分利用其激活免疫细胞的功能,本发明利用抗pd-l1抗体,封锁pd-1/pd-l1信号通路,激活t细胞自体免疫;与其融合单细胞因子il-2可与多种淋巴细胞表面受体结合,赋予il-2靶向性,募集于肿瘤部位,使机体适应性免疫和固有免疫相结合,加大抑瘤效果。
技术实现要素:
靶向pd-l1的单抗具有免疫激活功能,并已显示出对多种肿瘤的良好治疗效果。其主要作用原理是通过阻止肿瘤细胞表面pd-l1分子与t细胞表面pd-1分子的结合,从而解除其对t细胞的抑制作用,增强抗肿瘤免疫效应。但由于临床上某些肿瘤细胞表面pd-l1低表达,应用抗pd-l1单抗治疗的疗效较差,病情易反复。因此需要开发新型抗肿瘤免疫制剂来提高单抗疗效,改善肿瘤愈后。il-2因能激活体内多种免疫细胞而可以用来治疗肿瘤和炎症性疾病。但由于其受体广泛表达,系统性应用il-2会引起严重的不良反应。
因此,我们采用基因工程技术,将靶向pd-l1抗体的fc段与il-2分子融合,构建新型重组免疫细胞因子bipi。其目的是使之能特异性靶向肿瘤区域,避免非特异性结合导致的血管毒性;同时阻断pd-1/pd-l1结合,使机体的适应性免疫协同特异性免疫,激活t细胞,b细胞等多种淋巴细胞,释放细胞因子,以期达到对抗原表达量低,单用抗体效果不明显肿瘤的更好治疗效果。
本发明公开了:
一种靶向pd-l1与il-2受体的重组免疫细胞因子,其特征在于,与pd-l1结合,同时与il-2受体结合。
如上所述的一种靶向pd-l1与il-2受体的重组免疫细胞因子,其特征在于,重组免疫细胞因子的重链恒定区为igg1,重链fc段连接il-2分子。
如上所述的一种靶向pd-l1与il-2受体的重组免疫细胞因子,其特征在于,重组免疫细胞因子的轻链具有seqidno:2所述的氨基酸序列,重组免疫细胞因子的重链具有seqidno:4所述的氨基酸序列。
如上所述的一种组合物,含有如上所述的靶向pd-l1与il-2受体的重组免疫细胞因子,和药学上可接受的载体。
如上所述的靶向pd-l1与il-2受体的重组免疫细胞因子在制备抗体肿瘤药物中的应用。
如上所述的组合物在制备抗肿瘤药物中的应用。
如上所述的靶向pd-l1与il-2受体的重组免疫细胞因子在制造免疫调节药物上的应用。
如上所述的组合物在制造免疫调节药物上的应用。
如上所述的任一用途,还包括和其他的抗肿瘤药物联合使用。
利用全基因合成技术将抗pd-l1单抗的igg1亚类抗体的fc重链基因通过linker与突变后的il-2基因融合,连接至表达载体pc3.1dna,构建重链质粒。mpdl3280单抗的可变区基因连接至表达载体pcdna3.1构建轻链质粒,并与重链质粒共同转入cho细胞中进行表达。经过细胞批次培养,纯化上清得到新型重组免疫细胞因子,简称bipi。
利用以下手段对其进行表征鉴定:sds-page分析其分子量;流式细胞术(fcm)测定bipi的亲和力和抗原结合力;体外检测其对小鼠t淋巴细胞系ctll-2细胞的促增殖作用;在小鼠的转移性结直肠癌预防性肿瘤模型中评估其抑瘤活性,并在治疗过程中监测小鼠体重,评估bipi的安全性;利用抗体消除小鼠体内产生的cd8+t细胞,验证其在抑瘤过程中的作用;最后分离各治疗组小鼠脾细胞,用磁珠法从混合细胞中分离得到cd8+t细胞,测定并比较各组间cd8+t细胞数量上的差异和其ifn-γ分泌量的不同,验证bipi对体内免疫系统的激活作用并分析免疫细胞因子的作用机制。
获得新型免疫细胞因子,其纯度大于99%,经sds-page、fcm等试验证实bipi结构正确并且与抗原有较好的结合活性。体外实验中,含有与il-2同等摩尔质量的bipi与il-2标准品有几乎相同的促进ctll-2细胞生长的效能。在弱表达pd-l1的ct26转移性结直肠癌小鼠模型上,与对照组相比显著减少了肺表面肿瘤结节的数量,证实了免疫细胞因子bipi的抑瘤活性。在后续对其抑瘤机制研究的过程中发现,cd8+t细胞在bipi的抑瘤效果发挥上有重要作用;消除cd8+t细胞后,小鼠的生存期显著缩短,并且生存状态不佳。在治疗结束后,将cd8+t细胞分离后培养,检测其ifn-γ分泌量,结果显示bipi治疗组显著高于其他对照组,结果具有统计学意义。在体重监测过程中各组小鼠体重均未发生异常改变,说明治疗性药物并未对小鼠产生明显毒性,提示其安全性良好。
经过实验证实,我们所构建的新型重组免疫细胞因子bipi具有较好的抑瘤活性,尤其针对pd-l1弱表达的肿瘤类型,达到激活体内淋巴细胞,促进细胞因子分泌的作用。在临床上应用于pd-l1低表达肿瘤,可弥补由于抗原结合不足导致的单抗药物疗效差,易复发等问题。
附图说明
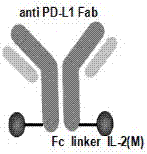
图1、bipi重组免疫细胞因子重链结构示意图
图2、bipi重组免疫细胞因子结构示意图
图3、bipi重组免疫细胞因子重链构建sds-page鉴定图
图4、筛选高表达bipi重组免疫细胞因子细胞株dot-blot图谱
图5、bipi重组免疫细胞因子分子量与纯度检测图谱
图6、bipi重组免疫细胞因子对抗原结合能力fcm检测图谱
图7、bipi与il-2标准品对ctll-2细胞促增殖作用检测图谱
图8、bipi双侧结合能力流式细胞术fcm检测图谱
图9、bipi在ct26模型中的疗效检测肺表面结节图谱
图10、bipi在ct26模型中的疗效检测肺表面结节统计分析图谱
图11、bipi治疗中小鼠体重变化趋势图谱
图12、bipi治疗组小鼠脾脏分离的cd8+t细胞数量统计图谱
图13、bipi治疗组小鼠脾脏分离的cd8+t细胞分泌ifn-γ含量分析图谱。
具体实施方式
实施例1、免疫细胞因子bipi的构建、表达与鉴定
利用基因工程的方法,将pd-l1单抗的重链恒定区基因与il-2突变后基因进行全基因合成,得到bipi全长重链片段,酶切后连接到pcdna3.1真核表达载体上。重链结构示意图如图1所示,和蛋白结构示意图如图2所示,原抗pd-l1抗体重链fc端通过(ggggs)3linker连接il-2基因。通过ecori和hindiii双酶切鉴定,结果见图3,证明已成功构建了bipi轻链质粒和bipi重链质粒。后经测序分别验证了轻链和重链序列的正确性。
将上述构建成功的bipi轻链和重链质粒转染cho细胞后,在培养基中加入g418进行抗性培养2周,利用有限稀释法对亚克隆进行筛选。共获得9株单克隆细胞,设置人igg点膜组为阳性对照进行用dot-blot试验,,检测各细胞株蛋白的表达量。在显色后,利用软件分析各克隆对相对于阳性对照隆显色深度百分比。如图4所示,b2单克隆具有最高的相对强度,表明此克隆的目的蛋白产量最多,故选择此克隆扩大培养。
将b2单克隆的细胞株通过细胞批次培养技术进行扩大培养后,将其上清通过proteina亲和层析柱纯化,得到目的蛋白bipi。通过sds-page鉴定了抗体的分子量。如图5所示(泳道1、3为抗pd-l1抗体,泳道2、4为bipi),将免疫细胞因子还原后重链和轻链的分子量分别为65.5kd和25kd。对照组为抗pd-l1单抗(mpdl3280a)。
实验例1、免疫细胞因子bipi对细胞表面pd-l1抗原的结合活性检测
(1)将重组的高表达pd-l1的cho细胞以2×105的密度重悬于1%fbspbs缓冲液中,每管100ul加入流式管。垂直滴入,勿将细胞悬液滴于管壁;
(2)稀释抗体:将纯化得到的bipi用pbs稀释到100ug/ml浓度,100ul/管加入流式管,三复孔。其他对照组(igg,pd-l1抗体和pbs)同上处理;用涡旋振荡器将各组流式管中溶液混匀,避光置于4℃冰箱静置1h;
(3)将流式管以1000r/min离心5min,迅速倾倒出上层液体,勿震荡以免管底部的细胞损失,之后以200ul/管加入含1%fbs的pbs溶液,震荡混匀后再次离心,重复该步骤2次,洗去细胞表面未结合的抗体;
(4)加入hrp-goatanti-humanigg(1:300稀释),100ul/管加入流式管,震荡混匀,于4℃避光孵育45min;
(5)重复步骤(3),待细胞洗涤2次后,用100ul1%fbs的pbs溶液重悬细胞,上机读数前需震荡使细胞混匀。
利用流式细胞检测法(fcm)初步测定bipi对抗原pd-l1的结合活性。图6的结果表明,bipi与抗pd-l1单抗(mpdl3280a)可同等程度地与高表达pd-l1的cho工程细胞结合,表明我们构建的免疫细胞因子依然具有较强的pd-l1抗原结合活性。
实验例2、免疫细胞因子bipi中il2生物活性检测
(1)取生长状态良好的ctll-2细胞,每孔1×104个细胞均匀铺于96孔细胞培养板中;
(2)每孔加入不同浓度的梯度倍比稀释的免疫细胞因子和重组人il-2,抗体以100ng/ml的浓度起始进行稀释,通过计算,使bipi中所含il-2与il-2标准品有相同的摩尔质量起始,进行2倍比稀释,共15个浓度,并作三复孔,于37℃、8%co2的孵箱中培养72h;
(3)每孔60ul加入1:6比例混合的aps和mts试剂进行显色。显色3h后将细胞培养板置于imark酶标仪中,设置波长为450nm并读取各组吸光度值;
(4)细胞活率百分比=[(试验组读数-空白组读数)∕(igg对照组读数-空白组读数)]×100。
验证免疫细胞因子中的il-2是否与重组人il-2标准品有相同的促进t细胞增殖的功能。由图7结果可知,在起始浓度为100ng/ml时,细胞增长达到平台期,与ril-2标准品有相似的促进t细胞增殖功能。ril-2组的ec50=0.4022;bipi组的ec50=0.2288。
通过计算,使bipi中il-2含量与重组ril-2具有相等摩尔质量,结果显示对ctll-2细胞有相近的促进增殖作用。其ec50值分别为:0.2288和0.4022。
实验例3、免疫细胞因子bipi的双特异性结合能力检测
(1)将生长状态良好的ct26细胞收集到15ml离心管中,1000rpm,离心5分钟,倒掉上层培养液,将细胞用含有1%fbs的pbs溶液重悬;
(2)对细胞进行计数,并以2×106个细胞/管的量加入流式管中,每管100ul,设置三复孔
(3)将bipi和igg分别以100ug/ml的浓度起始稀释,3倍比稀释,每组三复孔,共稀释12个浓度,将100ul稀释后的实验药物和对照分别垂直加入流式管,震荡3s,使细胞和抗体充分混合,用锡箔纸盖住,置于4°冰箱1小时;
(4)将流式管1000r/min离心5min,迅速倾倒出上层液体,勿震荡以免管底部的细胞损失,之后以200ul/管的体积加入含1%fbs的pbs溶液,震荡混匀后再次离心,重复该步骤2次,洗去细胞表面未结合的抗体;
(5)加入pe标记的兔抗人il-2二抗(1:300稀释),100ul/管加入流式管,震荡混匀,于4℃避光孵育45min;
(6)重复步骤(4),洗涤细胞2次后,用200ul含1%fbs的pbs溶液重悬细胞,震荡混匀后上机检测荧光读数。
在体外实验中验证bipi的双侧结合能力,从图8结果可知,bipi的fab段仍能保持与ct26细胞表面pd-l1的结合能力,并且在fc端的il-2也能与pe连接的il-2抗体进行结合。说明bipi可以靶向pd-l1,并且il-2可以与表达il-2r的淋巴细胞结合,募集淋巴细胞至肿瘤区域,发挥其抑瘤作用。
bipi可与ct26细胞结合,并且同时能和pe标记的il-2抗体结合,结合能力呈线性趋势,说明bipi有较好的的双侧结合能力,fab和fc部分均能结合目的蛋白
实验例4、免疫细胞因子bipi肿瘤预防和安全性评估
为评估bipi的体内抑瘤活性建立了肺静脉转移的预防性肿瘤模型。
如图9、图10所示(图9:igg治疗组肺表面呈现大小不等的结节,数量最多,在il-2和pa治疗组,虽然结节数量有所减少,但还是比较bipi治疗组有比较明显的转移结节,而bipi治疗组小鼠肺表面基本没有结节。每个治疗组选取一只小鼠肺脏作为代表。图10:对各组小鼠肺表面结节进行计数并做统计学分析。*p<0.05),在babl/c小鼠转移性结直肠癌肿瘤模型中,bipi可有效的抑制肿瘤细胞的肺转移,显著减少肺表面的肿瘤结节。
治疗21天后,对各组小鼠肺表面肿瘤结节进行计数,并进行统计学分析。结果表明,对照组(注射人igg)的肿瘤结节数量显著高于bipi组,p<0.01,并且虽然单独应用抗pd-l1抗体组和il-2组有一定的抑制趋势,但与bipi治疗组还是在治疗效果上形成了明显的差别。此外,在ct26小鼠模型的治疗过程中,我们对babl/c小鼠的体重变化进行了监测,以初步评估bipi的体内用药安全性。图11的结果表明,在治疗过程中监测并记录小鼠体重变化,各组之间没有显著差异。在注射免疫细胞因子bipi后,与对照组相比,实验小鼠的体重并未出现明显下降或升高,各治疗组体重变化趋势相近。表明免疫细胞因子bipi在小鼠体内并未产生较大毒副作用,安全性良好。
实验例5、免疫细胞因子bipi疗效评估
因为在细胞毒性t细胞发挥作用的过程中,会分泌多种细胞因子,共同发挥协同促进作用来抑制肿瘤增长和转移。ifn-γ是重要的抗肿瘤细胞因子,因此,检测ifn-γ含量是否在bipi治疗组中和其他对照组是否有差别。从图12、13中结果可知(图12:不同治疗组小鼠脾脏分离出的cd8+t细胞数量统计图,bipi治疗组显著高于其他三组。图13:elisa方法测定不同组别小鼠脾脏分离出的cd8+t细胞分泌ifn-γ含量,bipi治疗组远远高于其他各组,并且单独使用抗体和il-2进行治疗产生的ifn-γ与对照组无统计学差异,说明单独应用两种药物并不能引起细胞因子大量分泌,从而协助杀伤肿瘤细胞),从bipi治疗组小鼠脾细胞中获得的cd8+t细胞也在数量上远高于其他治疗组。而bipi治疗组小鼠体内分离的cd8+t细胞,在72小时培养后产生的ifn-γ量相较其他组显著增多,抗体治疗组和il-2组均较低。这一结果也从侧面验证了bipi治疗组疗效优于其他组的原因,cd8+t细胞会大量扩增,并且其分泌的ifn-γ在这一抑瘤过程中也发挥了重要协同促进作用。
sequencelisting
<110>上海张江生物技术有限公司
<120>一种靶向pd-l1与il-2受体的重组免疫细胞因子及其应用
<130>2016
<160>4
<170>patentinversion3.5
<210>1
<211>645
<212>dna
<213>人工序列
<400>1
gatatacagatgacccagagcccatcttctctcagcgcatctgtgggcgaccgtgtcacc60
atcacttgtcgtgccagtcaggacgtctccactgccgtcgcctggtaccaacagaagcct120
gggaaggcgccaaaactgttaatctactccgctagttttctctacagcggtgtgccatcc180
aggttcagcggttctgggtcgggcacagattttaccctgaccatcagctctctccagcct240
gaggacttcgctacctactattgtcagcagtacttgtaccaccctgctaccttcggccag300
gggacaaaggtggagatcaagcgcacagtcgctgccccttccgtgttcatttttcctccc360
tctgacgagcagctaaagagcggtaccgcttcagtcgtgtgtttactgaacaacttttac420
cctcgggaagccaaggtccagtggaaggttgacaacgcactccagtccggcaattcgcag480
gagtctgttaccgagcaagactccaaggacagcacatactccctttcatcaactttgacg540
ttgtctaaggctgactacgagaagcataaagtgtatgcttgcgaggtgacacatcagggc600
ctgagttccccagttaccaaatcattcaatcggggggaatgctga645
<210>2
<211>214
<212>prt
<213>人工序列
<400>2
aspileglnmetthrglnserproserserleuseralaservalgly
151015
aspargvalthrilethrcysargalaserglnaspvalserthrala
202530
valalatrptyrglnglnlysproglylysalaprolysleuleuile
354045
tyrseralaserpheleutyrserglyvalproserargphesergly
505560
serglyserglythraspphethrleuthrileserserleuglnpro
65707580
gluaspphealathrtyrtyrcysglnglntyrleutyrhisproala
859095
thrpheglyglnglythrlysvalgluilelysargthrvalalaala
100105110
proservalpheilepheproproseraspgluglnleulyssergly
115120125
thralaservalvalcysleuleuasnasnphetyrproarggluala
130135140
lysvalglntrplysvalaspasnalaleuglnserglyasnsergln
145150155160
gluservalthrgluglnaspserlysaspserthrtyrserleuser
165170175
serthrleuthrleuserlysalaasptyrglulyshislysvaltyr
180185190
alacysgluvalthrhisglnglyleuserserprovalthrlysser
195200205
pheasnargglyglucys
210
<210>3
<211>1450
<212>dna
<213>人工序列
<400>3
gctagcaccaagggcccatcggtcttccccctggcaccctcctccaagagcacctctggg60
ggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcg120
tggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctca180
ggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacc240
tacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagccc300
aaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctgggggga360
ccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccct420
gaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactgg480
tacgtggacggcgtggaggtgcataatgccaagacaaagccgcgggaagagcagtacgcc540
agcacgtaccgtgtggtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaag600
gagtacaagtgcaaggtctccaacaaagccctcccagcccccatcgagaaaaccatctcc660
aaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggag720
atgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatc780
gccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtg840
ctggactccgacggctccttcttcctctacagcaagctcaccgtggacaagagcaggtgg900
cagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacg960
cagaagagcctctccctgtctcccggtggcggcggtagcggcggagggggcagcggcggt1020
ggcggaagcggcggtgggggctccgcgccaaccagttcctccactaaaaagactcagctc1080
cagctggagcacttgctcctcacactccagatgatcctcaacgggatcaacaactacaag1140
aatcctaaactgacacgcatgctcaccttcaagttctatatgcctaagaaagcaactgaa1200
ctcaaacatctgcaatgcctcgaagaggaactgaagcccctggaggaggtgctcaatctc1260
gcacagtctaaaaacttccacctgcgccctcgcgacttaatctcaaacattaacgtgatc1320
gtgctagagcttaaaggttccgaaaccacattcatgtgtgagtatgctgatgaaacggct1380
actatcgtcgagttcctgaaccgctggatcacattctgtcagtctatcatctccactctg1440
acttgaattc1450
<210>4
<211>599
<212>prt
<213>人工序列
<400>4
gluvalglnleuvalgluserglyglyglyleuvalglnproglygly
151015
serleuargleusercysalaalaserglyphethrpheseraspser
202530
trpilehistrpvalargglnalaproglylysglyleuglutrpval
354045
alatrpileserprotyrglyglyserthrtyrtyralaaspserval
505560
lysglyargphethrileseralaaspthrserlysasnthralatyr
65707580
leuglnmetasnserleuargalagluaspthralavaltyrtyrcys
859095
alaargarghistrpproglyglypheasptyrtrpglyglnglythr
100105110
leuvalthrvalserseralaserthrlysglyproservalphepro
115120125
leualaproserserlysserthrserglyglythralaalaleugly
130135140
cysleuvallysasptyrpheprogluprovalthrvalsertrpasn
145150155160
serglyalaleuthrserglyvalhisthrpheproalavalleugln
165170175
serserglyleutyrserleuserservalvalthrvalproserser
180185190
serleuglythrglnthrtyrilecysasnvalasnhislysproser
195200205
asnthrlysvalasplyslysvalgluprolyssercysasplysthr
210215220
histhrcysproprocysproalaprogluleuleuglyglyproser
225230235240
valpheleupheproprolysprolysaspthrleumetileserarg
245250255
thrprogluvalthrcysvalvalvalaspvalserhisgluasppro
260265270
gluvallyspheasntrptyrvalaspglyvalgluvalhisasnala
275280285
lysthrlysproargglugluglntyralaserthrtyrargvalval
290295300
servalleuthrvalleuhisglnasptrpleuasnglylysglutyr
305310315320
lyscyslysvalserasnlysalaleuproalaproileglulysthr
325330335
ileserlysalalysglyglnproarggluproglnvaltyrthrleu
340345350
proproserarggluglumetthrlysasnglnvalserleuthrcys
355360365
leuvallysglyphetyrproseraspilealavalglutrpgluser
370375380
asnglyglnprogluasnasntyrlysthrthrproprovalleuasp
385390395400
seraspglyserphepheleutyrserlysleuthrvalasplysser
405410415
argtrpglnglnglyasnvalphesercysservalmethisgluala
420425430
leuhisasnhistyrthrglnlysserleuserleuserproglygly
435440445
glyglyserglyglyglyglyserglyglyglyglyserglyglygly
450455460
glyseralaprothrserserserthrlyslysthrglnleuglnleu
465470475480
gluhisleuleuleuthrleuglnmetileleuasnglyileasnasn
485490495
tyrlysasnprolysleuthrargmetleuthrphelysphetyrmet
500505510
prolyslysalathrgluleulyshisleuglncysleuglugluglu
515520525
leulysproleuglugluvalleuasnleualaglnserlysasnphe
530535540
hisleuargproargaspleuileserasnileasnvalilevalleu
545550555560
gluleulysglysergluthrthrphemetcysglutyralaaspglu
565570575
thralathrilevalglupheleuasnargtrpilethrphecysgln
580585590
serileileserthrleuthr
595
- 还没有人留言评论。精彩留言会获得点赞!