赋予对鞘翅目有害生物的抗性的核酸分子的制作方法
本申请是2011年12月30日提交的申请号为201180068808.8(国际申请号pct/us2011/068062)、发明名称为“赋予对鞘翅目有害生物的抗性的核酸分子”的发明专利申请的分案申请。
优先权要求
本申请要求2010年12月30日提交的美国临时专利申请流水号61/428,592关于“nucleicacidmoleculesthatconferresistancetocoleopteranpests”的申请日(filingdate)权益。
发明领域
一般而言,本发明涉及对由鞘翅目有害生物引起的植物损伤的遗传控制。在特定的实施方案中,本发明涉及靶编码和非编码序列的鉴定,及重组dna技术用于转录后阻抑或抑制鞘翅目有害生物细胞中靶编码和非编码序列的表达以提供植物保护效果的用途。
发明背景
西部玉米根虫(westerncornrootworm、wcr),即玉米根萤叶甲(diabroticavirgiferavirgiferaleconte)是北美洲最具破坏性的玉米根虫物种之一,并且是美国中西部玉米生长区的一项特定忧虑。北部玉米根虫(northerncornrootworm、ncr),即北美玉米根萤叶甲(diabroticabarberismithandlawrence)是一种与wcr共同驻留于大量相同范围的紧密相关物种。存在着作为北美洲重要的有害生物的几种其它相关根萤叶甲属亚种:墨西哥玉米根虫(mcr),即d.virgiferazeaekrysanandsmith;南方玉米根虫(scr),即d.undecimpunctatahowardibarber;d.balteataleconte;d.undecimpunctatatenella;和d.u.undecimpunctatamannerheim。美国农业部目前估计玉米根虫每年造成10亿美元收入损失,包括8亿美元产量损失和2亿美元处理成本。
wcr和ncr两者在夏季期间在土壤中以卵沉积。该昆虫遍及整个冬季保持为卵阶段。卵是椭圆的、白色、且长度小于0.004英寸(0.010cm)。幼虫在5月下旬或6月上旬孵出,卵孵出的精确时机由于温度差异和位置而在各年间有所变化。新孵出的幼虫是白色的蠕虫,其长度小于0.125英寸(0.3175cm)。一旦孵出,幼虫便开始以玉米根为食。玉米根虫经历三个幼虫龄。在进食几周后,幼虫蜕皮,进入蛹阶段。它们在土壤中化蛹,然后它们在7月和8月中以成虫自土壤出现。成年根虫长度是约0.25英寸(0.635cm)。
玉米根虫幼虫在玉米和几种其它禾本科物种上完成发育。黄色狐尾草(yellowfoxtail)上饲养的幼虫较晚出现,并且作为成虫比玉米上饲养的幼虫具有更小的头壳尺寸。ellsbury等(2005)environ.entomol.34:627-34。wcr成虫以暴露的穗尖端的谷粒、玉米穗丝、和花粉为食。若wcr成虫在存在玉米生殖组织前出现,则它们可以以叶组织为食,由此减缓植物生长,且偶而杀死宿主植物。然而,成虫会在优选的穗丝和花粉变得可得到时快速向其转移。ncr成虫也以玉米植物的生殖组织为食,但是比较而言,很少以玉米叶为食。
玉米中的大部分根虫损伤通过幼虫进食引起。新孵出的根虫最初以较细的玉米根毛为食,并且钻入根尖中。随着幼虫生长得更大,它们以初生根为食并钻入其中。在富含玉米根虫时,幼虫进食经常导致自始至终向玉米杆基部修剪根。重度根损伤干扰根将水和营养物转运到植物中的能力,降低植物生长,并且导致降低的谷粒生产,由此经常显著降低总体产量。重度根损伤还经常导致玉米植物的倒伏,其使收获变得更难,而且进一步降低产量。此外,成虫以玉米生殖组织为食可以导致穗尖端的穗丝修剪。若此“穗丝剪切”在花粉脱落期间足够严重,则传粉可能受到破坏。
可以通过作物轮种、化学杀昆虫剂、生物杀虫剂(例如,孢子形成革兰氏阳性细菌苏云金芽孢杆菌(bacillusthuringiensis))、或其组合尝试控制玉米根虫。作物轮种遭受对农田的使用放置不想要的限制的重大缺点。此外,一些根虫物种的产卵可以在大豆田间发生,由此降低用玉米和大豆实施的作物轮种的效率。
化学杀昆虫剂最严重依赖于实现玉米根虫控制的策略。化学杀昆虫剂用途(尽管)是一种不完美的玉米根虫控制策略;在将化学杀昆虫剂的成本添加到尽管使用杀昆虫剂仍可以发生的根虫损伤的成本时在美国每年由于玉米根虫可能损失超过10亿美元。高幼虫群体、大雨、和不适当的杀昆虫剂应用均可以导致不充分的玉米根虫控制。此外,杀昆虫剂的连续使用可以选择杀昆虫剂抗性根虫品系,以及由于它们中许多对非靶物种的毒性而提高重大的环境忧虑。
rna干扰(rnai)是一种利用内源细胞途径的方法,由此对足够大小的整个或任何部分靶基因序列特异性的干扰rna(irna)分子(例如,dsrna分子)导致由其编码的mrna的降解。在最近几年中,已经使用rnai在许多物种和实验系统,例如线虫秀丽隐杆线虫(c.elegans)、植物、昆虫胚胎、和组织培养物中的细胞)实施基因“敲低”。参见例如fire等(1998)nature391:806-11;martinez等(2002)cell110:563-74;mcmanus和sharp(2002)naturerev.genetics3:737-47。
rnai经由内源途径,包括dicer蛋白质复合物实现mrna的降解。dicer将较长的dsrna分子切割成约20个核苷酸的短片段,称作小干扰rna(sirna)。sirna解开成两个单链rna:乘客链(passengerstrand)和引导链(guidestrand)。乘客链受到降解,而引导链掺入rna诱导的沉默复合物(risc)中。微小抑制性核糖核酸(mirna)分子可以类似地掺入risc中。转录后基因沉默在引导链特异性结合mrna分子的互补序列并且诱导argonaute(risc复合物的催化组分)切割时发生。尽管在一些真核生物诸如植物、线虫类和一些昆虫中sirna和/或mirna的浓度最初是有限的,但已知此过程遍及整个生物体系统性扩散。
仅与sirna和/或mirna互补的转录物受到切割和降解,如此mrna表达的敲低是序列特异性的。在植物中,存在dicer基因的几个功能性组。rnai的基因沉默效应持续数天,并且在实验条件下,可以导致靶定转录物的丰度下降90%或更多,随之发生相应蛋白质的水平降低。
美国专利7,612,194和美国专利公开文本no.us2007/0050860、us2010/0192265、和us2011/0154545披露了自玉米根萤叶甲蛹分离的9112种表达序列标签(est)序列的文库。在美国专利7,612,194和美国专利公开文本no.us2007/0050860中提出将核酸分子与启动子可操作连接以在植物细胞中表达反义rna,所述核酸分子与其中披露的玉米根萤叶甲液泡型h+-atp酶(v-atp酶)的几个特定部分序列之一互补。美国专利公开文本no.us2010/0192265提出将启动子与核酸分子可操作连接以在植物细胞中表达反义rna,所述核酸分子与未知的且未披露的功能的玉米根萤叶甲基因的特定部分序列互补(该部分序列叙述为与秀丽隐杆线虫中的c56c10.3基因产物是58%相同的)。美国专利公开文本no.us2011/0154545提出将启动子与核酸分子可操作连接以在植物细胞中表达反义rna,所述核酸分子与玉米根萤叶甲外被体beta亚基基因的两个特定部分序列互补。此外,美国专利7,943,819披露了自玉米根萤叶甲幼虫、蛹、和切开的中肠分离的906种表达序列标签(est)序列的文库,并且提出了将启动子与核酸分子可操作连接以在植物细胞中表达双链rna,所述核酸分子与玉米根萤叶带电(charged)多泡体蛋白4b基因的特定部分序列互补。
在美国专利7,612,194和美国专利公开文本no.us2007/0050860、us2010/0192265和us2011/0154545中没有进一步提出在v-atp酶的几种特定部分序列和未知功能的基因的特定部分序列之外使用其中列出的超过9000种序列的任何特定序列进行rna干扰。此外,美国专利7,612,194、和美国专利公开文本no.us2007/0050860和us2010/0192265、以及us2011/0154545无一提供关于提供的超过9000种中哪些其它序列在用作dsrna或sirna时在玉米根虫物种中会是致命,或即使在其它方面有用的任何指导。美国专利7,943,819没有提出在带电多泡体蛋白4b基因的特定部分序列外使用其中列出的超过900种序列的任何特定序列进行rna干扰。此外,美国专利7,943,819没有提供关于提供的超过900种中哪些其它序列在用作dsrna或sirna时在玉米根虫物种中会是致命,或即使在其它方面有用的指导。
与玉米根虫dna(诸如前述dna)互补的绝大多数序列在用作dsrna或sirna时在玉米根虫物种中不是致命的。例如,baum等(2007)描述了通过rnai抑制几种wcr基因靶物的效果。这些作者报告了他们测试的26种靶基因中的8种在超过520ng/cm2的非常高的irna(例如dsrna)浓度不能提供实验显著性鞘翅目有害生物死亡率。
发明概述
本文中公开了用于控制鞘翅目有害生物的核酸分子(例如,靶基因、dna、dsrna、sirna、mirna、和hprna)及其使用方法,所述鞘翅目有害生物包括例如玉米根萤叶甲(西部玉米根虫、“wcr”);北美玉米根萤叶甲(北部玉米根虫、“ncr”);d.u.howardibarber(南部玉米根虫、“scr”);d.v.zeaekrysanandsmith(墨西哥玉米根虫、“mcr”);d.balteataleconte;d.u.tenella;及d.u.undecimpunctatamannerheim。在具体的例子中,公开了例示性的核酸分子,其与鞘翅目有害生物中的一种或多种天然核酸序列的至少一部分可以是同源的。
在这些和其它例子中,天然核酸序列可以是靶基因,其产物可以是例如但不限于:牵涉代谢过程;牵涉生殖过程;或者牵涉幼虫发育。在一些例子中,通过包含与靶基因同源的序列的核酸分子对靶基因表达的翻译后抑制在鞘翅目有害生物中可以是致命的,或者导致降低的生长和/或生殖。在具体的例子中,至少一种选自下列的基因可以选择作为转录后沉默的靶基因:d_vir_c47185_caf1180;d_vir_c1229_vatpasec;d_vir_c1319_vatpaseh;和contig_01_rho1_1-191_cdc42。在具体的例子中,可用于翻译后抑制的靶基因是本文中称为d_vir_c47185_caf1180的新颖基因。因此,本文中公开了包含d_vir_c47185_caf1180(seqidno:1)的分离的核酸分子、其互补物、和任一者的片段。
还公开了包含核苷酸序列的核酸分子,所述核苷酸序列编码与靶基因产物(例如,选自下组的基因的产物:d_vir_c47185_caf1180;d_vir_c1229_vatpasec;d_vir_c1319_vatpaseh;和contig_01_rho1_1-191_cdc42))内的氨基酸序列至少85%相同的多肽。在具体的例子中,核酸分子包含编码多肽或其同系物的核苷酸序列,所述多肽与d_vir_c47185_caf1180产物内的氨基酸序列是至少85%相同的。进一步公开了包含核苷酸序列的核酸分子,所述核苷酸序列是编码与靶基因产物内的氨基酸序列至少85%相同的多肽的核苷酸序列的反向互补物。
还公开了cdna序列,其可以用于生成与整个或部分鞘翅目有害生物靶基因,例如:d_vir_c47185_caf1180;d_vir_c1229_vatpasec;d_vir_c1319_vatpaseh;和/或contig_01_rho1_1-191_cdc42互补的irna(例如,dsrna、sirna、mirna、和hprna)分子。在特定的实施方案中,dsrna、sirna、mirna、和/或hprna在体外或通过经遗传修饰的生物体,诸如植物和细菌在体内生成。在具体的例子中,公开了cdna分子,其可以用于生成与整个或部分的d_vir_c47185_caf1180互补的irna分子。
进一步公开了用于抑制鞘翅目有害生物中必需基因的表达的手段,和用于对植物提供鞘翅目有害生物抗性的手段。用于抑制鞘翅目有害生物中必需基因表达的手段是单或双链rna分子,其由seqidno:7、8、10、11、49、50、86、87、91、92、93、94、95、96、97、98、99、100、101、102、103、104、105、106、107、108、109、110、111、112、113、114、115、116、127、128、129、130、131之任一,或其互补物组成。用于对植物提供鞘翅目有害生物抗性的手段是dna分子,其包含与启动子可操作连接的编码用于抑制鞘翅目有害生物中必需基因表达的手段的核酸序列,其中所述dna分子能够整合入玉米植物的基因组中。
公开了用于控制鞘翅目有害生物群体的方法,包括对鞘翅目有害生物提供irna(例如,dsrna、sirna、mirna、和hprna)分子,其在被鞘翅目有害生物摄取后发挥功能以抑制鞘翅目有害生物内的生物学功能,其中irna分子包含整个或部分的选自下组的核苷酸序列:seqidno:1;seqidno:1的互补物;seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;根萤叶甲属生物体(例如,wcr)的天然编码序列,其包含整个或部分的seqidno:1-4之任一;根萤叶甲属生物体的天然编码序列的互补物,所述天然编码序列包含整个或部分的seqidno:1-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含整个或部分的seqidno:1-4之任一;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含整个或部分的seqidno:1-4之任一。
在具体的例子中,公开了用于控制鞘翅目有害生物群体的方法,包括对鞘翅目有害生物提供irna(例如dsrna、sirna、mirna、和hprna)分子,其被鞘翅目有害生物摄取后发挥功能以抑制鞘翅目有害生物内的生物学功能,其中irna分子包含选自下组的核苷酸序列:整个或部分的seqidno:1;整个或部分的seqidno:1的互补物;seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;包含seqidno:1的根萤叶甲属生物体(例如wcr)的天然编码序列的整个或部分;包含seqidno:1的根萤叶甲属生物体的天然编码序列的整个或部分互补物;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的整个或部分,所述天然非编码序列包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的整个或部分互补物,所述天然非编码序列包含seqidno:1。
本文中还公开了如下的方法,其中可以在基于饮食的测定法中,或在表达dsrna、sirna、mirna和/或hprna的经遗传修饰的植物细胞中对鞘翅目有害生物提供dsrna、sirna、mirna、和/或hprna。在这些和其它例子中,鞘翅目有害生物幼虫可以摄取dsrna、sirna、mirna、和/或hprna。然后,本发明的dsrna、sirna、mirna、和/或hprna的摄取可以导致幼虫中的rnai,其继而可以导致对于鞘翅目有害生物的存活力必需的基因沉默,最终导致幼虫死亡率。如此,公开了如下的方法,其中对鞘翅目有害生物提供核酸分子,其包含可用于控制鞘翅目有害生物的例示性核酸序列。在具体的例子中,通过使用本发明的核酸分子控制的鞘翅目有害生物可以是wcr或ncr。
更具体地,本申请提供下述各项:
1.一种分离的多核苷酸,其包含至少一种选自下组的核苷酸序列:seqidno:1;seqidno:1的互补物;seqidno:1的至少19个连续核苷酸的片段;seqidno:1的至少19个连续核苷酸的片段的互补物;包含seqidno:1的根萤叶甲属生物体的天然编码序列;根萤叶甲属生物体的天然编码序列的互补物,所述天然编码序列包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,其包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,其包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,其包含seqidno:1。
2.项1的多核苷酸,其进一步包含至少一种选自下组的核苷酸序列:seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;seqidno:2-4之任一的至少19个连续核苷酸的片段;seqidno:2-4之任一的至少19个连续核苷酸的片段的互补物;根萤叶甲属生物体的天然编码序列,其包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,其包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,其包含seqidno:2-4之任一;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,其包含seqidno:2-4之任一。
3.项1的多核苷酸,其中所述至少一种核苷酸序列与异源启动子可操作连接。
4.项1的多核苷酸,其中所述多核苷酸选自下组:seqidno:49、seqidno:86、seqidno:91、seqidno:92、seqidno:95、seqidno:96、seqidno:97、和seqidno:110。
5.项1的多核苷酸,其中所述多核苷酸的长度是至少约50个核苷酸。
6.项1的多核苷酸,其中所述多核苷酸的长度是至少约100个核苷酸。
7.项6的多核苷酸,其中所述多核苷酸的长度是约100个核苷酸。
8.一种植物转化载体,其包含项1的多核苷酸。
9.项1的多核苷酸,其中所述根萤叶甲属生物体选自下组:玉米根萤叶甲(d.v.virgiferaleconte);北美玉米根萤叶甲(d.barberismithandlawrence);d.u.howardi;d.v.zeae;d.balteataleconte;d.u.tenella;和d.u.undecimpunctatamannerheim。
10.项1的多核苷酸,其中所述多核苷酸是核糖核酸(rna)分子。
11.项1的多核苷酸,其中所述多核苷酸是脱氧核糖核酸(dna)分子。
12.项11的多核苷酸,其中所述多核苷酸包含seqidno:136。
13.一种双链核糖核酸分子,其由表达项11的多核苷酸生成。
14.一种双链核糖核酸分子,其由表达项12的多核苷酸生成。
15.项13的双链核糖核酸分子,其中使所述多核苷酸序列与鞘翅目有害生物接触抑制与项1的多核苷酸序列特异性互补的内源核苷酸序列表达。
16.项15的双链核糖核酸分子,其中使所述核糖核苷酸分子与鞘翅目有害生物接触杀死或抑制所述鞘翅目有害生物的生长、繁殖、和/或进食。
17.项13的双链核糖核酸分子,其包含第一、第二和第三多核苷酸序列,其中所述第一多核苷酸序列包含项1的多核苷酸,其中所述第三多核苷酸序列通过所述第二多核苷酸序列与所述第一多核苷酸序列连接,且其中所述第三多核苷酸序列基本上是所述第一多核苷酸序列的反向互补物,使得所述第一和所述第三多核苷酸序列在转录成核糖核酸时杂交以形成所述双链核糖核苷酸分子。
18.项17的双链核糖核酸分子,其中所述第二多核苷酸序列包含seqidno:136。
19.项10的多核苷酸,其选自下组:长度介于约50和约300个核苷酸之间的双链核糖核酸分子和单链核糖核酸分子。
20.一种核糖核酸分子,其通过表达项11的多核苷酸生成,其中所述核糖核酸分子选自下组:长度介于约50和约300个核苷酸之间的双链核糖核酸分子和单链核糖核酸分子。
21.一种植物转化载体,其包含项1的多核苷酸,其中所述至少一种核苷酸序列与植物细胞中功能性的异源启动子可操作连接。
22.一种用项1的多核苷酸转化的细胞。
23.项22的细胞,其中所述细胞是原核细胞。
24.项22的细胞,其中所述细胞是真核细胞。
25.项24的细胞,其中所述细胞是植物细胞。
26.一种用项1的多核苷酸转化的植物。
27.项26的植物的种子,其中所述种子包含所述多核苷酸。
28.项26的植物,其中所述至少一种核苷酸序列在所述植物中以双链核糖核酸分子表达。
29.项25的细胞,其中所述细胞是玉蜀黍(zeamays)细胞。
30.项26的植物,其中所述植物是玉蜀黍。
31.项26的植物,其中所述至少一种核苷酸序列在所述植物中以核糖核酸分子表达,且所述核糖核酸分子在鞘翅目有害生物摄取所述植物的一部分时抑制与所述至少一种核苷酸序列特异性互补的内源鞘翅目有害生物核苷酸序列表达。
32.项1的多核苷酸,其包含超过一种选自下组的核苷酸序列:seqidno:1;seqidno:1的互补物;seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;seqidno:1-4之任一的至少19个连续核苷酸的片段;seqidno:1-4之任一的至少19个连续核苷酸的片段的互补物;根萤叶甲属生物体的天然编码序列,其包含seqidno:1-4之任一;根萤叶甲属生物体的天然编码序列的互补物,所述天然编码序列包含seqidno:1-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1-4之任一;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,其包含seqidno:1-4之任一。
33.一种植物转化载体,其包含项32的多核苷酸,其中所述核苷酸序列各与植物细胞中功能性的异源启动子可操作连接。
34.一种用项32的多核苷酸转化的细胞。
35.一种用项32的多核苷酸转化的植物。
36.项35的植物,其中所述超过一种核苷酸序列在所述植物细胞中以双链核糖核酸分子表达。
37.项34的细胞,其中所述细胞是玉蜀黍细胞。
38.项36的植物,其中所述植物是玉蜀黍。
39.项31的植物,其中所述鞘翅目有害生物是根萤叶甲属物种。
40.项39的植物,其中所述鞘翅目有害生物选自下组:玉米根萤叶甲(d.virgiferavirgiferaleconte)、北美玉米根萤叶甲(d.barberismithandlawrence)、d.virgiferazeaekrysanandsmith、d.undecimpunctatahowardibarber、d.balteataleconte、d.undecimpunctatatenella、和d.undecimpunctataundecimpunctatamannerheim。
41.一种自项30的植物生成的商品产品,其中所述商品产品包含可检测量的项1的多核苷酸。
42.一种用于控制鞘翅目有害生物群体的方法,包括提供包含双链核糖核酸分子的制剂,其在与所述鞘翅目有害生物接触后发挥功能以抑制所述鞘翅目有害生物内的生物学功能,其中所述制剂包含选自下组的核苷酸序列:seqidno:1;seqidno:1的互补物;seqidno:1的至少19个连续核苷酸的片段;seqidno:1的至少19个连续核苷酸的片段的互补物;包含seqidno:1的根萤叶甲属生物体的天然编码序列;包含seqidno:1的根萤叶甲属生物体的天然编码序列的互补物;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,其包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,其包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,其包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,其包含seqidno:1。
43.一种用于控制鞘翅目有害生物群体的方法,包括提供包含第一和第二多核苷酸序列的制剂,其在与所述鞘翅目有害生物接触后发挥功能以抑制所述鞘翅目有害生物内的生物学功能,其中所述第一多核苷酸序列包含与seqidno:1的约19至约30个连续核苷酸展现出约90%至约100%序列同一性的区域,且其中所述第一多核苷酸序列与所述第二多核苷酸序列特异性杂交。
44.依照项43的方法,其中所述第一多核苷酸序列进一步包含与选自下组的鞘翅目有害生物基因的约19至约30个连续核苷酸展现出约90%至约100%序列同一性的区域:液泡atp酶的c亚基;液泡atp酶的h亚基;和rho1小gtp结合蛋白。
45.一种用于控制鞘翅目有害生物群体的方法,该方法包括:
在鞘翅目有害生物的宿主植物中提供包含项1的多核苷酸的经转化的植物细胞,其中表达所述多核苷酸以生成核糖核酸分子,该核糖核酸分子在与属于该群体的鞘翅目有害生物接触后发挥功能以抑制所述鞘翅目有害生物内靶序列表达,并且相对于缺乏所述经转化的植物细胞的相同物种的宿主植物上的生长,导致所述鞘翅目有害生物或鞘翅目有害生物群体的生长降低。
46.依照项45的方法,其中所述核糖核酸分子是双链核糖核酸分子。
47.依照项45的方法,其中所述鞘翅目有害生物群体相对于侵袭缺乏所述经转化的植物细胞的相同物种的宿主植物的鞘翅目有害生物群体是减少的。
48.一种用于控制植物中植物鞘翅目有害生物侵袭的方法,包括在鞘翅目有害生物的饮食中提供项1的多核苷酸。
49.依照项48的方法,其中所述饮食包含经过转化以表达项1的多核苷酸的植物细胞。
50.一种用于改善玉米作物产量的方法,该方法包括:
将项1的多核苷酸导入玉米植物中以生成转基因玉米植物;并
栽培所述玉米植物以容许所述至少一种核苷酸序列的表达;其中所述至少一种核苷酸序列的表达抑制鞘翅目有害生物感染或生长和由于鞘翅目有害生物感染所致的产量损失。
51.依照项50的方法,其中所述至少一种核苷酸序列的表达生成rna分子,其抑制已经接触所述玉米植物的一部分的鞘翅目有害生物中的至少第一靶基因。
52.一种用于生成转基因植物细胞的方法:
用载体转化植物细胞,所述载体包含与启动子和转录终止序列可操作连接的项1的多核苷酸;
在如下的条件下培养所述经转化的植物细胞,所述条件足以容许形成包含多个经转化的植物细胞的植物细胞培养物;
选择经转化的植物细胞,该经转化的植物细胞已经将所述至少一种核苷酸序列整合入其基因组中;
对所述经转化的植物细胞筛选由所述至少一种核苷酸序列编码的核糖核酸分子表达;并
选择表达dsrna的植物细胞。
53.一种用于生成鞘翅目有害生物抗性转基因植物的方法,该方法包括:
提供通过依照项52的方法生成的转基因植物细胞;并
从所述转基因植物细胞再生转基因植物,其中由所述至少一种核苷酸序列编码的核糖核酸分子的表达足以调控接触所述经转化植物的鞘翅目有害生物中靶基因的表达。
54.一种用于生成转基因植物细胞的方法:
用包含多核苷酸的载体转化植物细胞,所述多核苷酸编码用于抑制鞘翅目有害生物中必需基因表达的手段,其中所述多核苷酸与启动子和转录终止序列可操作连接;
在如下的条件下培养经转化的植物细胞,所述条件足以容许包含多个经转化的植物细胞的植物细胞培养物的形成;
选择已经将所述多核苷酸整合入其基因组中的经转化的植物细胞;
对所述经转化的植物细胞筛选所述用于抑制鞘翅目有害生物中必需基因表达的手段的表达;并
选择表达所述用于抑制鞘翅目有害生物中必需基因表达的手段的植物细胞。
55.依照项54的方法,其中所述多核苷酸选自下组:seqidno:49、seqidno:86、seqidno:91、seqidno:92、seqidno:95、seqidno:96、seqidno:97、和seqidno:110。
56.一种植物细胞,其包含用于对植物提供鞘翅目有害生物抗性的手段。
57.项56的植物细胞,其中所述用于对植物提供鞘翅目有害生物抗性的手段是选自下组的多核苷酸:seqidno:49、seqidno:86、seqidno:91、seqidno:92、seqidno:95、seqidno:96、seqidno:97、和seqidno:110。
58.一种分离的核酸,其包含至少一种与选自下组的多核苷酸基本上同源的多核苷酸:seqidno:1;seqidno:1的至少19个连续核苷酸的片段;根萤叶甲属生物体的天然编码序列,其包含seqidno:1;和根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1。
59.项58的分离的核酸,其中所述多核苷酸与选自下组的多核苷酸是至少80%相同的:seqidno:1;seqidno:1的至少19个连续核苷酸的片段;根萤叶甲属生物体的天然编码序列,其包含seqidno:1;和根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1。
60.项58的分离的核酸,其中所述多核苷酸与选自下组的多核苷酸是至少90%相同的:seqidno:1;seqidno:1的至少19个连续核苷酸的片段;根萤叶甲属生物体的天然编码序列,其包含seqidno:1;和根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1。
61.项58的分离的核酸,其中所述多核苷酸与选自下组的多核苷酸是至少95%相同的:seqidno:1;seqidno:1的至少19个连续核苷酸的片段;根萤叶甲属生物体的天然编码序列,其包含seqidno:1;和根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1。
62.项58的分离的核酸,其中seqidno:1在中等严格性条件下与所述至少一种多核苷酸杂交。
63.项58的分离的核酸,其中seqidno:1在高严格性条件下与所述至少一种多核苷酸杂交。
64.一种植物细胞、植物组织、植物部分、或植物,其包含项58的核酸。
前述和其它特征从几个实施方案的以下详细描述看会变得更为显而易见,所述实施方案参照附图进行。
附图简述
图1包括用于提供dsrna生成的特异性模板的策略的描绘。
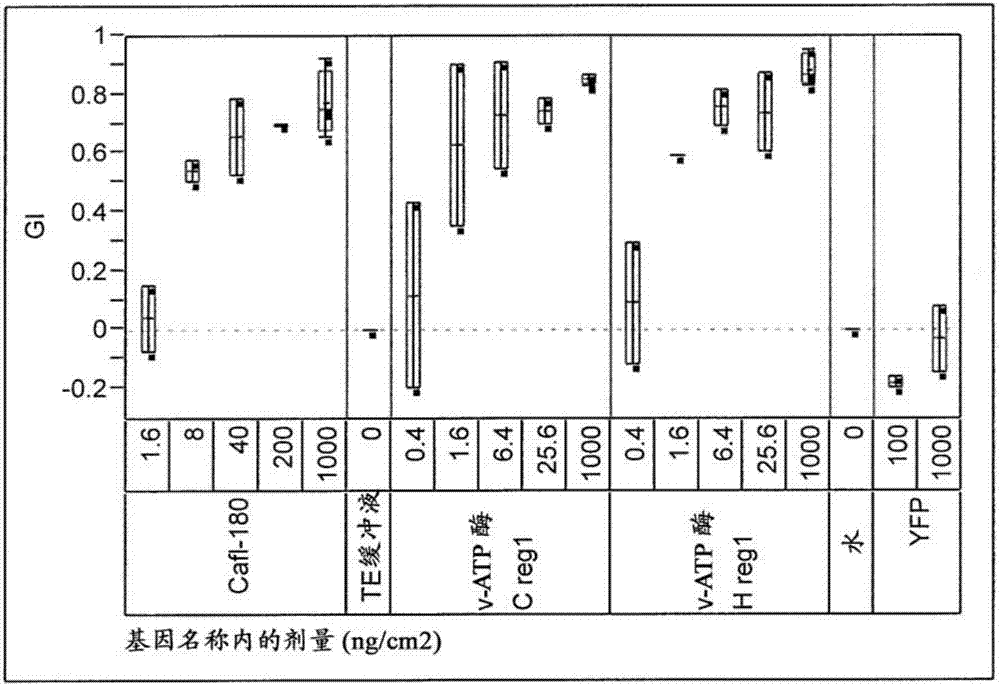
图2包括用例示性核酸分子处理的鞘翅目有害生物的生长抑制的变化性图。
图3包括用例示性核酸分子处理的鞘翅目有害生物的死亡率的变化性图。
图4包括用例示性核酸分子处理的鞘翅目有害生物的生长抑制的别的变化性图。
图5包括可以存在于本发明的某些核酸分子中的几种核苷酸序列的草图描绘。在标示的情况中,“(序列)”指具有任何长度和特定序列的描绘核苷酸序列的区域,例如但不限于seqidno:1-4之一的连续区段。描绘核苷酸序列的斜体化区域是任选的。
序列表
所附序列表中所列的核酸序列使用核苷酸碱基的标准字母缩写显示,如37c.f.r.§1.822中显示的。仅显示每个核酸序列的一条链,但是互补链和反向互补链理解为通过对展示链的任何参考而包括在内。在所附序列表中,
seqidno:1显示了例示性根萤叶甲属cdna序列,在一些地方中称为d_vir_c47185_caf1180,或caf1-180。
seqidno:2显示了例示性根萤叶甲属cdna序列,在一些地方中称为d_vir_c1229_vatpasec,或vatpasec。
seqidno:3显示了例示性根萤叶甲属cdna序列,在一些地方中称为d_vir_c1319_vatpaseh,或vatpaseh。
seqidno:4显示了例示性根萤叶甲属cdna序列,在一些地方中称为contig_01_rho1_1-191_cdc42,或rho1。
seqidno:5-8显示了根萤叶甲属液泡atp酶c亚基cdna的例示性非连续片段。
seqidno:9-11显示了根萤叶甲属液泡atp酶h亚基cdna的例示性非连续片段。
seqidno:12显示了t7噬菌体启动子序列。
seqidno:13-38显示了用于通过pcr扩增例示性靶基因的编码区的各部分的引物。
seqidno:39-48显示了用于扩增caf1-180和vatpasec的基因区以合成发夹rna的引物。
seqidno:49显示了含有st-ls1内含子(seqidno:136)的caf1-180发夹rna形成dna序列,用于含有玉米共有序列的型式1表达载体。
seqidno:50显示了含有st-ls1内含子(seqidno:136)的vatpasec发夹rna形成dna序列,用于含有玉米共有序列的型式1hprna表达载体。
seqidno:51显示了膜联蛋白区1的dna序列。
seqidno:52显示了膜联蛋白区2的dna序列。
seqidno:53显示了beta血影蛋白2区1的dna序列。
seqidno:54显示了beta血影蛋白2区2的dna序列。
seqidno:55显示了mtrp-l4区1的dna序列。
seqidno:56显示了mtrp-l4区2的dna序列。
seqidno:57显示了编码yfp的dna序列。
seqidno:58-85显示了用于扩增膜联蛋白、beta血影蛋白2、mtrp-l4、和yfp的基因区以合成dsrna的引物。
seqidno:86显示了d_vir_c47185_caf1-180cdna的例示性260bp扩增片段,其用作合成dsrna分子的模板。
seqidno:87-90显示了根萤叶甲属rho1cdna的例示性非连续片段。
seqidno:91显示了含有st-ls1内含子(seqidno:136)(包括在内含子侧翼的任选克隆位点)的例示性caf1-180发夹rna形成dna序列,用于含有玉米共有序列的型式1表达载体。
seqidno:92显示了含有st-ls1内含子(seqidno:136)的例示性caf1-180发夹rna形成dna序列,用于不含玉米共有序列的型式2表达载体。
seqidno:93显示了含有st-ls1内含子(seqidno:136)(包括在内含子侧翼的任选克隆位点)的例示性vatpasec发夹rna形成dna序列,用于含有玉米共有序列的型式1hprna表达载体。
seqidno:94显示了含有st-ls1内含子(seqidno:136)的例示性vatpasec发夹rna形成dna序列,用于不含玉米共有序列的型式2hprna表达载体。
seqidno:95显示了caf1-180dna有义链中含有st-ls1内含子(seqidno:136)的例示性区段,用于型式3表达载体。
seqidno:96显示了caf1-180dna反义链的例示性区段,用于型式3表达载体。
seqidno:97显示了含有st-ls1内含子(seqidno:136)的例示性caf1-180发夹rna形成dna序列,用于型式3表达载体。
seqidno:98显示了vatpasecdna有义链中含有st-ls1内含子(seqidno:136)的例示性区段,用于型式3表达载体。
seqidno:99显示了vatpasecdna反义链的例示性区段,用于型式3表达载体。
seqidno:100显示了含有st-ls1内含子(seqidno:136)的例示性vatpasec发夹rna形成dna序列,用于型式3表达载体。
seqidno:101显示了vatpasecdna有义链中含有st-ls1内含子(seqidno:136)的例示性区段,用于型式4表达载体。
seqidno:102显示了vatpasecdna反义链的例示性区段,用于型式4表达载体。
seqidno:103显示了含有st-ls1内含子(seqidno:136)的例示性vatpasec发夹rna形成dna序列,用于型式4表达载体。
seqidno:104显示了vatpasehdna有义链中含有st-ls1内含子(seqidno:136)的例示性区段,用于型式1表达载体。
seqidno:105显示了vatpasehdna反义链的例示性区段,用于型式1表达载体。
seqidno:106显示了含有st-ls1内含子(seqidno:136)的例示性vatpaseh发夹rna形成dna序列,用于型式1表达载体。
seqidno:107显示了rho1dna有义链中含有st-ls1内含子(seqidno:136)的例示性区段,用于型式1表达载体。
seqidno:108显示了rho1dna反义链的例示性区段,用于型式1表达载体。
seqidno:109显示了含有st-ls1内含子(seqidno:136)的例示性rho1发夹rna形成dna序列,用于型式1表达载体。
seqidno:110显示了caf-180dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:111显示了vatpasec区1dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:112显示了vatpasec区1(短)dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:113显示了vatpasec区2dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:114显示了vatpaseh区1dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:115显示了vatpaseh区2dna中用于在饮食喂养生物测定法中提供dsrna合成的模板的例示性区段。
seqidno:116显示了rho1dna中用于在饮食喂养生物测定法中提供dsrna合成模板的例示性区段。
seqidno:117显示了在饮食喂养生物测定法中用作dsrna的例示性vatpasecrna有义链(“vatpasec5’-15”)。
seqidno:118显示了在饮食喂养生物测定法中用作dsrna的别的例示性vatpasecrna有义链(“vatpasec5’-25”)。
seqidno:119显示了在饮食喂养生物测定法中用作dsrna的别的例示性vatpasecrna有义链(“vatpasec3’-15”)。
seqidno:120显示了在饮食喂养生物测定法中用作dsrna的别的例示性vatpasecrna有义链(“vatpasec3’-25”)。
seqidno:121-126显示了用于扩增根萤叶甲属vatpasec基因中作为dsrna合成模板的部分的引物。
seqidno:127显示了在饮食喂养生物测定法中用作dsrna模板的例示性vatpasec有义链(“vatpasec5’-50”)。
seqidno:128显示了在饮食喂养生物测定法中用作dsrna模板的例示性vatpasec有义链(“vatpasec5’-100”)。
seqidno:129显示了在饮食喂养生物测定法中用作dsrna模板的例示性vatpasec有义链(“vatpasec-174”)。
seqidno:130显示了在饮食喂养生物测定法中用作dsrna模板的例示性vatpasec有义链(“vatpasec3’-50”)。
seqidno:131显示了在饮食喂养生物测定法中用作dsrna的例示性vatpasec有义链(“vatpasec3’-100”)。
seqidno:132-135显示了用于转基因玉米的分子分析的引物。
seqidno:136显示了在一些实施方案中可用于形成发夹rna的st-ls1内含子。
seqidno:137-138显示了根萤叶甲属rho1cdna的例示性非连续片段。
发明详述
i.几个实施方案的概述
本文中公开了用于遗传控制鞘翅目有害生物侵袭的方法和组合物。还提供了用于鉴定一种或多种对于鞘翅目有害生物的生活史而言必需的基因以用作rnai介导的鞘翅目有害生物群体控制的靶基因的方法。编码dsrna分子的dna质粒载体可以设计为抑制一种或多种对于生长、存活、发育和/或生殖必需的靶基因。在一些实施方案中,提供了经由与鞘翅目有害生物中靶基因的编码或非编码序列互补的核酸分子来转录后阻抑靶基因表达或抑制靶基因的方法。在这些和其它实施方案中,鞘翅目有害生物可以摄取自与靶基因的编码或非编码序列互补的整个或部分核酸分子转录的一种或多种dsrna、sirna、mirna、和/或hprna分子,由此提供植物保护性效果。
如此,一些实施方案牵涉使用与靶基因的编码和/或非编码序列互补以至少实现对鞘翅目有害生物的部分控制的dsrna、sirna、mirna、和/或hprna对靶基因产物表达的序列特异性抑制。公开了一组分离的且纯化的核酸分子,其包含例如如seqidno:1-4之一中列出的核苷酸序列及其片段。在一些实施方案中,可以从这些序列、其片段、或包含这些序列之一的基因表达稳定化的dsrna分子以转录后沉默或抑制靶基因。在某些实施方案中,分离的且纯化的核酸分子包含整个或部分的seqidno:1。
一些实施方案牵涉在其基因组中具有编码至少一种irna(例如dsrna)分子的至少一种重组dna序列的重组宿主细胞(例如植物细胞)。在特定的实施方案中,dsrna分子可以在被鞘翅目有害生物摄取时表达以转录后沉默或抑制鞘翅目有害生物中靶基因的表达。重组dna序列可以包含例如seqid1-4之任一,seqid1-4之任一的片段,或包含seqid1-4之一的基因的部分序列,或其互补物。
特定的实施方案牵涉在其基因组中具有编码至少一种包含整个或部分的seqidno:1的irna(例如dsrna)分子的重组dna序列的重组宿主细胞。在被鞘翅目有害生物摄取时,irna分子可以沉默或抑制鞘翅目有害生物中包含seqidno:1的靶基因的表达,由此导致鞘翅目有害生物中生长、发育、生殖、和/或进食的停止。
在一些实施方案中,在其基因组中具有编码至少一种dsrna分子的至少一种重组dna序列的重组宿主细胞可以是经转化的植物细胞。一些实施方案牵涉包含此类经转化的植物细胞的转基因植物。在此类转基因植物外,任何转基因植物世代的后代植物、转基因种子、和转基因植物产物都有提供,其中每种包含重组dna序列。在特定的实施方案中,可以在转基因植物细胞中表达本发明的dsrna分子。因此,在这些和其它实施方案中,可以从转基因植物细胞分离本发明的dsrna分子。在特定的实施方案中,转基因植物是选自下组的植物:玉米(玉蜀黍(zeamays))、大豆(glycinemax)、和科禾本科(poaceae)的植物。
一些实施方案牵涉用于调控鞘翅目有害生物细胞中靶基因表达的方法。在这些和其它实施方案中,可以提供核酸分子,其中核酸分子包含编码dsrna分子的核苷酸序列。在特定的实施方案中,编码dsrna分子的核苷酸序列可以与启动子可操作连接,并且还可以与转录终止序列可操作连接。在特定的实施方案中,用于调控鞘翅目有害生物细胞中靶基因表达的方法可以包括:(a)用包含编码dsrna分子的核苷酸序列的载体转化植物细胞;(b)在足以容许形成包含多个经转化的植物细胞的植物细胞培养物的条件下培养经转化的植物细胞;(c)选择已经将载体整合入其基因组中的经转化的植物细胞;并(d)确定选择的经转化的植物细胞包含由载体的核苷酸序列编码的dsrna分子。可以从具有其基因组中整合的载体且包含由载体的核苷酸序列编码的dsrna分子的植物细胞再生植物。
如此,还公开了包含其基因组中整合的载体的转基因植物,所述载体具有编码dsrna分子的核苷酸序列,其中转基因植物包含由载体的核苷酸序列编码的dsrna分子。在特定的实施方案中,植物中dsrna分子的表达足以调控鞘翅目有害生物细胞中靶基因的表达,所述鞘翅目有害生物例如通过以经转化的植物、植物的一部分(例如根)或植物细胞为食而接触经转化的植物或植物细胞。本文中公开的转基因植物可以展现出对鞘翅目有害生物侵袭的抗性和/或增强的耐受性。特定的转基因植物可以展现出对一种或多种选自下组的鞘翅目有害生物的抗性和/或增强的耐受性:wcr;ncr;scr;mcr;d.balteataleconte;d.u.tenella;和d.u.undecimpunctatamannerheim。
本文中还公开了用于将控制剂,诸如irna分子投递至鞘翅目有害生物的方法。此类控制剂可以直接或间接引起对鞘翅目有害生物进食、生长能力的损伤或在其它方面引起对宿主的损伤。在一些实施方案中,提供了如下的方法,包括将稳定化的dsrna分子投递至鞘翅目有害生物以阻抑鞘翅目有害生物中的至少一种靶基因,由此降低或消除鞘翅目有害生物的植物损伤。在一些实施方案中,抑制鞘翅目有害生物中靶基因表达的方法可以导致鞘翅目有害生物中生长、发育、生殖和/或进食的停止。在一些实施方案中,所述方法最终可以导致鞘翅目有害生物死亡。
在一些实施方案中,提供了包含本发明的irna(例如dsrna)分子的组合物(例如,表面组合物),其在植物、动物、和/或植物或动物的环境中使用以实现鞘翅目有害生物侵袭的消除或降低。在特定的实施方案中,组合物可以是要对鞘翅目有害生物喂养的营养组合物或食物来源。一些实施方案包括使营养组合物或食物来源对于鞘翅目有害生物可得到。包含irna分子的组合物的摄入可以导致一个或多个鞘翅目有害生物细胞摄取该分子,这继而可以导致对鞘翅目有害生物细胞中至少一种靶基因表达的抑制。可以通过在鞘翅目有害生物宿主中提供一种或多种包含本发明的irna分子的组合物在存在鞘翅目有害生物的任何宿主组织或环境之中或之上限制或消除鞘翅目有害生物对植物或植物细胞的摄入或损伤。
本文中公开的组合物和方法可以与用于控制鞘翅目有害生物损害的其它方法和组合物组合一起使用。例如,可以在如下的方法中使用用于针对鞘翅目有害生物保护植物的如本文中所描述的irna分子,所述方法包括另外使用一种或多种针对鞘翅目有害生物有效的化学剂、针对鞘翅目有害生物有效的生物杀虫剂、作物轮种、或重组遗传技术,其展现出与本发明的rnai介导的方法和rnai组合物的特征不同的特征(例如,在植物中重组生成对于鞘翅目有害生物有害的蛋白质(例如,bt毒素))。
ii.缩写
dsrna双链核糖核酸
gi生长抑制
ncbi国立生物技术信息中心
gdna基因组dna
irna抑制性核糖核酸
orf可读框
rnai核糖核酸干扰
mirna微小抑制性核糖核酸
sirna小抑制性核糖核酸
hprna发夹核糖核酸
utr非翻译区
wcr西部玉米根虫(玉米根萤叶甲)
ncr北部玉米根虫(北美玉米根萤叶甲)
mcr墨西哥玉米根虫(diabroticavirgiferazeaekrysanandsmith)
pcr聚合酶链式反应
riscrna诱导的沉默复合物
scr南部玉米根虫(diabroticaundecimpunctatahowardibarber)
iii.术语
在以下描述和表中,使用许多术语。为了提供对说明书和权利要求书的清楚且一致的理解,包括要给予此类术语的范围,提供以下定义:
鞘翅目有害生物:如本文中所使用的,术语“鞘翅目有害生物”指以玉米和其它真正的禾本科为食的属根萤叶甲属昆虫。在具体的例子中,鞘翅目有害生物选自下组:玉米根萤叶甲(wcr);北美玉米根萤叶甲(ncr);d.u.howardi(scr);d.v.zeae(mcr);d.balteataleconte;d.u.tenella;和d.u.undecimpunctatamannerheim。
(与生物体)接触:如本文中所使用的,术语与生物体(例如鞘翅目有害生物)“接触”或被生物体“摄取”就核酸分子而言包括核酸分子内在化入生物体中,例如但不限于:生物体摄取分子(例如,通过进食进行);使生物体与包含核酸分子的组合物接触;以及用包含核酸分子的溶液浸泡生物体。
重叠群:如本文中所使用的,术语“重叠群”指自从单一遗传来源衍生的一组重叠dna区段重建的dna序列。
玉米植物:如本文中所使用的,术语“玉米植物”指物种玉蜀黍(玉米)的植物。
表达:如本文中所使用的,编码序列(例如,基因或转基因)的“表达”指核酸转录单元(包括例如基因组dna或cdna)的编码信息转化成细胞的操作、非操作、或结构部分的过程,经常包括蛋白质的合成。基因表达可以受到外部信号;例如细胞、组织或生物体对提高或降低基因表达的制剂的暴露影响。也可以在从dna至rna至蛋白质的途径中的任何地方调节基因的表达。例如,经由对转录、翻译、rna转运和加工、对中间分子诸如mrna的降解起作用的控制,或者经由活化、失活、区室化、或在已经生成特定蛋白质分子后对其的降解,或者通过其组合发生基因表达的调节。可以通过本领域中已知的任何方法,包括但不限于northern印迹、rt-pcr、western印迹、或体外、原位、或体内蛋白质活性测定法在rna水平或蛋白质水平上测量基因表达。
遗传材料:如本文中所使用的,术语“遗传材料”包括所有基因、和核酸分子,诸如dna和rna。
抑制:如本文中所使用的,术语“抑制”在用于描述对编码序列(例如基因)的影响时指自编码序列转录的mrna和/或编码序列的肽、多肽、或蛋白质产物的细胞水平的可测量降低。在一些例子中,可以抑制编码序列的表达,使得大致消除表达。“特异性抑制”指在没有因此影响实现特异性抑制的细胞中其它编码序列(例如基因)表达的情况下对靶编码序列的抑制。
分离的:“分离的”生物体组分(诸如核酸或蛋白质)已经与该组分天然存在的生物体细胞中的其它生物组分,即,其它染色体和染色体外dna和rna及蛋白质基本上分开、离开其生成、远离其纯化。已经“分离的”核酸分子和蛋白质包括通过标准纯化方法纯化的核酸分子和蛋白质。该术语还包括通过宿主细胞中的重组表达制备的核酸和蛋白质以及化学合成的核酸分子、蛋白质和肽。
核酸分子:如本文中所使用的,术语“核酸分子”可以指核苷酸的聚合形式,其可以包括rna的有义和反义链两者、cdna、基因组dna、和合成形式及上述物质的混合聚合物。核苷酸可以指核糖核苷酸、脱氧核糖核苷酸、或任一类型的核苷酸的经修饰形式。如本文中所使用的,“核酸分子”与“核酸”和“多核苷酸”是同义的。除非另有规定,核酸分子通常是至少10个碱基的长度。根据惯例,核酸分子的核苷酸序列从分子的5’至3’端阅读。核苷酸序列的“互补物”指与核苷酸序列的核碱基形成碱基对(即a-t/u,和g-c)的核碱基的5’至3’序列。核酸序列的“反向互补物”指与核苷酸序列的核碱基形成碱基对的核碱基的3’至5’序列。
“核酸分子”包括dna的单链和双链形式;rna的单链形式;和rna的双链形式(dsrna)。术语“核苷酸序列”或“核酸序列”指作为个别单链或在双链体中的核酸有义和反义链两者。术语“核糖核酸”(rna)包括irna(抑制性rna)、dsrna(双链rna)、sirna(小干扰rna)、mrna(信使rna)、mirna(微小rna)、hprna(发夹rna)、trna(转移rna)(无论是装载还是卸下相应的酰化氨基酸)、和crna(互补rna)。术语“脱氧核糖核酸”(dna)包括cdna、基因组dna、和dna-rna杂合物。术语“核酸区段”和“核苷酸序列区段”,或更一般地“区段”会被本领域技术人员理解为功能术语,其包含基因组序列、核糖体rna序列、转移rna序列、信使rna序列、操纵子序列、和较小的工程化核苷酸序列,其编码或可以改编为编码肽、多肽、或蛋白质。
寡核苷酸:寡核苷酸是一种短的核酸聚合物。寡核苷酸可以通过切割较长的核酸区段,或者通过聚合个别核苷酸前体形成。自动合成仪容许合成长度多至几百个碱基对的寡核苷酸。由于寡核苷酸可以结合互补核苷酸序列,它们可以用作检测dna或rna的探针。由dna(寡脱氧核糖核苷酸)构成的寡核苷酸可以用于pcr,即一种用于扩增dna和rna(逆转录成cdna)序列的技术。在pcr中,寡核苷酸通常称为“引物”,其容许dna聚合酶延伸寡核苷酸,并且复制互补链。
核酸分子可以包括通过天然存在的和/或非天然存在的核苷酸连接连接在一起的天然存在的和经修饰的核苷酸之任一或两者。核酸分子可以是化学或生物化学修饰的,或者可以含有非天然的或衍生化的核苷酸碱基,如本领域技术人员会容易领会的。此类修饰包括例如标记物、甲基化、用类似物取代一种或多种天然存在的核苷酸、核苷酸间修饰(例如,不带电荷的连接:例如,膦酸甲酯、磷酸三酯、氨基磷酸酯、氨基甲酸酯,等等;带电荷的连接:例如,硫代磷酸酯、二硫代磷酸酯,等等;悬垂模块:例如,肽;插入剂:例如,吖啶、补骨脂素,等等;螯合剂;烷化剂;和经修饰的连接:例如,alpha异头核酸等)。术语“核酸分子”还包括任何拓扑构象,包括单链、双链、部分双链体的、三链体的、发夹的、环状的、和挂锁的构象。
如本文中所使用的,就dna而言,术语“编码序列”、“结构核苷酸序列”、或“结构核酸分子”指在置于合适的调节序列控制下时经由转录和mrna最终翻译成多肽的核苷酸序列。就rna而言,术语“编码序列”指翻译成肽、多肽或蛋白质的核苷酸序列。通过5’端的翻译起始密码子和3’端的翻译终止密码子确定编码序列的边界。编码序列包括但不限于:基因组dna;cdna;est;和重组核苷酸序列。
基因组:如本文中所使用的,术语“基因组”指存在于细胞核内的染色体dna,并且也指存在于细胞的亚细胞组分内的细胞器dna。在本发明的一些实施方案中,可以将dna分子导入植物细胞中,使得dna分子整合入植物细胞的基因组中。在这些和其它实施方案中,dna分子可以整合入植物细胞的核dna中,或者整合入植物细胞的叶绿体或线粒体的dna中。术语“基因组”在其应用于细菌时指细菌细胞内的染色体和质粒两者。在本发明的一些实施方案中,可以将dna分子导入细菌中,使得dna分子整合入细菌的染色体中。在这些和其它实施方案中,dna分子可以是染色体整合的或者作为稳定的质粒定位或位于稳定的质粒中。
序列同一性:如本文中在两种核酸或多肽序列的背景中所使用的,术语“序列同一性”或“同一性”指两种序列中在规定的比较窗里为了实现最大对应性而比对时相同的残基。
如本文中所使用的,术语“序列同一性百分比”可以指通过在比对窗里比较两个最佳比对序列(例如,核酸序列)测定的数值,其中为了两个序列的最佳比对,比较窗中序列的部分与参照序列(其不包含添加或缺失)相比可以包含添加或缺失(即,缺口)。通过测定两个序列中存在相同核苷酸或氨基酸残基的位置的数目以产生匹配位置数目,将匹配位置的数目除以比较窗中位置的总数,并将结果乘以100以产生序列同一性百分比来计算百分比。与参照序列相比每个位置处相同的序列说成与参照序列是100%相同的,且反之亦然。
比对序列进行比较的方法是本领域中公知的。多种程序和比对算法记载于例如:smith和waterman(1981)adv.appl.math.2:482;needleman和wunsch(1970)j.mol.biol.48:443;pearson和lipman(1988)proc.natl.acad.sci.u.s.a.85:2444;higgins和sharp(1988)gene73:237-44;higgins和sharp(1989)cabios5:151-3;corpet等(1988)nucleicacidsres.16:10881-90;huang等(1992)comp.appl.biosci.8:155-65;pearson等(1994)methodsmol.biol.24:307-31;tatiana等(1999)femsmicrobiol.lett.174:247-50。序列比对方法和同源性计算的详细考虑可以参见例如altschul等(1990)j.mol.biol.215:403-10。
国立生物技术信息中心(ncbi)基本局部比对搜索工具(blasttm;altschul等(1990))可获自几个来源,包括国立生物技术信息中心(bethesda、md)以及在因特网上,其与几种序列分析程序结合使用。如何使用此程序来测定序列同一性的描述在因特网上在blasttm的“帮助”部分下可得到。为了比较核酸序列,可以采用blasttm(blastn)程序的“对2种序列blast”功能,其使用设置为缺省参数的缺省blosum62矩阵进行。与参照序列具有甚至更大相似性的核酸序列在通过此方法评估时会显示增加的百分比同一性。
特异性可杂交/特异性互补:如本文中所使用的,术语“特异性可杂交”和“特异性互补”是指示足够的互补性程度,使得在核酸分子和靶核酸分子之间发生稳定的且特异性的结合的术语。两种核酸分子间的杂交牵涉两种核酸分子的核酸序列间反平行对准的形成。然后,两种分子能够与相反链上的相应碱基形成氢键以形成双链体分子,若该双链体分子是足够稳定的,则其使用本领域中公知的方法是可检出的。核酸分子与要特异性可杂交的其靶序列不需要是100%互补的。然而,对于特异性的杂交必须存在的序列互补性量是使用的杂交条件的函数。
产生特定严格性程度的杂交条件会随选择的杂交方法的性质及杂交核酸序列的组成和长度而有所变化。一般地,杂交的温度和杂交缓冲液的离子强度(尤其是na+和/或mg++浓度)会决定杂交严格性,尽管清洗次数和清洗缓冲液组成也影响严格性。关于获得特定严格性程度需要的杂交条件的计算是本领域普通技术人员已知的,并且例如在sambrook等(编)molecularcloning:alaboratorymanual、第2版、第1-3卷、coldspringharborlaboratorypress、coldspringharbor、ny、1989、第9和11章;及hames和higgins(编)nucleicacidhybridization、irlpress、oxford、1985中讨论。关于核酸杂交的更为指示和指导可以参见例如tijssen、“overviewofprinciplesofhybridizationandthestrategyofnucleicacidprobeassays,”于laboratorytechniquesinbiochemistryandmolecularbiology-hybridizationwithnucleicacidprobes、部分i、第2章、elsevier、ny、1993;及ausubel等编、currentprotocolsinmolecularbiology、第2章、greenepublishingandwiley-interscience、ny、1995。
如本文中所使用的,“严格条件”涵盖仅在杂交分子与靶核酸分子内的同源序列之间存在有小于20%错配时发生杂交的条件。“严格条件”包括其它特定的严格性水平。如此,如本文中使用的,“中等严格性”条件是具有超过20%序列错配的分子不会杂交的那些条件;“高严格性”条件是具有超过10%错配的序列不会杂交的那些条件;而“非常高严格性”的条件是具有超过5%错配的序列不会杂交的那些条件。
以下内容是代表性的非限制性杂交条件。
高严格性条件(检出共享至少90%序列同一性的序列):于65℃在5xssc缓冲液中杂交16小时;在2xssc缓冲液中于室温清洗两次,各15分钟;并在0.5xssc缓冲液中于65℃清洗两次,各20分钟。
中等严格性条件(检出共享至少80%序列同一性的序列):在5x-6xssc缓冲液中于65-70℃杂交16-20小时;于室温在2xssc缓冲液中清洗两次,各5-20分钟;并在1xssc缓冲液中于55-70℃清洗两次,各30分钟。
非严格对照条件(共享至少50%序列同一性的序列会杂交):在6xssc缓冲液中于室温至55℃杂交16-20小时;于室温至55℃在2x-3xssc缓冲液中清洗至少两次,各20-30分钟。
如本文中所使用的,术语“基本上同源的”或“实质性同源性”就连续核酸序列而言指在严格条件下与参照核酸序列杂交的连续核苷酸序列。例如,与seqidno:1-4中任一项的参照核酸序列基本上同源的核酸序列是如下的那些核酸序列,其在严格条件(例如,所列的中等严格性条件,见上文)与seqidno:1-4中任一项的参照核酸序列杂交。基本上同源的序列可以具有至少80%序列同一性。例如,基本上同源的序列可以具有约80%至100%序列同一性,诸如约81%;约82%;约83%;约84%;约85%;约86%;约87%;约88%;约89%;约90%;约91%;约92%;约93%;约94%约95%;约96%;约97%;约98%;约98.5%;约99%;约99.5%;和约100%。实质性同源性的特性与特异性杂交紧密相关。例如,在存在有足够的互补性程度以避免在期望特异性结合的条件下,例如在严格杂交条件下核酸与非靶序列的非特异性结合时,核酸分子是特异性可杂交的。
如本文中所使用的,术语“直向同系物”指两种或更多种物种中已经从共同的祖先核苷酸序列进化,并且可以在所述两种或更多种物种中保留相同功能的基因。
如本文中所使用的,在以5’至3’方向阅读的序列的每个核苷酸与在以3’至5’方向阅读时的另一序列的每个核苷酸序列互补时,两种核酸序列分子说成展现出“完全互补性”。与参照核苷酸序列互补的核苷酸序列会展现出与参照核苷酸序列的反向互补序列相同的序列。这些术语和描述是本领域中定义明确的,并且是本领域普通技术人员容易理解的。
可操作连接的:当第一核酸序列在与第二核酸序列的功能性关系中时,第一核苷酸序列与第二核酸序列可操作连接。在重组生成时,可操作连接的核酸序列一般是连续的,并且在必需连接两个蛋白质编码区的情况中,在相同阅读框中(例如,在多顺反子orf中)。然而,为了是可操作连接的,核酸不需要是连续的。
术语“可操作连接”在提及调节序列和编码序列使用时意指调节序列影响连接的编码序列的表达。“调节序列”或“控制元件”指影响转录的时机和水平/量、rna加工或稳定性、或联合的编码序列的翻译的核苷酸序列。调节序列可以包括启动子;翻译前导序列;内含子;增强子;茎-环结构;阻抑物结合序列;终止序列;和多聚腺苷酸化识别位点等。特定调节序列可以位于与其可操作连接的编码序列上游和/或下游。还有,与编码序列可操作连接的特定调节序列可以位于双链核酸分子的关联互补链上。
启动子:如本文中所使用的,术语“启动子”指可以在转录起始上游,并且可以牵涉识别和结合rna聚合酶和其它蛋白质以启动转录的dna区。启动子可以与编码序列可操作连接以在细胞中表达,或者启动子可以与编码信号序列的核苷酸序列可操作连接,所述核苷酸序列可以与编码序列可操作连接以在细胞中表达。“植物启动子”可以是能够在植物细胞中启动转录的启动子。在发育控制下的启动子的例子包括优选在某些组织,诸如叶、根、种子、纤维、木质部导管、管胞、或厚壁组织中启动转录的启动子。此类启动子称为“组织优选性的”。仅在某些组织中启动转录的启动子称为“组织特异性的”。“细胞类型特异性”启动子主要在一种或多种器官中的某些细胞类型中,例如根或叶中的液泡细胞中驱动表达。“诱导型”启动子可以是可在环境控制下的启动子。可以通过诱导型启动子启动转录的环境条件的例子包括厌氧条件和光的存在。组织特异性、组织优先性、细胞类型特异性、和诱导型启动子构成“非组成性”启动子的类别。“组成性”启动子是可以在大多数环境条件下或在大多数细胞或组织类型中有活性的启动子。
可以在本发明的一些实施方案中使用任何诱导型启动子。参见ward等(1993)plantmol.biol.22:361-366。凭借诱导型启动子,转录速率响应诱导剂而升高。例示性的诱导型启动子包括但不限于:响应铜的的来自acei系统的启动子;响应苯磺酰胺除草剂安全剂的来自玉米的in2基因;来自tn10的tet阻抑物;和来自类固醇激素基因的诱导型启动子,其转录活性可以通过糖皮质类固醇激素诱导(schena等(1991)proc.natl.acad.sci.usa88:0421)。
例示性的组成性启动子包括但不限于:来自植物病毒的启动子,诸如来自花椰菜花叶病毒(camv)的35s启动子;来自稻肌动蛋白基因的启动子;泛素启动子;pemu;mas;玉米h3组蛋白启动子;和als启动子,即油菜(brassicanapus)als3结构基因5’的xba1/ncoi片段(或与所述xba1/ncoi片段的核苷酸序列类似性)(国际pct公开文本no.wo96/30530)。
另外,任何组织特异性或组织优选性启动子可以在本发明的一些实施方案中利用。用包含与组织特异性启动子可操作连接的编码序列的核酸分子转化的植物可以专门或优先在特定组织中生成编码序列产物。例示性的组织特异性或组织优选性启动子包括但不限于:根优选性启动子,诸如来自菜豆蛋白(phaseolin)基因的启动子;叶特异性且光诱导型启动子诸如来自cab或rubisco的启动子;花药特异性启动子诸如来自lat52的启动子;花粉特异性启动子诸如来自zm13的启动子;和小孢子优选性启动子诸如来自apg的启动子。
转化:如本文中所使用的,术语“转化”或“转导”指将一种或多种核酸分子转移到细胞中。当细胞通过将核酸分子掺入细胞基因组中,或者通过附加体复制而稳定复制核酸分子时,细胞通过转导入细胞中的核酸分子“转化”。如本文中所使用的,术语“转化”涵盖可以将核酸分子导入此类细胞中的所有技术。例子包括但不限于:用病毒载体转染;用质粒载体转化;电穿孔(fromm等(1986)nature319:791-3);脂转染(felgner等(1987)proc.natl.acad.sci.usa84:7413-7);显微注射(mueller等(1978)cell15:579-85);土壤杆菌介导的转移(fraley等(1983)proc.natl.acad.sci.usa80:4803-7);直接的dna摄取;和微粒轰击(klein等(1987)nature327:70)。
转基因:一种外源核酸序列。在一些例子中,转基因可以是编码dsrna分子的一条或两条链的序列,所述dsrna分子包含与存在于鞘翅目有害生物中的核酸分子互补的核苷酸序列。在其它例子中,转基因可以是反义核酸序列,其中反义核酸序列的表达抑制靶核酸序列的表达。在又一些例子中,转基因可以是基因序列(例如,除草剂抗性基因)、编码工业或药学有用的化合物的基因、或编码期望的农业性状的基因。在这些和其它例子中,转基因可以含有与转基因的编码序列可操作连接的调节序列(例如启动子)。
载体:导入细胞中,例如以生成经转化的细胞的核酸分子。载体可以包括容许其在宿主细胞中复制的核酸序列,诸如复制起点。载体的例子包括但不限于:质粒;粘粒;噬菌体;或将外源dna携带到细胞中的病毒。载体还可以包含一种或多种基因、反义序列、和/或选择标志基因和本领域中已知的其它遗传元件。载体可以转导、转化、或感染细胞,由此引起细胞表达由载体编码的核酸分子和/或蛋白质。任选地,载体包含帮助实现核酸分子进入细胞中的材料(例如,脂质体、蛋白质涂层等)。
产量:相对于在相同生长位置中在相同时间且在相同条件下生长的检验品种的产量,约100%或更大的稳定化产量。在特定的实施方案中,“改善的产量”或“改善产量”意指相对于在相同生长位置中在相同时间且在相同条件下生长的含有相当大密度的对该作物有害的鞘翅目有害生物的检验品种的产量,具有105%至115%或更大的稳定化产量的栽培种。
如本文中所使用的,除非明确指示或暗示,术语“一个/种”、和“所述/该”表示“至少一个/种”。
除非另有明确解释,本文中使用的所有技术和科学术语与本公开内容所属领域普通技术人员的通常理解具有相同的意义。分子生物学的常见术语的定义可以参见例如lewinb.、genesv、oxforduniversitypress、1994(isbn0-19-854287-9);kendrew等(编)、theencyclopediaofmolecularbiology、blackwellscienceltd.、1994(isbn0-632-02182-9);及meyersr.a.(编)、molecularbiologyandbiotechnology:acomprehensivedeskreference、vchpublishers、inc.、1995(isbn1-56081-569-8)。除非另有记录,所有百分比按重量计,而所有溶剂混合物比例按体积计。所有温度以摄氏度计。
iv.包含鞘翅目有害生物序列的核酸分子
a.概述
本文中描述了可用于控制鞘翅目有害生物的核酸分子。描述的核酸分子包括靶序列(例如,天然基因、和非编码序列)、dsrna、sirna、hprna、和mirna。例如,在一些实施方案中描述了dsrna、sirna、mirna和/或hprna分子,其可以与鞘翅目有害生物中的整个或部分的一种或多种天然核酸序列特异性互补。在这些和其它实施方案中,天然核酸序列可以是一种或多种靶基因,其产物可以是例如但不限于:牵涉代谢过程;牵涉生殖过程;或牵涉幼虫发育。本文中所描述的核酸分子在导入包含至少一种与该核酸分子特异性互补的天然核酸序列的细胞中时可以在细胞中启动rnai,随后降低或消除天然核酸序列的表达。在一些例子中,通过包含与靶基因特异性互补的序列的核酸分子降低或消除靶基因的表达在鞘翅目有害生物中可以是致死的,或者导致降低的生长和/或生殖。
在一些实施方案中,可以选择鞘翅目有害生物中的至少一种靶基因,其中靶基因包含选自下列的核苷酸序列:d_vir_c47185_caf1180(seqidno:1);d_vir_c1229_vatpasec(seqidno:2);d_vir_c1319_vatpaseh(seqidno:3);和contig_01_rho1_1-191_cdc42(seqidno:4)。在具体的例子中,选择鞘翅目有害生物中的靶基因,其中靶基因包含seqidno:1的新颖核苷酸序列。
在一些实施方案中,靶基因可以是包含编码多肽的核苷酸序列的核酸分子,所述多肽包含与d_vir_c47185_caf1180(seqidno:1);d_vir_c1229_vatpasec(seqidno:2);d_vir_c1319_vatpaseh(seqidno:3);和contig_01_rho1_1-191_cdc42(seqidno:4)之一的蛋白质产物的氨基酸序列至少85%相同(例如,约90%、约95%、约96%、约97%、约98%、约99%、约100%、或100%相同)的连续氨基酸序列。靶基因可以是鞘翅目有害生物中的任何核酸序列,对其的转录后抑制对鞘翅目有害生物具有有害的影响,或者对植物提供针对鞘翅目有害生物的保护益处。在具体的例子中,靶基因是包含编码多肽的核苷酸序列的核酸分子,所述多肽包含与新颖的核苷酸序列seqidno:1的蛋白质产物的氨基酸序列至少85%相同、约90%相同、约95%相同、约96%相同、约97%相同、约98%相同、约99%相同、约100%相同、或100%相同的连续氨基酸序列。
依照本发明提供了核苷酸序列,其表达生成包含核苷酸序列的rna分子,所述核苷酸序列与由鞘翅目有害生物中的编码序列编码的整个或部分天然rna分子特异性互补。在一些实施方案中,在鞘翅目有害生物摄入表达的rna分子后,可以获得鞘翅目有害生物细胞中编码序列的下调。在特定的实施方案中,鞘翅目有害生物细胞中编码序列的下调可以对鞘翅目有害生物的生长、存活力、增殖、和/或生殖产生有害的影响。
在一些实施方案中,靶序列包括转录的非编码rna序列,诸如5’utr;3’utr;剪接前导序列;内含子序列;末端内含子(outron)序列(例如,随后在反式剪接中修饰的5’utrrna);donatron序列(例如,提供反式剪接的供体序列需要的非编码rna);和靶鞘翅目有害生物基因的其它非编码转录rna。此类序列可以自单顺反子和多顺反子基因两者衍生。
如此,本文中结合一些实施方案还描述了包含至少一种核苷酸序列的irna分子(例如,dsrnas、sirna、mirna和hprna),所述核苷酸序列与鞘翅目有害生物中的整个或部分靶序列特异性互补。在一些实施方案中,irna分子可以包含与多种靶序列;例如2、3、4、5、6、7、8、9、10、或更多种靶序列的整个或部分互补的核苷酸序列。在特定的实施方案中,irna分子可以由经遗传修饰的生物体,诸如植物或细菌在体外或在体内生成。还公开了cdna序列,其可以用于生成与鞘翅目有害生物中的整个或部分靶序列特异性互补的dsrna分子、sirna分子、mirna和/或hprna分子。进一步描述了在实现特定宿主靶物的稳定转化中使用的重组dna构建体。经转化的宿主靶物可以从重组dna构建体表达有效水平的dsrna、sirna、mirna和/或hprna分子。因此,还描述了植物转化载体,其包含与在植物细胞中功能性的异源启动子可操作连接的至少一种核苷酸序列,其中核苷酸序列的表达生成包含与鞘翅目有害生物中的整个或部分靶序列特异性互补的核苷酸序列的rna分子。
在一些实施方案中,可用于控制鞘翅目有害生物的核酸分子可以包括:整个或部分的自根萤叶甲分离的天然核酸序列、d_vir_c47185_caf1180(seqidno:1);如下的核苷酸序列,其在表达时生成rna分子,该rna分子包含与整个或部分由d_vir_c47185_caf1180(seqidno:1)编码的天然rna分子特异性互补的核苷酸序列;irna分子(例如dsrna、sirna、mirna和hprna),其包含至少一种与整个或部分的d_vir_c47185_caf1180(seqidno:1)特异性互补的核苷酸序列;cdna序列,其可以用于生成与整个或部分的d_vir_c47185_caf1180(seqidno:1)特异性互补的dsrna分子、sirna分子、mirna和/或hprna分子;和在实现特定宿主靶物的稳定转化中使用的重组dna构建体,其中经转化的宿主靶物包含前述核酸分子中的一种或多种。
在这些和其它实施方案中,可用于控制鞘翅目有害生物的其它核酸分子可以包括:d_vir_c1229_vatpasec(液泡atp酶的c亚基)(seqidno:2);d_vir_c1319_vatpaseh(液泡atp酶的h亚基)(seqidno:3);和contig_01_rho1_1-191_cdc42(rho1小gtp结合蛋白)(seqidno:4);在表达时生成rna分子的核苷酸序列,所述rna分子包含如下的核苷酸序列,该核苷酸序列与由d_vir_c1229_vatpasec(csubunitofvacuolaratpase)(seqidno:2);d_vir_c1319_vatpaseh(hsubunitofvacuolaratpase)(seqidno:3);和contig_01_rho1_1-191_cdc42(rho1smallgtp-bindingprotein)(seqidno:4)编码的天然rna分子特异性互补;irna分子(例如,dsrna、sirna、mirna和hprna),其包含至少一种核苷酸序列,该核苷酸序列与d_vir_c1229_vatpasec(seqidno:2);d_vir_c1319_vatpaseh(seqidno:3);或contig_01_rho1_1-191_cdc42(seqidno:4)特异性互补;cdna序列,其可以用于生成与d_vir_c1229_vatpasec(seqidno:2);d_vir_c1319_vatpaseh(seqidno:3);或contig_01_rho1_1-191_cdc42(seqidno:4)特异性互补的dsrna分子、sirna分子、mirna和/或hprna分子;和重组dna构建体,其用于实现特定宿主靶物的稳定转化,其中经转化的宿主靶物包含一种或多种前述核酸分子。
b.核酸分子
本发明提供了抑制鞘翅目有害生物的细胞、组织或器官中的靶基因表达的irna(例如dsrna、sirna、mirna和hprna)分子;和能够在细胞或微生物中以irna分子表达以抑制鞘翅目有害生物的细胞、组织或器官中的靶基因表达的dna分子,等等。
本发明的一些实施方案提供了分离的核酸分子,其包含至少一种(例如,1、2、3、或更多种)选自下组的核苷酸序列:seqidno:1;seqidno:1的互补物;seqidno:1的至少19个连续核苷酸的片段;seqidno:1的至少19个连续核苷酸的片段的互补物;包含seqidno:1的根萤叶甲属生物体(例如wcr)的天然编码序列;包含seqidno:1的根萤叶甲属生物体的天然编码序列的互补物;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,所述天然非编码序列包含seqidno:1。在特定的实施方案中,分离的核酸序列与鞘翅目有害生物接触或被鞘翅目有害生物摄取抑制鞘翅目有害生物的生长、发育、生殖和/或进食。
在一些实施方案中,本发明的分离的核酸分子可以进一步包含至少一种(例如,1、2、3、或更多种)选自下组的核苷酸序列:seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;seqidno:2-4之任一的至少19个连续核苷酸的片段;seqidno:2-4之任一的至少19个连续核苷酸的片段的互补物;根萤叶甲属生物体的天然编码序列,其包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:2-4之任一;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,所述天然非编码序列包含seqidno:2-4之任一。在特定的实施方案中,分离的核酸序列与鞘翅目有害生物接触或被鞘翅目有害生物摄取抑制鞘翅目有害生物的生长、发育、生殖和/或进食。
在一些实施方案中,本发明的核酸分子可以包含至少一种(例如,1、2、3、或更多种)dna序列,其能够在细胞或微生物中以irna分子表达以抑制鞘翅目有害生物的细胞、组织或器官中的靶基因表达。此类dna序列可以与在包含该dna分子的细胞中发挥功能的启动子序列可操作连接以启动或增强编码的dsrna分子的转录。在一个实施方案中,至少一种(例如,1、2、3、或更多种)dna序列可以自seqidno:1衍生。seqidno:1的衍生物包括seqidno:1的片段。在一些实施方案中,seqidno:1的片段可以包含例如seqidno:1的至少约19个连续核苷酸,或其互补物。如此,seqidno:1的片段可以包含例如seqidno:1的19、20、21、22、23、24、25、26、27、28、29、或30个连续核苷酸,或其互补物。在这些和其它实施方案中,seqidno:1的片段可以包含例如seqidno:1的超过约19个连续核苷酸,或其互补物。如此,seqidno:1的片段可以包含例如seqidno:1的19、20、21、约25(例如22、23、24、25、26、27、28、和29)、约30、约40(例如35、36、37、38、39、40、41、42、43、44、和45)、约50、约60、约70、约80、约90、约100、约110、约120、约130、约140、约150、约160、约170、约180、约190、约200、或更多个连续核苷酸,或其互补物。
在特定的实施方案中,至少一种能够在细胞或微生物中以irna分子表达以抑制鞘翅目有害生物的细胞、组织或器官中的靶基因表达的dna序列可以进一步包含自选自下组的核苷酸序列衍生的dna序列:seqidno:2-4。选自包含seqidno:2-4的组的核苷酸序列的衍生物包括seqidno:2-4中一项或多项的片段。在一些实施方案中,seqidno:2-4中一项或多项的片段可以包含例如seqidno:2-4之任一的至少约19个连续核苷酸,或其互补物。如此,seqidno:2-4中一项或多项的片段可以包含例如seqidno:2-4之任一的19、20、21、22、23、24、25、26、27、28、29、或30个连续核苷酸,或其互补物。在这些和其它实施方案中,seqidno:2-4中一项或多项的片段可以包含例如seqidno:2-4之任一的超过约19个连续核苷酸,或其互补物。如此,seqidno:2-4中一项或多项的片段可以包含例如seqidno:2-4中的一项或多项的19、20、21、约25(例如22、23、24、25、26、27、28、和29)、约30、约40(例如35、36、37、38、39、40、41、42、43、44、和45)、约50、约60、约70、约80、约90、约100、约110、约120、约130、约140、约150、约160、约170、约180、约190、约200、或更多个连续核苷酸,或其互补物。
一些实施方案包括将部分或完全稳定化的dsrna分子导入鞘翅目有害生物中以抑制鞘翅目有害生物的细胞、组织或器官中靶基因的表达。在以irna分子(例如dsrna、sirna、mirna、和hprna)表达并被鞘翅目有害生物摄取时,包含seqidno:1-4之任一的一种或多种片段的核酸序列可以引起下列一项或多项:死亡、生长抑制、性别比变化、窝卵数(broodsize)减少、感染停止、和/或鞘翅目有害生物进食停止。例如,在一些实施方案中,提供了dsrna分子,其包含含有与鞘翅目有害生物靶基因序列基本上同源的约19至约300个核苷酸的核苷酸序列,且包含seqidno:1的一个或多个片段。包含至少一种与seqidno:1基本上同源的核苷酸序列的dsrna分子的具体例子可以是但不限于长度为至少19、20、21、约25(例如22、23、24、25、26、27、28、和29)、约30、约40(例如35、36、37、38、39、40、41、42、43、44、和45)、约50、约60、约70、约80、约90、约100、约110、约120、约130、约140、约150、约160、约170、约180、约190、约200、或更多个核苷酸。此类dsrna分子可以进一步包含seqidno:2-4的一种或多种片段。此类dsrna分子的表达可以例如导致摄取dsrna分子的鞘翅目有害生物中的死亡率。
在某些实施方案中,本发明提供的dsrna分子包含与包含seqidno:1的靶基因互补的核苷酸序列和/或与seqidno:1的片段互补的核苷酸序列,在鞘翅目有害生物中抑制所述靶基因导致对于鞘翅目有害生物生长、发育、或其它生物学功能必需的蛋白质或核苷酸序列制剂的降低或消除。选择的核苷酸序列可以与下列各项展现出约80%至约100%序列同一性:seqidno:1;seqidno:1中所列的核苷酸序列的连续片段;或任何前述物的互补物。例如,选择的核苷酸序列可以与下列各项展现出约81%;约82%;约83%;约84%;约85%;约86%;约87%;约88%;约89%;约90%;约91%;约92%;约93%;约94%约95%;约96%;约97%;约98%;约98.5%;约99%;约99.5%;或约100%序列同一性:seqidno:1;seqidno:1中所列的核苷酸序列的连续片段;或任何前述物的互补物。在特定的实施方案中,本发明提供的dsrna分子可以进一步包含一种或多种与包含seqidno:2-4之一的靶基因互补的核苷酸序列,在鞘翅目有害生物中抑制所述靶基因导致对于鞘翅目有害生物生长、发育、或其它生物学功能必需的蛋白质或核苷酸序列制剂的降低或消除。
在一些实施方案中,能够在细胞或微生物中以irna分子表达以抑制靶基因表达的dna分子可以包含与存在于一种或多种靶鞘翅目有害生物物种中的天然核酸序列的整个或部分特异性互补的单一核苷酸序列,或者可以从多个此类特异性互补序列将dna分子构建为嵌合物。
在一些实施方案中,核酸分子可以包含以“间隔物序列”分开的第一和第二核苷酸序列。间隔物序列可以是包含促进第一和第二核苷酸序列之间的二级结构形成(在这是期望的情况下)的任何核苷酸序列的区域。在一个实施方案中,间隔物序列是mrna的有义或反义编码序列的部分。或者,间隔物序列可以包含能够与核酸分子可操作连接的核苷酸或其同系物的任何组合。
例如,在一些实施方案中,dna分子可以包含编码一种或多种不同rna分子的核苷酸序列,其中每种不同rna分子包含第一核苷酸序列和第二核苷酸序列,其中第一和第二核苷酸序列彼此互补。第一和第二核苷酸序列在rna分子内可以通过间隔物序列连接。间隔物序列可以构成第一核苷酸序列或第二核苷酸序列的一部分。包含第一和第二核苷酸序列的rna分子的表达可以通过第一和第二核苷酸序列的特异性杂交而导致本发明的dsrna分子的形成。第一核苷酸序列或第二核苷酸序列可以与对于鞘翅目有害生物而言天然的核酸序列(例如,靶基因、或转录非编码序列)、其衍生物、或其互补序列基本上相同。
dsrna核酸分子包含双链的聚合核糖核苷酸序列,并且可以包含对磷酸-糖主链或核苷的修饰。可以改编rna结构的修饰以容许特异性抑制。在一个实施方案中,可以经由遍在的酶促过程修饰dsrna分子,使得可以生成sirna分子。此酶促过程可以利用体外或体内的rna酶iii酶,诸如真核生物中的dicer。参见elbashir等(2001)nature411:494-8;及hamilton和baulcombe(1999)science286(5441):950-2。dicer或功能等同性rna酶iii酶将较大的dsrna链和/或hprna分子切割成较小的寡核苷酸(例如sirnas),每个所述寡核苷酸长度是约19-25个核苷酸。由这些酶生成的sirna分子具有2至3个核苷酸3’突出物、和5’磷酸根和3’羟基末端。由rna酶iii酶生成的sirna分子在细胞中解开,并分成单链rna。然后,sirna分子与自靶基因转录的rna序列特异性杂交,并且随后,这两种rna分子受到固有的细胞rna降解机制降解。此过程可以导致由靶生物体中的靶基因编码的rna序列的有效降解或消除。结果是靶定基因的转录后沉默。在一些实施方案中,通过内源rna酶iii酶从异源核酸分子生成的sirna分子可以有效介导鞘翅目有害生物中靶基因的下调。
在一些实施方案中,本发明的核酸分子可以包括至少一种可以转录成单链rna分子的非天然存在的核苷酸序列,所述单链rna分子能够经由分子间杂交在体内形成dsrna分子。此类dsrna序列通常自身装配,并且可以在鞘翅目有害生物的营养来源中提供以实现靶基因的转录后抑制。在这些和其它实施方案中,本发明的核酸分子可以包含两种不同非天然存在的核苷酸序列,其中每种与鞘翅目有害生物中的不同靶基因特异性互补。在以dsrna分子对鞘翅目有害生物提供此类核酸分子时,dsrna分子抑制鞘翅目有害生物中至少两种不同靶基因的表达。
c.获得核酸分子
可以使用鞘翅目有害生物中的多种天然序列作为靶序列来设计本发明的核酸分子,诸如irna和编码irna的dna分子。然而,天然序列的选择不是直接的过程。鞘翅目有害生物中的仅少量天然序列会是有效的靶物。例如,不能肯定预测特定天然序列是否可以由本发明的核酸分子有效下调,或者特定天然序列的下调是否会对鞘翅目有害生物的生长、存活力、增殖和/或生殖具有有害影响。绝大多数天然鞘翅目有害生物序列,诸如自其分离的est(例如,如美国专利7,612,194和美国专利7,943,819中所列的)对鞘翅目有害生物,诸如wcr或ncr的生长、存活力、增殖和/或生殖没有有害影响。也不可预测可以对鞘翅目有害生物具有有害影响的天然序列中的哪些能够在重组技术中使用以在宿主植物中表达与此类天然序列互补的核酸分子,并且在进食后对鞘翅目有害生物提供有害影响而不对宿主植物引起损害。
在一些实施方案中,本发明的核酸分子(例如,要在鞘翅目有害生物的宿主植物中提供的dsrna分子)选择为靶向编码对于鞘翅目有害生物存活必需的蛋白质或蛋白质部分,诸如牵涉代谢或催化生物化学途径、细胞分裂、复制、能量代谢、消化、寄生等的氨基酸序列的cdna序列。如本文中所描述的,靶生物体摄入含有一种或多种dsrna的组合物可以导致靶物的死亡或其它抑制,所述dsrna的至少一个区段至少与靶病原体细胞中生成的rna的基本上相同的区段特异性互补。可以使用自鞘翅目有害生物衍生的核苷酸序列(dna或rna)来构建对鞘翅目有害生物侵袭有抗性的植物细胞。例如,可以转化鞘翅目有害生物(例如,玉蜀黍或大豆)的宿主植物以含有自鞘翅目有害生物衍生的如本文中提供的一种或多种核苷酸序列。转化入宿主中的核苷酸序列可以编码一种或多种rna,其在经转化宿主内的细胞或生物学流体中形成dsrna序列,如此若/在鞘翅目有害生物与转基因宿主形成营养关系时使dsrna可用。这可以导致鞘翅目有害生物细胞中一种或多种基因表达的阻抑,最终导致死亡或对其生长或发育的抑制。
如此,在一些实施方案中,靶向实质上牵涉鞘翅目有害生物的生长、发育和生殖的基因。本发明中使用的其它靶基因可以包括例如那些在鞘翅目有害生物存活力、运动、迁移、生长、发育、传染性、进食位置的建立和生殖中发挥重要作用的靶基因。因此,靶基因可以是持家基因或转录因子。另外,本发明中使用的天然鞘翅目有害生物核苷酸序列也可以自植物、病毒、细菌或昆虫基因的同系物(例如,直向同系物)衍生,所述同系物的功能对于本领域技术人员是已知的,且其核苷酸序列与靶鞘翅目有害生物的基因组中的靶基因特异性可杂交。通过杂交鉴定与已知核苷酸序列的基因同系物的方法是本领域技术人员已知的。
在一些实施方案中,本发明提供了用于获得包含用于生成irna(例如dsrna、sirna、mirna、和hprna)分子的核苷酸序列的核酸分子的方法。一种此类实施方案包括:(a)对一种或多种靶基因分析其在鞘翅目有害生物中在dsrna介导的基因阻抑后的表达、功能和表型;(b)用探针探查cdna或gdna文库,所述探针包含整个或部分来自靶定鞘翅目有害生物的核苷酸序列或其同系物,所述靶定鞘翅目有害生物在dsrna介导的阻抑分析中展现出改变的(例如,降低的)生长或发育表型;(c)鉴定与探针特异性杂交的dna克隆;(d)分离步骤(b)中鉴定的dna克隆;(e)对包含步骤(d)中分离的克隆的cdna或gdna片段测序,其中测序的核酸分子转录整个或实质性部分的rna序列或其同系物;并(f)化学合成整个或实质性部分的基因序列,或sirna或mirna或hprna或mrna或dsrna。
在别的实施方案中,用于获得包含用于生成实质性部分的irna(例如dsrna、sirna、mirna、和hprna)分子的核苷酸序列的核酸片段的方法包括:(a)合成第一和第二寡核苷酸引物,其与来自靶定鞘翅目有害生物的天然核苷酸序列的一部分特异性互补;并(b)使用步骤(a)的第一和第二寡核苷酸引物扩增克隆载体中存在的cdna或gdna插入物,其中扩增的核酸分子转录实质性部分的sirna或mirna或hprna或mrna或dsrna分子。
本发明的核酸可以通过多种办法分离、扩增、或生成。例如,可以通过pcr扩增自gdna或cdna文库衍生的靶核酸序列(例如,靶基因或靶转录非编码序列),或其部分获得irna(例如dsrna、sirna、mirna、和hprna)分子。可以从靶生物体提取dna或rna,并且可以使用本领域普通技术人员已知的方法自其制备核酸文库,并且可以使用自靶生物体生成的gdna或cdna文库进行靶基因的pcr扩增和测序。可以使用确认的pcr产物作为体外转录的模板以在最小程度启动子的情况下生成有义和反义rna。或者,可以使用标准化学,诸如亚磷酰胺化学通过许多技术(参见例如ozaki等(1992)nucleicacidsresearch、20:5205-5214;及agrawal等(1990)nucleicacidsresearch、18:5419-5423),包括使用自动化dna合成仪(例如p.e.biosystems、inc.(fostercity、calif.)392或394型dna/rna合成仪)合成核酸分子。参见例如beaucage等(1992)tetrahedron、48:2223-2311;美国专利4,980,460、4,725,677、4,415,732、4,458,066、和4,973,679。也可以采用备选化学,其生成非天然主链基团,诸如硫代磷酸酯、氨基磷酸酯,等等。
本领域技术人员可以经由手动或自动反应化学或酶促生成,或者在包含含有编码rna、dsrna、sirna、mirna、或hprna分子的序列的核酸分子的细胞中在体内生成本发明的rna、dsrna、sirna、mirna、或hprna分子。也可以通过部分或完全有机合成生成rna,可以通过体外酶促或有机合成引入任何经修饰的核糖核苷酸。可以通过细胞rna聚合酶或噬菌体rna聚合酶(例如t3rna聚合酶、t7rna聚合酶、和sp6rna聚合酶)合成rna分子。可用于克隆和表达核苷酸序列的表达构建体是本领域中已知的。参见例如国际pct公开文本no.wo97/32016;及美国专利5,593,874、5,698,425、5,712,135、5,789,214、和5,804,693。可以在导入细胞中前纯化以化学法或通过体外酶促合成合成的rna分子。例如,可以通过用溶剂或树脂提取、沉淀、电泳、化学层析、或其组合从混合物纯化rna分子。或者,可以在不纯化或最小程度纯化的情况下使用化学或通过体外酶促合成来合成的rna分子,例如以避免由于样品加工所致的损伤。可以将rna分子干燥以进行贮存或者在水性溶液中溶解。溶液可以含有缓冲剂或盐以促进dsrna分子双链体链的退火和/或稳定化。
在实施方案中,可以通过单一自身互补的rna链或者从两条互补rna链形成dsrna分子。dsrna分子可以在体内或在体外合成。细胞的内源rna聚合酶可以在体内介导一条或两条rna链的转录,或者可以使用克隆的rna聚合酶在体内或在体外介导转录。对鞘翅目有害生物中靶基因的转录后抑制通过宿主器官、组织、或细胞类型中的特异性转录(例如,通过使用组织特异性启动子进行);刺激宿主中的环境条件(例如,通过使用响应感染、应激、温度、和/或化学诱导物的诱导型启动子进行);和/或工程化改造宿主的某个发育阶段或发育龄的转录(例如,通过使用发育阶段特异性启动子进行)可以是宿主靶向的。形成dsrna分子的rna链(不论是体外还是体内转录的)可以是或不是多聚腺苷酸化的,并且可以能够或不能被细胞翻译装置翻译成多肽。d.重组载体和宿主细胞转化
在一些实施方案中,本发明还提供了用于导入细胞(例如,细菌细胞、酵母细胞、或植物细胞)中的dna分子,其中dna分子包含在表达为rna并被鞘翅目有害生物摄取后实现对鞘翅目有害生物的细胞、组织或器官中靶基因的阻抑的核苷酸序列。如此,一些实施方案提供重组核酸分子,其包含能够在植物细胞中以irna(例如,dsrna、sirna、mirna、和hprna)分子表达以抑制鞘翅目有害生物中的靶基因表达的核酸序列。为了启动或增强表达,此类重组核酸分子可以包含一种或多种调节序列,该调节序列可以与能够表达为irna的核酸序列可操作连接。在植物中表达基因阻抑分子的方法是已知的,并且可以用于表达本发明的核苷酸序列。参见例如国际pct公开文本no.wo06073727;及美国专利公开文本no.2006/0200878al)。
在特定的实施方案中,本发明的重组dna分子可以包含编码dsrna分子的核酸序列。此类重组dna分子可以编码dsrna分子,其能够在摄取后抑制鞘翅目有害生物细胞中内源靶基因的表达。在许多实施方案中,转录的dsrna分子可以以稳定化形式提供;例如,以发夹和茎和环结构提供。
在这些和其它实施方案中,可以通过自与选自下组的核苷酸序列基本上同源的核苷酸序列转录形成dsrna分子的一条链:seqidno:1;seqidno:1的互补物;seqidno:1的至少19个连续核苷酸的片段;seqidno:1的至少19个连续核苷酸的片段的互补物;包含seqidno:1的根萤叶甲属生物体(例如wcr)的天然编码序列;包含seqidno:1的根萤叶甲属生物体的天然编码序列的互补物;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含seqidno:1;根萤叶甲属生物体(例如wcr)的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,所述天然非编码序列包含seqidno:1。
也可以通过自与选自下组的核苷酸序列基本上同源的核苷酸序列转录形成dsrna分子的一条链:seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;seqidno:2-4之任一的至少19个连续核苷酸的片段;seqidno:2-4之任一的至少19个连续核苷酸的片段的互补物;根萤叶甲属生物体(例如wcr)的天然编码序列,其包含seqidno:2-4之任一;根萤叶甲属生物体(例如wcr)的天然编码序列的互补物,所述天然编码序列包含seqidno:2-4之任一;根萤叶甲属生物体的天然非编码序列,其转录成包含seqidno:2-4之任一的天然rna分子;根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列转录成包含seqidno:2-4之任一的天然rna分子;根萤叶甲属生物体(例如wcr)的天然编码序列的至少19个连续核苷酸的片段,其包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述片段包含seqidno:2-4之任一;根萤叶甲属生物体的非天然编码序列的至少19个连续核苷酸的片段,其转录成包含seqidno:2-4之任一的天然rna分子;和根萤叶甲属生物体的非天然编码序列的至少19个连续核苷酸的片段的互补物,所述片段转录成包含seqidno:2-4之任一的天然rna分子。
在特定的实施方案中,编码dsrna分子的重组dna分子可以包含至少两个核苷酸序列,编码序列排列为使得相对于至少一个启动子,一个核苷酸序列为有义取向,而另一个核苷酸序列为反义取向,其中有义核苷酸序列和反义核苷酸序列通过约五(约5)至约一千(约1000)个核苷酸的间隔物序列连接或相连。间隔物序列可以在有义和反义序列之间形成环。有义核苷酸序列或反义核苷酸序列可以与靶基因(例如,包含seqidno:1-4之一的基因)或其片段的核苷酸序列基本上同源。然而,在一些实施方案中,重组dna分子可以编码没有间隔物序列的dsrna分子。在实施方案中,有义编码序列和反义编码序列可以是不同长度。
可以经由在本发明的重组核酸分子中创建合适的表达盒将鉴定为对鞘翅目有害生物具有有害影响或者就鞘翅目有害生物而言的植物保护效应的序列容易地掺入表达的dsrna分子中。例如,可以如下将此类序列以具有茎和环结构的发夹表达,即采用第一区段,其对应于靶基因序列(例如,seqidno:1及其片段);将此序列与第二区段间隔物区连接,所述第二区段间隔物区与第一区段不是同源或互补的;并将这与第三区段连接,其中第三区段的至少一部分与第一区段基本上互补。此类构建体通过第一区段与第三区段的杂交而形成茎和环结构,其中环结构形式包含第二区段。参见例如美国专利公开文本no.us2002/0048814和us2003/0018993;及国际pct公开文本no.wo94/01550和wo98/05770。dsrna分子可以例如以双链结构诸如茎-环结构(例如发夹)形式生成,由此通过共表达例如在别的植物可表达盒上的靶定基因的片段增强靶定天然鞘翅目有害生物序列的sirna的生成,这导致增强的sirna生成,或者降低甲基化或防止dsrna发夹启动子的转录基因沉默。
本发明的实施方案包括将本发明的重组核酸分子导入植物中(即,转化)以实现鞘翅目有害生物抑制水平的一种或多种irna分子的表达。重组dna分子可以例如是载体,诸如线性或闭合环状质粒。载体系统可以是单一载体或质粒,或者共同含有要导入宿主基因组中的总dna的两个或更多个载体或质粒。另外,载体可以是表达载体。可以例如将本发明的核酸序列适当插入载体中,在一种或多种宿主中发挥功能以驱动连接的编码序列或其它dna序列表达的合适启动子控制下。许多载体可用于此目的,并且合适载体的选择会主要取决于要插入载体中的核酸的大小和要用载体转化的特定宿主细胞。根据其功能(例如dna扩增或dna表达)及与其相容的特定宿主细胞,每种载体含有各种组件。
为了对转基因植物赋予鞘翅目有害生物抗性,例如可以在重组植物的组织或流体内将重组dna转录成irna分子(例如,形成dsrna分子的rna分子)。irna分子可以包含与可以以宿主植物物种为食的鞘翅目有害生物内的相应转录核苷酸序列基本上同源且特异性可杂交的核苷酸序列。例如,鞘翅目有害生物可以通过摄取包含irna分子的转基因宿主植物的细胞或流体来接触在转基因宿主植物细胞中转录的irna分子。如此,靶基因的表达在侵袭转基因宿主植物的鞘翅目有害生物内受到irna分子阻抑。在一些实施方案中,对靶鞘翅目有害生物中靶基因表达的阻抑可以生成对有害生物有抗性的植物。
为了使irna分子能够投递至与已经用本发明的重组核酸分子转化的植物细胞在营养关系中的鞘翅目有害生物,需要在植物细胞中表达(即,转录)irna分子。如此,重组核酸分子可以包含与一种或多种调节序列诸如在宿主细胞中发挥功能的异源启动子序列可操作连接的本发明的核苷酸序列,所述宿主细胞诸如要扩增核酸分子的细菌细胞,或要表达核酸分子的植物细胞。
适合于用于本发明的核酸分子的启动子包括诱导型、病毒的、合成的、或组成性的那些启动子,它们都是本领域中公知的。描述此类启动子的非限制性例子包括美国专利no.6,437,217(玉米rs81启动子);5,641,876(稻肌动蛋白启动子);6,426,446(玉米rs324启动子);6,429,362(玉米pr-1启动子);6,232,526(玉米a3启动子);6,177,611(组成性玉米启动子);5,322,938、5,352,605、5,359,142、和5,530,196(35s启动子);6,433,252(玉米l3油质蛋白启动子);6,429,357(稻肌动蛋白2启动子和稻肌动蛋白2内含子);6,294,714(光诱导型启动子);6,140,078(盐诱导型启动子);6,252,138(病原性诱导型启动子);6,175,060(磷缺乏诱导型启动子);6,388,170(双向启动子);6,635,806(gamma薏苡辛(coixin)启动子);及美国专利申请流水号no.09/757,089(玉米叶绿体醛缩酶启动子)。其它启动子包括胭脂氨酸合成酶(nos)启动子(ebert等(1987)proc.natl.acad.sci.usa84(16):5745-9)和章鱼碱合酶(ocs)启动子(两者都在根癌土壤杆菌的肿瘤诱导质粒上携带);花椰菜花叶病毒(caulimovirus)启动子,诸如花椰菜花叶病毒(camv)19s启动子(lawton等(1987)plantmol.biol.9:315-24);及camv35s启动子(odell等(1985)nature313:810-2;玄参花叶病毒35s启动子(walker等(1987)proc.natl.acad.sci.usa84(19):6624-8);蔗糖合酶启动子(yang和russell(1990)proc.natl.acad.sci.usa87:4144-8);r基因复合物启动子(chandler等(1989)plantcell1:1175-83);叶绿素a/b结合蛋白基因启动子;camv35s(美国专利no.5,322,938、5,352,605、5,359,142、和5,530,196);fmv35s(美国专利no.6,051,753、和5,378,619);pc1sv启动子(美国专利no.5,850,019);scp1启动子(美国专利no.6,677,503);和agrtu.nos启动子(genbank登录号v00087;depicker等(1982)j.mol.appl.genet.1:561-73;bevan等(1983)nature304:184-7)。
在特定的实施方案中,本发明的核酸分子包含组织特异性启动子,诸如根特异性启动子。根特异性启动子驱动专门或优先在根组织中表达可操作连接的序列。根特异性启动子的例子是本领域中已知的。参见例如美国专利5,110,732;5,459,252和5,837,848;及opperman等(1994)science263:221-3;以及hirel等(1992)plantmol.biol.20:207-18。在一些实施方案中,可以在两个根特异性启动子间克隆依照本发明的用于鞘翅目有害生物控制的核苷酸序列或片段,所述根特异性启动子在转基因植物细胞中可操作,并且在其中表达以在转基因植物细胞中生成rna分子,随后其可以形成dsrna分子,如上文所描述的。鞘翅目有害生物可以摄取植物组织中表达的irna分子,从而实现对靶基因表达的阻抑。
可以任选地与感兴趣的核酸分子可操作连接的其它调节序列包括位于启动子序列和功能为翻译前导序列的编码序列之间的5’utr。翻译前导序列存在于完全加工的mrna中,并且其可以影响前转录物(primarytranscript)的加工和/或rna的稳定性。翻译前导序列的例子包括玉米和矮牵牛热休克蛋白前导物(美国专利no.5,362,865)、植物病毒壳体蛋白前导物、植物rubisco前导物等。见例如turnerandfoster(1995)molecularbiotech.3(3):225-36。5’utr的非限制性例子包括gmhsp(美国专利no.5,659,122);phdnak(美国专利no.5,362,865);atant1;tev(carrington和freed(1990)j.virol.64:1590-7);及agrtunos(genbank登录号v00087;及bevan等(1983)nature304:184-7)。
可以任选地与感兴趣的核酸分子可操作连接的其它调节序列还包括3’非翻译序列、3’转录终止区、或多腺苷酸化区。这些是位于核苷酸序列下游的遗传元件,并且包括提供多腺苷酸化信号的多核苷酸、和/或能够影响转录或mrna加工的其它调节信号。多腺苷酸化信号在植物中发挥功能以引起多腺苷酸化核苷酸添加至mrna前体的3’端。多腺苷酸化序列可以源自多种植物基因或t-dna基因。3’转录终止区的非限制性例子是胭脂氨酸合成酶3’区(nos3’;fraley等(1983)proc.natl.acad.sci.usa80:4803-7)。在ingelbrecht等、(1989)plantcell1:671-80中提供使用不同3’非翻译区的例子。多腺苷酸化信号的非限制性例子包括来自豌豆(pisumsativum)rbcs2基因(ps.rbcs2e9;coruzzi等(1984)emboj.3:1671-9)和agrtu.nos(genbank登录号e01312)的。
一些实施方案可以包括植物转化载体,该植物转化载体包含分离的且纯化的dna分子,该dna分子包含与本发明的一种或多种核苷酸序列可操作连接的至少一种上文描述的调节序列。在表达时,一种或多种核苷酸序列生成一种或多种包含核苷酸序列的rna分子,所述核苷酸序列与鞘翅目有害生物中的整个或部分的天然rna分子特异性互补。如此,核苷酸序列可以包含编码靶定鞘翅目有害生物rna转录物内存在的整个或部分核糖核苷酸序列的区段,并且可以包含整个或部分靶定鞘翅目有害生物转录物的反向重复。植物转化载体可以含有与超过一种靶序列特异性互补的序列,如此容许生成超过一种dsrna以抑制靶鞘翅目有害生物的一个或多个群体或物种的细胞中两种或更多种基因的表达。可以将与不同基因中存在的核苷酸序列特异性互补的核苷酸序列区段组合成单一复合核酸分子以在转基因植物中表达。此类区段可以是连续的或者由间隔物序列分开。
在一些实施方案中,可以通过在同一质粒中序贯插入别的核苷酸序列修饰已经含有本发明的至少一种核苷酸序列的本发明质粒,其中所述别的核苷酸序列与同初始的至少一种核苷酸序列相同的调节元件可操作连接。在一些实施方案中,核酸分子可以设计为抑制多种靶基因。在一些实施方案中,要抑制的多种基因可以获自相同鞘翅目有害生物物种,其可以增强核酸分子的效率。在其它实施方案中,可以从不同鞘翅目有害生物衍生基因,从而可以加宽药剂有效针对的鞘翅目有害生物的范围。在靶向多种基因以实现阻抑或表达和阻抑的组合时,可以制作多顺反子dna元件。
本发明的重组核酸分子或载体可以包含对经转化的细胞,诸如植物细胞赋予可选择表型的选择标志物。也可以使用选择标志物来选择包含本发明的重组核酸分子的植物或植物细胞。标志物可以编码杀生物剂抗性、抗生素抗性(例如,卡那霉素、遗传霉素(g418)、博来霉素、潮霉素,等等)、或除草剂抗性(例如,草甘膦等)。选择标志物的例子包括但不限于编码卡那霉素抗性并且可以使用卡那霉素、g418等选择的neo基因;编码双丙氨磷抗性的bar基因;编码草甘膦抗性的突变体epsp合酶基因;赋予对溴草腈(bromoxynil)的抗性的腈水解酶基因;赋予咪唑啉酮或磺酰脲抗性的突变体乙酰乳酸合酶基因(als);和甲氨蝶呤抗性dhfr基因。可用多种选择标志物,其赋予对氨苄青霉素、博来霉素、氯霉素、庆大霉素、潮霉素、卡那霉素、林可霉素、甲氨蝶呤、膦丝菌素、嘌呤霉素、大观霉素、利福平、链霉素和四环素等的抗性。此类选择标志物的例子例示于例如美国专利no.5,550,318;5,633,435;5,780,708和6,118,047。
本发明的重组核酸分子或载体还可以包含筛选标志物。可以使用筛选标志物来监测表达。例示性筛选标志物包括β-葡糖醛酸糖苷酶或uida基因(gus),其编码已知多种生色底物的酶(jefferson等(1987)plantmol.biol.rep.5:387-405);r-基因座基因,其编码调节植物组织中花色素苷色素(红色)生成的产物(dellaporta等(1988)“molecularcloningofthemaizer-njallelebytransposontaggingwithac.”于18thstadlergeneticssymposium、p.gustafson和r.appels编、(newyork:plenum)、第263-82页);β-内酰胺酶基因(sutcliffe等(1978)proc.natl.acad.sci.usa75:3737-41);编码已知多种生色底物的酶(例如,padac、生色头孢菌素);萤光素酶基因(ow等(1986)science234:856-9);xyle基因,其编码能转化生色儿茶酚的儿茶酚双加氧酶(zukowski等(1983)gene46(2-3):247-55);淀粉酶基因(ikatu等(1990)bio/technol.8:241-2);酪氨酸酶基因,其编码能够将酪氨酸氧化为dopa和多巴醌(dopaquinone)(其继而缩合成黑色素)的酶(katz等(1983)j.gen.microbiol.129:2703-14);和α-半乳糖苷酶。
在一些实施方案中,可以在用于创建转基因植物和在植物中表达异源核酸的方法中使用如上文所描述的重组核酸分子以制备展现出降低的对鞘翅目有害生物的易感性的转基因植物。例如,可以通过将编码irna分子的核酸分子插入植物转化载体中,并将这些导入植物中来制备植物转化载体。
适合于转化宿主细胞的方法包括可以将dna导入细胞中的任何方法,诸如通过转化原生质体(参见例如美国专利5,508,184),通过干燥(desiccation)/抑制介导的dna摄取(参见例如potrykus等(1985)mol.gen.genet.199:183-8),通过电穿孔(参见例如美国专利5,384,253),通过与碳化硅纤维一起搅拌(参见例如美国专利5,302,523和5,464,765),通过土壤杆菌介导的转化(参见例如美国专利5,563,055;5,591,616;5,693,512;5,824,877;5,981,840;和6,384,301)及通过加速dna包被的颗粒(参见例如美国专利no.5,015,580;5,550,318;5,538,880;6,160,208;6,399,861;和6,403,865)等进行。特别可用于转化玉米的技术记载于例如美国专利no.7,060,876和5,591,616;及国际pct公开文本wo95/06722。经由应用诸如这些的技术,可以稳定转化实际上任何物种的细胞。在一些实施方案中,在转化的dan整合入宿主细胞的基因组中。在多细胞物种的情况中,可以将转基因细胞再生成转基因生物体。可以使用任一种这些技术来生成例如在转基因植物的基因组中包含编码一种或多种irna分子的一种或多种核酸序列的转基因植物。
最广泛利用的用于将表达载体导入植物中的方法基于土壤杆菌的天然转化系统。根癌土壤杆菌(a.tumefaciens)和发根土壤杆菌(a.rhizogenes)是遗传转化植物细胞的植物病原性土壤细菌。根癌土壤杆菌和发根土壤杆菌分别的ti和ri质粒携带负责遗传转化植物的基因。ti(肿瘤诱导)质粒含有转移至经转化植物的较大的区段,称为t-dna。ti质粒的另一个区段vir区负责t-dna转移。t-dna区以末端重复为边界。在经修饰的二元载体中,已经删除肿瘤诱导基因,并且利用vir区的功能来转移以t-dna边界序列为边界的外来dna。t区还可以含有用于有效回收转基因植物和细胞的选择标志物和用于插入转移序列诸如dsrna编码核酸的多克隆位点。
如此,在一些实施方案中,植物转化载体源自根癌土壤杆菌的ti质粒(参见例如美国专利no.4,536,475、4,693,977、4,886,937、和5,501,967;及欧洲专利ep0122791)或发根土壤杆菌的ri质粒。别的植物转化载体包括例如但不限于那些由herrera-estrella等(1983)nature303:209-13;bevan等(1983)nature304:184-7;klee等(1985)bio/technol.3:637-42;及于欧洲专利ep0120516描述的载体,以及那些源自任一种前述载体的载体。可以修饰与植物天然相互作用的其它细菌诸如中华根瘤菌属(sinorhizobium)、根瘤菌属(rhizobium)、和中生根瘤菌属(mesorhizobium)以介导对许多多种多样的植物的基因转移。可以使这些植物关联共生细菌成为感受态,以通过获得解除的ti质粒和合适的二元载体两者进行基因转移。
在对接受细胞提供外源dna后,一般鉴定经转化的细胞以进一步培养和植物再生。为了改善鉴定经转化的细胞的能力,在用于生成转化体的转化载体的情况中可以期望采用选择或筛选标志物基因,如先前列出的。在使用选择标志物的情况中,可以通过将细胞暴露于一种或多种选择剂在潜在经转化的细胞群体内鉴定经转化的细胞。在使用筛选标志物的情况中,可以对细胞筛选期望的标志物基因性状。
可以将幸免于对选择剂暴露的细胞,或者已经在筛选测定法中评分呈阳性的细胞在支持植物再生的培养基中培养。在一些实施方案中,可以通过包括别的物质,诸如生长调节物改良任何合适的植物组织培养基(例如ms和n6培养基)。可以将组织在具有生长调节物的基础培养基上维持,直到足够的组织可用于开始植物再生努力,或者在重复的手动选择轮次后,直到组织形态学适合于再生(例如,至少2周),然后转移到有益于枝条形成的培养基。定期转移培养物,直到已经发生足够的枝条形成。一旦形成枝条,将它们转移到有益于根形成的培养基。一旦形成足够的根,可以将植物转移到土壤以进一步生长和成熟。
为了确认再生植物中存在感兴趣的核酸分子(例如,编码一种或多种irna分子的dna序列,所述irna分子抑制鞘翅目有害生物中靶基因表达),可以实施多种测定法。此类测定法包括例如:分子生物学测定法,诸如southern和northern印迹、pcr、和核酸测序;生物化学测定法,诸如检测蛋白质产物的存在,例如,其通过免疫学手段(elisa和/或western印迹)或者通过酶功能进行;植物部分测定法,诸如叶或根测定法;和对再生的全植物的表型的分析。
例如,可以使用例如对感兴趣的核酸分子特异性的寡核苷酸引物可以通过pcr扩增分析整合事件。pcr基因型分型理解为包括但不限于自预测含有整合入基因组中的感兴趣核酸分子的分离的宿主植物愈伤组织(callustissue)衍生的基因dna的聚合酶链式反应(pcr)扩增,接着是pcr扩增产物的标准克隆和序列分析。pcr基因型分型的方法已经充分描述(例如rios、g.等(2002)plantj.32:243-53),并且可以应用于自任何植物物种(例如玉蜀黍和大豆)或组织类型,包括细胞培养物衍生的基因组dna。
使用土壤杆菌依赖性转化方法形成的转基因植物通常含有插入一条染色体中的单一重组dna序列。单一重组dna序列称为“转基因事件”或“整合事件”。此类转基因植物在插入的外源序列方面是半合子的。在一些实施方案中,可以通过将含有单一外源基因序列的独立的分离转基因植物与自身,例如f0植物有性杂交(自交)以生成fl种子来获得就转基因而言纯合的转基因植物。生成的四分之一的fl种子就转基因而言会是纯合的。使fl种子萌发产生植物,通常可以使用snp测定法或容许区别杂合子和纯合子的热扩增测定(即,接合性测定法)对所述植物测试杂合性。
在特定的实施方案中,在植物细胞中生成至少2、3、4、5、6、7、8、9或10或更多种不同irna分子,其具有鞘翅目有害生物抑制效果。可以从不同转化事件中引入的多种核酸序列,或者从单一转化事件中引入的单一核酸序列表达irna分子(例如,dsrna分子)。在一些实施方案中,在单一启动子的控制下表达多个irna分子。在其它实施方案中,在多个启动子的控制下表达多个irna分子。可以表达单一irna分子,其包含多个核酸序列,该核酸序列各自与在相同鞘翅目有害生物物种的不同群体中或在鞘翅目有害生物的不用物种中的一种或多种鞘翅目有害生物内的不同基因座(例如,由seqidno:1和seqidno:2-4中的一项或多项限定的基因座)同源。
在用重组核酸分子直接转化植物外,可以通过将具有至少一种转基因事件的第一植物与缺乏此类事件的第二植物杂交来制备转基因植物。例如,可以将包含编码irna分子的核苷酸序列的重组核酸分子导入适合于转化的第一植物系中,以生成转基因植物,可以将该转基因植物与第二植物系杂交以将编码irna分子的核苷酸序列渐渗入第二植物系的遗传背景中。
本发明的一些实施方案还包括含有本发明的一种或多种序列的商品产品,其从含有本发明的一种或多种核苷酸序列的重组植物或种子生成。含有本发明的一种或多种序列的商品产品意图包括但不限于植物的粗粉、油、碾碎的或完整的谷粒或种子、或包含含有本发明的一种或多种序列的重组植物或种子的任何粗粉、油、或碾碎的或完整的谷粒的任何食物产品。检测本文中涵盖的一种或多种商品或商品产品中的本发明的一种或多种序列是实际的证据,即出于使用dsrna介导的基因阻抑方法控制植物疾病的目的,商品或商品产品由设计为表达本发明的一种或多种核苷酸序列的转基因植物构成。
在一些方面,包括由自经转化的植物细胞衍生的转基因植物生成的种子和商品产品,其中种子或商品产品包含可检测量的本发明的核酸序列。在一些实施方案中,例如,此类商品产品可以通过获得转基因植物并从它们制备食物或饲料来生产。包含本发明的一种或多种核酸序列的商品产品包括例如但不限于:植物的传粉、油、碾碎的或完整的谷粒或种子、和包含含有本发明的一种或多种序列的重组植物或种子的任何粗粉、油、或碾碎的或完整的谷粒的任何食物产品。检测一种或多种商品或商品产品中的本发明的一种或多种序列是实际的证据,即出于控制鞘翅目有害生物的目的,商品或商品产品由设计为表达本发明的一种或多种irna分子的转基因植物构成。
在一些实施方案中,包含本发明核酸分子的转基因植物或种子还可以在其基因组中包含至少一种其它转基因事件,包括但不限于:自其转录靶向鞘翅目有害生物中与由seqidno:1-4限定的基因座不同的基因座的irna分子的转基因事件;自其转录靶向与鞘翅目有害生物不同的生物体(例如,植物寄生性线虫)中的基因的irna分子的转基因事件;编码杀昆虫蛋白(例如,苏云金芽孢杆菌杀昆虫蛋白)的基因;除草剂耐受性基因(例如,提供对草甘膦的耐受性的基因);和促成转基因植物中期望的表型(诸如增加的产量、改变的脂肪酸代谢或细胞质雄性不育的恢复)的基因)。在特定的实施方案中,可以在植物中组合编码本发明irna分子的序列与疾病控制性状和其它昆虫控制性状以实现增强的植物疾病和昆虫损害控制的期望性状。组合采用独特作用模式的昆虫控制性状由于例如对会在田间形成对所述性状的抗性的概率降低而可以提供优于含有单一控制性状的植物的持久性的受保护的转基因植物。
v.鞘翅目有害生物中的靶基因阻抑
a.概述
在本发明的一些实施方案中,可以对鞘翅目有害生物提供至少一种可用于控制鞘翅目有害生物的核酸分子,其中所述核酸分子在鞘翅目有害生物中导致rnai介导的基因沉默。在特定的实施方案中,可以对鞘翅目昆虫宿主提供irna分子(例如dsrna、sirna、mirna、和hprna)。在一些实施方案中,可以通过使核酸分子与鞘翅目有害生物接触对鞘翅目有害生物提供可用于控制鞘翅目有害生物的核酸分子。在这些和其它实施方案中,可用于控制鞘翅目有害生物的核酸分子可以在鞘翅目有害生物的进食基质,例如营养组合物中提供。在这些和其它实施方案中,可以经由摄取被鞘翅目有害生物摄取的包含核酸分子的植物材料提供可用于控制鞘翅目有害生物的核酸分子。在某些实施方案中,核酸分子经由表达导入植物材料中的重组核酸序列而存在于植物材料中,所述导入例如通过用包含重组核酸序列的载体转化植物细胞,并从经转化的植物细胞再生植物材料或全植物来进行。
b.rnai介导的靶基因阻抑
在实施方案中,本发明提供了irna分子(例如,dsrna、sirna、mirna、和hprna),其可以设计用于靶向鞘翅目有害生物(例如wcr或ncr)的基因组和/或cdna文库中的必需天然核苷酸序列(例如,必需基因),例如通过设计包含至少一条包含与靶序列特异性互补的核苷酸序列的链的irna分子进行。如此设计的irna分子的序列可以与靶序列相同,或者可以掺入错配,该错配没有阻止irna分子与其靶序列之间的特异性杂交。
可以在用于鞘翅目有害生物中基因阻抑的方法中使用本发明的irna分子,由此降低由有害生物对植物(例如包含irna分子的受保护的经转化的植物)引起的损害水平或发生率。如本文中所使用的,术语“基因阻抑”指用于降低由于基因转录成mrna及随后mrna翻译生成的蛋白质水平,包括降低自基因或编码序列的蛋白质表达,包括转录后抑制表达和转录阻抑的任何公知方法。转录后抑制通过自靶向阻抑的基因或编码序列转录的整个或部分mrna与用于阻抑的相应irna分子之间的特异性同源性介导。并且,转录后抑制指通过核糖体结合的细胞中可用的mrna量的实质性和可测量降低。
在rnai分子是dsrna分子的实施方案,酶dicer可以将dsrna分子切割成短sirna分子(约20个核苷酸的长度)。通过dicer对dsrna分子的活性生成的双链sirna分子可以分成两个单链sirna;即“乘客链”和“引导链”。乘客链可以受到降解,而引导链可以掺入risc中。通过引导链与mrna分子的特异性互补序列的特异性杂交,随后酶argonaute(catalyticcomponentoftherisc复合物的催化组分)切割而发生转录后抑制。
在本发明的实施方案中,可以使用任何形式的irna分子。本领域技术人员会理解dsrna分子在制备过程中及在对细胞提供irna分子的步骤过程中通常比单链rna分子更稳定,而且在细胞中通常也是更稳定的。如此,尽管sirna和mirna分子例如可以是同等有效,但是在一些实施方案中,dsrna分子可以由于其稳定性而选定。
在特定的实施方案中,提供了包含核苷酸序列的核酸分子,所述核苷酸序列可以在体外表达以生成irna分子,该irna分子与由鞘翅目有害生物基因组内的核苷酸序列编码的核酸分子基本上同源。在某些实施方案中,体外转录的irna分子可以是包含茎-环结构的稳定化的dsrna分子。在鞘翅目有害生物接触体外转录的irna分子后,可以发生对鞘翅目有害生物中靶基因(例如,必需基因)的转录后抑制。
在本发明的一些实施方案中,在用于对鞘翅目有害生物中靶基因的转录后抑制的方法中使用包含核苷酸序列的至少19个连续核苷酸的核酸分子的表达,其中所述核苷酸序列选自下组:seqidno:1;seqidno:1的互补物;seqidno:1的至少19个连续核苷酸的片段;seqidno:1的至少19个连续核苷酸的片段的互补物;包含seqidno:1的根萤叶甲属生物体的天然编码序列;包含seqidno:1的根萤叶甲属生物体的天然编码序列的互补物;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含seqidno:1;根萤叶甲属生物体(例如wcr)的天然编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:1;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:1;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:1;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,所述天然非编码序列包含seqidno:1。在某些实施方案中,可以使用与前述任一项至少80%相同(例如,80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在这些和其它实施方案中,可以表达核酸分子,其与存在于至少一个鞘翅目有害生物细胞中的rna分子特异性杂交。
在这些和其它实施方案中,可以在用于转录后抑制鞘翅目有害生物中靶基因的方法中使用至少一种包含核苷酸序列的至少19个连续核苷酸的别的核酸分子的表达,其中所述核苷酸序列选自下组:seqidno:2;seqidno:2的互补物;seqidno:3;seqidno:3的互补物;seqidno:4;seqidno:4的互补物;seqidno:2-4之任一的至少19个连续核苷酸的片段;seqidno:2-4之任一的至少19个连续核苷酸的片段的互补物;根萤叶甲属生物体(例如wcr)的天然编码序列,其包含seqidno:2-4之任一;根萤叶甲属生物体(例如wcr)的天然编码序列的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列,其包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的互补物,所述天然非编码序列包含seqidno:2-4之任一;根萤叶甲属生物体(例如wcr)的天然编码序列的至少19个连续核苷酸的片段,所述天然编码序列包含seqidno:2-4之任一;根萤叶甲属生物体的天然编码序列的至少19个连续核苷酸的片段的互补物,所述天然编码序列包含seqidno:2-4之任一;转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段,所述天然非编码序列包含seqidno:2-4之任一;和转录成天然rna分子的根萤叶甲属生物体的天然非编码序列的至少19个连续核苷酸的片段的互补物,所述天然非编码序列包含seqidno:2-4之任一。在某些实施方案中,可以使用与前述任一项至少80%相同(例如,80%、约81%、约82%、约83%、约84%、约85%、约86%、约87%、约88%、约89%、约90%、约91%、约92%、约93%、约94%、约95%、约96%、约97%、约98%、约99%、约100%、和100%)的核酸分子的表达。在这些和其它实施方案中,可以表达核酸分子,其与存在于至少一个鞘翅目有害生物细胞中的rna分子特异性杂交。
本发明的一些实施方案的重要特征在于,转录后rnai抑制系统能够容许靶基因间由于遗传突变、品系多态性、或进化趋异而可以预期的序列变异。导入的核酸分子可以不需要是与靶基因的一级转录产物或经完全加工的mrna绝对同源的,只要导入的核酸分子与靶基因的一级转录产物或经完全加工的mrna特异性可杂交。此外,相对于靶基因的一级转录产物或经完全加工的mrna,导入的核酸分子可以不需要是全长。
使用本发明的irna技术抑制靶基因是序列特异性的;即靶向与irna分子基本上同源的核苷酸序列进行遗传抑制。在一些实施方案中,可以使用包含与靶基因序列的一部分相同的核苷酸序列的rna分子进行抑制。在这些和其它实施方案中,可以使用包含相对于靶基因序列具有一处或多处插入、缺失和/或点突变的核苷酸序列的rna分子。在特定的实施方案中,irna分子和靶基因的一部分可以共享例如至少自约80%、至少自约81%、至少自约82%,至少自约83%、至少自约84%、至少自约85%、至少自约86%、至少自约87%、至少自约88%、至少自约89%、至少自约90%、至少自约91%、至少自约92%、至少自约93%、至少自约94%、至少自约95%、至少自约96%、至少自约97%、至少自约98%、至少自约99%、至少自约100%、和100%序列同一性。或者,dsrna分子的双链体区可以与靶基因转录物的一部分特异性可杂交。在特异性可杂交的分子中,展现出较大同源性的小于全长的序列补偿较长的、不太同源的序列。dsrna分子的双链体区中与靶基因转录物的一部分相同的核苷酸序列的长度可以是至少约25、50、100、200、300、400、500、或至少约1000个碱基。在一些实施方案中,可以使用大于20-100个核苷酸的序列。在特定的实施方案中,可以使用大于约200-300个核苷酸的序列。在特定的实施方案中,根据靶基因的大小,可以使用大于约500-1000个核苷酸的序列。
在某些实施方案中,可以将鞘翅目有害生物中靶基因的表达在鞘翅目有害生物细胞内抑制至少10%;至少33%;至少50%;或至少80%,使得发生显著抑制。显著抑制指高于阈值的抑制,所述阈值导致可检测的表型(例如,生长停止、进食停止、发育停止、和昆虫死亡率,等等),或与所抑制的靶基因对应的rna和/或基因产物的可检测降低。虽然在本发明的某些实施方案中,在基本上所有鞘翅目有害生物细胞中发生抑制,在其它实施方案中,仅在表达靶基因的细胞子集中发生抑制。
在一些实施方案中,转录阻抑由细胞中dsrna分子的存在介导,所述dsrna分子与启动子dna序列或其互补物展现出实质性序列同一性以实现称为“启动子反式阻抑”的事情。基因阻抑针对鞘翅目有害生物中的靶基因可以是有效的,所述鞘翅目有害生物可以例如通过摄取或接触含有dsrna分子的植物材料而摄取或接触此类dsrna分子。启动子反式阻抑中使用的dsrna分子可以特异性设计为抑制或阻抑鞘翅目有害生物细胞中一种或多种同源或互补序列的表达。美国专利5,107,065;5,759,829;5,283,184;和5,231,020中披露了通过反义或有义取向的rna进行转录后基因阻抑以调节植物细胞中的基因表达。
c.对鞘翅目有害生物提供的irna分子的表达
可以在许多体外或体内形式之任一种中实施表达用于在鞘翅目有害生物中rnai介导的基因抑制的irna分子。然后,可以对鞘翅目有害生物提供irna分子,例如通过使irna分子与有害生物接触,或者通过引起有害生物摄取或以其它方式内在化irna分子进行。本发明的一些实施方案包括鞘翅目有害生物的经转化的宿主植物、经转化的植物细胞、和经转化的植物的后代。经转化的植物细胞和经转化的植物可以工程化改造为例如在异源启动子控制下表达一种或多种irna分子,以提供有害生物保护效应。如此,在鞘翅目有害生物在进食期间食用转基因植物或植物细胞时,有害生物可以摄取转基因植物或细胞中表达的irna分子。也可以将本发明的核苷酸序列导入极其多种原核和真核微生物宿主中以生成irna分子。术语“微生物”包括原核和真核物种,诸如细菌和真菌。
基因表达的调控可以包括对此类表达的部分或完全阻抑。在另一个实施方案中,用于阻抑鞘翅目有害生物中基因表达的方法包括在有害生物的宿主组织中提供基因阻抑量的至少一种自如本文中所描述的核苷酸序列转录的dsrna分子,其至少一个区段与鞘翅目有害生物细胞内的mrna序列互补。依照本发明的病原性微生物摄取的dsrna分子,包括其经修饰形式诸如sirna、mirna、或hprna分子可以与自包含选自下组的核苷酸序列的核酸分子转录的rna分子至少约80%、81%、82%、83%、84%、85%、86%、87%、88%、89%、90%、91%、92%、93%、94%、95%、96%、97%、98%、99%、或约100%相同:seqidno:1-4。因此,提供了用于转录本发明的dsrna分子的分离的且基本上纯化的核酸分子,包括但不限于非天然存在的核苷酸序列和重组dna构建体,其在导入鞘翅目有害生物时阻抑或抑制其中的内源编码序列或靶编码序列的表达。
特定的实施方案提供了用于投递irna分子以转录后抑制植物寄生性鞘翅目有害生物中的一种或多种靶基因并控制植物寄生性鞘翅目有害生物群体的投递系统。在一些实施方案中,所述投递系统包含摄取包含在宿主细胞中转录的rna分子的宿主转基因植物细胞或宿主细胞内容物。在这些和其它实施方案中,创建转基因植物细胞或转基因植物,其含有转录本发明的稳定化dsrna分子的重组dna构建体。可以如下生成包含编码特定irna分子的核酸序列的转基因植物细胞和转基因植物,即采用重组dna技术(该基础技术是本领域中公知的)以构建包含编码本发明的irna分子(例如,稳定化的dsrna分子)的核苷酸序列的植物转化载体;以转化植物细胞或植物;并且以生成含有转录的irna分子的转基因植物细胞或转基因植物。
为了对转基因植物赋予鞘翅目有害生物抗性,可以例如将重组dna分子转录成irna分子,诸如dsrna分子、sirna分子、mirna分子、或hprna分子。在一些实施方案中,自重组dna分子转录的rna分子可以在重组植物的组织或流体内形成dsrna分子。此类dsrna分子可以包含在核苷酸序列的一部分中,其与自可以寄生宿主植物的类型的鞘翅目有害生物内的dna序列转录的相应核苷酸序列相同。鞘翅目有害生物内靶基因的表达受到dsrna分子阻抑,并且鞘翅目有害生物中靶基因表达的阻抑生成对鞘翅目有害生物有抗性的转基因植物。已经显示了dsrna分子的调控效应可适用于有害生物中表达的多种基因,包括例如负责细胞代谢或细胞转化的内源基因,包括持家基因;转录因子;蜕皮相关基因;和编码牵涉细胞代谢或正常生长和发育的多肽的其它基因。
对于自体内转基因或表达构建体转录,可以在一些实施方案中使用调节区(例如,启动子、增强子、沉默子、和多聚腺苷酸化信号)以转录一条或多条rna链。因此,在一些实施方案中,如上文所列的,在生成irna分子中使用的核苷酸序列可以与在植物宿主细胞中功能性的一种或多种启动子序列可操作连接。启动子可以是通常驻留于宿主基因组中的内源启动子。在可操作连接的启动子序列控制下的本发明的核苷酸序列侧翼可以进一步有有利地影响其转录和/或所得转录物的稳定性的别的序列。此类序列可以位于可操作连接启动子的上游、表达构建体3’端的下游、并且可以同时存在于启动子的上游和表达构建体3’端的下游。
一些实施方案提供了用于降低由以植物为食的鞘翅目有害生物引起的对宿主植物(例如玉米植物)的损害的方法,其中所述方法包括在宿主植物中提供表达本发明的至少一种核酸分子的经转化的植物细胞,其中所述核酸分子在被鞘翅目有害生物摄取后发挥功能以抑制鞘翅目有害生物内靶序列的表达,该表达抑制导致鞘翅目有害生物的死亡率、降低的生长、和/或降低的生殖,由此降低由鞘翅目有害生物引起的对宿主植物的损害。在一些实施方案中,核酸分子包含dsrna分子。在这些和其它实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交的核苷酸序列。在一些实施方案中,核酸分子由一种核苷酸序列组成,所述核苷酸序列与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交。
在一些实施方案中,提供了用于提高玉米作物产量的方法,其中所述方法包括将本发明的至少一种核酸分子导入玉米植物中;栽培玉米植物以容许表达包含核酸序列的irna分子,其中包含核酸序列的irna分子的表达抑制鞘翅目有害生物损害和/或生长,由此降低或消除由于鞘翅目有害生物侵袭所致的产量损失。在一些实施方案中,irna分子是dsrna分子。在这些和其它实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交的核苷酸序列。在一些实施方案中,核酸分子由一种核苷酸序列组成,所述核苷酸序列与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交。
在一些实施方案中,提供了用于调控鞘翅目有害生物中靶基因表达的方法,所述方法包括:用包含编码本发明的至少一种核酸分子的核酸序列的载体转化植物细胞,其中所述核苷酸序列与启动子和转录终止序列可操作连接;在足以容许形成包含多个经转化的植物细胞的植物细胞培养物的条件下培养经转化的植物细胞;选择已经将核酸分子整合入其基因组中的经转化的植物细胞;对经转化的植物细胞筛选由整合的核酸分子表达的irna分子的表达;选择表达irna分子的转基因植物细胞;并以选择的转基因植物细胞喂养鞘翅目有害生物。也可以自表达由整合的核酸分子编码的irna分子的经转化的植物细胞再生植物。在一些实施方案中,irna分子是dsrna分子。在这些和其它实施方案中,核酸分子包含dsrna分子,所述dsrna分子各自包含超过一种与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交的核苷酸序列。在一些实施方案中,核酸分子由一种核苷酸序列组成,所述核苷酸序列与鞘翅目有害生物细胞中表达的核酸分子特异性可杂交。
可以将本发明的irna分子作为来自掺入植物细胞基因组中的重组基因的表达产物或者掺入应用于种植前种子的涂层或种子处理中在植物物种(例如玉米)的种子之内或之上掺入。认为包含重组基因的植物细胞是转基因事件。还包括用于将irna分子投递至鞘翅目有害生物的投递系统。例如,可以将本发明的irna分子直接导入鞘翅目有害生物细胞中。用于导入的方法可以包括直接混合irna与来自鞘翅目有害生物宿主的植物组织,以及对宿主植物组织应用包含本发明的irna分子的组合物。例如,将irna分子喷雾到植物表面上。或者,可以通过微生物表达irna分子,并且可以将微生物应用到植物表面上,或者通过物理手段诸如注射导入根或茎中。如上文所讨论的,也可以将转基因植物遗传工程化改造为以如下的量表达至少一种irna分子,所述量足以杀死已知侵袭植物的鞘翅目有害生物。通过化学或酶促合成生成的irna分子也可以以与常见的农业实践一致的方式配制,并且作为喷射产物使用以控制植物疾病。配制剂可以包含有效的叶覆盖需要的合适粘着剂(sticker)和湿润剂,以及保护irna分子(例如,dsrna分子)免于uv损伤的uv保护剂。此类添加剂通常在生物杀虫剂产业中使用,并且是本领域技术人员公知的。此类应用可以与其它喷射杀虫剂应用(基于生物学的或其它方面的)组合以增强针对鞘翅目有害生物的植物保护。
本文中引用的所有参考文献,包括出版物、专利、和专利申请在此通过提及以它们不与本公开内容的明确详细不一致的程度收录,并且因此以如下的程度收录,就像个别且明确指出每篇参考文献通过提及收录,并在本文中完整列出一样。本文中讨论的参考文献仅提供其在本申请提交日前的公开内容。凭借在先发明,本文中的任何文字都不应解释为承认发明人没有资格早于所述公开内容。
提供以下实施例以例示某些特定的特征和/或方面。这些实施例不应解释为将公开内容限制为所描述的特定特征或方面。
实施例
实施例1:材料和方法
样品制备和生物测定法。
合成许多dsrna分子(包括cafl-180;液泡atp酶(v-atpase)亚基c区1;v-atp酶亚基c区2;v-atp酶亚基h区1;v-atp酶亚基h区2;和rho1),并使用
在人工昆虫饮食上用新生昆虫幼虫进行的生物测定法中对样品测试昆虫活性。wcr卵获自cropcharacteristics、inc.(farmington、mn)。
在特别为昆虫生物测定法设计的128孔塑料盘(c-dinternational、pitman、nj)中进行生物测定法。每个孔含有约1.0ml为鞘翅目昆虫生长设计的饮食。通过移液到每孔的1.5cm2饮食表面(40μl/cm2)投递60μl蛋白质样品等分试样。dsrna样品浓度以孔中每立方厘米表面积的dsrna量(ng/cm2)计算。将经处理的盘在通风橱中保持,直到饮食表面上的液体蒸发或者吸收到饮食中。
在孵化的几个小时内,用湿润的骆驼毛刷挑出个别幼虫,并在经处理的饮食上放置(每孔1或2个幼虫)。然后,将128孔塑料盘的侵袭孔用透明塑料的粘合片密封,并开孔以容许气体交换。将生物测定盘在受控的环境条件(28°,约40%相对湿度,16:8(光照:黑暗))下保持9天,在该时间后,记录暴露于每种样品的昆虫总数、死亡昆虫数目、和存活昆虫重量。对每个处理计算百分比死亡率、平均活体重量、和生长抑制。萎缩(stunting)定义为平均活体重量的降低。生长抑制(gi)如下计算:
gi=[1–(twit/tnit)/(twibc/tnibc)]
其中twit是处理中的活体昆虫总重量,
tnit是处理中的昆虫总数,
twibc是背景检验(缓冲液对照)中的活体昆虫总重量,且
tnibc是背景检验(缓冲液对照)中的昆虫总数。
gi50测定为gi值是50%时饮食中样品的浓度。lc50(50%致死浓度)以50%测试昆虫被杀死时饮食中样品的浓度计算。使用jmp软件(sas、cary、nc)完成统计学分析。
重复的生物测定法表明样品的摄取导致玉米根虫幼虫的令人惊讶的且意料不到的死亡率和生长抑制。
实施例2:候选靶基因的鉴定
选择第一龄wcr幼虫进行转录物组分析,因为在此生长阶段通过转基因昆虫抗性技术的控制会是有利的。
从约0.9gm整个第一龄wcr幼虫(玉米根萤叶甲;孵化后4至5天;于16℃保持)分离总rna,并使用以下基于酚/tri
将幼虫于室温在具有10mltri
将上清液小心取出并弃去,并将rna团粒通过用75%乙醇涡旋振荡清洗两次,在每次清洗后通过以7,500xg(4℃或25℃)离心5分钟回收。将乙醇小心除去,并容许团粒风干3至5分钟,然后在无核酸酶的无菌水中溶解。通过于260nm和280nm测量吸光度(a)测定rna浓度。自约0.9g幼虫的典型提取产生超过1mg总rna,其中a260/a280比是1.9。如此提取的rna于-80℃贮存,直到进一步加工。
通过将等分试样运行通过1%琼脂糖凝胶确定rna质量。在经高温灭菌的容器中使用用经depc(二乙基焦碳酸盐)处理的水稀释的经高温灭菌的10倍tae缓冲液(tris乙酸盐edta、1倍浓度是0.04mtris乙酸盐,1mmedta(乙二胺四乙酸钠盐,ph8.0)生成琼脂糖凝胶溶液。使用1倍tae作为运行缓冲液。在使用前,用rnaseawaytm(invitrogeninc.、carlsbad、ca)清洁电泳槽和孔形成梳。将2μlrna样品与8μlte缓冲液(10mmtrishclph7.0;1mmedta)和10μlrna样品缓冲液(
标准化的cdna文库使用随机化引发由商业服务提供商(eurofinsmwgoperon、huntsville、al)自幼虫总rna制备。
标准化的幼虫cdna文库由eurofinsmwgoperon的gsflx454titaniumtmserieschemistry以1/2板规模测序,其产生具有平均阅读长度348bp的超过600,000个阅读。将350,000个阅读装配成超过50,000个重叠群。使用公众可得到的程序formatdb(可获自国立生物信息中心(ncbi))将未装配的阅读和重叠群两者转换成可blast的数据库。
使用关于特定基因在其它昆虫诸如果蝇和拟谷盗属(tribolium)中的致死rnai效应的信息选择rnai靶向的候选基因。假设这些基因对于鞘翅目昆虫的存活和生长是必需的。在转录物组序列数据库中鉴定选择的靶基因同系物,如下文所描述的。通过pcr扩增靶基因的全长或部分序列以制备双链rna(dsrna)生成的模板。
针对含有未装配的根萤叶甲序列阅读或配装的重叠群的可blast的数据库运行使用候选蛋白质编码序列的tblastn搜索。针对ncbi非冗余数据库使用blastx确认显著命中根萤叶甲序列(定义为对于重叠群同源性,好于e-20,且对于未装配的序列阅读同源性,好于e-10)。此blastx搜索的结果确认tblastn搜索中鉴定的根萤叶甲同系物候选基因序列的确包含根萤叶甲基因,或者是根萤叶甲序列中对非根萤叶甲候选基因序列的可得最佳命中。在大多数情况中,注释为编码蛋白质的拟谷盗候选基因给出与根萤叶甲转录物组序列中的一种或多种序列的明确序列同源性。在几个情况中,清楚的是通过与非根萤叶甲候选基因的同源性选择的一些根萤叶甲重叠群或未装配的序列阅读重叠,而且重叠群的装配已经不能连接这些重叠。在那些情况中,使用sequenchertmv4.9(genecodescorporation、annarbor、mi)来将序列装配成更长的重叠群。
多种候选靶基因鉴定为可以导致鞘翅目有害生物死亡率或wcr中的生长、发育、或生殖抑制的基因,包括seqidno:1-4。使用根萤叶甲属候选基因同系物序列的全长或部分克隆来生成pcr扩增子以合成dsrna。
seqidno:1对应于d_vir_c47185,其是caf1-180的同系物,并且在本文中称为caf1-180。已经显示了果蝇(drosophila)caf1-180在将组蛋白投递至新合成的dna和caf1-180上方面发挥功能。已经显示了果蝇caf1-180的功能丧失突变引起幼虫生长和发育的停滞,和幼虫死亡率(klapholz等(2009)chromosoma118(2):235-48;tyler等(2001)mol.cellbiol.21(19):6574-84;song等(2007)dev.biol.311(1):213-22)。
seqidno:2对应于d_vir_c1229,其是液泡atp酶亚基c的同系物,并且在本文中称为vatpasec。seqidno:5-8是vatpasec的非连续序列片段。
seqidno:3对应于d_vir_c1319,其是液泡atp酶亚基h的同系物,并且在本文中称为vatpaseh。seqidno:9-11是vatpaseh的非连续序列片段。
果蝇液泡atp酶是一种牵涉notch信号传导的多亚基质子转运蛋白(vaccari等(2010)development137(11):1825-32)。已经显示了果蝇液泡atp酶亚基的功能丧失突变是隐性致死的(allan等(2005)physiol.genomics22(2):128-38;及davies等(1996)j.biol.chem.271(48):30677-84)。vatpasec(vha44)和vatpaseh(vhasfd)是果蝇液泡atp酶的两个亚基。
seqidno:4是contig_01_rho1_1-191_cdc42,本文中称为rho1。果蝇rho1在调节肌动蛋白丝的装配和拆卸中发挥功能,并且果蝇rho1的功能丧失突变对细胞组分运动、突触重塑、幼虫发育、和对应激的响应具有重大影响(fox等(2005)development132(21):4819-31;kadandale等(2010)proc.natl.acad.sci.u.s.a.107(23):10502-7;rosales-nieves等(2006)nat.cellbiol.8(4):367-76;magie和parkhurst(2005)dev.biol.278(1):144-54;及xu等(2008)dev.biol.321(1):88-100)。
实施例3:靶基因的扩增
引物设计为通过pcr扩增每个靶基因的编码区的部分。参见表1。在适当的情况下,将t7噬菌体启动子序列(ttaatacgactcactatagggaga(seqidno:12))掺入扩增的有义或反义链的5’端中。参见表1。自wcr提取基因组dna,并使用pcr引物经由pcr反应从基因组dna扩增整个或部分天然靶基因序列。
表1:用于制备供dsrna生成用的模板的pcr引物的序列和配对。
实施例4:rnai构建体
通过pcr制备模板和dsrna合成。
图1中显示了用于提供供体外dsrna生成用的特异性模板的策略。使用表1中的引物对和(作为pcr模板)从分离自wcr一龄幼虫的总rna制备的第一链cdna通过pcr制备意图在dsrna合成中使用的模板dna。对于每个选择的靶基因区,实施两个分开的pcr扩增。第一个pcr扩增在扩增的有义链的5’端引入t7启动子序列。第二个反应在反义链的5’端掺入t7启动子序列。然后,将每个靶基因区的两个pcr扩增片段以大致相等的量混合,并使用混合物作为转录模板以生成dsrna。图1。合成双链rna,并使用
植物转化载体的构建:方法1
使用标准的多位点
还构建为生成hprna设计的第三类进入载体,其利用caf1-180基因或vatpasec基因的片段。使用表2中所列的引物扩增caf1-180和vatpasec的靶基因区以合成发夹。由rna前转录物进行的分子内发夹形成通过以彼此相反的取向排列(在单一转录单元内)两个拷贝的靶基因片段促进,两个片段以st-ls1内含子序列分开(vancanneyt等(1990)mol.gen.genet.220(2):245-50)。通过玉米泛素1启动子(美国专利5,510,474)拷贝的驱动mrna前转录物的生成。如此,mrna前转录物含有作为彼此的大反向重复的两个基因片段序列,其以内含子序列分开。使用包含来自玉米肪酶基因的3’非翻译区(zmlip3’utr、美国专利7,179,902)的片段终止基因的转录。
表2:用于制备供玉米转化用的发夹构建体的引物的序列和配对。
seqidno:49代表caf1-180发夹形成序列,而seqidno:50代表vatpasec发夹形成序列。
对caf1-180和vatpasec之每种构建hprna表达进入载体的两种型式。在进入载体的第一种型式中,玉米共有翻译起始背景序列存在于mrna5’非翻译前导序列中,所述mrna5’非翻译前导序列与第一靶基因片段序列的5’端相邻,所述第一靶基因片段序列以“直接”取向(也就是相对于启动子处于有义取向)存在。在第二种型式中省略玉米共有翻译起始序列。
使用来自invitrogen的lrclonaseiiplustm用3种进入载体和一种目的地载体(destinationvector)(pdab104124)实施标准的
在已经用bcli(对于caf1-180构建体)或bbsi加afei(对于vatpasec构建体)消化yfp/hprna/pat表达载体dna以除去载体主链中存在的细菌壮观霉素抗性基因后通过高压液相层析(hplc)以制备规模完成片段纯化,用于whiskerstm介导的转化。
植物转化载体的构建:方法2
whiskerstm目的地载体:使用标准的一位点
构建为生成hprna设计的第一种进入载体,其利用携带caf1-180v3有义序列(+st-ls1内含子)(seqidno:95)的dna片段和携带caf1-180v3反义序列(seqidno:96)的片段。这些片段分别由商业厂商(dna2.0;menlopark、ca)合成。通过标准的克隆方法以合适的取向连接合成的各部分以形成seqidno:97的发夹形成构建体,并在进入载体中在启动子和3’utr序列之间克隆构建体。mrna前转录物的生成由玉米泛素1启动子(美国专利5,510,474)拷贝的驱动,而使用包含来自玉米过氧化物酶5基因的3’非翻译区(zmper53’utrv2、美国专利6,699,984)的序列终止靶基因片段的转录。由rna前转录物进行的分子内发夹形成通过以彼此相反的取向排列(在单一转录单元内)两个拷贝的靶基因片段促进,两个片段以st-ls1内含子序列分开(vancanneyt等(1990)mol.gen.genet.220(2):245-50)。如此,mrna前转录物含有作为彼此的大反向重复的两个基因片段序列,其以内含子序列分开。
使用来自invitrogen的lrclonaseiiplustm用构建的caf-180v3发夹进入载体和目的地载体pdab108916实施标准的
以类似的方式,使用标准的一位点
通过标准克隆方法构建为生成hprna设计的第二种进入载体,其利用包含以合适的取向连接的vatpasecv3有义序列(+st-ls1内含子)序列(seqidno:98)和vatpasecv3反义序列(seqidno:99)的合成片段(seqidno:100)。使用vaptasecv3进入载体进行用目的地载体(pdab108916)的
通过标准克隆方法构建为hprna生成设计的第三种进入载体,其利用包含以合适的取向连接的vatpasecv4有义(+st-ls1内含子)序列(seqidno:101)和vatpasecv4反义序列(seqidno:102)的合成片段(seqidno:103)。使用vatpasecv4进入载体进行用目的地载体(pdab108916)的
通过标准克隆方法构建为hprna生成设计的第四种进入载体,其利用包含以合适的取向连接的vatpasehv1有义(+st-ls1内含子)序列(seqidno:104)和vatpasehv1反义序列(seqidno:105)的合成片段(seqidno:106)。使用vatpasehv1进入载体进行用目的地载体(pdab108916)的
通过标准克隆方法构建为hprna生成设计的第五种进入载体,其利用包含以合适的取向连接的rho1v1有义(+st-ls1内含子)序列(seqidno:107)和rho1v1反义序列(seqidno:108)的合成片段(seqidno:109)。使用rho1v1进入载体进行用目的地载体(pdab108916)的
使用纯化的环状质粒dna完成whiskerstm介导的转化。实施例9.
土壤杆菌目的地载体:使用标准的
为了转化玉米,将hprna二元表达载体导入解除的(disarmed)根癌土壤杆菌(agrobacteriumtumefaciens)菌株dat13192(如记载于2011年7月29日提交的pct国际申请no.pct/us11/046028的)中。
实施例5:候选靶基因的筛选
设计为抑制实施例2中鉴定的一些但不是所有靶基因序列的合成dsrna在基于饮食的测定法中在对wcr施用时引起死亡率和生长抑制。观察到cafl-180;vatpasec;vatpaseh;和rho1在此测定法中比筛选的其它dsrna展现出大大升高的效力。
重复的生物测定法证明摄取转录成cafl-180(seqidno:110)dsrna;vatpasec区1(seqidno:111)dsrna;vatpasec区1短(seqidno:112)dsrna;vatpasec区2(seqidno:113)dsrna;vatpaseh区1(seqidno:114)dsrna;vatpaseh区2(seqidno:115)dsrna;及rho1(seqidno:116)dsrna的构建体各自产生强力得令人惊讶的西部玉米根虫幼虫死亡率和生长抑制。表3和4显示了对这些dsrna暴露9天后wcr幼虫的基于饮食的进食生物测定法的结果。图2-4显示了用例示性核酸分子处理的鞘翅目有害生物的死亡率和生长抑制的变化性图。
表3:西部玉米根虫饮食进食生物测定法(进食9天后)的结果。
*剂量单位是ng/cm2
**nd=未测定
表4:西部玉米根虫饮食进食生物测定法(进食9天后)的结果。
*rho1死亡率计数与te缓冲液或水计数显著不同(似然比p<0.0001)。
先前已经提出了可以采用根萤叶甲属物种的某些基因进行rnai介导的昆虫控制。参见美国专利公开文本no.2007/0124836(其披露了906种基因),及美国专利7,614,924(其披露了9,112种基因)。然而,确定的是,提示具有rnai介导的昆虫控制效用的许多基因在控制根萤叶甲中不是有效的。还确定与提示具有rnai介导的昆虫控制效用的其它基因相比,cafl-180、vatpasec、vatpaseh、和rho1各自提供对根萤叶甲的令人惊讶的且意料不到的卓越控制。
美国专利7,614,924中提出膜联蛋白、beta血影蛋白2、和mtrp-l4各自在rnai介导的昆虫控制中是有效的。seqidno:51是膜联蛋白区1的dna序列,而seqidno:52是膜联蛋白区2的dna序列。seqidno:53是beta血影蛋白2区1的dna序列,而seqidno:54是beta血影蛋白2区2的dna序列。seqidno:55是mtrp-l4区1的dna序列,而seqidno:56是mtrp-l4区2的dna序列。还使用yfp序列(seqidno:57)来生成dsrna作为阴性对照。
使用这些序列之每种通过实施例4的方法生成dsrna,并且dsrna各自通过上文所描述的相同的基于饮食的生物测定法测试。表5列出了用于生成膜联蛋白、beta血影蛋白2、和mtrp-l4dsrna分子的引物的序列。表6呈现了在对这些dsrna分子暴露9天后wcr幼虫的基于饮食的进食生物测定法的结果。重复生物测定法证明了摄取这些dsrna没有产生对上述西部玉米根虫幼虫的死亡率或生长抑制,这用te缓冲液、yfp、或水对照样品看到。
表5:用于扩增膜联蛋白、beta血影蛋白2、mtrp-l4、和yfp的基因区以合成dsrna的引物的序列和配对。
表6:西部玉米根虫幼虫dsrna饮食进食生物测定法(进食9天后)的结果。
实施例6:序列长度对vatpasecdsrna效力的影响
大小范围为349至500bp的液泡atp酶亚基c(vatpasec)的dsrna在针对西部玉米根虫幼虫的饮食进食测定法中是有活性的。表3。为了测定dsrna分子效力的长度依赖性,改变vatpasecdsrna的长度,并测试。序列选择为代表vatpasec编码序列(seqidno:2)的174个碱基对区段的5’端15、25、50和100个碱基、和3’端100、50、25、和15个碱基。
代表15bp和25bp5’和3’vatpasec序列的dsrna片段由商业厂商(integrateddnatechnologies;coralville、ia)合成。表7。使用表8中列出的引物使用t7rna聚合酶遵循megascripttmrnai试剂盒(
表7:设计用于充当15bp和25bp(平端的)vatpasecdsrna的合成rna。
表8:用于生成各种长度的包含t7启动子序列(加下划线)的vatpasecdna模板以合成dsrna的pcr引物。
表9:在pcr中用于生成vatpasecdna模板以合成各种长度的dsrna的引物组合
*在两种引物的5'端添加的t7启动子序列(24bp),导致对扩增子添加48bp。
**在转录期间在dsrna的两条链的5’端添加来自t7启动子序列的gggaga,导致对转录物添加12bp。
用西部玉米根虫幼虫使用饮食进食测定法以单剂(1000ng/cm2)对vatpasecdsrna(即vatpasec5'-15、vatpasec5'-25、vatpasec5'-50、vatpasec5'-100、vatpasec3'-15、vatpasec3'-25、vatpasec3'-50、vatpasec3'-100、和vatpasec-174)各自测试活性(致死性)。如表10中显示的,这些dsrna样品的活性(如通过进食第9天的百分比幼虫死亡率测量的)是不同的。进食15bp和25bpvatpasecdsrna样品(5'和3'两者)没有诱导显著的幼虫死亡率,其是与使用yfpdsrna的阴性对照相似的结果。然而,100bp的5'和3'vaptasecdsrna,以及174bp的5'和3'vaptasecdsrna两者产生类似地意料不到的高幼虫死亡率。两种50bpdsrna样品在进食测定法中显示不同结果;vatpasec5'-50dsrna样品没有产生显著的幼虫死亡率,但是显著显著的萎缩,而vaptasec3'-50dsrna样品引起几乎60%的死亡率,其是几乎与用100bp和174bpdsrna的样品获得的数值一样高的水平。表10。
此外,在六种测试剂量(1.0、3.9、62.5、250、和1000ng/cm2)测定vatpasecdsrna的活性以获得相对lc50和gi50值。表11。用vatpasec3'-100样品获得的lc50值是3.01ng/cm2,其是观察到的最低数值(即,最高效力)。特别注意到用vatpasec-174样品获得的lc50值9.81ng/cm2。设计为在玉米植物中生成vatpasecv4发夹的构建体在用温室培养的植物测试时显示活性。实施例10;表18。所述vaptasecv4序列片段(166bp)与本文描述的vatpasec-174序列几乎是相同的;本文描述的vatpasec-174序列在166bpvaptasecv4序列片段的5'端仅缺乏8bp(atgactga)。
表10:以1000ng/cm2利用不同长度的vatpasecdsrna的西部玉米根虫幼虫dsrna饮食进食生物测定法(进食9天后)的结果。
*后面有相同字母的均值%死亡率数值在p=0.05水平彼此不是显著不同的(如anova/tukey均值比较计算的)。没有dsrna的阴性对照(te缓冲液和水)的死亡率数值小于20%,并且没有显示。
**后面有相同字母的均值%gi值在p=0.05水平彼此不是显著不同的(通过anova/tukey均值比较计算)。使用阴性对照yfpdsrna喂养的幼虫的幼虫重量作为gi计算的基础。
表11:以1.0、3.9、62.5、250、和1000ng/cm2对西部玉米根虫幼虫提供的不同长度的vatpasecdsrna(在食用9天后)的lc50和gi50值。
实施例7:dsrna序列变异对vatpasecdsrna效力的影响
与靶序列不完全互补的dsrna分子(例如,与靶mrna仅具有95%同一性)有效控制鞘翅目有害生物。将非互补碱基引入vatpasecdsrna的有义链或反义链中,导致dsrna的有义和反义链之间的错配(因此,导致dsrna和体内靶mrna之间的错配)。在饮食生物测定法中测量对针对西部玉米根虫幼虫的不完全匹配的dsrna的杀昆虫活性的影响。
一次仅突变编码vatpasecdsrna的两条dna链之一,并且在每个突变位点处人工引入单一非互补核苷酸。突变位点沿着编码vatpasecdsrna的dna的突变链的整个长度以20个核苷酸的间隔任意定位。从不互补突变位点相反的非突变链上的核苷酸的突变位点任一侧的近侧核苷酸产生选择用于替换天然核苷酸的核苷酸的身份。在三个或更多个核苷酸在给定位置中相同的事件中,选择最近的不同核苷酸替换天然核苷酸。
设计两种不完全互补的、基本上同源的vatpasec(v4)dsrna(即v4.1和v4.2),将其生成,并测试杀昆虫活性。这两种dsrna都包含166bp的vatpasecv4序列,及每条rna链5'端的6个核苷酸(gggaga),其是通过t7启动子进行转录的典型产物。如此,每种dsrna的总长度是178bp。
设计并生成基本上同源的vatpasecv4.1dsrna,其具有导入反义链中的非互补碱基。此dsrna通过下列步骤实现:(1)反义链序列设计为具有在每20个核苷酸处引入的突变;(2)将包含突变反义链dna序列或天然有义链dna序列(即,没有突变)的dna片段化学合成(idt);(3)将包含突变反义链的dna寡核苷酸和包含有义链的寡核苷酸退火以形成双链分子;(3)采用包含5'端t7启动子序列的引物使用pcr扩增将t7启动子序列掺入两条链的5'端中,由此为dsrna合成提供足够的模板;并(4)使用t7rna聚合酶和
通过上述方法的合适修改获得基本上同源的vatpasecv4.2dsrna,其在有义链序列中具有20个碱基间隔处引入的突变(但是具有天然反义链序列)。此外,将包含天然序列(即,具有完全互补的有义和反义链)的vatpasecv4dsrna构建,并作为对照使用,用于在针对西部玉米根虫幼虫的基于饮食的进食测定法中(使用先前描述的方法进行)与基本上同源的vatpasecv4.1和v4.2dsrna的效力比较。
用西部玉米根虫第一龄幼虫在进食生物测定法中测试7种剂量的这些dsrna(0.2、1.0、3.9、15.6、62.5、250、和1000ng/cm2)以获得lc50和gi50值。将该测定法独立进行两次,并且每个实验具有两个重复。
对基本上同源的vatpasecv4.1(反义链错配的)dsrna计算的lc50是780.9ng/cm2,其比对天然(即,非突变的)vatpasecv4dsrna得到的lc50高7.9倍。表12。此结果指示即使vatpasecv4.1dsrna的178个碱基的反义链含有8个突变碱基,存在着与鞘翅目有害生物靶mrna链的足够同源性以在摄取dsrna后诱导致死效应。对基本上同源的vatpasecv4.2(有义链错配的)dsrna计算的lc50值是868.6ng/cm2,其与用vatpasecv4.1获得的数值相似,且比在这些测定法中对vatpasecv4dsrna获得的数值高8.8倍。这些结果证明包含dsrna有义链错配的基本上同源的dsrna与鞘翅目有害生物靶mrna链保持足够的同源性以在摄取dsrna后诱导致死效应。进一步发现了,在这些进食测定法中,vatpasecv4.1和v4.2dsrna具有的gi50值比对天然(即,非突变)vatpasecv4dsrna计算的数值高。表12。因此,看出vatpasecv4.1和4.2dsrna在被鞘翅目有害生物摄取后能够诱导改善的生长抑制和改善的死亡率两者。
表12:基本上同源的vatpasecdsrna(v4.1和v4.2),和天然vatpasecv4dsrna针对西部玉米根虫幼虫的杀昆虫活性。
实施例8:土壤杆菌介导的包含杀昆虫发夹dsrna的转基因玉米组织的生成
穗灭菌和胚分离。从温室中种植的玉蜀黍近交系b104植物获得未成熟的玉米胚,并将其自花或近缘传粉以生成穗。在传粉后约9至12天收获穗。在实验当天,通过在次氯酸钠(6.15%)的20%溶液中浸没,并摇动20至30分钟,接着在无菌水中漂洗三次将穗表面灭菌。灭菌后,将未成熟的合子胚(1.5至2.4mm)自每个穗中在无菌条件下切出,并随机分配到微量离心管中,所述微量离心管含有液体接种培养基(2.2gm/lms盐(frame等、2011、见上文);1倍isu改良ms维生素(frame等(2011)“genetictransformationusingmaizeimmaturezygoticembryos,”于plantembryoculturemethodsandprotocols:methodsinmolecularbiology.t.a.thorpe和e.c.yeung、(编)、springerscienceandbusinessmedia、llc.、第327-341页);68.4gm/l蔗糖;36gm/l葡萄糖;115mg/ll-脯氨酸;100mg/l肌肉肌醇(myo-inositol);和200μm乙酰丁香酮(在dmso中制备);ph5.4)。对于给定的实验组,使用来自合并的穗的胚进行每次转化。
土壤杆菌培养启动。将含有上文在实施例4中描述的二元转化载体的土壤杆菌菌株dat13192的甘油储液在含有合适抗生素的ab基本培养基板(watson等(1975)j.bacteriol.123:255-64)上划线,并于20℃培养3至4天。从每块板挑出单菌落,划线到含有合适抗生素的yep板(10gm/l酵母提取物;10gm/l蛋白胨;和5gm/lnacl)上,并于20℃温育1-2天。
土壤杆菌培养物和共培养。从yep板采集土壤杆菌菌落,在50ml一次性管中的10ml接种培养基中悬浮,并使用分光光度计将细胞密度调节至0.2至0.4的od550(于550nm测量的光密度)。将土壤杆菌培养物在旋转摇动器上以125rpm(室温)温育,期间实施胚切出。将未成熟的合子胚(先前从灭菌过的玉米粒分离,并在1ml接种培养基中放置)在接种培养基中清洗一次。将2ml土壤杆菌悬浮液添加至胚的每个管,并将管在摇动器平台上放置10-15分钟。将胚转移到共培养培养基(4.33gm/lms盐;1倍isu改良ms维生素;30gm/l蔗糖;700mg/ll-脯氨酸;koh中的3.3mg/l麦草畏(3,6-二氯-o-茴香酸或3,6-二氯-2-甲氧基苯甲酸);100mg/l肌肉肌醇;100mg/l酪蛋白酶水解物;15mg/lagno3;dmso中的100μm乙酰丁香酮;和3gm/lgelzantm
愈合组织选择和推定事件的再生:在共培养期后,将胚转移到静止培养基(restingmedium)(4.33gm/lms盐;1倍isu改良ms维生素;30gm/l蔗糖;700mg/ll-脯氨酸;koh中的3.3mg/l麦草畏;100mg/l肌肉肌醇;100mg/l酪蛋白酶水解物;15mg/lagno3;0.5gm/lmes(2-(n-吗啉代)乙磺酸一水合物;phytotechnologies
将胚转移到选择培养基1(其由静止培养基(上述)及100nmr-吡氟氯禾灵酸(haloxyfopacid)(0.0362mg/l)组成)上,并在黑暗中或在24小时光照下以50μem-2sec-1光强度于28℃温育7至14天。将增殖的胚胎发生愈合组织转移到选择培养基2(其由静止培养基(上述)及500nmr-吡氟氯禾灵酸(haloxyfopacid)(0.1810mg/l)组成)上,并在24小时光照下以50μem-2sec-1光强度于28℃温育14至21天。此选择步骤容许转基因愈伤组织进一步增殖和分化。
将增殖的胚胎发生愈合组织转移到再生前培养基(4.33gm/lms盐;1倍isu改良ms维生素;45gm/l蔗糖;350mg/ll-脯氨酸;100mg/l肌肉肌醇;50mg/l酪蛋白酶水解物;1.0mg/lagno3;0.25gm/lmes;naoh中的0.5mg/l萘乙酸;乙醇中的2.5mg/l脱落酸;1mg/l6-苯甲酸嘌呤;250mg/l羧苄青霉素;2.5gm/lgelzantm;和500nmr-吡氟氯禾灵酸;ph5.8)上,并在24小时光照下以50μem-2sec-1光强度于28℃温育7天。
将具有枝条样芽的胚胎发生愈伤组织转移到再生培养基i(4.33gm/lms盐;1倍isu改良ms维生素;60gm/l蔗糖;100mg/l肌肉肌醇;125mg/l羧苄青霉素;3.0gm/lgelzantm;和500nmr-吡氟氯禾灵酸;ph5.8)上,并在24小时光照下以50μem-2sec-1光强度温育7天。
将具有初生根的小枝条转移至phytatraystm(phytotechnologies
种子生成:将植物移植到metro-mixtm360无土生长培养基(sungrohorticulture;bellevue、wa)中,并在生长室中茁壮生长。然后,将植物移植到sunshinecustomblendtm160土壤混合物中,并在温室中种植至开花。进行受控传粉以生产种子。
通过实施例12中描述的方法进行对温室种植的植物的西部玉米根虫生物测定法。将具有根评级0.75或更好的植物立即移植至5加仑(18.92升)盆以生产种子。用杀虫剂处理移植物以防止进一步根虫损害和温室中的昆虫释放。将b104植物与非转基因b104(花粉供体)植物异型杂交以生产种子。保存由这些植物生产的种子以在植物的t1和后续世代时评估。
使用包含编码caf1-180v3发夹dsrna的植物可表达构建体(实施例4;质粒pdab109817中的构建体)的土壤杆菌菌株dat13192来生成b104转基因玉米植物。通过实施例12中描述的方法进行对这些温室种植的转基因植物的西部玉米根虫生物测定法。使用用类似质粒(pdab101556)(其含有yfp生成的基因,但是不含dsrna生成的基因)转化的植物作为阴性对照。表13中显示了这些生物测定法的结果。
表13:表达caf1-180v3发夹dsrna的b104转基因玉米事件的西部玉米根虫生物测定法的结果。
*玉米根损害评级量表:0–1.0
使用包含编码vatpasecv3发夹dsrna的植物可表达构建体(实施例4;质粒pdab109815中的构建体)的土壤杆菌菌株dat13192来生成b104转基因玉米植物。通过实施例12中描述的方法进行对这些温室种植的转基因植物的西部玉米根虫生物测定法。使用用类似质粒(pdab101556)(其含有yfp生成的基因,但是不含dsrna生成的基因)转化的植物作为阴性对照。表14中显示了这些生物测定法的结果。
表14:表达vatpasecv3发夹dsrna的b104转基因玉米事件的西部玉米根虫生物测定法的结果。
*玉米根损害评级量表:0-1.0
使用包含编码vatpasehv1发夹dsrna的植物可表达构建体(实施例4;质粒pdab109816中的构建体)的土壤杆菌菌株dat13192来生成b104转基因玉米植物。通过实施例12中描述的方法进行对这些温室种植的转基因植物的西部玉米根虫生物测定法。使用用类似质粒(pdab101556)(其含有yfp生成的基因,但是不含dsrna生成的基因)转化的植物作为阴性对照。表15中显示了这些生物测定法的结果。
表15:表达vatpasehv1发夹dsrna的b104转基因玉米事件的西部玉米根虫生物测定法的结果。
*玉米根损害评级量表:0–1.0
实施例9:whiskers介导的包含杀昆虫caf1-180v3发夹dsrna的转基因玉米组织的生成
生成如下的植物,其经由稳定整合入植物基因组中的嵌合基因的表达而生成一种或多种杀昆虫dsrna分子(例如,至少一种dsrna分子,其包含靶向包含seqidno:1-4之一的基因的dsrna分子)。制备供植物转化用的dna分子,如实施例4中所描述的,并经由whiskers介导的转化投递至hi-i悬浮细胞培养物中的玉米细胞中(基本上如记载于美国专利5,302,523和5,464,765;美国专利公开文本no.2008/0182332;及petolino和arnold(2009)m.paulscott(编)methodsinmolecularbiology:transgenicmaize、第526卷、humanapress、ny、第59-67页的)。
植物细胞转化和选择:用于whiskers介导的转化的所有规程使用标准无菌技术实施,并且一般遵循由petolino和arnold(2009)、见上文描述的方法。如下将玉蜀黍hiii胚胎发生细胞悬浮液(armstrong等(1991)maizegenet.coop.newslett.65:92-3)传代培养,即将30ml沉积的悬浮细胞,及70ml条件化培养基(即,已经培养过细胞的培养基)转移到175mlnalgenetm一次性圆锥底离心管(thermofisherscientific;产品目录编号3145-0175)中进入含有5%椰子汁(sigmaaldrich、st.louis、mo;产品目录编号c5915)的200ml新鲜h9cp(4.33gm/lms基础盐(phytotechnologieslaboratories、产品目录编号m524);30.0gm/l蔗糖;100.0mg/l肌肉肌醇;200mg/l酪蛋白酶水解物;2.0mg/l2,4-二氯苯氧基乙酸;2.0mg/l萘乙酸;691mg/ll-脯氨酸;2.0mg/l甘氨酸;0.5mg/l硫胺hcl;0.5mg/l吡哆醇hcl;0.05mg/l烟酸;ph6.0)培养基中。然后,将稀释的细胞在无菌1l一次性细胞培养烧瓶中在黑暗中于28℃在具有1英寸(2.54cm)抛掷半径(throwradius)的陀螺旋转(gyro-rotary)摇动器上以120rpm温育,并每3.5天将细胞传代培养,如上文所描述的。
在传代培养后一天,且在转染规程前18至24小时,将含有30ml沉积细胞的50mlh9cp培养基添加至无菌1l烧瓶中的150ml新鲜gn6培养基(3.99gm/lchun6基础盐及维生素(phytotechnologieslaboratories、产品目录编号c167);30gm/l蔗糖;100mg/l肌肉肌醇;2mg/l2,4-二氯苯氧基乙酸;ph6.0),并将培养物在陀螺旋转摇动器上温育,如上文所描述的。
gn6培养基中温育18至24小时后,将整个细胞悬浮液转移至无菌175ml一次性圆锥底离心管,并容许细胞沉积1-3分钟,产生约40ml的细胞体积。将用过的培养基小心弃去,并通过移液管取出残留的液体以生成潮湿的细胞块。将细胞在离心管中的约180ml高渗透培养基(gn6-sm;补充有45gm/l山梨糖醇和45gm/l甘露醇的gn6培养基)中重悬,并将培养物在桌面振荡器摇动器上于室温(23℃至25℃)放置约30分钟,但不超过45分钟。30分钟渗透处理后,容许细胞在离心管中沉积3-5分钟。然后,经由移液管将液体小心取出,向下至离心管的50ml标记,注意不扰乱细胞。
对于将质粒dna投递至玉米悬浮培养细胞,通过将合适量的gn6-sm培养基添加至经灭菌的(经高温灭菌的)干燥晶须制备5ml5%w/wbiogradetmsc09碳化硅晶须(whisker)(advancedcompositematerials、greer、sc;批次981011-101)悬浮液。将合适量的pdab109830(实施例4)的dna添加至离心管(通常为80μg/40ml细胞的50ml悬浮液),将帽紧密密封,并将管温和旋动以混合内容物。将离心管在修改为牢固保持管的商业涂料混合器(paintmixer)(reddeviltm5400型;minneapolis、mn)中牢固固定,并摇动10-20秒。在用约150ml新鲜培养基(gn6-sm:gn6;2:1v/v)稀释至终体积约200ml以降低培养基的渗量,将细胞于rt在桌面振荡器摇动器上温育约1小时。
然后,通过移液管在约8.3ml的等分试样中将经转染的细胞转移到无菌70mmwhatmantm#4滤纸(thermofisherscientific)上,注意将细胞在滤纸上均匀分配。将滤纸在100x20mm塑料板中的gn6琼脂培养基上放置,然后,在塑料盒中在黑暗中于28℃温育7天。
在转化后一周,将保持细胞的滤纸转移到100x20mm板中含有1.0mg/lbialaphos的gn6-1h(补充有2.5gm/lgelzantm的gn6培养基)固体琼脂培养基的新鲜板,并在黑暗中于28℃温育。bialaphos以
一周后,如下将细胞在软琼脂中包埋,即于37℃至40℃将每张滤纸的细胞内容物刮入含有15mlgn6软琼脂糖培养基(具有7.0gm/lseaplaquetm琼脂糖的gn6培养基;lonza、rockland、me)的50ml无菌一次性离心管中,有力摇动加盖的管,然后将管的内容物均匀倒入4个含有gn61h固体琼脂培养基的100x25mm塑料板上。将每块板搅动以将表面包被以均匀的细胞悬浮液层,并在凝固后,将板于28℃在黑暗中温育。
在温育6至10周后,将良好生长的出现的菌落转移到含有gn61h琼脂培养基的新鲜板。容许这些经候选物转化的菌落在选择培养基上生长2-4周以建立具有约50至200mg组织块的稳定事件,然后,将其进行分子分析。
将来自候选事件的0.1ml压缩愈伤组织细胞的样品取样,并在1.2mlcostartm聚丙烯簇管(corning、inc.;corningny)中放置,并于-80℃冷冻。
分子分析:通过per53'utr的实时定量pcr对愈伤组织细胞事件分析全长转录物的相对表达,并通过与内部玉米基因(tip41样蛋白)比较评估拷贝数。使用rneasytm96试剂盒(qiagen、valencia、ca)分离rna。在第一次清洗(rw1)后,用缓冲液“rdd”中的qiagen无rna酶的dna酶(依照试剂盒提出的备选方案)处理柱。使用在具有5μl变性rna的10μl反应体积中的高性能cdna合成试剂盒(invitrogen),基本上依照制造商推荐的方案制备第一链cdna。将方案略微修改为包括将10μl100μmt20vn寡核苷酸(idt)添加至1ml随机引物储液混合物管中,以制备组合的随机引物和寡聚dt的工作储液。
cdna合成后,将样品用无核酸酶的水以1:3稀释,并于-20℃贮存,直到测定。在lightcyclertm480(rochediagnostics、indianapolis、in)上在10μl反应体积中实施实时pcr。所有测定法包括无模板的阴性对照(仅混合物)。对于标准曲线,来源板中还包含空白(来源孔中的水)以检查样品交叉污染。用0.5μm的rocheuniversalprobetm和10μm的针对靶物和参照基因的引物运行反应。表16中列出了引物序列。pcr反应条件如下:(1)于95℃进行靶物活化10分钟;(2)43个循环的(于95℃变性10秒和于60℃延伸);(3)于72℃获得1秒;并(4)于40℃冷冻10秒。
表16:用于转基因玉米的分子分析的引物序列。
*tip41样蛋白;通过tblastx得到的at4g34270的玉米同系物(74%同一性)
数据分析:使用lightcyclertm软件v1.5分析数据,其通过使用用于计算cq值的二次导数最大值算法依照供应商的推荐的相对量化进行。对于表达分析,使用δct法(即,2-(cqtarget–cqref))计算表达数值,其依赖于两种靶物间cq值差的比较,其中在如下的假设下选择基值(basevalue)为2:对于优化的pcr反应,产物每个循环加倍。
转基因植物再生:将一些稳定转化的事件再生成植物,用于植物体内昆虫生物测定法。
在选择过程后,将愈伤组织培养物转移至再生前28培养基(4.33gm/l具有维生素的ms基础培养基;30.0gm/l蔗糖;5mg/l6-苯甲酸嘌呤;25μg/l2,4-二氯苯氧基乙酸;2.5gm/lgelzantm;和1.0mg/lbialaphos)。将转移的愈伤组织培养物于28℃在连续白色荧光(约50μem-2s-1)下温育7天。
对于再生,将培养物转移至再生培养基36(4.33gm/l具有维生素的ms基础培养基;30gm/l蔗糖;2.5gm/lgelzantm;和1.0mg/lbialaphos),并容许小植物生成,并于28℃在连续白色荧光下生长多至3周。在小植物达到合适的生长阶段时,用镊子和解剖刀将它们切出,转移至含有琼脂的20x100mm试管,并在光中放置2天。
对转基因植物指派独特的标识符,并转移至受控的环境室(约28℃日间温度;24℃夜间温度;以16:8补充照明光周期)。将转基因植物从管移植到3.5英寸(8.89cm)盆,并返回受控的环境室达1-2周以帮助驯化植物。然后,将转基因植物移动至温室,并以每个rootrainertm每个事件一个植物从小盘移植到rootrainerstm(式样tinustm350-4;spencer-lemaireindustries;acheson、alberta、canada),用于昆虫进食生物测定法。移植到roottrainerstm后约4天,t0植物准备好昆虫进食生物测定法。通过实施例12中描述的方法进行昆虫进食生物测定法。
生成发夹dsrna的转基因的存在和表达导致wcr根修剪的降低,如在那些展现出高表达水平的dsrna转录物(如通过per53'utr的相对pcr评分测量的)的植物中明显的。表17中显示了对含有rnai发夹的那些事件的西部玉米根虫生物测定法的结果,所述rnai发夹包含seqidno:97,且比内部tip41样参照基因具有高16倍的相对表达水平。结果指示令人惊讶的结果,即与仅3%的测试非转基因对照植物相比,36%的转基因rnai植物表明降低的wcr损害。虽然生物学响应在t0转基因玉米植物内可变,但是与非转基因对照植物相比,转基因植物显示可觉察的wcr修剪降低。
表17:表达caf1-180发夹dsrna的hi-ii转基因玉米事件的西部玉米根虫生物测定法的结果。
实施例10:whiskers介导的包含杀昆虫vatpasecv3和vatpasecv4发夹dsrna的转基因玉米组织的生成
使用实施例9中描述的转化方法来提供表达vatpasecv3发夹dsrna或vatpasecv4发夹dsrna的hiii玉米转基因植物,其通过用质粒pdab109828和pdab109838(实施例4)转化进行。依照实施例9中描述的筛选方法实施分子筛选,并依照实施例12中描述的生物测定方法实施西部玉米根虫生物测定法。
生成发夹dsrna的转基因的存在和表达导致wcr根修剪的降低,如在那些展现出高表达水平的dsrna转录物(如通过per53'utr的相对pcr评分测量的)的植物中明显的。表18中显示了对含有rnai发夹的那些事件的西部玉米根虫生物测定法的结果,所述rnai发夹包含seqidno:100(vatpasecv3)或seqidno:103(vatpasecv4),且比内部tip41样参照基因具有高20倍的相对表达水平。结果指示令人惊讶的结果,即与0%的测试非转基因对照植物相比,43%的转基因rnai植物表明降低的wcr损害。虽然生物学响应在t0转基因玉米植物内可变,但是仅转基因植物显示可觉察的wcr修剪降低。
表18:表达vatpasecv3或v4发夹dsrna的hi-ii转基因玉米事件的西部玉米根虫生物测定法的结果。
实施例11:whiskers介导的包含杀昆虫vatpasehv1发夹dsrna的转基因玉米组织的生成
使用实施例9中描述的转化方法来提供表达vatpasehv1发夹dsrna的hiii玉米转基因植物,其通过用质粒pdab109829(实施例4)转化进行。依照实施例9中描述的筛选方法实施分子筛选,并依照实施例12中描述的生物测定方法实施西部玉米根虫生物测定法。
生成发夹dsrna的转基因的存在和表达导致wcr根修剪的降低,如在那些展现出高表达水平的dsrna转录物(如通过per53'utr的相对pcr评分测量的)的植物中明显的。表19中显示了对含有rnai发夹的那些事件的西部玉米根虫生物测定法的结果,所述rnai发夹包含seqidno:106,且比内部tip41样参照基因具有高18倍的相对表达水平。结果指示令人惊讶的结果,即与0%的测试非转基因对照植物相比,60%的转基因rnai植物表明降低的wcr损害。虽然生物学响应在t0转基因玉米植物内可变,但是仅转基因植物显示可觉察的wcr修剪降低。
表19:表达vatpaseh发夹dsrna的hi-ii转基因玉米事件的西部玉米根虫生物测定法的结果。
实施例12:转基因玉米的昆虫生物测定法
西部玉米根虫(wcr,玉米根萤叶甲)卵在土壤中自cropcharacteristics(farmington、mn)接收。将卵于28℃温育10-11天。将卵从土壤洗出,放入0.15%琼脂溶液中,并将浓度调节至每0.25ml等分试样约75-100个卵。在培养皿中用卵悬浮液的等分试样制备孵化板以监测孵化率。
用150-200个wcr卵侵袭玉米植物周围的土壤。容许昆虫进食2周,在该时间后,对每个植物给出“根评级”。利用节损伤量表进行评级。oleson等(2005)j.econ.entomol.98(1):1-8。
将具有根评级0.75或更好的植物立即移植到5加仑(18.92升)盆以生成种子。用杀虫剂处理移植物以防止进一步根虫损害和温室中的昆虫释放。将hiii植物与玉米近交系5xh751(花粉供体)异型杂交以生产种子。保存由这些植物生产的种子以在植物的t1和后续世代时评估。
实施例13:包含鞘翅目有害生物序列的转基因玉蜀黍
生成10至20个转基因t0玉蜀黍植物,如实施例8和9中描述的。从玉米根虫攻击获得另外的10-20个t1玉蜀黍独立系,其表达用于rnai构建体的发夹dsrna,如seqidno:1-4中所列的。这些经由rt-pcr确认。任选地,使用来自选择的独立t1系的总rna用引物进行rt-pcr,所述引物设计为在每种rnai构建体中的发夹盒的pdk内含子中结合。另外,任选地,使用针对rnai构建体中每种靶基因的特异性引物来扩增并确认植物体内sirna生成需要的加工前mrna的生成。每种靶基因的期望条带的扩增确认每种转基因玉蜀黍中发夹rna的表达。随后,任选地,使用rna印迹杂交在独立的转基因系中确认将靶基因的dsrna发夹加工成sirna。
转基因rnai系和野生型玉蜀黍的表型比较。
选择用于创建发夹dsrna的靶鞘翅目有害生物基因或序列与任何已知的植物基因或序列没有相似性。因此,没有预期通过靶向这些基因或序列的构建体的rnai生成或活化(系统性)对转基因植物具有具有任何有害的影响。然而,与野生型植物比较转基因系的发育和形态学特征,以及用空发夹载体转化的转基因系的那些发育和形态学特征。比较植物根、枝条、叶和生殖特征。转基因和野生型植物的根长度和生长样式没有观察到的差异。植物枝条特征诸如高度、叶数目和大小、开花时间、花大小和外观是相似的。一般地,在体外和温室中的土壤中培养时,转基因系和那些没有靶irna分子表达的系之间没有观察到的形态学差异。
实施例14:包含鞘翅目有害生物序列和其它rnai构建体的转基因玉蜀黍
经由土壤杆菌或whiskerstm介导的转化转化转基因玉蜀黍植物以生成一种或多种杀昆虫dsrna分子(例如,至少一种dsrna分子,其包括靶向包含seqidno:1-4之一的基因的dsrna分子),所述转基因玉蜀黍植物在其基因组中包含异源编码序列,其转录成靶向与鞘翅目有害生物不同的生物体的irna分子。将基本上如实施例4中所描述的那样制备的植物转化dna分子的制备物投递至自转基因玉蜀黍植物获得的玉米hi-ii悬浮细胞培养物中,所述转基因玉蜀黍植物在其基因组中包含异源编码序列,其转录成靶向与鞘翅目有害生物不同的生物体的irna分子。
实施例15:转基因鞘翅目有害生物抗性植物
与靶基因对应的dsrna、sirna或mirna的植物inplanta投递及随后鞘翅目有害生物经由进食摄取经由rna介导的基因沉默导致靶基因的下调。在靶基因的功能是重要的时,鞘翅目有害生物的生长、发育和生殖受到影响,并且在wcr、ncr、scr、mcr、d.balteataleconte、d.u.tenella、和d.u.undecimpunctatamannerheim中至少一种的情况中导致不能成功寄生、进食、发育、和/或生殖,或导致一个或多个发育阶段的鞘翅目有害生物的死亡。然后,使用靶基因的选择和rnai的成功应用来控制鞘翅目有害生物。用玉米根虫物种攻击每种rnai构建体的10-20个独立的t1玉蜀黍转基因系的5至10个重复。对每种玉米根虫物种重复攻击。使rnai系的t1种子萌发,并在萌发后10-15天将抗性植物转移至改良的knop氏培养基。同时萌发野生型对照玉蜀黍种子,并用于玉米根虫感染。
在对照上比在含有一种或多种rnai构建体的转基因玉蜀黍系上有显著更多的(大于50%)存活玉米根虫。使用定量实时rt-pcr在以野生型和转基因植物的根为食的玉米根虫中测量irna丰度。在含有一种或多种rnai构建体的转基因玉蜀黍系中比在对照植物中找到显著更多irna分子。这些结果指示转基因系加工与靶基因对应的sirna,并且这些sirna可用于被进食的玉米根虫摄取。更重要地,结果指示rnai介导的对所有靶基因的抑制影响靶玉米根虫的生长、发育、和存活力。此外,具有与靶基因具有超过80%序列同一性的错配序列的rnai分子以与野生型序列相似的方式影响玉米根虫。错配序列与天然序列的配对以在相同rnai构建体中形成发夹dsrna投递经植物加工的sirna,其能够影响进食鞘翅目有害生物的生长、发育和存活力。
序列表
<110>陶氏益农公司(dowagrosciences,llc)
narva,kennethe
li,huarong
geng,chaoxian
larrinua,ignacio
olson,monicab
elango,narvin
<120>赋予对鞘翅目有害生物的抗性的核酸分子
<130>2971-p10251.1us
<150>us61/428,592
<151>2010-12-30
<160>138
<170>patentinversion3.4
<210>1
<211>635
<212>dna
<213>diabroticavirgifera
<220>
<221>misc_feature
<222>(466)..(466)
<223>n是a,c,g,或t
<220>
<221>misc_feature
<222>(576)..(576)
<223>n是a,c,g,或t
<220>
<221>misc_feature
<222>(628)..(628)
<223>n是a,c,g,或t
<400>1
ttcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaaagtctaaa60
aacttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttgggagaaac120
tatttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtacttccatga180
caatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgttaaaccaag240
aaaacctttggctgaagataaaaaacatttttaaattatgaaggtaggatttcagatgat300
gattgggaggaagaggaacaaggtgagtctmttaatggttcagaagatgaagctgagaag360
gataatgacgatgacaaggatgattatgaagttgataatgaattctttgttccacatggg420
cacttaagtgatgatgaggtggatgatgaagaactgagtcgtttanccccagattctttg480
aaacaaaaacttaaacttcttaaagaagagtttgatcaagatatgaagtctaaaactcag540
aaactaaagacctaggtccatggtttgcatatggtnttaacaaagaataagtagaggagt600
atagtaggaggagagtgtgtgtagtatntactact635
<210>2
<211>1387
<212>dna
<213>diabroticavirgifera
<220>
<221>misc_feature
<222>(1382)..(1382)
<223>n是a,c,g,或t
<400>2
agggcwgactcagttcattggggagtggttgaaaggggtgttttttggagttattccatt60
atttgaaggagtgataaagaagatcttggaagccaccatagacttaattcacatagaacc120
agcatactttctgatcaccaagaagaaatgactgagtattggttgatatctgccccaggg180
gataagacctgtcaacagacatgggacacgatgaataatatgacaagtaaacagaataat240
ttgtcaaccaactacaagtttcaaatcccggacttgaaagttggtactctggatcagtta300
gtaggactttctgatgatctgggcaagcttgatgggttcgtggagcaggttaccaggaag360
gtagctcagtaccttggagaagtacttgaagaacagagggacaagtggtcagaaaacctg420
caggccaataacagcgatttgcccacttatttaacccgatttacttgggacatcgccaaa480
tatcccatcaagcaatcgctccgtaacatcgccgatataatcagcaaacaagttggccaa540
attgacgctgatctgaagaccaaatcgtctgcgtacaataacttgaaaggaagtcttcag600
aatttggagaagaaacagacaggaagtttactaacaagaaacttggcagatttggtgaaa660
aaggaacattttattttggattctgaatatttgcagacattgttggttattgttccaaaa720
gcccaattcaacgaatggaatgcgacatacgaaaagattaccgacatgatagtccctcgt780
tcttcacaactgattaaacaggataacgaatacggtctatatactgtatccctattcaaa840
aaggtcgtcgaagaattcaagttacacgctagagaaaagaagttcatcgttcgtgatttc900
atatataacgaagaggaactggcagctggaaagaacgagataaccaaactcgtcaccgat960
aaaaagaagcaatttggtccattagttagatggcttaaagtcaacttcagcgagtgtttc1020
tgcgcctggattcacgttaaagcgttgagggtatttgttgaatctgttttaagatatggc1080
ctcccgtcaatttccaagctattttaatccacccaataagcaacaattaaaacgtttaag1140
agatgtcttgaaccaactttacggccacctcgatagtagtgctgccctttcaggacctaa1200
cgttgatagcgttgacattccaggcctcggattcggccaatcagagtattacccatacgt1260
ttactacaagctgaacgtcgacatgttagaatcgaagatctaaactcacgcatcaatcca1320
agagtttgttattaataggttccaaacaattttaaaaaaacgtacttacgcaagtgagtg1380
anaggcc1387
<210>3
<211>1701
<212>dna
<213>diabroticavirgifera
<400>3
aattawattttatacctcgatccaaaaatattttcgcaaaattgtagtacactgaatttt60
aactatcatgattgttccagatatgttggctgctacaagtgtattgcagcaaagagctac120
agatataagaaatcaacatgtaaactggcaatcctatttccagtcacaaatgatttcgca180
ggaagactacaacttcatagtagcttttgatgtgactgatagtgcaaaaagagaagctct240
cctgaaaagcgacagaaaccaatgtgctcaaacattgttgaaccttttggggcatgtatc300
caaggaccaaactctgcagtacattttagttttggttgatgatatgttacaggagaacag360
aagtcgtgtagagatatttcacgaatatgccaaccaaaagaaagaatctgtttggggtcc420
atttttgaaccttcttaaccgccaggatggatttattaccaacatgacttctaggatcat480
cgctaaaattgcttgttggtcgcagacacttatggatcgttctgaccttcatttttactt540
gacctggttaaaagaccaactaaaaactcagaacaacgaatacatccaatcggttgcaag600
atgccttcaaatgatgctccgaattgacgactaccgttttgccttcgtgtccgtcgatgg660
aatatccactctgttgtcggtactctctggaagagtaaatttccaagttcagtaccagct720
gatattctgcttgtgggtactcactttcaacccgttgttagccgaaaagatgaacaaatt780
caacgttattcctatattggcagatattttgagcgattcggtaaaggagaaagtaaccag840
aattattttagctgttttcaggaatttgatcgaaaaacccgaagatcaacaagtttcaaa900
agaacattgtatagctatggtgcaatgcaaagtattgaaacaattggccattttagaaca960
gcgcaagttcgatgatgaagatgtgacagctgatgtcgaatttctaactgaaaaactaca1020
gagttctgttcaggatctgagttccttcgatgaatattcaactgaagtcaaatctggacg1080
tttggaatggtctccagtacacaagagcaagttctggcgggaaaatgcgcagcgcctcaa1140
cgagaaaaactacgaattactccgtattctcattcatttgttagaaaccagcaaggaccc1200
cttggtattgagcgtagctagtttcgatattggagagtatgttcgccattatcctcgcgg1260
caaacacataatcgagcagttgggaggaaagcaattagttatgcaactcttagcccacga1320
agatcccaatgtacgatatgaagctctgttggcagtccaaaaattgatggtccacaattg1380
ggaataccttggtcgtcaacttgaaaaagagcaaagtacagataaaaacgcgccaaaggg1440
tggagctcctgtcgcaggcaaggcgtaaatactaccatgaaaagtatttaaagatattta1500
ctgcaataattgtagaaactttaaaaataattgtaaaaattatatcgatttctgtacata1560
gctgtataggtatacattcctgtcatattttattattgttacgataaccaataatcaacc1620
atgcacattttaaacatttaaaaatattgcttttaaagttaamtttttttacttgtattg1680
ttagtattataataagtatat1701
<210>4
<211>1001
<212>dna
<213>diabroticavirgifera
<400>4
aaattggactaaacttttttgctcgtttwgttaatcttatccattaatttctaaaattgt60
atgattttgttagaaagtggtagaataacgctaaccaatttaggagtaacactgcaaaga120
gactctctgaatgttttgttgctgttccgataaggaggactacggaaaatttttgcttga180
tccttatctgtctgttatccccgctcttacttcgtaatttatttaatgtccgccattttc240
gttcgtagttgaataggcgttaactttgtgtgttttggcaggtccgatggctgcaataag300
aaaaaaattggtgattgttggtgacggagcatgtggaaaaacatgtcttttaatagtatt360
tagtaaagatcagttcccagaagtatacgtgcccactgtattcgaaaactatgtggcgga420
tattgaagtagatggaaagcaggtagaactcgccttatgggataccgcaggtcaagaaga480
ttatgatcgtctgcggcccctttcatatccagacaccgatgtgattttaatgtgcttctc540
agtagactcccccgactcattagaaaacatcccagaaaaatggaccccagaggtaaaaca600
cttttgtccaaatgtgccaattattctggtaggcaataagaaggatctgcgtaacgatcc660
caataccattaacgaacttaaaaagatgaaacaagaaccagtaaaacctcaagacggaag720
atccatggcagagaaaatcaatgcttttgcatatctagaatgttctgccaaaagtaagga780
gggtgtaagagaggtttttgaaaatgctacccgagctgcactgcaagtcaaaaagaagaa840
gaagccccgttgtgtaatgttttgagccactacatattctatattttatattaataatcg900
ttacgtgtgttaattaatttactaagggggacgttctgtcatgatgtattgacaaggggg960
cttctgcatgatgtattgactaacaaaaataaaccccwtta1001
<210>5
<211>338
<212>dna
<213>diabroticavirgifera
<400>5
agggcwgactcagttcattggggagtggttgaaaggggtgttttttggagttattccatt60
atttgaaggagtgataaagaagatcttggaagccaccatagacttaattcacatagaacc120
agcatactttctgatcaccaagaagaaatgactgagtattggttgatatctgccccaggg180
gataagacctgtcaacagacatgggacacgatgaataatatgacaagtaaacagaataat240
ttgtcaaccaactacaagtttcaaatcccggacttgaaagttggtactctggatcagtta300
gtaggactttctgatgatctgggcaagcttgatgggtt338
<210>6
<211>21
<212>dna
<213>diabroticavirgifera
<220>
<221>misc_feature
<222>(16)..(16)
<223>n是a,c,g,或t
<400>6
tacgcaagtgagtganaggcc21
<210>7
<211>460
<212>dna
<213>diabroticavirgifera
<400>7
agaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacagac60
atgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagtt120
tcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatct180
gggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggaga240
agtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgattt300
gcccacttatttaacccgatttacttgggacatcgccaaatatcccatcaagcaatcgct360
ccgtaacatcgccgatataatcagcaaacaagttggccaaattgacgctgatctgaagac420
caaatcgtctgcgtacaataacttgaaaggaagtcttcag460
<210>8
<211>500
<212>dna
<213>diabroticavirgifera
<400>8
agaaacagacaggaagtttactaacaagaaacttggcagatttggtgaaaaaggaacatt60
ttattttggattctgaatatttgcagacattgttggttattgttccaaaagcccaattca120
acgaatggaatgcgacatacgaaaagattaccgacatgatagtccctcgttcttcacaac180
tgattaaacaggataacgaatacggtctatatactgtatccctattcaaaaaggtcgtcg240
aagaattcaagttacacgctagagaaaagaagttcatcgttcgtgatttcatatataacg300
aagaggaactggcagctggaaagaacgagataaccaaactcgtcaccgataaaaagaagc360
aatttggtccattagttagatggcttaaagtcaacttcagcgagtgtttctgcgcctgga420
ttcacgttaaagcgttgagggtatttgttgaatctgttttaagatatggcctcccgtcaa480
tttccaagctattttaatcc500
<210>9
<211>77
<212>dna
<213>diabroticavirgifera
<400>9
aattawattttatacctcgatccaaaaatattttcgcaaaattgtagtacactgaatttt60
aactatcatgattgttc77
<210>10
<211>445
<212>dna
<213>diabroticavirgifera
<400>10
tcatgattgttccagatatgttggctgctacaagtgtattgcagcaaagagctacagata60
taagaaatcaacatgtaaactggcaatcctatttccagtcacaaatgatttcgcaggaag120
actacaacttcatagtagcttttgatgtgactgatagtgcaaaaagagaagctctcctga180
aaagcgacagaaaccaatgtgctcaaacattgttgaaccttttggggcatgtatccaagg240
accaaactctgcagtacattttagttttggttgatgatatgttacaggagaacagaagtc300
gtgtagagatatttcacgaatatgccaaccaaaagaaagaatctgtttggggtccatttt360
tgaaccttcttaaccgccaggatggatttattaccaacatgacttctaggatcatcgcta420
aaattgcttgttggtcgcagacact445
<210>11
<211>570
<212>dna
<213>diabroticavirgifera
<400>11
agaacattgtatagctatggtgcaatgcaaagtattgaaacaattggccattttagaaca60
gcgcaagttcgatgatgaagatgtgacagctgatgtcgaatttctaactgaaaaactaca120
gagttctgttcaggatctgagttccttcgatgaatattcaactgaagtcaaatctggacg180
tttggaatggtctccagtacacaagagcaagttctggcgggaaaatgcgcagcgcctcaa240
cgagaaaaactacgaattactccgtattctcattcatttgttagaaaccagcaaggaccc300
cttggtattgagcgtagctagtttcgatattggagagtatgttcgccattatcctcgcgg360
caaacacataatcgagcagttgggaggaaagcaattagttatgcaactcttagcccacga420
agatcccaatgtacgatatgaagctctgttggcagtccaaaaattgatggtccacaattg480
ggaataccttggtcgtcaacttgaaaaagagcaaagtacagataaaaacgcgccaaaggg540
tggagctcctgtcgcaggcaaggcgtaaat570
<210>12
<211>24
<212>dna
<213>人工的
<220>
<223>t7噬菌体启动子序列(t7phagepromotersequence)
<400>12
ttaatacgactcactatagggaga24
<210>13
<211>50
<212>dna
<213>人工的
<220>
<223>引物caf-ft7
<400>13
ttaatacgactcactatagggagattcggaagcttcatatttaaaagatc50
<210>14
<211>24
<212>dna
<213>人工的
<220>
<223>引物caf-r
<400>14
tatcttcagccaaaggttttcttg24
<210>15
<211>26
<212>dna
<213>人工的
<220>
<223>引物caf-f
<400>15
ttcggaagcttcatatttaaaagatc26
<210>16
<211>48
<212>dna
<213>人工的
<220>
<223>引物caf-rt7
<400>16
ttaatacgactcactatagggagatatcttcagccaaaggttttcttg48
<210>17
<211>46
<212>dna
<213>人工的
<220>
<223>引物atp.c-f1t7
<400>17
ttaatacgactcactatagggagaagaagaaatgactgagtattgg46
<210>18
<211>20
<212>dna
<213>人工的
<220>
<223>引物atp.c-r1
<400>18
ctgaagacttcctttcaagt20
<210>19
<211>22
<212>dna
<213>人工的
<220>
<223>引物atp.c-f1
<400>19
agaagaaatgactgagtattgg22
<210>20
<211>44
<212>dna
<213>人工的
<220>
<223>引物atp.c-r1t7
<400>20
ttaatacgactcactatagggagactgaagacttcctttcaagt44
<210>21
<211>48
<212>dna
<213>人工的
<220>
<223>引物atp.c-r1shortt7
<400>21
ttaatacgactcactatagggagagatgggatatttggcgatgtccca48
<210>22
<211>24
<212>dna
<213>人工的
<220>
<223>引物atp.c-r1short
<400>22
gatgggatatttggcgatgtccca24
<210>23
<211>46
<212>dna
<213>人工的
<220>
<223>atp.c-f2t7
<400>23
ttaatacgactcactatagggagaagaaacagacaggaagtttact46
<210>24
<211>25
<212>dna
<213>人工的
<220>
<223>引物atp.c-r2
<400>24
ggattaaaatagcttggaaattgac25
<210>25
<211>22
<212>dna
<213>人工的
<220>
<223>引物atp.c-f2
<400>25
agaaacagacaggaagtttact22
<210>26
<211>49
<212>dna
<213>人工的
<220>
<223>引物atp.c-r2t7
<400>26
ttaatacgactcactatagggagaggattaaaatagcttggaaattgac49
<210>27
<211>48
<212>dna
<213>人工的
<220>
<223>引物atp.h-f1t7
<400>27
ttaatacgactcactatagggagatcatgattgttccagatatgttgg48
<210>28
<211>20
<212>dna
<213>人工的
<220>
<223>引物atp.h-r1
<400>28
agtgtctgcgaccaacaagc20
<210>29
<211>24
<212>dna
<213>人工的
<220>
<223>引物atp.h-f1
<400>29
tcatgattgttccagatatgttgg24
<210>30
<211>44
<212>dna
<213>人工的
<220>
<223>引物atp.h-r1t7
<400>30
ttaatacgactcactatagggagaagtgtctgcgaccaacaagc44
<210>31
<211>46
<212>dna
<213>人工的
<220>
<223>引物atp.h-f2t7
<400>31
ttaatacgactcactatagggagaagaacattgtatagctatggtg46
<210>32
<211>20
<212>dna
<213>人工的
<220>
<223>引物atp.h-r2
<400>32
atttacgccttgcctgcgac20
<210>33
<211>22
<212>dna
<213>人工的
<220>
<223>引物atp.h-f2
<400>33
agaacattgtatagctatggtg22
<210>34
<211>44
<212>dna
<213>人工的
<220>
<223>引物atp.h-r2t7
<400>34
ttaatacgactcactatagggagaatttacgccttgcctgcgac44
<210>35
<211>46
<212>dna
<213>人工的
<220>
<223>引物rho1-ft7
<400>35
ttaatacgactcactatagggagacaggtccgatggctgcaataag46
<210>36
<211>20
<212>dna
<213>人工的
<220>
<223>引物rho1-r
<400>36
gacttgcagtgcagctcggg20
<210>37
<211>22
<212>dna
<213>人工的
<220>
<223>引物rho1-f
<400>37
caggtccgatggctgcaataag22
<210>38
<211>44
<212>dna
<213>人工的
<220>
<223>引物rho1-rt7
<400>38
ttaatacgactcactatagggagagacttgcagtgcagctcggg44
<210>39
<211>39
<212>dna
<213>人工的
<220>
<223>引物hpcaf-f
<400>39
gagaggtacctcggaagcttcatatttaaaagatctgtc39
<210>40
<211>42
<212>dna
<213>人工的
<220>
<223>引物hpcaf-r
<400>40
ctctggatccaaaatgttttttatcttcagccaaaggttttc42
<210>41
<211>42
<212>dna
<213>人工的
<220>
<223>引物hp-invcaf-f
<400>41
agagccatggaaaatgttttttatcttcagccaaaggttttc42
<210>42
<211>39
<212>dna
<213>人工的
<220>
<223>引物hp-invcaf-r
<400>42
ctctgagctctcggaagcttcatatttaaaagatctgtc39
<210>43
<211>43
<212>dna
<213>人工的
<220>
<223>引物st-ls1-f
<400>43
agagggatccaggcctaggtatgtttctgcttctacctttgat43
<210>44
<211>47
<212>dna
<213>人工的
<220>
<223>引物st-ls1-r
<400>44
ctctccatggaccggtatttaaatacctgcacatcaccatgttttgg47
<210>45
<211>40
<212>dna
<213>人工的
<220>
<223>引物hpatpasec-f
<400>45
agagggtaccagaagaaatgactgagtattggttgatatc40
<210>46
<211>32
<212>dna
<213>人工的
<220>
<223>引物hpatpasec-r
<400>46
ctctggatccgatgggatatttggcgatgtcc32
<210>47
<211>32
<212>dna
<213>人工的
<220>
<223>引物hp-invatpasec-f
<400>47
agagccatgggatgggatatttggcgatgtcc32
<210>48
<211>39
<212>dna
<213>人工的
<220>
<223>引物hp-invatpasec-r
<400>48
gctgagctcagaagaaatgactgagtattggttgatatc39
<210>49
<211>729
<212>dna
<213>人工的
<220>
<223>caf1-180形成发夹序列(caf1-180hairpin-formingsequence)
<400>49
tcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaaagtctaaaa60
acttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttgggagaaact120
atttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtacttccatgac180
aatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgttaaaccaaga240
aaacctttggctgaagataaaaaacattttgtatgtttctgcttctacctttgatatata300
tataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaa360
agaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttat420
aacttttctaatatatgaccaaaacatggtgatgtgcagaaaatgttttttatcttcagc480
caaaggttttcttggtttaacaaatttgctttttttcctccatgtgccataatatgcagg540
tctacgattgtcatggaagtacaaaaacttggctttgtgtctttgaaggccagatttgtc600
ttcgcaaatagtttctcccaaatcaacaagaatgatttcatcatcaagagattcttcata660
tggccaagtttttagacttttacctatagcctttcctgatgacagatcttttaaatatga720
agcttccga729
<210>50
<211>889
<212>dna
<213>人工的
<220>
<223>vatpasec形成发夹序列
<400>50
aagaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacaga60
catgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagt120
ttcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatc180
tgggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggag240
aagtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgatt300
tgcccacttatttaacccgatttacttgggacatcgccaaatatcccatcgtatgtttct360
gcttctacctttgatatatatataataattatcactaattagtagtaatatagtatttca420
agtatttttttcaaaataaaagaatgtagtatatagctattgcttttctgtagtttataa480
gtgtgtatattttaatttataacttttctaatatatgaccaaaacatggtgatgtgcagg540
atgggatatttggcgatgtcccaagtaaatcgggttaaataagtgggcaaatcgctgtta600
ttggcctgcaggttttctgaccacttgtccctctgttcttcaagtacttctccaaggtac660
tgagctaccttcctggtaacctgctccacgaacccatcaagcttgcccagatcatcagaa720
agtcctactaactgatccagagtaccaactttcaagtccgggatttgaaacttgtagttg780
gttgacaaattattctgtttacttgtcatattattcatcgtgtcccatgtctgttgacag840
gtcttatcccctggggcagatatcaaccaatactcagtcatttcttctt889
<210>51
<211>447
<212>dna
<213>diabroticavirgifera
<400>51
gctccaacagtggttccttatccagatttcaatgccgaagaagatgcaaaggctctaaaa60
caagcattcaagggtttcggaacggacgaagaagctgtaatagaaattatcacaaaaagg120
agtaatgaacaaagacgtgaaatagcgattacattcaaaacaatgtatggcaaggatctc180
atcaaagaactgaaaagtgaacttagaggaaatttcgaagatgccatcgtagctctgatg240
acagagcccatcgagtttcaagccaaacagttgcataaagctatcagcggattgggaact300
gatgaaagtacaatmgtmgaaattttaagtgtmcacaacaacgatgagattataagaatt360
tcccaggcctatgaaggattgtaccaacgmtcattggaatctgatatcaaaggagatacc420
tcaggaacattaaaaaagaattattag447
<210>52
<211>424
<212>dna
<213>diabroticavirgifera
<220>
<221>misc_feature
<222>(393)..(395)
<223>n是a,c,g,或t
<400>52
ttgttacaagctggagaacttctctttgctggaaccgaagagtcagtatttaatgctgta60
ttctgtcaaagaaataaaccacaattgaatttgatattcgacaaatatgaagaaattgtt120
gggcatcccattgaaaaagccattgaaaacgagttttcaggaaatgctaaacaagccatg180
ttacaccttatccagagcgtaagagatcaagttgcatatttggtaaccaggctgcatgat240
tcaatggcaggcgtcggtactgacgatagaactttaatcagaattgttgtttcgagatct300
gaaatcgatctagaggaaatcaaacaatgctatgaagaaatctacagtaaaaccttggct360
gataggatagcggatgacacatctggcgactannnaaaagccttattagccgttgttggt420
taag424
<210>53
<211>397
<212>dna
<213>diabroticavirgifera
<400>53
agatgttggctgcatctagagaattacacaagttcttccatgattgcaaggatgtactga60
gcagaatagtggaaaaacaggtatccatgtctgatgaattgggaagggacgcaggagctg120
tcaatgcccttcaacgcaaacaccagaacttcctccaagacctacaaacactccaatcga180
acgtccaacaaatccaagaagaatcagctaaacttcaagctagctatgccggtgatagag240
ctaaagaaatcaccaacagggagcaggaagtggtagcagcctgggcagccttgcagatcg300
cttgcgatcagagacacggaaaattgagcgatactggtgatctattcaaattctttaact360
tggtacgaacgttgatgcagtggatggacgaatggac397
<210>54
<211>490
<212>dna
<213>diabroticavirgifera
<400>54
gcagatgaacaccagcgagaaaccaagagatgttagtggtgttgaattgttgatgaacaa60
ccatcagacactcaaggctgagatcgaagccagagaagacaactttacggcttgtatttc120
tttaggaaaggaattgttgagccgtaatcactatgctagtgctgatattaaggataaatt180
ggtcgcgttgacgaatcaaaggaatgctgtactacagaggtgggaagaaagatgggagaa240
cttgcaactcatcctcgaggtataccaattcgccagagatgcggccgtcgccgaagcatg300
gttgatcgcacaagaaccttacttgatgagccaagaactaggacacaccattgacgacgt360
tgaaaacttgataaagaaacacgaagcgttcgaaaaatcggcagcggcgcaagaagagag420
attcagtgctttggagagactgacgacgttcgaattgagagaaataaagaggaaacaaga480
agctgcccag490
<210>55
<211>330
<212>dna
<213>diabroticavirgifera
<400>55
agtgaaatgttagcaaatataacatccaagtttcgtaattgtacttgctcagttagaaaa60
tattctgtagtttcactatcttcaaccgaaaatagaataaatgtagaacctcgcgaactt120
gcctttcctccaaaatatcaagaacctcgacaagtttggttggagagtttagatacgata180
gacgacaaaaaattgggtattcttgagctgcatcctgatgtttttgctactaatccaaga240
atagatattatacatcaaaatgttagatggcaaagtttatatagatatgtaagctatgct300
catacaaagtcaagatttgaagtgagaggt330
<210>56
<211>320
<212>dna
<213>diabroticavirgifera
<400>56
caaagtcaagatttgaagtgagaggtggaggtcgaaaaccgtggccgcaaaagggattgg60
gacgtgctcgacatggttcaattagaagtccactttggagaggtggaggagttgttcatg120
gaccaaaatctccaacccctcatttttacatgattccattctacacccgtttgctgggtt180
tgactagcgcactttcagtaaaatttgcccaagatgacttgcacgttgtggatagtctag240
atctgccaactgacgaacaaagttatatagaagagctggtcaaaagccgcttttgggggt300
ccttcttgttttatttgtag320
<210>57
<211>503
<212>dna
<213>人工的
<220>
<223>yfp序列
<400>57
caccatgggctccagcggcgccctgctgttccacggcaagatcccctacgtggtggagat60
ggagggcaatgtggatggccacaccttcagcatccgcggcaagggctacggcgatgccag120
cgtgggcaaggtggatgcccagttcatctgcaccaccggcgatgtgcccgtgccctggag180
caccctggtgaccaccctgacctacggcgcccagtgcttcgccaagtacggccccgagct240
gaaggatttctacaagagctgcatgcccgatggctacgtgcaggagcgcaccatcacctt300
cgagggcgatggcaatttcaagacccgcgccgaggtgaccttcgagaatggcagcgtgta360
caatcgcgtgaagctgaatggccagggcttcaagaaggatggccacgtgctgggcaagaa420
tctggagttcaatttcaccccccactgcctgtacatctggggcgatcaggccaatcacgg480
cctgaagagcgccttcaagatct503
<210>58
<211>46
<212>dna
<213>人工的
<220>
<223>引物ann-f1t7
<400>58
ttaatacgactcactatagggagagctccaacagtggttccttatc46
<210>59
<211>29
<212>dna
<213>人工的
<220>
<223>引物ann-r1
<400>59
ctaataattcttttttaatgttcctgagg29
<210>60
<211>22
<212>dna
<213>人工的
<220>
<223>引物ann-f1
<400>60
gctccaacagtggttccttatc22
<210>61
<211>53
<212>dna
<213>人工的
<220>
<223>引物ann-r1t7
<400>61
ttaatacgactcactatagggagactaataattcttttttaatgttcctgagg53
<210>62
<211>48
<212>dna
<213>人工的
<220>
<223>引物ann-f2t7
<400>62
ttaatacgactcactatagggagattgttacaagctggagaacttctc48
<210>63
<211>24
<212>dna
<213>人工的
<220>
<223>引物ann-r2
<400>63
cttaaccaacaacggctaataagg24
<210>64
<211>24
<212>dna
<213>人工的
<220>
<223>引物ann-f2
<400>64
ttgttacaagctggagaacttctc24
<210>65
<211>48
<212>dna
<213>人工的
<220>
<223>引物ann-r2t7
<400>65
ttaatacgactcactatagggagacttaaccaacaacggctaataagg48
<210>66
<211>47
<212>dna
<213>人工的
<220>
<223>引物betasp2-f1t7
<400>66
ttaatacgactcactatagggagaagatgttggctgcatctagagaa47
<210>67
<211>22
<212>dna
<213>人工的
<220>
<223>引物betasp2-r1
<400>67
gtccattcgtccatccactgca22
<210>68
<211>23
<212>dna
<213>人工的
<220>
<223>引物betasp2-f1
<400>68
agatgttggctgcatctagagaa23
<210>69
<211>46
<212>dna
<213>人工的
<220>
<223>引物betasp2-r1t7
<400>69
ttaatacgactcactatagggagagtccattcgtccatccactgca46
<210>70
<211>46
<212>dna
<213>人工的
<220>
<223>引物betasp2-f2t7
<400>70
ttaatacgactcactatagggagagcagatgaacaccagcgagaaa46
<210>71
<211>22
<212>dna
<213>人工的
<220>
<223>引物betasp2-r2
<400>71
ctgggcagcttcttgtttcctc22
<210>72
<211>22
<212>dna
<213>人工的
<220>
<223>引物betasp2-f2
<400>72
gcagatgaacaccagcgagaaa22
<210>73
<211>46
<212>dna
<213>人工的
<220>
<223>引物betasp2-r2t7
<400>73
ttaatacgactcactatagggagactgggcagcttcttgtttcctc46
<210>74
<211>51
<212>dna
<213>人工的
<220>
<223>引物l4-f1t7
<400>74
ttaatacgactcactatagggagaagtgaaatgttagcaaatataacatcc51
<210>75
<211>26
<212>dna
<213>人工的
<220>
<223>引物l4-r1
<400>75
acctctcacttcaaatcttgactttg26
<210>76
<211>27
<212>dna
<213>人工的
<220>
<223>引物l4-f1
<400>76
agtgaaatgttagcaaatataacatcc27
<210>77
<211>50
<212>dna
<213>人工的
<220>
<223>引物l4-r1t7
<400>77
ttaatacgactcactatagggagaacctctcacttcaaatcttgactttg50
<210>78
<211>50
<212>dna
<213>人工的
<220>
<223>引物l4-f2t7
<400>78
ttaatacgactcactatagggagacaaagtcaagatttgaagtgagaggt50
<210>79
<211>25
<212>dna
<213>人工的
<220>
<223>引物l4-r2
<400>79
ctacaaataaaacaagaaggacccc25
<210>80
<211>26
<212>dna
<213>人工的
<220>
<223>引物l4-f2
<400>80
caaagtcaagatttgaagtgagaggt26
<210>81
<211>49
<212>dna
<213>人工的
<220>
<223>引物l4-r2t7
<400>81
ttaatacgactcactatagggagactacaaataaaacaagaaggacccc49
<210>82
<211>47
<212>dna
<213>人工的
<220>
<223>引物yfp-ft7
<400>82
ttaatacgactcactatagggagacaccatgggctccagcggcgccc47
<210>83
<211>23
<212>dna
<213>人工的
<220>
<223>引物yfp-r
<400>83
agatcttgaaggcgctcttcagg23
<210>84
<211>23
<212>dna
<213>人工的
<220>
<223>引物yfp-f
<400>84
caccatgggctccagcggcgccc23
<210>85
<211>47
<212>dna
<213>人工的
<220>
<223>引物yfp-rt7
<400>85
ttaatacgactcactatagggagaagatcttgaaggcgctcttcagg47
<210>86
<211>260
<212>dna
<213>diabroticavirgifera
<400>86
ttcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaaagtctaaa60
aacttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttgggagaaac120
tatttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtacttccatga180
caatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgttaaaccaag240
aaaacctttggctgaagata260
<210>87
<211>551
<212>dna
<213>diabroticavirgifera
<400>87
caggtccgatggctgcaataagaaaaaaattggtgattgttggtgacggagcatgtggaa60
aaacatgtcttttaatagtatttagtaaagatcagttcccagaagtatacgtgcccactg120
tattcgaaaactatgtggcggatattgaagtagatggaaagcaggtagaactcgccttat180
gggataccgcaggtcaagaagattatgatcgtctgcggcccctttcatatccagacaccg240
atgtgattttaatgtgcttctcagtagactcccccgactcattagaaaacatcccagaaa300
aatggaccccagaggtaaaacacttttgtccaaatgtgccaattattctggtaggcaata360
agaaggatctgcgtaacgatcccaataccattaacgaacttaaaaagatgaaacaagaac420
cagtaaaacctcaagacggaagatccatggcagagaaaatcaatgcttttgcatatctag480
aatgttctgccaaaagtaaggagggtgtaagagaggtttttgaaaatgctacccgagctg540
cactgcaagtc551
<210>88
<211>19
<212>dna
<213>diabroticavirgifera
<400>88
gttaatcttatccattaat19
<210>89
<211>104
<212>dna
<213>diabroticavirgifera
<400>89
tttttgcttgatccttatctgtctgttatccccgctcttacttcgtaatttatttaatgt60
ccgccattttcgttcgtagttgaataggcgttaactttgtgtgt104
<210>90
<211>98
<212>dna
<213>diabroticavirgifera
<400>90
gttacgtgtgttaattaatttactaagggggacgttctgtcatgatgtattgacaagggg60
gcttctgcatgatgtattgactaacaaaaataaacccc98
<210>91
<211>745
<212>dna
<213>人工的
<220>
<223>caf1-180形成发夹序列与内部克隆位点(caf1-180hairpin-formingsequencewithinternalcloningsites)
<400>91
tcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaaagtctaaaa60
acttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttgggagaaact120
atttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtacttccatgac180
aatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgttaaaccaaga240
aaacctttggctgaagataaaaaacattttaggcctaggtatgtttctgcttctaccttt300
gatatatatataataattatcactaattagtagtaatatagtatttcaagtatttttttc360
aaaataaaagaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattt420
taatttataacttttctaatatatgaccaaaacatggtgatgtgcaggtaccggtaaaat480
gttttttatcttcagccaaaggttttcttggtttaacaaatttgctttttttcctccatg540
tgccataatatgcaggtctacgattgtcatggaagtacaaaaacttggctttgtgtcttt600
gaaggccagatttgtcttcgcaaatagtttctcccaaatcaacaagaatgatttcatcat660
caagagattcttcatatggccaagtttttagacttttacctatagcctttcctgatgaca720
gatcttttaaatatgaagcttccga745
<210>92
<211>929
<212>dna
<213>人工的
<220>
<223>2版本的caf1-180发夹rna-形成序列与st-ls1内含子和没有玉米共有序列(version2caf1-180hairpinrna-formingsequencewithst-ls1intronandwithoutmaizeconsensussequence)
<400>92
cagaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacaga60
catgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagt120
ttcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatc180
tgggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggag240
aagtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgatt300
tgcccacttatttaacccgatttacttgggacatcgccaaatatcccatcggatccaggc360
ctaggtatgtttctgcttctacctttgatatatatataataattatcactaattagtagt420
aatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttt480
tctgtagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaaca540
tggtgatgtgcaggtatttaaataccggtccatgggatgggatatttggcgatgtcccaa600
gtaaatcgggttaaataagtgggcaaatcgctgttattggcctgcaggttttctgaccac660
ttgtccctctgttcttcaagtacttctccaaggtactgagctaccttcctggtaacctgc720
tccacgaacccatcaagcttgcccagatcatcagaaagtcctactaactgatccagagta780
ccaactttcaagtccgggatttgaaacttgtagttggttgacaaattattctgtttactt840
gtcatattattcatcgtgtcccatgtctgttgacaggtcttatcccctggggcagatatc900
aaccaatactcagtcatttcttctgagct929
<210>93
<211>905
<212>dna
<213>人工的
<220>
<223>vatpasec形成发夹序列与内部克隆位点和玉米共有序列
<400>93
aagaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacaga60
catgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagt120
ttcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatc180
tgggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggag240
aagtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgatt300
tgcccacttatttaacccgatttacttgggacatcgccaaatatcccatcaggcctaggt360
atgtttctgcttctacctttgatatatatataataattatcactaattagtagtaatata420
gtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctgta480
gtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggtga540
tgtgcaggtaccggtgatgggatatttggcgatgtcccaagtaaatcgggttaaataagt600
gggcaaatcgctgttattggcctgcaggttttctgaccacttgtccctctgttcttcaag660
tacttctccaaggtactgagctaccttcctggtaacctgctccacgaacccatcaagctt720
gcccagatcatcagaaagtcctactaactgatccagagtaccaactttcaagtccgggat780
ttgaaacttgtagttggttgacaaattattctgtttacttgtcatattattcatcgtgtc840
ccatgtctgttgacaggtcttatcccctggggcagatatcaaccaatactcagtcatttc900
ttctt905
<210>94
<211>929
<212>dna
<213>人工的
<220>
<223>2版本的vatpasec发夹rna-形成序列与st-ls1内含子和没有玉米共有序列(version2vatpasechairpinrna-formingsequencewithst-ls1intronandwithoutmaizeconsensussequence)
<400>94
cagaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacaga60
catgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagt120
ttcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatc180
tgggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggag240
aagtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgatt300
tgcccacttatttaacccgatttacttgggacatcgccaaatatcccatcggatccaggc360
ctaggtatgtttctgcttctacctttgatatatatataataattatcactaattagtagt420
aatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttt480
tctgtagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaaca540
tggtgatgtgcaggtatttaaataccggtccatgggatgggatatttggcgatgtcccaa600
gtaaatcgggttaaataagtgggcaaatcgctgttattggcctgcaggttttctgaccac660
ttgtccctctgttcttcaagtacttctccaaggtactgagctaccttcctggtaacctgc720
tccacgaacccatcaagcttgcccagatcatcagaaagtcctactaactgatccagagta780
ccaactttcaagtccgggatttgaaacttgtagttggttgacaaattattctgtttactt840
gtcatattattcatcgtgtcccatgtctgttgacaggtcttatcccctggggcagatatc900
aaccaatactcagtcatttcttctgagct929
<210>95
<211>501
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的caf1-180有意义链的片段(segmentofacaf1-180sensestrandcontaininganst-ls1intron)
<400>95
ggtacctaatcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaa60
agtctaaaaacttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttg120
ggagaaactatttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtac180
ttccatgacaatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgtt240
aaaccaagaaaacctttggctgaagataaaaaacattttgactagtaccggttgggaaag300
gtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaata360
tagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctg420
tagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggt480
gatgtgcaggttgatccgcgg501
<210>96
<211>286
<212>dna
<213>人工的
<220>
<223>caf1-180dna反义链片段
<400>96
ccgcggttaaaaatgttttttatcttcagccaaaggttttcttggtttaacaaatttgct60
ttttttcctccatgtgccataatatgcaggtctacgattgtcatggaagtacaaaaactt120
ggctttgtgtctttgaaggccagatttgtcttcgcaaatagtttctcccaaatcaacaag180
aatgatttcatcatcaagagattcttcatatggccaagtttttagacttttacctatagc240
ctttcctgatgacagatcttttaaatatgaagcttccgaagagctc286
<210>97
<211>781
<212>dna
<213>人工的
<220>
<223>caf1-180发夹rna-形成dna序列含有st-ls1内含子
<400>97
ggtacctaatcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaa60
agtctaaaaacttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttg120
ggagaaactatttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtac180
ttccatgacaatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgtt240
aaaccaagaaaacctttggctgaagataaaaaacattttgactagtaccggttgggaaag300
gtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaata360
tagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctg420
tagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggt480
gatgtgcaggttgatccgcggttaaaaatgttttttatcttcagccaaaggttttcttgg540
tttaacaaatttgctttttttcctccatgtgccataatatgcaggtctacgattgtcatg600
gaagtacaaaaacttggctttgtgtctttgaaggccagatttgtcttcgcaaatagtttc660
tcccaaatcaacaagaatgatttcatcatcaagagattcttcatatggccaagtttttag720
acttttacctatagcctttcctgatgacagatcttttaaatatgaagcttccgaagagct780
c781
<210>98
<211>511
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的vatpasecdna有意义链的片段(segmentofavatpasecdnasensestrandcontaininganst-ls1intron)
<400>98
ggtacctaaactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagttt120
caaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatctg180
ggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggagaa240
gtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaatagactagtaccg300
gttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaatt360
agtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctat420
tgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgacc480
aaaacatggtgatgtgcaggttgatccgcgg511
<210>99
<211>296
<212>dna
<213>人工的
<220>
<223>vatpasecdna反义链片段(segmentofavatpasecdnaantisensestrand)
<400>99
ccgcggttatattggcctgcaggttttctgaccacttgtccctctgttcttcaagtactt60
ctccaaggtactgagctaccttcctggtaacctgctccacgaacccatcaagcttgccca120
gatcatcagaaagtcctactaactgatccagagtaccaactttcaagtccgggatttgaa180
acttgtagttggttgacaaattattctgtttacttgtcatattattcatcgtgtcccatg240
tctgttgacaggtcttatcccctggggcagatatcaaccaatactcagtagagctc296
<210>100
<211>801
<212>dna
<213>人工的
<220>
<223>vatpasec发夹rna-形成dna序列含有st-ls1内含子
<400>100
ggtacctaaactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagttt120
caaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatctg180
ggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggagaa240
gtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaatagactagtaccg300
gttgggaaaggtatgtttctgcttctacctttgatatatatataataattatcactaatt360
agtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctat420
tgcttttctgtagtttataagtgtgtatattttaatttataacttttctaatatatgacc480
aaaacatggtgatgtgcaggttgatccgcggttatattggcctgcaggttttctgaccac540
ttgtccctctgttcttcaagtacttctccaaggtactgagctaccttcctggtaacctgc600
tccacgaacccatcaagcttgcccagatcatcagaaagtcctactaactgatccagagta660
ccaactttcaagtccgggatttgaaacttgtagttggttgacaaattattctgtttactt720
gtcatattattcatcgtgtcccatgtctgttgacaggtcttatcccctggggcagatatc780
aaccaatactcagtagagctc801
<210>101
<211>402
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的vatpasecdna有意义链的片段(segmentofavatpasecdnasensestrandcontaininganst-ls1intron)
<400>101
ggtacctaaactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagttt120
caaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatctg180
gactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataat240
tatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtag300
tatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttct360
aatatatgaccaaaacatggtgatgtgcaggttgatccgcgg402
<210>102
<211>187
<212>dna
<213>人工的
<220>
<223>vatpasecdna反义链片段
<400>102
ccgcggttacagatcatcagaaagtcctactaactgatccagagtaccaactttcaagtc60
cgggatttgaaacttgtagttggttgacaaattattctgtttacttgtcatattattcat120
cgtgtcccatgtctgttgacaggtcttatcccctggggcagatatcaaccaatactcagt180
agagctc187
<210>103
<211>583
<212>dna
<213>人工的
<220>
<223>vatpasec发夹rna-形成dna序列含有st-ls1内含子
<400>103
ggtacctaaactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagttt120
caaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatctg180
gactagtaccggttgggaaaggtatgtttctgcttctacctttgatatatatataataat240
tatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaaagaatgtag300
tatatagctattgcttttctgtagtttataagtgtgtatattttaatttataacttttct360
aatatatgaccaaaacatggtgatgtgcaggttgatccgcggttacagatcatcagaaag420
tcctactaactgatccagagtaccaactttcaagtccgggatttgaaacttgtagttggt480
tgacaaattattctgtttacttgtcatattattcatcgtgtcccatgtctgttgacaggt540
cttatcccctggggcagatatcaaccaatactcagtagagctc583
<210>104
<211>531
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的vatpasehdna有意义链的片段
<400>104
ggtacctaagtattgcagcaaagagctacagatataagaaatcaacatgtaaactggcaa60
tcctatttccagtcacaaatgatttcgcaggaagactacaacttcatagtagcttttgat120
gtgactgatagtgcaaaaagagaagctctcctgaaaagcgacagaaaccaatgtgctcaa180
acattgttgaaccttttggggcatgtatccaaggaccaaactctgcagtacattttagtt240
ttggttgatgatatgttacaggagaacagaagtcgtgtagagatatttcacgaatatgcc300
aaccaaaaggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatata360
tataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaa420
agaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttat480
aacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcgg531
<210>105
<211>316
<212>dna
<213>人工的
<220>
<223>vatpasehdna反义链片段(segmentofavatpasehdnaantisensestrand)
<400>105
ccgcggttacttttggttggcatattcgtgaaatatctctacacgacttctgttctcctg60
taacatatcatcaaccaaaactaaaatgtactgcagagtttggtccttggatacatgccc120
caaaaggttcaacaatgtttgagcacattggtttctgtcgcttttcaggagagcttctct180
ttttgcactatcagtcacatcaaaagctactatgaagttgtagtcttcctgcgaaatcat240
ttgtgactggaaataggattgccagtttacatgttgatttcttatatctgtagctctttg300
ctgcaatacagagctc316
<210>106
<211>841
<212>dna
<213>人工的
<220>
<223>vatpaseh发夹rna-形成dna序列含有st-ls1内含子(vatpasehhairpinrna-formingdnasequencecontaininganst-ls1intron)
<400>106
ggtacctaagtattgcagcaaagagctacagatataagaaatcaacatgtaaactggcaa60
tcctatttccagtcacaaatgatttcgcaggaagactacaacttcatagtagcttttgat120
gtgactgatagtgcaaaaagagaagctctcctgaaaagcgacagaaaccaatgtgctcaa180
acattgttgaaccttttggggcatgtatccaaggaccaaactctgcagtacattttagtt240
ttggttgatgatatgttacaggagaacagaagtcgtgtagagatatttcacgaatatgcc300
aaccaaaaggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatata360
tataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaa420
agaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttat480
aacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttacttttg540
gttggcatattcgtgaaatatctctacacgacttctgttctcctgtaacatatcatcaac600
caaaactaaaatgtactgcagagtttggtccttggatacatgccccaaaaggttcaacaa660
tgtttgagcacattggtttctgtcgcttttcaggagagcttctctttttgcactatcagt720
cacatcaaaagctactatgaagttgtagtcttcctgcgaaatcatttgtgactggaaata780
ggattgccagtttacatgttgatttcttatatctgtagctctttgctgcaatacagagct840
c841
<210>107
<211>531
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的rho1dna有意义链片段
<400>107
ggtacctaacatgaggaaaaacatgtcttttaatagtatttagtaaagatcagttcccag60
aagtatacgtgcccactgtattcgaaaactatgtggcggatattgaagtagatggaaagc120
aggtagaactcgccttatgggataccgcaggtcaagaagattatgatcgtctgcggcccc180
tttcatatccagacaccgatgtgattttaatgtgcttctcagtagactcccccgactcat240
tagaaaacatcccagaaaaatggaccccagaggtaaaacacttttgtccaaatgtgccaa300
ttattctgggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatata360
tataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaa420
agaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttat480
aacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcgg531
<210>108
<211>316
<212>dna
<213>人工的
<220>
<223>rho1dna反义链片段
<400>108
ccgcggttaccagaataattggcacatttggacaaaagtgttttacctctggggtccatt60
tttctgggatgttttctaatgagtcgggggagtctactgagaagcacattaaaatcacat120
cggtgtctggatatgaaaggggccgcagacgatcataatcttcttgacctgcggtatccc180
ataaggcgagttctacctgctttccatctacttcaatatccgccacatagttttcgaata240
cagtgggcacgtatacttctgggaactgatctttactaaatactattaaaagacatgttt300
ttcctcatgagagctc316
<210>109
<211>841
<212>dna
<213>人工的
<220>
<223>含有st-ls1内含子的rho1发夹rna-形成dna序列
<400>109
ggtacctaacatgaggaaaaacatgtcttttaatagtatttagtaaagatcagttcccag60
aagtatacgtgcccactgtattcgaaaactatgtggcggatattgaagtagatggaaagc120
aggtagaactcgccttatgggataccgcaggtcaagaagattatgatcgtctgcggcccc180
tttcatatccagacaccgatgtgattttaatgtgcttctcagtagactcccccgactcat240
tagaaaacatcccagaaaaatggaccccagaggtaaaacacttttgtccaaatgtgccaa300
ttattctgggactagtaccggttgggaaaggtatgtttctgcttctacctttgatatata360
tataataattatcactaattagtagtaatatagtatttcaagtatttttttcaaaataaa420
agaatgtagtatatagctattgcttttctgtagtttataagtgtgtatattttaatttat480
aacttttctaatatatgaccaaaacatggtgatgtgcaggttgatccgcggttaccagaa540
taattggcacatttggacaaaagtgttttacctctggggtccatttttctgggatgtttt600
ctaatgagtcgggggagtctactgagaagcacattaaaatcacatcggtgtctggatatg660
aaaggggccgcagacgatcataatcttcttgacctgcggtatcccataaggcgagttcta720
cctgctttccatctacttcaatatccgccacatagttttcgaatacagtgggcacgtata780
cttctgggaactgatctttactaaatactattaaaagacatgtttttcctcatgagagct840
c841
<210>110
<211>260
<212>dna
<213>人工的
<220>
<223>caf-180dna片段用于提供dsrna合成模板(segmentofacaf-180dnausedtoprovideatemplatefordsrna
synthesis)
<400>110
ttcggaagcttcatatttaaaagatctgtcatcaggaaaggctataggtaaaagtctaaa60
aacttggccatatgaagaatctcttgatgatgaaatcattcttgttgatttgggagaaac120
tatttgcgaagacaaatctggccttcaaagacacaaagccaagtttttgtacttccatga180
caatcgtagacctgcatattatggcacatggaggaaaaaaagcaaatttgttaaaccaag240
aaaacctttggctgaagata260
<210>111
<211>460
<212>dna
<213>人工的
<220>
<223>vatpasec区1dna片段用于提供dsrna合成模板(segmentofavatpasecregion1dnausedtoprovideatemplatefordsrnasynthesis)
<400>111
agaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacagac60
atgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagtt120
tcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatct180
gggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggaga240
agtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgattt300
gcccacttatttaacccgatttacttgggacatcgccaaatatcccatcaagcaatcgct360
ccgtaacatcgccgatataatcagcaaacaagttggccaaattgacgctgatctgaagac420
caaatcgtctgcgtacaataacttgaaaggaagtcttcag460
<210>112
<211>349
<212>dna
<213>人工的
<220>
<223>vatpasec区1(短的)dna片段用于提供dsrna合成模板
<400>112
agaagaaatgactgagtattggttgatatctgccccaggggataagacctgtcaacagac60
atgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagtt120
tcaaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatct180
gggcaagcttgatgggttcgtggagcaggttaccaggaaggtagctcagtaccttggaga240
agtacttgaagaacagagggacaagtggtcagaaaacctgcaggccaataacagcgattt300
gcccacttatttaacccgatttacttgggacatcgccaaatatcccatc349
<210>113
<211>500
<212>dna
<213>人工的
<220>
<223>vatpasec区2dna片段用于提供dsrna合成模板
<400>113
agaaacagacaggaagtttactaacaagaaacttggcagatttggtgaaaaaggaacatt60
ttattttggattctgaatatttgcagacattgttggttattgttccaaaagcccaattca120
acgaatggaatgcgacatacgaaaagattaccgacatgatagtccctcgttcttcacaac180
tgattaaacaggataacgaatacggtctatatactgtatccctattcaaaaaggtcgtcg240
aagaattcaagttacacgctagagaaaagaagttcatcgttcgtgatttcatatataacg300
aagaggaactggcagctggaaagaacgagataaccaaactcgtcaccgataaaaagaagc360
aatttggtccattagttagatggcttaaagtcaacttcagcgagtgtttctgcgcctgga420
ttcacgttaaagcgttgagggtatttgttgaatctgttttaagatatggcctcccgtcaa480
tttccaagctattttaatcc500
<210>114
<211>445
<212>dna
<213>人工的
<220>
<223>vatpaseh区1dna片段用于提供dsrna合成模板
<400>114
tcatgattgttccagatatgttggctgctacaagtgtattgcagcaaagagctacagata60
taagaaatcaacatgtaaactggcaatcctatttccagtcacaaatgatttcgcaggaag120
actacaacttcatagtagcttttgatgtgactgatagtgcaaaaagagaagctctcctga180
aaagcgacagaaaccaatgtgctcaaacattgttgaaccttttggggcatgtatccaagg240
accaaactctgcagtacattttagttttggttgatgatatgttacaggagaacagaagtc300
gtgtagagatatttcacgaatatgccaaccaaaagaaagaatctgtttggggtccatttt360
tgaaccttcttaaccgccaggatggatttattaccaacatgacttctaggatcatcgcta420
aaattgcttgttggtcgcagacact445
<210>115
<211>570
<212>dna
<213>人工的
<220>
<223>vatpaseh区2dna片段用于提供dsrna合成模板
<400>115
agaacattgtatagctatggtgcaatgcaaagtattgaaacaattggccattttagaaca60
gcgcaagttcgatgatgaagatgtgacagctgatgtcgaatttctaactgaaaaactaca120
gagttctgttcaggatctgagttccttcgatgaatattcaactgaagtcaaatctggacg180
tttggaatggtctccagtacacaagagcaagttctggcgggaaaatgcgcagcgcctcaa240
cgagaaaaactacgaattactccgtattctcattcatttgttagaaaccagcaaggaccc300
cttggtattgagcgtagctagtttcgatattggagagtatgttcgccattatcctcgcgg360
caaacacataatcgagcagttgggaggaaagcaattagttatgcaactcttagcccacga420
agatcccaatgtacgatatgaagctctgttggcagtccaaaaattgatggtccacaattg480
ggaataccttggtcgtcaacttgaaaaagagcaaagtacagataaaaacgcgccaaaggg540
tggagctcctgtcgcaggcaaggcgtaaat570
<210>116
<211>551
<212>dna
<213>人工的
<220>
<223>rho1dna片段用于提供dsrna合成模板
<400>116
caggtccgatggctgcaataagaaaaaaattggtgattgttggtgacggagcatgtggaa60
aaacatgtcttttaatagtatttagtaaagatcagttcccagaagtatacgtgcccactg120
tattcgaaaactatgtggcggatattgaagtagatggaaagcaggtagaactcgccttat180
gggataccgcaggtcaagaagattatgatcgtctgcggcccctttcatatccagacaccg240
atgtgattttaatgtgcttctcagtagactcccccgactcattagaaaacatcccagaaa300
aatggaccccagaggtaaaacacttttgtccaaatgtgccaattattctggtaggcaata360
agaaggatctgcgtaacgatcccaataccattaacgaacttaaaaagatgaaacaagaac420
cagtaaaacctcaagacggaagatccatggcagagaaaatcaatgcttttgcatatctag480
aatgttctgccaaaagtaaggagggtgtaagagaggtttttgaaaatgctacccgagctg540
cactgcaagtc551
<210>117
<211>15
<212>rna
<213>人工的
<220>
<223>vatpasecrna有意义链("vatpasec5'-15")用作dsrna
<400>117
augacugaguauugg15
<210>118
<211>25
<212>rna
<213>人工的
<220>
<223>vatpasecrna有意义链("vatpasec5'-25")用作dsrna
<400>118
augacugaguauugguugauaucug25
<210>119
<211>15
<212>rna
<213>人工的
<220>
<223>vatpasecrna有意义链("vatpasec3'-15")用作dsrna
<400>119
cuuucugaugaucug15
<210>120
<211>25
<212>rna
<213>人工的
<220>
<223>vatpasecrna有意义链("vatpasec3'-25")用作dsrna
<400>120
guuaguaggacuuucugaugaucug25
<210>121
<211>50
<212>dna
<213>人工的
<220>
<223>引物vatpasec-ft7
<400>121
ttaatacgactcactatagggagaatgactgagtattggttgatatctgc50
<210>122
<211>45
<212>dna
<213>人工的
<220>
<223>引物vatpasec-r50t7
<400>122
ttaatacgactcactatagggagatgttgacaggtcttatcccct45
<210>123
<211>54
<212>dna
<213>人工的
<220>
<223>引物vatpasec-r100t7
<400>123
ttaatacgactcactatagggagattgacaaattattctgtttacttgtcatat54
<210>124
<211>51
<212>dna
<213>人工的
<220>
<223>引物vatpasec-rt7
<400>124
ttaatacgactcactatagggagacagatcatcagaaagtcctactaactg51
<210>125
<211>48
<212>dna
<213>人工的
<220>
<223>引物vatpasec-f50t7
<400>125
ttaatacgactcactatagggagagacttgaaagttggtactctggat48
<210>126
<211>52
<212>dna
<213>人工的
<220>
<223>引物vatpasec-f100t7
<400>126
ttaatacgactcactatagggagagacaagtaaacagaataatttgtcaacc52
<210>127
<211>62
<212>dna
<213>人工的
<220>
<223>vatpasec有意义链("vatpasec5'-50")用作dsrna模板
<400>127
gggagaatgactgagtattggttgatatctgccccaggggataagacctgtcaacatctc60
cc62
<210>128
<211>112
<212>dna
<213>人工的
<220>
<223>vatpasec有意义链("vatpasec5'-100")用作dsrna模板
<400>128
gggagaatgactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaatctccc112
<210>129
<211>186
<212>dna
<213>人工的
<220>
<223>vatpasec有意义链("vatpasec-174")用作dsrna模板
<400>129
gggagaatgactgagtattggttgatatctgccccaggggataagacctgtcaacagaca60
tgggacacgatgaataatatgacaagtaaacagaataatttgtcaaccaactacaagttt120
caaatcccggacttgaaagttggtactctggatcagttagtaggactttctgatgatctg180
tctccc186
<210>130
<211>63
<212>dna
<213>人工的
<220>
<223>vatpasec有意义链("vatpasec3'-50")用作dsrna模板
<400>130
gggagagacttgaaagttggtactctggatcagttagtaggactttctgatgatctgtct60
ccc63
<210>131
<211>112
<212>dna
<213>人工的
<220>
<223>vatpasec有意义链("vatpasec3'-100")用作dsrna模板
<400>131
gggagagacaagtaaacagaataatttgtcaaccaactacaagtttcaaatcccggactt60
gaaagttggtactctggatcagttagtaggactttctgatgatctgtctccc112
<210>132
<211>20
<212>dna
<213>人工的
<220>
<223>引物mztipu67f
<400>132
agccaagccagtggtacttc20
<210>133
<211>22
<212>dna
<213>人工的
<220>
<223>引物mztipu67r
<400>133
tcgcagacaaagtagcaaatgt22
<210>134
<211>20
<212>dna
<213>人工的
<220>
<223>引物p5u76s
<400>134
ttgtgatgttggtggcgtat20
<210>135
<211>24
<212>dna
<213>人工的
<220>
<223>引物p5u76a
<400>135
tgttaaataaaaccccaaagatcg24
<210>136
<211>189
<212>dna
<213>人工的
<220>
<223>st-ls1内含子
<400>136
gtatgtttctgcttctacctttgatatatatataataattatcactaattagtagtaata60
tagtatttcaagtatttttttcaaaataaaagaatgtagtatatagctattgcttttctg120
tagtttataagtgtgtatattttaatttataacttttctaatatatgaccaaaacatggt180
gatgtgcag189
<210>137
<211>48
<212>dna
<213>diabroticavirgifera
<400>137
aaattggactaaacttttttgctcgtttwgttaatcttatccattaat48
<210>138
<211>102
<212>dna
<213>diabroticavirgifera
<400>138
gttacgtgtgttaattaatttactaagggggacgttctgtcatgatgtattgacaagggg60
gcttctgcatgatgtattgactaacaaaaataaaccccwtta102
- 还没有人留言评论。精彩留言会获得点赞!
- 诱导果实抗性控制病害的方法和所用制剂与流程
- 水稻白叶枯抗性基因Xa27的分子标记引物及其应用的制作方法与工艺
- 赋予对鞘翅目有害生物的抗性的核酸分子的制造方法与工艺
- 避免抗微生物剂抗性的抗腹泻制剂的制造方法与工艺
- 一种选育小麦条锈病持久抗性材料的方法与流程
- 一种产生PVY抗性的NteIF4E‑1基因突变体及其应用的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体GAMMA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体BETA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体ALPHA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体DELTA亚单位核酸分子的制造方法与工艺
- 一种选育小麦条锈病持久抗性材料的方法与流程
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体GAMMA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体BETA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体ALPHA亚单位核酸分子的制造方法与工艺
- 赋予鞘翅目和半翅目害虫抗性的COPI外被体DELTA亚单位核酸分子的制造方法与工艺
- 一个影响猪生长和PRRSV抗性的SLA‑DRB1启动子区分子标记及其引物和应用的制造方法与工艺
- 一个影响猪生长和PRRSV抗性的分子标记、检测方法及其应用与制造工艺
- 赋予鞘翅目害虫抗性的DRE4核酸分子的制造方法与工艺
- 一种提高茶树抗性且肥效高的茶树肥料及其制备方法与制造工艺
- 抗穿透的复合件的制作方法