使用谷氨酰胺合成酶基因内互补载体直接选择表达高水平异聚蛋白的细胞的制作方法
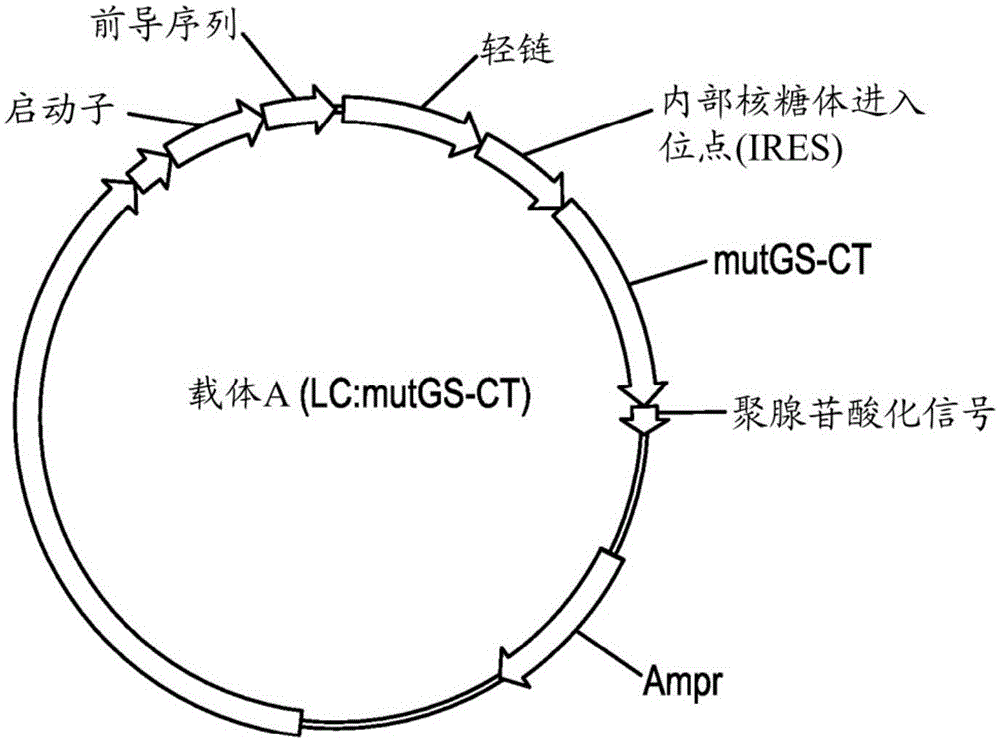
本申请要求2016年5月11日提交的美国临时申请号62/334,966的权益,所述临时申请特此以引用的方式整体并入。以电子方式提交的文本文件的描述本申请含有已经以ascii格式电子提交且特此以引用的方式整体并入的序列表。创建于2017年5月8日的序列表的计算机可读格式拷贝名为a-2011-wo-pct_seq_st25.txt且大小为79.1千字节。本发明涉及哺乳动物细胞培养物中的多肽的重组表达的一般领域。更具体地说,本发明涉及经设计以表达异聚多肽的重组工程化载体的细胞中的选择改进。发明背景异聚重组蛋白的表达通常需要组分链的类似表达,以避免不能并入至成熟蛋白中的链的不必要的合成。另外,大多数用于产生异聚重组蛋白的方法需要筛选许多克隆,以鉴别表达高水平的成熟异聚蛋白的每条链的相当高表达克隆。一种断裂选择性标记系统(在所述系统中选择性标记断裂成两个节段并且每个片段连接至异聚蛋白的一条链的表达)可用于鉴别表达两个选择性标记片段且因此表达异聚蛋白的两条链的那些细胞。谷氨酰胺合成酶(gs)通过氨与谷氨酸盐的缩合来催化谷氨酰胺生物合成。哺乳动物gs酶是由两个堆叠的五聚环组成的十聚体,其中十个活性位点定位在亚基的接点处。每个活性位点通过来自一个亚基的n-末端结构域(β-抓握结构域,其由残基25-112组成)的残基和通过来自相邻亚基的c-末端结构域(催化结构域,其由残基113-373组成)的残基来形成。酿酒酵母菌(s.cerevisiae)和大肠杆菌(e.coli)的遗传研究显示,某些gs突变展示出基因内互补,其中映射至gs基因的5′端的一些突变可与3′端的突变互补(mitchell,a.p.genetics111,243-258(1985);macneil,t.,等人journalofbacteriology150,1302-1313(1982))。因此,鉴别基于代谢酶的选择系统(诸如利用谷氨酰胺合成酶的基因内互补的系统)可证明适用于表达具有多于两条独特多肽链的分子(例如单克隆抗体和其他重组蛋白)。技术实现要素:本发明的一个实施方案提供一种载体,其包含:a.第一核酸,所述第一核酸编码第一多肽,和b.第二核酸,所述第二核酸编码第二多肽,其中所述第二多肽是选择性标记的初始突变体亚基,其中所述第一核酸的转录可操作地连接至所述第二核酸的转录,所述载体还包含:c.第三核酸,所述第三核酸编码第三多肽,其中所述第三多肽能够与所述第一多肽缔合以形成异聚复合物,以及d.第四核酸,所述第四核酸编码第四多肽,其中所述第四多肽是所述选择性标记的互补突变体亚基,其中所述第三核酸的转录可操作地连接至所述第四核酸的转录,其中所述选择性标记的所述初始突变体亚基和所述互补突变体亚基相互作用以提供可选择活性,并且进一步其中所述载体能够转染到哺乳动物细胞中并改进转染的细胞的选择。本发明的另一个实施方案提供一种载体,其包含:e.第一核酸,所述第一核酸编码第一多肽,和f.第二核酸,所述第二核酸编码第二多肽,其中所述第二多肽是选择性标记的初始片段,其中所述第一核酸的转录可操作地连接至所述第二核酸的转录,所述载体还包含:g.第三核酸,所述第三核酸编码第三多肽,其中所述第三多肽能够与所述第一多肽缔合以形成异聚复合物,以及h.第四核酸,所述第四核酸编码第四多肽,其中所述第四多肽是所述选择性标记的互补片段,其中所述第三核酸的转录可操作地连接至所述第四核酸的转录,其中所述选择性标记的所述初始突变体亚基和所述互补突变体亚基相互作用以提供可选择活性,并且进一步其中所述载体能够转染到哺乳动物细胞中并改进转染的细胞的选择。在另一实施方案中,异聚复合物是免疫球蛋白。在一个实施方案中,第一核酸编码免疫球蛋白重链,并且第三核酸编码免疫球蛋白轻链,在一个替代实施方案中,第一核酸编码免疫球蛋白轻链,并且第三核酸编码免疫球蛋白重链。在前述实施方案中,选择性标记是选自由以下组成的组的代谢酶:谷氨酰胺合成酶、苏氨酸脱水酶、腺苷酸基琥珀酸合成酶和谷氨酸脱氢酶。本发明可包括内部核糖体进入位点(ires);在一个实施方案中,ires发生在选自由以下组成的组的位点:在第一核酸与第二核酸之间的位点;在第三核酸与第四核酸之间的位点;以及在第一核酸与第二核酸之间和在第三核酸与第四核酸之间的位点。在另一实施方案中,内部核糖体进入位点包含seqidno:23。对于本文所述的实施方案中的任一个,当选择性标记的初始突变体亚基是谷氨酰胺合成酶的n-末端端突变体(mutgs-nt)时,选择性标记的互补突变体亚基是谷氨酰胺合成酶的c-末端突变体(mutgs-ct)。在本发明的一个这样的方面中,mutgs-nt包含两个或更多个选自由以下组成的组的突变:w60an61ad63ad63rs66a和d76a,并且mutgs-ct包含一个或多个选自由以下组成的组的突变:e134ae136ae196ae203an248ah253an255ar319a和r324a。在一个实施方案中,mutgs-nt是选自由以下组成的组:w60an61ad63a(mutgs-nt1;seqidno:6)、w60an61ad63as66a(mutgs-nt2;seqidno:10)w60an61ad63as66ad76a(mutgs-nt3;seqidno:25)和w60an61ad63rs66a(mutgs-nt4;seqidno:19);并且mutgs-ct是选自由以下组成的组:e134ae136ae196ae203a(mutgs-ct1;seqidno:21)和n248ah253an255a(mutgs-ct2;seqidno:22)。在其他实施方案中,谷氨酰胺合成酶的n-末端片段和c-末端片段通过在谷氨酰胺合成酶蛋白的一个或多个氨基酸处断裂所述蛋白来产生。在本发明的一个这样的方面中,谷氨酰胺合成酶的n-末端片段和/或c-末端片段在谷氨酰胺合成酶的选自由以下组成的组的氨基酸处断裂:e110、y104、s125、n126、e264、t111、n105、n126、q127以及n265本发明还提供一种分离的宿主细胞,其用如上文所述的载体转染、转化或转导。所述宿主细胞可选自由以下组成的组:cho、vero、bhk、hela、cos、mdck、293、3t3、骨髓瘤细胞系以及wi38细胞。本发明的一个方面提供一种产生异聚复合物的方法,所述方法包括在由所述宿主细胞表达所述异聚复合物的条件下培养这种宿主细胞的步骤。在一个实施方案中,异聚复合物是抗体。权利要求的本发明方法还可包括分离异聚复合物。本发明的一个实施方案提供一种表达系统,其包含:a)第一载体,所述第一载体编码双顺反子转录物,所述第一载体包含编码第一多肽的第一核酸,所述第一核酸可操作地连接至编码第二多肽的第二核酸,其中所述第二多肽是选择性标记的初始突变体亚基,和b)第二载体,所述第二载体编码双顺反子转录物,所述第二载体包含编码第三多肽的第三核酸,所述第三核酸可操作地连接至编码第四多肽的第四核酸,其中所述第四多肽是所述选择性标记的互补突变体亚基,所述互补突变体亚基能够与所述选择性标记的所述初始突变体亚基缔合以提供可选择活性,进一步其中所述第三多肽能够与所述第一多肽缔合以形成异聚复合物;并且进一步其中所述表达系统能够转染到哺乳动物细胞中并改进所述细胞的选择。本发明的另一个实施方案提供一种表达系统,其包含:c)第一载体,所述第一载体编码双顺反子转录物,所述第一载体包含编码第一多肽的第一核酸,所述第一核酸可操作地连接至编码第二多肽的第二核酸,其中所述第二多肽是选择性标记的初始片段,知d)第二载体,所述第二载体编码双顺反子转录物,所述第二载体包含编码第三多肽的第三核酸,所述第三核酸可操作地连接至编码第四多肽的第四核酸,其中所述第四多肽是所述选择性标记的互补片段,所述互补片段能够与所述选择性标记的所述初始片段缔合以提供可选择活性,进一步其中所述第三多肽能够与所述第一多肽缔合以形成异聚复合物;并且进一步其中所述表达系统能够转染到哺乳动物细胞中并改进所述细胞的选择。在另一实施方案中,异聚复合物是免疫球蛋白。在一个实施方案中,第一核酸编码免疫球蛋白重链,并且第三核酸编码免疫球蛋白轻链,在一个替代实施方案中,第一核酸编码免疫球蛋白轻链,并且第三核酸编码免疫球蛋白重链。在前述实施方案中,选择性标记是选自由以下组成的组的代谢酶:谷氨酰胺合成酶、苏氨酸脱水酶、腺苷酸基琥珀酸合成酶和谷氨酸脱氢酶。上文所述的本发明表达系统可包含内部核糖体进入位点(ires);在一个实施方案中,ires发生在选自由以下组成的组的位点:在第一核酸与第二核酸之间的位点;在第三核酸与第四核酸之间的位点;以及在第一核酸与第二核酸之间和在第三核酸与第四核酸之间的位点。在另一实施方案中,内部核糖体进入位点包含seqidno:23。在本文所述的表达系统中,当选择性标记的初始突变体亚基是谷氨酰胺合成酶的n-末端突变体(mutgs-nt)时,选择性标记的互补突变体亚基是谷氨酰胺合成酶的c-末端突变体(mutgs-ct)。在本发明的一个这样的方面中,mutgs-nt包含两个或更多个选自由以下组成的组的突变:w60an61ad63ad63rs66a和d76a,并且mutgs-ct包含一个或多个选自由以下组成的组的突变:e134ae136ae196ae203an248ah253an255ar319a和r324a。在一个实施方案中,mutgs-nt是选自由以下组成的组:w60an61ad63a(mutgs-nt1;seqidno:6)、w60an61ad63as66a(mutgs-nt2;seqidno:10)w60an61ad63as66ad76a(mutgs-nt3;seqidno:25)和w60an61ad63rs66a(mutgs-nt4;seqidno:19);并且mutgs-ct是选自由以下组成的组:e134ae136ae196ae203a(mutgs-ct1;seqidno:21)和n248ah253an255a(mutgs-ct2;seqidno:22)。在其他实施方案中,谷氨酰胺合成酶的n-末端片段和c-末端片段通过在谷氨酰胺合成酶蛋白的一个或多个氨基酸处断裂所述蛋白来产生。在本发明的一个这样的方面中,谷氨酰胺合成酶的n-末端片段和/或c-末端片段在谷氨酰胺合成酶的选自由以下组成的组的氨基酸处断裂:e110、y104、s125、n126、e264、t111、n105、n126、q127和n265。本发明还提供一种分离的宿主细胞,其用如上文所述的载体转染、转化或转导。所述宿主细胞可选自由以下组成的组:cho、vero、bhk、hela、cos、mdck、293、3t3、骨髓瘤细胞系以及wi38细胞。本发明的一个方面提供一种产生异聚复合物的方法,所述方法包括在由所述宿主细胞表达所述异聚复合物的条件下培养这种宿主细胞的步骤。在一个实施方案中,异聚复合物是抗体。权利要求的本发明方法还可包括分离异聚复合物。在一个实施方案中,本发明提供一种表达系统,其包含:第一载体,所述第一载体包含编码抗体的重链的第一核酸,所述第一核酸可操作地连接至编码n-末端突变体谷氨酰胺合成酶(mutgs-nt)的第二核酸;以及第二载体,所述第二载体包含编码抗体的轻链的第三核酸,所述第三核酸可操作地连接至编码c-末端突变体谷氨酰胺合成酶(mutgs-ct)的第四核酸,其中谷氨酰胺合成酶的每个突变体亚基当单独表达时不具有可选择活性,并且所述mutgs-nt与所述mutgs-ct的共表达提供谷氨酰胺合成酶活性,其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择。在一个实施方案中,本发明提供一种表达系统,其包含:第一载体,所述第一载体包含编码抗体的重链的第一核酸,所述第一核酸可操作地连接至编码谷氨酰胺合成酶的n-末端片段的第二核酸;以及第二载体,所述第二载体包含编码抗体的轻链的第三核酸,所述第三核酸可操作地连接至编码谷氨酰胺合成酶的c-末端片段的第四核酸,其中谷氨酰胺合成酶的每个突变体亚基当单独表达时不具有可选择活性,并且谷氨酰胺合成酶的所述n-末端片段与所述c-末端片段的共表达提供谷氨酰胺合成酶活性,其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择。在另一实施方案中,本发明提供一种表达系统,其包含:第一载体,所述第一载体包含编码抗体的轻链的第一核酸,所述第一核酸可操作地连接至编码n-末端突变体谷氨酰胺合成酶(mutgs-nt)的第二核酸;以及第二载体,所述第二载体包含编码抗体的重链的第三核酸,所述第三核酸可操作地连接至编码c-末端突变体谷氨酰胺合成酶(mutgs-ct)的第四核酸,其中谷氨酰胺合成酶的每个突变体亚基当单独表达时不具有可选择活性,并且所述mutgs-nt与所述mutgs-ct的共表达提供谷氨酰胺合成酶活性,其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择。在另一个实施方案中,本发明提供一种表达系统,其包含:第一载体,所述第一载体包含编码抗体的轻链的第一核酸,所述第一核酸可操作地连接至编码谷氨酰胺合成酶的n-末端片段的第二核酸;以及第二载体,所述第二载体包含编码抗体的重链的第三核酸,所述第三核酸可操作地连接至编码谷氨酰胺合成酶的c-末端片段的第四核酸,其中谷氨酰胺合成酶的每个片段当单独表达时不具有可选择活性,并且谷氨酰胺合成酶的所述n-末端片段与所述c-末端片段的共表达提供谷氨酰胺合成酶活性,其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择。对于前述实施方案中的任一个,在本发明的一个方面中,mutgs-nt包含两个或更多个选自由以下组成的组的突变:w60an61ad63ad63rs66a和d76a,并且mutgs-ct包含一个或多个选自由以下组成的组的突变:e134ae136ae196ae203an248ah253an255ar319a和r324a。在一个实施方案中,mutgs-nt是选自由以下组成的组:w60an61ad63a(mutgs-nt1;seqidno:6)、w60an61ad63as66a(mutgs-nt2;seqidno:10)w60an61ad63as66ad76a(mutgs-nt3;seqidno:25)和w60an61ad63rs66a(mutgs-nt4;seqidno:19);并且mutgs-ct是选自由以下组成的组:e134ae136ae196ae203a(mutgs-ct1;seqidno:21)和n248ah253an255a(mutgs-ct2;seqidno:22)。在其他实施方案中,谷氨酰胺合成酶的n-末端片段和c-末端片段通过在谷氨酰胺合成酶蛋白的一个或多个氨基酸处断裂所述蛋白来产生。在本发明的一个这样的方面中,谷氨酰胺合成酶的n-末端片段和/或c-末端片段在谷氨酰胺合成酶的选自由以下组成的组的氨基酸处断裂:e110、y104、s125、n126、e264、t111、n105、n126、q127和n265。类似地,本发明还提供一种分离的宿主细胞,其用如上文所述的载体转染、转化或转导。所述宿主细胞可选自由以下组成的组:cho、vero、bhk、hela、cos、mdck、293、3t3、骨髓瘤细胞系以及wi38细胞。本发明的一个方面提供一种产生异聚复合物的方法,所述方法包括在由所述宿主细胞表达所述异聚复合物的条件下培养这种宿主细胞的步骤。在一个实施方案中,异聚复合物是抗体。权利要求的本发明方法还可包括分离异聚复合物。本发明的其他方面包括一种表达系统,其包含:第一载体,所述第一载体编码双顺反子转录物,所述第一载体包含编码所需多肽的第一核酸,所述第一核酸可操作地连接至编码选择性标记的初始突变体亚基的第二核酸;以及第二载体,所述第二载体编码双顺反子转录物,所述第二载体包含第三核酸,所述第三核酸可操作地连接至编码选择性标记的互补突变体亚基的第四核酸,其中所述选择性标记的所述第一突变体亚基和所述互补突变体亚基缔合以提供可选择活性,并且其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择,并且进一步其中所述第一核酸编码抗体重链且所述第三核酸编码抗体轻链。在一个实施方案中,选择性标记是谷氨酰胺合成酶。本发明的其他方面包括一种表达系统,其包含:第一载体,所述第一载体编码双顺反子转录物,所述第一载体包含编码所需多肽的第一核酸,所述第一核酸可操作地连接至编码选择性标记的初始片段的第二核酸;以及第二载体,所述第二载体编码双顺反子转录物,所述第二载体包含第三核酸,所述第三核酸可操作地连接至编码选择性标记的互补片段的第四核酸,其中所述选择性标记的所述第一片段和互补片段缔合以提供可选择活性,并且其中所述表达系统能够转染到哺乳动物细胞中并改进转染的细胞的选择,并且进一步其中所述第一核酸编码抗体重链且所述第三核酸编码抗体轻链。在一个实施方案中,选择性标记是谷氨酰胺合成酶。在本发明的一个这样的方面中,谷氨酰胺合成酶包含两个互补突变;因此,mutgs-nt包含两个或更多个选自由以下组成的组的突变:w60an61ad63ad63rs66a和d76a,并且mutgs-ct包含一个或多个选自由以下组成的组的突变:e134ae136ae196ae203an248ah253an255ar319a和r324a。在一个实施方案中,mutgs-nt是选自由以下组成的组:w60an61ad63a(mutgs-nt1;seqidno:6)、w60an61ad63as66a(mutgs-nt2;seqidno:10)w60an61ad63as66ad76a(mutgs-nt3;seqidno:25)和w60an61ad63rs66a(mutgs-nt4;seqidno:19);并且mutgs-ct是选自由以下组成的组:e134ae136ae196ae203a(mutgs-ct1;seqidno:21)和n248ah253an255a(mutgs-ct2;seqidno:22)。在其他此类实施方案中,谷氨酰胺合成酶的n-末端片段和c-末端片段通过在谷氨酰胺合成酶蛋白的一个或多个氨基酸处断裂所述蛋白来产生。在本发明的一个这样的方面中,谷氨酰胺合成酶的n-末端片段和/或c-末端片段在谷氨酰胺合成酶的选自由以下组成的组的氨基酸处断裂:e110、y104、s125、n126、e264、t111、n105、n126、q127和n265。本发明还提供一种分离的宿主细胞,其用如上文所述的载体转染、转化或转导。所述宿主细胞可选自由以下组成的组:cho、vero、bhk、hela、cos、mdck、293、3t3、骨髓瘤细胞系以及wi38细胞。本发明的一个方面提供一种产生异聚复合物的方法,所述方法包括在由所述宿主细胞表达所述异聚复合物的条件下培养这种宿主细胞的步骤。在一个实施方案中,异聚复合物是抗体。权利要求的本发明方法还可包括分离异聚复合物。附图说明图1是实施例1-5中所利用的核酸构建体的示意性图示,所述核酸构建体各自包含选择性标记的亚基并表达不同多肽,所述多肽可缔合以在细胞中形成异聚复合物。缩写是如下:mugs,突变体谷氨酰胺合成酶;ampr,氨比西林抗性。图1(a)描绘载体a(lc:mutgs-ct),并且图1(b)描绘载体b(hc:mutgs-nt)。图2是示出拯救(rescue)实验1的结果的图。将每种型式(1-5)的构建体a和b转染到cho-k1gs敲除细胞中并测试其在无谷氨酰胺的培养基中拯救细胞的能力。将表达全长gs酶的质粒用作对照。图3是示出单独a或b片段均不能拯救ko细胞系的图,从而确认使gs酶活性恢复需要这两种片段。图4是实施例6中所使用的表达载体的示意性图示,所述表达载体各自含有抗体轻链或重链、ires和谷氨酰胺合成酶片段。图5是示出gs片段载体在抗体lc和hc基因的下游的使用的图。载体组合2a-lc和2b-hc能够拯救gs敲除细胞。图6是示出以下的图:在10d进料分批生产率测定中的来自构建体2选定池的细胞的生产率,并将其与使用表达全长谷氨酰胺合成酶的载体选择的对照池进行比较。具体实施方式重组异聚复合物在细胞中的有效产生在所述复合物的每种组分成比例地且大量地表达的情况下改进。本发明通过提供选择重组工程化细胞的方法和组合物来改进转染的细胞的选择,所述重组工程化细胞以成比例量表达多于一种异聚多肽,以使得所述多肽可有效缔合以形成表达水平高于传统制备的异聚复合物的异聚复合物。本发明的优点还在于,其减少选择表达高水平的所需重组异聚多肽复合物的细胞所需的时间,这是本发明所提供的改进转染的细胞的选择的另一方面。尽管本申请中所使用的术语在本领域内是标准的,但是本文中提供某些术语的定义以确保权利要求中的含义的清楚性和明确性。单位、前缀、和符号都以其si(国际单位制)可接受的形式表示。本文中所述的数值范围包括限定所述范围的数字,并且包括并支持所限定的范围内的每个整数。除非另外指明,否则本文中所述的方法和技术通常是根据本领域中熟知的常规方法且如本说明书通篇所引用和讨论的各种一般性和更特定参考文献中所述来执行。参见例如,sambrook等人.molecularcloning:alaboratorymanual,第3版,coldspringharborlaboratorypress,coldspringharbor,n.y.(2001)和ausubel等人,currentprotocolsinmolecularbiology,greenepublishingassociates(1992)以及harlow和laneantibodies:alaboratorymanualcoldspringharborlaboratorypress,coldspringharbor,n.y.(1990)。所公开的方法可适用于在搅拌的槽式反应器(包括传统分批和进料分批细胞培养,其可包含但不必包含旋转过滤器)、灌注系统(包括交替切向流(“atf”)培养、声学(acoustic)灌注系统、深层过滤器(depthfilter)灌注系统和其他系统)、中空纤维生物反应器(hfb,其在一些情况下可用于灌注过程中)中生长的粘附培养物或悬浮培养物,以及各种其他细胞培养方法(参见例如,tao等人,(2003)biotechnol.bioeng.82:751-65;kuystermans和al-rubeai,(2011)“bioreactorsystemsforproducingantibodvfrommammaliancells”在antibodyexpressionandproduction中,cellengineering7:25-52,al-rubeai(ed)springer;catapano等人,(2009)“bioreactordesignandscale-up”在cellandtissuereactionengineering:principlesandpractice中,eibl等人(编)springer-verlag,所述文献均以引用的方式整体并入本文)。除非另外具体地指明,否则如本文所用,术语“一个/种(a/an)”是指一个或多个(种)。此外,除非另外为上下文所需,否则单数术语将包括复数且复数术语将包括单数。一般来说,结合本文所述的细胞和组织培养、分子生物学、免疫学、微生物学、遗传学和蛋白质和核酸化学和杂交使用的命名法是本领域中所熟知且常用的那些。本公开提供调节表达“目标蛋白质”的细胞培养物的性质的方法;“目标蛋白质”包括天然存在的蛋白质、重组蛋白和工程化的蛋白质(例如,自然界中不存在且已由人设计和/或创造的蛋白质)。目标蛋白质可为(但不必为)已知或怀疑具有治疗相关性的蛋白质。目标蛋白质的具体实例包括:抗原结合蛋白(如本文所述和定义)、肽体(即,包含直接或间接地融合至其他分子诸如抗体的fc域的一个或多个肽的分子,其中所述肽部分特异性结合至所需靶标;所述一个或多个肽可融合至fc区或插入到fc-环中,或修饰的fc分子,例如,如美国专利临时申请号us2006/0140934中所述,所述临时申请以引用的方式整体并入本文)、融合蛋白(例如,fc融合蛋白,其中fc片段融合至蛋白质或肽,包括肽体)、细胞因子、生长因子、激素和其他天然存在的分泌蛋白质以及天然存在的蛋白质的突变形式。术语“异聚复合物”是指包括通过至少两个不同分子的缔合所形成的分子复合物。缔合可为非共价相互作用或共价连接,例如,二硫键。两个不同分子通常为两种不同多肽,然而,本发明考虑到在多肽与核酸之间以及在不同核酸之间的异聚复合物。在一个实施方案中,异聚复合物提供功能性活性,诸如,结合底物的能力(例如,能够结合对应抗原的免疫球蛋白)、酶活性等。在一个实施方案中,本发明的异聚复合物被分泌到产生其的宿主细胞的培养基中。如本文所用,术语“缔合”或“相互作用”是指描述至少两个分子之间的关系,其中一个分子结合至其他分子和/或影响其他分子的活性。相互作用可包括两种多肽(或多肽和核酸)的直接或间接结合、或分子活性通过另一分子功能性活化或抑制。在一个具体实施方案中,异聚复合物是免疫球蛋白分子。脊椎动物系统中的免疫球蛋白为由两条相同轻链和两条相同重链构成的抗体。这四条链通过二硫键连接在一起,以使得每条轻链与重链连接,并且重链跨其全部尾部连接,从而形成y形异聚复合物。可操纵编码免疫球蛋白分子的dna以产生能够编码重组蛋白(诸如亲和力增强的抗体)或其他基于抗体的多肽的dna的许多技术是已知的(参见例如,larrick等人(1989),biotechnology7:934-938;reichmann等人(1988),nature332:323-327;roberts等人(1987),nature328:731-734;verhoeyen等人(1988),science239:1534-1536;chaudhary等人(1989),nature339:394-397)。本发明中还可使用产生人抗体的重组细胞(诸如使用抗体文库和/或转基因动物来制备,和任选地在体外进一步修饰),以及人源化抗体。参见例如,cabilly等人,美国专利号4,816,567;cabilly等人,欧洲专利号0,125,023b1;boss等人,美国专利号4,816,397;boss等人,欧洲专利号0,120,694b1;neuberger等人,wo86/01533;neuberger等人,欧洲专利号0,194,276b1;winter,美国专利号5,225,539;winter,欧洲专利号0,239,400b1;queen等人,欧洲专利号0,451,216b1;以及padlan等人,欧洲专利号0,519,596a1。例如,本发明可用于诱导免疫特异性地识别特异性细胞靶标的人和/或人源化抗体的表达,所述靶标例如人egf受体、her-2/neu抗原、cea抗原、前列腺特异性膜抗原(psma)、cd5、cd11a、cd18、ngf、cd20、cd45、ep-cam、其他癌症表面分子、tnf-α、tgf-b1、vegf、其他细胞因子、α4β7整联蛋白、ige、病毒蛋白(例如巨细胞病毒)等,仅举几个例子。除免疫球蛋白之外,异聚复合物的实例包括但不限于任何异二聚或异寡聚蛋白,例如,bmp2/bmp7、成骨蛋白、白介素1转化酶(ice)、各种白介素受体(例如,il-18受体、il-13受体、il-4受体和il-7受体)、细胞核的受体诸如类视黄醇受体、t细胞受体、整联蛋白(诸如细胞粘附分子)、整联蛋白、肿瘤坏死因子受体以及i类和ii类主要组织相容性复合物蛋白(mhc)的可溶性形式和膜结合形式。对于为受体的异聚复合物,本发明涵盖多肽的可溶性形式和膜结合形式两者。可根据本发明产生的额外异聚蛋白的描述可见于例如以下中:humancytokines:handbookforbasicandclinicalresearch,第ii卷(aggarwal和gutterman,编blackwellsciences,cambridgemass.,1998);growthfactors:apracticalapproach(mckay和leigh,编oxforduniversitypressinc.,newyork,1993);以及thecytokinehandbook(awthompson,编;academicpress,sandiegocalif.;1991)。如本文所用,术语“融合蛋白”是指融合至异源蛋白或肽的蛋白质或蛋白质的结构域(例如,可溶性细胞外结构域)。此类融合蛋白的实例包括表达为与免疫球蛋白分子的一部分的融合的蛋白、表达为与拉链部分的融合蛋白的蛋白以及多功能性蛋白诸如细胞因子和生长因子的融合蛋白(即,gm-csf和il-3、mgf和il-3)。wo93/08207和wo96/40918描述了被称为cd40l的分子的各种可溶性寡聚形式的制备,分别包括免疫球蛋白融合蛋白和拉链融合蛋白;其中所讨论的技术可适用于其他蛋白质。本文所述的分子中的任一种均可表达为融合蛋白,包括但不限于细胞受体分子的细胞外结构域、酶、激素、细胞因子、免疫球蛋白分子的一部分、拉链结构域以及表位。术语“抗原结合蛋白”是以其最广泛意义使用并且是指包含结合至抗原或靶标的一部分并且任选地包含允许抗原结合部分采用促进抗原结合蛋白与抗原的结合的构象的支架或框架部分的蛋白质。抗原结合蛋白的实例包括人抗体、人源化抗体;嵌合抗体;重组抗体;单链抗体;双抗体;三抗体;四抗体;fab片段;f(ab′)2片段;igd抗体;ige抗体;igm抗体;igg1抗体;igg2抗体;igg3抗体;或igg4抗体,以及其片段。抗原结合蛋白可包含例如具有移植的cdr或cdr衍生物的替代蛋白支架或人工支架。此类支架包括但不限于,抗体来源的支架,其包含被引入以例如使抗原结合蛋白的三维结构稳定的突变;以及完全合成的支架,其包含例如生物相容性聚合物。参见例如,korndorfer等人,2003,proteins:structure,function,andbioinformatics,53(1):121-129(2003);roque等人,biotechnol.prog.20:639-654(2004)。此外,可使用肽抗体模拟物(“pam”),以及使用纤连蛋白组分作为支架的基于抗体模拟物的支架。抗原结合蛋白可具有例如天然存在的免疫球蛋白的结构。“免疫球蛋白”是四聚体分子。在天然存在的免疫球蛋白中,每个四聚体由两对相同的多肽链组成,每对具有一条“轻”链(约25kda)和一条“重”链(约50-70kda)。每条链的氨基末端部分包括主要负责抗原识别的具有约100至110个或更多个氨基酸的可变区。每条链的羧基末端部分限定主要负责效应子功能的恒定区。人轻链被分类为κ和λ轻链。重链被分类为μ、δ、γ、α或ε,并且将抗体的同种型分别定义为igm、igd、igg、iga和ige。天然存在的免疫球蛋白链展现出通过三个高变区连接的相对保守的框架区(fr)的相同通用结构,所述高变区又称为互补决定区或cdr。从n-末端至c-末端,轻链和重链两者均包含结构域fr1、cdr1、fr2、cdr2、fr3、cdr3以及fr4。每个结构域的氨基酸的指定可根据kabat等人在sequencesofproteinsofimmunologicalinterest,第5版,usdept.ofhealthandhumanservices,phs,nih,nih出版号91-3242,(1991)中的定义来进行。根据需要,cdr还可根据替代命名方案来重新定义,诸如chothia的命名方案(参见chothia和lesk,(1987)j.mol.biol.196:901-917;chothia等人,(1989)nature342:878-883;或honegger和pluckthun,(2001)j.mol.biol.309:657-670)。在本公开的上下文中,抗原结合蛋白描述为当解离常数(kd)≤10-8m时“特异性结合”或“选择性结合”其靶抗原。所述抗体当kd≤5x10-9m时以“高亲和力”特异性结合抗原,并且当kd≤5x10-10m时以“极高亲和力”特异性结合抗原。除非另外说明,否则术语“抗体”包括对任何同种型或亚类的糖基化和非糖基化的免疫球蛋白或与完整抗体竞争以特异性结合的其抗原结合区的提及。此外,除非另外说明,否则术语“抗体”是指完整免疫球蛋白或与完整抗体竞争以特异性结合的其抗原结合部分。抗原结合部分可通过重组dna技术或通过完整抗体的酶促或化学裂解来产生,并且可形成目标蛋白质的元件。抗原结合部分尤其包括fab、fab’、f(ab’)2、fv、结构域抗体(dab)、包括互补决定区(cdr)的片段、单链抗体(scfv)、嵌合抗体、双抗体、三抗体、四抗体以及含有足以赋予与多肽的特异性抗原结合的至少一部分免疫球蛋白的多肽。术语“抗体”是指结构和/或功能是基于抗体(例如,全长或全免疫球蛋白分子)的结构和/或功能,和/或由抗体或其片段的可变重链(vh)和/或可变轻链(vl)引起的分子。抗体构建体因此能够结合至其特异性靶标或抗原。此外,根据本发明的抗体构建体的结合结构域包含抗体的允许靶标结合的最低结构要求。这种最低要求可例如通过存在至少三个轻链cdr(即,vl区的cdr1、cdr2和cdr3)和/或三个重链cdr(即,vh区的cdr1、cdr2和cdr3),优选所有六个cdr来限定。限定抗体的最低结构要求的替代方法是分别限定特定靶标的结构内的抗体的表位(包含表位区(表位簇)的靶蛋白的蛋白质结构域),或通过参考与限定的抗体的表位竞争的特定抗体。根据本发明的构建体所基于的抗体包括例如单克隆抗体、重组抗体、嵌合抗体、去免疫化抗体、人源化抗体和人抗体。fab片段是具有vl、vh、cl和ch1结构域的单价片段;f(ab′)2片段是具有通过铰链区的二硫桥连接的两个fab片段的二价片段;fd片段具有vh和ch1结构域;fv片段具有抗体的单臂的vl和vh结构域;dab片段具有vh结构域、vl结构域、或vh或vl结构域的抗原结合片段;分离的互补决定区(cdr);以及单链fv(scfv)(美国专利号6,846,634、6,696,245、美国申请公布号05/0202512、04/0202995、04/0038291、04/0009507、03/0039958、ward等人,(1989)nature341:544-546)。单链抗体(scfv)是其中vl和vh区经由接头(例如,氨基酸残基的合成序列)连接以形成连续蛋白链的抗体,其中接头足够长以允许蛋白链自身折叠并形成单价抗原结合位点(参见例如,bird等人,science242:423-26(1988)和huston等人,(1988)proc.natl.acad.sci.usa85:5879-83)。双抗体是包含两条多肽链的二价抗体,其中每条多肽链包含通过接头连接的vh和vl结构域,所述接头太短以致不允许同一链上两个结构域之间的配对,因此允许每个结构域与另一多肽链上的互补结构域配对(参见例如,holliger等人,(1993)proc.natl.acad.sci.usa90:6444-48;和poljak等人,(1994)structure2:1121-23)。如果双抗体的两条多肽链是相同的,则由其配对所产生的双抗体将具有两个相同的抗原结合位点。具有不同序列的多肽链可用于制备具有两个不同抗原结合位点的双抗体。类似地,三抗体和四抗体是分别包含三条多肽链和四条多肽链并且分别形成三个抗原结合位点和四个抗原结合位点的抗体,所述抗原结合位点可相同或不同。此外,术语“抗体”包括单价构建体、二价构建体和多价(polyvalent/multivalent)构建体和因此双特异性构建体(其特异性结合至仅两种抗原结构)、以及多特异性(polyspecific/multispecific)构建体(其通过不同结合域特异性结合多于两种抗原结构,例如三种、四种或更多种)。此外,术语“抗体”包括由仅一条多肽链组成的分子以及由多于一条链组成的分子,所述链可为相同的(同二聚体、同三聚体或同寡聚物)或不同的(异二聚体、异三聚体或异寡聚物)。上文所鉴别的抗体和变体或其衍生物的实例尤其描述于以下中:harlow和lane,antibodiesalaboratorymanual,cshlpress(1988)和usingantibodies:alaboratorymanual,cshlpress(1999);kontermann和dübel,antibodyengineering,springer,第2版2010;以及little,recombinantantibodiesforimmunotherapy,cambridgeuniversitypress2009。如本文所用,术语“双特异性”是指为“至少双特异性”的抗体,即,其包含至少第一结合结构域和第二结合结构域,其中所述第一结合结构域结合至一种抗原或靶标(在此:靶细胞表面抗原),并且所述第二结合结构域结合至另一抗原或靶标。因此,双特异性抗体构建体包含针对至少两种不同抗原或靶标的特异性。双特异性抗体还可涵盖多特异性抗体构建体,诸如三特异性抗体构建体(后者包括三个结合结构域)或具有多于三种(例如四种、五种......)特异性的构建体。一个或多个cdr可共价地或非共价地并入到分子中以使所述分子为抗原结合蛋白。抗原结合蛋白可并入一个或多个cdr作为较大多肽链的一部分,可将一个或多个cdr共价地连接至另一多肽链,或可非共价地并入一个或多个cdr。cdr允许抗原结合蛋白特异性结合至特定目标抗原。抗原结合蛋白可具有一个或多个结合位点。如果存在多于一个结合位点,则所述结合位点可彼此相同或可不同。例如,天然存在的人免疫球蛋白通常具有两个相同的结合位点,而“双特异性”或“双功能性”抗体具有两个不同的结合位点。为清楚起见,并且如本文所述,应指出,抗原结合蛋白可以但不必源自人(例如,人抗体),并且在一些情况下将包含非人蛋白质,例如大鼠或鼠类蛋白,并且在其他情况下,抗原结合蛋白可包含人蛋白质和非人蛋白质的杂合体(例如,人源化抗体)。目标蛋白质可包含人抗体。术语“人抗体”包括具有来源于人免疫球蛋白序列的一个或多个可变区和恒定区的所有抗体。在一个实施方案中,所有可变结构域和恒定结构域都来源于人免疫球蛋白序列(完全人抗体)。此类抗体可以多种方式制备,包括通过用小鼠的目标抗原免疫,所述小鼠经遗传修饰以表达源自人重链和/或轻链编码基因的抗体,诸如来源于ultimabtm或系统的小鼠。还可采用基于噬菌体的方法。可替代地,目标蛋白质可包含人源化抗体。“人源化抗体”具有与来源于非人物种的抗体的序列相差一个或多个氨基酸取代、缺失和/或添加的序列,以使得当将人源化抗体施用至人受试者时,相较于非人物种抗体,所述人源化抗体不太可能诱导免疫应答,和/或诱导不太严重的免疫应答。在一个实施方案中,非人物种抗体的重链和/或轻链的框架和恒定结构域中的某些氨基酸被突变以产生人源化抗体。在另一实施方案中,将人抗体的一个或多个恒定结构域融合至非人物种的一个或多个可变结构域。如何制备人源化抗体的实例可见于美国专利号6,054,297、5,886,152和5,877,293中。如本文中所使用的术语“fc”区包含两个重链片段,其包含抗体的ch2和ch3结构域。将两个重链片段通过两个或更多个二硫键且通过ch3结构域的疏水相互作用来保持在一起。包含fc区的目标蛋白质(包括抗原结合蛋白和fc融合蛋白)形成本公开的另一方面。“半抗体(hemibody)”是免疫功能性免疫球蛋白构建体,其包含完整的重链、完整的轻链和与完整的重链的fc区配对的第二重链fc区。可以但不必采用接头来连接重链fc区和第二重链fc区。在具体实施方案中,半抗体是本文中所公开的抗原结合蛋白的单价形式。在其他实施方案中,可采用带电残基对来将一个fc区与第二fc区缔合。半抗体可为本公开的上下文中的目标蛋白质。术语“宿主细胞”是指经遗传工程化以表达具有商业或科学利益的多肽的细胞。遗传工程化细胞系涉及以编码重组多核苷酸分子(“目标基因”)的核酸转染、转化或转导细胞,和/或以其他方式改变(例如,通过同源重组和重组细胞与非重组小的基因活化或融合),以便导致宿主细胞表达所需的重组多肽。用于遗传工程化细胞和/或细胞系以表达目标多肽的方法和载体是本领域技术人员熟知的;例如,各种技术说明于以下中:currentprotocolsinmolecularbiology,ausubel等人,编(wiley&sons,newyork,1988,和季度更新);sambrook等人,molecularcloning:alaboratorymanual(coldspringlaboratorypress,1989);kaufman,r.j.,largescalemammaliancellculture,1990,第15-69页。所述术语包括亲本细胞的子代,无论所述子代在形态上或在遗传构成上是否与原始亲本细胞相同,只要目标基因存在即可。细胞培养物可包含一个或多个宿主细胞。术语“杂交瘤”是指由永生化细胞和抗体产生细胞的融合产生的细胞或细胞的子代。所得杂交瘤是产生抗体的永生化细胞。用以产生杂交瘤的单独细胞可来自任何哺乳动物来源,包括但不限于仓鼠、大鼠、猪、兔、绵羊、山羊和人。所述术语还涵盖当异杂交骨髓瘤融合时产生的三源杂交瘤细胞系,三源杂交瘤细胞系是人细胞与小鼠骨髓瘤细胞系之间的融合的产物,随后与浆细胞融合。所述术语旨在包括产生抗体的任何永生化杂交细胞系,例如像四源杂交瘤(quadroma)(参见例如,milstein等人,(1983)nature,537:3053)。术语“培养物”和“细胞培养物”可互换使用,并且是指在适合于细胞群体的存活和/或生长的条件下维持于培养基中的细胞群体。如本领域技术人员将清楚的,这些术语还指包含细胞群体和所述群体所悬浮的培养基的组合。术语“多肽”和“蛋白质”(例如,如目标蛋白质或目标多肽的上下文中所使用)在本文中可互换地用以指氨基酸残基的聚合物。所述术语还适用于一个或多个氨基酸残基为对应天然存在的氨基酸的类似物或模拟物的氨基酸聚合物,以及天然存在的氨基酸聚合物。所述术语还可涵盖经修饰(例如通过添加碳水化合物残基以形成糖蛋白)或磷酸化的氨基酸聚合物。多肽和蛋白质可由天然存在且非重组细胞产生,或多肽和蛋白质可通过遗传工程化或重组细胞产生。多肽和蛋白质可包含具有天然蛋白质的氨基酸序列的分子、或具有天然序列的一个或多个氨基酸的缺失、添加和/或取代的分子。术语“多肽”和“蛋白质”涵盖仅包含天然存在的氨基酸的分子,以及包含非天然存在的氨基酸的分子。非天然存在的氨基酸(根据需要,其可取代本文所公开的任何序列中所见的任何天然存在的氨基酸)的实例包括(但不限于):4-羟基脯氨酸、γ-羧基谷氨酸盐、ε-n,n,n-三甲基赖氨酸、ε-n-乙酰基赖氨酸、o-磷酸丝氨酸、n-乙酰基丝氨酸、n-甲酰基甲硫氨酸、3-甲基组氨酸、5-羟基赖氨酸、σ-n-甲基精氨酸和其他类似氨基酸和亚氨基酸(例如,4-羟基脯氨酸)。在本文所使用的多肽记法中,根据标准用法和惯例,左侧方向是氨基末端方向,并且右侧方向是羧基末端方向。“细胞培养”或“培养”是指细胞在多细胞生物体或组织外的生长和繁殖。哺乳动物的合适的培养条件是本领域中已知的。参见例如,animalcellculture:apracticalapproach,d.rickwood,编,oxforduniversitypress,newyork(1992)。哺乳动物细胞可以悬浮的方式或在连接至固体底物的同时培养。可使用流体化床生物反应器、中空纤维生物反应器、滚瓶、摇瓶或搅拌的槽式生物反应器(具有或不具有微载体)。在一个实施方案中,使用500l至2000l生物反应器。在一个实施方案中,使用1000l至2000l生物反应器。术语“细胞培养基(cellculturingmedium)”(还称为“培养基(culturemedium)”、“细胞培养基(cellculturemedia)”、“组织培养基”)是指用于生长细胞(例如,动物或哺乳动物细胞),并且一般提供来自以下的至少一种或多种组分的任何营养素溶液:能源(通常呈碳水化合物诸如葡萄糖的形式);所有必需氨基酸的一种或多种,并且一般是二十种碱性氨基酸加半胱氨酸;维生素和/或通常以低浓度需要的其他有机化合物;脂质或游离脂肪酸;以及痕量要素,例如,通常以低浓度(通常在微摩尔范围内)需要的无机化合物或天然存在的要素。营养素溶液可任选地补充有额外任选组分以使细胞的生长优化,诸如激素和其他生长因子,例如,胰岛素、转铁蛋白、表皮生长因子、血清等;盐,例如,钙、镁和磷酸盐,以及缓冲液,例如,hepes;核苷和碱,例如,腺苷、胸苷、次黄嘌呤;以及蛋白质和组织水解产物,例如,水解的动物或植物蛋白(蛋白胨或蛋白胨混合物,其可获自动物副产品、纯化的明胶或植物物质);抗生素,例如,庆大霉素;细胞保护剂或表面活性剂,诸如f68(还称为f68和p188);非离子三嵌段共聚物,其由侧接两条亲水性聚氧丙烯(聚(氧化丙烯))链的中心疏水性聚氧丙烯(聚(氧化丙烯))链组成;聚胺,例如,腐胺、亚精胺和精胺(参见例如,wipo公布号wo2008/154014);以及丙酮酸盐(参见例如美国专利号8053238),其取决于待培养的细胞的需要和/或所需的细胞培养参数。细胞培养基包括任何细胞培养过程通常采用的和/或已知与任何细胞培养过程(诸如但不限于,细胞的分批、延长分批、补料分批和/或灌注或连续培养)一起使用的那些细胞培养基。“基础”或(分批)细胞培养基是指通常用以起始细胞培养且足够完整以支持细胞培养的细胞培养基。“生长”细胞培养基是指通常用于指数生长的时期期间(“生长期”)的细胞培养中,并且足够完整以支持在此期期间的细胞培养的细胞培养基。生长细胞培养基还可含有选择剂,其向并入宿主细胞系中的选择性标记赋予抗性或存活。此类选择剂包括但不限于遗传霉素(g4118)、新霉素、潮霉素b、嘌呤霉素、博莱霉素、甲硫氨酸亚砜亚胺、甲氨蝶呤、无谷氨酰胺细胞培养基,缺乏甘氨酸、次黄嘌呤和胸苷或单独缺乏胸苷的细胞培养基。“产生”细胞培养基是指通常用于在当指数生长结束且蛋白质产生开始时的过渡期间(“过渡”和/或“产物”期)的细胞培养中,并且足够完整以维持在此阶段期间的所需的细胞密度、存活力和/或产物效价的细胞培养基。“灌注”细胞培养基是指通常用于通过灌注培养或连续培养方法维持的细胞培养中,并且足够完全支持此过程期间的细胞培养的细胞培养基。灌注细胞培养基制剂可比基础细胞培养基制剂更富集或更浓缩以适应用来移除废培养基的方法。灌注细胞培养基可在生长期和产生期期间使用。浓缩细胞培养基可含有维持细胞培养所必需的营养素中的一些或所有;具体地说,浓缩培养基可含有鉴别或已知在细胞培养的产生期的时程期间被消耗的营养素。浓缩培养基可基于几乎任何细胞培养基制剂。这种浓缩进料培养基可含有以例如约2x、3x、4x、5x、6x、7x、8x、9x、10x、12x、14x、16x、20x、30x、50x、100x、200x、400x、600x、800x、或甚至约1000x组分的正常量的细胞培养基的组分中的一些或所有。用于制备细胞培养基的组分可完全研磨成粉末培养基制剂;与根据需要添加至细胞培养基的液体补充物一起部分研磨;或以完全液体形式添加至细胞培养物中。细胞培养物还可补充有可能难以配制或在细胞培养物中快速耗减的特定营养素的独立浓缩进料。此类营养素可以是氨基酸,诸如酪氨酸、半胱氨酸和/或胱氨酸(参见例如,wipo公布号2012/145682)。在一个实施方案中,浓缩酪氨酸溶液独立地进料至生长于含有酪氨酸的细胞培养基中的细胞培养物,以使得细胞培养物中酪氨酸的浓度不超过8mm。在另一个实施方案中,浓缩酪氨酸和胱氨酸溶液独立地进料至生长于缺乏酪氨酸、胱氨酸或半胱氨酸的细胞培养基中的细胞培养物。独立进料可在产生期之前或开始时开始。独立进料可通过在与浓缩进料培养基相同或不同天分批进料至细胞培养基来实现。独立进料还可在与灌注培养基相同或不同天灌注。“无血清”应用于不含有动物血清(诸如胎牛血清)的细胞培养基。各种组织培养基(包括限定的培养基)是可商购的,例如,可使用以下细胞培养基中的任一种或组合:rpmi-1640培养基、rpmi-1641培养基、杜氏改良的伊格尔氏培养基(dmem)、最低必需培养基eagle、f-12k培养基、汉姆氏f12培养基、伊思考夫氏改良的杜氏培养基、麦考伊氏5a培养基、莱柏维兹氏l-15培养基和无血清培养基诸如ex-celltm300系列(jrhbiosciences,lenexa,kansas)以及其他细胞培养基。还可获得此类细胞培养基的无血清型式。细胞培养基可补充有额外或浓度增加的组分,诸如氨基酸、盐、糖、维生素、激素、生长因子、缓冲液、抗生素、脂质、痕量要素等,其取决于待培养的细胞的需要和/或所需的细胞培养参数。术语“生物反应器”是指可用于细胞培养物的生长的任何容器。本公开的细胞培养物可生长于生物反应器中,其可基于通过生物反应器中的细胞生长所产生的目标蛋白质的应用来选择。生物反应器可具有任何大小,只要其可用于细胞的培养即可;通常,生物反应器的大小适合于生长于其内部的细胞培养物的体积。通常,生物反应器将为至少1升,并且可以是2升、5升、10升、50升、100升、200升、250升、500升、1,000升、1500升、2000升、2,500升、5,000升、8,000升、10,000升、12,000升或更大,或可为之间的任何体积。生物反应器的内部条件(包括但不限于ph和温度)可在培养期期间控制。本领域普通技术人员基于相关考虑应意识到并且应该能够选择用于实践本发明的合适的生物反应器。“细胞密度”是指给定体积的培养基中细胞的数目。“活细胞密度”是指给定体积的培养基中活细胞的数目,如通过标准存活力测定(诸如台盼蓝染料排阻方法)所确定。术语“细胞存活力”是指培养物中的细胞在一组给定培养条件或实验变化下存活的能力。术语还指在特定时间为活着的细胞相对于在所述时间培养物中细胞(活的和死亡的)的总数目的部分。“填充细胞体积”(pcv)(还称为“百分比填充细胞体积”(%pcv))是细胞所占据的体积与细胞培养物的总体积的比率,表达为百分比(参见stettler,等人,(2006)biotechnolbioeng.dec20:95(6):1228-33)。填充细胞体积是细胞密度和细胞直径的函数;填充细胞体积的增加可起因于细胞密度或细胞直径或两者的增加。填充细胞体积是细胞培养物中固体含量的量度。固体在收集和下游纯化期间被移除。更多固体是指在收获和下游纯化步骤期间将固体物质与所需产物分离的更多努力。此外,所需产物在收获过程期间可变得捕获在固体中并损失,从而导致产物产量降低。因为宿主细胞大小不同且细胞培养物还含有死亡和濒死细胞和其他细胞碎片,所以填充细胞体积是比细胞密度或活细胞密度更准确描述细胞培养物中固体含量的方式。例如,取决于细胞的大小,具有50x106个细胞/ml的细胞密度的2000l培养物可具有许多不同的填充细胞体积。此外,一些细胞当处于生长停滞状态时大小将增加,所以在生长停滞之前和在生长停滞之后的填充细胞体积将可能是不同的,其归因于细胞大小增加导致生物质增加。“生长停滞”(其还可称为“细胞生长停滞”)是细胞的数目停止增加或当细胞周期不再进展时的点。生长停滞可通过确定细胞培养物的活细胞密度来监测。生长停滞状态的一些细胞大小可增加,但数目不增加,所以生长停滞培养物的填充细胞体积可增加。如果细胞不是健康衰退,则生长停滞可通过逆转引起生长停滞的条件来在一定程度上逆转。术语“效价”是指给定量培养基体积中由细胞培养物所产生的目标多肽或蛋白质(其可为天然存在的或目标重组蛋白)的总量。效价可以每毫升(或体积的其他量度)培养基毫克或微克多肽或蛋白质的单位表达。“累积效价”是在培养的时程期间细胞所产生的效价,且可例如通过测量每日效价并使用所述值计算累积效价来确定。术语“进料分批培养”是指悬浮培养的形式,并且是指在培养过程开始后一次或数次向培养物提供额外组分的培养细胞的方法。所提供的组分通常包含在培养过程期间已耗减的细胞的营养补充物。另外或可替代地,额外组分可包括辅助组分(例如,细胞周期抑制化合物)。进料分批培养通常在某一点处停止,并且将培养基中的细胞和/或组分收获并任选地纯化。术语“积分活细胞密度”或“ivcd”可互换使用,并且是指在培养的过程内活细胞的平均密度乘以培养进行的时间的量。“累积活细胞密度”(cvcd)是通过将两个时间点之间的平均活细胞密度(vcd)与那两个时间点之间的持续时间相乘来计算。cvcd是通过对vcd对时间绘图所形成的曲线下面积。本发明利用进行基因内互补以使用两载体系统直接选择编码异聚蛋白(诸如抗体)的表达构建体的选择性标记。“基因内互补”是指遗传物质(即,核酸)的节段之间的互补,所述节段各自在相同基因座内具有不同缺陷。由遗传物质编码的单独蛋白质是有缺陷的且非功能性的,但两种单独缺陷型蛋白质可缔合以形成活性蛋白。如果单独缺陷型蛋白质能够缔合并展现活性,则所述单独缺陷型蛋白质称为“互补”。对于由多个相同亚基(诸如gs)构成的选择性标记蛋白而言,基因内互补可当突变体亚基为互补时发生。“基因内互补”还可发生在由遗传物质所编码的单独蛋白质的片段之间,所述片段缔合以形成活性蛋白。如本文所述,高等真核生物中的成熟gs酶呈由两个五聚环组成的十聚复合物形式存在。本发明证明了,gs的基因内互补可成功地用作用于产生表达重组蛋白的细胞系的选择系统的一部分。所述系统在遗传修饰为谷氨酰胺合成酶缺失的cho细胞中重组抗体的高水平表达方面的实用性。通过在编码核酸中引入一个或多个突变来使由遗传物质所编码的单独蛋白质是有缺陷的且非功能性的,所述一个或多个突变改变蛋白质的结构足以使其为非功能性的。所述一个或多个突变可通过分子生物学领域中已知的任何方法引入。可替代地,可使用已使得是有缺陷的且非功能性的蛋白片段。在这种情况下,两个蛋白片段可通过基因内互补来缔合以形成活性蛋白。构建了编码多肽和选择性标记的突变体亚基的核酸分子,其以突变体亚基的表达与多肽的表达相关联的方式布置。因此,当将编码互补突变体亚基的核酸分子引入至细胞中并应用选择性条件时,所述互补突变体亚基各自的大致相等且高水平的表达将提供最高可选择活性。此外,可操作连接的多肽将以几乎相等且高的量表达,因此使表达相等且高水平所需多肽的细胞的选择优化。在本发明的一个实施方案中,选择性标记是谷氨酰胺合成酶(gs),其活性位点定位在gs的两个亚基的接点处,所以互补、功能上受损的突变体亚基的共表达应得到一些完整功能性活性位点。个别突变体亚基单独不具有显著的可选择活性,但当与其互补突变体亚基共表达时提供可选择活性。亚基的最佳活性可取决于其相互作用,且因而可通过相互作用结构域来促进。此类相互作用结构域可内源于亚基,或其可异源于亚基。可用于本发明中的额外选择性标记包括精氨基琥珀酸裂解酶、腺苷酸基琥珀酸合成酶和谷氨酸脱氢酶,可鉴别其互补突变体。例如,与精氨基琥珀酸裂解酶突变体的基因内互补可通过不存在精氨酸的情况下的生长来确定,而与腺苷酸基琥珀酸合成酶突变体的基因内互补可通过不存在腺苷的情况下的生长来确定。在一个非限制性实施方案中,本发明需要使用选择性标记的两个片段,所述片段各自可表达为相互作用结构域的融合蛋白。当表达时,相互作用结构域促进两个片段的缔合或二聚化,从而允许亚基起作用并提供可选择活性(例如,但不限于pelletier等人(1998),proc.natl.acad.sci.,95:12141-12146所描述者)。合适的选择性标记包括gs和上文所述的其他选择性标记,以及选择性标记,诸如赋予通常为毒性的具体药物的抗性的那些,和在规定条件下赋予细胞存活或诱导细胞死亡的代谢酶,如本文中所公开。“相互作用结构域”是包括但不限于能够促进两个或更多个同源或异源多肽的相互作用或缔合的多肽的结构域。在一个实施方案中,相互作用结构域是二聚化结构域。二聚化结构域可为能够诱导两个多肽的相互作用或缔合的多肽。存在两种类型的二聚体,能够形成同二聚体(与相同序列)或异二聚体(与另一序列)的那些。在一个说明性但非限制性实施方案中,相互作用结构域是亮氨酸拉链卷曲螺旋多肽。亮氨酸拉链通常包含约35个氨基酸,其含有特征的七残基重复,其中疏水性残基在重复的第一和第四残基处(harbury等人(1993),science262:1401)。因此,亮氨酸拉链适合于出于使多肽寡聚合的目的融合至多肽,因为其是小分子且与较大相互作用结构域相比较不可能破坏多肽正常功能。亮氨酸拉链的实例包括但不限于来自多肽的亮氨酸拉链结构域,诸如gcn4、c/ebp、c-fos、c-jun、c-myc和c-max。二聚化结构域的额外实例包括螺旋-环-螺旋结构域(murre等人(1989),cell58:537-544)。视黄酸受体、甲状腺激素受体、其他细胞核激素受体(kurokawa等人(1993),genesdev.7:1423-1435)和酵母转录因子gal4和hap1(marmonstein等人(1992),nature356:408-414;zhang等人(1993),proc.natl.acad.sci.usa90:2851-2855;美国专利号5,624,818)所有都具有带有此基序的二聚化结构域。在另一个实施方案中,相互作用结构域是四聚化结构域,其是能够结合三个其他四聚化结构域以形成四聚复合物的多肽。含有四聚化结构域的蛋白质的实例包括但不限于大肠杆菌乳糖阻遏物(氨基酸46-360;chakerian等人(1991),j.biol.chem.266:1371;alberti等人(1993),emboj.12:3227;和lewis等人(1996),nature271:1247),和在残基322-355处的p53四聚化结构域(clore等人(1994),science265:386;harbury等人(1993),science262:1401;美国专利好5,573,925)。在一个替代实施方案中,本发明需要使用选择性标记的三个亚基,所述亚基各自表达为相互作用结构域的融合蛋白,从而增强缔合以提供可选择活性。在此实施方案中,存在异聚复合物的三种组分。在一个或多个表达的载体中,其各自的编码序列可操作地连接至选择性标记的相应亚基各自的编码序列,例如表达单一重链和两条不同轻链的双特异性抗体,其中所述两条轻链均能够与所述重链缔合。本发明还涵盖使用已知的或有待公开的具有四个或甚至更多个亚基的选择性标记。本发明还可用于制备异多聚体(或异寡聚)蛋白,其含有三个或更多个亚基,诸如本文中先前所描述者。这包括双特异性抗体,和本领域中已知的其他异多聚体蛋白。例如,双特异性抗体可通过使用以下来表达:如本文所述的能够基因内互补的第一选择性标记(例如,gs),以表达来自第一抗体的重链和轻链;和不同的选择性标记诸如断裂代谢酶或抗性标记、或第二选择性标记(其不同于第一选择性标记且能够基因内互补),以表达第二抗体的重链和轻链,所述第二抗体能够与所述第一抗体缔合以形成双特异性抗体。或者,可使用如本文所述的能够基因内互补的第一选择性标记(例如,gs)表达双特异性抗体的两条不同重链,其中所述重链能够彼此缔合。然后可使用不同的选择性标记表达轻链。在一些情况下,双特异性抗体的两个臂的轻链可相同,其指明单一选择性标记(代谢酶或抗性标记、或选择性标记的其他形式)可用于表达轻链。可替代地,不同轻链可与双特异性抗体的每条不同重链缔合,在此情况下,使用不同的选择性标记诸如断裂代谢酶或抗性标记、或第二选择性标记(其不同于第一选择性标记且能够基因内互补)表达第一轻链和第二轻链。如以下将在实施例中显示,已发现,本发明的方法和组合物减少选择表达高水平异多聚体蛋白的所需细胞所必需的时间的量。因此,在另一个实施方案中,本发明涵盖选择表达高水平重组多肽(具体地说,异多聚体多肽)的细胞。另外,本发明在改进经由细胞培养过程产生异聚复合物的方面具有特别实用性。在一些实施方案中,编码突变体标记亚基的核酸同框融合至编码接头的核酸,其随后同框融合至编码相互作用结构域的核酸。接头可包括允许相互作用结构域相互作用并允许亚基起作用以提供可选择活性的任何相对短的、柔性序列。接头的实例在相关技术中是丰富的,并且可包含ggpgg、gpggg,其中在单一字母氨基酸代码中,g是甘氨酸且p是脯氨酸。在一个实施方案中,接头是例如curtis等人(1991;procnatlacadsci88(13):5809-5813)所描述的一系列甘氨酸和丝氨酸残基。在一个实施方案中,两个互补突变体亚基由两种载体表达,其中第一载体包含编码第一多肽的第一核酸,并且其中第一核酸可操作地连接至编码选择性标记的突变体亚基的第二核酸。第二载体包含编码多肽的第三核酸,所述多肽能够与第一核酸所编码的第一多肽缔合,其中所述第三核酸可操作地连接至编码选择性标记的互补突变体亚基的第四核酸。因此,两种载体同时引入到细胞群体中,并且应用基因内互补的选择。在另一个实施方案中,两个互补片段由两种载体表达,其中第一载体包含编码第一多肽的第一核酸,并且其中第一核酸可操作地连接至编码选择性标记的片段的第二核酸。第二载体包含编码多肽的第三核酸,所述多肽能够与第一核酸所编码的第一多肽缔合,其中所述第三核酸可操作地连接至编码选择性标记的互补片段的第四核酸。因此,两种载体同时引入到细胞群体中,并且应用基因内互补的选择。例如,表达抗体的两载体系统是通过制备包含编码抗体的一条链的第一核酸的第一载体来构建,其中所述第一核酸可操作地连接至编码mutgs-nt的第二核酸。第二载体包含编码抗体的另一条链的第三核酸,其中所述第三核酸可操作地连接至编码mutgs-ct的第四核酸。因此,在一个实施方案中,所述第一核酸编码免疫球蛋白重链,并且所述第三核酸编码免疫球蛋白轻链。或者,所述第一核酸编码免疫球蛋白轻链,并且所述第三核酸编码免疫球蛋白重链。在另一个实施方案中,两个互补片段由两种载体表达,其中第一载体包含编码第一多肽的第一核酸,并且其中第一核酸可操作地连接至编码选择性标记的片段的第二核酸。第二载体包含编码多肽的第三核酸,所述多肽能够与第一核酸所编码的第一多肽缔合,其中所述第三核酸可操作地连接至编码选择性标记的互补片段的第四核酸。因此,两种载体同时引入到细胞群体中,并且应用基因内互补的选择。在这种情况下,表达抗体的两载体系统是通过制备包含编码抗体的一条链的第一核酸的第一载体来构建,其中所述第一核酸可操作地连接至编码n-末端谷氨酰胺合成酶片段的第二核酸。第二载体包含编码抗体的另一条链的第三核酸,其中所述第三核酸可操作地连接至编码c-末端谷氨酰胺合成酶片段的第四核酸。因此,在一个实施方案中,所述第一核酸编码免疫球蛋白重链,并且所述第三核酸编码免疫球蛋白轻链。或者,所述第一核酸编码免疫球蛋白轻链,并且所述第三核酸编码免疫球蛋白重链。额外实施方案包括使用允许mutgs亚基中的一者或两者的表达的一个或多个ires。在本发明的上下文中,“ires”是指内部核糖体进入位点,其促进mrna从内部位点(即,不是mrna的5′端的位点)的翻译的起始。合适的ires的一个实例是脑心肌炎病毒(ecmv)的ires,如jang和wimmergenes&development41560(1990)以及jang,davies,kaufman和wimmerj.vir.631651(1989)中所描述。emcv的残基335-848形成合适的ires;ecmvires的其他变体或部分是已知的且将合适用于本发明中。ires的合适的部分或变体是将赋予第二开放阅读框(orf)的充分翻译者。另外,ires的3′端可改变(或突变)以降低翻译效率,从而提供增强选择和/或扩增方法的手段。例如,ires的效率可通过使用先前显示以允许有效选择和扩增的序列(aldrich等人,biotechnolprog19,1433;2003)来减小。替代序列是已知的,或可通过本领域技术人员确定。在另一个实施方案中,除选择性标记的亚基和异聚复合物的多肽之外,本发明还包含一种编码不同功能性选择性标记的核酸。出于本文中的目的,“不同功能性选择性标记”不是能够基因内互补的选择性标记的突变体亚基,而是具有不同可选择活性的蛋白质。熟知的标记诸如博来霉素(zeomycin)、新霉素、嘌呤霉素、杀稻瘟菌素s、或赋予霉酚酸的抗性的gpt等可用作不同功能性选择性标记。另外,还可使用能够基因内互补的不同选择性标记的互补突变体。在此实施方案中,本发明包含两种载体,其中所述载体各自包含:编码可形成异聚复合物的多肽的第一核酸,所述第一核酸可操作地连接至编码选择性标记的至少一个突变体亚基的第二核酸;以及编码不同功能性选择性标记的核酸。此外,由每种载体的第一核酸所编码的相应多肽可缔合以形成复合物,并且每种载体的第二核酸所编码的一个或多个亚基可缔合以提供可选择活性,且第三核酸所编码的多肽提供不同于第二核酸所编码的亚基的可选择活性的可选择活性。例如,第一载体可编码新霉素的抗性,并且第二载体可编码博来霉素的抗性,或仅一种载体可含有额外不同功能性选择性标记。因此,将一种载体转染到细胞系中,并应用选择(即,将药物g418添加至抗新霉素细胞)。在选择之后,可使用常规方法确定是否存在载体,和载体上的核酸所编码的多肽的表达水平,例如通过pcr、dna印迹、elisa、蛋白质印迹等。一旦已获得高水平表达,就将第二载体转染到细胞系中。在维持第一载体的选择的同时,应用第二选择性标记的选择(即,博来霉素抗性),且评估第二载体的存在和相应载体编码的蛋白质的表达。在此实施方案中,一旦已确定存在两种载体,就应用如本文中所述的已缔合于细胞中以提供可选择活性的亚基的表达的选择。在一个替代实施方案中,将本发明的编码独立的可选择活性的两种核酸同时转染,并同时应用选择。一旦已确定存在两种载体,就应用已缔合于细胞中以提供可选择活性的亚基的表达的选择,如上文所述。在另一个实施方案中,将编码独立的可选择活性的本发明载体各自转染到分开的细胞系中。一旦应用选择,并且已鉴别出表达高水平的由每个所需载体所编码的蛋白质的克隆,就将细胞如hori等人(美国专利号5,916,771)中所述来融合。一旦融合完成,就应用亚基所提供的可选择活性的选择。在另一个实施方案中,将任选地不含有独立的可选择活性的本发明核酸用第三载体同时转染。第三载体编码分开的可选择活性诸如例如新霉素抗性或β半乳糖苷酶,从而可允许成功转染的细胞的初步选择。一旦已执行这种初步选择,可应用突变体亚基的可选择活性的选择。在此实施方案中,转染相等量的两个表达载体,同时以前两种载体的三分之一浓度转染第三载体(例如,3∶3∶1或6∶6∶1的比率,或类似比率)。本领域技术人员应认识到,所述比率的变化在本发明的范围内。编码所需异聚复合物的组分的核酸可通过本领域中已知的方法呈cdna或呈基因体dna获得。例如,编码所需组分的信使rna可采用rna分离的标准技术从合适的来源分离出,并使用寡-dt纤维素色谱法以隔离聚-amrna。当待表达的异聚复合物是抗体时,所需核酸的合适的来源可从成熟b细胞或杂交瘤培养物分离出。此外,用于本发明中的核酸可通过化学合成来获得。除编码异聚复合物的所需组分的核酸之外,载体构建体可包括促进原核细胞和/或真核细胞中的复制、将构建体整合至真核染色体中的额外组分,和有助于选择和/或筛选含有所述构建体的细胞的标记。本发明的载体是重组dna载体,其包括但不限于质粒、噬菌体、噬菌粒、粘粒、病毒、逆转录病毒等,所述载体将所需核酸插入到细胞中。当核酸放置成与另一核酸呈功能性关系时,其是“可操作地连接”的。更具体地说,可操作地连接是指编码不同多肽的两个不同核酸使转录被同时诱导。可操作地连接还意图是指,连接的核酸在单一转录单元中可为连续的,同时从一个或多个核糖体开始位点(例如,内部核糖体进入位点)引导翻译。编码两个不同多肽的可操作地连接的转录单元称为“双顺反子转录物”。本发明的方法还可结合已知的或有待发现的诱导重组蛋白的产生的方法使用。“诱导条件”是指增加所需重组蛋白的每细胞相对产量的技术。此类技术包括冷温度变换和添加化学品和任何已知的或有待发现的技术的组合,仅举几个例子,以及任何有待描述和/或发现的诱导技术。通常,诱导高密度的细胞的分批培养物或灌注培养物以产生重组蛋白。常常,其他细胞过程(诸如生长和分化)被抑制,以便将大部分细胞能量引导至重组蛋白产生中。具有亚基的任何选择性标记均可用于本发明的方法和组合物中。如本文所用,术语“亚基”当指代选择性标记时是指选择性标记的一部分。此外,选择性标记的第一突变体亚基(本文中称为“初始突变体亚基”)可与相同可选择标记的第二互补突变体亚基(本文中称为“互补突变体亚基”)一起表达,以提供单独任一突变体亚基中不存在的可选择活性水平。亚基还可指具有由另一突变的多肽互补的突变的多肽,其还为选择性标记的不同亚基。赋予对动物细胞通常为毒性的具体药物的抗性的选择性标记可用于本发明的方法和组合物中。例如,以下是抗性选择性标记的非限制性实例:博来霉素(zeo);嘌呤霉素(pac);杀稻瘟菌素s(blas)、gpt(其赋予霉酚酸的抗性)(mulligan和berg(1981),proc.natl.acad.sci.usa78:2072);新霉素抗性基因,其赋予氨基糖苷g-418的抗性(colberre-garapin等人(1981),j.mol.biol.150:1);和hygro,其赋予潮霉素的抗性(santerre等人(1984),gene30:147)。在规定条件下赋予细胞存活或诱导细胞死亡的代谢酶还可用于本发明的方法和组合物中。实例包括但不限于:二氢叶酸还原酶(dhfr);单纯疱疹病毒胸苷激酶(tk)(wigler等人(1977),cell11:223)、次黄嘌呤-鸟嘌呤磷酸核糖基转移酶(hgprt)(szybalska和szybalski(1962),proc.natl.acad.sci.usa48:2026)以及腺嘌呤磷酸核糖基转移酶(aprt)(lowy等人(1980),cell22:817),其是可分别用于缺乏tk、hgprt或aprt的细胞中的基因。在一个实施方案中,谷氨酰胺合成酶(gs)是本发明的方法和组合物中所使用的选择性标记。gs包含相同亚基的两个堆叠的五聚环,其中十个活性位点是由一个亚基的n-末端残基和相邻亚基的c-末端残基形成。制备n-末端(mutgs-nt)和c-末端(mutgs-ct)突变体构建体并评估互补。mutgs-nt和mutgs-ct两者均不允许缺乏谷氨酰胺的培养基中的细胞生长,然而,当将gs突变体细胞用两个构建体转染时,以类似水平用两个构建体转染的那些细胞在缺乏谷氨酰胺的培养基中存活。当发生以两个此类突变体共转染时,所述突变体称为“互补”。或者,可使用表达内源gs的细胞,并且转染子可通过赋予增加的甲硫氨酸磺酰亚胺(msx)的毒性水平的抗性来选择。基于颜色选择的选择性标记还可用于本发明的方法和组合物中。在一个具体实例中,可使用β-半乳糖苷酶(blau等人,wo98/44350)。荧光标记还可用于本发明的方法中,例如,gfp已用于细胞的克隆选择以测量蛋白质-片段互补测定中的蛋白质相互作用(remy和michnick(1999),proc.natl.acad.sci.,96:5394-5399)。类似地,荧光素缀合的甲氨蝶呤可用以检测表达互补dhfr片段的细胞(remy和michnick(2001),proc.natl.acad.sci.,98:7678-83)。荧光标记的优点在于,这种选择可在任何动物细胞类型中进行,并且不限于在代谢路径中具有缺乏的那些动物细胞类型,或不需要例如新霉素的药物敏感性。如本文中所使用的术语“多肽”包括天然存在的蛋白质或仅重组表达的蛋白质(包括翻译前和翻译后加工)或其片段,它们通常保留二级结构。蛋白质是具有高分子量(从小蛋白[具有50-100个氨基酸]的约10,000至某些形式的大于1,000,000)的大分子;其由不同量的相同的20种氨基酸组成,完整蛋白质中的所述氨基酸通过称为肽键的共价化学键联合并。连接在一起的氨基酸形成称为多肽链的线性未分支聚合结构;此类链可含有数百个氨基酸残基;这些氨基酸残基针对给定种类的蛋白质以特定次序布置。术语“肽”包括多肽或蛋白质的短片段,其长度通常小于20个氨基酸。术语“细胞培养”是指包括细胞在多细胞生物体或组织外的生长和繁殖。通常,细胞培养是在无菌、受控的温度和大气条件下在组织培养板(例如,10-cm板、96孔板等等)中执行,或其他粘附培养(例如,在微载体珠粒上)或以悬浮培养的形式(诸如在滚瓶中)。培养物可生长于摇瓶、小规模生物反应器和/或大规模生物反应器中。生物反应器是用以培养细胞的装置,在所述装置中可监测并调整环境条件诸如温度、气氛、搅拌和/或ph。许多公司(例如,abs公司,wilmington,del.;celltrends公司,middletown,md.)以及大学和/或政府资助组织(例如,细胞培养中心,minneapolis,minn.)提供基于合同的细胞培养服务。培养物与选择可选择活性的试剂接触的最佳时期长于一个正常生长周期的典型时期(例如,对于中国仓鼠卵巢细胞(cho细胞),其中一个生长周期据报道是大约20-22小时(rasmussen等人(1998),cytotechnology,28:31-42))。因此,在一个实施方案中,培养物包含可选择条件,例如,药物、代谢物或彩色底物,优选持续至少约一天、更优选持续至少约3天、且甚至更优选持续至少约7天。适合于在培养物中生长的多种动物细胞系可购自例如美国典型培养物保藏中心(atcc,manassas,va.)和nrrl(peoria,ill.)。通常用于工业或学院实验室中的更多已建立细胞系中的一些包括cho、vero、bhk、hela、cos、cv1、mdck、293、3t3、pc12、骨髓瘤(mycloma)(例如,nso)、以及wi38细胞系,仅举几个例子。在其他实施方案中,非动物细胞系可用于本发明的方法中,例如,植物细胞系、昆虫细胞系(例如,sf9)、酵母细胞或细菌细胞诸如大肠杆菌。在一个实施方案中,使用gs-缺陷型cho细胞诸如gs-/-zfn-修饰的cho细胞(sigmaaldrichfinechemicals,st.louismo)。gs-缺陷型cho细胞(或其他哺乳动物细胞)还可使用本领域中已知的方法制备。另外,cho细胞易于呈粘附培养物或悬浮培养物操纵,且展现相对良好的遗传稳定性。此外,可使用本领域技术人员熟知的技术(例如,通过转化、病毒感染和/或选择)建立新的动物细胞系。如上文所指出,多种宿主-表达载体系统可用于表达本发明的异聚复合物。当异聚复合物可溶时,肽或多肽可自培养物回收,即在不分泌异聚复合物的情况下从宿主细胞回收,且在细胞分泌异聚复合物的情况下从培养基回收。然而,表达系统还涵盖表达锚定于细胞膜中的异聚复合物的工程化的宿主细胞。纯化或富集来自此类表达系统的异聚复合物可使用适当清洁剂和脂质胶束以及本领域技术人员熟知的方法来实现。然而,此类工程化的宿主细胞本身可用于不仅保留异聚复合物的结构和功能特性是重要的、而且例如在药物筛选测定中评估生物活性也是重要的情况中。通过本发明的方法所表达的蛋白质可被收集。此外,所述蛋白质可使用已知方法从此类培养物或组分(例如,从培养基或细胞萃取物或体液)纯化或部分纯化。短语“部分纯化的”是指,已进行了某一或某些分级分离程序,但存在多于所需蛋白质的多肽种类(至少10%)。“纯化的”是指,蛋白质是基本上均质的,即,存在小于1%的污染蛋白。分级分离程序可包括但不限于以下的一个或多个步骤:过滤、离心、沉淀、相分离、亲和纯化、凝胶过滤、离子交换色谱法、尺寸排阻色谱法(sec)、疏水性相互作用色谱法(hic;使用如苯醚、丁醚或丙醚的树脂)、hplc或上述的一些组合。本发明还任选地涵盖进一步配制所述蛋白。术语“配制”是指所述蛋白质可为最终使用者经缓冲液交换、杀菌、松散填充和/或填充。出于本发明的目的,术语“无菌松散形式”是指,制剂不含或基本上不含微生物污染(达到诸如食品和/或药物目的可接受的程度),并且具有限定的组成和浓度。术语“无菌单位剂量形式”是指适合于顾客和/或患者施用或消耗的形式。此类组合物可包含有效量的蛋白质,与其他组分诸如生理学上可接受的稀释剂、载体或赋形剂的组合。术语“生理学上可接受”是指不会干扰一种或多种活性成分的生物活性的有效性的非毒性物质。针对本发明,开发了多组编码哺乳动物gs蛋白的n-末端改变(mutgs-nt)的不活化突变,以及多组编码所述蛋白的c-末端改变(mutgs-ct)的不活化突变。使gs催化位点进行丙氨酸扫描,在所述位点中突变单独地且组合地发生于涉及结合谷氨酸盐、atp和/或氨的所有进化上保守的丙氨酸残基。将大多数残基基于来自公开的gs酶的x射线结晶学和生物化学研究来选择,但将活性位点中在的底物/配体/金属内的所有残基评估为潜在的候选者。鉴别n-末端和c末端催化受损但结构上完整的gs突变体的组合,并评估它们在共表达时彼此互补以恢复一些gs活性的能力。通过将抗体的一条链的表达与mutgs-nt或n-末端gs片段连接并将另一条链的表达与mutgs-ct或c-末端gs片段连接来构建两载体系统。构建一种表达载体,其具有驱动抗体重链(hc)在单一mrna上的表达的启动子/增强子和允许mutgs-nt或n-末端gs片段的表达的ires。分开地,通过构建具有驱动lc的表达的启动子/增强子和允许mutgs-ct或c-末端gs片段的表达的ires的载体,将抗体的轻链(lc)的表达连接至mutgs-ct或c端gs片段。mutgs-nt和mutgs-ct、n-末端或c-末端gs片段自身均不允许缺乏谷氨酰胺的培养基中的细胞生长。然而,当以两种构建体转染gs突变体细胞时,那些以类似水平用两种构建体转染的细胞允许缺乏谷氨酰胺的培养基中的存活。那些细胞还产生类似量的目标抗体的lc和hc。可通过降低ires的效率,导致mutgs选择性标记的翻译减少,或可能通过添加gs抑制剂l-甲硫氨酸磺酰亚胺(msx)来进一步增加选择的严格性。本发明的范围不限于本文中所述的意图作为本发明的单独方面的单一说明的特定实施方案,且在功能上等效的方法和组分均在本发明的范围内。事实上,除本文中所示和所述的修改之外,本领域技术人员根据前述描述和附图将明白对本发明的各种修改。此类修改意图属于所附权利要求书的范围。实施例实施例1此实施例描述鉴别为可能可用于基因内互补的谷氨酰胺合成酶(gs)突变体的选择和制备。选择4个鼠类gs的n-末端残基(w60、n61、d63和s66)和11个c-末端残基(e134、e136、e196、e203、n248、h253、n255、r319、r324、e338和r340)用于丙氨酸突变扫描中的评估。单独和组合突变体构建体产生于pcdna3.3载体(新霉素抗性,invitrogengrandisland,ny)中,得到总计9个n-末端突变体构建体和16个c-末端突变体构建体。针对cho-k1细胞系中完整gs蛋白的表达来评估突变体构建体。在36℃、5%co2、70%相对湿度下,于排气型摇瓶中的含有谷氨酰胺的非选择性化学性限定生长培养基中培养细胞,并于加湿孵育箱中以150-160rpm振荡。以vicell自动细胞计数器(beckman-coulter公司,brea,ca)测量活细胞密度(vcd)和存活力。通过每3-4天稀释来使培养物传代。通过电穿孔将每种条件下一百万个细胞以10微克线性化dna转染(生物学一式三份)。根据制造商的建议,在37℃下,将dna以pvui-hf限制酶(newenglandbiolabs,ipswich,ma)线性化过夜。使用琼脂糖凝胶电泳确定各质粒的完全线性化。在限制消化之后,在-70℃下,通过向各消化物中添加10%反应体积的3m乙酸钠和200%反应体积的100%乙醇来将dna沉淀过夜。在乙醇沉淀之后,dna集结成粒,将球粒以70%乙醇洗涤一次,风干,且再悬浮于hepes缓冲盐水中。在nanodrop2000(thermoscientific,wilmington,de)上测量dna浓度,且以hepes缓冲盐水调整至1微克/微升。对于基因内互补测试,使用各自5毫克的两种质粒以及10微克剪切的鲑精dna(invitrogen)。通过以下建立转染:将来自第3天生长培养物的每种条件下一百万个细胞旋转沉降(spinningdown);将细胞再悬浮于150微升hepes缓冲盐水中;添加总计10微克的质粒dna(各质粒5微克)和10微克;且将混合物转移至96孔电穿孔板(bio-radlaboratories公司,hercules,ca)的孔中。电穿孔是使用genepulsermxcelltm(bio-radlaboratories公司,hercules,ca)于96孔板中执行,其中设置如下:指数波,电压=290v,电容=950microf,电阻=950欧姆。在电穿孔之后,将细胞转移至排气型24深孔板(24dwp)中的2ml/孔预热的非选择性生长培养基中,且在加湿kühner孵育箱(kühnerag,basel,switzerland)中以220rpm振荡。在转染后三天,将细胞计数,且以1x106个活细胞/ml转移至选择性培养基(缺乏谷氨酰胺的化学性限定生长培养基)中。通过每3-4天使细胞在选择性培养基中连续传代直至所恢复的培养物存活力高于90%来产生稳定池。于试剂中孵育细胞(将培养物1∶10稀释至工作浓度(1∶50pbs稀释液)的viacountflex试剂(emdmillipore,billerica,ma)中)15分钟之后,在guavaeasycytetmht流式细胞仪(emdmillipore,billerica,ma)上使用viacount测定模块执行细胞计数。使所有池恢复至至少90%存活力,之后在功能测定中测试。在池产生之后,执行一式三份池的蛋白质印迹分析,以确定各构建体的gs蛋白过表达的水平并确定是否有任何突变干扰了完整gs蛋白的产生。对于蛋白质印迹法分析,将3x106个细胞于冰上溶解于补充有无edtahalt蛋白酶和磷酸酶抑制剂混合物(thermoscientific,rockford,il)的100微升ripa缓冲液(thermoscientificpierce,rockford,il)中持续30分钟。通过以13,000rpm离心清除溶解产物,添加4x负载缓冲液(invitrogen,grandisland,ny),且将样品在95℃下煮沸5分钟。将15μl各样品在e-page48凝胶(invitrogen,grandisland,ny)上运行,且使用iblot(invitrogen,grandisland,ny)转移至pvdf膜。在4℃下以1∶500小鼠抗谷氨酰胺合成酶抗体(cat#610518,bdbiosciences,sanjose,ca)探测各膜过夜,接着以1∶2,000抗小鼠alexafluor680第二抗体(lifetechnologies,grandisland,ny)孵育。在odyssey红外成像系统(li-cor,lincoln,ne)上扫描印迹,且使用imagej软件定量。表1:所测试的鼠类谷氨酰胺合成酶突变体尽管初始分析指明氨基酸残基e388和r340为突变的候选残基,但是e388a和r340ags突变体的过表达(呈单一突变或结合其他突变)并不产生完整gs蛋白。因此,不再考虑含有这些突变的构建体。因为稳定的突变体gs池产生于野生型cho-k1亲代细胞系中,所述细胞系具有功能性gs酶,所以在缺乏谷氨酰胺且补充有25micromgs抑制剂l-甲硫氨酸磺酰亚胺(msx)的选择性培养基中以生长/存活力筛选执行突变体活性评估。野生型cho-k1细胞在选择性培养基中4次传代之后死亡。某些n-末端和c-末端突变阻止选择性培养基中的培养物生长,但是即使生长受损的池也保留了相对高的存活力。用单独gsn-末端突变体转染的细胞的生长类似于野生型构建体,所以不进一步测试这些构建体。然而,两种n-末端多重突变体构建体w60an61ad63a(mutgs-nt1;seqidno:6)和w60an61ad63as66a(mutgs-nt2;seqidno:10)产生高水平的过表达的gs蛋白且在功能测定中表现得最接近于空载体阴性对照,并且选择它们用于进一步分析。此外,两种其他n-末端突变体构建体w60an61ad63as66ad76a(mutgs-nt3;seqidno:25)和w60an61ad63rs66a(mutgs-nt4;seqidno:19)是为了尽力增加互补作用的严格性而制备,并且对它们进行评估。在后一构建体中,天冬氨酸-63突变成精氨酸而非丙氨酸,因为鱼腥藻(anabenaazollae)中的类似突变体完全消除生物合成活性且表达与野生型相同,然而丙氨酸的突变保留6.5%的生物合成活性(eur.j.biochem.266,1202(1999)crespo等人)。虽然若干单一c-末端突变体构建体通过了功能评估,但是选择含有多重突变的两种构建体e134ae136ae196ae203a(mutgs-ct1;seqidno:21)和n248ah253an255a(mutgs-ct2;seqidno:22)用于进一步分析。实施例2此实施例描述gs中的突变对重组异二聚蛋白(即,单克隆抗体(mab))的表达的作用。将实施例1中所述的选择的n-末端和c-末端突变体克隆至mab表达载体中以构建两载体系统的多重组合,其中轻链(lc)的表达经由ires连接至mutgs-ct,且重链(hc)连接至mutgs-nt。根据制造商的说明书(sigma-aldrichst.louis,mo),使用knockoutzfnkit-chogs构建cho-k1gs敲除细胞系。将cho-k1谷氨酰胺合成酶敲除(cho-k1gs-ko)细胞系如先前所述培养。使用含有连接至野生型gs(阳性对照)或选择的n-末端和c-末端突变体的mablc或hc的载体执行一式三份的cho-k1gs敲除转染,并且在缺乏谷氨酰胺的培养基中培养。将含有野生型gs的初始pcdna3.3载体用作额外阳性对照。通过在无谷氨酰胺的培养基中传代20天,pcdna3.3野生型gs池和mab连接的野生型gs池恢复高于90%的存活力。与此相反,在无谷氨酰胺的培养基中17天之后,模拟转染的池和所有mab连接的突变体gs池死亡,从而确认选择的突变体产生功能上受损的gs蛋白产物。最后,为了在cho-k1gs敲除细胞系中产生产mab的池,针对在两载体表达系统内的gs基因内互补评估经选择的n-末端和c-末端突变体。将这些细胞以在gs序列中具有指示的突变的基因内互补载体a(lc:mutgs-ct)和b(hc:mutgs-nt)转染(示出于以下表2中)。所有经评估的mutgs-ct/mutgs-nt组合均能够在无谷氨酰胺的培养基中25天内恢复至大致90%的存活力。两载体lc:mutgs-ct/hc:mutgs-nt池的存活力下降较大,且比含有野生型gs的单一载体阳性对照需要更长时间在选择性培养基中恢复。以10天进料分批产生测定针对mab表达进一步评估所产生的一式三份的稳定的池。产生测定是由以1∶5分开的第4天生长培养物通过稀释至24dwp中的化学性限定产生培养基中(约0.5x106个活细胞/ml)建立。使用guavaviacount测定(millipore,billerica,ma),在第3天、第6天、第8天、和第10天检测培养物生长和存活力。以化学性限定进料培养基在第3天、第6天和第8天执行推注进料。使用比色polychem(polymedco,cortlandtmanor,ny)试剂在进料当天测量培养基葡萄糖浓度,且以50%葡萄糖储备溶液调整至12g/l。在第10天通过使用porosa/20蛋白a柱的亲和高效液相色谱法(hplc)确定培养物上清液的产生结束效价。以含有经由ires连接至单独gs突变体的单独mab链的gs基因内互补两载体系统所产生的所有池均产生可检测水平的抗体;值示出于以下表2中(实施例3)。实施例3此实施例描述通过减小iresgs接点效率来增加选择的严格性从而增加转染的池中的抗体效价的方法。初始构建体中的序列是直到atg-12的野生型emcv序列,atg-12融合至gs的起始密码子(davies和kaufman,jvirol66,1924;1992)。ires的效率通过使用先前显示以允许有效选择和扩增的序列(aldrich等人,biotechnolprog19,1433;2003)来减小。将更有效的(ed3)接合中接点的dna序列与下文不太有效的接合(317)的dna序列比较。ed3序列的下的数字指示wtemcv中所见的atg。最后一个atg表示gs的起始密码子。表达载体与图1中所示的那些相同,不是与ires和gs的接点处的序列。以具有不太有效的ires-gs接点的两个质粒转染细胞得到转染后较低的存活力和较长的恢复时间。三种ct1-nt4转染中仅两种ct1-nt4转染恢复,且其他mutires组合中没有恢复。然而,对于恢复的那些mutires组合,10天进料分批培养得到0.55g/l(一个池)和0.17g/l(另一池)。这些池的比生产率分别为16p/c/d和6.9p/c/d。0.55g/l效价超过先前所报道的效价,且不需要实现那些效价的facs分选(ye,j.等人biotechnolprog26,1431;2010)。表2:具有不同ires-gs接点的基因内gs转染子的表达。标记ct1-nt1-4和ct2-nt1-4的所有样品均使用ped3接点(seqidno:24)。标记mutires的样品具有seqidno:23中所示的不太有效的ires。实施例4此实施例描述增加这些培养物的选择性的另一策略。转染是如实施例2中所描述执行,但在选择期间,将gs抑制剂msx添加至培养物中以选择较高水平的gs表达。具有更有效的(即,非突变体)ires的三种ct2-nt4培养物中有一种恢复,而ct1-nt4或具有突变体ires的转染子均没有恢复。对于恢复的单一培养物,10天进料分批培养得到0.29g/l,其中比生产率为9.18p/c/d(参见上文表2)。实施例5此实施例描述大量mutiresct1-nt4转染的结果。这些构建体具有seqidno:23中所示的不太有效的ires。使用含有mutiresct1-nt4的载体执行总计88种cho-k1gs敲除转染。88种转染中总计10种在选择中存活,且在大约25天的培养之后得到茁壮生长的转染池,且行为类似于实施例2中的较少量转染。在无谷氨酰胺培养基中17天之后,模拟转染的池gs池死亡。以10天进料分批产生测定针对mab表达进一步评估一式两份的稳定的池。产生测定是由第4天生长培养物通过以0.6x106个活细胞/ml接种至24dwp中的化学性限定产生培养基中建立。使用guavaviacount测定(millipore,billerica,ma),在第3天、第6天、第8天和第10天检测培养物生长和存活力。以化学性限定进料培养基在第3天、第6天和第8天执行推注进料。使用比色polychem(polymedco,cortlandtmanor,ny)试剂在进料当天测量培养基葡萄糖浓度,且以50%葡萄糖储备溶液调整至12g/l。在第10天通过使用porosa/20蛋白a柱的亲和高效液相色谱法(hplc)确定培养物上清液的产生结束效价。用含有经由ires连接至单独gs突变体的单独mab链的gs基因内互补两载体系统所产生的所有池均产生相对高水平的抗体,其中两种池平均超过1g/l;值示出于表3中。表3:具有mutires-gs接点的基因内ct1-nt4gs转染子的表达。实施例6此实施例描述谷氨酰胺合成酶的分子互补方法。将谷氨酰胺合成酶表达为两个独立的片段,并且通过两个片段的遗传互补而完全重构酶活性。产生一系列构建体,所述构建体在各种残基处断裂谷氨酰胺合成酶。这些点是基于来自酶的分子结构的信息。将这些构建体在基于细胞的测定中测试其拯救缺乏谷氨酰胺合成酶活性的细胞系的能力。根据此测试,鉴别出若干片段,所述片段能够充分拯救谷氨酰胺合成酶缺乏细胞,并且因此证明,这些片段可缔合以产生完全功能性谷氨酰胺合成酶。许多先前关于蛋白片段互补的工作已使用呈功能性单体存在且重新组装仅需要设计的片段的蛋白质。此工作证明了使用分子互补以再现gs单体的活性和十聚功能性酶复合物的随后形成的能力。通过使用分子模型化方法鉴别互补蛋白片段,以寻找gs单体中可允许将蛋白断裂的区。还将gcn4亮氨酸拉链(lz)和柔性接头添加至分开的片段以帮助重新组装。所测试的片段示出于表4中,并且这些研究中所使用的鼠类gs蛋白的序列示出于表5中。将gs片段克隆至哺乳动物表达载体中,且转染至缺乏gs活性的cho细胞中。这些细胞通常可仅存活于补充有谷氨酰胺的细胞培养基中。通过将谷氨酰胺从培养基移除来测试这些构建体拯救这些细胞的能力。表4:gs分子互补片段设计:n末端lz=msdrmkqledkveellskvyhlenevarlkklvgerggggsgggggsc末端lz=ggggsgggggssdrmkqledkveellskvyhlenevarlkklvger-stop表5:这些研究中所使用的小鼠gs酶的序列和突出显示的断裂点y104e110s125n126e264如图2中所示,构建体1、2、3和5能够成功拯救谷氨酰胺合成酶缺乏细胞。这证明,这些片段能够在体内形成完全功能性谷氨酰胺合成酶。拯救结果的概述示出于表6中。作为对照,评估单独a或b片段中的任一者是否能够拯救谷氨酰胺合成酶缺乏细胞系。如图4中所示,单独a或b片段均不能拯救谷氨酰胺合成酶缺乏细胞系,从而确认使谷氨酰胺合成酶活性恢复需要这两种片段。表6:表4中所描述的gs片段的拯救动力学。构建体cho-k1gsko的拯救达到>90%存活力的天数wt4/4141.2/4约302.4/4183.4/4约244.0/4无恢复5.4/4约24接着在含有抗体轻链和重链基因的表达载体的情况下测试这些细胞互补构建体,以评估其在细胞系发展选择系统中的实用性。将两种表达载体改造成含有如表4中所示的互补谷氨酰胺合成酶片段,一个含有抗体轻链基因,并且第二个含有抗体重链基因(图4)。还将ires序列替换成通过框内atg的突变所减毒的型式(biotechniques20:102-1101996年1月)。这将相较于抗体基因进一步降低表达的谷氨酰胺合成酶片段的水平,并且增加系统的选择严格性,以实现偏向于在高转录活性位点处基因组整合的选择性压力水平。所述研究的结果示出于图6中。构建体1、2、3和5在此测定中进行测试,并且以两种定向将重链和轻链基因连接至谷氨酰胺合成酶a和b片段的中的每一个。来自构建体2的含有谷氨酰胺合成酶片段的载体能够在无谷氨酰胺培养基中的选择中存活。在10d进料分批生产率测定中评估来自构建体2选择池的细胞的生产率,并与使用表达全长gs酶的载体选择的对照池进行比较。如图6中所示,来自构建体2的细胞展示高生产率。当前第1页12
相关技术
网友询问留言
已有0条留言
- 还没有人留言评论。精彩留言会获得点赞!
1