一种促进番茄叶绿素合成及光合效率的基因及应用的制作方法
本发明涉及基因工程、分子生物学及生理学等领域,具体是涉及一种促进番茄叶绿素合成及光合效率的基因及应用。
背景技术:
番茄是茄科茄属中的一年生或多年生草本植物,是世界上栽培最广泛、消费量最大的蔬菜作物之一。番茄品种类型多样、营养丰富(如番茄红素可预防癌症),深受消费者喜爱,所以提高番茄的产量和品质至关重要。番茄起源于亚热带地区,是喜光喜温植物,而设施生产中,低温弱光现象频繁发生,严重限制番茄等园艺作物的产量。因此,研究光对番茄叶绿素合成及光合作用的影响机制,挖掘调控番茄叶绿素合成和提高光合效率的关键基因,对提高番茄的产量有着重要的应用价值。
叶绿素作为植物进行光合作用的重要色素,影响着番茄等园艺作物的产量。光是植物叶绿素生物合成必不可少的环境因素,它不仅为植物提供光合作用的能量,而且还作为一种重要的信号参与植物叶绿素生物的合成。在光信号转导中,亮氨酸拉链(bzip)类的转录因子hy5(longhypocotyl5)位于许多光受体的下游,将光信号传递给下游作用元件,对植物的光形态建成起着重要的作用。研究发现,hy5编码一个由168个氨基酸构成的分子量为18.5kda的蛋白质,它是c末端具有bzip结构域的转录因子,可以直接与相关基因启动子中的acgt的顺式作用元件结合,激活启动子并诱导基因表达,进而调控植物的生长发育及逆境抗性。有研究报道,hy5可以提高植物中次生代谢物质如类黄酮和花青素的含量,说明光信号对植物代谢物质的含量有着重要的影响作用,然而slhy5转录因子对番茄叶绿素合成及光合效率的影响作用及调控机制尚未见报道。
参考文献:
(1)fillattijj,kiserj,roser等,1987,efficienttransferofaglyphosatetolerancegeneintotomatousingabinaryagrobacteriumtumefaciensvector,bio-technol5:726-730。
(2)lixj,chenxj,guox,yinll等,2016,dwarfoverexpressioninducesalterationinphytohormonehomeostasis,development,architectureandcarotenoidaccumulationintomato,plantbiotechnolj,14:1021-1033。
(3)lixj,guox,zhouyh等,2016,overexpressionofabrassinosteroidbiosyntheticgenedwarfenhancesphotosyntheticcapacitythroughactivationofcalvincycleenzymesintomato,bmcplantbiol,16:33。
(4)wangy,caisy,yinll等,2015,tomatohsfa1aplaysacriticalroleinplantdroughttolerancebyactivatingatggenesandinducingautophagy,autophagy,11:2033-2047。
(5)zhangdw,yuans,xuf等,2016,lightintensityaffectschlorophyllsynthesisduringgreeningprocessbymetabolitesignalfrommitochondrialalternativeoxidaseinarabidopsis,plantcellenviron,39:12-25。
(6)zhanggx,liuyf,niy等,2014,exogenouscalciumalleviateslownighttemperaturestressonthephotosyntheticapparatusoftomatoleaves,plosone,9:e97322。
技术实现要素:
为了弥补上述现有技术的不足,本发明的目的是提出一种促进番茄叶绿素合成及光合效率的基因及应用,通过对slhy5基因的克隆、转基因技术培育番茄slhy5过表达植株,明确了其在提高番茄叶绿素含量及光合效率中的作用,为培育高叶绿素含量番茄新品种提供了基因资源,具有良好的应用价值,为研究光信号对植物叶绿素合成及光合作用的调控机制奠定理论基础。
本发明的目的是通过以下技术方案实现的:
本发明提供了一种番茄slhy5基因,该基因具有seqidno:1所示的核苷酸序列。
本发明还提供所述基因番茄slhy5基因编码的蛋白质,该蛋白质具有seqidno:2所示的蛋白质序列。
本发明提供了番茄slhy5基因或基因番茄slhy5基因编码的蛋白质在提高番茄植物叶绿素合成和促进番茄植物叶片光合作用中的应用。
本发明还提供一种获得植物叶绿素和光合速率提高的转基因番茄植株的方法,具体方法为:通过基因过表达技术使得所述番茄slhy5基因的表达水平上升,从而提高番茄植物的叶绿素;
所述基因过表达技术的具体内容为:提取番茄叶片总rna,反转录获得cdna,以cdna为模板,slhy5-oe-f和slhy5-oe-r为引物,扩增slhy5基因,将扩增产物构建到植物过表达载体上,然后用获得的重组表达载体导入宿主细胞中,接着用转化的农杆菌侵染番茄子叶,进行组织培养,筛选阳性转基因植株,获得叶绿素含量提高的转基因番茄;其中,所述引物slhy5-oe-f和slhy5-oe-r的核苷酸序列如seqidno:3和4所示。
进一步的,所述植物表达载体为具有35s启动子的表达载体pfgc1008-ha,所述表达载体pfgc1008-ha具体为:在过表达载体pfgc1008中加入了ha标签蛋白,所述ha标签蛋白为植物可选择性标记,以便于对转基因番茄植株进行鉴定及筛选。
进一步的,过表达载体质粒为pfgc1008::slhy5-ha。
进一步的,所述的宿主细胞为大肠杆菌细胞或农杆菌细胞,优选所述农杆菌为eha105。
本发明的有益效果如下:
(1)通过转基因手段构建番茄slhy5过表达植株来研究其对番茄叶绿素合成及光合效率的调控机制,结果发现,番茄slhy5过表达可以提高植物的叶绿素含量及光合效率。同时,番茄slhy5过表达植株中赤霉素(ga)含量降低,slpro基因(即番茄中的ga负调控因子)明显上调。另外,与野生番茄(wt)比,番茄pro突变体中叶绿素含量及光合效率明显降低。因此,番茄slhy5基因通过诱导体内ga负调控因子slpro基因的表达,从而促进植物的叶绿素合成,并提高植物的光合效率。
(2)本发明提供的slhy5基因为培育高叶绿素含量番茄新品种提供了基因资源,具有良好的应用价值,为研究光信号对植物叶绿素合成及光合作用的调控机制奠定理论基础。
附图说明
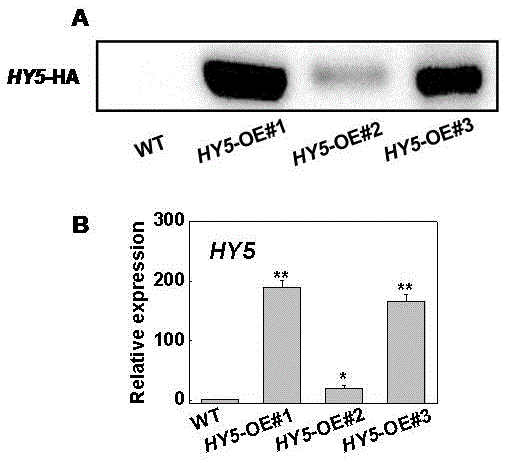
图1为本发明实施例2中slhy5基因过表达番茄株系和野生型的植物蛋白westernblot及基因表达的检测结果;其中,a为slhy5蛋白含量的水平,b为slhy5基因的表达水平;
图2为本发明实施例3中番茄slhy5过表达株系和野生型的叶绿素含量及其表型;其中,a为slhy5过表达株系和野生型的叶绿素含量,b为slhy5过表达株系和野生型番茄叶片的表型;
图3为本发明实施例3中番茄slhy5过表达株系和野生型的光系统ⅱ最大光化学效率(φpsii)和光合速率(pn);其中,a为slhy5过表达株系和野生型的光系统ⅱ最大光化学效率,b为slhy5过表达株系和野生型番茄叶片的光合速率;
图4为本发明实施例4中番茄slhy5过表达株系和野生型的赤霉素(ga)的含量;其中,a为ga44,b为ga19,c为ga20,d为ga1;
图5为本发明实施例4中番茄slhy5过表达株系和野生型中ga负调控因子slpro的基因表达水平;
图6为本发明实施例5中番茄pro突变体及野生型中叶绿素含量和表型;其中,a为番茄pro突变体及野生型的叶绿素含量,b为番茄pro突变体及野生型的表型;
图7为本发明实施例5中番茄pro突变体及野生型中光系统ⅱ最大光化学效率(φpsii)和光合速率(pn)的变化;其中,a为番茄pro突变体及野生型的光系统ⅱ最大光化学效率,b为番茄pro突变体及野生型番茄叶片的光合速率。
具体实施方式
下面结合具体实施例对本发明作进一步说明,但本发明并不限于以下实施例。本发明旨在涵盖本发明公开的范围和精神内的其他更改和变型。
除非另有说明,本发明的实施将使用本领域技术人员显而易见的植物学常规技术、组织培养、分子生物学、生物生理生化、dna重组及生物信息学技术。这些技术在文献中进行了充分解释(zhangetal.,2014;wangetal.,2015;lietal.,2016a,2016b;zhangetal.,2016);番茄(品种ailsacraig:世界番茄遗传资源中心购买获得,网址为:http://tgrc.ucdavis.edu)。
实施例1:slhy5基因过表达载体的构建
为了明确光信号对番茄植物叶绿素合成的调控机制,提取番茄叶片总rna,反转录获得cdna,以cdna为模板,从番茄基因组中克隆了slhy5基因。
克隆slhy5基因及构建过表达载体的具体方法:根据编码区序列分析,设计特异性引物并在引物上分别加上限制性酶切位点(asci和sali),引物slhy5-oe-f和slhy5-oe-r的序列分别为ttggcgcgccatgcaagagcaagcgacgag(如seqidno:3所示)和acgcgtcgaccttcctcccttcctgtgcac(如seqidno:4所示)。用primerstar高保真酶(北京天根科技有限公司)pcr克隆slhy5片段,然后对pcr克隆片段及pfgc1008-ha载体(biovectorntcc中国质粒载体菌种细胞基因保藏中心获得)进行双酶切,利用t4连接酶将slhy5片段连接到pfgc1008-ha上(wangetal.,2015),得到过表达载体pfgc1008::slhy5-ha。将上述重组质粒送到擎科公司测序确认。
所测基因slhy5的核苷酸序列如seqidno:1所示:
atgcaagagcaagcgacgagttctattgccgctagttcactaccttcaagtagtgagaga
tcatctagttcagctttacatcatgaactcaaagaaggtatggagagtgatgatgagatc
agaagagtgccggagatgggcggagaagcgacggggacaacgtcagcttctggaagagat
ggagtatcggccgccggtcaagctcaaccatcagctgggactcaaaggaagagaggaaga
agcccagctgacaaagaaaacaaaaggttaaaaagattgttgagaaatagagtatcagca
caacaagcaagggagaggaagaaagcatatttgatagatctggaagcaagggtgaaggaa
ttggaaacaaagaatgcagaacttgaagagaggttgtctactttgcaaaatgagaaccaa
atgcttagacatatactgaagaacacaacagcaggtgcacaggaagggaggaagtag;
该基因编码的蛋白质的氨基酸序列如seqidno:2所示:
metglngluglnalathrserserilealaalaserserleuproser
sersergluargserserserseralaleuhishisgluleulysglu
glymetgluseraspaspgluileargargvalproglumetglygly
glualathrglythrthrseralaserglyargaspglyvalserala
alaglyglnalaglnproseralaglythrglnarglysargglyarg
serproalaasplysgluasnlysargleulysargleuleuargasn
argvalseralaglnglnalaarggluarglyslysalatyrleuile
aspleuglualaargvallysgluleugluthrlysasnalagluleu
glugluargleuserthrleuglnasngluasnglnmetleuarghis
ileleulysasnthrthralaglyalaglngluglyarglys。
实施例2:番茄slhy5过表达转基因材料的构建与检测
将过表达载体pfgc1008::slhy5-ha转化农杆菌eha105,并对番茄(品种ailsacraig:世界番茄遗传资源中心购买获得,http://tgrc.ucdavis.edu)进行子叶侵染,通过植物组织培养技术,诱导愈伤,抗性诱导分化以及生根培养,获得组培苗(fillattietal.,1987),将t2代种子分别进行氯霉素抗性测试,选择3/4具有抗性而其余1/4没有抗性的株系,说明在该株系中连有目的基因的过表达载体以单拷贝形式插入。将这些植株移出,再进行单株收种。利用westernblot验证slhy5过表达阳性转基因植株(wangetal.,2015),结果显示野生型没有蛋白条带,而过表达株系有slhy5-ha的条带(图1a),再利用实时荧光定量qpcr技术验证阳性slhy5过表达转基因植株(lietal.,2016a),发现番茄slhy5-oe#1和slhy5-oe#3株系中slhy5基因的表达量相对于野生型显著提高,约150倍以上,而slhy5-oe#2株系中slhy5基因表达水平的提高倍数较低(图1b),所以后续实验选择番茄slhy5-oe#1和slhy5-oe#3株系。
实施例3:番茄slhy5转基因材料中叶绿素含量及光合效率的检测
番茄幼苗植株在人工气候室中进行培养,期间昼夜温度为25℃/20℃,光照时间为12h,光强为600μmolm-2s-1。待番茄幼苗长到五叶一心时,检测野生型番茄幼苗(wt)和实施例2中所得的slhy5基因过表达株系的叶绿素含量(图2;zhangetal.,2016)、光系统ⅱ最大光化学效率(φpsii,图3a;zhangetal.,2014;lietal.,2016b)和光合速率(pn,图3b;zhangetal.,2014;lietal.,2016b),结果显示,与野生型番茄(wt)相比,slhy5基因过表达显著提高了番茄叶片中的叶绿素含量(图2),同时,番茄植株中φpsii和pn均显著提高(图3)。由此可见,番茄slhy5通过促进植物叶绿素的合成,提高植物光系统ⅱ最大光化学效率和光合速率。
实施例4:番茄slhy5转基因材料体内赤霉素含量及slpro基因表达水平的检测
番茄幼苗植株在人工气候室中进行培养,期间昼夜温度为25℃/20℃,光照时间为12h,光强为600μmolm-2s-1。待番茄幼苗长到五叶一心时,检测野生型番茄幼苗(wt)和实施例2中所得的slhy5基因过表达株系叶片激素ga含量(lietal.,2016a;图4)和ga负调控因子slpro的基因表达水平(wangetal.,2015;lietal.,2016a;图5)。以上结果显示,番茄slhy5基因过表达株系叶片中的ga含量(ga44、ga19、ga20、ga1,图4)显著降低,而slpro基因的表达水平明显升高(图5)。这说明番茄slhy5通过抑制ga含量来诱导ga信号负调控因子slpro的表达。
实施例5:番茄slpro基因突变体材料中叶绿素含量及光合效率的检测
番茄幼苗植株在人工气候室中进行培养,期间昼夜温度为25℃/20℃,光照时间为12h,光强为600μmolm-2s-1。待番茄幼苗长到五叶一心时,检测野生型番茄幼苗(wt)和slpro基因突变体材料(pro)中的叶绿素含量(图6;zhangetal.,2016)、光系统ⅱ最大光化学效率(φpsii,图7a;zhangetal.,2014;lietal.,2016b)和光合速率(pn,图7b;zhangetal.,2014;lietal.,2016b),结果显示,与野生型番茄(wt)相比,pro突变体中番茄叶绿素含量明显降低(图6),同时,番茄植株中φpsii和pn均显著下降(图7)。由此可见,番茄slpro可以促进植物叶绿素的合成,同时提高植物光系统ⅱ最大光化学效率和光合速率。
由此可见,番茄slhy5通过抑制ga,诱导植物slpro基因表达,进而促进植物叶绿素合成及提高植物的光化学效率和光合速率。
应当理解,本领域技术人员可根据本发明公开的氨基酸序列,在不影响其活性的前提下,取代、缺失和/或增加一个或几个氨基酸,得到所述蛋白的突变序列。应理解,考虑到密码子的简并性及不同物种密码子的偏爱性,本领域技术人员可以根据需要使用适合特定物种表达的密码子。
虽然,上文中已经用一般性说明及具体实施方案对本发明作了详尽的描述,但本发明不限于以上实施例,还可以有许多变形或改进,这对本领域技术人员而言是显而易见的。因此,在不偏离本发明精神的基础上所做的这些修改或改进,均属于本发明要求保护的范围。
序列表
<110>沈阳农业大学
<120>一种促进番茄叶绿素合成及光合效率的基因及应用
<160>4
<170>siposequencelisting1.0
<210>1
<211>477
<212>dna
<213>番茄(solanumlycopersicum)
<400>1
atgcaagagcaagcgacgagttctattgccgctagttcactaccttcaagtagtgagaga60
tcatctagttcagctttacatcatgaactcaaagaaggtatggagagtgatgatgagatc120
agaagagtgccggagatgggcggagaagcgacggggacaacgtcagcttctggaagagat180
ggagtatcggccgccggtcaagctcaaccatcagctgggactcaaaggaagagaggaaga240
agcccagctgacaaagaaaacaaaaggttaaaaagattgttgagaaatagagtatcagca300
caacaagcaagggagaggaagaaagcatatttgatagatctggaagcaagggtgaaggaa360
ttggaaacaaagaatgcagaacttgaagagaggttgtctactttgcaaaatgagaaccaa420
atgcttagacatatactgaagaacacaacagcaggtgcacaggaagggaggaagtag477
<210>2
<211>158
<212>prt
<213>番茄(solanumlycopersicum)
<400>2
metglngluglnalathrserserilealaalaserserleuproser
151015
sersergluargserserserseralaleuhishisgluleulysglu
202530
glymetgluseraspaspgluileargargvalproglumetglygly
354045
glualathrglythrthrseralaserglyargaspglyvalserala
505560
alaglyglnalaglnproseralaglythrglnarglysargglyarg
65707580
serproalaasplysgluasnlysargleulysargleuleuargasn
859095
argvalseralaglnglnalaarggluarglyslysalatyrleuile
100105110
aspleuglualaargvallysgluleugluthrlysasnalagluleu
115120125
glugluargleuserthrleuglnasngluasnglnmetleuarghis
130135140
ileleulysasnthrthralaglyalaglngluglyarglys
145150155
<210>3
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>3
ttggcgcgccatgcaagagcaagcgacgag30
<210>4
<211>30
<212>dna
<213>人工序列(artificialsequence)
<400>4
acgcgtcgaccttcctcccttcctgtgcac30
- 还没有人留言评论。精彩留言会获得点赞!