一种去氢表雄酮的制备方法及其制备用酶与流程
本发明涉及生物医药技术领域,特别涉及一种去氢表雄酮的制备方法及其制备用酶。
背景技术:
去氢表雄酮(脱氢表雄酮)(dehyroepiandrosterone,dhea)是一种c19类固醇载体化合物,化学名称为3β-羟基雄甾-5-烯-17-酮,分子式c19h28o2。dhea是人血浆中大量共存在的肾上腺甾体化合物,具有抗炎、抗增殖及抗抑郁等活性,其衍生物在实验中能促进动物免疫应答,而且具有很好的神经保护作用。此外,dhea还是合成甾体药物的重要中间体。近几年来,dhea凭借其重要研究价值,已逐渐发展成为最具热门的研究对象之一。
传统的dhea的合成大多采用化学方法,例如以孕甾双烯醇酮或醋酸妊娠双烯醇酮为原料,经过肟化、重排与水解后得到产物。然而所述合成方法分3步以上化学反应完成,原料价格高昂,并且工艺中使用苯、卤代烷或吡啶等溶剂,对环境伤害较大。此外,传统工艺中的产品纯度低、效率低,后期提纯操作繁琐。
因此,开发一种步骤少、成本低、产率高且绿色环保的dhea合成方法具有重要意义。
技术实现要素:
为了解决上述技术问题,本发明提供了一种去氢表雄酮的制备方法及其制备用酶,所述去氢表雄酮的制备方法共涉及二步反应,大大缩少了传统方法制备过程中繁多的合成步骤,同时本发明提供的制备用酶使所述去氢表雄酮的合成工艺具备高收率、低成本且绿色环保的特点。
第一方面,本发明提供了一种去氢表雄酮的制备方法,包括:
(1)在保护气氛下,向叔丁醇中加入叔丁醇钾,搅拌均匀后加入4-雄烯二酮得到混合料,将所述混合料滴加至含有抗坏血酸钠的乙酸溶液中反应得到5-雄烯二酮;
(2)将步骤(1)中所述5-雄烯二酮溶于有机溶剂中,并添加酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶得到混合液,控制所述混合液的ph为6.0-6.3,于22-26℃下搅拌反应1-6小时,得到反应液,将所述反应液分离提纯后得到去氢表雄酮,所述酮还原酶和所述葡萄糖脱氢酶由微生物菌株共表达产生并以粗酶液形式添加,所述酮还原酶的来源于领地伯克霍尔德菌属、红球菌属和鞘氨醇单胞菌属中的一种或多种。
本发明中,所述去氢表雄酮的制备方法的具体工艺路线如下所示:
其中,所述工艺中共包括两步反应,第一步为化学法,第二步为生物酶法,所述4-雄烯二酮(4-androstene-3,17-dione,4-ad)如所式(ⅰ)所示,所述5-雄烯二酮(5-androstene-3,17-dione,5-ad)如式(ⅱ)所示,所述去氢表雄酮如式(ⅲ)所示。
可选地,所述步骤(1)中,所述保护气氛是指所述充满氮气或惰性气体的环境下。所述惰性气体包括氦(he)、氖(ne)、氩(ar)、氪(kr)和氙(xe)中的一种或多种。
可选地,所述步骤(1)中,所述混合料的具体制备过程包括:在室温下向叔丁醇中加入叔丁醇钾,然后升温至55-65℃,搅拌0.3-0.8小时至所述叔丁醇钾完全溶解后,将温度维持在25-30℃下,加入4-雄烯二酮搅拌0.3-0.8小时后,冷却得到所述混合料。本发明中,在加入叔丁醇钾后,将温度控制在55-65℃,可以大大缩短叔丁醇钾的溶解时间,可以将传统3小时以上的溶解时间缩短至0.5小时左右;随后将温度维持在25-30℃下,可以促进所述4-雄烯二酮高效地正向反应,若温度过高容易逆反应生成4-雄烯二酮,若温度过低则该反应过程耗时较长,生产效率低。
可选地,所述步骤(1)中,所述将所述混合料滴加至含有抗坏血酸钠的乙酸溶液中反应得到5-雄烯二酮的过程中,反应温度为25-30℃,反应时间为0.3-0.8小时。进一步优选地,所述反应温度可以为26-30℃,反应时间为0.5-0.8小时。例如,所述反应温度为30℃。
可选地,步骤(1)中,所述4-雄烯二酮和所述叔丁醇钾的摩尔比为1:(0.75-1.2)。进一步可选地,所述4-雄烯二酮和所述叔丁醇钾的摩尔比为1:(0.8-1.2)。
可选地,所述步骤(2)中,所述有机溶剂包括疏水性溶剂。进一步可选地,所述有机溶剂包括乙酸乙酯。本发明步骤(2)中优选的乙酸乙酯溶剂可以有效提高所述去氢表雄酮的产量。
可选地,步骤(1)中,所述4-雄烯二酮、所述乙酸和所述抗坏血酸钠的摩尔比为1:(0.5-1):(0.04-0.4)。进一步可选地,所述4-雄烯二酮、所述乙酸和所述抗坏血酸钠的摩尔比为1:(0.6-1):(0.05-0.2)。
可选地,所述步骤(2)中,所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶为水溶液或缓冲溶液。进一步可选地,所述缓冲溶液包括磷酸盐缓冲液、tris-hcl缓冲液或其他缓冲溶液,所述缓冲溶液的ph为6.0-7.0。例如所述缓冲溶液为磷酸钠缓冲液。可选地,所述缓冲溶液的浓度为50-120mmol/l。进一步可选地,所述缓冲溶液的浓度为60-100mmol/l。优选的,所述缓冲溶液的浓度为80-120mmol/l。例如,所述缓冲溶液的浓度为60mmol/l,或为70mmol/l,或为90mmol/l,或为100mmol/l,或为120mmol/l。
可选地,所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶这四种组分的添加形式可以是将酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶这四种混合后同时添加;或单独添加所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶,其中,当单独添加时,所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶按顺序依次加入且添加顺序不做限定。所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶的添加形式还可以为:取所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶中的一种或多种,并与所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶中剩下的组分混合后进行添加至所述含有5-雄烯二酮的有机溶剂中。例如,所述酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶中的所述酮还原酶和所述葡萄糖脱氢酶先混合在一起,然后在酮还原酶和葡萄糖脱氢酶的混合组分中再添加剩下的葡萄糖和氧化还原辅酶这两种组分,得到了含有酮还原酶、葡萄糖脱氢酶、葡萄糖和氧化还原辅酶的混合组分,并将其添加至所述含有5-雄烯二酮的有机溶剂中得到混合液。
可选地,所述步骤(2)中,所述葡萄糖的浓度为0.04-0.12mg/ml。进一步可选地,所述葡萄糖的浓度为0.08-0.1mg/ml。例如,所述葡萄糖在所述缓冲溶液中的浓度为0.1mg/ml,或为0.12mg/ml。
可选地,所述步骤(2)中,所述微生物菌株包括rosetta(de3)大肠杆菌和bl21(de3)大肠杆菌中的一种或两种。进一步可选地,所述微生物菌株包括rosetta(de3)大肠杆菌。
可选地,所述步骤(2)中,所述微生物菌株共表达产生所述酮还原酶和所述葡萄糖脱氢酶的具体过程包括:构建一重组表达质粒,将所述重组表达质粒转染至所述微生物菌株内后,在微生物培养液中诱导含有所述重组表达质粒的微生物菌株表达所述酮还原酶和所述葡萄糖脱氢酶,其中所述重组表达质粒包括所述酮还原酶和所述葡萄糖脱氢酶的基因编码序列。所述酮还原酶和所述葡萄糖脱氢酶可以在所述微生物菌株的胞内生成,也可以由所述微生物菌株的胞内生成后在释放在所述微生物培养液中。通过对所述能够表达酮还原酶和葡萄糖脱氢酶的微生物菌株进行收集、清洗、破胞后收集得到所述粗酶液,所述粗酶液中包含有所述酮还原酶和所述葡萄糖脱氢酶。
本发明中,所述酮还原酶和所述葡萄糖脱氢酶是两个独立的蛋白分子,并通过上述共表达的方式得到,这样操作既可以节省生产成本,又能简化实验过程。所述重组表达质粒可以同时含有所述酮还原酶和所述葡萄糖脱氢酶的基因编码序列,并能够同时表达所述酮还原酶和所述葡萄糖脱氢酶两个蛋白分子。本发明中,以粗酶液的形式参与所述步骤(2)中的反应,例如所述粗酶液可以是由大肠杆菌rosetta(de3)诱导表达所述酮还原酶和所述葡萄糖脱氢酶后,对所述大肠杆菌rosetta(de3)进行离心收集、清洗后,将菌体悬浮于缓冲溶液中并进行超声破胞后,所述酮还原酶和所述葡萄糖脱氢酶被释放至缓冲溶液中,得到所述粗酶液。本发明通过采用共表达形式得到酮还原酶和葡萄糖脱氢酶粗酶液参与dhea的制备,不仅精简制备工艺,而且还提高产量,节约成本,绿色环保。进一步地,本发明还可以对所述酮还原酶和葡萄糖脱氢酶提纯并参与合成反应。
可选地,所述酮还原酶的基因编码序列包括如seqidno:1、seqidno:2和seqidno:3所示的核苷酸序列中的任意一种。可选地,所述葡萄糖脱氢酶的基因编码序列包括如seqidno:4所示的核苷酸序列。本发明优选地具有上述包括如seqidno:1、seqidno:2和seqidno:3所示的核苷酸序列中的任意一种的酮还原酶可以在所述微生物菌株中高效表达,所述粗酶液具有较高浓度的所述酮还原酶。
可选地,所述步骤(2)中,所述酮还原酶、所述葡萄糖脱氢酶和所述4-雄烯二酮的摩尔比为(0.015-0.04):(0.01-0.04):1。
所述步骤(2)中,所述氧化还原辅酶包括nad+、nadp+、nadh和nadph中的一种或多种。进一步可选地,所述步骤(2)中,所述氧化还原辅酶包括nad+。可选地,所述步骤(2)中,所述氧化还原辅酶的浓度为0.04-0.4mg/ml。进一步可选地,所述氧化还原辅酶的浓度为0.08-0.2mg/ml。例如,所述氧化还原辅酶在所述缓冲溶液中的浓度为0.1mg/ml,或为0.15mg/ml,或为0.2mg/ml。
其中,所述nad+为烟酰胺腺嘌呤二核苷酸(nicotinamideadeninedinucleotide,nad+),也称为氧化型辅酶ⅰ;所述nadh为烟酰胺腺嘌呤二核苷酸的还原态,也称为还原型辅酶ⅰ;所述nadp+为烟酰胺腺嘌呤二核苷酸磷酸(nicotinamideadeninedinucleotidephosphate,nadp+),也称为氧化型辅酶ⅱ;所述nadph为烟酰胺腺嘌呤二核苷酸磷酸的还原态,也称为还原型辅酶ⅱ。在存在氧气(o2)的条件下,所述氧化还原辅酶,葡萄糖脱氢酶以及葡萄糖之间形成一个辅酶再生体系,该体系是一电子循环体系,在酮还原酶作用下,可以有效使5-雄烯二酮还原生成去氢表雄酮,提高去氢表雄酮的产量。本发明体系中,可以仅仅使用所述nad+、nadp+、nadh和nadph中的nad+就能具有实现循环,且所述去氢表雄酮的具有很高产率。
可选地,所述步骤(2)中分离提纯的过程包括依次进行的过滤、清洗和萃取过程。进一步可选地,所述步骤(2)之后还包括对所述去氢表雄酮进行重结晶。
可选地,所述步骤(2)中分离提纯的具体过程包括:将所述反应液加至硅藻土中过滤得到滤液和滤饼,将滤饼用乙酸乙酯洗涤后得到洗涤液,收集并混合所述洗涤液和所述滤液,得到水相和有机相,对所述水相用乙酸乙酯萃取两或三次后得到萃取液,将所述萃取液与所述有机相混合并经浓缩、过滤、清洗、洗涤干燥后得到去氢表雄酮。
本方面第一方面所提供的去氢表雄酮的制备方法,所述制备方法工艺简单,省时高效、成本低廉、绿色环保,该制备方法可适用于大规模工业化生产;由所述制备方法制得的去氢表雄酮具有极高的产率。
第二方面,本发明提供了一种酮还原酶,所述酮还原酶包括酮还原酶kred3(简写成kred3)、酮还原酶kred4(简写成kred4)和酮还原酶kred5(简写成kred5)中的一种或多种,所述酮还原酶kred3的基因编码序列包括如seqidno:1所示的核苷酸序列,所述酮还原酶kred4的基因编码序列包括如seqidno:2所示的核苷酸序列,所述酮还原酶kred5的基因编码序列包括如seqidno:3所示的核苷酸序列。
可选地,所述kred3的基因编码序列还包括如seqidno:1所示的核苷酸序列至少95%同源性的核苷酸序列,所述kred4的基因编码序列还包括如seqidno:2所示的核苷酸序列至少95%同源性的核苷酸序列,所述kred5的基因编码序列还包括如seqidno:3所示的核苷酸序列至少95%同源性的核苷酸序列。
其中,所述kred3的氨基酸序列包括如seqidno:5所示的氨基酸序列,所述kred4的氨基酸序列包括如seqidno:6所示的氨基酸序列,所述kred5的氨基酸序列包括如seqidno:7所示的氨基酸序列。可选地,所述葡萄糖脱氢酶的氨基酸序列包括如seqidno:8所示的氨基酸序列。
可选地,本发明所述酮还原酶kred3的基因来源于领地伯克霍尔德菌(burkholderiaterritorii),所述酮还原酶kred4的基因来源于红球菌属(rhodococcus),所述酮还原酶kred5的基因来源于鞘氨醇单胞菌属(sphingomonassp.)。所述葡萄糖脱氢酶的基因来源于巨大芽孢杆菌(bacillusmegaterium)。本发明优选地酮还原酶具有很强的化学选择性和区域选择性,在一定条件下,可以高效地将所述5-雄烯二酮转化为所述去氢表雄酮。
可选地,所述酮还原酶kred3、kred4和kred5中的任意一个蛋白酶的基因片段上还包括his标签。进一步可选地,所述酮还原酶kred3、kred4和kred5中的任意一个蛋白酶的基因片段上还包括8×his标签。所述葡萄糖脱氢酶的基因片段上还包括6×his标签。随着所述his标签的his数目的增加,所述蛋白的结合效率,纯化收率增加;本发明可以通过对所述所述酮还原酶和所述葡萄糖脱氢酶上连接不同his个数的his标签来分别提纯。
本发明第二方面提供的酮还原酶kred3、kred4和kred5为本发明第一方面所述去氢表雄酮的制备方法的制备用酶之一,在所述基因片段的上增设his标签(组氨酸标签)的核苷酸序列,能使表达后的蛋白带上his标签,his标签有利于表达后蛋白的分离纯化,及在实验中的分析和追踪,比如用于免疫印迹实验时的分析。
第三方面,本方面还提供了一种酮还原酶在催化酮或醛转化为手性醇中的应用,所述酮还原酶的基因编码序列包括如seqidno:1、seqidno:2和seqidno:3所示的核苷酸序列中的任意一种。即本发明所述酮还原酶可以高效、有选择性地将醛或酮上的羰基催化还原为手性羟基。
可选地,所述催化酮或醛转化为手性醇中的应用包括催化5-雄烯二酮转化为去氢表雄酮中的应用。本发明优选地酮还原酶具有很强的化学选择性和区域选择性,在一定条件下,可以高效地、有选择性地将所述5-雄烯二酮转化为所述去氢表雄酮。
第四方面,本方面还提供了一种重组质粒,所述重组质粒包括酮还原酶的基因编码序列,所述酮还原酶的基因编码序列包括kred3的基因编码序列、kred4的基因编码序列和kred5的基因编码序列中的一种或多种,所述kred3的基因编码序列包括如seqidno:1所示的核苷酸序列,所述kred4的基因编码序列包括如seqidno:2所示的核苷酸序列,所述kred5的基因编码序列包括如seqidno:3所示的核苷酸序列。
可选地,所述重组质粒还包括葡萄糖脱氢酶的基因编码序列,所述葡萄糖脱氢酶的基因编码序列与所酮还原酶的基因编码序列之间还插入有rbs序列,所述葡萄糖脱氢酶的基因编码序列包括如seqidno:4所示的核苷酸序列,所述rbs序列包括如seqidno:17所示的核苷酸序列。可选地,所述重组质粒包括从5’端到3’端顺次连接的酮还原酶的基因编码序列、rbs序列和葡萄糖脱氢酶的基因编码序列。本发明所述rbs序列为核糖体结合位点(ribosomebindingsite,rbs)序列,可以有效促进两个同源基因(酮还原酶基因和葡萄糖脱氢酶基因)进行独立的转录及翻译。
本发明第四方面所述重组质粒可用于共表达酮还原酶和葡萄糖脱氢酶,该酮还原酶和葡萄糖脱氢酶具有很好的活性,可广泛应用于生物制药、蛋白生产等领域。
第五方面,本发明还提供了一种重组质粒的制备方法,包括:
(1)提供或制备包含酮还原酶和葡萄糖脱氢酶的基因模板;
(2)制备所述酮还原酶和所述葡萄糖脱氢酶的基因的上游引物和下游引物,扩增所述酮还原酶和所述葡萄糖脱氢酶的基因片段,所述酮还原酶的基因编码序列包括如seqidno:1、seqidno:2和seqidno:3所示的核苷酸序列中的任意一种,所述葡萄糖脱氢酶的基因编码序列包括如seqidno:4所示的核苷酸序列;
(3)取pet22b(+)载体质粒,将步骤(2)扩增得到的所述酮还原酶和所述葡萄糖脱氢酶基因片段插入至所述pet22b(+)载体质粒,纯化回收后进行连接,得到所述重组质粒。
可选地,所述酮还原酶的基因插入到pet22b(+)载体质粒中ndei和ecori酶切位点之间。所述酮还原酶的基因插入到pet22b(+)载体质粒时,所述酮还原酶的基因编码序列的3’端可加入终止密码子(如taa)与pet22b(+)载体质粒中ecorⅰ酶切位点相连。
所述葡萄糖脱氢酶的基因插入到pet22b(+)载体质粒中ecori和xhoi酶切位点之间。所述葡萄糖脱氢酶的基因插入到pet22b(+)载体质粒时,所述葡萄糖脱氢酶的基因编码序列的3’端可加入终止密码子(如taa)与pet22b(+)载体质粒中xhoi酶切位点相连。
可选地,在所述酮还原酶和所述葡萄糖脱氢酶基因编码序列之间插入rbs序列,所述rbs序列包括如seqidno:17所示的核苷酸序列可选地,所述酮还原酶的基因编码序列包括如seqidno:1、seqidno:2和seqidno:3所示的核苷酸序列中的任意一种,所述葡萄糖脱氢酶的基因编码序列包括如seqidno:4所示的核苷酸序列。所述酮还原酶和所述葡萄糖脱氢酶基因编码序列在所述pet22b(+)载体质粒的先后顺序可以是所述酮还原酶基因的3’端位于所述葡萄糖脱氢酶基因的5’端之前,或所述酮还原酶基因的5’端位于所述葡萄糖脱氢酶基因的3’端之后。
进一步可选地,所述rbs序列插入至所述葡萄糖脱氢酶基因序列的5’端前。所述rbs序列的3’端与所述葡萄糖脱氢酶基因序列的5’端之间还可以设置一段linker,便于所述葡萄糖脱氢酶的翻译识别,有利于酮还原酶和葡萄糖脱氢酶能够在同一载体中分别进行翻译表达。优选地,所述linker的序列可以为“atatacat”。
可选地,所述步骤(3)中,所示pet22b(+)载体质粒采用的酶切位点包括ndei内切酶和xhoi内切酶。
本发明中,所述制备方法可以制备仅含有酮还原酶或葡萄糖脱氢酶的重组质粒,也可以制备同时含有酮还原酶和葡萄糖脱氢酶的重组质粒。可选地,将所述重组质粒转染至微生物菌株中诱导表达可以获得本发明所述酮还原酶和葡萄糖脱氢酶中的一种或两种。
本发明的有益效果包括以下几个方面:
1、本发明所述去氢表雄酮的制备方法,共包括两步反应,以4-雄烯二酮为初始原料,并采用酮还原酶等生物酶,生产成本低廉,工艺简单,省时高效,反应条件温和,大大提高了产品的收率;
2、本发明所述去氢表雄酮的制备方法,绿色环保,从根源上避免了环境污染问题,进一步降低成本,可以广泛适用于工业化规模生产;
3、由本发明所述的方法制备的去氢表雄酮,纯度高,可在制药领域或生物医学领域中有广泛的应用。
附图说明
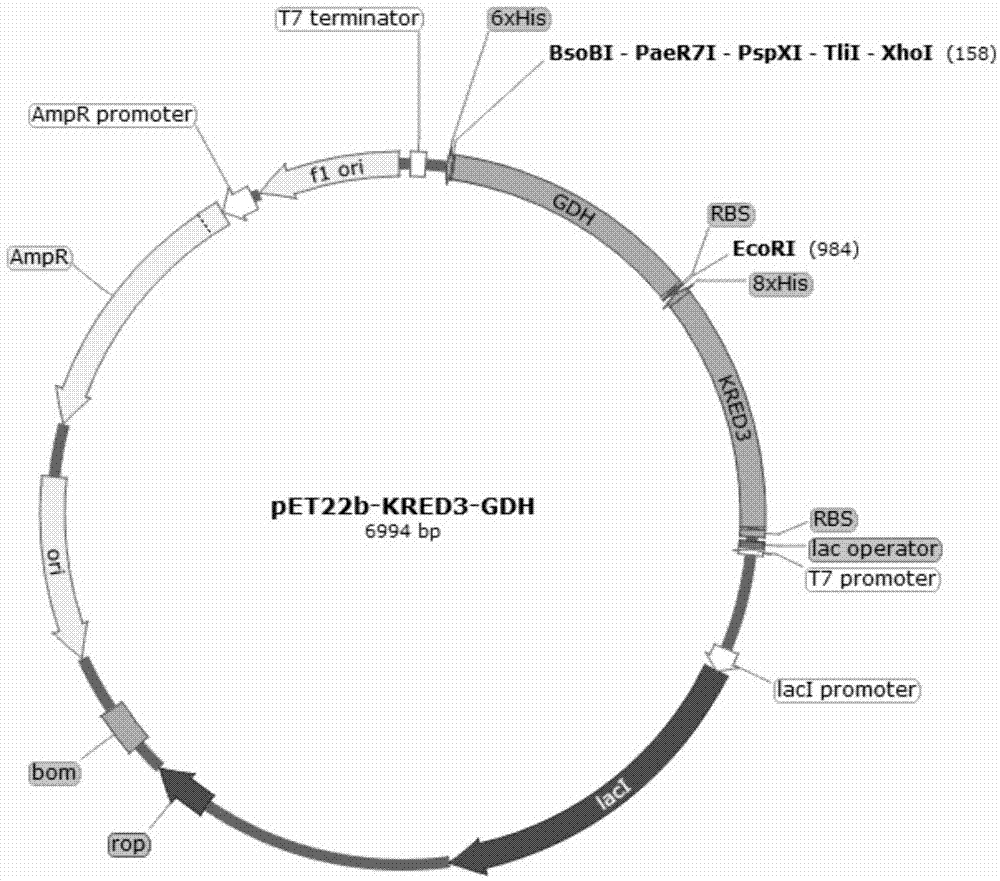
图1为本发明一实施例提供的pet22b-kred3-gdh重组质粒的质粒图谱;
图2为本发明一实施例提供的pet22b-kred4-gdh重组质粒的质粒图谱;
图3为本发明一实施例提供的pet22b-kred5-gdh重组质粒的质粒图谱;
图4为本发明一实施例提供的kred3-gdh粗酶液的凝胶电泳图;
图5为本发明一实施例提供的kred4-gdh粗酶液的凝胶电泳图;
图6为本发明一实施例提供的kred5-gdh粗酶液的凝胶电泳图;
图7为本发明一实施例提供的4-雄烯二酮的高效液相色谱图;
图8为本发明一实施例提供的5-雄烯二酮的高效液相色谱图;
图9为本发明一实施例提供的去氢表雄酮标准品的高效液相色谱图;
图10为本发明一实施例提供的去氢表雄酮粗产品的高效液相色谱图。
具体实施方式
以下所述是本发明实施例的优选实施方式,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明实施例原理的前提下,还可以做出若干改进和润饰,这些改进和润饰也视为本发明实施例的保护范围。
若无特别说明,本发明实施例所采用的原料及其它化学试剂皆为市售商品。
本发明实施例提供了一种酮还原酶和葡萄糖脱氢酶的制备方法,所述酮还原酶和葡萄糖脱氢酶为所述去氢表雄酮的制备方法中的制备用酶,具体的实验步骤包括:
(1)构建pet22b-kred3/4/5-gdh重组质粒
提供上游引物和下游引物,采用pcr扩增实验得到酮还原酶kred3、酮还原酶kred4、酮还原酶kred5和葡萄糖脱氢酶的基因编码序列。其中,所述酮还原酶kred3的基因编码序列如seqidno:1所示,所述酮还原酶kred4的基因编码序列如seqidno:2所示,所述酮还原酶kred5的基因编码序列如seqidno:3所示,所述葡萄糖脱氢酶的基因编码序列如seqidno:4所示。所述酮还原酶kred3对应的上游引物的碱基序列如seqidno:9所示,下游引物的碱基序列如seqidno:10所示。所述酮还原酶kred3的基因对应的上游引物的碱基序列如seqidno:9所示,下游引物的碱基序列如seqidno:10所示。所述酮还原酶kred4的基因对应的上游引物的碱基序列如seqidno:11所示,下游引物的碱基序列如seqidno:12所示。所述酮还原酶kred5的基因对应的上游引物的碱基序列如seqidno:13所示,下游引物的碱基序列如seqidno:14所示。所述葡萄糖脱氢酶的基因对应的上游引物的碱基序列如seqidno:15所示,下游引物的碱基序列如seqidno:16所示。在所述葡萄糖脱氢酶的基因的5’端前插入rbs序列,并可通过上下游引物进行将其与插入至质粒pet22b(+)中,所述rbs序列如seqidno:17所示。
通过对应的上游引物和下游引物分别进行pcr扩增体系,配置扩增体系如下:
pcr扩增程序为:98℃预变性2min;98℃变性10s;60℃退火30s;72℃延伸1min;30个循环后,72℃延伸10min。pcr产物经胶回收试剂盒纯化后,分别利用限制性内切酶ndei和ecori、ecori和xhoi进行酶切,酶切后用t4连接酶连接已用ndei和xhoi酶切处理的质粒pet22b(+)。连接产物转入大肠杆菌dh5α,经过氨苄抗性(amp+)的筛选后挑取菌落送测序,测序成功后即得到酮还原酶和葡萄糖脱氢酶共表达的重组质粒pet22b-kred3/4/5-gdh(包括pet22b-kred3-gdh、pet22b-kred4-gdh或pet22b-kred5-gdh)。所述重组质粒pet22b-kred3-gdh的质粒图谱如图1所示;所述重组质粒pet22b-kred4-gdh的质粒图谱如图2所示;所述重组质粒pet22b-kred5-gdh的质粒图谱如图3所示。本实施方式中所述dna模板分别为质粒pet22b-kred3、pet22b-kred4、pet22b-kred5和pet22b-gdh。
(2)表达酮还原酶和葡萄糖脱氢酶
将构建的重组质粒pet22b-kred3-gdh、pet22b-kred4-gdh和pet22b-kred5-gdh中的一种或多种转入大肠杆菌rosetta(de3)中,得到能够表达酮还原酶和葡萄糖脱氢酶的大肠杆菌rosetta(de3)。将含有重组质粒pet22b-kred3-gdh、pet22b-kred4-gdh或pet22b-kred5-gdh的大肠杆菌以1%的接种量接种至含有10ml的lb培养基(100μg/ml氨苄青霉素(amp))的50ml三角瓶中,维持恒定的37℃,200rpm的摇晃速率,过夜培养后,将菌液以1%的接种量转接到含有1l的lb培养基(100μg/mlamp)的2l三角瓶中,继续37℃恒温培养至培养基中的od600值达到0.6左右,加入异丙基-β-d-硫代半乳糖苷(iptg)诱导剂(体系终浓度0.5mm),在37℃条件培养8小时后离心收集菌体。将菌体分别用4倍质量的100mm的磷酸钾缓冲液(ph=6.5)进行重悬并超声破碎20min,得到kred3-gdh粗酶液、kred4-gdh粗酶液或kred5-gdh粗酶液。所述kred3-gdh粗酶液中含有酮还原酶kred3和葡萄糖脱氢酶。所述kred4-gdh粗酶液中含有酮还原酶kred4和葡萄糖脱氢酶。所述kred5-gdh粗酶液中含有酮还原酶kred5和葡萄糖脱氢酶。
将表达获得的kred3-gdh粗酶液、kred4-gdh粗酶液或kred5-gdh粗酶液分别进行sds-聚丙烯酰胺凝胶电泳(sds-page)鉴定。图4为所述kred3-gdh粗酶液的凝胶电泳图,其中泳道m为蛋白marker(thermoscientificpageruler),泳道1为菌体破胞后总蛋白,泳道2为菌体破胞后上清液,其中kred3的分子量约为28kda,gdh的分子量为26kda。图5为所述kred4-gdh粗酶液的凝胶电泳图,其中泳道m为蛋白marker(thermoscientificpageruler),泳道1为菌体破胞后上清液,泳道2为菌体破胞后总蛋白,其中kred4的分子量为29kda,gdh的分子量为26kda。图6为所述kred5-gdh粗酶液的凝胶电泳图,其中泳道m为蛋白marker(thermoscientificpageruler),泳道1为菌体破胞后上清液,泳道2为菌体破胞后总蛋白,其中kred5的分子量为28kda,gdh的分子量为26kda。所述酮还原酶kred3、kred4和kred5,以及gdh的分子均于相应蛋白的理论计算值相近。
实施例一
一种去氢表雄酮的制备方法,包括以下步骤:
(1)在氮气保护下,在388ml的叔丁醇中加入82.5g叔丁醇钾,然后升温到60℃,搅拌0.5h至叔丁醇钾完全溶解,降温至30℃,持续氮气保护中加入100g的4-ad,持续搅拌0.5h后,冷却至室温得到混合料。在氮气保护下,在205.3g的26.8%的乙酸水溶液中,加入6.0g抗坏血酸钠,得到含有抗坏血酸钠的乙酸溶液。在氮气保护下,30℃下,将所述混合料在30min内缓慢滴加到含有抗坏血酸钠的乙酸溶液中,维持在温度30℃下续搅拌0.5h,室温冷却,得到含有5-ad的反应液。
(2)将500ml乙酸乙酯加入所述5-ad的反应液中,搅拌均匀,加入400ml1.8g/lkred3和gdh的粗酶液,300ml100mmol/l磷酸钠缓冲液(ph6.5),100g葡萄糖,200mgnad+,于25℃搅拌至完全溶解,加入含有5-ad的反应液,控制ph为6.3,温度26℃,以200rpm的速率搅拌3h,得到酶反应液;将酶反应液加入至硅藻土中搅拌、过滤后收集滤饼,并用乙酸乙酯对滤饼洗涤两次,并将洗涤液和滤液混合,分开水相和有机相,水相用乙酸乙酯萃取两次,萃取液与有机相混合,将有机相减压浓缩后,加入600g水并充分搅拌后过滤,滤饼用120g水洗涤两次,将洗涤后的滤饼真空干燥得dhea粗产品。将dhea粗产品转入至干净的容器中,加入234g丙酮溶解后再加200g水,重复上述操作进行重结晶,得到78.02gdhea最终产物,本实施例中,dhea的收率为78.02%。
分别对两步反应进行取样检测,检测条件:费罗门phenyl-hexyl250×4.6mm色谱柱,波长uv210nm,流动相为50%乙腈水溶液,流速1.0ml/min,温度25℃。其中,从4-ad的液相色谱图(见图7)和5-ad合成过程中的液相色谱图(见图8)对比发现,4-ad可以高效地转化生产5-ad;将dhea粗产品的液相色谱图(见图10)与dhea标准品液相色谱图(见图9)进行比对,粗产品中含有纯度较高的dhea。
实施例二
一种去氢表雄酮的制备方法,包括以下步骤:
(1)在氮气保护下,在388ml的叔丁醇中加入80.0g叔丁醇钾,然后升温到55℃,搅拌0.8h至叔丁醇钾完全溶解,降温至30℃,持续氮气保护中加入100g的4-ad,持续搅拌0.6h后,冷却至室温得到混合料。在氮气保护下,在200.0g的26.8%的乙酸水溶液中,加入6.0g抗坏血酸钠,得到含有抗坏血酸钠的乙酸溶液。在氮气保护下,30℃下,将所述混合料在30min内缓慢滴加到含有抗坏血酸钠的乙酸溶液中,维持在温度30℃下续搅拌0.5h,室温冷却,得到含有5-ad的反应液。
(2)将500ml乙酸乙酯加入所述5-ad的反应液中,搅拌均匀,加入400ml1.5g/lkred4和gdh的粗酶液,300ml80mmol/l磷酸钠缓冲液(ph6.0),100g葡萄糖,200mgnad+,于25℃搅拌至完全溶解,加入含有5-ad的反应液,控制ph为6.1,温度22℃,以180rpm的速率搅拌5h,得到酶反应液;将酶反应液加入至硅藻土中搅拌、过滤后收集滤饼,并用乙酸乙酯对滤饼洗涤两次,并将洗涤液和滤液混合,分开水相和有机相,水相用乙酸乙酯萃取两次,萃取液与有机相混合,将有机相减压浓缩后,加入600g水并充分搅拌后过滤,滤饼用120g水洗涤两次,将洗涤后的滤饼真空干燥得dhea粗产品。将dhea粗产品转入至干净的容器中,加入234g丙酮溶解后再加200g水,重复上述操作进行重结晶,得到75.16gdhea最终产物,本实施例中,dhea的收率为75.16%。
实施例三
一种去氢表雄酮的制备方法,包括以下步骤:
(1)在氮气保护下,在400ml的叔丁醇中加入90.0g叔丁醇钾,然后升温到65℃,搅拌0.3h至叔丁醇钾完全溶解,降温至30℃,持续氮气保护中加入100g的4-ad,持续搅拌0.5h后,冷却至室温得到混合料。在氮气保护下,在230g的25%的乙酸水溶液中,加入6.5g抗坏血酸钠,得到含有抗坏血酸钠的乙酸溶液。在氮气保护下,30℃下,将所述混合料在30min内缓慢滴加到含有抗坏血酸钠的乙酸溶液中,维持在温度28℃下续搅拌0.5h,室温冷却,得到含有5-ad的反应液。
(2)将500ml乙酸乙酯加入所述5-ad的反应液中,搅拌均匀,加入400ml2.0g/lkred5和gdh的粗酶液,300ml100mmol/l磷酸钠缓冲液(ph7.0),100g葡萄糖,180mgnadh,于25℃搅拌至完全溶解,加入含有5-ad的反应液,控制ph为6.2,温度26℃,以220rpm的速率搅拌2h,得到酶反应液;将酶反应液加入至硅藻土中搅拌、过滤后收集滤饼,并用乙酸乙酯对滤饼洗涤两次,并将洗涤液和滤液混合,分开水相和有机相,水相用乙酸乙酯萃取两次,萃取液与有机相混合,将有机相减压浓缩后,加入600g水并充分搅拌后过滤,滤饼用120g水洗涤两次,将洗涤后的滤饼真空干燥得dhea粗产品。将dhea粗产品转入至干净的容器中,加入250g丙酮溶解后再加200g水,重复上述操作进行重结晶,得到77.53gdhea最终产物,本实施例中,dhea的收率为77.53%。
以上所述实施例仅表达了本发明的几种实施方式,其描述较为具体和详细,但并不能因此而理解为对本发明专利范围的限制。应当指出的是,对于本领域的普通技术人员来说,在不脱离本发明构思的前提下,还可以做出若干变形和改进,这些都属于本发明的保护范围。因此,本发明专利的保护范围应以所附权利要求为准。
序列表
<110>邦泰生物工程(深圳)有限公司
<120>一种去氢表雄酮的制备方法及其制备用酶
<150>pct/cn2018/075822
<151>2018-02-08
<160>17
<170>siposequencelisting1.0
<210>1
<211>774
<212>dna
<213>人工序列(artificialsequence)
<400>1
atgggccgtctggaaggtaaagttgcaattgtgaccggtggtgcacgcggcatgggcgca60
gcaacctgccgtctgtttgtggaagaaggtgcacgcgttgtgattggtgacgtgctggat120
gcagaaggcgaagcactggcccgtgaactgggcgatgccgcacgttttatgcgtctggat180
gttagtgatgaagcaaattgggcccgcgttgccgatgccaccgttgaacagtttggccgt240
attgatgttctggtgaataatgccgccgtgctgacctttggtgcaattaccgaactgagc300
aaacgtgattttgaacgcgccgttagtattaatctggtgggcacctttgtgggtattcgt360
accattgcaccgcgcatgattgcacagaaacgtggcagcattgtgaatattagcagcgtt420
gatggtctgcgcggtgtgaatgcactggccgcctatgtgagcagcaaatggggtgttcgc480
ggtctgaccaaagttgcagcactggaactgggtcatcagggtgttcgtgttaatagcatt540
catccgggtggtgtgaataccgccatgagcaatccgaccggcgcaccgctggatgaagtg600
aataagcattatacccgcgtgccgctgcagcgcgttggcttaccggatgaaattgcacgt660
gcaaccctgtttctggcaagtgatgaagcgagctattgtaatggcgcagaactgagcgtt720
gatggcggtatggcagcaggtgcatattatccgggtctgccgggtgcaccgttt774
<210>2
<211>789
<212>dna
<213>人工序列(artificialsequence)
<400>2
atggcacgtctggaaggtaaagtggcaattgttaccggcgccgcccagggtatgggtgca60
gctaccgcacgtctgtttgtgcatgaaggcgcccgtgtggttctgggtgacgttctggaa120
gaaaaaggccgtgccctggccgccgaactgggcgacgcagcaatttttaccccgctggat180
gttagtgatgaaagcagctgggaaagtgcagtggccgtggccgttgatcgttttggtggt240
ctggatattctggtgaataatgcaggtgttatgcattgggccccgattgaagatctggat300
gtggcccgcaccgaacgtctgctggatattaatgttctgggcaatctgctgggcgccaaa360
gccgtggttccgaccatgaaaaaagccggtcgtggtgtgattgttaatattagtagtgtg420
gatggcctgcgcggcgttaatggtctggcagcctataccgcaagtaaatgggcagtgcgc480
ggtctgaccaaagcactggcatacgaactgggtccggcaggtattcgcgtgtgtagtgtt540
catccgggcggcgttgataccaccctgggtaatccgggtggcctggtgggcgatgatctg600
cagagtaaatatgtgggtgttccgctgcagcgcattggtgaaagtgaagatattgcccgc660
gccaccctgtttgttgccagtgatgaagcaagttatattagcggcgccgaactggcagtg720
gatggcggttggagcgccggtacctattatccgggtctgccgggtaccccgccggcatta780
atgccgaat789
<210>3
<211>786
<212>dna
<213>人工序列(artificialsequence)
<400>3
atgggccgtctggatggtaaagttgcaattgtgaccggtggcgcccagggtatgggtgca60
gcaaccgcacgcctgtttgccgccgaaggtgcccgtgttgttattggtgacgttattgaa120
gataaaggcgccgaagttgcaaccgaaattggtgacaatgcccgttttagtcgtctggat180
gttcgcagcgaagatgattgggccgccgcagttagcgcagccgtggaagcctttggtacc240
attgatattctggttaataacgcagccgttgttcattttaccccgattgaaatgctggat300
gcaagtgaagcagaacgtgtgctggcaattaatgttctgggcgcaatgctgggcgccaaa360
catgtgaccccggttatgacccgcgccggccatggcgttattgttaatattagcagcgtg420
gatggcctgcgcggctgtaatggtctgagtgtttataccgccagtaaatgggccgtgcgt480
ggcctgagcaaaacccaggccatggaactgggtccgcgtggtattcgcgtgtgtagtatt540
catccgggcggtgttaatacccagatgggtaatccgatgggtatgaccggtgacgccctg600
aataaggattataaacgtgttccgctgcagcgcattggtgaaccggaagaaattgcccgt660
gcaaccctgtttattgcaagtgatgaagccagttatattagtggtgccgaactggccgtt720
gatggcggctggagcgccggttattatcagccggtgctgccgggtgccccggaaggttta780
ggtgca786
<210>4
<211>783
<212>dna
<213>人工序列(artificialsequence)
<400>4
atgtatacagatttaaaagataaagtagtagttgtaacaggcggatcaaaaggattgggt60
cgcgcaatggccgttcgttttggtcaagagcagtcaaaagtggttgtaaactaccgcagc120
aatgaagaagaagcgctagaagtaaaaaaagaaattgaacaagctggcggccaagcaatt180
attgttcgaggcgacgtaacaaaagaggaagacgttgtgaatcttgtagagacagctgtt240
aaagagtttggcacattagacgttatgattaacaatgctggtgttgaaaacccggttcct300
tcacatgaattatcgttagaaaactggaatcaagtaatcgatacaaacttaacaggcgcg360
tttttaggaagccgcgaagcgattaaatattttgttgaaaatgatattaaaggaaacgtt420
attaacatgtccagcgttcacgagatgattccttggccactatttgttcactatgcagca480
agtaaaggcggtatgaaactaatgacagaaacattggctcttgaatatgcgccaaaaggt540
atccgcgtaaataacattggaccaggcgcgatcgatacgccaatcaacgctgaaaaattc600
gcagatccggaacagcgtgcagacgtagaaagcatgattccaatgggctacatcggcaac660
ccggaagaaattgcatcagttgcagcattcttagcatcgtcacaagcaagctacgtaaca720
ggtattacactatttgctgatggcggtatgacaaaatatccttctttccaaacgggaaga780
ggc783
<210>5
<211>258
<212>prt
<213>领地伯克霍尔德菌(burkholderiaterritorii)
<400>5
metglyargleugluglylysvalalailevalthrglyglyalaarg
151015
glymetglyalaalathrcysargleuphevalglugluglyalaarg
202530
valvalileglyaspvalleuaspalagluglyglualaleualaarg
354045
gluleuglyaspalaalaargphemetargleuaspvalseraspglu
505560
alaasntrpalaargvalalaaspalathrvalgluglnpheglyarg
65707580
ileaspvalleuvalasnasnalaalavalleuthrpheglyalaile
859095
thrgluleuserlysargaspphegluargalavalserileasnleu
100105110
valglythrphevalglyileargthrilealaproargmetileala
115120125
glnlysargglyserilevalasnileserservalaspglyleuarg
130135140
glyvalasnalaleualaalatyrvalserserlystrpglyvalarg
145150155160
glyleuthrlysvalalaalaleugluleuglyhisglnglyvalarg
165170175
valasnserilehisproglyglyvalasnthralametserasnpro
180185190
thrglyalaproleuaspgluvalasnlyshistyrthrargvalpro
195200205
leuglnargvalglyleuproaspgluilealaargalathrleuphe
210215220
leualaseraspglualasertyrcysasnglyalagluleuserval
225230235240
aspglyglymetalaalaglyalatyrtyrproglyleuproglyala
245250255
prophe
<210>6
<211>263
<212>prt
<213>红球菌属(rhodococcus)
<400>6
metalaargleugluglylysvalalailevalthrglyalaalagln
151015
glymetglyalaalathralaargleuphevalhisgluglyalaarg
202530
valvalleuglyaspvalleugluglulysglyargalaleualaala
354045
gluleuglyaspalaalailephethrproleuaspvalseraspglu
505560
sersertrpgluseralavalalavalalavalaspargpheglygly
65707580
leuaspileleuvalasnasnalaglyvalmethistrpalaproile
859095
gluaspleuaspvalalaargthrgluargleuleuaspileasnval
100105110
leuglyasnleuleuglyalalysalavalvalprothrmetlyslys
115120125
alaglyargglyvalilevalasnileserservalaspglyleuarg
130135140
glyvalasnglyleualaalatyrthralaserlystrpalavalarg
145150155160
glyleuthrlysalaleualatyrgluleuglyproalaglyilearg
165170175
valcysservalhisproglyglyvalaspthrthrleuglyasnpro
180185190
glyglyleuvalglyaspaspleuglnserlystyrvalglyvalpro
195200205
leuglnargileglyglusergluaspilealaargalathrleuphe
210215220
valalaseraspglualasertyrileserglyalagluleualaval
225230235240
aspglyglytrpseralaglythrtyrtyrproglyleuproglythr
245250255
proproalaleumetproasn
260
<210>7
<211>262
<212>prt
<213>鞘氨醇单胞菌属(sphingomonassp.)
<400>7
metglyargleuaspglylysvalalailevalthrglyglyalagln
151015
glymetglyalaalathralaargleuphealaalagluglyalaarg
202530
valvalileglyaspvalilegluasplysglyalagluvalalathr
354045
gluileglyaspasnalaargpheserargleuaspvalargserglu
505560
aspasptrpalaalaalavalseralaalavalglualapheglythr
65707580
ileaspileleuvalasnasnalaalavalvalhisphethrproile
859095
glumetleuaspalaserglualagluargvalleualaileasnval
100105110
leuglyalametleuglyalalyshisvalthrprovalmetthrarg
115120125
alaglyhisglyvalilevalasnileserservalaspglyleuarg
130135140
glycysasnglyleuservaltyrthralaserlystrpalavalarg
145150155160
glyleuserlysthrglnalametgluleuglyproargglyilearg
165170175
valcysserilehisproglyglyvalasnthrglnmetglyasnpro
180185190
metglymetthrglyaspalaleuasnlysasptyrlysargvalpro
195200205
leuglnargileglygluproglugluilealaargalathrleuphe
210215220
ilealaseraspglualasertyrileserglyalagluleualaval
225230235240
aspglyglytrpseralaglytyrtyrglnprovalleuproglyala
245250255
progluglyleuglyala
260
<210>8
<211>261
<212>prt
<213>巨大芽孢杆菌(bacillusmegaterium)
<400>8
mettyrthraspleulysasplysvalvalvalvalthrglyglyser
151015
lysglyleuglyargalametalavalargpheglyglngluglnser
202530
lysvalvalvalasntyrargserasnglugluglualaleugluval
354045
lyslysgluilegluglnalaglyglyglnalaileilevalarggly
505560
aspvalthrlysglugluaspvalvalasnleuvalgluthralaval
65707580
lysglupheglythrleuaspvalmetileasnasnalaglyvalglu
859095
asnprovalproserhisgluleuserleugluasntrpasnglnval
100105110
ileaspthrasnleuthrglyalapheleuglyserargglualaile
115120125
lystyrphevalgluasnaspilelysglyasnvalileasnmetser
130135140
servalhisglumetileprotrpproleuphevalhistyralaala
145150155160
serlysglyglymetlysleumetthrgluthrleualaleuglutyr
165170175
alaprolysglyileargvalasnasnileglyproglyalaileasp
180185190
thrproileasnalaglulysphealaaspprogluglnargalaasp
195200205
valglusermetileprometglytyrileglyasnproglugluile
210215220
alaservalalaalapheleualaserserglnalasertyrvalthr
225230235240
glyilethrleuphealaaspglyglymetthrlystyrproserphe
245250255
glnthrglyarggly
260
<210>9
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>9
cgccatatgatgggccgtctggaaggt27
<210>10
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>10
ccggaattcttaatgatgatggtgatgat29
<210>11
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>11
cgccatatgatggcacgtctggaaggt27
<210>12
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>12
ccggaattcttaatgatgatggtgatgat29
<210>13
<211>27
<212>dna
<213>人工序列(artificialsequence)
<400>13
cgccatatgatgggccgtctggatggt27
<210>14
<211>29
<212>dna
<213>人工序列(artificialsequence)
<400>14
ccggaattcttaatgatgatggtgatgat29
<210>15
<211>45
<212>dna
<213>人工序列(artificialsequence)
<400>15
ccggaattcaaggagatatacatatgatgtatacagatttaaaag45
<210>16
<211>28
<212>dna
<213>人工序列(artificialsequence)
<400>16
ccgctcgagttagcctcttcccgtttgg28
<210>17
<211>6
<212>dna
<213>人工序列(artificialsequence)
<400>17
aaggag6
- 还没有人留言评论。精彩留言会获得点赞!